cellular liaisons of natural killer lymphocytes in immunology and immunotherapy of cancer
TRANSCRIPT

Review
10.1517/14712598.7.5.599 © 2007 Informa UK Ltd ISSN 1471-2598 599
Cytokines & Chemokines
Cellular liaisons of natural killer lymphocytes in immunology and immunotherapy of cancerAinhoa Arina, Oihana Murillo, Juan Dubrot, Arantza Azpilikueta, Carlos Alfaro, Jose L Pérez-Gracia, Maurizio Bendandi, Belen Palencia, Sandra Hervás-Stubbs & Ignacio Melero†
†University of Navarra, Centro de Investigación Médica Aplicada (CIMA) and Clinica Universitaria, Gene Therapy Unit, Avda. Pio XII 55, 31008, Pamplona, Spain
There is compelling evidence for the role of natural killer (NK) cells in tumorimmunosurveillance and their beneficial effects on many experimentallysuccessful immunotherapy strategies. NK cells mediate cellcontact-dependent cellular cytotoxicity and produce pro-inflammatorycytokines, but do not rearrange antigen receptors. Their activation dependson various germline-encoded receptors, including CD16, which mediatesrecognition of antibody-coated target cells. NK cytotoxicity is checked by arepertoire of inhibitory receptors that scan adequate expression of majorhistocompatibility complex class I molecules on the potential target cell.Functional cross-talk of NK and dendritic cells suggests a critical role forNK cells in the initiation and regulation of cellular immunity. Considerableknowledge on the molecular basis of NK recognition/activation contrastswith a lack of successful translational research on these matters. However,there is plenty of opportunity for targeted intervention ofinhibitory/activatory surface receptors and for adoptive cell therapy withautologous or allogeneic NK cells.
Keywords: adaptive immunity, cancer immunology, DC, immunotherapy, NK cell, NKG2D
Expert Opin. Biol. Ther. (2007) 7(5):599-615
1. Introduction
Natural killer (NK) cells were initially discovered because of their ability to kill tumorcells without the need for previous antigen exposure or immunization and withoutapparent specificity. These lymphocytes are now considered as key regulators not onlyof innate, but also of adaptive immune responses [1]. NK cells are found at theirhighest frequency in the peripheral blood (5 – 20% of the circulating lymphocytes),spleen (3% of mice splenocytes) and liver (10% of hepatic lymphocytes) [2,3].
Regarding the expression of surface markers, mouse NK cells are often defined asNK1.1+CD3- or DX5+CD3- cells. Until recently, to unambiguously identify amurine NK cell, a combination of three antibodies could be used: NK1.1/DX5,CD3 and CD122 [4]. CD122 (IL-2Rβ) responds to interleukin (IL)-15 and -2, andis essential for NK cell ontogeny, which implies that all NK cells express thatmolecule. However, the characterization of a new subpopulation of immune cells,closely related to NK cells (IFN-producing killer dendritic cell [IKDC] [5,6]), makesits necessary to add a fourth antibody: B220. Expression of B220 by IKDCs and notby NK cells may be used to distinguish both populations. In humans, CD56, CD16and CD94 have been considered as the most typical NK cell surface markers,although they are shared by many T lymphocytes. Human NK cells are best defined
1. Introduction
2. Effector functions of NK cells
3. NK cell receptors involved in
tumor target recognition
4. NK cells in cancer immunology
and immunotherapy: studies in
mouse models
5. Cross-regulation of NK cells, DCs
and T cells
6. Role of NKG2D and its ligands in
cancer immunology
7. NK cells in immunotherapy of
human malignancies
8. Expert opinion and conclusion
For reprint orders, please contact:[email protected]
Exp
ert O
pin.
Bio
l. T
her.
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
idad
De
Nav
arra
Fo
r pe
rson
al u
se o
nly.

Cellular liaisons of natural killer lymphocytes in immunology and immunotherapy of cancer
600 Expert Opin. Biol. Ther. (2007) 7(5)
as CD3-negative cells that express CD56, CD16 andCD94 [7], whereas human counterparts for IKDC have notbeen identified yet.
This article surveys the molecular mechanisms of NK cellsregarding their potential usage for immunotherapy againstcancer by directly manipulating NK cell functions or byunderstanding how the interaction of NK cells with adaptiveleukocytes would improve antitumor immunity.
2. Effector functions of NK cells
The primary functions of NK cells are the cytolysis of bothtransformed and infected cells, and the regulation of innateand adaptive immune responses through the production ofcytokines. Although mature NK cells are functionally active,for the elaboration of full effector functions, NK cells requirean activating stimulus that can be provided by cytokines (suchas IL-15, -2, -12, -18 and -21) or by direct interaction withanother cell (for example, with mature dendritic cells[DCs]) [3,8]. The effector mechanisms used by activatedNK cells include:
2.1 Cytolysis2.1.1 Perforin/granzyme-mediated natural cytotoxicityResting NK cells contain preformed granules that storeperforin and granzymes A and B. When the proper conditionsexist (for example, loss of self-major histocompatibilitycomplex [MHC] molecules in the presence of activatoryligands on the same cell), the granules containing perforin andgranzymes are mobilized and directed towards the membraneof the target cell at the site of intercellular contact [8].
2.1.2 TRAIL/Fas ligand-mediated apoptosisTNF-related apoptosis-inducing ligand (TRAIL) and Fasligand (FasL), are members of the TNF family of cytokinesand are expressed by effector lymphocytes, serving as inducersof apoptosis in the course of immune responses. In mice,immature NK cells constitutively expressing TRAIL havebeen identified in bone marrow and liver. Liver NK cellantimetastatic capacity has been shown to beTRAIL-dependent in a murine model [9,10]. Furthermore,TRAIL is induced in most activated NK cells in humans andmice [11]. NK cells can also directly promote upregulation ofFas expression in cancer cells and then kill them in aFas-dependent manner [12].
2.1.3 Antibody-dependent cell-mediated cytotoxicity Using this system, NK cells can kill targets that havepreviously been recognized by antibodies. When an IgGantibody binds to its recognized antigen, its Fc portion isbound by CD16 (FcγRIII) present on NK cells, leading toactivation and proliferation of NK cells, target-celldestruction and production of cytokines. This phenomenon iscalled antibody-dependent cell-mediated cytotoxicity(ADCC) [13]. This mechanism has important therapeutic
implications with the advent of monoclonal antibodies todestroy malignant cells. For example, the main lymphomacell-destroying mechanism of anti-CD20 monoclonalantibody (rituximab) is believed to be mediated by ADCC.When treating with these mAbs, administration ofNK-activating cytokines can be advisable to enhanceADCC [14] and, conceivably, the overall therapeutic activity.
2.2 Cytokine secretionActivation of NK cell production of T helper (Th) type 1(IFN-γ) and Th2 (IL-4, -5 and -13) cytokines occurs at distinctstages of NK cell differentiation. Mature NK cells lose the abilityto produce the type-2 cytokines and acquire the capacity tosecrete IFN-γ when primed [15]. Further activation of NK cellsleads to the production of a conspicuous battery of cytokines,chemokines and growth factors: tumor necrosis factor (TNF),interferon (IFN)-α, interleukin (IL)-1, -3 and -6, macrophageinflammatory protein (MIP)-1α and -1β, lymphotactin,RANTES and granulocyte-macrophage colony-stimulatingfactor (GM-CSF) [8]. Among all of these cytokines, IFN-γ playsthe most important role in protection against cancer [16].
