bba rebound paper 2014-1
TRANSCRIPT

Biological Conservation 169 (2014) 319–333
Contents lists available at ScienceDirect
Biological Conservation
journal homepage: www.elsevier .com/ locate /biocon
Black-browed albatross numbers in Chile increase in responseto reduced mortality in fisheries
0006-3207/$ - see front matter � 2013 Elsevier Ltd. All rights reserved.http://dx.doi.org/10.1016/j.biocon.2013.12.002
⇑ Corresponding author at: Australian Antarctic Division, 203 Channel Highway,Kingston, Tasmania 7050, Australia. Tel.: +61 3 62 323 337.
E-mail address: [email protected] (G. Robertson).1 Current address. Skadia Pty Ltd, GIP Business Centre, Horsham, Victoria 3409,
Australia.2 Current address. Department of Animal Ecology and Systematics, Justus Liebig
University Giessen, Heinrich-Buff-Ring 38, 35392 Giessen, Germany.
Graham Robertson a,⇑, Carlos Moreno b, Javier A. Arata c, Steven G. Candy a, Kieran Lawton a,e,1,Jose Valencia c, Barbara Wienecke a, Roger Kirkwood a, Phil Taylor d, Cristián G. Suazo b,f,2
a Australian Antarctic Division, 203 Channel Highway, Kingston, Tasmania 7050, Australiab Instituto de Ciencias Ambientales y Evolutivas, Facultad de Ciencias Campus Isla Teja, Valdivia, Chilec Instituto Antarctico Chileno, Plaza Benjamin Muñoz Gamero 1055, Punta Arenas, Chiled Birdlife International, Wellbrook Court, Girton Road, Cambridge CB3 0NA, UKe Skadia Pty Ltd, GIP Business Centre, Horsham, Victoria 3409, Australiaf Department of Animal Ecology and Systematics, Justus Liebig University Giessen, Heinrich-Buff-Ring 38, 35392 Giessen, Germany
a r t i c l e i n f o
Article history:Received 28 August 2013Received in revised form 29 November 2013Accepted 1 December 2013
Keywords:Albatross population trendsLongline fisheriesTrawl fisheriesSeabird bycatchInnovationCo-operative research
a b s t r a c t
Black-browed albatrosses (Thalassarche melanophrys) are a common victim of incidental mortality incommercial fishing operations. Chile holds globally important populations of black-browed albatrossesand grey-headed (Thalassarche chrysostoma) albatrosses with about 85% of the former species breedingat the Diego Ramirez and Ildefonso Archipelagos. In the nine years between 2002 and 2011 the numberof black-browed albatrosses at these sites increased by 52% and 18%, respectively, or 23% for both sitescombined. In the same period the population size of grey-headed albatrosses showed no signs of change.To explain the reason for the increases we examined trends in fishing effort for the five main longline andtrawl fisheries with a history of interactions with seabirds; the extent of overlap between the fisheriesand albatross foraging ranges; and albatross bycatch rates for each fishery. The circumstantial evidenceand evidence from the SEABIRD population model suggest the most likely reason for the populationincrease was reduced mortality in the industrial longline fishery for Patagonian toothfish. In 2002 Span-ish system vessels in this fishery took an estimated 1555 black-browed albatrosses. In 2006 and 2007 thefleet converted to a new method of fishing, called the Chilean system (or trotline-with-nets), whichreduced seabird mortality to zero. Developed to minimise depredation of toothfish by sperm whalesand killer whales, the re-designed gear configuration resulted in a fivefold increase in mean sink ratesof baited hooks (from 0.15 m/s to 0.80 m/s), making them inaccessible to seabirds. The Chilean systemis an example of innovation yielding both economic benefits to the fishing industry and conservationbenefits to seabirds. The ability of black-browed albatrosses to respond to reduced mortality levelsencourages continued efforts to implement seabird-friendly gears and practices in other fisheries withdetrimental impacts on seabirds.
� 2013 Elsevier Ltd. All rights reserved.
1. Introduction
Many populations of albatrosses and petrels face an uncertainfuture due to high levels of mortality associated with commercialfishing operations. The main commercial fishing methods of con-cern are pelagic and demersal longlining (see Robertson and Gales,1997 and references therein; Delord et al., 2005; Anderson et al.,
2011) and trawling (González-Zevallos and Yorio, 2006; Sullivanet al., 2006a). Incidental mortality in these fisheries is consideredthe principal reason most of the world’s 22 species of albatrossesare threatened with extinction (IUCN Red List of ThreatenedSpecies, 2012). The detrimental impact of fisheries on seabirdshas attracted considerable international attention, most notablyfrom the UNs Food and Agriculture Organisation (FAO, 1999), theCommission for the Conservation of Antarctic Marine LivingResources (CCAMLR), various regional fisheries managementorganisations (Small, 2005) and fisheries in the national economiczones of many coastal States. The Agreement on the Conservationof Albatrosses and Petrels (ACAP; http://www.acap.aq) is the mostrecent initiative of significance to be established in recognition ofthe pervasive impacts of fisheries (and other agents of mortality)

320 G. Robertson et al. / Biological Conservation 169 (2014) 319–333
on migratory albatrosses and petrels. The Agreement currently has13 signatories (nations) which are required to conduct research toimprove understanding of the conservation status of species listedunder the Agreement and implement measures to reduce impactson populations. Currently the Agreement applies to 30 species ofalbatrosses, petrels and shearwaters, the majority of which arethreatened by mortality in commercial fishing operations.
The black-browed albatross (Thalassarche melanophrys) is themost abundant albatross species in the southern hemisphere(Tickell, 2000) and is a common victim of commercial fishingoperations. Black-browed albatrosses are taken incidentally inpelagic longline fisheries (Jiménez et al., 2010; Rolland et al.,2009; González et al., 2012; Yeh et al., 2012), demersal longlinefisheries (Moreno et al., 1996; Otley, 2005; Favero et al., 2013)and trawl fisheries for various pelagic and deep water species(SC-CCAMLR, 2006; Sullivan et al., 2006a). Nearly all of the world’sblack-browed albatrosses reside at the Falkland Islands/IslasMalvinas, South Georgia and the islands of southern Chile. Populationdecreases were detected at South Georgia in the 1970s (Croxallet al., 1997) and have continued ever since (Poncet et al., 2006;R. Phillips, personal communication). Populations at the Falklands/Malvinas Island decreased in the 1980s and 1990s but haverecently shown signs of recovery (Catry et al., 2011; Wolfaardt,2013). Detection of trends with the Chilean populations is lessstraightforward than at the other two sites because systematiccensuses in the region only commenced in 2001 and 2002 whenthree of the six known breeding sites were unknown to science.Nonetheless, the early surveys included the three most populoussites in the region – Isla Diego de Almagro (surveyed in 2001), IslasDiego Ramirez (2002), Islas Ildefonso (2002) – which together hold�95% of total numbers (ACAP, 2010). The latter two sites werere-surveyed in 2011 to determine how the albatrosses had faredin the nine years since the previous censuses. The surveys revealeda substantial increase in numbers. This increase was unexpected
Diego RamArchipela
Islote Evangel
80W
60S
Isla Diegode Almag
I
Is. Cincloid
1 km
Is. Boina
Cabo Gruta
Pta Falsa Lucia
Colony ‘Tooluka’
Pta Filon Santa Lucia
Roca Puntiaguda
Isla Diego de Almagro
10 km
Is. NorteIs. Schlatter
Is. Martinez
Is. Mendoza
Is. Santander
Is. Bartolome
Is. EsterIs. Gonzalo
1 km
Northern
Group
Diego RamirezArchipelago
50S
Islote L
Fig. 1. Locations of the six known albatross breeding sites in Chile in relation to the ChAlmagro shows the six albatross colonies from the census in 2001 (Lawton et al., 2003). Tthe archipelagos, their proximity to one another and relative sizes.
and prompted a second survey in 2012, the objective being toverify the accuracy of the results in 2011 and determine if changesrecorded in the two south-most sites (Diego Ramirez andIldefonso) were evident at the four other breeding sites in Chile.
In this paper we report the results of trends in black-browedand grey-headed (T. chrysostoma) albatross numbers between theinitial surveys in 2001/02 and those in 2011 and 2012. To explainthe likely reasons for observed population increases we examined:(i) albatross foraging distribution (from tracking studies) in fivebreeding seasons from 1997 to 2011; (ii) the distribution and effortfrom all longline and trawl fisheries in Chile operating within alba-tross breeding season foraging ranges; (iii) albatross bycatch esti-mates for each fishery in which seabird mortality has beenrecorded; and (iv) trends in bycatch associated with recentchanges to fishing gear (see below) in the industrial longline fish-ery for Patagonian toothfish (Dissostichus eleginoides). Trends inchlorophyll-a density (a proxy indicator of marine productivity)in the black-browed albatross foraging range was also examinedto determine if variation in marine productivity was implicatedin the population increases. Finally, we used the fishing effort dataand albatross bycatch data in combination with albatross life his-tory parameters from the published literature to construct a modelto determine if reduced bycatch was the most likely reason for theobserved increases in albatross numbers.
2. Methods
2.1. Breeding sites and population estimates
In Chile black-browed albatrosses breed at six known sites: IslasDiego Ramirez (56�310S, 68�430W); Islas Ildefonso (55�480S,69�240W); Islote Albatros (54�270S, 69�010W); Islote Leonard(53�230S; 74�040W), Islote Evangelistas (52�240S, 75�050W) and IslaDiego de Almagro (51�320S, 75�200W) (Fig. 1). Except for a few pairs
irezgo
Ildefonso Archipelago
Islote Albatros
istas
Falkland Islands/
Islas Malvinas
SouthAmerica
Antarctic
Peninsula
50W60W70W
Drake Passage
ro
s. Norte
Is. SquareIs. Spirit
Is. Sur
Is. Grande
es
Ildefonso Archipelago
.eonard
Northern
Group
ilean mainland, Drake Passage and Antarctic Peninsula. The map of Islas Diego dehe maps of Diego Ramirez and Ildefonso show the northern and southern groups in

0
10
20
30
40
50
60
70
toothfish –
1986
1988
1990
1992
1994
1996
1998
2000
2002
2004
2006
2008
2010
2012
(millions)
Year
Patagonian toothfish – m
congrio –
Fig. 2. Trend in fishing effort for the industrial Austral hake and congrio longlinefishery, and the Patagonian toothfish longline fishery. Effort for the Patagoniantoothfish fishery is divided into hook numbers set by the traditional Spanish systemand the Chilean system. Data sources: the Institute for Fishery Research (Australhake and congrio) and the Chilean Sub-Secretary of Fisheries (Patagonian toothfish).
G. Robertson et al. / Biological Conservation 169 (2014) 319–333 321
at Ildefonso (Robertson et al., 2007), the grey-headed albatrosspopulation is confined to the Diego Ramirez archipelago.
Populations at Diego de Almagro were surveyed in October 2001(Lawton et al., 2003) and those at Ildefonso (Robertson et al., 2008a)and Diego Ramirez (Robertson et al., 2007) in October 2002. Thecolonies at Islote Evangelistas were surveyed for the first time inOctober 2002 (Arata et al., 2003). The tiny colony at Islote Albatros,which lies deep inside the Chilean channels (in Seno Almirantazgo),was discovered and surveyed in January 2003 during chick rearing(Aguayo et al., 2003). Finally, the small colony at Islote Leonard wasdiscovered and surveyed in November 2005 (Marin and Oehler,2007). The first survey during the incubation period at IsloteAlbatros was conducted in October 2006 (Moreno and Robertson,2008) and repeated in October 2011. Partial surveys (selectedislands only, for comparative assessment) of Ildefonso and DiegoRamirez were completed in 2006, 2011 and 2012.
