charged residues are involved in membrane fusion mediated by a hydrophilic peptide located in...
TRANSCRIPT

Charged residues are involved in membrane fusion mediated by ahydrophilic peptide located in vesicular stomatitis virus G protein
FABIANA A. CARNEIRO1, GUY VANDENBUSSCHE2, MARIA A. JULIANO3,
LUIZ JULIANO3, JEAN-MARIE RUYSSCHAERT2, & ANDREA T. DA POIAN1
1Instituto de Bioquımica Medica, Programa de Biologia Molecular e Biotecnologia, Universidade Federal do Rio de Janeiro,
Rio de Janeiro, Brazil, 2Structure et Fonction des Membranes Biologiques, Centre de Biologie Structurale et de
Bioinformatique, Universite Libre de Bruxelles, Brussels, Belgium, and 3Departamento de Biofısica, Escola Paulista de
Medicina, UNIFESP, Sao Paulo, Brazil
(Received 11 October 2005; and in revised form 27 April 2006)
AbstractMembrane fusion is an essential step of the internalization process of the enveloped animal viruses. Vesicular stomatitis virus(VSV) infection is mediated by virus spike glycoprotein G, which induces membrane fusion at the acidic environment of theendosomal compartment. In a previous work, we identified a specific sequence in VSV G protein, comprising the residues145 to 164, directly involved in membrane interaction and fusion. Unlike fusion peptides from other viruses, this sequenceis very hydrophilic, containing six charged residues, but it was as efficient as the virus in catalyzing membrane fusion at pH6.0. Using a carboxyl-modifying agent, dicyclohexylcarbodiimide (DCCD), and several synthetic mutant peptides, wedemonstrated that the negative charges of peptide acidic residues, especially Asp153 and Glu158, participate in the formationof a hydrophobic domain at pH 6.0, which is necessary to the peptide-induced membrane fusion. The formation of thehydrophobic region and the membrane fusion itself were dependent on peptide concentration in a higher than linearfashion, suggesting the involvement of peptide oligomerization. His148 was also necessary to hydrophobicity and fusion,suggesting that peptide oligomerization occurs through intermolecular electrostatic interactions between the positively-charged His and a negatively-charged acidic residue of two peptide molecules. Oligomerization of hydrophilic peptidescreates a hydrophobic region that is essential for the interaction with the membrane that results in fusion.
Keywords: Membrane fusion, fusion peptide, vesicular stomatitis virus, dicyclohexylcarbodiimide, hydrophobicity
Introduction
Virus replication depends on the transfer of viral
genome and accessory proteins to the cytosol or to
the nucleus of a host cell. In the case of enveloped
virus, this entry process involves the fusion of the
virus envelope with the plasma or the endosomal
membranes of the host cell [1]. The membrane
fusion reaction is catalyzed by viral surface glyco-
proteins, which undergo conformational changes
triggered by either their interaction with a cellular
receptor or by the acidification of the endosomal pH.
Viral fusion glycoproteins contain a short sequence
directly involved in the interaction with the target
membrane during the fusion reaction, known as the
fusion peptide. Fusion peptides from several viruses
have been identified by mutagenesis experiments, in
which a single amino acid change abolished the
fusion activity of the glycoprotein. The sequence of
the fusion peptides is generally conserved within the
viral family, but not among different families.
Based on structural differences, the viral fusion
proteins were classified into two groups. Class I
fusion proteins form trimeric spikes predominantly
folded as a-helices with a hydrophobic fusion pep-
tide located at the N-terminal end of the protein [2].
After binding to a cellular receptor or on exposure to
low pH, the protein forms an extended conforma-
tion and the fusion peptide inserts into the target
membrane. The post-fusion conformation is a hair-
pin-like structure in which the fusion peptide and
the membrane anchor are at the same end [2]. In
class II fusion proteins three domains folded largely
on b-sheets are arranged in a continuous protein
lattice formed by dimers [3]. The fusion peptide is
an internal loop between two b-strands, buried in
the dimer interface. The determination of the post-
Correspondence: Andrea T. Da Poian, Instituto de Bioquımica Medica, Universidade Federal do Rio de Janeiro, Av. Bauhinia, 400,
Bl. H, s 22, Rio de Janeiro, RJ 21941-590, Brazil. Tel: 55 21 22706264. Fax: 55 21 22708647. E-mail: [email protected]
Molecular Membrane Biology, September�October 2006; 23(5): 396�406
ISSN 0968-7688 print/ISSN 1464-5203 online # 2006 Informa UK Ltd
DOI: 10.1080/09687860600780892

fusion structure of class II fusion proteins revealed a
surprising convergence of the class I and class II
fusion mechanisms [4]. The acidic pH of the
endosome induces a disassembly of envelope pro-
teins dimers, which rearrange in trimers with the
fusion peptide loops clustered at one end of an
elongated molecule.
The viruses that belong to the Rhabdoviridae
family are widely distributed in nature and their
hosts range from vertebrates and invertebrates
animals to many species of plants. All the rhabdo-
viruses present a bullet-shaped structure that is
formed by two major components: the nucleocapsid
and the envelope. The envelope is a lipid bilayer
derived from the host cell containing trimeric
transmembrane spikes composed by the viral surface
glycoprotein G. Vesicular stomatitis virus (VSV) is
the prototype of the Rhabdoviridae family. VSV G
protein is involved both in the cell recognition and in
the membrane fusion reaction, which occurs in the
acidic environment of the endosome after virus
internalization. Mutagenesis experiments have
shown that substitution of conserved Gly, Pro, or
Asp located in the region between amino acids 117
and 137 either abolished the fusion ability of G
protein or shifted the optimum pH of fusion [5�7].
This led the authors to propose that this segment
would be the VSV G protein putative fusion peptide,
although direct evidence that this particular region
interacts with the target membrane is still lacking.
Studying the requirement for PS in the target
membrane [8] and the crucial role of G protein
His residues for VSV fusion [9], we found another
candidate to be the VSV fusion peptide, the PS
binding site of the rhabdoviruses G protein [10].
This segment was firstly characterized for viral
hemorrhagic septicemia virus (VHSV), a rhabdo-
virus of salmonids [11,12], and then was found
among all rhabdoviruses [10]. For VSV, it corre-
sponds to the sequence between amino acid 145 and
164 (sequence VTPHHVLVDEYTGEWVDSQF).
We have demonstrated that a synthetic peptide
corresponding to this sequence was as efficient as
the whole virus in catalyzing fusion, whereas the
putative fusion peptide failed to induce fusion [9].