3. NK cell receptors involved in tumor target recognition
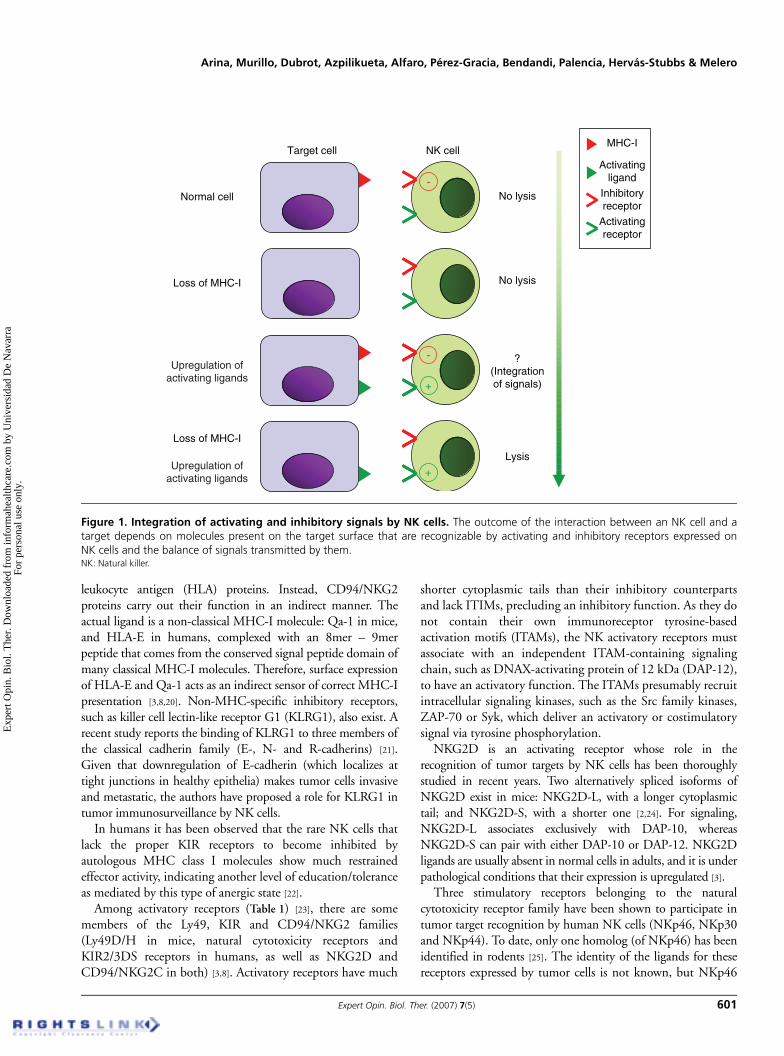
The initial observation that NK cells preferentially kill cells thatlack MHC-I expression led to the missing-self hypothesis [17],which predicted the existence of NK inhibitory receptorsspecific for MHC-I molecules. The co-regulation of NK cellfunction by NK-activating receptors acting in the opposite waywas much less expected. However, the missing-self hypothesiscannot account for all NK cell-mediated cytotoxicity; indeed,under certain conditions, NK cells can kill evenMHC-I-positive tumor cells. NK-activating receptors canpartially explain this fact in as much as they can recognizeligands that are upregulated in tumoral and stressed cells orviral glycoproteins [18]. It is now believed that NK cell functionis controlled by a refined balance of NK cell inhibitory andactivating signals (Figure 1) [1,2].
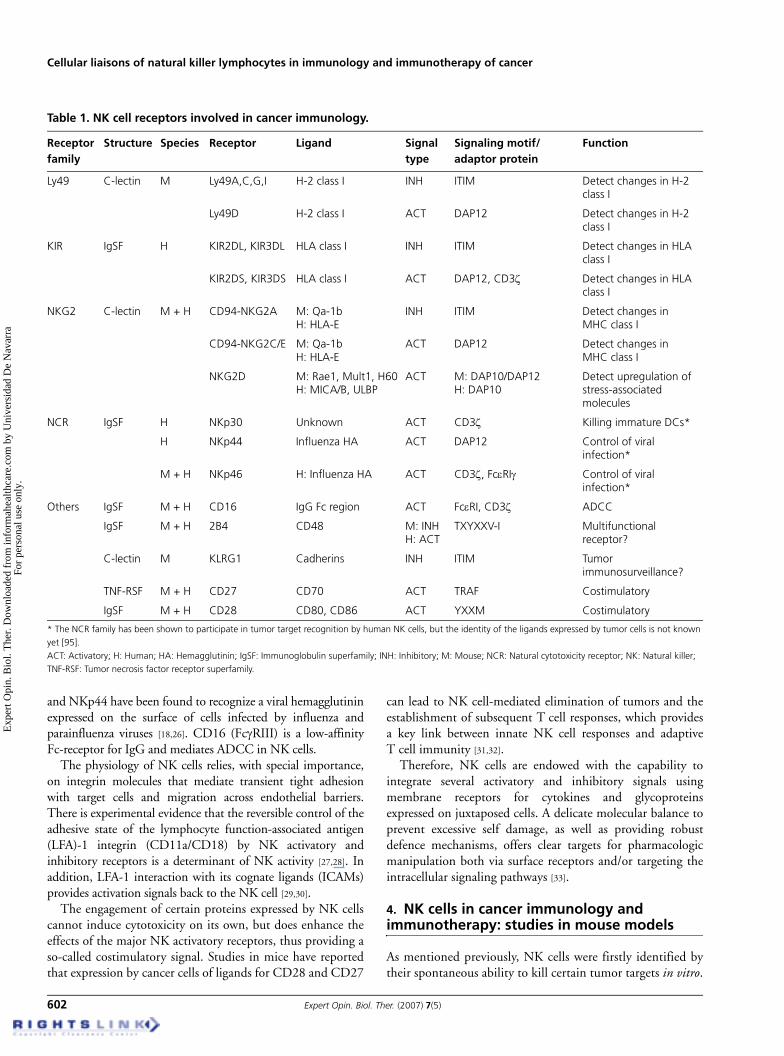
There are different classes of inhibitory receptors specific forMHC (Table 1), including the Ly49 family in mice, the killercell immunoglobulin-like receptor (KIR) L family in humansand the CD94–NKG2A complex in both species. The set ofinhibitory receptors expressed is maintained by the clonalprogeny of a mature NK cell, such that a stable clonal repertoiredefined by coexpression of a set of inhibitory receptors of NKcells is originated [19]. These receptors contain a signalingimmunoreceptor tyrosine-based inhibitory motif (ITIM) intheir cytoplasmic domains, allowing for the inhibition of NKfunction. Engagement of an inhibitory receptor with its cognateMHC-I ligand results in tyrosine phosphorylation of the ITIMand subsequent recruitment of SHP-1 or -2 tyrosinephosphatases, an event which ultimately leads to inhibition ofNK effector functions. Different members of the Ly49/KIRfamilies directly bind to different MHC-I H-2/human
Exp
ert O
pin.
Bio
l. T
her.
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
idad
De
Nav
arra
Fo
r pe
rson
al u
se o
nly.
Arina, Murillo, Dubrot, Azpilikueta, Alfaro, Pérez-Gracia, Bendandi, Palencia, Hervás-Stubbs & Melero
Expert Opin. Biol. Ther. (2007) 7(5) 601
leukocyte antigen (HLA) proteins. Instead, CD94/NKG2proteins carry out their function in an indirect manner. Theactual ligand is a non-classical MHC-I molecule: Qa-1 in mice,and HLA-E in humans, complexed with an 8mer – 9merpeptide that comes from the conserved signal peptide domain ofmany classical MHC-I molecules. Therefore, surface expressionof HLA-E and Qa-1 acts as an indirect sensor of correct MHC-Ipresentation [3,8,20]. Non-MHC-specific inhibitory receptors,such as killer cell lectin-like receptor G1 (KLRG1), also exist. Arecent study reports the binding of KLRG1 to three members ofthe classical cadherin family (E-, N- and R-cadherins) [21].Given that downregulation of E-cadherin (which localizes attight junctions in healthy epithelia) makes tumor cells invasiveand metastatic, the authors have proposed a role for KLRG1 intumor immunosurveillance by NK cells.
In humans it has been observed that the rare NK cells thatlack the proper KIR receptors to become inhibited byautologous MHC class I molecules show much restrainedeffector activity, indicating another level of education/toleranceas mediated by this type of anergic state [22].