The census at Diego de Almagro in 2001 was conducted usingyatch-based photography (Lawton et al., 2003). The censuses atIldefonso and Diego Ramirez in 2002 and 2011, and at Evangelistasin 2002, were made using photography from a twin otter aircraft(Robertson et al., 2007; Arata et al., 2003). The population at IsloteAlbatros was surveyed in 2003 using yatch-based photography(Aguayo et al., 2003), and by aerial photography in 2006 and2011 (Moreno and Robertson, 2008; this report). The populationat Islote Leonard was assessed by ground count in 2005 (Marinand Oehler, 2007). The methods adopted for the air surveys werekept as consistent as possible in all years and at all locations. Weused the same aircraft, photographic altitude (300–500 m) and an-gle (perpendicular to the landscape), photographer (GR), albatrosscounter (KL) and counting methods (see Lawton et al., 2003;Robertson et al., 2007). For reasons of accuracy every effort wasmade to ensure the air surveys were conducted between 1 h beforeand 4 h after solar noon, when the proportion of birds sitting onnests is highest and the proportion of birds not on nests (partnerbirds and non-breeders) is lowest (Robertson et al., 2008a).
2.2. Fisheries and albatross bycatch estimations
All fisheries operating within or adjacent to the Chilean eco-nomic exclusive zone (EEZ) with the potential to interact fatallywith black-browed and grey-headed albatrosses were included inthe investigation. Relevant information about the fisheries andthe seabird bycatch levels are presented briefly below.
2.2.1. Pelagic longlineThis fishery principally targets broad billed swordfish (Xiphias
gladius) and tunas (Thunnus spp.) with the American and Spanishlongline systems (see Vega and Licandeo, 2009). The fleet includesvessels <28 m long and varying in size from 12 vessels in 2000 toseven vessels in 2011/12. Prior to 2000 four high seas vessels(>40 m in length) operated in the fishery outside the Chilean EEZsetting �4 million hooks annually (R. Vega, personal communica-tions). The domestic fishery is concentrated at 19–39�S and de-ploys up to 1.2 million hooks annually with most effort occurringbetween Coquimbo and the Juan Fernandez archipelago, includingEaster, Salas and Gómez islands and the adjacent open ocean(Azócar et al., 2011; González et al., 2012). Line setting nominallycommences at 1700 h (peak time of setting: 1900 h) and ends atabout 2130 h (Azócar et al., 2008). Albatross mortality estimateswere available for 2005–2010 (Azócar et al., 2011).
2.2.2. Demersal artisanal longlineThis fishery targets Patagonian toothfish, Austral hake (Merluc-
cius australis) and congrio dorado (Genypterus blacodes, hereafterreferred to as ‘‘congrio’’). Gear configurations differ slightly withspecies targeted but all are variations of a single demersal longline
(see Moreno et al., 2006). The artisanal Patagonian toothfish fisheryis concentrated along the Chile-Perú trench in waters 1000–2000 m deep. In the past, this fishery comprised >300 vessels upto 18 m in length. Currently <200 artisanal vessels operate in thefishery, mostly from 41 to 47�S and 18 to 30 km off the southernChilean archipelagos. Occasionally vessels operate illegally southof 47�S in the zone specified for industrial vessels (Oyarzún et al.,2003). Lines are typically set during the day. Effort ranged from17 to 20 million hooks annually in the early 2000s to <0.5 millionhooks in 2011. The artisanal fishery for Austral hake and congrio isconcentrated in the inner channels south of Puerto Montt (41.5–45.9�S) in waters >100 m to <500 m deep. Effort varied from 0.9million hooks/year prior to 2002 to 1.8 million/year from 2002 to2011/12 (Moreno et al., 2006). Lines for Austral hake and congrioare typically set at night. Seabird mortality was estimated in1999 (Austral hake/congrio) and 2002 (Patagonian toothfish) (seeMoreno et al., 2006).
2.2.3. Demersal industrial longline – Austral hake and congrioThis fishery commenced in 1987 and operates inside the Chil-
ean channels and in the open ocean from 45 to 57�S at depths from200 to 600 m. The number of vessels peaked at 52 in 1990 and de-creased thereafter (Subsecretaría de Pesca, 2012a). Hook effort ran-ged from 9.1 million in 1987 to 65.5 million in 1990 (Fig. 2). In2010 only eight vessels operated in the fishery with a fishing effortof 13.5 million hooks. Vessels use the Spanish system (as for Pata-gonian toothfish) with floats attached mid-way between weightson the hook line to raise sections of the line off the seabed. Linesof �15,000 hooks are currently set almost exclusively at night, typ-ically near midnight, although some lines may be set near dawnand in daylight. Observations on 96 sets on two vessels in Octo-ber/November 2003 and March 2004 suggest that sets undertakenin daylight or near dawn could result in significant albatross by-catch. An average of 0.0181 black-browed albatrosses/1000 hooksand no grey-headed albatrosses were killed (unpublished datafrom Moreno et al., 2003 and Moreno and Arata, 2004). This esti-mate was scaled on hooking effort and extrapolated for the period1987–2010 to derive an overall estimate of albatross mortality forthe fishery.
2.2.4. Demersal industrial longline – Patagonian toothfishThis fishery is one of the most important in terms of risks to
albatrosses given its proximity to albatross breeding islands. Thefishery commenced in 1988 under an exploratory regime and by1994 up to 26 vessels operated in the fishery due to the inclusion
322 G. Robertson et al. / Biological Conservation 169 (2014) 319–333
of vessels from Spain under a UNCLOS agreement between Chileand Spain. This number subsequently decreased and from 1997to 2006 14 or 15 vessels operated in the fishery. That number grad-ually reduced to 11 vessels in 2007–2009, then to nine in 2010 andseven in 2011 and 2012. Typically, lines are set in daylight and twi-light but due to operational and weather-related events may be setat any time of the day/night cycle. Effort ranged from 499,377hooks in 1989 to 16.392 million hooks in 1994 (Fig. 2). Effort isconcentrated (by law) south of 47�S.
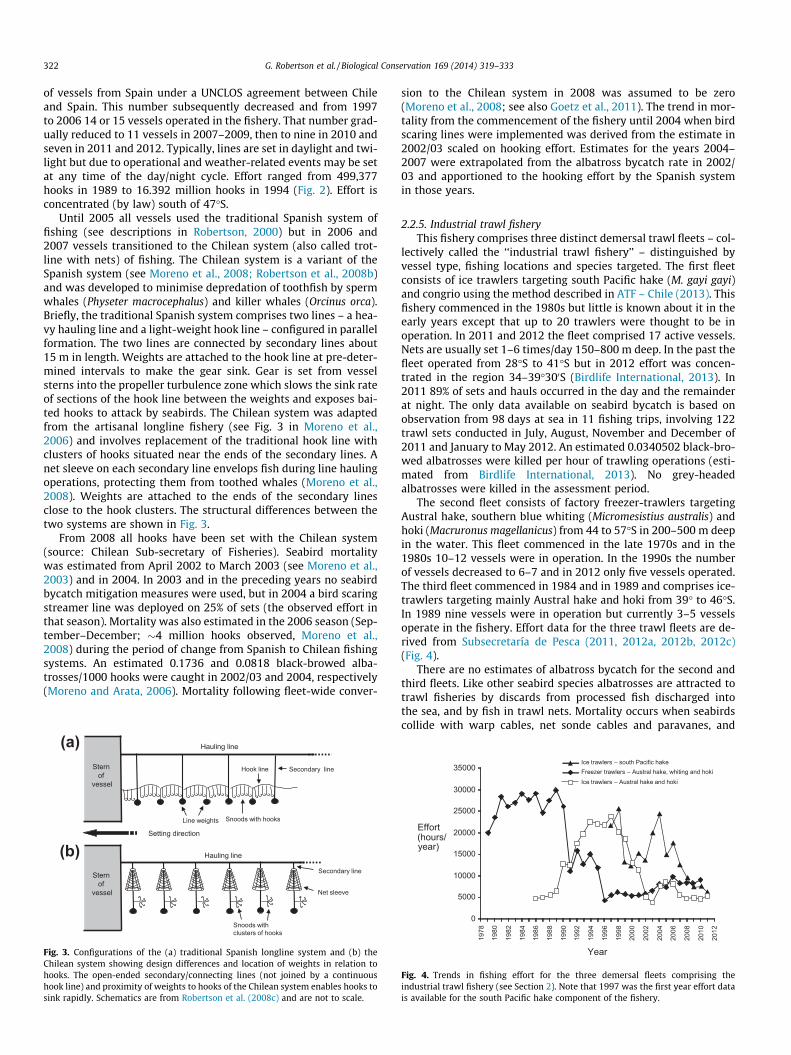
Until 2005 all vessels used the traditional Spanish system offishing (see descriptions in Robertson, 2000) but in 2006 and2007 vessels transitioned to the Chilean system (also called trot-line with nets) of fishing. The Chilean system is a variant of theSpanish system (see Moreno et al., 2008; Robertson et al., 2008b)and was developed to minimise depredation of toothfish by spermwhales (Physeter macrocephalus) and killer whales (Orcinus orca).Briefly, the traditional Spanish system comprises two lines – a hea-vy hauling line and a light-weight hook line – configured in parallelformation. The two lines are connected by secondary lines about15 m in length. Weights are attached to the hook line at pre-deter-mined intervals to make the gear sink. Gear is set from vesselsterns into the propeller turbulence zone which slows the sink rateof sections of the hook line between the weights and exposes bai-ted hooks to attack by seabirds. The Chilean system was adaptedfrom the artisanal longline fishery (see Fig. 3 in Moreno et al.,2006) and involves replacement of the traditional hook line withclusters of hooks situated near the ends of the secondary lines. Anet sleeve on each secondary line envelops fish during line haulingoperations, protecting them from toothed whales (Moreno et al.,2008). Weights are attached to the ends of the secondary linesclose to the hook clusters. The structural differences between thetwo systems are shown in Fig. 3.
From 2008 all hooks have been set with the Chilean system(source: Chilean Sub-secretary of Fisheries). Seabird mortalitywas estimated from April 2002 to March 2003 (see Moreno et al.,2003) and in 2004. In 2003 and in the preceding years no seabirdbycatch mitigation measures were used, but in 2004 a bird scaringstreamer line was deployed on 25% of sets (the observed effort inthat season). Mortality was also estimated in the 2006 season (Sep-tember–December; �4 million hooks observed, Moreno et al.,2008) during the period of change from Spanish to Chilean fishingsystems. An estimated 0.1736 and 0.0818 black-browed alba-trosses/1000 hooks were caught in 2002/03 and 2004, respectively(Moreno and Arata, 2006). Mortality following fleet-wide conver-
(a)
(b)
vessel
vessel
Fig. 3. Configurations of the (a) traditional Spanish longline system and (b) theChilean system showing design differences and location of weights in relation tohooks. The open-ended secondary/connecting lines (not joined by a continuoushook line) and proximity of weights to hooks of the Chilean system enables hooks tosink rapidly. Schematics are from Robertson et al. (2008c) and are not to scale.
sion to the Chilean system in 2008 was assumed to be zero(Moreno et al., 2008; see also Goetz et al., 2011). The trend in mor-tality from the commencement of the fishery until 2004 when birdscaring lines were implemented was derived from the estimate in2002/03 scaled on hooking effort. Estimates for the years 2004–2007 were extrapolated from the albatross bycatch rate in 2002/03 and apportioned to the hooking effort by the Spanish systemin those years.
2.2.5. Industrial trawl fisheryThis fishery comprises three distinct demersal trawl fleets – col-
lectively called the ‘‘industrial trawl fishery’’ – distinguished byvessel type, fishing locations and species targeted. The first fleetconsists of ice trawlers targeting south Pacific hake (M. gayi gayi)and congrio using the method described in ATF – Chile (2013). Thisfishery commenced in the 1980s but little is known about it in theearly years except that up to 20 trawlers were thought to be inoperation. In 2011 and 2012 the fleet comprised 17 active vessels.Nets are usually set 1–6 times/day 150–800 m deep. In the past thefleet operated from 28�S to 41�S but in 2012 effort was concen-trated in the region 34–39�300S (Birdlife International, 2013). In2011 89% of sets and hauls occurred in the day and the remainderat night. The only data available on seabird bycatch is based onobservation from 98 days at sea in 11 fishing trips, involving 122trawl sets conducted in July, August, November and December of2011 and January to May 2012. An estimated 0.0340502 black-bro-wed albatrosses were killed per hour of trawling operations (esti-mated from Birdlife International, 2013). No grey-headedalbatrosses were killed in the assessment period.