Moreover, as found for VSV-induced membrane
fusion, the fusion induced by the peptide was
dependent on pH and on the presence of PS in the
target membrane.
An interesting feature of VSV peptide145�164 is
that it contains four acidic amino acid residues, two
aspartic and two glutamic acids, which would be
negatively charged at the fusion pH. Using the
carboxyl-modifying agent dicyclohexylcarbodiimide
(DCCD) and several mutant peptides, we showed in
this work that the acidic residues, especially Asp153
and Glu158, participate in the formation of a hydro-
phobic domain at pH 6.0, which is necessary to the
peptide-induced membrane fusion. In addition, the
formation of this hydrophobic region as well as the
membrane fusion itself, are highly dependent on
peptide concentration, suggesting the involvement of
peptide oligomerization.
Materials and methods
Chemicals
Phosphatidylserine (PS) and phosphotidylcholine
(PC) from bovine brain, and dicyclohexylcarbodii-
mide (DCCD) were purchased from Sigma Chemi-
cal Co., St Louis, MO, USA. N-(lissamine
Rhodamine B sulfonyl) phosphatidylethanolamine
(Rh-PE), N-(7- nitro-2,1,3-benzoxadiazol-4-yl)
phosphatidylethanolamine (NBD-PE) and 8-anili-
nonaphthalene-1-sulfonate (ANS) were purchased
from Molecular Probes Inc., Eugene, OR, USA. All
other reagents were of analytical grade.
Peptides synthesis. VSV peptide145�164 (sequence
VTPHHVLVDEYTGEWVDSQF) and histidine
and acid residues mutants were synthesized by solid
phase using the Fmoc methodology and all protected
amino acids were purchased from Calbiochem-
Novabiochem (San Diego, CA) or from Neosystem
(Strasbourg, France). The syntheses were done in an
automated bench-top simultaneous multiple solid-
phase peptide synthesizer (PSSM 8 system from
Shimadzu, Tokyo, Japan). The final deprotected
peptides were purified by semipreparative HPLC
using an Econosil C-18 column (10 mm, 22.5�/250
mm) and a two-solvent system: (A) trifluoroacetic
acid/H2O (1:1000, v/v) and (B) trifluoroacetic acid/
acetonitrile/H2O (1:900:100, v/v/v). The column
was eluted at a flow rate of 5 ml.min�1 with a 10
or 30 to 50 or 60% gradient of solvent B over 30 or
45 min. Analytical HPLC was performed using a
binary HPLC system from Shimadzu with a SPD-
10AV Shimadzu UV/vis detector, coupled to an
Ultrasphere C-18 column (5 mm, 4.6�/150 mm),
which was eluted with solvent systems A1 (H3PO4/
H2O, 1:1000, v/v) and B1 (acetonitrile/H2O/H3PO4,
900:100:1, v/v/v) at a flow rate of 1.7 ml.min�1 and
a 10�80% gradient of B1 over 15 min. The
HPLC column eluted materials were monitored
by their absorbance at 220 nm. The molecular
mass and purity of synthesized peptides were
checked by MALDI-TOF mass spectrometry (Tof-
Spec-E, Micromass) and/or peptide sequencing
using a protein sequencer PPSQ-23 (Shimadzu
Tokyo, Japan).
G protein charged residues in VSV-induced membrane fusion 397

Peptide modification with DCCD. A solution of
dicyclohexylcarbodiimide (DCCD) was freshly pre-
pared by dilution of the reagent in ethanol.
Peptide145�164 was diluted in 20 mM MES, 30
mM Tris buffer, pH 6.0 and incubated for 1 h at
room temperature with DCCD, in a molar ratio of
DCCD/peptide of 40.
Preparation of liposomes. PC and PS at a molar ratio
of 1:3 were dissolved in chloroform and evaporated
under nitrogen. The lipid film formed was resus-
pended in 20 mM MES, 30 mM Tris buffer (pH
indicated in the Figure legends) at a final concentra-
tion of 1 mM. The suspension was vortexed
vigorously for 5 min. Small unilamellar vesicles
(SUV) were obtained by sonicating the turbid
suspension using a Branson Sonifier (Sonic Power
Company, Danbury, CT) equipped with a titanium
microtip probe. Sonication was performed in an ice
bath, alternating cycles of 30 sec at 20% full power,
with 60-sec resting intervals until a transparent
solution was obtained (approx. 10 cycles). For
fusion assays, 1 mol% of each Rh-PE and NBD-
PE was incorporated in the lipid films.
Liposome fusion assay. Liposomes composed of PC-
PS (1:3) containing equal amounts of unlabeled
vesicles and vesicles labeled with Rh-PE and NBD-
PE were prepared in 20 mM MES, 30 mM Tris
buffer (pH indicated in the Figure legends), at a final
phospholipid concentration of 0.1 mM. The fusion
reaction was initiated by addition of the peptide.
Fusion was followed by the resonance energy
transfer assay as described in Struck et al. (1981)
[13]. The samples were excited at 470 nm and the
fluorescence intensity was collected at 530 nm, using
a Hitachi F-4500 Fluorescence Spectrophotometer.
Mass spectrometry. A stock solution of VSV
peptide145�164 in ethanol was diluted at different
concentrations (as indicated in the Figure legends)
in 20 mM MES, 30 mM Tris buffer pH 6.0, and
incubated with DCCD for 1 h at room temperature.
Before analysis by mass spectrometry, the buffer was
removed by loading the peptide on a ZipTip C18
(Millipore, Billerica, USA). The peptide was washed
with TFA 0.1% (v/v) and eluted in 5 ml acetonitrile
50%/formic acid 1% (v/v). The samples were loaded
into a nanoflow capillary (Proxeon, Odense, Den-
mark). ESI mass spectra were acquired on a quad-
rupole time-of-flight instrument (Q-Tof Ultima �Micromass/Waters, Manchester, UK) operating in
the positive ion mode, equipped with a Z-spray
nanoelectrospray source. Capillary voltages of
1.1�1.5 kV and cone voltage of 50 V typically were
used. The source temperature was held at 808C. The
spectra represent the average of 1 sec scans. Data
acquisition was performed with a MassLynx 4.0
system. The exact mass of the peptide was deter-
mined after processing of the spectra by the software
Transform (Micromass/Waters, Manchester, UK).