Among activatory receptors (Table 1) [23], there are somemembers of the Ly49, KIR and CD94/NKG2 families(Ly49D/H in mice, natural cytotoxicity receptors andKIR2/3DS receptors in humans, as well as NKG2D andCD94/NKG2C in both) [3,8]. Activatory receptors have much
shorter cytoplasmic tails than their inhibitory counterpartsand lack ITIMs, precluding an inhibitory function. As they donot contain their own immunoreceptor tyrosine-basedactivation motifs (ITAMs), the NK activatory receptors mustassociate with an independent ITAM-containing signalingchain, such as DNAX-activating protein of 12 kDa (DAP-12),to have an activatory function. The ITAMs presumably recruitintracellular signaling kinases, such as the Src family kinases,ZAP-70 or Syk, which deliver an activatory or costimulatorysignal via tyrosine phosphorylation.
NKG2D is an activating receptor whose role in therecognition of tumor targets by NK cells has been thoroughlystudied in recent years. Two alternatively spliced isoforms ofNKG2D exist in mice: NKG2D-L, with a longer cytoplasmictail; and NKG2D-S, with a shorter one [2,24]. For signaling,NKG2D-L associates exclusively with DAP-10, whereasNKG2D-S can pair with either DAP-10 or DAP-12. NKG2Dligands are usually absent in normal cells in adults, and it is underpathological conditions that their expression is upregulated [3].
Three stimulatory receptors belonging to the naturalcytotoxicity receptor family have been shown to participate intumor target recognition by human NK cells (NKp46, NKp30and NKp44). To date, only one homolog (of NKp46) has beenidentified in rodents [25]. The identity of the ligands for thesereceptors expressed by tumor cells is not known, but NKp46
Figure 1. Integration of activating and inhibitory signals by NK cells. The outcome of the interaction between an NK cell and atarget depends on molecules present on the target surface that are recognizable by activating and inhibitory receptors expressed onNK cells and the balance of signals transmitted by them. NK: Natural killer.
Target cell NK cell
Normal cell
Loss of MHC-I
-
+
+
-
No lysis
?(Integrationof signals)
Lysis
MHC-I
Activatingligand
Inhibitoryreceptor
Activatingreceptor
Loss of MHC-I
Upregulation ofactivating ligands
Upregulation ofactivating ligands
No lysis
Exp
ert O
pin.
Bio
l. T
her.
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
idad
De
Nav
arra
Fo
r pe
rson
al u
se o
nly.
Cellular liaisons of natural killer lymphocytes in immunology and immunotherapy of cancer
602 Expert Opin. Biol. Ther. (2007) 7(5)
and NKp44 have been found to recognize a viral hemagglutininexpressed on the surface of cells infected by influenza andparainfluenza viruses [18,26]. CD16 (FcγRIII) is a low-affinityFc-receptor for IgG and mediates ADCC in NK cells.
The physiology of NK cells relies, with special importance,on integrin molecules that mediate transient tight adhesionwith target cells and migration across endothelial barriers.There is experimental evidence that the reversible control of theadhesive state of the lymphocyte function-associated antigen(LFA)-1 integrin (CD11a/CD18) by NK activatory andinhibitory receptors is a determinant of NK activity [27,28]. Inaddition, LFA-1 interaction with its cognate ligands (ICAMs)provides activation signals back to the NK cell [29,30].
The engagement of certain proteins expressed by NK cellscannot induce cytotoxicity on its own, but does enhance theeffects of the major NK activatory receptors, thus providing aso-called costimulatory signal. Studies in mice have reportedthat expression by cancer cells of ligands for CD28 and CD27
can lead to NK cell-mediated elimination of tumors and theestablishment of subsequent T cell responses, which providesa key link between innate NK cell responses and adaptiveT cell immunity [31,32].
Therefore, NK cells are endowed with the capability tointegrate several activatory and inhibitory signals usingmembrane receptors for cytokines and glycoproteinsexpressed on juxtaposed cells. A delicate molecular balance toprevent excessive self damage, as well as providing robustdefence mechanisms, offers clear targets for pharmacologicmanipulation both via surface receptors and/or targeting theintracellular signaling pathways [33].
4. NK cells in cancer immunology and immunotherapy: studies in mouse models
As mentioned previously, NK cells were firstly identified bytheir spontaneous ability to kill certain tumor targets in vitro.
Table 1. NK cell receptors involved in cancer immunology.
Receptor family
Structure Species Receptor Ligand Signal type
Signaling motif/adaptor protein
Function
Ly49 C-lectin M Ly49A,C,G,I H-2 class I INH ITIM Detect changes in H-2 class I
Ly49D H-2 class I ACT DAP12 Detect changes in H-2 class I
KIR IgSF H KIR2DL, KIR3DL HLA class I INH ITIM Detect changes in HLA class I
KIR2DS, KIR3DS HLA class I ACT DAP12, CD3ζ Detect changes in HLA class I
NKG2 C-lectin M + H CD94-NKG2A M: Qa-1bH: HLA-E
INH ITIM Detect changes in MHC class I
CD94-NKG2C/E M: Qa-1bH: HLA-E
ACT DAP12 Detect changes in MHC class I
NKG2D M: Rae1, Mult1, H60H: MICA/B, ULBP
ACT M: DAP10/DAP12H: DAP10
Detect upregulation of stress-associated molecules
NCR IgSF H NKp30 Unknown ACT CD3ζ Killing immature DCs*
H NKp44 Influenza HA ACT DAP12 Control of viral infection*
M + H NKp46 H: Influenza HA ACT CD3ζ, FcεRIγ Control of viral infection*
Others IgSF M + H CD16 IgG Fc region ACT FcεRI, CD3ζ ADCC
IgSF M + H 2B4 CD48 M: INHH: ACT
TXYXXV-I Multifunctional receptor?
C-lectin M KLRG1 Cadherins INH ITIM Tumor immunosurveillance?
TNF-RSF M + H CD27 CD70 ACT TRAF Costimulatory
IgSF M + H CD28 CD80, CD86 ACT YXXM Costimulatory
* The NCR family has been shown to participate in tumor target recognition by human NK cells, but the identity of the ligands expressed by tumor cells is not known yet [95].ACT: Activatory; H: Human; HA: Hemagglutinin; IgSF: Immunoglobulin superfamily; INH: Inhibitory; M: Mouse; NCR: Natural cytotoxicity receptor; NK: Natural killer; TNF-RSF: Tumor necrosis factor receptor superfamily.
Exp
ert O
pin.
Bio
l. T
her.
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
idad
De
Nav
arra
Fo
r pe
rson
al u
se o
nly.

Arina, Murillo, Dubrot, Azpilikueta, Alfaro, Pérez-Gracia, Bendandi, Palencia, Hervás-Stubbs & Melero
Expert Opin. Biol. Ther. (2007) 7(5) 603
The classical cell line used as target for mouse NK cells isYAC-1 (a Moloney virus-induced lymphoma from A/Snmice). When activated with IL-2 or type-I IFN, NK cellsshow a greater cytotoxic capacity, allowing them to kill abroader spectrum of tumor targets that are otherwiseresistant to freshly isolated NK cells [2]. These ex vivoactivated cells are called ‘lymphokine-activated killer’ cells.An extensive series of experiments have confirmed theparticipation of NK cells in protection against tumors inmouse models: a number of representative studies are shownin Table 2. Among these reports and depending on thequestions addressed regarding NK cell function, it is possibleto find different sorts of models:
• Experiments aimed at ascertaining the NK cell role inprotection against tumor initiation, incarcinogen-induced tumorigenesis.
• Experiments that assess NK involvement in tumor growthand metastasis development, using models of establishedtransplantable tumors.
• Experiments directed to determine the implication ofcertain molecules necessary for NK cell effector functionsin protection against cancer.
In order to achieve these objectives, several differentexperimental approaches can also be listed (Table 2):
• Depletion of NK cells using anti-asialoGM1 antisera. Thismethod, however, is not completely specific, as theasialoGM1 glycolipid can also be expressed by CTLs andmacrophages. The alternative consists of depletion ofNK1.1+ cells using anti-NK1.1-specific monoclonalantibodies. Anti-NK1.1 efficiently depletes NK cells, butonly in selected mouse strains; furthermore, NK1.1 is alsopresent in a small subset of T cells, named natural killer T(NKT) cells, which have also been found to mediateantitumor activity.
• The employment of mouse strains with a defective NKfunction, such as non-obese diabetic (NOD) and beigemice. These strains have a normal number of NK cells, butexhibit an impaired cytolytic capacity that also affects othercytotoxic cell subsets.