The second fleet consists of factory freezer-trawlers targetingAustral hake, southern blue whiting (Micromesistius australis) andhoki (Macruronus magellanicus) from 44 to 57�S in 200–500 m deepin the water. This fleet commenced in the late 1970s and in the1980s 10–12 vessels were in operation. In the 1990s the numberof vessels decreased to 6–7 and in 2012 only five vessels operated.The third fleet commenced in 1984 and in 1989 and comprises ice-trawlers targeting mainly Austral hake and hoki from 39� to 46�S.In 1989 nine vessels were in operation but currently 3–5 vesselsoperate in the fishery. Effort data for the three trawl fleets are de-rived from Subsecretaría de Pesca (2011, 2012a, 2012b, 2012c)(Fig. 4).
There are no estimates of albatross bycatch for the second andthird fleets. Like other seabird species albatrosses are attracted totrawl fisheries by discards from processed fish discharged intothe sea, and by fish in trawl nets. Mortality occurs when seabirdscollide with warp cables, net sonde cables and paravanes, and
0
5000
10000
15000
20000
25000
30000
35000
19
78
19
80
19
82
19
84
19
86
19
88
19
90
19
92
19
94
19
96
19
98
20
00
20
02
20
04
20
06
20
08
20
10
20
12
– hoki
I – hoki
–
Year
Effort(hours/year)
Fig. 4. Trends in fishing effort for the three demersal fleets comprising theindustrial trawl fishery (see Section 2). Note that 1997 was the first year effort datais available for the south Pacific hake component of the fishery.

G. Robertson et al. / Biological Conservation 169 (2014) 319–333 323
when they become entangled in nets (Weimerskirch et al., 2000;Sullivan et al., 2006a, 2006b; ATF – Chile, 2013). All three fleetsare similar in their offal discharge practices, length of warp cables(15–20 m), net mesh sizes (6–10 cm), and all conduct trawlingoperations during the day (Céspedes et al., 2011). The main differ-ence is the absence of net sonde cables in the factory freezer fleet.Because of these similarities and distribution of fishing effort inrelation to albatross breeding season foraging ranges (see above)the black-browed albatross mortality rate for the second and thirdfleets was assumed to be the same as for the south Pacific hakefishery.
2.3. Overlaps with fisheries
2.3.1. Albatross trackingAlbatross foraging ranges and spatial and temporal overlaps
with fisheries were assessed in five breeding seasons between1997 and 2011. The critical details about the tracking studies areshown in Table 1. Albatrosses were tracked at Diego Ramirez, Ilde-fonso and Diego de Almagro (not at the three other sites). In 1997,1999, 2000 and 2001 albatrosses were tracked with Sirtrack ST-10satellite transmitters using the Argos CLS satellite system. In 2011the albatrosses at Diego Ramirez were tracked with Sirtrack G1H134A and i-gotU GT-120 microGPS tags (20 g) with a 30 min sam-pling interval (for methods see Arata, 2004; Lawton et al., 2008).Briefly, the majority of albatrosses tracked were captured and fit-ted with instruments as they departed colonies following nestchange over. When time to observe changeovers was limited(e.g., Diego de Almagro, due to colony remoteness and time con-straints) devices were fitted to incubating albatrosses. On theseoccasions (and when retrieving devices) birds were not removedfrom their nest to minimise the chances of abandonment. All track-ing devices were attached to back feathers between the shoulderblades using #4651 Tesa tape and a thin smear of Loctite 401 glue.A total of 152 individual black-browed albatrosses and 70 individ-ual grey-headed albatrosses were tracked at the three islands inthe five study years.
2.3.2. Albatross foraging ranges and fishery distributionsTracking data were projected into a Lambert Equal-Area Azi-
muthal projection centred on the midpoint of all locations, to en-sure the accuracy of all spatial analyses. Erroneous locationswere removed from the data using a speed filter adapted fromMcConnell et al. (2002). Locations were removed iteratively by iso-lating the point with the maximum velocity, and testing it againsta maximum likely velocity – in this instance 100 km/h (BirdLife
Table 1Number of individual black-browed and grey-headed albatrosses at each breeding site tracwere satellite transmitters (ST) and GPSs. Breeding stages are incubation (Inc.), brood/gua
Species Location Year
Black-browed albatross Diego Ramirez 19971999200020012011
Ildefonso 2001Diego de Almagro 2001
Total
Grey-headed albatross Diego Ramirez 1997199920002001
Ildefonso 2001
Total
International, 2004). About 30% of points were removed from theArgos data. No GPS data showed movements above maximumvelocity. Locations were interpolated to make the two data typescomparable to prevent sample bias. Locations were interpolatedto represent hourly locations for the tracked individual assuminga straight line of flight between to known locations. Thus addi-tional points were added to the satellite data but GPS data, col-lected at a higher frequency, were thinned.
Data were assembled into data groups by breeding population.A kernel density estimation (KDE) was applied to each data groupusing a smoothing factor of 110 km (BirdLife International, 2004).These KDE analyses resulted in density surfaces of bird hours indi-cating the distribution of birds from each population studied. Eachdensity surface was then normalised to remove any bias in thetracking survey effort apparent in the data and utilisation distribu-tions (UDs; the proportion of time spent within a given area) werecalculated for each island population on 3-degree grid squares. UDsfor the Chilean population as a whole were then calculated. Esti-mates of breeding population sizes were used to calculate the pro-portion of the total Chilean population represented by eachindividual island. Finally, using these abundance-weighted distri-butions the density surfaces were combined to create an estimateof the Chilean population UDs.
To examine the extent of overlap between albatrosses andfisheries, fishing effort for the five fisheries mentioned above wasoverlaid on the albatross foraging distributions using the same 3-degree grid squares. Only fishing effort data coincident with theyears of albatross tracking studies shown in Table 1 were consid-ered. The number of longline and trawl sets was summed for eachgrid square to give a map of relative fishing effort for each fisherywhich pertains to the number of sets of the longline for the fourtypes of longline fisheries and the number of sets of the net forthe trawl fishery.
2.4. Population modelling using SEABIRD
The purpose of the population modelling was to evaluate thehypothesis that reduced mortality in fisheries was the principalreason for the observed increase in black-browed albatross num-bers. For the reasons outlined below (see Section 4) only data forthe industrial longline fishery for Patagonian toothfish were usedin the model. It was assumed that environmental factors affectingsurvival and reproductive output and the extent of supplementaryfeeding from discharged offal (e.g., Thompson and Riddy, 1995) didnot change greatly in the life of the fishery. Our hypothesis was as-sessed with a stage-structured population model implemented in
ked during the study. Year denotes year at beginning of the breeding season. Devicesrd (bg) and post-brood/guard (pbg).
Device Breeding stage Individuals tracked (n)
ST Inc. 25ST Inc., bg 15ST Inc., bg, pbg 22ST Inc., bg, pbg 25GPS Inc. 26ST Inc. 26ST Inc. 13
152
ST Inc., bg 17ST Inc., bg, pbg 12ST Inc., bg, pbg 22ST Inc., bg, pbg 18ST Inc. 1
70

324 G. Robertson et al. / Biological Conservation 169 (2014) 319–333
the program SEABIRD (Fu and Francis, 2008), using the demo-graphic statistics for black-browed albatross in Appendix A. Cen-suses in 2002 and 2011 were only completed at two sites, DiegoRamirez and Ildefonso (see below). Only islands in each of theselocations surveyed in both 2002 and 2011 were included in themodel. This included all islands at Ildefonso and Diego Ramirez ex-cept Isla Bartolomé, Isla Santander, and Islote Ester. The combinedtotal numbers of black-browed albatrosses at Ildefonso and DiegoRamirez in 2002 and 2011 included in the model were 58,357and 71,726 birds, respectively, representing 55.1% of the total com-bined populations in 2002. This proportion was used to apportionthe estimated numbers of black-browed albatrosses killed inciden-tally from 1989 to 2011. It was assumed that all black-browedalbatrosses killed in the fishery were taken in equal proportionto their abundance and the bycatch rate for the modelled portionof the populations (see above and Table 3) was the same for bothbreeding sites. The northern group of islets at Ildefonso was re-sur-veyed in 2012 to determine population change since 2011 and toenable comparison with predictions from the model.
A baseline model was defined that predicted the population tra-jectory during two different periods (see Appendix A). The firstperiod went from the year (1988) before the fishery commenceduntil 2002, when the albatrosses were first censused and whenalbatross bycatch was estimated in the fishery. The second periodwas from 2002 to 2011, the year of the final albatross census. Thisperiod includes the time when bycatch by Spanish system vesselsoccurred (up until 2008) and then dropped to zero (2008 onwards).Because the two demographic parameters N0 and SEB (see Appen-dix A) were estimated from just two albatross census years, thisbaseline model will give an almost perfect prediction, meaningthat it was not possible to separate the effect of bycatch from nat-ural demographic parameters on population increase between thetwo census years. To tease out the effect of bycatch from naturaldemographic parameters on population increase two additionalmodels based on artificial (and contrasting) bycatch scenarios weredeveloped for 2002–2011. Bycatch scenario 1 applies the same by-catch estimated for the baseline model up until 2008 but from2008 to 2011 applies an artificial bycatch of 2000 black-browedalbatrosses per year. Bycatch scenario 2 is identical to scenario 1with the exception that the artificial bycatch was 1000 black-bro-wed albatrosses per year (see Appendix B). These values wereapportioned to the modelled fraction of the population only. Thesame demographic parameters and bycatch rates until 2007 (theyear before the change in fishing gear) were used for all models,so all models give the same prediction for all years up to andincluding 2007 (Appendix B). By this approach the degree to whichbycatch scenarios 1 and 2 gave reduced population estimates sug-gests that the population increase from 2002 to 2011 was primar-ily due to reduced levels of bycatch. The dates critical to the modelare summarised in Table 2.
Table 2List of key dates and events relevant to the model for the industrial longline fishery for P
Years Events
1989 First year of the fishery1989–2005 All hooks set by traditional Spanish2006 and 2007 Transition from Spanish to Chilean fi2008–2011 All hooks set by Chilean system; by2002 First albatross bycatch estimate (Spa2004 Second albatross bycatch estimate (2006 Third albatross bycatch estimate (Ch1989–2001; 2003–2011 Bycatch estimate based on hooking2002 and 2011 Years of main albatross censuses1989–2007 Period in which baseline, bycatch sc2008–2011 Period in which predictions of popu2012 Albatross census at northern group
Black-browed albatross bycatch rates from 1989 to 2011 wereobtained by multiplying annual hook effort for the Spanish systemwith the observed bycatch rate in 2002. From 2008 to 2011 mortal-ity was assumed to be zero (see Moreno et al., 2006). The estimatedbycatch was scaled by multiplying by 0.551 to account for the pop-ulation fraction being modelled.
2.5. Trend in marine productivity
To investigate whether variations in marine productivity mayhave contributed to the observed increase in black-browedalbatross numbers, we used SeaWiFS estimates of surface chloro-phyll-a (Feldman and McClain, 2010). Monthly mean chlorophyll-a values were extracted and converted into seasonal compositesspanning September–May of each austral summer from 1997/98to 2009/10. The means and interquartile ranges of the seasonalchlorophyll-a values within the 95% UD of black-browed alba-trosses were then collated.
3. Results
3.1. Albatross population trends
In the nine years between 2002 and 2011 the number of black-browed albatrosses at selected islands at Diego Ramirez increasedby 52% at an average annual rate of 4.8% (Table 3). In the same timeperiod the population at Ildefonso increased by 18% and averaged1.8%/year. Between 2011 and 2012 the number of black-browedalbatrosses at the northern group of Ildefonso increased by 6.8%(from 23,819 to 25,444). Overall, the number of albatrosses inthe surveyed portions of Diego Ramirez and Ildefonso increasedby 23% (from 58,357 to 71,726) in the nine years between surveys(annual average rate: 2.32%). From 2006 to 2011 the small colonyon Islote Albatros increased in number by 71%. Note, however, thatthe observed annual growth rate (11.3%) for this colony should betreated with caution because of possible immigration. Efforts in2011 and 2012 to survey the populations of black-browed alba-trosses at Diego de Almagro, Islote Evangelistas and Islote Leonardproved unsuccessful (see below). In the same time period that theDiego Ramirez black-browed albatross numbers increased thenumbers of grey-headed albatrosses remained stable (Table 3).