For MS/MS studies, the quadrupole was used to
select the charged parent ion, which was subse-
quently fragmented in a hexapole collision cell using
argon as collision gas and an appropriate collision
energy. MS/MS data were processed by a maximum
entropy data enhancement program, MaxEnt 3
(Micromass/Waters, Manchester, UK). Amino acid
sequence was semi-automatically deduced with the
peptide sequencing program, PepSeq.
Infrared spectroscopy. ATR-FTIR (Attenuated Total
Reflection Fourier Transform infrared spectroscopy)
spectra were recorded on a Bruker IFS-55 FTIR
spectrophotometer (Bruker, Karlsruhe, Germany)
equipped with a liquid nitrogen-cooled mercury-
cadmium-telluride detector [14�16]. The spectro-
photometer was continuously purged with dried air.
The internal reflection element (ATR) was a germa-
nium plate (50�/20�/2 mm) with an aperture angle
of 458, yielding 25 internal reflections. Samples were
deposited on the germanium element. Films were
formed by slowly evaporating the sample on one side
of the ATR plate under a stream of nitrogen.
Samples were rehydrated by flushing D2O-saturated
N2 for 30 min at room temperature.
Results
Effects of Glu and/or Asp modification on peptide-
induced membrane fusion
We have previously shown that fusion induced by
peptide145�164 depends on the protonation of its
His148 and His149 residues, which confer positive
charges to the peptide at pH below 6.0 [12].
However, the peptide also contains four negatively
charged amino acid residues (Asp153, Glu154, Glu158
and Asp161) and no other positively charged residue.
To investigate the role of these negative charges in
peptide-induced fusion, we used DCCD, a com-
pound that labels Asp and Glu residues located in
hydrophobic environments [17]. Peptide-induced
membrane fusion was quantified by NBD-PE/Rh-
PE energy transfer assay (Figure 1a). DCCD label-
ing completely abolished the ability of the peptide to
mediate membrane fusion. To confirm the impor-
tance of the negative charges of these residues,
fusion was assayed at pH 4.0, a pH close to the
pKa of the carboxyl groups of lateral chain of the
acidic residues (Figure 1b). Fusion did not occur at
pH 4.0, even when the peptide was not incubated
398 F. A. Carneiro et al.
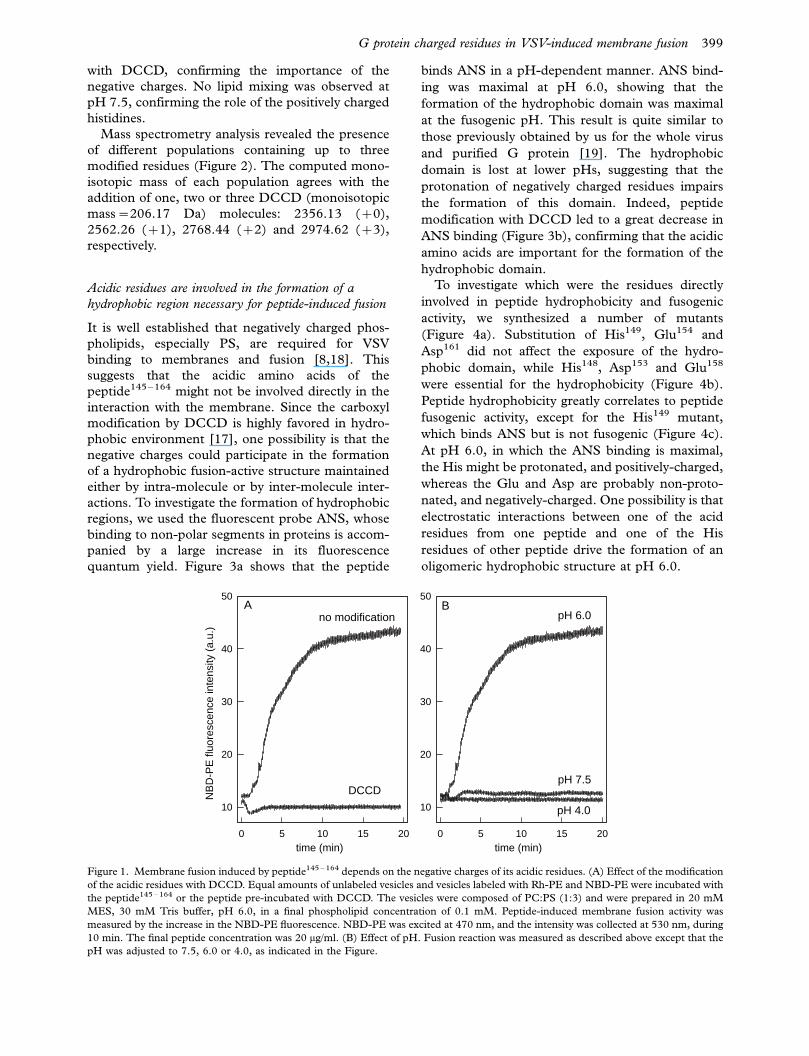
with DCCD, confirming the importance of the
negative charges. No lipid mixing was observed at
pH 7.5, confirming the role of the positively charged
histidines.
Mass spectrometry analysis revealed the presence
of different populations containing up to three
modified residues (Figure 2). The computed mono-
isotopic mass of each population agrees with the
addition of one, two or three DCCD (monoisotopic
mass�/206.17 Da) molecules: 2356.13 (�/0),
2562.26 (�/1), 2768.44 (�/2) and 2974.62 (�/3),
respectively.
Acidic residues are involved in the formation of a
hydrophobic region necessary for peptide-induced fusion
It is well established that negatively charged phos-
pholipids, especially PS, are required for VSV
binding to membranes and fusion [8,18]. This
suggests that the acidic amino acids of the
peptide145�164 might not be involved directly in the
interaction with the membrane. Since the carboxyl
modification by DCCD is highly favored in hydro-
phobic environment [17], one possibility is that the
negative charges could participate in the formation
of a hydrophobic fusion-active structure maintained
either by intra-molecule or by inter-molecule inter-
actions. To investigate the formation of hydrophobic
regions, we used the fluorescent probe ANS, whose
binding to non-polar segments in proteins is accom-
panied by a large increase in its fluorescence
quantum yield. Figure 3a shows that the peptide
binds ANS in a pH-dependent manner. ANS bind-
ing was maximal at pH 6.0, showing that the
formation of the hydrophobic domain was maximal
at the fusogenic pH. This result is quite similar to
those previously obtained by us for the whole virus
and purified G protein [19]. The hydrophobic
domain is lost at lower pHs, suggesting that the
protonation of negatively charged residues impairs
the formation of this domain. Indeed, peptide
modification with DCCD led to a great decrease in
ANS binding (Figure 3b), confirming that the acidic
amino acids are important for the formation of the
hydrophobic domain.