• The development of ‘knock-out’ strains of mice withtargeted mutations on the genes of interest: STAT1, IFN-γ,IFN-γ receptor α chain (IFN-γR1), perforin and TRAIL.An alternative approach consists of the temporary blockadeof the target molecule through in vivo administration ofblocking monoclonal antibodies.
• The administration of cytokines (such as IL-2, -12, -15,-18 and -21), chemokines or monoclonalimmunostimulating antibodies known to potentiateNK cell effector functions, as an indirect way to determineNK-mediated action.
Some general conclusions about NK cell activity in the cancerimmune response can be inferred from these experiments [2].As expected, NK cells are more efficient at eliminating tumor
targets that express either low levels of MHC-I molecules orthat do not express them at all. Another general rule is thatNK cells are usually able to prevent the outgrowth ofNK cell-sensitive tumors when challenges consisting of lowtumor cell numbers are injected, but this ability can beoverwhelmed by sufficiently increasing the tumor cell charge.In tumor models, NK cells have not exhibited a memorycapacity, that is, following a repeated exposure to a tumor thathas been eliminated by NK cells, there is not an accelerated oraugmented increase in the pool of NK cells that respond tothat tumor. NK cells are usually necessary, but not sufficient, toreject a number of murine tumors in vivo, with the exceptionof MHC-I-deficient tumor cells, such as RMA-S [17].
Thus, the evidence for an involvement of NK cellimmunotherapy and immunosurveillance in mouse models iscompelling. Although there is solid experimental evidencegenerated with knockout mice and monoclonal antibodiespointing at the key involvement of some molecules, there isconsiderable lack of knowledge about how, when and wherethese NK mechanisms are executed. In particular, the study ofthe interplay of NK cells in networks with otherleukocytes [34] is offering important pieces of informationwith potential translational research value.
5. Cross-regulation of NK cells, DCs and T cells
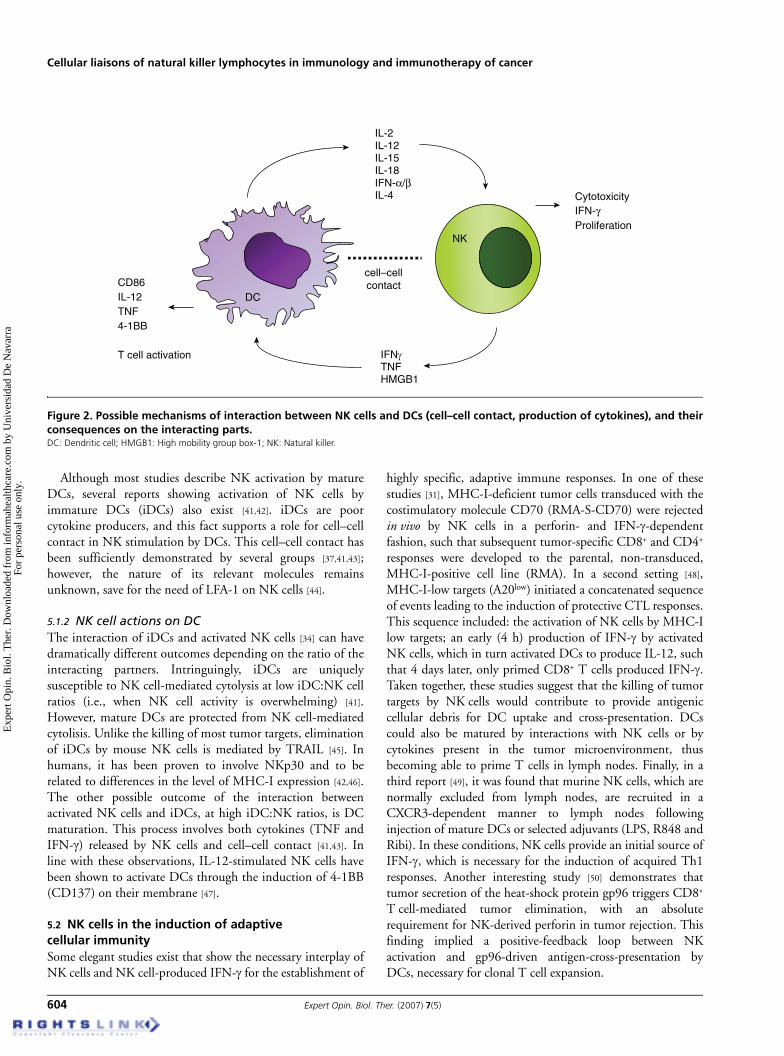
5.1 DC–NK cross-talkA significative number of reports provide evidence of thereciprocal influences between DCs and NK cells (reviewedin [35]) (Figure 2), a possibility first proposed in 1985. BothDCs and NK cells can be considered part of the innate arm ofimmunity and, therefore, their role at initiating immuneresponses is deemed essential. In addition, DCs colocalizewith NK cells in the T cell areas of human lymph nodes, asshown in an interesting study by Ferlazzo et al. [36], suggestingthat the DC–NK interaction may have further implicationsregarding T cell responses.
5.1.1 DC actions on NK cellsBoth DC-derived cytokines and cell–cell contact contributeto enhancement of NK function by DCs. Among cytokines,IL-12 and -2 appear to be necessary for mature DC triggeringof IFN-γ production by NK cells, depending on the stimulusused to induce DC maturation [36-38]. In contrast, andfollowing data presented in the same studies, type-I IFNsseem to be crucial for stimulation of NK cytotoxicity, whereasDC-membrane-bound IL-15 is essential for NK cellproliferation and survival. IL-4 can also promoteDC-mediated activation of NK cells through DAP12triggering [39]. A recent report proposes a DC directionalsecretion of IL-18 as an important mechanism acting at theDC–NK immunological synapse, thereby inducing NK cellsto release high mobility group box-1 (HMGB1), which inturn enhances DC maturation, thus establishing abidirectional cross-talk [40].
Exp
ert O
pin.
Bio
l. T
her.
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
idad
De
Nav
arra
Fo
r pe
rson
al u
se o
nly.
Cellular liaisons of natural killer lymphocytes in immunology and immunotherapy of cancer
604 Expert Opin. Biol. Ther. (2007) 7(5)
Although most studies describe NK activation by matureDCs, several reports showing activation of NK cells byimmature DCs (iDCs) also exist [41,42]. iDCs are poorcytokine producers, and this fact supports a role for cell–cellcontact in NK stimulation by DCs. This cell–cell contact hasbeen sufficiently demonstrated by several groups [37,41,43];however, the nature of its relevant molecules remainsunknown, save for the need of LFA-1 on NK cells [44].
5.1.2 NK cell actions on DCThe interaction of iDCs and activated NK cells [34] can havedramatically different outcomes depending on the ratio of theinteracting partners. Intringuingly, iDCs are uniquelysusceptible to NK cell-mediated cytolysis at low iDC:NK cellratios (i.e., when NK cell activity is overwhelming) [41].However, mature DCs are protected from NK cell-mediatedcytolisis. Unlike the killing of most tumor targets, eliminationof iDCs by mouse NK cells is mediated by TRAIL [45]. Inhumans, it has been proven to involve NKp30 and to berelated to differences in the level of MHC-I expression [42,46].The other possible outcome of the interaction betweenactivated NK cells and iDCs, at high iDC:NK ratios, is DCmaturation. This process involves both cytokines (TNF andIFN-γ) released by NK cells and cell–cell contact [41,43]. Inline with these observations, IL-12-stimulated NK cells havebeen shown to activate DCs through the induction of 4-1BB(CD137) on their membrane [47].