3.2. Foraging ranges and fisheries overlaps
Tracking data were compiled of all available tracking informa-tion from Diego Ramirez, Ildefonso and Diego de Almagro for theincubation period (Figs. 5 and 6). In general, black-browed alba-trosses from the three sites aggregated over continental shelf andslope waters of central (north to 35�S) and southern Chile to�58�S and made sporadic trips into deeper oceanic waters to the
atagonian toothfish.
systemshing systems
catch drops to zeronish method)
Spanish method, introduction of bird scaring lines)ilean method)
effort
enarios 1 and 2 models gave the same prediction of population sizelation size from actual (baseline) and bycatch scenarios 1 and 2 were comparedof Ildefonso

Table 3Estimated numbers of black-browed (BBA) and grey-headed (GHA) albatrosses at selected locations in the Diego Ramirez and Idlefonso archipelagos, and at Islote Albatros, in thecensus years shown. In 2002 Isla Bartolome and Islote Ester in the Diego Ramirez group were surveyed using a mix of census methods and therefore were not included in aerialcensuses in 2011 and 2012. Islote Santander was also not surveyed from the air in 2011 and 2012. In 2002 Bartolome, Ester and Santander held 47,453 of a total of 56,000 (raw,uncorrected, count) black-browed albatrosses at Diego Ramirez (Robertson et al., 2007). Therefore the estimates below for black-browed albatrosses at Diego Ramirez indicatetrends only, not the size of the total population. The estimates for Ildefonso in 2002 and 2011 include all islets in the archipelago and are indicative of the total population sizes inthose years. All numbers are raw, uncorrected, counts (see Robertson et al., 2007, 2008a). The estimate for Islote Albatros in 2006 is from Moreno and Robertson (2008). Meanchange/year calculated as (e.g., Total Diego Ramirez) 100 � ((13,549/8889)^(1/9) � 1). Ns = not surveyed.
Species Location Group/islet Year Change (%)
2002 2006 2011 2012 Total Mean/y
BBA Diego Ramirez Northern groupIslote Cabezas 0 0 nsIslote Penailillo 0 0 nsIslote Norte 1316 3826 nsIslote Schlatter 178 204 nsIslote Martinez 286 227 nsIslote Mendoza 143 128 nsTotal northern group 1923 4385 128 9.6
Southern groupIsla Gonzalo 6966 7807 9164 32a 3.1a
Total Diego Ramirez (see caption) 8889 13,549 52 4.8
Ildefonso Northern groupIsla Norte 10,920 13,920 14,799 27a 2.7a
Isla Cinclodes 775 871 1029 12a 1.3a
Isla Square 488 528 593 8a 0.9a
Isla Spirit 1383 1447 1747 5a 0.5a
Isla Sur 5222 7053 7276 35a 3.4a
Total northern group 18,788 23,819 25,444 27a 2.7a
6.8b 6.8b
Southern groupIsla Grande 30,680 31,770 34,358 ns 12a 1.3a
Total Ildefonso (see caption) 49,468 58,177 18 1.8
Islote Albatros ns 65 111 ns 71c 11.3c
GHA Diego Ramirez Northern groupIslote Cabeza 0 0 nsIslote Penailillo 0 0 nsIslote Norte 463 476 nsIslote Schlatter 97 76 nsIslote Martinez 69 49 nsIslote Mendoza 174 279 nsTotal northern group 803 870
Southern groupIsla Gonzalo 4523 4122 4413 nsTotal Diego Ramirez (see caption) 5326 5293 0
a Change from 2002 to 2011.b Change between 2011 and 2012.c Change from 2006 to 2011.
G. Robertson et al. / Biological Conservation 169 (2014) 319–333 325
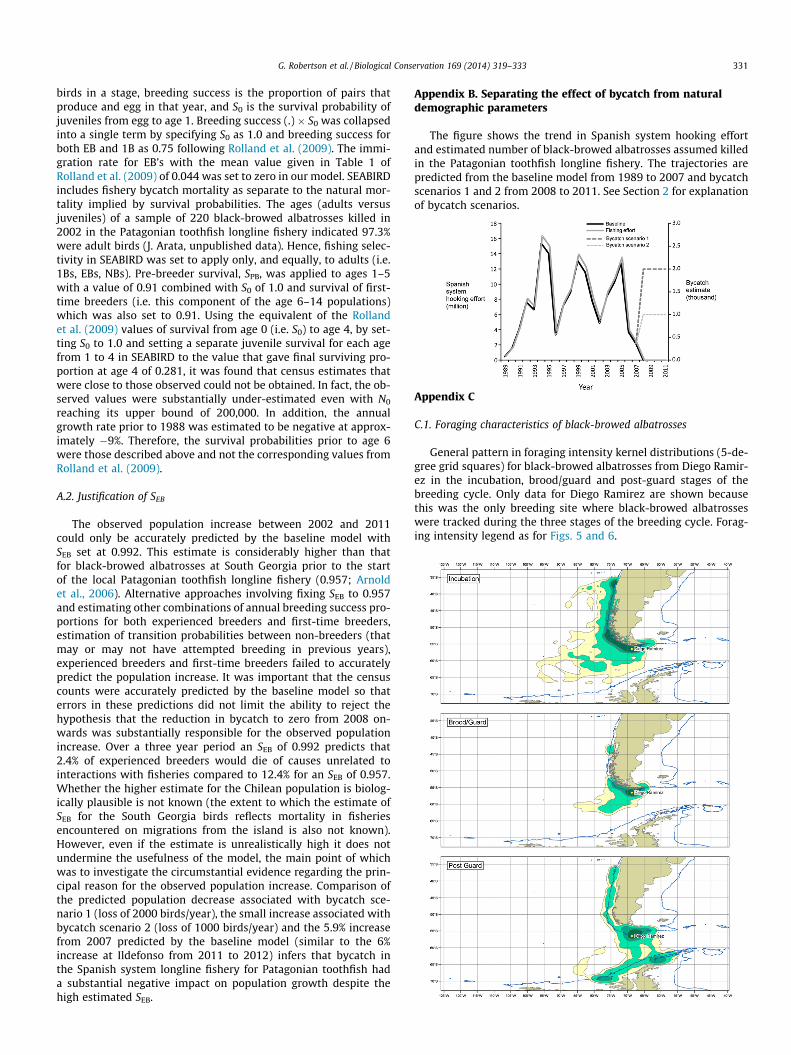
west of Chile. A total of 83% of foraging time occurred within theChilean EEZ. Black-browed albatrosses from Diego Ramirez, wheretracking occurred at various stages of the breeding cycle (AppendixC.1), concentrated during the incubation period along the Chileancoast off the Arauco Gulf (37�S), Chiloé Island (43�S) and southernChile (52–57�S). During the brood/guard stage the birds were ac-tive mainly around the southern tip of the Chilean continental shelfand during the post-guard stage aggregations occurred aroundsouthern Chile with excursions to waters to the north along thecoast of Chile and along the continental shelf and slope of the wes-tern side of the Antarctic Peninsula.
The foraging range of grey-headed albatrosses from Diego Ra-mirez (only one bird tracked from Ildefonso) differed markedlyto that of black-browed albatrosses. In general, during incubationgrey-headed albatrosses moved over a vast area of the ocean from45�W to 115�W and from �46�S to 66�S. The southernmost pointcoincided with the waters of the Antarctic Polar Front. Duringthe brood/guard stages activity was greatest closer to Diego Ramir-ez and near the edge of the pack-ice zone on the western edge ofthe Antarctic Peninsula, from �57�W to 90�W and from �56�S to70�S. The same pattern occurred in the post-guard stage except
birds spent more time near the edge of the Antarctic pack-ice zoneand over the Antarctic continental shelf than in the other two peri-ods (Appendix C.2).
Figs. 5 and 6 also show the distribution of relative fishing effortfor the five fisheries analysed in relation to albatross foragingranges during incubation. Since the fishing grounds remained rel-atively unchanged during the fishing seasons, albatross/fisheriesoverlaps in the non-incubation periods can be inferred fromAppendix C.1 (black-browed albatrosses) and C.2 (grey-headedalbatrosses). During incubation there was extensive overlap be-tween black-browed albatrosses from the three breeding sitesand four of the five fisheries (Table 4). Overlap was greatest withthe industrial longline fisheries for Patagonian toothfish and Aus-tral hake and congrio, and the industrial trawl fishery. There wassubstantial overlap between black-browed albatrosses and theartisanal longline fisheries and no overlap with the pelagic longlinefishery in the breeding season (see Fig. 5). The foraging areas ofgrey-headed albatrosses in the breeding season did not overlapwith the pelagic longline, artisanal longline and industrial longline(hake and congrio) fisheries. Overlap was greatest with the indus-trial longline fishery for Patagonian toothfish (41% overlap) and the
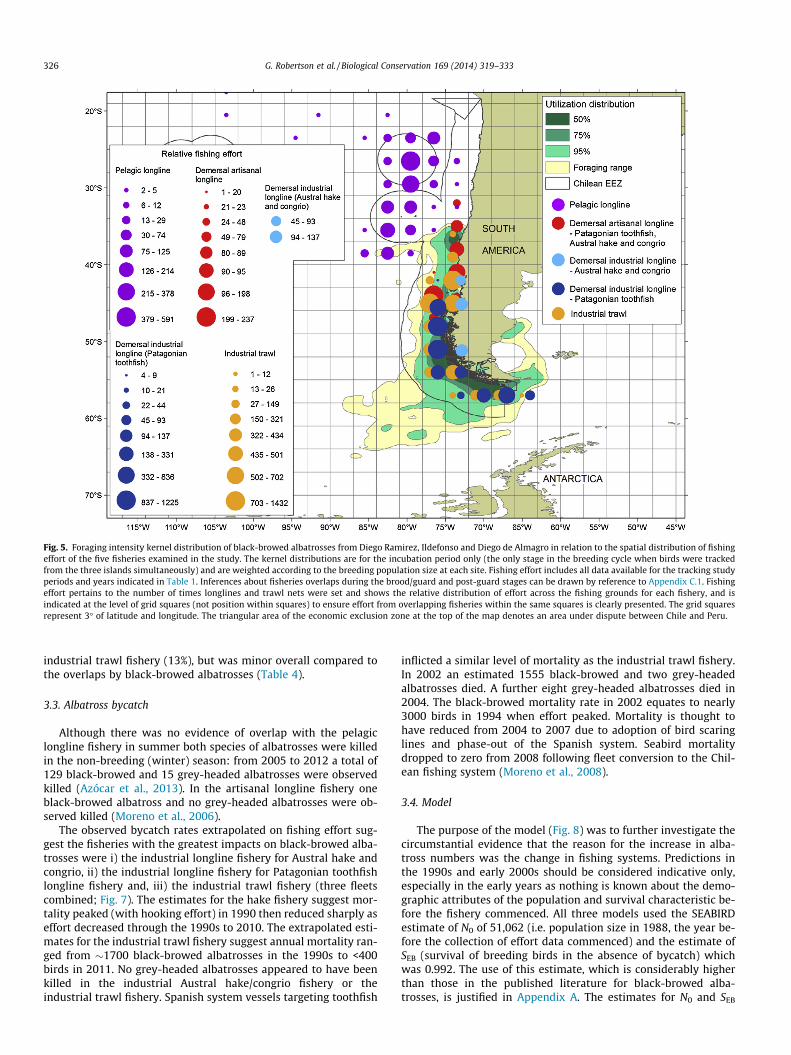
Fig. 5. Foraging intensity kernel distribution of black-browed albatrosses from Diego Ramirez, Ildefonso and Diego de Almagro in relation to the spatial distribution of fishingeffort of the five fisheries examined in the study. The kernel distributions are for the incubation period only (the only stage in the breeding cycle when birds were trackedfrom the three islands simultaneously) and are weighted according to the breeding population size at each site. Fishing effort includes all data available for the tracking studyperiods and years indicated in Table 1. Inferences about fisheries overlaps during the brood/guard and post-guard stages can be drawn by reference to Appendix C.1. Fishingeffort pertains to the number of times longlines and trawl nets were set and shows the relative distribution of effort across the fishing grounds for each fishery, and isindicated at the level of grid squares (not position within squares) to ensure effort from overlapping fisheries within the same squares is clearly presented. The grid squaresrepresent 3� of latitude and longitude. The triangular area of the economic exclusion zone at the top of the map denotes an area under dispute between Chile and Peru.
