chronic stress during paradoxical sleep deprivation increases paradoxical sleep rebound: association...
TRANSCRIPT

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Chronic stress during paradoxical sleep deprivationincreases paradoxical sleep rebound: Association withprolactin plasma levels and brain serotonin content
Ricardo Borges Machado, Sergio Tufik, Deborah Suchecki *
Department of Psychobiology, Universidade Federal de Sao Paulo, Sao Paulo, Brazil
Received 26 December 2007; received in revised form 6 June 2008; accepted 19 June 2008
1. Introduction
Numerous studies have pointed out to a sleep rebound-inducing effect of acute stress (Cespuglio et al., 1994;Rampin et al., 1991). This effect follows a temporal pat-
Psychoneuroendocrinology (2008) 33, 1211—1224
KEYWORDSParadoxical sleepdeprivation;Footshock stress;Sleep homeostasis;Heart rate;Prolactin;Corticosterone;Dopamine;Serotonin
Summary Previous studies suggest that stress associated to sleep deprivation methods canaffect the expression of sleep rebound. In order to examine this association and possiblemechanisms, rats were exposed to footshock stress during or immediately after a 96-h periodof paradoxical sleep deprivation (PSD) and their sleep and heart rate were recorded. Control rats(maintained in individual home cages) and paradoxical sleep-deprived (PS-deprived) rats weredistributed in three conditions (1) no footshock — NF; (2) single footshock — SFS: one singlefootshock session at the end of the PSD period (6—8 shocks per minute; 100 ms; 2 mA; for 40 min);and (3) multiple footshock — MFS: footshock sessions with the same characteristics as describedabove, twice a day throughout PSD (at 7:00 h and 19:00 h) and one extra session before therecovery period. After PSD, animals were allowed to sleep freely for 72 h. Additional groups weresacrificed at the end of the sleep deprivation period for blood sampling (ACTH, corticosterone,prolactin and catecholamine levels) and brain harvesting (monoamines andmetabolites). NeitherSFS nor MFS produced significant alterations in the sleep patterns of control rats. All PS-deprivedgroups exhibited increased heart rate which could be explained by increased dopaminergicactivity in the medulla. As expected, PS deprivation induced rebound of paradoxical sleep in thefirst day of recovery; however, PSD + MFS group showed the highest rebound (327.3% above thebaseline). This group also showed intermediate levels of corticosterone and the highest levels ofprolactin, which were positively correlated with the length of PS episodes. Moreover, paradoxicalsleep deprivation resulted in elevation of the serotonergic turnover in the hypothalamus, whichpartly explained the hormonal results, and in the hippocampus, which appears to be related toadaptive responses to stress. The data are discussed in the realm of a prospective importance ofparadoxical sleep for processing of traumatic events.# 2008 Elsevier Ltd. All rights reserved.
* Corresponding author at: Department of Psychobiology — UNI-FESP, Rua Napoleao de Barros 925, Sao Paulo — SP 04024-002, Brazil.Tel.: +55 11 2149 0159; fax: +55 11 5572 5092.
E-mail address: [email protected] (D. Suchecki).
ava i lab le at www.sc ienced i rect .com
journa l homepage: www.e l sev ie r.com/locate/psyneuen
0306-4530/$ — see front matter # 2008 Elsevier Ltd. All rights reserved.doi:10.1016/j.psyneuen.2008.06.007

Author's personal copy
tern insofar as both slow wave sleep (SWS) and paradoxicalsleep (PS) are reduced during stress exposure (Marinescoet al., 1999) and remain so inhibited for some hoursafter the end of the stimulus (Tiba et al., 2004); 2—3 hafter the end of the stimulus, however, sleep beginsto rise, and reaches the highest levels during the darkphase of the cycle (Dewasmes et al., 2004; Koehl et al.,2002; Tiba et al., 2003, 2004). Sleep rebound, and espe-cially, increased slow wave activity during SWS, is believedto play a recovery function that helps restore internalbalance after a traumatic event (Meerlo et al., 1997,2001a). Nevertheless, long periods of immobilizationstress (4 h) have been reported to induce less sleeprebound than shorter periods (1 h or 2 h), suggestingthat prolonged secretion of corticosterone could beinvolved in the suppression of sleep rebound (Marinescoet al., 1999).
Sleep deprivation using instrumental methods, but notstimulation of the midbrain reticular formation, results insleep rebound (Kovalzon and Tsibulsky, 1984). Due to thisdissociation, some authors have claimed that sleeprebound is a consequence of the stress inherent to thesemethods (Coenen and Van Luijtelaar, 1985; Jouvet, 1994;Rechtschaffen et al., 1989). However, when two relatedmethods of PS deprivation (PSD) were compared, itbecame clear that both completely suppressed PS andreduced the amount of SWS to the same extent; none-theless, there was a difference in the resulting sleeprebound between the methods. Thus, rats submitted toPSD by the modified multiple platform method (MMPM)displayed a lager amount of PS rebound than rats PS-deprived by the single platform method (SPM) (Machadoet al., 2004). Interestingly, we have shown previously thatthe immediate activation of the hypothalamic—pituitary—adrenal (HPA) axis after 96 h of PSD is similar for bothmethods (Suchecki et al., 2002a), but the behavioral andphysiological responses of the rats to a psychologicalstressor (the elevated plus maze) is smaller in ratsdeprived by the MMPM than in rats deprived by the SPM(Suchecki et al., 2002b).
Based on this evidence we sought to examine whethera high intensity stressor would be capable to preventthe sleep rebound that follows a prolonged period ofPSD. We hypothesized that acute application of stresswould impair the sleep rebound, whereas chronically itwould not interfere with PS rebound that typically followsa prolonged period of sleep deprivation in the SPM,because animals would have habituated to the stressor.To test this hypothesis, we chose a stressor that had beenshown in previous studies to impair sleep. From the lit-erature data, acute footshock appears to be the mostdetrimental to sleep, increasing sleep latency and reducingthe time spent in PS (Palma et al., 2000; Pawlyk et al.,2005; Sanford et al., 2003a; Vazquez-Palacios and Velaz-quez-Moctezuma, 2000). Therefore, in the present studywe assessed the effects of footshock stress on the expres-sion of sleep rebound and heart rate of PS-deprived rats.The secretion of hormones of the HPA, of prolactin and ofperipheral catecholamines and central monoamines andtheir metabolites were determined in discrete areas ofthe brain to unravel the mechanisms responsible for theobserved changes.
2. Methods
2.1. Subjects
Groups containing 8—10 adult Wistar rats (300—400 g) fromour own colony were used after prior approval from the EthicsResearch Committee of Universidade Federal de Sao Paulowas issued in accordance with international guidelines forcare in animal research. Rats were allowed free access towater and food during all experiments. A constant 12 h lightto 12 h dark cycle was maintained in the experimental roomswith fluorescent white lamps (lights on at 7:00 h) during theentire study. Temperature in the experimental rooms waskept at 20—22 8C.
2.2. Electrophysiological procedures
After anaesthesia with ketamine-diazepam (140.0—5.5 mg/kg, i.p.), the ratswere fittedwith electrodes tomonitor sleep.Two bipolar electrodes placed ipsilaterally with stainless-steelmicro-screws (1 0.9 mm) were used for EEG monitoring: onepair on the right lateral parieto-parietal (for minimum thetaactivity EEG) and the other on the left medial fronto-parietal(for maximum theta activity EEG) areas (Rosenberg et al.,1976; Timo-Iaria et al., 1970). One pair of insulated nickel—chromium flexible fine wire electrodes was implanted in thedorsal neckmuscle for EMG recording and other additional pairof the same material was positioned on either side of the ribcage, bilaterally, to record heart rate (EKG). After the surgicalprocedure,antibiotics (Pentabiotico1Fort-Dodge)and sodiumdiclofenac were administered and the animal was allowed torecover from surgery for 15 days.
Electrophysiological signals were recorded on a digitalpolygraph (Neurofax QP 223 A #Nihon Kohden Co., Tokyo,Japan) and recordings were displayed on 30 s epochs andsubmitted to visual scoring routine, as described previously(Machado et al., 2005). Behavioral states were classified aswakefulness (W); low amplitude slow wave sleep (LSWS), highamplitude slow wave sleep (HSWS) and paradoxical sleep.LSWS is classified by the presence of a voltage in the EEG of20.0—30.0 mV (usually 200—400 mV, peak to peak) and lowfrequency activity (delta waves) in less than 50% of the epoch.HSWSwascharacterizedbyaveragevoltageof30.0 mV (usually250—500 mV, peak to peak) and presence of delta waves in 50%or more of the epoch. Additionally, transition-type sleep(spindles and fast high voltage activity) was considered asHSWS. The sleep—wake states were analyzed off-line in 3 hblocks. The following parameters were compared within eachrecording period throughout the study:
1. Sleep time (latency, total time of sleep represented aspercentage of recording time).
2. Slow wave sleep (time of low [LSWS] and high [HSWS]amplitude fractions, and total time of slow wave sleep,presented as percentage of sleep time).
3. Paradoxical sleep (latency, total time, represented aspercentage of sleep time, number and duration of epi-sodes).
Control and paradoxical sleep-deprived rats submitted tothe single footshock session were not recorded during thesleep deprivation period, because they were exactly the
1212 R.B. Machado et al.

Author's personal copy
same as their non-stressed counterparts, which providedsleep data during the deprivation procedure.
Six seconds of electrocardiograms recordings with nor-mal QRS complex morphology and cycle lengths wereselected for each sleep state (i.e. LSWS, HSWS and PS,when present) at every 3 h and were included in the analysisof sleep heart rate index. Acquisition of the heart rateduring wakefulness was avoided, because of the presence ofinterference and artifacts on EKG signal, especially duringsleep deprivation.
Animals were habituated to the cables and to the record-ing environment for 3 days before baseline recording. Base-line sleep was recorded on two consecutive days (2 � 24-h)and the parameters are represented by the average of these 2days. After the baseline recording, in the period that pre-ceded paradoxical sleep deprivation, animals were adaptedto the sleep deprivation chambers for 30 min per day forthree consecutive days.
2.3. Sleep deprivation procedure
It was accomplished by the single platform method, inwhich each animal was placed onto a narrow cylindricalplatform, 6.5 cm in diameter surrounded by water about1 cm below the platform surface. Temperature of thewater inside the water tanks was similar to the environ-mental temperature (22 8C). Food and water were pro-vided ad libitum throughout the whole study. Hanging foodcontainers were placed inside the water chambers andfilled whenever necessary. Water bottles were fixed onthe grid that covers the water chamber, cleaned andrefilled daily. In addition, the water in the tanks waschanged everyday. After 4 days under this protocol, ratswere allowed to sleep freely in their individual home cages(recovery period) for 3 days (R1—R3), being continuouslymonitored.