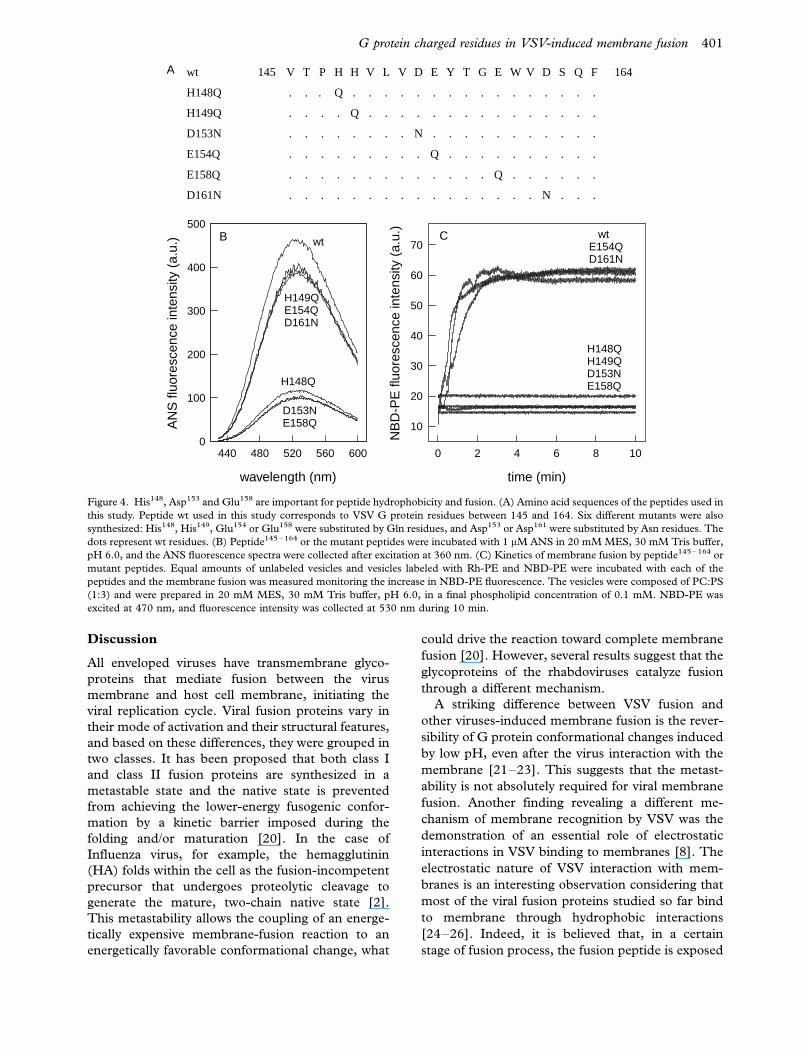
To investigate which were the residues directly
involved in peptide hydrophobicity and fusogenic
activity, we synthesized a number of mutants
(Figure 4a). Substitution of His149, Glu154 and
Asp161 did not affect the exposure of the hydro-
phobic domain, while His148, Asp153 and Glu158
were essential for the hydrophobicity (Figure 4b).
Peptide hydrophobicity greatly correlates to peptide
fusogenic activity, except for the His149 mutant,
which binds ANS but is not fusogenic (Figure 4c).
At pH 6.0, in which the ANS binding is maximal,
the His might be protonated, and positively-charged,
whereas the Glu and Asp are probably non-proto-
nated, and negatively-charged. One possibility is that
electrostatic interactions between one of the acid
residues from one peptide and one of the His
residues of other peptide drive the formation of an
oligomeric hydrophobic structure at pH 6.0.
time (min)0 5 10 15 20
NB
D-P
E fl
uore
scen
ce in
tens
ity (
a.u.
)
10
20
30
40
50
DCCD
no modification
time (min)0 5 10 15 20
10
20
30
40
50
pH 6.0
pH 7.5
pH 4.0
A B
Figure 1. Membrane fusion induced by peptide145 �164 depends on the negative charges of its acidic residues. (A) Effect of the modification
of the acidic residues with DCCD. Equal amounts of unlabeled vesicles and vesicles labeled with Rh-PE and NBD-PE were incubated with
the peptide145 �164 or the peptide pre-incubated with DCCD. The vesicles were composed of PC:PS (1:3) and were prepared in 20 mM
MES, 30 mM Tris buffer, pH 6.0, in a final phospholipid concentration of 0.1 mM. Peptide-induced membrane fusion activity was
measured by the increase in the NBD-PE fluorescence. NBD-PE was excited at 470 nm, and the intensity was collected at 530 nm, during
10 min. The final peptide concentration was 20 mg/ml. (B) Effect of pH. Fusion reaction was measured as described above except that the
pH was adjusted to 7.5, 6.0 or 4.0, as indicated in the Figure.
G protein charged residues in VSV-induced membrane fusion 399

Peptide oligomerization
To evaluate the requirement of peptide oligomeriza-
tion for hydrophobicity and fusion, we checked the
dependence on peptide concentration (Figure 5).
The extent of lipid mixing varied with the peptide
concentration in a higher than linear fashion (Figure
5a and 5b), supporting the hypothesis that peptide
promotes membrane fusion as oligomers. A drastic
increase in ANS binding occurred at the peptide
concentration range required for fusion (Figure 5c),
suggesting that peptide hydrophobicity is a conse-
quence of its oligomerization. The threshold of
aggregation was neither altered for the fusogenic
mutants E154Q and D161N nor for the non-
fusogenic H149Q mutant (Figure 5c).
Analyses by mass spectrometry indicated that
DCCD labels the peptide only at concentrations
that favor oligomerization (Figure 6). Since DCCD
labeling occurs in hydrophobic environments, this is
another evidence that oligomerization contributes to
the formation of a hydrophobic region that is
probably necessary for the interaction with the
membrane and consequently, for fusion.
We analysed peptide secondary structure at pH
7.5 and 6.0 by infrared spectroscopy (Figure 7). The
1600�1700 cm�1 region (amide I) corresponding to
the C�/O stretching vibration is the most sensitive to
the secondary structure of the proteins and each
secondary structure absorbs at different wave-
lengths. The peak at 1650 cm�1 is characteristic of
a peptide folded in an a-structure [14]. More
importantly, the fold is not affected by pH (Figure
7a), suggesting that the hydrophobicity is a conse-
quence of reorganization of the oligomeric state of
the peptides rather than due to changes in their
secondary structure. IR spectra of the non-fusogenic
mutants did not reveal any structural changes as
compared with wild type spectrum (Figure 7b),
excluding the possibility that the mutations disrupt
the structure and not only the generation of the
hydrophobicity.
pH2345678
AN
S fl
uore
scen
ce in
tens
ity (
a.u.