5.2 NK cells in the induction of adaptive cellular immunitySome elegant studies exist that show the necessary interplay ofNK cells and NK cell-produced IFN-γ for the establishment of
highly specific, adaptive immune responses. In one of thesestudies [31], MHC-I-deficient tumor cells transduced with thecostimulatory molecule CD70 (RMA-S-CD70) were rejectedin vivo by NK cells in a perforin- and IFN-γ-dependentfashion, such that subsequent tumor-specific CD8+ and CD4+
responses were developed to the parental, non-transduced,MHC-I-positive cell line (RMA). In a second setting [48],MHC-I-low targets (A20low) initiated a concatenated sequenceof events leading to the induction of protective CTL responses.This sequence included: the activation of NK cells by MHC-Ilow targets; an early (4 h) production of IFN-γ by activatedNK cells, which in turn activated DCs to produce IL-12, suchthat 4 days later, only primed CD8+ T cells produced IFN-γ.Taken together, these studies suggest that the killing of tumortargets by NK cells would contribute to provide antigeniccellular debris for DC uptake and cross-presentation. DCscould also be matured by interactions with NK cells or bycytokines present in the tumor microenvironment, thusbecoming able to prime T cells in lymph nodes. Finally, in athird report [49], it was found that murine NK cells, which arenormally excluded from lymph nodes, are recruited in aCXCR3-dependent manner to lymph nodes followinginjection of mature DCs or selected adjuvants (LPS, R848 andRibi). In these conditions, NK cells provide an initial source ofIFN-γ, which is necessary for the induction of acquired Th1responses. Another interesting study [50] demonstrates thattumor secretion of the heat-shock protein gp96 triggers CD8+
T cell-mediated tumor elimination, with an absoluterequirement for NK-derived perforin in tumor rejection. Thisfinding implied a positive-feedback loop between NKactivation and gp96-driven antigen-cross-presentation byDCs, necessary for clonal T cell expansion.
Figure 2. Possible mechanisms of interaction between NK cells and DCs (cell–cell contact, production of cytokines), and theirconsequences on the interacting parts.DC: Dendritic cell; HMGB1: High mobility group box-1; NK: Natural killer.
IL-2IL-12IL-15IL-18IFN-α/βIL-4 Cytotoxicity
IFN-γProliferation
IFNγTNFHMGB1
CD86IL-12TNF4-1BB
T cell activation
DC
NK
cell–cellcontact
Exp
ert O
pin.
Bio
l. T
her.
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
idad
De
Nav
arra
Fo
r pe
rson
al u
se o
nly.
Arina, Murillo, Dubrot, Azpilikueta, Alfaro, Pérez-Gracia, Bendandi, Palencia, Hervás-Stubbs & Melero
Expert Opin. Biol. Ther. (2007) 7(5) 605
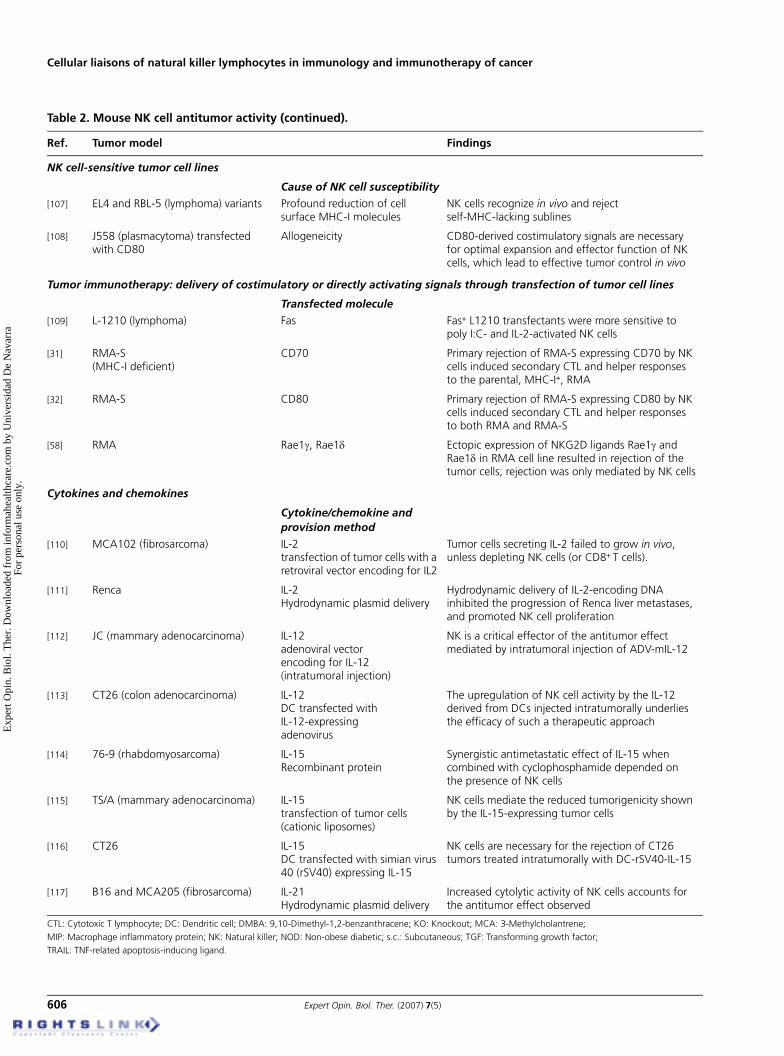
Table 2. Mouse NK cell antitumor activity .
Ref. Tumor model Findings
NK cell depletion
Depleting antibody[96] Chemical carcinogenesis (MCA s.c.) Anti-asialoGM1, anti-NK1.1 Both NK and NKT cells are essential in host
immunity against MCA-induced sarcoma
Mouse strains with defective NK function
Strain
[97] Chemical carcinogenesis by intragastric admistration of DMBA
Beige mice (reduced lytic NK activity, impairment of macrophages and neutrophils)
Earlier and/or higher incidence of non-thymic lymphomas and of epithelial and non-epithelial cutaneous tumors
[98] Spontaneous metastatic pancreatic cancer
T Ag/beige mice Crossing of T Ag mice to beige backgroung greatly elevated the frequency of mice containing metastases induced by SV40 T Ag gene
[99] Spontaneous tumorigenesis NOD/LtSz-scid (lacking functional NK, T and B cells)
These animals have a short lifespan due to a high incidence of thymic lymphomas in specific pathogen-free conditions.
[100] B16 melanoma and RMA-S lymphoma (MHC-I deficient)
Tg mice (selective deficiency in NK cells)
Tg mice show a higher rate of B16 lung metastasis and fail to reject minimal doses of RMA-S s.c.
Effector molecule gene-targeting or blockade
Targeted molecule and method[101] RMA-S Perforin:
Perforin KO miceGrowth of RMA-S tumor cells in non-primed mice is controlled by NK cells through perforin-dependent cytotoxicity
[102] • Spontaneous tumorigenesis• Chemical carcinogenesis (MCA s.c.)
IFN-γ:p53/IFN-γR double KO mice IFN-γR KO and STAT1 KO mice
Acceleration in spontaneous tumor development (lymphoid and non-lymphoid)Both strains develop fibrosarcomas more rapidly and with greater frequency
[103] K1735 melanoma FasL: gld mice (FasL KO) Increased number and incidence of metastases
[104] • Chemical carcinogenesis (MCA s.c.)• DA3 (mammary) and RM-1 (prostate) carcinomas
Perforin and IFN-γ:Perforin/IFN-γ double KO mice
Perforin and IFN-γ independently contribute antitumor effects, that together control the initiation, growth and metastasis
[105] • Spontaneous tumorigenesis• Chemical carcinogenesis (MCA s.c.)
IFN-γ (and lymphocytes):RAG2/STAT1 double KO mice
These mice present higher rates of MCA-induced sarcomas and a very high incidence of spontaneous epithelial carcinomas in specific pathogen-free conditions
[10] • Chemical carcinogenesis (MCA s.c.)• Renca (renal) and 4T1 (mammary) carcinomas
TRAIL: TRAIL KO mice These mice are more susceptible to experimental and spontaneous tumor metastasis, as well as to MCA-induced fibrosarcoma
[106] Renca TRAIL: in vivo blockade The antimetastatic activity of IL-12-activated liver NK cells involves TRAIL function
[57] EL4 (thymoma), RMA and B16 transduced with NKG2D ligands
NKG2D: in vivo blockade Ectopic expression of NKG2D ligands results in potent priming of CTLs and in vivo sensitisation of NK cells
[60] Chemical carcinogenesis (MCA s.c.) NKG2D: in vivo blockade NKG2D has a perforin-dependent important role in host protection from de novo tumorigenesis induced by MCA
CTL: Cytotoxic T lymphocyte; DC: Dendritic cell; DMBA: 9,10-Dimethyl-1,2-benzanthracene; KO: Knockout; MCA: 3-Methylcholantrene; MIP: Macrophage inflammatory protein; NK: Natural killer; NOD: Non-obese diabetic; s.c.: Subcutaneous; TGF: Transforming growth factor; TRAIL: TNF-related apoptosis-inducing ligand.