2.4. Footshock stress
The footshock stress was administered by placing eachanimal in an individual chamber (30 cm � 20 cm � 30 cm)with electrified floor that delivered unavoidable shocks(6—8 shocks per minute; 100 ms; 2 mA) for 40 min (Palmaet al., 2000). Control and PS-deprived groups were eithersubmitted to: (1) single footshock stress (SFS), appliedimmediately after the end of PSD or at the correspondingtime for control rats. These groups were not recordedduring the PSD period; (2) multiple footshock stress(MFS), applied twice a day, immediately before light shift(at 7:00 h and 19:00 h) to prevent the introduction of apossible additional zeitgeber. Footshock sessions weredelivered during the 4 days of PSD and one extra sessionwas given immediately before the recovery period, sum-ming up nine footshock sessions; (3) no footshock stress(NFS): rats were placed inside the chamber onto theelectrified grid also for 40 min, but no footshock wasdelivered; this group served as a control for the stressprocedure. Therefore, this study was composed by sixgroups: (1) control no footshock stress (CTL + NFS); (2)CTL single footshock stress (CTL + SFS); (3) CTL multiplefootshock stress (CTL + MFS); (4) paradoxical sleep depri-vation NFS (PSD + NFS); (5) PSD + SFS; (6) PSD + MFS.
2.5. Blood analyses
Trunk blood was obtained approximately 2 h after the endof stress procedures from matched groups, run simulta-neously with the sleep study. To prevent animals fromsleeping during these 2 h, and to avoid the effects of sleeprebound on these measurements, rats were placed back intheir previous environments until decapitation was per-formed. Blood was collected in chilled 7.5% EDTA-contain-ing vials, and was centrifuged at 2300 rpm at 4 8C for20 min; plasma was collected and separated for subsequentdetermination of prolactin, ACTH and corticosterone(CORT) levels. All assays were performed in duplicate.Prolactin concentrations were determined by a specificimmunenzimatic commercial kit, based on AChE competi-tive substrate (SPI Bio — Bertin Group, Montigny-le-Breton-neux, France). Plasma ACTH was determined by sequentialimmunometric assay (DPC Immulite, Los Angeles, CA); whennecessary, samples were diluted with appropriate solutionto avoid high-dose hook effect. Corticosterone levels wereevaluated by specific radioimmunoassay (INC Biomedicals,Costa Mesa, CA).
2.6. Monoamine concentrations in discrete brainareas
After 96 h of sleep deprivation and/or stress procedures,animals were sacrificed by decapitation, their brains wereremoved and the frontal and parietal cortex, striatum, hip-pocampus, hypothalamus, medulla and pons regions, sepa-rately, were dissected on a cold surface. The tissue sampleswere weighed individually and homogenized by sonication in500 mL of extraction solution (0.1 M perchloric acid contain-ing 0.4 mM sodium metabissulfite and 0.2 mM EDTA). Thehomogenates were centrifuged at 20,000 � g for 10 min,then filtered through 0.22 mm membrane and stoked at�80 8C for further analysis. Precipitates were dissolved in0.1N NaOH and assayed to protein estimation (Bicinchoninicacid method, Pierce Chemical, Rockford, IL).
Supernatants were submitted to fast isocratic separationthrough a C18 HPLC reversed-phase column system (Spheri-5,C18, ODS, 5 mm, 25 cm � 4.6 mm column; linked to a New-Guard Cartridge Column, RP-18, 7 mm pre-column; PerkinEl-mer Brownlee Columns, Shelton, CT) and electrochemicallydetected using an amperometric detector (L-ECD-6A, Shi-madzu, Japan), by oxidation on glass carbon electrode at+850 mV in relation to an Ag—AgCl reference electrode. Themobile phase consisted of 0.163 M citric acid, 0.06 M sodiumphosphate dibasic anhydrous, 0.69 mM octyl sodium sulfate,12 mM EDTA, acetonitrile 4%, tetrahydrofuran 1.7% andorthophosphoric acid sufficient to bring the pH to 2.85,diluted in double distilled water. The mobile phase wasfiltered through a 0.2 mm filter membrane, degassed underhelium and delivered at a flow rate of 1.4 mL/min. Eachsample was analyzed in duplicate for concentrations ofnorepinephrine (NE), dopamine (DA), serotonin (5-HT) andtheir non-conjugated metabolites 3,4-dihydroxyphenilaceticacid (DOPAC), homovanilic acid (HVA) and 5-hydroxyindolea-cetic acid (5-HIAA). The recovery of the analytes was deter-mined by adding a fixed concentration of internal standardDHBA (dihydroxybenzylamine) before tissue homogenization.All standards and salts were purchased from Sigma (St. Louis,
Chronic stress during paradoxical sleep deprivation increases rebound of paradoxical sleep 1213

Author's personal copy
MO) and the solvents (HPLC grade) were purchased fromCarlo Erba (Italy).
Although monoamines were measured in various brainregions, only data regarding those areas directly involvedwith the behavioral, physiological and endocrine changesobserved in the present study will be presented.
2.7. Statistical analyses
EEG data were analyzed by a two-way ANOVA for repeatedmeasures,withmain factorsGroup (Control [CTL], Paradoxicalsleep deprivation [PSD]), Stress (no footshock stress [NFS],single footshock stress [SFS] and multiple footshock stress[MFS]) and Day (repeated measure: Baseline, Recovery day1 [R1], Recovery day 2 [R2] andRecoveryday3 [R3]). Sleepwasalso recorded during the sleep deprivation period; howeverthesedatawill notbepresentedbecauseno relevant effects offootshockwereobserved. For EKGdata,ANOVAwas performedin two steps: during the deprivation period, the main factorswere Group (CTL, PSD), Stress (NFS, MFS) and Day (repeatedmeasure: Baseline, Day 1 [D1], Day 2 [D2], Day 3 [D3] and Day 4[D4]); during the rebound period, the factors were Group,Stress (no footshock stress [NFS], single footshock stress [SFS]and multiple footshock stress [MFS]) and Day (repeated mea-sure: Baseline, Recovery day 1 [R1], Recovery day 2 [R2] andRecovery day 3 [R3]). This two-step analysis was necessarybecause CTL + SFS and PSD + SFS groups were not recordedduring the sleep deprivation period, since these groups wereexactly the same as CTL + NFS and PSD + NFS. Hormone valuesof non-stressed groups (basal levels) were initially analyzed bya Student’s t-test. Hormonal and neurochemical data werecarried out by a two-way ANOVA, with main factors Group(CTL, PSD) and Stress (NFS, SFS, MFS). Post-hoc analysis wasperformed by the Newman—Keuls test. Finally, Pearson’s cor-relation tests were performed between the percentage of PSobtained in thefirst 12-hof recoveryperiodandeachoneof thehormone levels for CTL and PS-deprived animals, separately;between plasma hormone concentrations and monoaminecontents in the hypothalamus, and between monoamine con-tents and sleepparameters. The level of significancewas set atp � 0.05.
3. Results
3.1. Heart rate during sleep (Table 1)
A three-way interaction during sleep deprivation was shownF(4,128) = 2.76; p � 0.05. Regardless of the stress regimen, PS-deprived rats exhibited higher heart rate throughout theentire sleep deprivation period when compared to theirrespective CTL counterparts ( p � 0.005) and to their base-line heart rate ( p � 0.001). No differences were observed inCTL rats.
During the recovery period, heart rate was influenced byan interaction of all factors F(6,138) = 6.59; p � 0.0001. Ana-lysis of this interaction showed that all PS-deprived ratsexhibited higher heart rate on R1 than on baseline( p � 0.05). Moreover, a difference due to stress regimenwas also observed, in which the heart rate of PSD + MFS ratswas lower than that of PSD + NFS rats on R1 ( p � 0.01).Finally, PS-deprived rats showed higher heart rate than theirrespective CTL counterparts on R1 ( p � 0.05).
Table
1Heartrate
duringsleepin
PS-deprive
dandCTLrats,submittedornotto
singleormultiple
footshock
stress
Group
Stress
Baseline
D1
D2
D3
D4
R1
R2
R3
CLT
NFS
304.42�
5.9
293.25�
2.9
289.17�
3.0
295.58�
4.2
290
.83�
5.0
288.21�
11.6
286.38�
4.6
289.13�
3.0
SFS
296.38�
7.2
NR
NR
NR
NR
300.21�
8.9
306.30�
9.2
297.14�
8.9
MFS
291.82�
4.71
294.38�
4.2
287.08�
4.8
287.76�
3.5
291
.93�
6.2
289.06�
7.0
281.04�
4.0
288.59�
2.6
PSD
NFS
296.63�
7.8
365.75�
6.0
a,b
401.69�
12.0
a,b
448.75�
9.4a,b
467
.56�
9.1a,b
378.54�
2.7a,b
343.38�
10.6
325.63�
5.3
SFS
294.53�
4.9
NR
NR
NR
NR
357.40�
3.1a,b
331.04�
4.2
324.17�
3.0
MFS
315.63�
5.0
372.19�
5.3a,b
410.23�
5.9a,b
441.56�
7.4a,b
462
.58�
6.0a,b
335.99�
4.8a,b,c
318.80�
3.8
311.82�
3.2
Theva
luesareexp
ressedin
beatsperminute
(b.p.m
.)�
S.E.M
.of8—
10rats/g
roup.CLT,home-cag
eco
ntrols;PSD
,parad
oxica
lsleep-deprive
d;NFS,non-footshock
stress;SFS,
singlefootshock
stress;MFS,multiple
footshock
stress;D,sleepdepriva
tionperiod;R,reco
very
period;NR,non-reco
rded.
aDifferentfrom
baselineleve
ls.
bDifferentfrom
respectiveCTLgroup.
cDifferentfrom
respectiveNFS
group.
1214 R.B. Machado et al.

Author's personal copy
3.2. Sleep parameters
3.2.1. Latency to sleep (Fig. 1)Analysis of the light phase showed a three-way interactionF (6,138) = 2.6376; p � 0.02, in which SFS delayed the sleeponset of CTL rats compared to baseline ( p � 0.01), but didnot alter this parameter in PS-deprived rats.
During the dark phase there was a three way interactionF (6,138) = 2.905; p � 0.01. Analysis of this interaction showedthat the sleep latency of CTL + NFS rats was reduced in allrecovery nights, when compared to baseline ( p � 0.01).