326 G. Robertson et al. / Biological Conservation 169 (2014) 319–333
industrial trawl fishery (13%), but was minor overall compared tothe overlaps by black-browed albatrosses (Table 4).
3.3. Albatross bycatch
Although there was no evidence of overlap with the pelagiclongline fishery in summer both species of albatrosses were killedin the non-breeding (winter) season: from 2005 to 2012 a total of129 black-browed and 15 grey-headed albatrosses were observedkilled (Azócar et al., 2013). In the artisanal longline fishery oneblack-browed albatross and no grey-headed albatrosses were ob-served killed (Moreno et al., 2006).
The observed bycatch rates extrapolated on fishing effort sug-gest the fisheries with the greatest impacts on black-browed alba-trosses were i) the industrial longline fishery for Austral hake andcongrio, ii) the industrial longline fishery for Patagonian toothfishlongline fishery and, iii) the industrial trawl fishery (three fleetscombined; Fig. 7). The estimates for the hake fishery suggest mor-tality peaked (with hooking effort) in 1990 then reduced sharply aseffort decreased through the 1990s to 2010. The extrapolated esti-mates for the industrial trawl fishery suggest annual mortality ran-ged from �1700 black-browed albatrosses in the 1990s to <400birds in 2011. No grey-headed albatrosses appeared to have beenkilled in the industrial Austral hake/congrio fishery or theindustrial trawl fishery. Spanish system vessels targeting toothfish
inflicted a similar level of mortality as the industrial trawl fishery.In 2002 an estimated 1555 black-browed and two grey-headedalbatrosses died. A further eight grey-headed albatrosses died in2004. The black-browed mortality rate in 2002 equates to nearly3000 birds in 1994 when effort peaked. Mortality is thought tohave reduced from 2004 to 2007 due to adoption of bird scaringlines and phase-out of the Spanish system. Seabird mortalitydropped to zero from 2008 following fleet conversion to the Chil-ean fishing system (Moreno et al., 2008).
3.4. Model
The purpose of the model (Fig. 8) was to further investigate thecircumstantial evidence that the reason for the increase in alba-tross numbers was the change in fishing systems. Predictions inthe 1990s and early 2000s should be considered indicative only,especially in the early years as nothing is known about the demo-graphic attributes of the population and survival characteristic be-fore the fishery commenced. All three models used the SEABIRDestimate of N0 of 51,062 (i.e. population size in 1988, the year be-fore the collection of effort data commenced) and the estimate ofSEB (survival of breeding birds in the absence of bycatch) whichwas 0.992. The use of this estimate, which is considerably higherthan those in the published literature for black-browed alba-trosses, is justified in Appendix A. The estimates for N0 and SEB

Fig. 6. Foraging intensity kernel distribution of grey-headed albatrosses from Diego Ramirez and Ildefonso (one bird only) during the incubation period in relation to thespatial distribution of relative fishing effort of the five fisheries examined in the study. The information on relative fishing effort is as for Fig. 5.
Table 4Comparative overlap between core foraging areas of black-browed albatrosses (BBA)from Diego Ramirez, Ildefonso and Diego de Almagro, and grey-headed albatrosses(GHA) from Diego Ramirez and Ildefonso (one bird only) and the five fisheriesexamined in the study. Core foraging areas were taken to be the 75% utilisationdistributions from Figs. 5 and 6. Shown is the fishing effort as percentage of total setsof longlines and sets of trawl nets. Foraging area/fisheries overlaps pertain to theincubation period when birds from the three locations were tracked concurrently, andare weighted by population size estimated in 2002 (see Robertson et al., 2007).LL = longline.
Fishery Foraging area overlap (%)
BBA GHA
Pelagic LL 0 0Demersal artisanal LL 48 0Demersal industrial LL – hake and congrio 81 0Demersal industrial LL – Patagonian toothfish 97 41
Industrial trawl 83 13
0
500
1000
1500
2000
2500
3000
19
78
19
81
19
84
19
87
19
90
19
93
19
96
19
99
20
02
20
05
20
08
20
11
Year
killed
rawl
longline
toothfish)
longline
congrio)
Fig. 7. Extrapolated trend in black-browed albatross mortality in the industriallongline fisheries for Austral hake/congrio (Spanish system) and Patagoniantoothfish by Spanish system vessels, and the industrial trawl fishery (three fleetscombined). The extrapolations are based on bycatch estimates in 2002 and 2004 forthe Austral hake/congrio longline fishery; 2002 and 2004 for the Patagoniantoothfish longline fishery by Spanish system vessels and 2006 for Patagoniantoothfish longline fishery by Chilean system vessels; and 2011/12 for the trawlfishery. The extrapolations for the former and latter fisheries are tentative andshould be interpreted with caution (see Section 4).
G. Robertson et al. / Biological Conservation 169 (2014) 319–333 327
were obtained by fitting the baseline model. The estimated rate ofincrease from 1988 to 2002 was 1.1% per annum. The estimatedannual rates of increase from 2007 to 2011 for the baseline, by-catch scenario 1 and bycatch scenario 2 models were 6.0%,�1.8%, and 2.1%, respectively. The estimated annual rates of in-crease overall between 2002 and 2011 for the baseline, bycatchscenario 1 and bycatch scenario 2 models were 2.4%, �1.0%, and0.7%, respectively. The differences between these estimates, thefirst with a mix of bycatch (2002–2007) and zero bycatch (2008onwards), and the latter two with the same bycatch between
2002 and 2007 but artificial (and substantial) annual levels of by-catch from 2008 to 2011 (see above) suggests that reduced bycatchin the Patagonian toothfish longline fishery by Spanish system ves-sels was the most compelling reason for the 2.5% observed increase

albatrosses
(thousand)
Year
45
50
55
60
65
70
75
1989
1991
1993
1995
1997
1999
2001
2003
2005
2007
2009
2011
Fig. 8. Population trajectories for the sampled portion (see Section 2) of black-browed albatrosses at Diego Ramirez and Ildefonso predicted by the baseline model(1988–2011; dashed black line) and bycatch scenarios 1 (grey solid line; 2000 birdsper year) and 2 (black solid line; 1000 birds per year) during the years of fishing bythe Spanish system (1989–2005), Chilean system (2008–2011) and transitionbetween the two (2006 and 2007). Also shown is the number of black-browedalbatrosses in the censused portions of the populations (see Section 2 and Table 3)in 2002 and 2011 (black diamonds). The baseline, scenario 1 and scenario 2 modeltrajectories were constrained to be identical for the period 1988–2007 (seeSection 2). The difference between the trajectory for the baseline model and thosefor the scenario 1 and 2 models indicates the observed population increase was dueprimarily to reduced bycatch levels from 2008 onwards in the Patagonian toothfishlongline fishery by Spanish system vessels.
328 G. Robertson et al. / Biological Conservation 169 (2014) 319–333
in albatross numbers at Diego Ramirez and Ildefonso from 2002 to2011. The lambda parameter obtained from SEABIRD, which is theratio of the total population for the last year of the initialisation runof 50 years (default value in SEABIRD) in the absence of fisheriesmortality (with the final year corresponding to 1988) and thesecond-last year of the run (gives one year of growth prior to by-catch being imposed), was 1.0593, an annual increase of 5.9%. Thisestimate compares well to the observed increase between 2011and 2012 at the northern group of Ildefonso, when in the absenceof bycatch in the toothfish fishery the population increased by 6.8%(Table 3) and is similar to the predicted annual increase of 6.0%from 2007 to 2011 for the baseline model, also in the absence ofbycatch.
3.5. Marine productivity
The linear trend term (regression fitted with a 1st order autore-gressive error structure) in year-specific average surface chloro-phyll-a values from 1998 to 2010 was not statistically significant(P > 0.1). Sub-sections of the overall trend (assessed by the additionof a cubic spline term; Verbyla et al., 1999), including the period2002–2010 during which bycatch fell to zero, were also not signif-icant (P > 0.1). The absence of a discernible trend over the wholetime series or sub-sections of the series suggests that the observedpopulation increases at Diego Ramirez and Ildefonso were not ex-plained by variation in marine productivity.
4. Discussion
4.1. Difficulty with the overflights
In 2011 and 2012 37 days were dedicated to re-surveying Diegode Almagro and Islote Evangelistas from the air, and to survey
Islote Leonard from the air for the first time. These islands lie muchfurther north than Diego Ramirez (e.g., 725 km for Diego de Almag-ro). Unfortunately the re-surveys proved impossible due to persis-tent strong wind, turbulence and rain in the northern regions. Thisexperience lies in stark contrast with 2001 when a yacht-basedcensus at Diego de Almagro (Lawton et al., 2003) and aerial censusat Evangelistas (Arata et al., 2003) were completed in the albatrossincubation period when strong winds are the norm. Prolongeddelays have important logistical and financial implications andshould be considered when future surveys are planned at Diegode Almagro and Evangelistas. Because of the inability to repeatthe surveys it remains unclear if the observed increases at DiegoRamirez and Ildefonso were restricted to those sites or occurredgenerally throughout the region.
4.2. Population increases
In the nine years between the two censuses the number ofblack-browed albatrosses increased by 18% at Ildefonso and 52%at Diego Ramirez (23% at both sites combined). There was consid-erable variation within individual islets of both archipelagos: num-bers at the northern groups of Diego Ramirez and Ildefonsoincreased by 128% and 27%, respectively. These changes shouldbe interpreted with caution as the islets form a contiguous chainand it is not known if birds move between them. However, it is un-likely albatrosses move between Ildefonso and Diego Ramirez be-cause these archipelagos lie �90 km apart. The large difference inthe proportional increases at these two sites may indicate differentbycatch levels, availability of nesting space or other factors. Thebulk of the birds in the censuses were at Isla Gonzalo (Diego Ramir-ez) and Isla Grande (Ildefonso; see Fig. 1). Isla Gonzalo is exten-sively covered by soil and grass and nest space does not appearto be limiting, but Isla Grande is predominantly bare rock with lit-tle apparent available space to support population expansion. Ifpopulation growth continues at the current rate (6.8%/year) forthe northern group of Ildefonso, maximum nesting capacity willsoon be reached (if it has not already) and continued growthshould result in dispersal to other known sites or the establish-ment of new colonies in the region.
During the period of the black-browed albatross increases thenumber of grey-headed albatrosses at Diego Ramirez varied by<1%. Grey-headed albatrosses breed predominantly every secondyear (Ryan et al., 2007) and little can be concluded from two esti-mates in just nine years, other than that there was no evidence ofsignificant change. In 2002 and 2004 in the Patagonian toothfishlongline fishery by Spanish system vessels only 10 grey-headedalbatrosses were estimated killed compared to 2444 black-browedalbatrosses. No grey-headed albatrosses were taken in 2011/12 inthe south Pacific hake trawl fishery. The contrasting mortality lev-els for these two species likely reflects differences in foraging strat-egy, at-sea threats and possibly inter-specific competition aroundfishing vessels. Black-browed albatrosses frequented waters ofthe southern Chilean continental shelf and shelf slope, whereasthe main aggregations of grey-headed albatrosses occurred nearthe Antarctic Polar Front and the western side of the Antarctic Pen-insula. The diets reflect the at-sea segregation of the two species.The squid Martialia hyadesi, prevalent at the APF (Rodhouse et al.,1996), dominated the diet of grey-headed albatrosses (Arataet al., 2004) whereas black-browed diet was dominated by speciesassociated with commercial trawl fisheries (Arata and Xavier,2003). The tendency to favour ocean frontal areas over continentalshelves is typical for grey-headed albatrosses (Croxall et al., 2005;BirdLife International, 2006) and must be a key reason for the lowincidence of mortality of this species in the Chilean fisheries. To

G. Robertson et al. / Biological Conservation 169 (2014) 319–333 329
our knowledge there are no legal toothfish vessels operating in thebirds’ foraging ranges outside the Chilean EEZ and in the AntarcticPeninsula region. Toothfish stocks on the western side of the Pen-insula are not considered commercially viable (Arana and Vega,1999).