Exp
ert O
pin.
Bio
l. T
her.
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
idad
De
Nav
arra
Fo
r pe
rson
al u
se o
nly.
Cellular liaisons of natural killer lymphocytes in immunology and immunotherapy of cancer
606 Expert Opin. Biol. Ther. (2007) 7(5)
NK cell-sensitive tumor cell lines
Cause of NK cell susceptibility[107] EL4 and RBL-5 (lymphoma) variants Profound reduction of cell
surface MHC-I moleculesNK cells recognize in vivo and reject self-MHC-lacking sublines
[108] J558 (plasmacytoma) transfected with CD80
Allogeneicity CD80-derived costimulatory signals are necessary for optimal expansion and effector function of NK cells, which lead to effective tumor control in vivo
Tumor immunotherapy: delivery of costimulatory or directly activating signals through transfection of tumor cell lines
Transfected molecule
[109] L-1210 (lymphoma) Fas Fas+ L1210 transfectants were more sensitive to poly I:C- and IL-2-activated NK cells
[31] RMA-S(MHC-I deficient)
CD70 Primary rejection of RMA-S expressing CD70 by NK cells induced secondary CTL and helper responses to the parental, MHC-I+, RMA
[32] RMA-S CD80 Primary rejection of RMA-S expressing CD80 by NK cells induced secondary CTL and helper responses to both RMA and RMA-S
[58] RMA Rae1γ, Rae1δ Ectopic expression of NKG2D ligands Rae1γ and Rae1δ in RMA cell line resulted in rejection of the tumor cells; rejection was only mediated by NK cells
Cytokines and chemokines
Cytokine/chemokine and provision method
[110] MCA102 (fibrosarcoma) IL-2transfection of tumor cells with a retroviral vector encoding for IL2
Tumor cells secreting IL-2 failed to grow in vivo, unless depleting NK cells (or CD8+ T cells).
[111] Renca IL-2Hydrodynamic plasmid delivery
Hydrodynamic delivery of IL-2-encoding DNA inhibited the progression of Renca liver metastases, and promoted NK cell proliferation
[112] JC (mammary adenocarcinoma) IL-12adenoviral vectorencoding for IL-12(intratumoral injection)
NK is a critical effector of the antitumor effect mediated by intratumoral injection of ADV-mIL-12
[113] CT26 (colon adenocarcinoma) IL-12DC transfected withIL-12-expressingadenovirus
The upregulation of NK cell activity by the IL-12 derived from DCs injected intratumorally underlies the efficacy of such a therapeutic approach
[114] 76-9 (rhabdomyosarcoma) IL-15Recombinant protein
Synergistic antimetastatic effect of IL-15 when combined with cyclophosphamide depended on the presence of NK cells
[115] TS/A (mammary adenocarcinoma) IL-15transfection of tumor cells (cationic liposomes)
NK cells mediate the reduced tumorigenicity shown by the IL-15-expressing tumor cells
[116] CT26 IL-15DC transfected with simian virus 40 (rSV40) expressing IL-15
NK cells are necessary for the rejection of CT26 tumors treated intratumorally with DC-rSV40-IL-15
[117] B16 and MCA205 (fibrosarcoma) IL-21Hydrodynamic plasmid delivery
Increased cytolytic activity of NK cells accounts for the antitumor effect observed
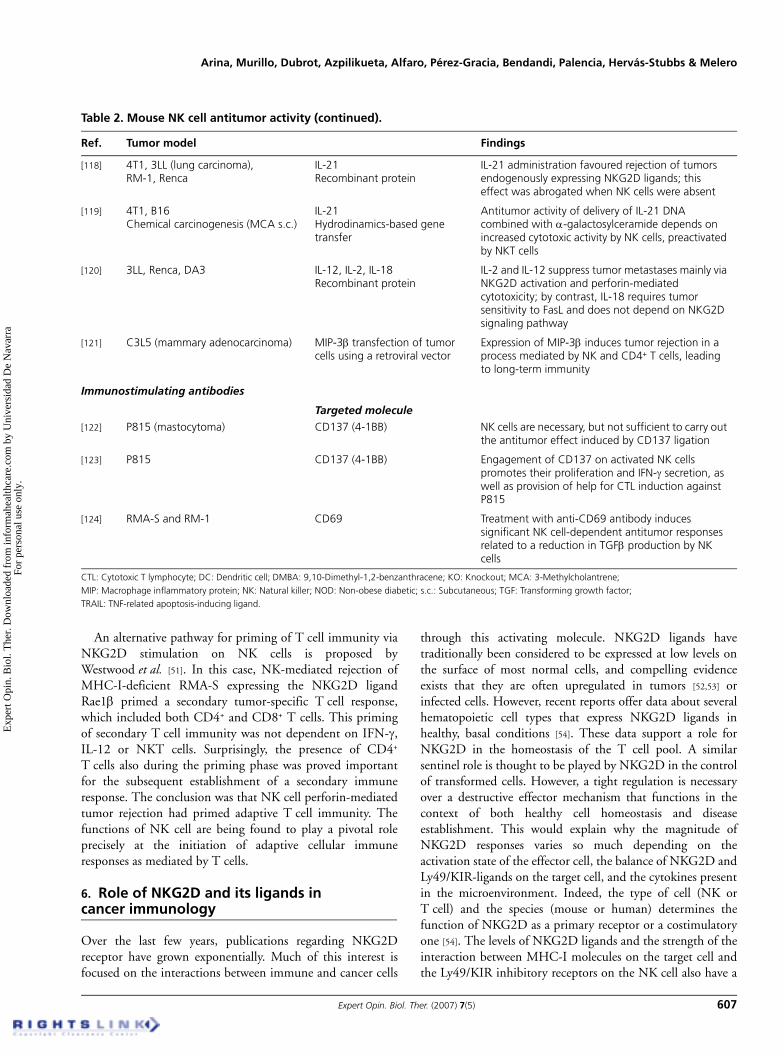
Table 2. Mouse NK cell antitumor activity (continued).
Ref. Tumor model Findings
CTL: Cytotoxic T lymphocyte; DC: Dendritic cell; DMBA: 9,10-Dimethyl-1,2-benzanthracene; KO: Knockout; MCA: 3-Methylcholantrene; MIP: Macrophage inflammatory protein; NK: Natural killer; NOD: Non-obese diabetic; s.c.: Subcutaneous; TGF: Transforming growth factor; TRAIL: TNF-related apoptosis-inducing ligand.
Exp
ert O
pin.
Bio
l. T
her.
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
idad
De
Nav
arra
Fo
r pe
rson
al u
se o
nly.
Arina, Murillo, Dubrot, Azpilikueta, Alfaro, Pérez-Gracia, Bendandi, Palencia, Hervás-Stubbs & Melero
Expert Opin. Biol. Ther. (2007) 7(5) 607
An alternative pathway for priming of T cell immunity viaNKG2D stimulation on NK cells is proposed byWestwood et al. [51]. In this case, NK-mediated rejection ofMHC-I-deficient RMA-S expressing the NKG2D ligandRae1β primed a secondary tumor-specific T cell response,which included both CD4+ and CD8+ T cells. This primingof secondary T cell immunity was not dependent on IFN-γ,IL-12 or NKT cells. Surprisingly, the presence of CD4+
T cells also during the priming phase was proved importantfor the subsequent establishment of a secondary immuneresponse. The conclusion was that NK cell perforin-mediatedtumor rejection had primed adaptive T cell immunity. Thefunctions of NK cell are being found to play a pivotal roleprecisely at the initiation of adaptive cellular immuneresponses as mediated by T cells.