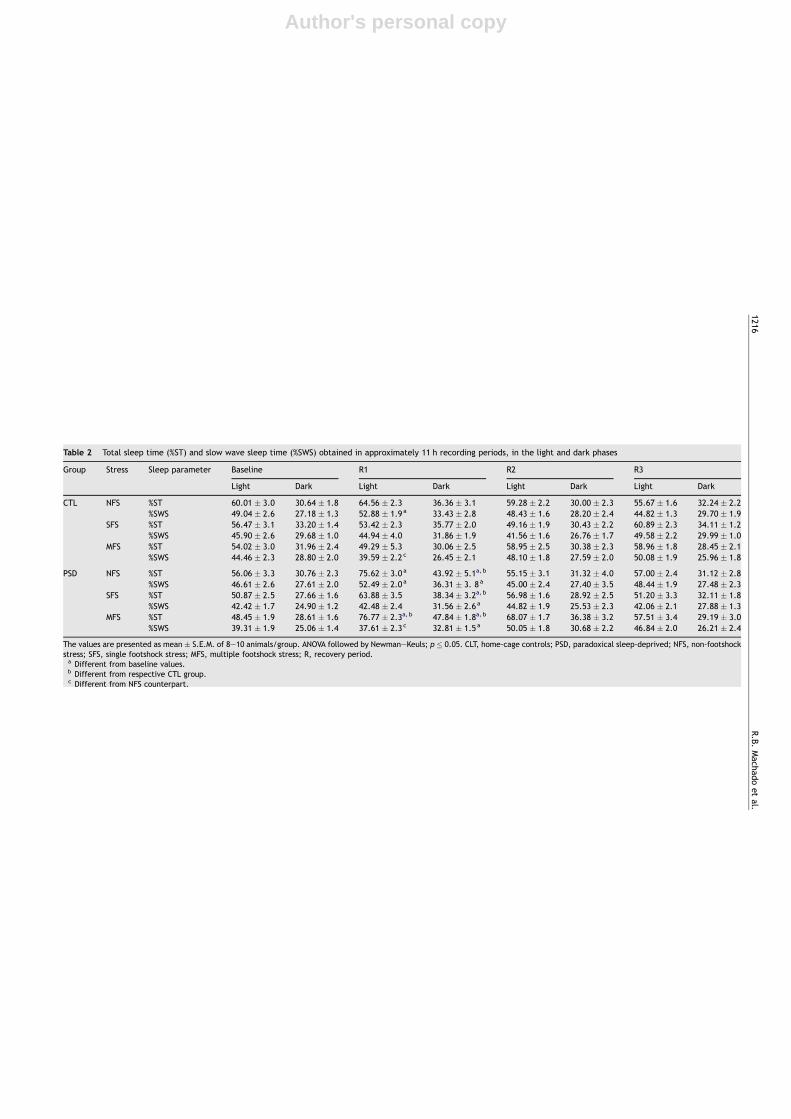
3.2.2. Total sleep time (Table 2)A three-way interaction was revealed during the light phaseF (6,138) = 3.531; p � 0.005. PSD resulted in greater sleep timeduring recovery day 1 (R1) than baseline; in addition, SFSapplied immediately before the rebound period preventedthe increase in sleep time, whereas MFS led to the highestsleep efficiency in R1 ( p � 0.01); this increase was stillsignificant during R2 ( p � 0.05).
In the dark phase there was a Day by Group interactionF (3,138) = 9.643; p � 0.001, in which PS-deprived rats sleptmore than control animals in R1 ( p � 0.001).
3.2.3. Slow wave sleep (Table 2)Because the statistical analysis did not indicate any substan-tial impact of the stress procedures on HSWS and LSWS, bothfractions were pooled together to represent total slow wavesleep.
During the light phase, an interaction between Stress andDay was observed F (6,138) = 8.879; p � 0.0001, and rats sub-mitted to MFS exhibited less slow wave sleep than thosewhich were not submitted to footshock ( p � 0.001). Duringthe dark phase, a Group by Day interaction was detectedF (6,138) = 3.005; p � 0.05 and the post hoc analysis revealed
that PSD rats exhibited more SWS in R1 compared to baseline( p � 0.0001), but were not different from CTL rats.
3.2.4. Paradoxical sleep3.2.4.1. Latency to paradoxical sleep. Similarly to theeffects described for sleep latency, an interaction betweenStress and Day was detected F (3,138) = 5.682; p � 0.001 in thelight phase, indicating that single footshock stress induced adelayed onset of PS ( p � 0.005), with a subsequent return tobaseline latency afterwards (R2). In the dark phase there wasa main effect of Day F (3,138) = 3.858; p � 0.01, in which PSlatency in the first dark period was shortened compared tobaseline ( p � 0.01).
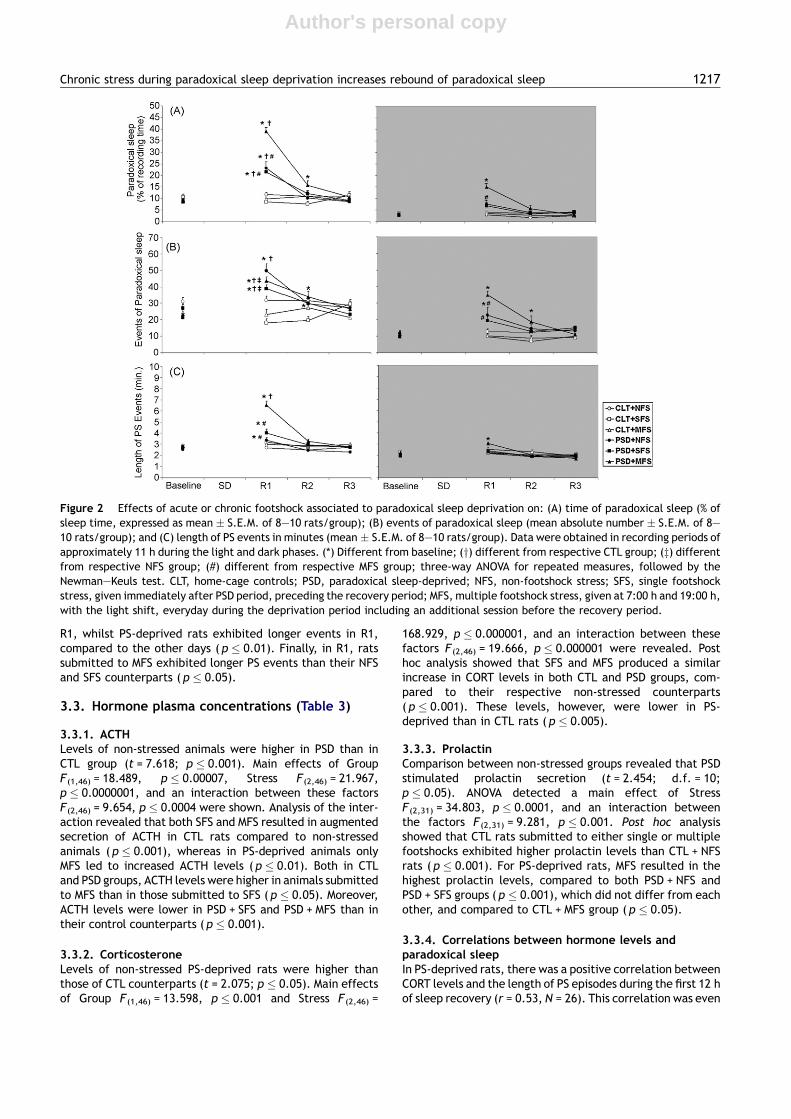
3.2.4.2. Percentage of paradoxical sleep (Fig. 2A). Lightphase: During the rebound period, there was a three-wayinteraction F(6,138) = 7.207; p � 0.001. All PS-deprived ratsexhibited more paradoxical sleep than CTL rats ( p � 0.001)and than their respective basal levels ( p � 0.0005) in R1;however, the association of sleep deprivation with repeatedfootshock stress produced a further increase in this para-meter when compared to the other PS-deprived groups( p � 0.0005); this increase persisted until R2 ( p � 0.001,compared to baseline).
Dark phase: An interaction Stress by Group by Day wasshown F(6,138) = 4.789; p � 0.001. All PS-deprived groupsexhibited more PS during R1 than baseline ( p � 0.001);however, this increase was even higher in PSD + MFS ratsthan in PSD + SFS, PSD + NFS and CTL + MFS rats ( p � 0.001).
3.2.4.3. Events of paradoxical sleep (Fig. 2B). Light phase:Interactions between Day and Group F(3,138) = 29.119;p � 0.001 and between Day and Stress F (3,127) = 3.127;p � 0.01 were obtained. Post hoc analysis indicated thatPS-deprived rats exhibited more events of PS in R1 and R2,than baseline and R3 ( p � 0.05). Compared to controlgroups, PS-deprived rats showed more PS events in R1( p � 0.05). Both stress regimens reduced the number of PSevents in R1, compared to non footshock stress ( p � 0.05).For NFS rats, PS events were higher in R1 than in all other days( p � 0.05). Rats treated with MFS exhibited more PS eventsin R1 and R2 than baseline and R3 ( p � 0.05).
Dark phase: A three way interaction during sleep recoveryperiod was observed F(6,138) = 2.637; p � 0.05. The New-man—Keuls analysis revealed that PSD + MFS rats exhibitedmore events of PS in R1 than in all other recovery days( p � 0.001) and than all other groups ( p � 0.01), and thisincrease persisted until R2 ( p � 0.001). In addition, thegroup PSD + NFS also displayed more events of PS in R1 thanin all other days ( p � 0.05).
3.2.4.4. Length of PS events (Fig. 2C). Light phase: A three-way interaction was obtained F(6,138) = 9.525; p � 0.001.Analysis of this interaction showed that all PS-deprivedgroups exhibited longer PS events during R1 than baseline,R2 and R3 ( p � 0.005). During the first day of recovery, thelength of PS events was higher in PSD + MFS rats than inPSD + NFS and PSD + SFS and CTL + MFS rats ( p � 0.005).
Dark phase: Interactions between Day and GroupF(6,138) = 3.116; p � 0.05 and between Day and StressF(6,138) = 2.684; p � 0.01 were observed. For CTL groups,the length of PS events was reduced in R3, compared to
Figure 1 Sleep latency (time elapsed between the onset ofsleep recording and sleep onset, taken as the first three epochsof continuous sleep)measured in R1, immediately after the end ofPSD or at the corresponding time for CTL rats or immediately afterthe end of footshock session (CTL + SFS, CTL + MFS, PSD + SFS,PSD + MFS). The latency is presented in min as mean � S.E.M. of8—10 animals/group. (*) Different from baseline values; three-way ANOVA for repeated measures, followed by the Newman—Keuls test. CLT, home-cage controls; PSD, paradoxical sleep-deprived; NFS, non-footshock stress; SFS, single footshock stress,given immediately after PSD period, preceding the recoveryperiod; MFS, multiple footshock stress, given at 7:00 h and19:00 h, with the light shift, everyday during the deprivationperiod including an additional session before the recovery period.