With respect to competition around fishing vessels, before sea-bird conservation measures were widely adopted in CCAMLRwaters and in Chile, grey-headed albatrosses suffered higher mor-tality where black-browed albatrosses were less abundant (e.g.,Nel et al., 2002). For example, more grey-headed albatrosses werekilled in the Kerguelen/Crozet fishery (Weimerskirch et al., 2000)than in this study. Kerguelen/Crozet holds three-times moregrey-headed than black-browed albatrosses, but at Diego Ramirezthat species ratio is reversed (Robertson et al., 2007). In the earlyyears of the South Georgia (black-browed albatrosses about 50%more numerous; Poncet et al., 2006) toothfish fishery, fewergrey-headed than black-browed albatrosses were taken (Morenoet al., 1996), which is more similar to the current situation inChile. Although grey-headed albatrosses attended fishing vessels(see Moreno et al., 2003) the numerical superiority (and aggres-siveness) of black-browed albatrosses may have limited theirinteractions. The lack of significant mortality in local fisheriesand apparent absence of commercial fisheries in foraging rangesduring the breeding season suggests grey-headed albatrossnumbers at Diego Ramirez could be influenced more by intrinsicdemographic factors and environmental processes than by mortal-ity in fisheries.
4.3. Albatross bycatch estimates and caveats
Black-browed albatross bycatch appeared to be significant inonly three of the five fisheries examined: the industrial longlinefishery for Patagonian toothfish; the industrial longline fisheryfor Austral hake and congrio; and the industrial trawl fishery.However, the bycatch extrapolations for the Austral hake/congriolongline fishery and the trawl fishery could be affected by thefollowing factors. Longlines were intentionally set at night tomaximise fish catch, but seven of the 10 black-browed alba-trosses observed were caught on the three sets (of 96 sets intotal) made in daylight/twilight, when seabird capture is morelikely. Also, the bycatch estimate was based mainly on datagathered in summer when night setting operations are morelikely to extend into dawn in the region where the fisheryoperated (45–57�S). Hence, bycatch in this fishery could be over-estimated. In the industrial trawl fishery, the Austral hake, south-ern blue whiting and hoki fleet commenced in 1979, 32 yearsbefore the first estimate was made of seabird bycatch in theindustrial trawl fishery in Chile. Back-extrapolation over such along period of time cast doubt over the validity of the bycatchestimates for this component of the industrial trawl fishery. Fur-ther , bycatch was estimated from 34� to 39�300S, the area ofgreatest effort, and extrapolated across the three trawl fleets. Thisarea lies north of the main foraging range of black-browedalbatrosses during incubation (50% UD in Fig. 5) and even moreso later in the breeding season when foraging activity contractedto the higher latitudes (see Appendix C.1). Due to these concernsthe bycatch estimates for the both these fisheries should beconsidered tentative. We have more confidence in the estimatesfor industrial longline fishery for Patagonian toothfish. Thisfishery operated as a single fleet between the latitudinalbands (47–56�S) that encompass all six albatross breeding sites.Observer coverage was the highest (40% of fleet) of all fisheriesexamined and the bycatch estimates the most comprehensive,being derived from monthly weighted averages. This last point
minimises the effects of temporal variation in fisheries/albatrossdistributions mentioned for the trawl fishery.
Annual mortality in the Austral hake/congrio longline fishery,though high in 1990 (when effort peaked), was minor in compari-son to the other two fisheries and remained relatively stable in theperiod of the albatross population increases. The highest mortalityoccurred in the Spanish-rig toothfish longline and industrial trawlfisheries. From 1989 to 2007, years when both fisheries were inoperation, 44% and 41% of total mortalities were attributed to theSpanish-rig toothfish longline and trawl fisheries, respectively. Asmany as 2800 albatrosses may have been taken in the former fish-ery in 1994 and 1750 in 1998 in the latter fishery. The high levelsof mortality in these fisheries almost certainly reflect the absenceof effective seabird bycatch mitigation measures. Measures criticalto the toothfish longline fishery include bird scaring lines andappropriate line weighting (and night setting). The absence ofscaring lines in Spanish system longline fisheries can have disas-trous consequences for black-browed albatrosses (Otley, 2005;Robertson et al., 2008c). The weighting regimes adopted byCCAMLR (8.5 kg every 40 m) yield faster sink rates (Robertsonet al., 2008b) and significantly lower mortality rates of black-browed albatrosses (Agnew et al., 2000) than the weighting regime(4–8 kg/50 m) used by Spanish-rig vessels in this study. In trawlfisheries scaring lines (or equivalent) around warp cables arehighly effective in minimising the mortality of black-browedalbatrosses (Sullivan et al., 2006b).
4.4. Model results and reason for population increases
The circumstantial evidence suggests the black-browed alba-tross populations at Diego Ramirez and Ildefonso increased princi-pally because of reduced mortality in the Patagonian toothfishlongline fishery by Spanish-rig vessels. This conclusion is sup-ported by the trend in the provisional bycatch estimates for thetrawl fishery: the number of albatrosses taken did not vary greatlyfor the duration of the fishery (see Fig. 7) and though decreasing inthe latter years remained at �400 albatrosses at the end of thestudy. This contrasts with bycatch levels by Spanish system long-line vessels which decreased substantially in the 2000s. The firstphase of the decrease in albatross bycatch occurred following thephased introduction of bird scaring lines from 2002 onwards. The3.6% increase in numbers of black-browed albatrosses at IslaGrande, Ildefonso, from 2002 and 2006 (see Table 3) could indi-rectly indicate use of these devices. The second phase of the de-crease occurred with the transition to the Chilean system offishing. The third phase occurred in 2008 when all hooks wereset with the Chilean system and mortality of black-browed alba-trosses (and other seabird species) was reduced to zero. The ob-served increase at Isla Grande from 2006 to 2011 was higher(8.1%) than from 2002 to 2006, which is consistent with continuedreduced bycatch following the phase out of the Spanish system.
This conclusion is supported by the output of the model. Themodel does not predict well from 2003 to 2006 which includesthe period when bycatch is thought to have decreased due to useof bird scaring lines. This could reflect the assumptions associatedwith the model construction and bycatch estimates. However, thekey components of the model are the periods 2002–2011 and2007–2011. In the former period the model predicted growth of2.4% with a mix of bycatch (2002–2007) and no bycatch (2007–2011), which is similar to the observed growth (2.5%) in themodelled portion of the albatross population at Diego Ramirezand Ildefonso for the same period. In the latter period the modelpredicts 5.9% growth in the absence of bycatch, similar to the ob-served population increase (6.8%) at Ildefonso from 2011 to 2012,

330 G. Robertson et al. / Biological Conservation 169 (2014) 319–333
also in the absence of bycatch. This supports the circumstantialevidence that reduced mortality in the Patagonian toothfish long-line fishery was the principal reason for the observed populationincreases at Diego Ramirez and Ildefonso.
4.5. Chilean system versus Spanish system
The open-ended configuration of the secondary lines (not joinedby a continuous length of hook line; see Fig. 3) and the closeproximity of weights to hooks with the Chilean system increasedfivefold the sink rates to 15 m depth compared to Spanish systemgear (means: 0.15 m/s versus 0.8 m/s; Robertson et al., 2008b). Thisdepth exceeds the known maximum dive depth (4.5 m) ofblack-browed albatrosses (Prince et al., 1994) which at 6 knots(3.1 m/s) vessel setting speed would be reached just 17 m asternand still within the zone of turbulent and aerated water (reducesbait visibility) from the propeller. These characteristics preventedthe incidental capture of albatrosses and petrels (see Morenoet al., 2008; Goetz et al., 2011), including white-chinned petrels(Procellaria aequinoctialis) during line setting in the absence of con-ventional seabird deterrents (Moreno et al., 2003). White-chinnedpetrels can dive �13 m deep (Huin, 1994) and are one of the mostdifficult species to deter – and most commonly caught – in demer-sal longline fisheries (e.g., Delord et al., 2005). The Chilean systemshould be equally effective in preventing captures during line haul-ing because hooks are enveloped in the net sleeves, protectingthem from attack by seabirds.
4.6. Comparison with other sites
As mentioned above nearly all the world’s black-browed alba-trosses breed at the Falkland islands/Islas Malvinas, South Georgiaand the islands of southern Chile (ACAP, 2010) with the FalklandIslands holding �70% of global numbers (Wolfaardt, 2013). TheFalklands population decrease in the 1980s and 1990s but is cur-rently increasing at about 4%/year (Catry et al., 2011; Wolfaardt,2013). In contrast, the South Georgia population has been decreas-ing since the 1970s (Croxall et al., 1997; Poncet et al., 2006) andcontinues to decrease (R. Phillips, personal communication). Birdsfrom the Falklands and South Georgia range, for the most part, indifferent areas of the south Atlantic region (Birdlife International,2006) and different threat levels in fisheries encountered is a likelyreason for the contrasting population trends. The population ofgrey-headed albatrosses at South Georgia is decreasing at a similarrate to black-browed albatrosses (Poncet et al., 2006). Birds fromthe South Georgia range widely in the Southern Ocean and sufferhigh levels of juvenile mortality (Croxall et al., 1997), most likelyin pelagic longline fisheries (Prince et al., 1997; see Azócar et al.,2013). Given their preference for oceanic frontal zones, also fa-voured by fishers, the continued decrease in grey-headed albatrossnumbers at South Georgia suggests the no-use of effective mitiga-tion measures by tuna and swordfish vessels operating on the highseas. Judging by the contrasting population trends it seems theat-sea risks are greater for grey-headed albatrosses from SouthGeorgia than from Diego Ramirez. The same can be said aboutblack-browed albatrosses.
4.7. Future priorities
The improved situation for the black-browed albatrosses inChile depends on the continued use of the Chilean system in theindustrial toothfish fishery and the adoption of effective seabirddeterrent measures in all three fleets of the industrial trawl fishery.Given the rapid increase in black-browed albatross numbers we
recommend the population surveys be repeated at no more thanfive-year intervals until such time as the trends appear to stabilise.Continued growth could lead to the colonisation of new breedingsites which should be located and surveyed at an appropriate timein the future. Islas Diego de Almagro, Islote Evangelistas and IsloteLeonard (not re-surveyed in 2011 and 2012) should be surveyedagain for evidence of growth and possible colonisation of new sites.Further tracking studies are needed, especially of juvenile alba-trosses (both species) during maiden flights and adults duringnon-breeding seasons and non-breeding years. Demographic stud-ies of both species of albatrosses are also required at one breedingsite at least (Diego Ramirez). The demographic studies will providemuch-needed sensitivity in assessments of population trends andthe tracking studies will provide valuable insights into unknownaspects of foraging ecology, identify fisheries-related threats tosurvival and highlight the need for conservation actions by rele-vant fisheries management organisations.
Acknowledgements
We are grateful to the Armada de Chile for transport to andfrom Diego Ramirez and for providing logistical support on the is-land during the albatross tracking studies. We are grateful to VictorRaimilla for assistance with the field work in 2011 and to Jose Oje-da for help with the fleet descriptions of the trawl fishery. Wethank Roger Wallis and Ben Wallis for transport and logisticalassistance with the projects on Diego de Almagro, Ildefonso andDiego Ramirez, David Smith for assistance preparing Figs. 5 and 6and Ben Raymond for providing the estimates of surface chloro-phyll-a distribution. The aircraft overflights were conducted withAerovias DAP, Pty Ltd, Chile. In 2011 the overflights were fundedby the GAP Planeterra Foundation, Toronto, Canada and in 2011by the Munha Broadcasting Corporation, Republic of South Korea,in collaboration with the Foundation for Antarctic Research. Wethank two reviewers for suggesting improvements to themanuscript.