)
200
250
300
350
400
450B
wavelength (nm)440 480 520 560 600
0
100
200
300
400
500A
DCCD
no modification
Figure 3. Acidic residues are involved in the formation of a hydrophobic region. (A) ANS binding as a function of pH. Peptide145 �164 was
diluted to a final concentration of 20 mg/ml in 20 mM MES, 30 mM Tris buffer, pH as indicated in the Figure, and incubated with 1 mM
ANS. ANS was excited at 360 nm and the emission was collected at 492 nm. (B) Peptide145 �164 without modification (*) or modified with
DCCD (. . .) were incubated with 1 mM ANS in 20 mM MES, 30 mM Tris buffer, pH 6.0, and the ANS fluorescence spectra were collected
after excitation at 360 nm.
Figure 2. Labeling of the peptide145 �164 with DCCD. ESI-MS
spectra were recorded to monitor the addition of DCCD groups
to the Asp and Glu residues of the peptide145 �164. The peptide
(20 mg/ml) was incubated for 1 h with DCCD in 20 mM MES, 30
mM Tris buffer, pH 6.0. Before analysis, the buffer was removed
on ZipTip C18 and the peptide was solubilized in 50% acetoni-
trile/ 1% formic acid (v/v). The different populations observed
(arrows) correspond to the addition of 1, 2 and 3 DCCD
molecules, respectively. The accuracy of mass measurement was
in the order of 8 ppm.
400 F. A. Carneiro et al.
Discussion
All enveloped viruses have transmembrane glyco-
proteins that mediate fusion between the virus
membrane and host cell membrane, initiating the
viral replication cycle. Viral fusion proteins vary in
their mode of activation and their structural features,
and based on these differences, they were grouped in
two classes. It has been proposed that both class I
and class II fusion proteins are synthesized in a
metastable state and the native state is prevented
from achieving the lower-energy fusogenic confor-
mation by a kinetic barrier imposed during the
folding and/or maturation [20]. In the case of
Influenza virus, for example, the hemagglutinin
(HA) folds within the cell as the fusion-incompetent
precursor that undergoes proteolytic cleavage to
generate the mature, two-chain native state [2].
This metastability allows the coupling of an energe-
tically expensive membrane-fusion reaction to an
energetically favorable conformational change, what
could drive the reaction toward complete membrane
fusion [20]. However, several results suggest that the
glycoproteins of the rhabdoviruses catalyze fusion
through a different mechanism.
A striking difference between VSV fusion and
other viruses-induced membrane fusion is the rever-
sibility of G protein conformational changes induced
by low pH, even after the virus interaction with the
membrane [21�23]. This suggests that the metast-
ability is not absolutely required for viral membrane
fusion. Another finding revealing a different me-
chanism of membrane recognition by VSV was the
demonstration of an essential role of electrostatic
interactions in VSV binding to membranes [8]. The
electrostatic nature of VSV interaction with mem-
branes is an interesting observation considering that
most of the viral fusion proteins studied so far bind
to membrane through hydrophobic interactions
[24�26]. Indeed, it is believed that, in a certain
stage of fusion process, the fusion peptide is exposed
wt 145 V T P H H V L V D E Y T G E W V D S Q F 164
H148Q . . . Q . . . . . . . . . . . . . . . .
H149Q . . . . Q . . . . . . . . . . . . . . .
D153N . . . . . . . . N . . . . . . . . . . .
E154Q . . . . . . . . . Q . . . . . . . . . .
E158Q . . . . . . . . . . . . . Q . . . . . .
D161N . . . . . . . . . . . . . . . . N . . .
A
wavelength (nm)
440 480 520 560 600
AN
S fl
uore
scen
ce in
tens
ity (
a.u.
)
0
100
200
300
400
500
wt
H149QE154QD161N
D153NE158Q
H148Q
B C
time (min)
0 2 4 6 8 10
NB
D-P
E fl
uore
scen
ce in
tens
ity (
a.u.
)
10
20
30
40
50
60
70 wtE154QD161N
H148QH149QD153NE158Q
Figure 4. His148, Asp153 and Glu158 are important for peptide hydrophobicity and fusion. (A) Amino acid sequences of the peptides used in
this study. Peptide wt used in this study corresponds to VSV G protein residues between 145 and 164. Six different mutants were also
synthesized: His148, His149, Glu154 or Glu158 were substituted by Gln residues, and Asp153 or Asp161 were substituted by Asn residues. The
dots represent wt residues. (B) Peptide145 �164 or the mutant peptides were incubated with 1 mM ANS in 20 mM MES, 30 mM Tris buffer,
pH 6.0, and the ANS fluorescence spectra were collected after excitation at 360 nm. (C) Kinetics of membrane fusion by peptide145 �164 or
mutant peptides. Equal amounts of unlabeled vesicles and vesicles labeled with Rh-PE and NBD-PE were incubated with each of the
peptides and the membrane fusion was measured monitoring the increase in NBD-PE fluorescence. The vesicles were composed of PC:PS
(1:3) and were prepared in 20 mM MES, 30 mM Tris buffer, pH 6.0, in a final phospholipid concentration of 0.1 mM. NBD-PE was
excited at 470 nm, and fluorescence intensity was collected at 530 nm during 10 min.
G protein charged residues in VSV-induced membrane fusion 401

and inserted into the membrane of the target cell
[27] and several studies using isolated fusion pep-
tides have evaluated peptide insertion and orienta-
tion into the lipid bilayer [28�32]. However, in the
case of the peptide145�164, a linear peptide used in
this study, the mode of interaction with the lipid
bilayer might probably be different since this peptide
is very hydrophilic. This is clearly shown in Figure 8,
which compares the plots of average hydropathies of
the amino acid residues between VSV peptide and
HIV-1 fusion peptide using the hydropathy scale of
Kyte and Doolittle [33]. While HIV-1 fusion peptide
is very hydrophobic, presenting most of the hydro-
pathies above zero, VSV peptide hydropathies are
time (min)0 2 4 6 8 10
NB
D-P
E fl
uore
scen
ce in
tens
ity (
a.u.