Chronic stress during paradoxical sleep deprivation increases rebound of paradoxical sleep 1215
Author's personal copy
Table 2 Total sleep time (%ST) and slow wave sleep time (%SWS) obtained in approximately 11 h recording periods, in the light and dark phases
Group Stress Sleep parameter Baseline R1 R2 R3
Light Dark Light Dark Light Dark Light Dark
CTL NFS %ST 60.01 � 3.0 30.64 � 1.8 64.56 � 2.3 36.36 � 3.1 59.28 � 2.2 30.00 � 2.3 55.67 � 1.6 32.24 � 2.2%SWS 49.04 � 2.6 27.18 � 1.3 52.88 � 1.9a 33.43 � 2.8 48.43 � 1.6 28.20 � 2.4 44.82 � 1.3 29.70 � 1.9
SFS %ST 56.47 � 3.1 33.20 � 1.4 53.42 � 2.3 35.77 � 2.0 49.16 � 1.9 30.43 � 2.2 60.89 � 2.3 34.11 � 1.2%SWS 45.90 � 2.6 29.68 � 1.0 44.94 � 4.0 31.86 � 1.9 41.56 � 1.6 26.76 � 1.7 49.58 � 2.2 29.99 � 1.0
MFS %ST 54.02 � 3.0 31.96 � 2.4 49.29 � 5.3 30.06 � 2.5 58.95 � 2.5 30.38 � 2.3 58.96 � 1.8 28.45 � 2.1%SWS 44.46 � 2.3 28.80 � 2.0 39.59 � 2.2c 26.45 � 2.1 48.10 � 1.8 27.59 � 2.0 50.08 � 1.9 25.96 � 1.8
PSD NFS %ST 56.06 � 3.3 30.76 � 2.3 75.62 � 3.0a 43.92 � 5.1a, b 55.15 � 3.1 31.32 � 4.0 57.00 � 2.4 31.12 � 2.8%SWS 46.61 � 2.6 27.61 � 2.0 52.49 � 2.0a 36.31 � 3. 8a 45.00 � 2.4 27.40 � 3.5 48.44 � 1.9 27.48 � 2.3
SFS %ST 50.87 � 2.5 27.66 � 1.6 63.88 � 3.5 38.34 � 3.2a, b 56.98 � 1.6 28.92 � 2.5 51.20 � 3.3 32.11 � 1.8%SWS 42.42 � 1.7 24.90 � 1.2 42.48 � 2.4 31.56 � 2.6a 44.82 � 1.9 25.53 � 2.3 42.06 � 2.1 27.88 � 1.3
MFS %ST 48.45 � 1.9 28.61 � 1.6 76.77 � 2.3a, b 47.84 � 1.8a, b 68.07 � 1.7 36.38 � 3.2 57.51 � 3.4 29.19 � 3.0%SWS 39.31 � 1.9 25.06 � 1.4 37.61 � 2.3c 32.81 � 1.5a 50.05 � 1.8 30.68 � 2.2 46.84 � 2.0 26.21 � 2.4
The values are presented as mean � S.E.M. of 8—10 animals/group. ANOVA followed by Newman—Keuls; p � 0.05. CLT, home-cage controls; PSD, paradoxical sleep-deprived; NFS, non-footshockstress; SFS, single footshock stress; MFS, multiple footshock stress; R, recovery period.a Different from baseline values.b Different from respective CTL group.c Different from NFS counterpart.
121
6R.B.Mach
adoetal.
Author's personal copy
R1, whilst PS-deprived rats exhibited longer events in R1,compared to the other days ( p � 0.01). Finally, in R1, ratssubmitted to MFS exhibited longer PS events than their NFSand SFS counterparts ( p � 0.05).
3.3. Hormone plasma concentrations (Table 3)
3.3.1. ACTHLevels of non-stressed animals were higher in PSD than inCTL group (t = 7.618; p � 0.001). Main effects of GroupF(1,46) = 18.489, p � 0.00007, Stress F(2,46) = 21.967,p � 0.0000001, and an interaction between these factorsF(2,46) = 9.654, p � 0.0004 were shown. Analysis of the inter-action revealed that both SFS and MFS resulted in augmentedsecretion of ACTH in CTL rats compared to non-stressedanimals (p � 0.001), whereas in PS-deprived animals onlyMFS led to increased ACTH levels (p � 0.01). Both in CTLand PSD groups, ACTH levels were higher in animals submittedto MFS than in those submitted to SFS (p � 0.05). Moreover,ACTH levels were lower in PSD + SFS and PSD + MFS than intheir control counterparts (p � 0.001).
3.3.2. CorticosteroneLevels of non-stressed PS-deprived rats were higher thanthose of CTL counterparts (t = 2.075; p � 0.05). Main effectsof Group F (1,46) = 13.598, p � 0.001 and Stress F (2,46) =
168.929, p � 0.000001, and an interaction between thesefactors F (2,46) = 19.666, p � 0.000001 were revealed. Posthoc analysis showed that SFS and MFS produced a similarincrease in CORT levels in both CTL and PSD groups, com-pared to their respective non-stressed counterparts( p � 0.001). These levels, however, were lower in PS-deprived than in CTL rats ( p � 0.005).
3.3.3. ProlactinComparison between non-stressed groups revealed that PSDstimulated prolactin secretion (t = 2.454; d.f. = 10;p � 0.05). ANOVA detected a main effect of StressF(2,31) = 34.803, p � 0.0001, and an interaction betweenthe factors F (2,31) = 9.281, p � 0.001. Post hoc analysisshowed that CTL rats submitted to either single or multiplefootshocks exhibited higher prolactin levels than CTL + NFSrats ( p � 0.001). For PS-deprived rats, MFS resulted in thehighest prolactin levels, compared to both PSD + NFS andPSD + SFS groups ( p � 0.001), which did not differ from eachother, and compared to CTL + MFS group ( p � 0.05).
3.3.4. Correlations between hormone levels andparadoxical sleepIn PS-deprived rats, there was a positive correlation betweenCORT levels and the length of PS episodes during the first 12 hof sleep recovery (r = 0.53, N = 26). This correlation was even
Figure 2 Effects of acute or chronic footshock associated to paradoxical sleep deprivation on: (A) time of paradoxical sleep (% ofsleep time, expressed as mean � S.E.M. of 8—10 rats/group); (B) events of paradoxical sleep (mean absolute number � S.E.M. of 8—10 rats/group); and (C) length of PS events in minutes (mean � S.E.M. of 8—10 rats/group). Data were obtained in recording periods ofapproximately 11 h during the light and dark phases. (*) Different from baseline; (y) different from respective CTL group; (z) differentfrom respective NFS group; (#) different from respective MFS group; three-way ANOVA for repeated measures, followed by theNewman—Keuls test. CLT, home-cage controls; PSD, paradoxical sleep-deprived; NFS, non-footshock stress; SFS, single footshockstress, given immediately after PSD period, preceding the recovery period; MFS, multiple footshock stress, given at 7:00 h and 19:00 h,with the light shift, everyday during the deprivation period including an additional session before the recovery period.
Chronic stress during paradoxical sleep deprivation increases rebound of paradoxical sleep 1217

Author's personal copy
stronger for prolactin levels (r = 0.71, N = 19). Moreover,there was a significant correlation between prolactin levelsand total time of paradoxical sleep during the first 12 h ofsleep rebound (r = 0.53, N = 19).
3.4. Monoamine concentrations in discrete brainareas (Fig. 3 and Table 4)
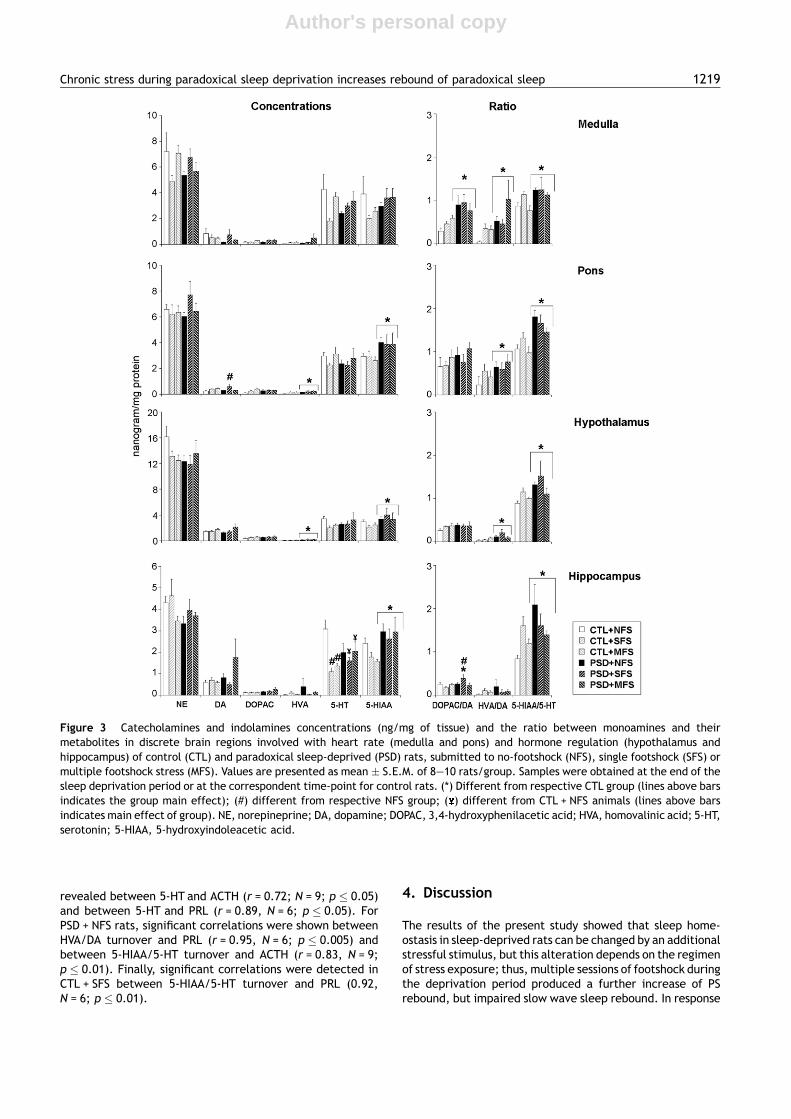
3.4.1. MedullaA main effect of group was detected for DOPAC/DA ratio(F(1,46) = 12.6287, p � 0.001), HVA/DA ratio (F(1,46) =8.06549, p � 0.01) and 5-HIAA/5-HT ratio (F(1,45) = 6.7713,p � 0.05). For all indexes, PSD rats exhibited higher turnoverthan CTL rats (p � 0.01).