6. Role of NKG2D and its ligands in cancer immunology
Over the last few years, publications regarding NKG2Dreceptor have grown exponentially. Much of this interest isfocused on the interactions between immune and cancer cells
through this activating molecule. NKG2D ligands havetraditionally been considered to be expressed at low levels onthe surface of most normal cells, and compelling evidenceexists that they are often upregulated in tumors [52,53] orinfected cells. However, recent reports offer data about severalhematopoietic cell types that express NKG2D ligands inhealthy, basal conditions [54]. These data support a role forNKG2D in the homeostasis of the T cell pool. A similarsentinel role is thought to be played by NKG2D in the controlof transformed cells. However, a tight regulation is necessaryover a destructive effector mechanism that functions in thecontext of both healthy cell homeostasis and diseaseestablishment. This would explain why the magnitude ofNKG2D responses varies so much depending on theactivation state of the effector cell, the balance of NKG2D andLy49/KIR-ligands on the target cell, and the cytokines presentin the microenvironment. Indeed, the type of cell (NK orT cell) and the species (mouse or human) determines thefunction of NKG2D as a primary receptor or a costimulatoryone [54]. The levels of NKG2D ligands and the strength of theinteraction between MHC-I molecules on the target cell andthe Ly49/KIR inhibitory receptors on the NK cell also have a
[118] 4T1, 3LL (lung carcinoma),RM-1, Renca
IL-21Recombinant protein
IL-21 administration favoured rejection of tumors endogenously expressing NKG2D ligands; this effect was abrogated when NK cells were absent
[119] 4T1, B16Chemical carcinogenesis (MCA s.c.)
IL-21Hydrodinamics-based gene transfer
Antitumor activity of delivery of IL-21 DNA combined with α-galactosylceramide depends on increased cytotoxic activity by NK cells, preactivated by NKT cells
[120] 3LL, Renca, DA3 IL-12, IL-2, IL-18Recombinant protein
IL-2 and IL-12 suppress tumor metastases mainly via NKG2D activation and perforin-mediated cytotoxicity; by contrast, IL-18 requires tumor sensitivity to FasL and does not depend on NKG2D signaling pathway
[121] C3L5 (mammary adenocarcinoma) MIP-3β transfection of tumor cells using a retroviral vector
Expression of MIP-3β induces tumor rejection in a process mediated by NK and CD4+ T cells, leading to long-term immunity
Immunostimulating antibodies
Targeted molecule[122] P815 (mastocytoma) CD137 (4-1BB) NK cells are necessary, but not sufficient to carry out
the antitumor effect induced by CD137 ligation
[123] P815 CD137 (4-1BB) Engagement of CD137 on activated NK cells promotes their proliferation and IFN-γ secretion, as well as provision of help for CTL induction against P815
[124] RMA-S and RM-1 CD69 Treatment with anti-CD69 antibody induces significant NK cell-dependent antitumor responses related to a reduction in TGFβ production by NK cells
Table 2. Mouse NK cell antitumor activity (continued).
Ref. Tumor model Findings
CTL: Cytotoxic T lymphocyte; DC: Dendritic cell; DMBA: 9,10-Dimethyl-1,2-benzanthracene; KO: Knockout; MCA: 3-Methylcholantrene; MIP: Macrophage inflammatory protein; NK: Natural killer; NOD: Non-obese diabetic; s.c.: Subcutaneous; TGF: Transforming growth factor; TRAIL: TNF-related apoptosis-inducing ligand.
Exp
ert O
pin.
Bio
l. T
her.
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
idad
De
Nav
arra
Fo
r pe
rson
al u
se o
nly.
Cellular liaisons of natural killer lymphocytes in immunology and immunotherapy of cancer
608 Expert Opin. Biol. Ther. (2007) 7(5)
decisive influence on the functional outcome of theencounter [55,56]. Finally, exposure to several cytokines canalter surface NKG2D expression either at the protein (IL-2,IL-15 or TGF-β) or at the transcriptional level (IFN) [54].
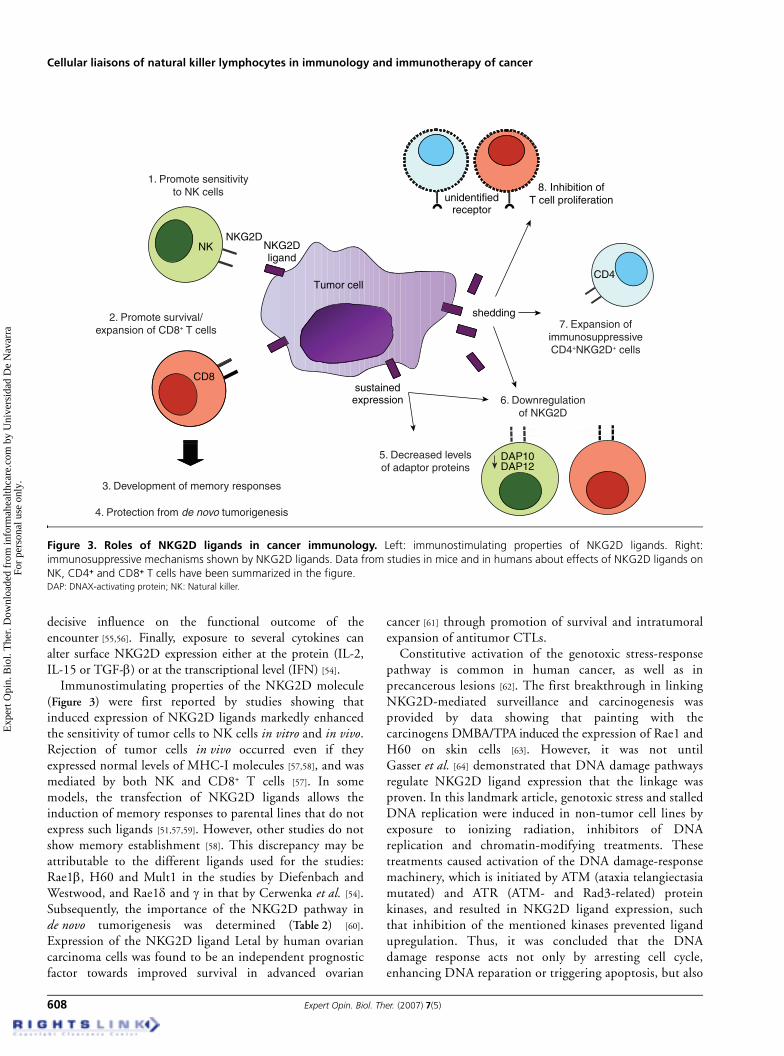
Immunostimulating properties of the NKG2D molecule(Figure 3) were first reported by studies showing thatinduced expression of NKG2D ligands markedly enhancedthe sensitivity of tumor cells to NK cells in vitro and in vivo.Rejection of tumor cells in vivo occurred even if theyexpressed normal levels of MHC-I molecules [57,58], and wasmediated by both NK and CD8+ T cells [57]. In somemodels, the transfection of NKG2D ligands allows theinduction of memory responses to parental lines that do notexpress such ligands [51,57,59]. However, other studies do notshow memory establishment [58]. This discrepancy may beattributable to the different ligands used for the studies:Rae1β, H60 and Mult1 in the studies by Diefenbach andWestwood, and Rae1δ and γ in that by Cerwenka et al. [54].Subsequently, the importance of the NKG2D pathway inde novo tumorigenesis was determined (Table 2) [60].Expression of the NKG2D ligand Letal by human ovariancarcinoma cells was found to be an independent prognosticfactor towards improved survival in advanced ovarian
cancer [61] through promotion of survival and intratumoralexpansion of antitumor CTLs.