)
10
20
30
40
50
60
70
12 µ g
6 µ g
3 µ g
100 µ g
30 µ g
18 µ g
A C
[peptide] (µg/ml)0 20 40 60 80 100
% fu
sion
0
20
40
60
80 B
[peptide] (µg/ml)0 20 40 60 80 100
AN
S fl
uore
scen
ce in
tesi
ty (
a.u.
)
100
200
300
400
500
H148Q
D153N
E158Q
wt
H149Q
E154Q
D161N
Figure 5. Peptide oligomerization is required for hydrophobicity and fusion. (A) Kinetics of membrane fusion at different peptide
concentration. Equal amounts of unlabeled vesicles and vesicles labeled with Rh-PE and NBD-PE were incubated with the peptide145 �164
in a final concentration of 3, 6, 12, 18, 30 and 100 mg/ml, and the membrane fusion was measured monitoring the increase in NBD-PE
fluorescence. The vesicles were composed of PC:PS (1:3) and were prepared in 20 mM MES, 30 mM Tris buffer, pH 6.0, in a final
phospholipid concentration of 0.1 mM. NBD-PE was excited at 470 nm, and the fluorescence intensity was collected at 530 nm during 10
min. (B) Percentage of fusion after 10 min as a function of peptide concentration. (C) ANS binding as a function of peptide concentration.
Peptide145 �164 (m) and the mutants H148Q (k), H149Q (j), D153N (%), E154Q (^), E158Q (%), and D161N (\) were diluted in 20
mM MES, 30 mM Tris buffer, pH 6.0, to a final concentration as indicated in the Figure, and incubated with 1 mM ANS. ANS was excited
at 360 nm and the emission was collected at 492 nm.
Figure 6. DCCD labeling is dependent on peptide concentration. Peptide145 �164 diluted to a final concentration of 3 mg/ml (A) or
100 mg/ml (B) was modified for 1 h with DCCD at pH 6.0 and ESI-MS spectra were recorded. No labeling was observed with 3 mg/ml and a
labeling comparable to the one described in Figure 3 was observed for 100 mg/ml.
402 F. A. Carneiro et al.

mostly below zero, what characterizes a hydrophilic
peptide.
An important question raised from these data is:
how could this very hydrophilic peptide mediate
membrane fusion? We believe that the answer to this
question was found when we showed that the
oligomerization of the peptide confers hydrophobi-
city to the oligomer. The increase in hydrophobicity
correlates to the peptide fusogenic activity. Both are
maximal at pH 6.0 and decrease as pH is increased
to 7.5 or decreased to 4.0. Thus, we propose that the
self-association of the peptides at pH 6.0 promotes
the formation of a hydrophobic region important for
the interaction with the target membrane.
It has already been shown that other fusion
peptides associate at the membrane surface. For
example, comparing the membrane interaction
properties of a synthetic coiled-coil Influenza he-
magglutinin fusion peptide with the monomeric
peptide, Lau et al. [34] showed that the trimeriza-
tion of the peptide increased lipid mixing, liposome
leakage and membrane destabilization, suggesting an
important role for the oligomerization of fusogenic
peptides in the fusion mechanism. Studies using
different peptide constructs suggested that the
oligomers are formed as a result of membrane
association and that small oligomers may be more
fusogenic than monomers or large aggregates [35].
For HIV-1 fusion peptide, the ability of the peptide
to form aggregates was also correlated to its ability to
induce membrane fusion [28].
In most cases, the oligomerization of the fusion
peptides is driven by changes in their secondary
structure. Influenza hemagglutinin fusion peptide
exists in at least two interconvertible forms at
membrane surface: monomeric a-helical peptides
that insert into the bilayer, and self-associated
peptides adopting a b-sheet structure [36]. The
equilibrium between these two forms is dependent
on the pH and on the ionic strength. HIV-1 fusion
peptides form oligomeric b-strand structures when
associated to membranes [37]. GALA, a synthetic
fusogenic peptide, undergoes a conformational
change to an amphipathic helix when the pH is
reduced [38]. IR spectroscopic data demonstrated
that the peptide inserted deeply into the lipid bilayer,
oriented parallel with respect to the lipid acyl chains
[39]. In our case, however, we demonstrated by
infra-red spectroscopy that there is no change in
peptide secondary structure induced by lowering the
Figure 7. Secondary structure measurements. (A) ATR-FTIR
spectra were recorded from thin films obtained by slowly
evaporating a sample containing 10 mg of peptide145 �164 (at pH
7.5 or 6.0, as indicated in the Figure) on an attenuated total
reflection element. The samples were rehydrated by flushing D2O-
saturated N2 for 30 min at room temperature. (B) The wild-type
spectrum was compared with the spectra obtained, at pH 6.0, for
the mutants H148Q, H149Q, D153N and E158Q, as indicated in
the Figure.
0 2 4 6 8 10 12 14 16 18 20-3
-2
-1
0
1
2
3VSV peptide
amino acid sequence0 2 4 6 8 10 12 14 16 18 20
-3
-2
-1
0
1
2
3HIV peptide
hydr
opho
bici
ty
Figure 8. Kyte-Doolittle plots of fusion peptide hydropathy.
Hydrophobic profile of VSV peptide[145�164] and HIV-1 fusion
peptide (sequence VGIGALFLGFLGAAGSTHGA). The hydro-
pathy of these peptides was plotted from the amino terminus to
the carboxy terminus by averaging hydropathy values over a
window of 5 residues. More positive values are assigned to more
hydrophobic residues.
G protein charged residues in VSV-induced membrane fusion 403

pH from 7.5�6.0. Thus, rather than favored by
conformational changes, the oligomerization of
peptide145�164 seems to be triggered by the proto-
nation of His residues at the pH range of fusion,
which creates positive charges in the peptides
probably involved in electrostatic interactions with
the negatively-charged residues. This explanation is
in agreement with our previous results showing the
importance of the electrostatic interactions for VSV
fusion [8], and adds to our previous findings the idea
that these electrostatic interactions are responsible
for the formation of a hydrophobic region directed
involved in VSV interaction with the membrane
during fusion. This mechanism is different from that
observed for WAE, an amphipathic negatively
charged peptide that induces fusion of liposomal
phosphatidylcholine membranes. Pecheur et al.
(1999) provided evidence that it is the peptide
penetration rather than peptide oligomerization
that modulates peptide-induced fusion [40].