3.4.2. PonsNo effects were observed in NA and 5-HT levels and in theDOPAC/DA ratio.
DA: A main effect of stress was detected F (2,46) = 5.2860,p � 0.01, and rats submitted to SFS showed higher levels ofthis neurotransmitter than NFS rats, regardless of the group(+86.48%; p � 0.01).
DOPAC: ANOVA showed a main effect of stressF(2,46) = 4.0688, p � 0.05; in this case, rats exposed to MFSshowed an 85.11% increase above NFS levels ( p < 0.05).
HVA: Main effects of group F(1,46) = 4.18543, p � 0.05 andstress F(2,46) = 3.67107, p � 0.05 were revealed. PSD ratsshowed higher levels of HVA than CTL rats (+62.1%;p � 0.05) and animals submitted to both forms of stressexhibited higher levels than non-stressed rats ( p � 0.05).
5-HIAA: There was an effect of group F (1,46) = 6.1135,p � 0.05 on this metabolite, and again PSD rats exhibitedhigher levels than CTL rats (+37.69%; p � 0.05).
HVA/DA ratio: A main effect of Group was detectedF(1,46) = 4.16770, p � 0.05, and PSD rats showed increaseddopamine turnover when compared to CTL rats (+68.27%;p � 0.05).
5-HIAA/5-HT ratio: Again, a main effect of Group wasrevealed F(1,46) = 21.6665, p � 0.0005 and augmented sero-tonin turnover was shown in PSD compared to CTL rats(+46.91%).
3.4.3. HypothalamusNo changes were observed in NA, DOPAC and 5-HT concen-trations and in the DOPAC/DA ratio.
DA: Only a main effect of stress was detectedF (2,46) = 3.9701, p � 0.05. Rats submitted to MFS showedhigher concentrations of the neurotransmitter than ratssubmitted to SFS (+41.64%; p � 0.05), which in turn werehigher than NFS rats (+34.52%; p < 0.05).
HVA: A main effect of group was revealedF (1,46) = 9.82643, p � 0.005; in this case, PSD rats exhibitedhigher levels of the metabolite than CTL rats (+139.33%).
5-HIAA: A main effect of group was detectedF (1,46) = 4.9575, p � 0.05, in which metabolite levels werehigher in PSD than CTL rats (+42.11%).
HVA/DA ratio: A main effect of Group was shownF (1,46) = 8.33912, p � 0.01. Once again, PSD rats exhibitedhigher ratio than CTL rats (+169.65%).
5-HIAA/5-HT ratio: ANOVA detected a main effect ofGroup F(1,45) = 7.2032, p � 0.01, in which PSD rats exhibitedincreased serotonergic turnover compared to CTL rats(+39.87%).
3.4.4. HippocampusNo differences were found in NA, DA and HVA levels, and inthe HVA/DA ratio.
DOPAC: Main effect of group was shown F(1,46) = 5.62480,p � 0.05. Higher metabolite levels were found in PSD than inCTL rats (+64.35%).
5-HT: There was a significant interaction between groupand stress F (2,46) = 4.4155, p � 0.05). Post-hoc analysisdetected a reduction of 5-HT levels in the hippocampus ofstressed animals (CTL + SFS, PSD + SFS, CTL + MFS andPSD + MFS) compared to CTL + NFS animals ( p � 0.05).
DOPAC/DA ratio: ANOVA revealed a significant interactionbetween the factors F(2,46) = 3.7324, p � 0.05). Newman—Keuls analysis of this interaction showed that PSD + SFSexhibited an increase in dopaminergic turnover comparedto PSD + NFS and CTL + SFS groups (+55.98% and + 122.88%,respectively; p � 0.05).
5-HIAA/5-HT: Main effect of group was shownF (1,46) = 3.9099, p � 0.05, with PSD rats showing increasedserotonergic turnover than CTL rats (+39.94%).
3.4.5. Correlations between concentrations ofhypothalamic neurotransmitters and pituitaryhormonesPearson’s correlation tests were carried out for each group,individually. Significant correlations in CTL + NFS group were
Table 3 Plasma levels of ACTH, corticosterone, prolactin and catecholamines in control (CLT) or PS-deprived (PSD) rats submittedto single (SFS), multiple sessions (MFS) of footshock stress or non-footshock stress (NFS)
Group Stress ACTH (pg/mL) Corticosterone (mg/dL) Prolactin (ng/mL)
CLT NFS 21.80 � 3.4 8.88 � 2.0 4.38 � 0.8SFS 839.57 � 77.5a 76.26 � 3.6a 61.48 � 15.5a
MFS 1066.20 � 187.0a, b 80.29 � 3.2a 65.23 � 10.8a
PSD NFS 134.18 � 14.3c 23.04 � 4.8c 24.12 � 15.6c
SFS 244.64 � 60.7c 53.21 � 5.0c, a 26.47 � 13.6c
MFS 433.80 � 27.7c,a, b 59.00 � 3.9c, a 89.30 � 23.0c,a, b
Values are presented as mean � S.E.M. of 6—10 animals/group. CLT, home-cage controls; PSD, paradoxical sleep-deprived; NFS, non-footshock stress; SFS, single footshock stress; MFS, multiple footshock stress.a Different from respective NFS animals.b Different from respective SFS rats.c Different from respective CTL group.
1218 R.B. Machado et al.
Author's personal copy
revealed between 5-HT and ACTH (r = 0.72; N = 9; p � 0.05)and between 5-HT and PRL (r = 0.89, N = 6; p � 0.05). ForPSD + NFS rats, significant correlations were shown betweenHVA/DA turnover and PRL (r = 0.95, N = 6; p � 0.005) andbetween 5-HIAA/5-HT turnover and ACTH (r = 0.83, N = 9;p � 0.01). Finally, significant correlations were detected inCTL + SFS between 5-HIAA/5-HT turnover and PRL (0.92,N = 6; p � 0.01).
4. Discussion
The results of the present study showed that sleep home-ostasis in sleep-deprived rats can be changed by an additionalstressful stimulus, but this alteration depends on the regimenof stress exposure; thus, multiple sessions of footshock duringthe deprivation period produced a further increase of PSrebound, but impaired slow wave sleep rebound. In response
Figure 3 Catecholamines and indolamines concentrations (ng/mg of tissue) and the ratio between monoamines and theirmetabolites in discrete brain regions involved with heart rate (medulla and pons) and hormone regulation (hypothalamus andhippocampus) of control (CTL) and paradoxical sleep-deprived (PSD) rats, submitted to no-footshock (NFS), single footshock (SFS) ormultiple footshock stress (MFS). Values are presented as mean � S.E.M. of 8—10 rats/group. Samples were obtained at the end of thesleep deprivation period or at the correspondent time-point for control rats. (*) Different from respective CTL group (lines above barsindicates the group main effect); (#) different from respective NFS group; ( ) different from CTL + NFS animals (lines above barsindicates main effect of group). NE, norepineprine; DA, dopamine; DOPAC, 3,4-hydroxyphenilacetic acid; HVA, homovalinic acid; 5-HT,serotonin; 5-HIAA, 5-hydroxyindoleacetic acid.
Chronic stress during paradoxical sleep deprivation increases rebound of paradoxical sleep 1219
Author's personal copy
to the additional forms of stress, PS-deprived rats displayedsmaller ACTH and CORT responses than their CTL counter-parts, despite the fact that PSD alone induced higher levels ofthese hormones compared to CTL non-stressed rats. Inter-estingly, the increase in heart rate induced by PSD was notaltered by either single or multiple footshock stress, indicat-ing that sleep deprivation might have induced a ceilingeffect, which persisted during the first day of recovery.
The persistent tachycardia herein reported is in accor-dance with previous publications using either prolonged PSD(De Mesquita and Hale, 1992) or short (48 h) periods of totalsleep deprivation (Sgoifo et al., 2006). According to theauthors of the latter study, sleep deprivation causes ablunted vagal tone following the sympathetic activation(Sgoifo et al., 2006). This is a possible explanation for ourresults, although we did not find increased catecholaminesperipheral levels in PSD rats (data not shown). In the brain,however, PSD deprivation produced a significant increase indopaminergic turnover in the medulla and pons (HVA/DA andDOPAC/DA), which seemed to be implicated in the augmen-ted heart rate observed in all PS-deprived rats, regardless ofthe stress regimen. It has been shown that local administra-tion of dopamine in the nucleus tratus solitarius producestachycardia and hypertension in rats (Granata and Woodruff,1982), possibly acting at the D2 receptors (Yang et al., 1990).Likewise, painful mechanical stimulation increases heartrate and mean arterial pressure and that microdyalisatestaken from the rostral ventrolateral medulla show increased5-HTand DA, and reduced NA concentrations (Karlsson et al.,2006). Moreover, local administration of serotonin in therostral part of the ventrolateral medulla leads to tachycardiain rats (Lovick, 1989), although administration of 5-HT1Aagonists in the intermediate part results in low blood pres-sure and bradycardia (Mandal et al., 1990).
Regarding the sleep pattern, the main effect of chronicfootshock during sleep deprivation was a robust increase inthe homeostatic drive to paradoxical sleep, due especially toa major lengthening of PS events. During the first 3 h ofrebound we observed events of approximately 12 min inaverage, which correspond to the duration of a whole sleep
cycle in naıve rats (Zepelin, 1994). Such specific effect hasonly been reported after i.c.v. administration of corticotro-phin-like intermediate lobe peptide (CLIP or ACTH18—39)(Chastrette et al., 1990; Wetzel et al., 1997), which isprocessed from the ACTH sequence of pro-opiomelanocortin(POMC) in two distinct locations of the brain, the nucleustractus solitarius and the arcuate nucleus of the hypothala-mus (Leger et al., 1990). The lower levels of ACTH seen inPSD + SFS and PSD + MFS could be due to a shift in pro-opiomelanocortin processing from ACTH to CLIP production.However, the hypothesis about the involvement of CLIP in theexpression of PS rebound is speculative and will soon betested. A more straightforward explanation for the lowerACTH levels in stressed PSD rats is the down-regulation oftype 1 corticotropin-releasing factor (CRF) receptors (CRFR1)in the pituitary resulting in reduced pituitary responsivenessto CRF stimulation. In a recent study (unpublished data), weobserved that chronic administration of CRF to PS-deprivedrats yielded a similar pattern of ACTH secretion, whereasremoval of the glucocorticoid negative feedback by means ofmetyrapone chronic administration did not alter the capacityof pituitary to secrete ACTH, suggesting that PSD may have agreater impact on CRF than on glucocorticoid receptors.These results are in agreement with Fadda and Fratta’sfindings (1997) of a 38% reduction of CRF receptors in thepituitary of paradoxical sleep-deprived rats.