Appendix A. Parameters for the model SEABIRD
A.1. General
Due to the absence of relevant information on the demographicparameters of black-browed albatrosses in Chile, information forthe stage-structured model was drawn from Rolland et al. (2009)for black-browed albatrosses at Illes Kerguelen in the southern In-dian Ocean. The life stages modelled were pre-breeders aged 1–14 years, first-time breeders, experienced breeders, andnon-breeding adults that had bred in a previous year. The resultsobtained are indicative only because there are only two populationsurvey data points available to calibrate the model. We used a judi-cious choice of only one parameter from the full set of 19 param-eters described in Rolland et al. (2009) plus the initial populationsize, required by SEABIRD. The parameters estimated were the ini-tial population size in 1988, N0, the year before the first availableeffort data for the Patagonian toothfish longline fishery (seeFig. 2), and the survival probability of experienced adult breeders(EB), SEB, where survival excludes the effect of fishery bycatch mor-tality. The other parameter values taken from Table 1 of Rollandet al. (2009) were transition probabilities (W’s) and survival prob-abilities of non-breeders and first-time breeders (excluding fisherybycatch mortality). Age at first breeding was taken as in Fig. 1 ofRolland et al. (2009) to be 5 years. The number of recruits wascalculated as 0.5 � N(EB) � breeding success (EB) � S0 + 0.5� N(1B) � breeding success (1B) � S0, where N(.) is the number of
G. Robertson et al. / Biological Conservation 169 (2014) 319–333 331
birds in a stage, breeding success is the proportion of pairs thatproduce and egg in that year, and S0 is the survival probability ofjuveniles from egg to age 1. Breeding success (.) � S0 was collapsedinto a single term by specifying S0 as 1.0 and breeding success forboth EB and 1B as 0.75 following Rolland et al. (2009). The immi-gration rate for EB’s with the mean value given in Table 1 ofRolland et al. (2009) of 0.044 was set to zero in our model. SEABIRDincludes fishery bycatch mortality as separate to the natural mor-tality implied by survival probabilities. The ages (adults versusjuveniles) of a sample of 220 black-browed albatrosses killed in2002 in the Patagonian toothfish longline fishery indicated 97.3%were adult birds (J. Arata, unpublished data). Hence, fishing selec-tivity in SEABIRD was set to apply only, and equally, to adults (i.e.1Bs, EBs, NBs). Pre-breeder survival, SPB, was applied to ages 1–5with a value of 0.91 combined with S0 of 1.0 and survival of first-time breeders (i.e. this component of the age 6–14 populations)which was also set to 0.91. Using the equivalent of the Rollandet al. (2009) values of survival from age 0 (i.e. S0) to age 4, by set-ting S0 to 1.0 and setting a separate juvenile survival for each agefrom 1 to 4 in SEABIRD to the value that gave final surviving pro-portion at age 4 of 0.281, it was found that census estimates thatwere close to those observed could not be obtained. In fact, the ob-served values were substantially under-estimated even with N0
reaching its upper bound of 200,000. In addition, the annualgrowth rate prior to 1988 was estimated to be negative at approx-imately �9%. Therefore, the survival probabilities prior to age 6were those described above and not the corresponding values fromRolland et al. (2009).
A.2. Justification of SEB
The observed population increase between 2002 and 2011could only be accurately predicted by the baseline model withSEB set at 0.992. This estimate is considerably higher than thatfor black-browed albatrosses at South Georgia prior to the startof the local Patagonian toothfish longline fishery (0.957; Arnoldet al., 2006). Alternative approaches involving fixing SEB to 0.957and estimating other combinations of annual breeding success pro-portions for both experienced breeders and first-time breeders,estimation of transition probabilities between non-breeders (thatmay or may not have attempted breeding in previous years),experienced breeders and first-time breeders failed to accuratelypredict the population increase. It was important that the censuscounts were accurately predicted by the baseline model so thaterrors in these predictions did not limit the ability to reject thehypothesis that the reduction in bycatch to zero from 2008 on-wards was substantially responsible for the observed populationincrease. Over a three year period an SEB of 0.992 predicts that2.4% of experienced breeders would die of causes unrelated tointeractions with fisheries compared to 12.4% for an SEB of 0.957.Whether the higher estimate for the Chilean population is biolog-ically plausible is not known (the extent to which the estimate ofSEB for the South Georgia birds reflects mortality in fisheriesencountered on migrations from the island is also not known).However, even if the estimate is unrealistically high it does notundermine the usefulness of the model, the main point of whichwas to investigate the circumstantial evidence regarding the prin-cipal reason for the observed population increase. Comparison ofthe predicted population decrease associated with bycatch sce-nario 1 (loss of 2000 birds/year), the small increase associated withbycatch scenario 2 (loss of 1000 birds/year) and the 5.9% increasefrom 2007 predicted by the baseline model (similar to the 6%increase at Ildefonso from 2011 to 2012) infers that bycatch inthe Spanish system longline fishery for Patagonian toothfish hada substantial negative impact on population growth despite thehigh estimated SEB.
Appendix B. Separating the effect of bycatch from naturaldemographic parameters
The figure shows the trend in Spanish system hooking effortand estimated number of black-browed albatrosses assumed killedin the Patagonian toothfish longline fishery. The trajectories arepredicted from the baseline model from 1989 to 2007 and bycatchscenarios 1 and 2 from 2008 to 2011. See Section 2 for explanationof bycatch scenarios.
Appendix C
C.1. Foraging characteristics of black-browed albatrosses
General pattern in foraging intensity kernel distributions (5-de-gree grid squares) for black-browed albatrosses from Diego Ramir-ez in the incubation, brood/guard and post-guard stages of thebreeding cycle. Only data for Diego Ramirez are shown becausethis was the only breeding site where black-browed albatrosseswere tracked during the three stages of the breeding cycle. Forag-ing intensity legend as for Figs. 5 and 6.

332 G. Robertson et al. / Biological Conservation 169 (2014) 319–333
C.2. Foraging characteristics of grey-headed albatrosses
General pattern in foraging intensity kernel distributions (5-de-gree grid squares) for grey-headed albatrosses from Diego Ramirezand Ildefonso (one bird only) in the incubation, brood/guard andpost-guard stages of the breeding cycle. Foraging intensity legendas for Figs. 5 and 6.
References
ACAP (Agreement on the Conservation of Albatrosses and Petrels), 2010. ACAPSpecies Assessment: Black-browed Albatross Thalassarche melanophrys. <http://www.acap.aq> (Downloaded 28.06.13).
Agnew, D.J., Black, A.D., Croxall, J.P., Parkes, G.B., 2000. Experimental evaluation ofthe effectiveness of weighting regimes in reducing seabird by-catch in thelongline toothfish fishery around South Georgia. CCAMLR Sci. 7, 119–131.
Aguayo, A., Acevedo, J., Acuña, P., 2003. Nuevo sitio de anidamiento del albatros cejanegra, Diomedea melanophris Temmink 1828, en el Seno Almirantazgo, Tierradel Fuego, Chile. Anal. Inst. Patagonia (Chile) 31, 91–96.
Anderson, A., Small, C., Croxall, J., Dunn, E., Sullivan, B., Yates, O., Black, A., 2011.Global seabird bycatch in longline fisheries. Endang. Species Res. 14, 91–106.
Arana, P.M., Vega, R., 1999. Exploratory fishing for Dissostichus spp. in the Antarcticregion (subareas 48.1, 48.2 and 88.3). CCAMLR Sci. 6, 1–17.
Arata, J., Robertson, G., Valencia, J., Lawton, K., 2003. The Evangelistas Islets, Chile: anew breeding site for black-browed albatrosses. Polar Biol. 26, 687–690.
Arata, J., Xavier, J., 2003. The diet of black-browed albatrosses at the Diego Ramirezislands, Chile. Polar Biol. 26, 638–647.
Arata, J.A., 2004. Análisis comparative de la ecología trófica y biologia reproductivede los albatross de ceja negra (Thalassarche melanophrys (Temminck, 1839)) ycabeza gris (T. chrysostoma (Foster, 1785)) en Isla Gonzalo, Archipiélago DiegoRamírez. Tesis Doctoral, Universidad Austral de Chile, pp 152.
Arata, J., Robertson, G., Valencia, J., Xavier, J.C., Moreno, C.A., 2004. Diet of Grey-headed Albatrosses at Diego Ramirez Islands, Chile: ecological implications.Antarct. Sci. 16, 263–275.
Arnold, J.M., Brault, S., Croxall, J.P., 2006. Albatross populations in peril: apopulation trajectory for black-browed albatrosses at South Georgia. Ecol.Appl. 16, 419–432.
ATF – Chile, 2013. Demersal Trawl Report (2011–2012). Comité Nacional ProDefensa de la Fauna y Flora; Subsecretaria de Pesca, Gobierno de Chile andSociedad Nacional de Pesca A.G.
Azócar, J.R., Moreno, C.A., Vega, R., 2008. Seguimiento del plan de acción nacionalaves marinas año 2008. Informe final, Mayo 2010, Proyecto FIP 2008–55, pp.310.
Azócar, J.R., Saavedra, J.C., Wiff, R., Barría, P.M., 2011. Interactions between seabirdsand the industrial longline fleet targeting swordfish in Chile. In: Fourth Meetingof the Seabird Bycatch Working Group of the Agreement on the Conservation ofAlbatrosses and Petrels, Guayaquil, Ecuador, 22–24 August 2011.
Azócar, J.R., Savedra-Nievas, J.C., Devia, D., 2013. Interacciones entre aves marinas yla flota palangrera industrial. In: Barría, P., Azócar, J., Bernal, C., González, A.,Mora, S., Acuicultura, F. (Eds.), 2012. Actividad 8: Recursos Pelágicos.Investigación situación de pesquerías de recursos altamente migratorios,2011. Informe Final. IFOP-SUBPESCA, Anexo 8, 143 p.
BirdLife International, 2004. Tracking Ocean Wanderers: The global distribution ofalbatrosses and petrels. Results from the Global Procellariiform TrackingWorkshop, 1–5 September, Gordon’s Bay, South Africa. BirdLife International,Cambridge, UK.
BirdLife International, 2006. Analysis of albatross and petrel distribution within theCCAMLR Convention Area: results from the Global Procellariiform TrackingDatabase. CCAMLR Sci. 13, 143–174.
BirdLife International, 2013. Seabird mortality in the Chilean demersal hake trawlfishery and the effect of bird-scaring lines as a mitigation measure. In: FifthMeeting of the Seabird Bycatch Working Group of the Agreement on theConservation of Albatrosses and Petrels, La Rochelle, France, 1–3 May 2013.
Catry, P., Forcada, J., Almeida, A., 2011. Demographic parameters of black-browedalbatrosses Thalassarche melanophris from the Falkland Islands. Polar Biol. 34,1221–1229.
Céspedes, R.L., Chong, V., Ojeda, L., Adasme, L., Muñoz, K., Hunt, L., Cid, M., Miranda,A., Villalón, 2011. Asesoría integral para la toma de decisiones en pesca yacuicultura 2010. Actividad 2: Peces Demersales: Seguimiento Demersal yAguas Profundas 2010. Sección III: Demersales Sur Austral. Informe Final,Instituto de Fomento Pesquero. 277 pp.
Croxall, J.P., Prince, P.A., Rothery, P., Wood, A.G., 1997. Population changes inalbatrosses at South Georgia. In: Robertson, G., Gales, R. (Eds.), Albatross Biologyand Conservation. Surrey Beatty & Sons, Chipping Norton, pp. 69–83.
Croxall, J.P., Silk, J.R.D., Phillips, R.A., Afanasyev, V., Briggs, D.R., 2005. Globalcircumnavigations: tracking year-round ranges of nonbreeding albatrosses.Science 307, 249–250.
Delord, K., Gasco, N., Weimerskirch, H., Barbraud, C., 2005. Seabird mortality in thePatagonian toothfish longline fishery around Crozet and Kerguelen islands,2001–2003. CCAMLR Sci. 12, 53–80.
FAO, 1999. International Plan of Action for Reducing Incidental Catch of Seabirds inLongline Fisheries. International Plan of Action for the Conservation andManagement of Sharks. International Plan of Action for the Management ofFishing Capacity. FAO, Rome, p. 26.
Favero, M., Blanco, B., Copello, S., Seco Pon, J.P., Patterlini, C., Mariano-Jelicich, R.,García, G., Paula Berón, M., 2013. Seabird bycatch in the Argentinean demersallongline fishery, 2001–2010. Endang. Species Res. 19, 187–199.