Using peptide mutants we demonstrated that
His148, Asp153 and Glu158 are essential for the
hydrophobicity and fusion. Asp153 is conserved
among at least 13 different animal rhabdoviruses
representing four recognized genera [41]. His148 is
also conserved except that it is substituted in the
Novirhabdovirus genus for a lysine, which should
also be protonated at the fusion pH (pK 10.53), or
for a serine in the Lyssavirus genus. Glu158 probably
has a specific role in VSV fusion since it is not
conserved among the rhabdovirus G proteins. We
found that His149 is important for fusion but not for
hydrophobicity, suggesting that this residue is prob-
ably involved in the direct binding to PS in the target
membrane.
Taken together, our data suggest that the negative
charges of peptide acidic residues participate in
intermolecular electrostatic interactions with posi-
tively-charged His residues, leading to the formation
of a hydrophobic domain at pH 6.0, which is
necessary to the peptide-induced membrane fusion.
This suggests a mechanism of membrane interaction
and destabilization resembling that one promoted by
the antimicrobial peptides acting through a carpet
model [42]. This model predicts an initial interac-
tion between the hydrophilic portion of the peptide
and negatively-charged membranes through electro-
static interactions. Then a reorientation of the
hydrophobic residues of the peptide toward the
hydrophobic core of the membrane occurs, causing
membrane disintegration by disrupting the bilayer
curvature.
Secondary structure prediction using the program
PSIPRED Protein Structure Prediction Server
(us.expasy.org) suggests that a loop is formed
between the residues Glu154 and Val160. The forma-
tion of this loop is supported by the presence of
residues Tyr155 and Trp159, which have large side
chains, and by the Gly157 in the middle, allowing the
structure to bend at this point. A similar loop was
found in the fusion peptide of dengue virus E
glycoprotein between residues 99 and 105 [43],
suggesting that some similarity might occur between
VSV and class II fusion peptides. One important
question to be posed is whether the data obtained
using the peptide145�164 could be correlated to the
role of the peptide when taking part of the whole G
glycoprotein. The peptide145�164 is located within
the domain shown to interact with the target
membrane at the fusogenic pH [44]. In addition,
its His residues (His148 and His149) were shown to
be modified after treatment of the whole virus with
DEPC, leading to the inhibition of virus-induced
fusion [9]. These observations support the hypoth-
esis that the peptide145-164 is also active within the
whole protein.
Acknowledgements
We would like to thank Adriana S. de Melo for
technical assistance, Dr Erik Goormaghtigh for
helpful assistance with ATR-FTIR experiments
and Dr Fabio C. Almeida for helpful suggestions.
This work was supported by grants from Conselho
Nacional de Desenvolvimento Cientıfico e Tecnolo-
gico (CNPq), Centro Argentino-Brasileiro de Bio-
tecnologia (CABBIO), Fundacao Carlos Chagas
Filho de Amparo a Pesquisa do Estado do Rio de
Janeiro (FAPERJ) and Fundacao de Amparo a
Pesquisa do Estado de Sao Paulo (FAPESP). F. A.
C. was recipient of PDEE fellowship from Coorde-
nacao de Aperfeicoamento de Pessoal de Nıvel
Superior (CAPES). G.V. thanks Action de Re-
cherches Concertees (ARC) for financial support.
References
[1] Hernandez LD, Hoffman LR, Wolfsberg TG, White JM.
Virus-cell and cell-cell fusion. Ann Rev Cell Dev Biol
1996;12:627�661.
[2] Skehel JJ, Wiley DC. Receptor binding and membrane
fusion in virus entry: the influenza hemagglutinin. Ann Rev
Biochem 2000;69:531�569.
[3] Heinz FX, Allison SL. The machinery for flavivirus fusion
with host cell membranes. Curr Opin Microbiol
2001;4:450�455.
[4] Jardetzky TS, Lamb RA. Virology: a class act. Nature
2004;427:307�308.
[5] Li Y, Drone C, Sat E, Ghosh HP. Mutational analysis of the
vesicular stomatitis virus glycoprotein G for membrane
fusion domains. J Virol 1993;67:4070�4077.
[6] Zhang L, Ghosh HP. Characterization of the putative
fusogenic domain in vesicular stomatitis virus glycoprotein
G. J Virol 1994;68:2186�2193.
404 F. A. Carneiro et al.

[7] Fredericksen B, Whitt MA. Vesicular stomatitis virus glyco-
protein mutations that affect membrane fusion activity and
abolish virus infectivity. J Virol 1995;69:1435�1443.
[8] Carneiro FA, Bianconi ML, Weissmuller G, Stauffer F, Da
Poian AT. Membrane recognition by vesicular stomatitis
virus involves enthalpy-driven protein-lipid interactions. J
Virol 2002;76:3756�3764.
[9] Carneiro FA, Stauffer F, Lima CL, Juliano MA, Juliano L,
Da Poian AT. Membrane fusion induced by vesicular
stomatitis virus depends on histidine protonation. J Biol
Chem 2003;278:13789�13794.
[10] Coll JM. Synthetic peptides from the heptad repeats of
the glycoproteins of rabies, vesicular stomatitis and
fish rhabdoviruses bind phosphatidylserine. Arch Virol
1997;142:2089�2097.
[11] Estepa A, Coll JM. Pepscan mapping and fusion-related
properties of the major phosphatidylserine-binding domain
of the glycoprotein of viral hemorrhagic septicemia virus, a
salmonid rhabdovirus. Virology 1996;216:60�70.
[12] Coll JM. Heptad-repeat sequences in the glycoprotein of
rhabdoviruses. Virus Genes 1995;10:107�114.
[13] Struck DK, Hoekstra D, Pagano RE. Use of resonance
energy transfer to monitor membrane fusion. Biochemistry
1981;20:4093�4099.
[14] Goormaghtigh E, Raussens V, Ruysschaert J-M. Attenuated
total reflection infrared spectroscopy of proteins and
lipids in biological membranes. Biochim Biophys Acta
1999;1422:105�185.
[15] Grimard V, Li C, Ramjeesingh M, Bear CE, Goormaghtigh
E, Ruysschaert J-M. Phosphorylation-induced conforma-
tional changes of cystic fibrosis transmembrane conductance
regulator monitored by attenuated total reflection-Fourier
transform IR spectroscopy and fluorescence spectroscopy. J
Biol Chem 2004;279:5528�5536.
[16] Manciu L, Chang X-B, Buyse F, Hou YX, Gustot A,
Riordan JR, Ruysschaert J-M. Intermediate structural states
involved in MRP1-mediated drug transport. Role of glu-
tathione. J Biol Chem 2003;278:3347�3356.