Glucocorticoids possess permissive or suppressive effectson many functions, depending on the circulating levels.Optimal concentrations, i.e., sufficient to saturate highaffinity mineralocorticoid receptors and approximately 50%of low affinity glucocorticoid receptors (Lupien et al., 2007),are required for optimal outcome. Thus, in response tostress, intermediate levels of circulating glucocorticoids,favor behavioral and physiological processes including learn-ing and memory, the activity of the immune system andsleep, whereas both very low and very high concentrationscan cause impairment (Sapolsky et al., 2000). This glucocor-ticoid-induced PS sleep phenomenon is clearly seen in ratssubmitted to different lengths of immobilization stress (Mar-inesco et al., 1999) and in Addison’s patients, whose impaired
Table 4 Summary of the findings of neurotransmitter concentrations in several brain areas, determined by HPLC in 10 animals/group
Pons Medulla Hypothalamus Hippocampus
NE — — — —DA Stress: MFS > NFS — Stress: MFS > SFS > NFS —HVA Group: PSD > CTL;
Stress: SFS and MFS > NFS— Group: PSD > CTL —
DOPAC Stress: MFS > NFS — — Group: PSD > CTL5-HT — — — Interaction: PSD + SFS and
PSD + MFS < CTL + NFS; CTL+ MFS and SFS < CTL + NFS
5-HIAA Group: PSD > CTL — Group: PSD > CTL Group: PSD > CTLHVA/DA Group: PSD > CTL Group: PSD > CTL Group: PSD > CTL —DOPAC/DA — Group: PSD > CTL — Interaction: PSD + SFS >
PS + NFS and CTL + SFS5-HIAA/5-HT Group: PSD > CTL Group: PSD > CTL Group: PSD > CTL Group: PSD > CTL
CTL, home-cage controls; PSD, paradoxical sleep-deprived; NFS, non-footshock stress; SFS, single footshock stress; MFS, multiple footshockstress; NE, norepinephrine; DA, dopamine; DOPAC, 3,4-hydroxyphenilacetic acid; HVA, homovalinic acid; 5-HT, serotonin; 5-HIAA,5-hydroxyindoleacetic acid.
1220 R.B. Machado et al.

Author's personal copy
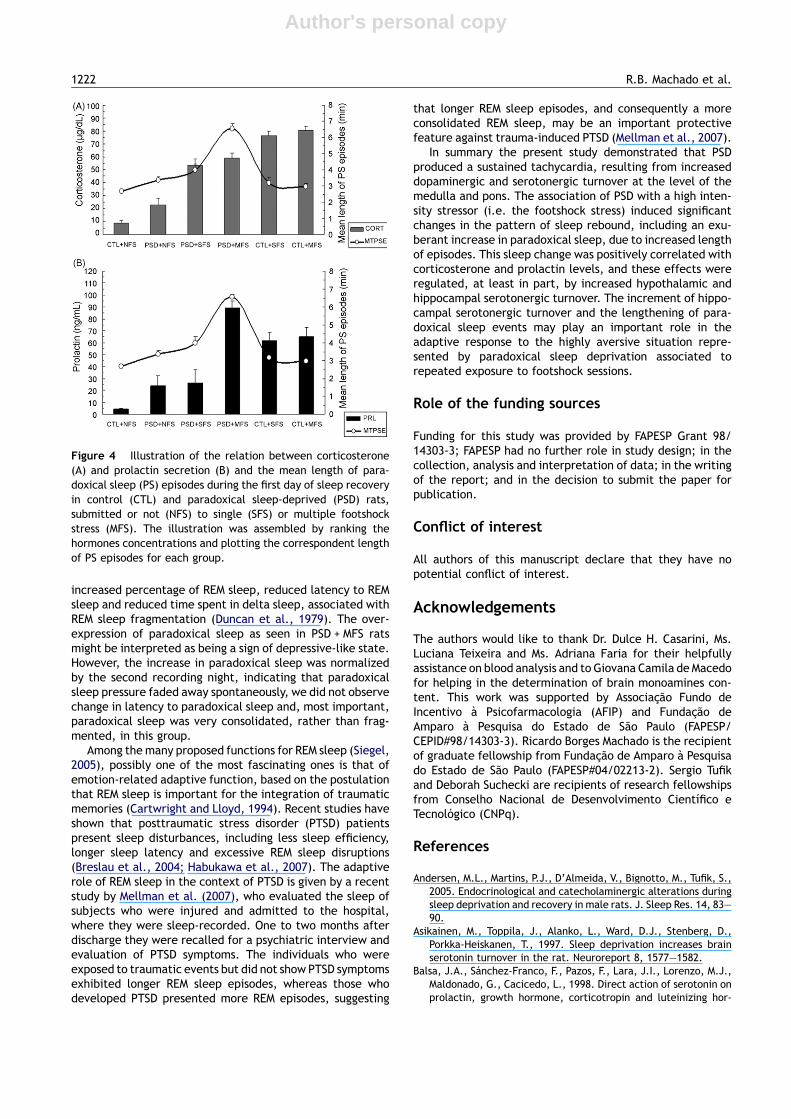
REM sleep is successfully corrected by administering hydro-cortisone before bed time (Garcıa-Borreguero et al., 2000).In the present study, corticosterone plasma levels in PS-deprived rats were positively correlated with the length ofPS episodes, indicating that corticosterone participates inthe expression of PS rebound. The fact that in non sleep-deprived rats either low or high corticosterone concentra-tions did not influence the length of paradoxical sleep epi-sodes, suggest that this hormone is involved in the sleephomeostatic process in animals with high propensity forparadoxical sleep (for instance, paradoxical sleep-deprivedrats). Recent data from our laboratory showed that corticos-terone supplementation during sleep deprivation resulted inincreased length of PS episodes, corroborating the role of thishormone in the expression of PS in sleep-deprived rats(unpublished data).
Nonetheless, prolactin was the hormone more stronglycorrelated with PS rebound, with a highly significant correla-tion with the length of PS episodes. Previous studies from ourlaboratory had shown that 96 h of PSD performed by themodified multiple platform method result in elevated levelsof prolactin (Andersen et al., 2005). PSD alone induced higherprolactin levels than those seen in the CTL group, whereaschronic stress in PS-deprived rats led to a 3.7-fold increasecompared to the effects of acute stress. In human beings,prolactin is mainly secreted during the second half of thenight (Sassin et al., 1972), with a very clear circadian rhythm(Van Cauter, 1990). Experimental studies demonstrate thatprolactin is, directly or indirectly, involved in the regulationof paradoxical sleep (Obal et al., 1989, 1994; Roky et al.,1993). Thus, systemic administration of prolactin induces aclear increase of PS in rodents, mainly during the light phase(Roky et al., 1995), whilst anti-prolactin suppresses PS in rats(Obal et al., 1997); prolactin deficient mice exhibit less PS(Obal et al., 2005) and ether stress increases both prolactinand paradoxical sleep in rats (Bodosi et al., 2000). It isimportant to mention that in the present study, the strongassociation between PRL and PS was true only for PS-deprivedrats, since in response to footshock, CTL rats displayed amajor increase in PRL levels, but no changes in PS.
A surprising finding in the present study was the lack ofnegative correlation between hypothalamic dopamine con-centration and prolactin levels. In general, a main effect ofstress on hypothalamic dopamine levels was observed, i.e.,both forms of footshock increase hypothalamic dopaminelevels, but this effect was not reflected on prolactin secre-tion. Classically, dopamine is considered to be the majorinhibitor of prolactin release, acting via D2 receptors locatedin pituitary lactotrophs, whereas the activity of these dopa-minergic neurons at the lactotroph is inhibited by D1 dopa-minergic receptors (Freeman et al., 2000). In a previous studyfrom our laboratory, 96 h of PSD did not result in changes in D1
or D2 receptors density in the hypothalamus, except for thesuprachiasmatic nucleus. Unfortunately, dopaminergicreceptors in the pituitary were not assessed, making difficultto draw any conclusion (Nunes et al., 1994). It is important tobare in mind, however, that 5-HT is the major stimulator ofPRL secretion; this action seems to be modulated by vasoac-tive intestinal peptide — VIP (Balsa et al., 1998). Interest-ingly, PSD causes an accumulation of VIP levels in thecerebrospinal fluid (Jimenez-Anguiano et al., 1993) and anupregulation of VIP receptors in several brain regions, some
of which are closely related to PS (Jimenez-Anguiano et al.,1996). Nevertheless, it is important to mention that withinthe constraints of the present protocol HPLC was performedin whole hypothalamus, thus providing a general idea of themonoamine concentrations in this brain region. It has beenreported in a recent study, that in female rats, the prolactinsurge that occurs during estrous is mediated by reduceddopamine concentrations and increased dopamine turnoverin the mediobasal hypothalamus and the medial preopticarea (Szawka et al., 2007). Therefore, had we performedmicrodissections of the hyptothalamus or microdyalisis, wemight have been able to estimate more accurately howparadoxical sleep deprivation and footshock stress alteredthe role of each biogenic amine in the hypothalamic regula-tion of prolactin secretion.
From the results of the present study, it appears as thoughthe most likely mediating process for PSD + MFS-induced suchrobust lengthening of PS episodes involves optimum corti-costerone associated with maximum prolactin plasma levels,as is illustrated in Fig. 4. Corroborating this idea, a study bySanford and co-workers report that mice displaying differentlevels of anxiety respond differently to footshock, i.e., themore anxious-type strain BALB/cJ mice exhibit intense PSinhibition following footshock and re-exposure to the stresschamber than the less anxious-type C57BL/C6 (Sanford et al.,2003b). Interestingly, it is also the less anxious-type strainwhich secretes more prolactin and exhibit more PS inresponse to a 1 h restraint stress (Meerlo et al., 2001b),despite a similar corticosterone response of both strains,suggesting that PRL plays a major role as a mediator ofstress-induced PS rebound.
Serotonin is involved with the stress response, sleep andbehavioral adaptation to stress (Linthorst and Reul, 2008).Neurons in the dorsal raphe nucleus fire in a behavioral state-dependent manner and hippocampal mycrodialysis studieshave shown that maximum 5-HT release occurs during wake,followed by a reduction during slow wave sleep and a furtherreduction during paradoxical sleep (Park et al., 1999; Penalvaet al., 2003). Short-term sleep deprivation increases 5-HTlevels and turnover in the hippocampus, which is rapidlyreversed by sleep rebound (Asikainen et al., 1997; Penalvaet al., 2003), whereas long-term (96 h) paradoxical sleepdeprivation increases 5-HIAA levels and serotonergic turn-over in the hypothalamus and hippocampus (Youngbloodet al., 1997). In the present study we observed the samechanges as Youngblood et al.’s (1997), indicating higheractivity of this system in PS-deprived than in control rats.These effects on serotonergic activity may be explained bythe down-regulation of 5-HT1A receptors in the dorsal raphenucleus (Evrard et al., 2006; Gardner et al., 1997) andsubsensitivity to systemic administration of 8-OH-DPAT(Roman et al., 2005; Roman et al., 2006) produced by short-or long-term sleep deprivation.