Feldman, G.C., McClain, C.R., 2010. In: Kuring, N., Bailey, S.W. (Eds.), Ocean ColorWeb, SeaWiFS Reprocessing. NASA Goddard Space Flight Center. <http://oceancolor.gsfc.nasa.gov/>.
Fu, D., Francis, R.I.C.C., 2008. SEABIRD Draft User Manual v1.00-2008/06/18. PaperWG-SAM-08/P3 of the Working Group on Statistics, Assessments, andModelling, CCAMLR Secretariat, Hobart, Australia.
Goetz, S., Laporta, M., Martinez Portela, J., Begoña Santos, M., Pierce, G.J., 2011.Experimental fishing with an ‘umbrella-and-stones’ system to reduceinteractions of sperm whales (Physeter macrocephalus) and seabirds withbottom-set longlines for Patagonian toothfish (Dissostichus eleginoides) in theSouthwest Atlantic. ICES J. Mar. Sci. 68, 228–238.
González-Zevallos, D., Yorio, P., 2006. Seabird use of discards and incidentalcaptures at the Argentinian hake trawl fishery in the Golfo San Jorge, Argentina.Mar. Ecol.-Prog. Ser. 316, 175–183.
González, A., Vega, R., Barbieri, M.A., Yáñez, E., 2012. Determinación de los factoresque inciden en la captura incidental de aves marinas en la flota palangrerapelágica chilena. Latin Am. J. Aquat. Resour. 40, 786–799.
Huin, N., 1994. Diving depths of white-chinned petrels. Condor 96, 1111–1113.IUCN, 2012. IUCN Red List of Threatened Species. Version 2012.1.
<www.iucn.redlist.org>.Jiménez, S., Abreu, M., Pons, M., Ortiz, M., Domingo, A., 2010. Assessing the impact
of the pelagic longline fishery on albatrosses and petrels in the southwestAtlantic. Aquat. Living Resour. 23, 49–64.
Lawton, K., Kirkwood, R., Robertson, G., Raymond, B., 2008. Preferred foraging areasof Heard Island albatrosses during chick raising and implications for themanagement of incidental mortality in fisheries. Aquat. Conserv. – Mar. Freshw.Ecosyst. 18, 309–320.
Lawton, K., Robertson, G., Valencia, J., Wienecke, B., Kirkwood, R., 2003. The status ofBlack-browed Albatrosses Thalassarche melanophrys at Diego de Almagro Island,Chile. Ibis 145, 502–505.
McConnell, B., Fedak, M., Burton, H.R., Engelhard, G.H., Reijnders, P.J.H., 2002.Movements and foraging areas of naive, recently weaned southern elephantseal pups. J. Anim. Ecol. 71, 65–78.
Marin, M., Oehler, D., 2007. Una nueva colonia de anidamiento para el albatros deceja negra (Thalassarche melanophrys) para Chile. Anal. Inst. Patagonia (Chile)35, 29–33.
Moreno, C.A., Rubilar, P.S., Marschoff, E., Benzaquen, L., 1996. Factors affecting theincidental mortality of seabirds in the Dissostuchus eleginoides fishery in thesouth-west Atlantic (subarea 48.3, 1995 season). CCAMLR Sci. 3, 79–91.

G. Robertson et al. / Biological Conservation 169 (2014) 319–333 333
Moreno, C.A., Hucke-Gaete, R., Arata, J., 2003. Interacción de la pesquería de bacalaode profundidad con mamíferos y aves marinas. Informe final del Proyecto FIP2001–31. Universidad Austral de Chile, pp. 199.
Moreno, C.A., Arata, J., 2004. Informe final del Proyecto FIP 2003–21. Plan de AcciónNacional de Chile para mitigar efectos de la pesca de palangre sobre AvesMarinas (PAN-AM). Final report, Project FIP 2003–21, pp. 132.
Moreno, C.A., Arata, J., 2006. Efectos de las pesquerías industriales de palangre sobreaves marinas y el camino a su mitigación en Chile. In: Saball, P., Arroyo, M.K.,Castilla, J.C., Estades, C., Ladrón de Guevara, J.M., Larraín, S., Moreno, C.A., Rivas,F., Rovira, J., Sánchez, A., Sierralta, L. (Eds.), Biodiversidad de Chile: Patrimonio ydesafíos. CONAMA-Chile, Santiago, Chile, pp. 462–467. ISBN: 978-956-8018-56-6.
Moreno, C.A., Arata, J., Rubilar, P., Hucke-Gaete, R., Robertson, G., 2006. Artisanallongline fisheries in Southern Chile: lessons to be learned to avoid incidentalseabird mortality. Biol. Conserv. 127, 27–36.
Moreno, C.A., Castro, R., Mujica, L.J., Reyes, P., 2008. Significant conservationbenefits obtained from the use of a new fishing gear in the Chilean PatagonianToothfish fishery. CCAMLR Sci. 15, 79–91.
Moreno, C.A., Robertson, G., 2008. ¿Cuántos albatros de ceja negra, Thalassarchemelanophrys (Temminck, 1828) anidan en Chile? Anal. Inst. Patagonia (Chile)36, 89–91.
Nel, D.C., Ryan, P.G., Watkins, B.P., 2002. Seabird mortality in the Patagoniantoothfish longline fishery around the Prince Edward Islands, 1996–2000.Antarct. Sci. 14, 151–161.
Otley, H., 2005. Seabird Mortality Associated with Patagonian Toothfish Longlinersin Falkland Island Waters during 2002/03 and 2003/04. Fisheries Department,Falkland Islands Government, Falkland Islands, pp. 45.
Oyarzún, C., Gacitúa, S., Araya, M., Cubillos, L., Galleguillos, R., Pino, C., Aedo, G.,Salamanca, M., Pedraza, M., Lamilla, J., 2003. Monitoreo de la pesqueríaartesanal de bacalao de profundidad entre la VIII y XI regiones. Final report,Project FIP 2001–16, Sub-secretary of fisheries, Chile. <www.fip.cl>.
Poncet, S., Robertson, G., Phillips, R.A., Lawton, K., Phalan, B., Trathan, P., Croxall, J.P.,2006. Status and distribution of wandering, black-browed and grey-headedalbatrosses at South Georgia. Polar Biol. 29, 772–781.
Prince, P.A., Huin, N., Weimerskirch, H., 1994. Diving depths of albatrosses. Antarct.Sci. 6, 353–354.
Prince, P.A., Croxall, J.P., Trathan, P.N., Wood, A.G., 1997. The pelagic distribution ofSouth Georgia albatrosses and their relationship with fisheries. In: Robertson,G., Gales, R. (Eds.), Albatross Biology and Conservation. Surrey Beatty & Sons,Chipping Norton, pp. 137–163.
Robertson, G., Gales, R., 1997. Albatross Biology and Conservation. Surrey Beatty &Sons, Chipping Norton.
Robertson, G., 2000. Effect of line sink rate on albatross mortality in the Patagoniantoothfish longline fishery. CCAMLR Sci., 133–150.
Robertson, G., Moreno, C.A., Lawton, K., Arata, J., Valencia, J., Kirkwood, R., 2007. Anestimate of the population sizes of black-browed (Thalassarche melanophrys)and grey-headed (T. chrysostoma) albatrosses at the Diego Ramirez Archipelago.Chile Emu 107, 239–244.
Robertson, G., Moreno, C.A., Lawton, K., Kirkwood, R., Valencia, J., 2008a.Comparison of census methods for black-browed albatrosses breeding at theIldefonso Archipelago, Chile. Polar Biol., 153–162.
Robertson, G., Moreno, C.A., Gutiérrez, E., Candy, S.G., Melvin, E.F., Seco Pon, J.P.,2008b. Line weights of constant mass (and sink rates) for Spanish-rigPatagonian toothfish longline vessels. CCAMLR Sci. 15, 93–106.
Robertson, G., Moreno, C.A., Crujeiras, J., Wienecke, B., Gandini, P., McPherson, G.,Seco Pon, J.P., 2008c. An experimental assessment of factors affecting the sink
rates of Spanish-rig longlines to minimise impacts on seabirds. Aquat. Conserv.:Mar. Freshw. Ecosyst. 17, S102–S121.
Rodhouse, P.G., Prince, P.A., Trathan, P.N., Hatfield, E.M.C., Watkins, J.L., Bone, D.G.,Murphy, E.J., White, M.G., 1996. Cephalopods and mesoscale oceanography atthe Antarctic Polar Front: satellite tracked predators locate pelagic trophicinteractions. Mar. Ecol. Prog. Ser. 136, 37–50.
Rolland, V., Nevoux, M., Barbraud, C., Weimerskirch, H., 2009. Respective impact ofclimate and fisheries on the growth of an albatross population. Ecol. Appl. 19,1336–1346.
Ryan, P.G., Phillips, R.A., Nel, D.C., Wood, A., 2007. Breeding frequency in Grey-headed Albatrosses Thalassarche chrysostoma. Ibis 149, 45–52.
SC-CCAMLR, 2006. Scientific Committee for the Conservation of Antarctic MarineLiving Resources, 2006. Report of the 25th Meeting of the Scientific Committee.CCAMLR, Hobart.
Small, C.J., 2005. Regional Fisheries Management Organisations: Their Duties andPerformance in Reducing Bycatch of Albatrosses and Other Species. BirdLifeInternational, Cambridge, UK, pp. 103.
Subsecretaría de Pesca, 2011. Cuota global anual de captura de merluza común(Merluccius gayi gayi), año 2012. Inf. Tec. (R. Pesq.) N� 117/2011, Subsecretaríade Pesca, Valparaíso, 46 pp.
Subsecretaría de Pesca, 2012a. Cuota global anual de captura de merluza del sur(Merluccius australis), año 2013. Inf. Tec. (R. Pesq.) N� 216/2012, Subsecretaríade Pesca, Valparaíso, 77 pp.
Subsecretaría de Pesca, 2012b. Cuota global anual de captura para las unidades depesquería de merluza de cola (Macruronus magellanicus), año 2013. Inf. Tec. (R.Pesq.) N� 205/2012, Subsecretaria de Pesca, Valparaíso, 59 pp.
Subsecretaría de Pesca, 2012c. Cuota global anual de captura de merluza de tresaletas, (Micromesistius australis), año 2013. Inf. Tec. (R. Pesq.) N� 202/2012,Subsecretaría de Pesca, Valparaíso, 37 pp.
Sullivan, B.J., Reid, T.A., Bugoni, L., 2006a. Seabird mortalilty on factory trawlers inthe Falkland Islands and beyond. Biol. Conserv. 131, 495–504.
Sullivan, B.J., Brickle, P., Reid, T.A., Bone, D.G., Middleton, A.J., 2006b. Mitigation ofseabird mortality on factory trawlers: trials of three devices to reduce warpcable strikes. Polar Biol. 29, 745–753.
Tickell, W.L.N., 2000. Albatrosses. Pica Press, Sussex, pp. 448.Thompson, K.R., Riddy, M.D., 1995. Utilization of offal and discards from ‘‘finfish’’
trawlers around the Falkland Islands by Black-browed Albatrosses Diomedeamelanophris. Ibis 137, 198–206.
Vega, R., Licandeo, R., 2009. The effect of American and Spanish longline systems ontarget and non-target species in the eastern South Pacific swordfish fishery.Fish. Res. 98, 22–32.
Verbyla, A.P., Cullis, B.R., Kenward, M.G., Welham, S.J., 1999. The analysis ofdesigned experiments and longitudinal data using smoothing splines (withdiscussion). Appl. Stat. 48, 269–311.
Weimerskirch, H., Capdeville, D., Duhamel, G., 2000. Factors affecting the numberand mortality of seabirds attending trawlers and longliners in the Kerguelenarea. Polar Biol. 23, 236–249.
Wolfaardt, A., 2013. An assessment of the population trends and conservationstatus of black-browed albatrosses in the Falkland Islands. In: First Meeting ofthe Population and Conservation Status Working Group of the Agreement onthe Conservation of Albatrosses and Petrels. La Rochelle, France, 29–30 April2013. PCSWG1 Doc 14.
Yeh, T.-M., Huang, H.-W., Dietrich, K.S., Melvin, E., 2012. Estimates of seabirdincidental catch by pelagic longline fisheries in the South Atlantic Ocean. Anim.Conserv. 16, 141–152.