[17] Carraway KL, Koshland DE Jr. Carbodiimide modification
of proteins. Meth Enzymol 1972;25:616�623.
[18] Eidelman O, Schlegel R, Tralka TS, Blumenthal R. pH-
dependent fusion induced by vesicular stomatitis virus
glycoprotein reconstituted into phospholipid vesicles. J Biol
Chem 1984;259:4622�4628.
[19] Carneiro FA, Ferradosa AS, Da Poian AT. Low pH-induced
conformational changes in vesicular stomatitis virus glyco-
protein involve dramatic structure reorganization. J Biol
Chem 2001;276:62�67.
[20] Carr CM, Chaudhry C, Kim PS. Influenza hemagglutinin is
spring-loaded by a metastable native conformation. Proc
Natl Acad Sci USA 1997;94:14306�14313.
[21] Gaudin Y, Tuffereau C, Segretain D, Knossow M, Flamand
A. Reversible conformational changes and fusion activity of
rabies virus glycoprotein. J Virol 1991;65:4853�4859.
[22] Pak CC, Puri A, Blumenthal R. Conformational changes
and fusion activity of vesicular stomatitis virus glycoprotein:
[125I]iodonaphthyl azide photolabeling studies in biological
membranes. Biochemistry 1997;36:8890�8896.
[23] Gaudin Y. Rabies virus-induced membrane fusion pathway. J
Cell Biol 2000;150:601�611.
[24] Harter C, James P, Bachi T, Semenza G, Brunner J.
Hydrophobic binding of the ectodomain of influenza he-
magglutinin to membranes occurs through the ‘‘fusion
peptide’’. J Biol Chem 1989;264:6459�6464.
[25] Rabenstein M, Shin Y-K. A peptide from the heptad repeat
of human immunodeficiency virus gp41 shows both mem-
brane binding and coiled-coil formation. Biochemistry
1995;34:13390�13397.
[26] Adam B, Lins L, Stroobant V, Thomas A, Brasseur R.
Distribution of hydrophobic residues is crucial for the
fusogenic properties of the Ebola virus GP2 fusion peptide.
J Virol 2004;78:2131�2136.
[27] Nieva JL, Agirre A. Are fusion peptides a good model to
study viral cell fusion? Biochim Biophys Acta
2003;1614:104�115.
[28] Kliger Y, Aharoni A, Rapaport D, Jones P, Blumenthal R,
Shai Y. Fusion peptides derived from the HIV type 1
glycoprotein 41 associate within phospholipid membranes
and inhibit cell-cell Fusion. Structure-function study. J Biol
Chem 1997;272:13496�13505.
[29] Tatulian SA, Jones LR, Reddy LG, Stokes DL, Tamm LK.
Secondary structure and orientation of phospholamban
reconstituted in supported bilayers from polarized attenu-
ated total reflection FTIR spectroscopy. Biochemistry
1995;34:4448�4456.
[30] Martin I, Dubois MC, Defrise-Quertain F, Saermark T,
Burny A, Brasseur R, Ruysschaert JM. Correlation between
fusogenicity of synthetic modified peptides corresponding
to the NH2-terminal extremity of simian immunodeficiency
virus gp32 and their mode of insertion into the lipid
bilayer: an infrared spectroscopy study. J Virol
1994;68:1139�1148.
[31] Martin I, Schaal H, Scheid A, Ruysschaert JM. Lipid
membrane fusion induced by the human immunodeficiency
virus type 1 gp41 N-terminal extremity is determined
by its orientation in the lipid bilayer. J Virol 1996;70:298�304.
[32] Ghosh JK, Shai Y. Direct evidence that the N-terminal
heptad repeat of Sendai virus fusion protein participates in
membrane fusion. J Mol Biol 1999;292:531�546.
[33] Kyte J, Doolittle RF. A simple method for displaying the
hydropathic character of a protein. J Mol Biol
1982;157:105�132.
[34] Lau WL, Ege DS, Lear JD, Hammer DA, DeGrado WF.
Oligomerization of fusogenic peptides promotes membrane
fusion by enhancing membrane destabilization. Biophys J
2004;86:272�284.
[35] Yang J, Prorok M, Castellino FJ, Weliky DP. Oligomeric
beta-structure of the membrane-bound HIV-1 fusion pep-
tide formed from soluble monomers. Biophys J
2004;87:1951�1963.
[36] Han X, Tamm LK. pH-dependent self-association of
influenza hemagglutinin fusion peptides in lipid bilayers. J
Mol Biol 2000;304:953�965.
[37] Yang J, Weliky DP. Solid-state nuclear magnetic resonance
evidence for parallel and antiparallel strand arrangements in
the membrane-associated HIV-1 fusion peptide. Biochem-
istry 2003;42:11879�11890.
[38] Li W, Nicol F, Szoka FC Jr. GALA: a designed synthetic
pH-responsive amphipathic peptide with applications in
drug and gene delivery. Adv Drug Deliv Rev
2004;56:967�985.
[39] Goormaghtigh E, De Meutter J, Szoka F, Cabiaux V,
Parente RA, Ruysschaert J-M. Secondary structure and
orientation of the amphipathic peptide GALA in lipid
structures. An infrared-spectroscopic approach. Eur J Bio-
chem 1991;195:421�429.
[40] Pecheur EI, Sainte-Marie J, Bienvenue A, Hoekstra D. Lipid
headgroup spacing and peptide penetration, but not peptide
oligomerization, modulate peptide-induced fusion. Bio-
chemistry 1999;38:364�373.
G protein charged residues in VSV-induced membrane fusion 405

[41] Walker PJ, Kongsuwan K. Deduced structural model for
animal rhabdovirus glycoproteins. J Gen Virol 1999;80:
1211�1220.
[42] Shai Y. Mechanism of the binding, insertion and destabiliza-
tion of phospholipid bilayer membranes by alpha-helical
antimicrobial and cell non-selective membrane-lytic pep-
tides. Biochim Biophys Acta 1999;1462:55�70.
[43] Modis Y, Ogata S, Clements D, Harrison SC. Structure of
the dengue virus envelope protein after membrane fusion.
Nature 2004;427:313�319.
[44] Durrer P, Gaudin Y, Ruigrok WH, Graf R, Brunner J.
Photolabeling identifies a putative fusion domain in the
envelope glycoprotein of rabies and vesicular stomatitis
viruses. J Biol Chem 1995;270:17575�17581.
This paper was first published online on prEview on 27 June 2006.
406 F. A. Carneiro et al.