That dysfunctions of the hippocampal serotonergic systemare involved with depression is corroborated by reportsof reduced concentrations of 5-HIAA in a subpopulation ofdepressed patients and indications of altered expressionof serotonergic receptors in post-mortem tissue studies ofdepressed patients (Maes and Meltzer, 1995). An activeserotonergic system, therefore, is believed to underlie adap-tive responses to chronic stress. One of the hallmarks ofdepression is the change in sleep architecture that includes
Chronic stress during paradoxical sleep deprivation increases rebound of paradoxical sleep 1221
Author's personal copy
increased percentage of REM sleep, reduced latency to REMsleep and reduced time spent in delta sleep, associated withREM sleep fragmentation (Duncan et al., 1979). The over-expression of paradoxical sleep as seen in PSD + MFS ratsmight be interpreted as being a sign of depressive-like state.However, the increase in paradoxical sleep was normalizedby the second recording night, indicating that paradoxicalsleep pressure faded away spontaneously, we did not observechange in latency to paradoxical sleep and, most important,paradoxical sleep was very consolidated, rather than frag-mented, in this group.
Among the many proposed functions for REM sleep (Siegel,2005), possibly one of the most fascinating ones is that ofemotion-related adaptive function, based on the postulationthat REM sleep is important for the integration of traumaticmemories (Cartwright and Lloyd, 1994). Recent studies haveshown that posttraumatic stress disorder (PTSD) patientspresent sleep disturbances, including less sleep efficiency,longer sleep latency and excessive REM sleep disruptions(Breslau et al., 2004; Habukawa et al., 2007). The adaptiverole of REM sleep in the context of PTSD is given by a recentstudy by Mellman et al. (2007), who evaluated the sleep ofsubjects who were injured and admitted to the hospital,where they were sleep-recorded. One to two months afterdischarge they were recalled for a psychiatric interview andevaluation of PTSD symptoms. The individuals who wereexposed to traumatic events but did not show PTSD symptomsexhibited longer REM sleep episodes, whereas those whodeveloped PTSD presented more REM episodes, suggesting
that longer REM sleep episodes, and consequently a moreconsolidated REM sleep, may be an important protectivefeature against trauma-induced PTSD (Mellman et al., 2007).
In summary the present study demonstrated that PSDproduced a sustained tachycardia, resulting from increaseddopaminergic and serotonergic turnover at the level of themedulla and pons. The association of PSD with a high inten-sity stressor (i.e. the footshock stress) induced significantchanges in the pattern of sleep rebound, including an exu-berant increase in paradoxical sleep, due to increased lengthof episodes. This sleep change was positively correlated withcorticosterone and prolactin levels, and these effects wereregulated, at least in part, by increased hypothalamic andhippocampal serotonergic turnover. The increment of hippo-campal serotonergic turnover and the lengthening of para-doxical sleep events may play an important role in theadaptive response to the highly aversive situation repre-sented by paradoxical sleep deprivation associated torepeated exposure to footshock sessions.
Role of the funding sources
Funding for this study was provided by FAPESP Grant 98/14303-3; FAPESP had no further role in study design; in thecollection, analysis and interpretation of data; in the writingof the report; and in the decision to submit the paper forpublication.
Conflict of interest
All authors of this manuscript declare that they have nopotential conflict of interest.
Acknowledgements
The authors would like to thank Dr. Dulce H. Casarini, Ms.Luciana Teixeira and Ms. Adriana Faria for their helpfullyassistance on blood analysis and to Giovana Camila deMacedofor helping in the determination of brain monoamines con-tent. This work was supported by Associacao Fundo deIncentivo a Psicofarmacologia (AFIP) and Fundacao deAmparo a Pesquisa do Estado de Sao Paulo (FAPESP/CEPID#98/14303-3). Ricardo Borges Machado is the recipientof graduate fellowship from Fundacao de Amparo a Pesquisado Estado de Sao Paulo (FAPESP#04/02213-2). Sergio Tufikand Deborah Suchecki are recipients of research fellowshipsfrom Conselho Nacional de Desenvolvimento Cientıfico eTecnologico (CNPq).
References
Andersen, M.L., Martins, P.J., D’Almeida, V., Bignotto, M., Tufik, S.,2005. Endocrinological and catecholaminergic alterations duringsleep deprivation and recovery in male rats. J. Sleep Res. 14, 83—90.
Asikainen, M., Toppila, J., Alanko, L., Ward, D.J., Stenberg, D.,Porkka-Heiskanen, T., 1997. Sleep deprivation increases brainserotonin turnover in the rat. Neuroreport 8, 1577—1582.
Balsa, J.A., Sanchez-Franco, F., Pazos, F., Lara, J.I., Lorenzo, M.J.,Maldonado, G., Cacicedo, L., 1998. Direct action of serotonin onprolactin, growth hormone, corticotropin and luteinizing hor-
Figure 4 Illustration of the relation between corticosterone(A) and prolactin secretion (B) and the mean length of para-doxical sleep (PS) episodes during the first day of sleep recoveryin control (CTL) and paradoxical sleep-deprived (PSD) rats,submitted or not (NFS) to single (SFS) or multiple footshockstress (MFS). The illustration was assembled by ranking thehormones concentrations and plotting the correspondent lengthof PS episodes for each group.
1222 R.B. Machado et al.

Author's personal copy
mone release in cocultures of anterior and posterior pituitarylobes: autocrine and/or paracrine action of vasoactive intestinalpeptide. Neuroendocrinology 68, 326—333.
Bodosi, B., Obal, F., Gardi, J., Komlodi, J., Fang, J., Krueger, J.M.,2000. An ether stressor increases REM sleep in rats: possible roleof prolactin. Am. J. Physiol. 279, R1590—R1598.
Breslau, N., Roth, T., Burduvali, E., Kapke, A., Schultz, L., Roehrs, T.,2004. Sleep in lifetime posttraumatic stress disorder: a commu-nity-based polysomnographic study. Arch. Gen. Psychiatry 61,508—511.
Cartwright, R.D., Lloyd, S.R., 1994. Early REM sleep: a compensatorychange in depression? Psychiatry Res. 51, 245—252.
Cespuglio, R., Baubet, V., Font, C., Marinesco, S., El Kafi, B., Jouvet,M., 1994. Possible involvement of the arcuate nucleus in the sleeprebound induced by an immobilization stress. J. Sleep Res. 3(Suppl.), 41.
Chastrette, N., Cespuglio, R., Jouvet, M., 1990. ProopiomelanocortinPOMC-derived peptides and sleep in the rat. Part I — Hypnogenicproperties of ACTH derivatives. Neuropeptides 15, 61—74.
Coenen, A.M.L., Van Luijtelaar, E.L.M., 1985. Stress Induced by threeprocedures of deprivation of paradoxical sleep. Physiol. Behav.35, 501—504.
De Mesquita, S., Hale, G.A., 1992. Cardiopulmonary regulation afterrapid-eye-movement sleep deprivation. J. Appl. Physiol. 72,970—976.
Dewasmes, G., Loos, N., Delanaud, S., Dewasmes, D., Ramadan, W.,2004. Pattern of rapid-eye movement sleep episode occurrenceafter an immobilization stress in the rat. Neurosci. Lett. 355,17—20.
Duncan Jr., W.C., Pettigrew, K.D., Gillin, J.C., 1979. REM architec-ture changes in bipolar and unipolar depression. Am. J. Psychiatry136, 1424—1427.
Evrard, A., Barden, N., Hamon, M., Adrien, J., 2006. Glucocorticoidreceptor-dependent desensitization of 5-HT1A autoreceptors bysleepdeprivation: studies inGR-i transgenicmice.Sleep29,31—36.
Fadda, P., Fratta, W., 1997. Stress-induced sleep deprivation modi-fies corticotropin releasing factor (CRF) levels and CRF binding inrat brain and pituitary. Pharmacol. Res. 35, 436—443.
Freeman, M.E., Kanyicska, B., Lerant, A., Nagy, G., 2000. Prolactin:structure, function, and regulation of secretion. Physiol. Rev. 80,1523—1631.
Garcıa-Borreguero, D., Wehr, T.A., Larrosa, O., Granizo, J.J., Hard-wick, D., Chrousos, G.P., Friedman, T.C., 2000. Glucocorticoidreplacement is permissive for rapid eye movement sleep andsleep consolidation in patients with adrenal insufficiency. J. Clin.Endocrinol. Metab. 85, 4201—4206.
Gardner, J.P., Fornal, C.A., Jacobs, B.L., 1997. Effects of sleepdeprivation on serotonergic neuronal activity in the dorsal raphenucleus of the freely moving cat. Neuropsychopharmacology 17,72—81.
Granata, A.R., Woodruff, G.N., 1982. Dopaminergic mechanisms inthe nucleus tractus solitarius and effects on blood pressure. BrainRes. Bull. 8, 483—488.
Habukawa, M., Uchimura, N., Maeda, M., Kotorii, N., Maeda, H.,2007. Sleep findings in young adult patients with posttraumaticstress disorder. Biol. Psychiatry 62, 1179—1182.
Jimenez-Anguiano, A., Baez-Saldana, A., Drucker-Colın, R., 1993.Cerebrospinal fluid (CSF) extracted immediately after REM sleepdeprivation prevents REM rebound and contains vasoactive intest-inal peptide (VIP). Brain Res. 631, 345—348.
Jimenez-Anguiano, A., Garcıa-Garcıa, F., Mendoza-Ramırez, J.L.,Duran-Vazquez, A., Drucker-Colın, R., 1996. Brain distributionof vasoactive intestinal peptide receptors following REM sleepdeprivation. Brain Res. 728, 37—46.
Jouvet, M., 1994. Paradoxical sleep mechanisms. Sleep 17 (8 Suppl.),S77—S83.
Karlsson, G.A., Preuss, C.V., Chaitoff, K.A., Maher, T.J., Ally, A.,2006. Medullary monoamines and NMDA-receptor regulation of
cardiovascular responses during peripheral nociceptive stimuli.Neurosci. Res. 55, 316—326.
Koehl, M., Bouyer, J.J., Darnaudery, M., Le Moal, M., Mayo, W., 2002.The effect of restraint stress on paradoxical sleep is influenced bythe circadian cycle. Brain Res. 937, 45—50.
Kovalzon, V.M., Tsibulsky, V.L., 1984. REM-sleep deprivation, stressand emotional behavior in rats. Behav. Brain Res. 14, 235—245.
Leger, L., Lema, F., Chastrette, N., Charnay, Y., Cespuglio, R., Mazie,J.C., Jouvet, M., 1990. A monoclonal antibody directed againstCLIP (ACTH18-39) anatomical distribution of immunoreactivity inthe rat brain and hypophysis with quantification of the hypotha-lamic cell group. J. Chem. Neuroanat. 3, 297—308.
Linthorst, A.C., Reul, J.M., 2008. Stress and the brain: solvingthe puzzle using microdialysis. Pharmacol. Biochem. Behav. 90,163—173.
Lovick, T.A., 1989. Cardiovascular responses to 5-HT in the ventro-lateral medulla of the rat. J. Auton. Nerv. Syst. 28, 35—41.
Lupien, S.J., Maheu, F., Tu, M., Fiocco, A., Schramek, T.E., 2007.The effects of stress and stress hormones on human cognition:implications for the field of brain and cognition. Brain Cogn. 65,209—237.
Machado, R.B., Hipolide, D.C., Benedito-Silva, A.A., Tufik, S., 2004.Sleep deprivation induced by modified multiple platform techni-que: quantification of sleep loss and recovery. Brain Res. 1004,45—51.
Machado, R.B., Suchecki, D., Tufik, S., 2005. Sleep homeostasis inrats assessed by a long-term intermittent paradoxical sleepdeprivation protocol. Behav. Brain Res. 160, 356—364.
Maes, M., Meltzer, H.Y., 1995. The serotonin hypothesis of depres-sion. In: Bloom, F.E., Kupfer, D.J. (Eds.), Psychopharmacology:The Fourth Generation of Progress, pp. 933—944.
Mandal, A.K., Zhong, P.Y., Kellar, K.J., Gillis, R.A., 1990. Ventrolat-eral medulla: an important site of action for the hypotensiveeffect of drugs that activate serotonin-1A receptors. J. Cardio-vasc. Pharmacol. 7, S49—60.
Marinesco, S., Bonnet, C., Cespuglio, R., 1999. Influence of stressduration on the sleep rebound induced by immobilization in therat: a possible role for corticosterone. Neuroscience 92, 921—933.
Meerlo, P., de Bruin, E.A., Strijkstra, A.M., Daan, S., 2001a. A socialconflict increases EEG slow-wave activity during subsequentsleep. Physiol. Behav. 73, 331—335.
Meerlo, P., Easton, A., Bergmann, B.M., Turek, F.W., 2001b. Restraintincreases prolactin and REM sleep in C57BL/6J mice but not inBALB/cJ mice. Am. J. Physiol. 281, R846—R854.
Meerlo, P., Pragt, B.J., Daan, S., 1997. Social stress induces highintensity sleep in rats. Neurosci. Lett. 225, 41—44.
Mellman, T.A., Pigeon, W.R., Nowell, P.D., Nolan, B., 2007. Relation-ships between REM sleep findings and PTSD symptoms during theearly aftermath of trauma. J. Trauma. Stress 20, 893—901.
Nunes Jr., G.P., Tufik, S., Nobrega, J.N., 1994. Autoradiographicanalysis of D1 and D2 dopaminergic receptors in rat brain afterparadoxical sleep deprivation. Brain Res. Bull. 34, 453—456.
Obal Jr., F., Bodosi, B., Szilagyi, A., Kacsoh, B., Krueger, J.M., 1997.Antiserum to growth hormone decreases sleep in the rat. Neu-roendocrinology 66, 9—16.
Obal Jr., F., Garcia-Garcia, F., Kacsoh, B., Taishi, P., Bohnet, S., Horse-man, N.D., Krueger, J.M., 2005. Rapid eye movement sleep isreduced inprolactin-deficientmice.J.Neurosci. 25, 10282—10289.
Obal Jr., F., Opp, M.R., Cady, A.B., Johannsen, L., Krueger, J.M.,1989. Prolactin, vasoactive intestinal peptide, and peptide his-tidine methionine elicit selective increases in REM sleep in rab-bits. Brain Res. 490, 292—300.
Obal Jr., F., Payne, L., Opp,M.R., Grosvenor, C.E., J.M., 1994. Involve-ment of prolactin in the REM sleep-promoting activity of systemicvasoactive intestinal peptide. Brain Res. 645, 143—149.
Palma, B.D., Suchecki, D., Tufik, S., 2000. Differential effectsof acute cold and footshock on the sleep of rats. Brain Res.861, 97—104.
Chronic stress during paradoxical sleep deprivation increases rebound of paradoxical sleep 1223

Author's personal copy
Park, S.P., Lopez-Rodriguez, F., Wilson, C.L., Maidment, N., Matsu-moto, Y., Engel Jr., J., 1999. In vivo microdialysis measures ofextracellular serotonin in the rat hippocampus during sleep-wakefulness. Brain Res. 833, 291—296.
Pawlyk, A.C., Jha, S.K., Brennan, F.X., Morrison, A.R., Ross, R.J.,2005. A rodent model of sleep disturbances in posttraumaticstress disorder: the role of context after fear conditioning. Biol.Psychiatry 57, 268—277.
Penalva, R.G., Lancel, M., Flachskamm, C., Reul, J.M., Holsboer, F.,Linthorst, A.C., 2003. Effect of sleep and sleep deprivation onserotonergic neurotransmission in the hippocampus: a combinedin vivo microdialysis/EEG study in rats. Eur. J. Neurosci. 17,1896—1906.
Rampin, C., Cespuglio, R., Chastrette, N., Jouvet, M., 1991. Immo-bilization stress induces a paradoxical sleep rebound in rat.Neurosci. Lett. 126, 113—118.
Rechtschaffen, A., Bergmann, B.M., Everson, C.A., Kushida, C.A.,Gilliland, M., 1989. Sleep deprivation in the rat: X Integration anddiscussion of the findings. Sleep 12, 68—87.
Roky, R., Obal Jr., F., Valatx, J.L., Bredow, S., Fang, J., Pagano, L.P.,Krueger, J.M., 1995. Prolactin and rapid eye movement sleepregulation. J. Neurophysiol. 18, 536—542.
Roky, R., Valatx, J.L., Jouvet, M., 1993. Effect of prolactin on thesleep-wake cycle in the rat. Neurosci. Lett. 156, 117—120.
Roman, V., Hagewoud, R., Luiten, P.G., Meerlo, P., 2006. Differentialeffects of chronic partial sleep deprivation and stress on seroto-nin-1A and muscarinic acetylcholine receptor sensitivity. J. SleepRes. 15, 386—394.
Roman, V., Walstra, I., Luiten, P.G., Meerlo, P., 2005. Too little sleepgradually desensitizes the serotonin 1A receptor system. Sleep28, 1505—1510.
Rosenberg, R.S., Bergmann, B.M., Rechtschaffen, A., 1976. Varia-tions in slow wave activity during sleep in the rat. Physiol. Behav.17, 931—938.
Sanford, L.D., Fang, J., Tang, X., 2003a. Sleep after differingamounts of conditioned fear training in BALB/cJ mice. Behav.Brain Res. 147, 193—202.
Sanford, L.D., Tang, X., Ross, R.J., Morrison, A.R., 2003b. Influence ofshock training and explicit fear-conditioned cues on sleep archi-tecture in mice: strain comparison. Behav. Genet. 33, 43—58.
Sapolsky, R.M., Romero, M.L., Muck, A., 2000. How do glucocorticoidsinfluence stress responses? Integrating permissive, suppressive,stimulatory, and preparative actions. Endocr. Rev. 21, 55—89.
Sassin, J.F., Frantz, A.G., Weitzman, E.D., Kapen, S., 1972. Humanprolactin: 24-hour pattern with increased release during sleep.Science 177, 1205—1207.
Sgoifo, A., Buwalda, B., Roos, M., Costoli, T., Merati, G., Meerlo, P.,2006. Effects of sleep deprivation on cardiac autonomic andpituitary-adrenocortical stress reactivity in rats. Psychoneuroen-docrinology 31, 197—208.
Siegel, J.M., 2005. Clues to the functions of mammalian sleep.Nature 437, 1264—1271.
Suchecki, D., Tiba, P.A., Tufik, S., 2002a. Paradoxical sleep depriva-tion facilitates subsequent corticosterone response to a mildstressor in rats. Neurosci. Lett. 320, 45—48.
Suchecki, D., Tiba, P.A., Tufik, S., 2002b. Hormonal and behaviouralresponses of paradoxical sleep-deprived rats to the elevated plusmaze. J. Neuroendocrinol. 14, 549—554.
Szawka, R.E., Rodovalho, G.V., Helena, C.V., Franci, C.R., Anselmo-Franci, J.A., 2007. Prolactin secretory surge during estrus coin-cides with increased dopamine activity in the hypothalamus andpreoptic area and is not altered by ovariectomy on proestrus.Brain Res. Bull. 73, 127—134.
Tiba, P.A., Palma, B.D., Tufik, S., Suchecki, D., 2003. Effects of earlyhandling on basal and stress-induced sleep parameters in rats.Brain Res. 975, 158—166.
Tiba, P.A., Tufik, S., Suchecki, D., 2004. Effects of maternal separa-tion on baseline sleep and cold stress-induced sleep rebound inadult Wistar rats. Sleep 27, 1146—1153.
Timo-Iaria, C., Negrao, N., Schimidek, R.W., Hoshino, K., Menezes,C.E.L., Da Rocha, T.L., 1970. Phases and states of sleep in the rat.Physiol. Behav. 5, 1057—1062.
Van Cauter, E., 1990. Diurnal and ultradian rhythms in human endo-crine function: a minireview. Horm. Res. 34, 45—53.
Vazquez-Palacios, G., Velazquez-Moctezuma, J., 2000. Effect ofelectric foot shocks immobilization and corticosterone adminis-tration on the sleep-wake pattern in the rat. Physiol. Behav. 71,23—28.
Wetzel, W., Wagner, T., Vogel, D., Demuth, H.U., Balschun, D., 1997.Effects of the CLIP fragment ACTH20-24 on the duration of REMsleep episodes. Neuropeptides 31, 41—45.
Yang, R.H., Igarashi, Y., Wyss, J.M., Chen, Y.F., 1990. Dopamine D2receptors in the posterior region of the nucleus tractus solitariusmediate the central pressor action of quinpirole (LY171555). BrainRes. Bull. 24, 97—103.
Youngblood, B.D., Zhou, J., Smagin, G.N., Ryan, D.H.,Harris, R.B., 1997. Sleep deprivation by the ‘‘flower pot’’technique and spatial reference memory. Physiol. Behav. 61,249—256.
Zepelin, H., 1994. Mammalian sleep. In: Kryger, M.H., Roth, T., De-ment, W.C. (Eds.), Principles and Practice of Sleep Medicine. WBSaunders, Philadelphia, pp. 69—80.
1224 R.B. Machado et al.