Constitutive activation of the genotoxic stress-responsepathway is common in human cancer, as well as inprecancerous lesions [62]. The first breakthrough in linkingNKG2D-mediated surveillance and carcinogenesis wasprovided by data showing that painting with thecarcinogens DMBA/TPA induced the expression of Rae1 andH60 on skin cells [63]. However, it was not untilGasser et al. [64] demonstrated that DNA damage pathwaysregulate NKG2D ligand expression that the linkage wasproven. In this landmark article, genotoxic stress and stalledDNA replication were induced in non-tumor cell lines byexposure to ionizing radiation, inhibitors of DNAreplication and chromatin-modifying treatments. Thesetreatments caused activation of the DNA damage-responsemachinery, which is initiated by ATM (ataxia telangiectasiamutated) and ATR (ATM- and Rad3-related) proteinkinases, and resulted in NKG2D ligand expression, suchthat inhibition of the mentioned kinases prevented ligandupregulation. Thus, it was concluded that the DNAdamage response acts not only by arresting cell cycle,enhancing DNA reparation or triggering apoptosis, but also
Figure 3. Roles of NKG2D ligands in cancer immunology. Left: immunostimulating properties of NKG2D ligands. Right:immunosuppressive mechanisms shown by NKG2D ligands. Data from studies in mice and in humans about effects of NKG2D ligands onNK, CD4+ and CD8+ T cells have been summarized in the figure.DAP: DNAX-activating protein; NK: Natural killer.
NK
CD8sustainedexpression
shedding
CD4
DAP10DAP12
unidentifiedreceptor
8. Inhibition ofT cell proliferation
NKG2DNKG2Dligand
Tumor cell
1. Promote sensitivityto NK cells
2. Promote survival/expansion of CD8+ T cells
3. Development of memory responses
4. Protection from de novo tumorigenesis
7. Expansion ofimmunosuppressiveCD4+NKG2D+ cells
6. Downregulationof NKG2D
5. Decreased levelsof adaptor proteins
Exp
ert O
pin.
Bio
l. T
her.
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
idad
De
Nav
arra
Fo
r pe
rson
al u
se o
nly.

Arina, Murillo, Dubrot, Azpilikueta, Alfaro, Pérez-Gracia, Bendandi, Palencia, Hervás-Stubbs & Melero
Expert Opin. Biol. Ther. (2007) 7(5) 609
by alerting the immune system to the presence ofpotentially dangerous cells.
Interestingly, the expression of Rae1 has been recentlyshown to be induced in tumor cells by the adenovirusserotype 5 E1A oncogene [65]. Given that adenoviruses cantransform in vitro human cells, but are not oncogenic inhumans, the authors hypothesize that sensitization of tumorcells by E1A to NK-mediated lysis can act protecting the hostfrom tumor development.
Notwithstanding their immunostimulatory properties, theexistence of many NKG2D ligand-expressing tumor cell linesand primary tumors indicates that response evasion is frequent.Indeed, tumors often subvert pre-existing natural hostmechanisms to render them favorable to their growth. In thecase of NKG2D-mediated protection, different immune escapemechanisms have also been described (Figure 3). There exists acorrelation between downregulation of NKG2D ontumor-infiltrating lymphocytes and peripheral blood T cellsand expression of MICA/B in the malignant cells of MIC+
tumor-bearing patients [66]. The same authors found that it wascirculating tumor-derived soluble MICA that caused thisdownregulation of NKG2D and the subsequent impairment intumor-specific effector T cell responsiveness. Shedding of bothMIC and ULBP2 proteins from the tumor cell surface seems toresult from the action of metalloproteinases [67,68]. Along thesame line, sustained expression of NKG2D ligands and,therefore, sustained stimulation of NKG2D promotedNKG2D downregulation and hyporesponsiveness of NK cellsin several mouse models. NK cells in NOD mice uniquelyexpress NKG2D ligands themselves and, as a consequence,exhibit a functional impairment of NK cell function [69].Transgenic mice expressing Rae1ε, either ubiquitously or in atissue-specific way, also shared the same phenotype and showeda higher susceptibility to skin cancer induced by chemicalcarcinogens [70]. Coudert et al. [71] demonstrated in a murinemodel that prolonged encounter with tumor cell-bound, butnot soluble, ligand can completely uncouple the NKG2Dreceptor from downstream signaling by decreasing protein lev-els of DAP10 and DAP12 adaptors.
A new immunoregulatory effect, mediated by NKG2Dand affecting CD4+NKG2D+ cells, has recently beendescribed. CD4+ T cells do not usually express NKG2D anduntil the publication of these new data, this case had onlybeen reported in patients with rheumatoid arthritis [72].However, in subsequent studies the same group found anincreased frequency of CD4+NKG2D+ cells in late-stagehuman tumor settings [73]. This finding led them to thedemonstration that TCR-activated and MICA-costimulatedCD4+NKG2D+ cells produce FasL, but are themselvesresistant to Fas-mediated effects. In turn, the released FasLacts in a paracrine way, inducing growth arrest of otherT cells that do not express NKG2D.
NKG2D ligands can also exert immunosuppressive effectsthrough non-NKG2D dependent mechanisms. Certainligands (H60 in mice and MICA in humans) strongly inhibit
T cell proliferation through an IL-10- but notNKG2D-mediated mechanism [74].
The tumor’s potential to adapt to the conditions imposedby the immune system is also shown by the fact that3-methylcholantrene (MCA)-induced sarcomas from‘non-NKG2D-edited’ mice (mice in which the NKG2Dsystem was not operational during carcinogenesis), such asthose treated with anti-NKG2D monoclonal antibody or inperforin-deficient mice, express Rae1 at higher frequenciesthan tumors derived from wild-type mice [75].
7. NK cells in immunotherapy of human malignancies
Data unambiguously linking NK cell involvement inantitumor surveillance and effector activity in humans arelimited because patients lacking NK cells or bearing perforinmutations that impair NK cytotoxic function die at an earlyage [1,15]. However, several lines of evidence suggest a similarrole to that displayed by murine NK cells:
• NK cells from cancer patients show impairedcytotoxicity, proliferation and response to IFNs. In linewith this observation, low levels of NK activity atdiagnosis in patients with cancer appear to predictsubsequent metastases [76].
• Administration of cytokines that activate endogenousNK cells, such as IL-2, achieved some success in patientswith renal cell carcinoma [77] and metastaticmelanoma [78,79]; however, IL-2 treatment-accompanyingtoxicity limits its therapeutical potential [79]. This problemcan be slightly ameliorated by a combination of IFN-α andlow-dose IL-2, with similar results to those obtained withhigh-dose IL-2 alone [80]. IFN-α has also proved useful forthe treatment of chronic myelogenous leukemia [81].
• Bispecific monoclonal antibodies that recognize both theHodgkin’s disease-associated antigen CD30 and the NK cellreceptor CD16 have resulted in several NK cell-mediatedclinical responses in Hodgkin’s disease patients [82].
• The administration of ex vivo-expanded autologouslymphokine-activated killer cells has been used in clinicaltrials [83]; however, this strategy is controversial becausethese cells localize poorly to tumors.
• NK cell infiltration is a positive prognostic parameter inseveral cancer types, including gastric carcinoma, squamouscell lung carcinoma and colorectal cancer.
• HLA-matched allogeneic hematopoietic transplantation isused in the clinical practice to treat leukemia.
A revolutionary report by Ruggeri et al. [84] showed thathaploidentical transplant, with MHC-I mismatches,generates an alloreactive NK cell response towards thegraft-versus-host direction, eradicates leukemia, improvesengraftment and protects from T cell-mediatedgraft-versus-host disease. A mismatch in the HLA-C genesstudied, and consequently in graft-versus-host alloreactivity,
Exp
ert O
pin.
Bio
l. T
her.
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
idad
De
Nav
arra
Fo
r pe
rson
al u
se o
nly.

Cellular liaisons of natural killer lymphocytes in immunology and immunotherapy of cancer
610 Expert Opin. Biol. Ther. (2007) 7(5)
was necessary for this favorable outcome. The effectsdescribed were attributed to the killing of hostantigen-presenting cells (APCs) and leukemia cells byalloreactive NK cells, which was demonstrated in a mousemodel. APCs were previously known to initiategraft-versus-host disease in allogeneic bone marrowtransplantation by presenting host alloantigens to donorT cells [85]. The results obtained by Ruggeri and colleaguesare controversial to some extent because another report [86]
did not find advantage for KIR-ligand incompatibility inunrelated donor transplants, although in a slightly differentclinical setting. However, a new clinical study byGiebel et al. [87] supported the conclusions obtained inRuggeri’s work. Discrepancy with Davies et al. was attributedto a conscientious pretransplantation lymphoablativeconditioning and a larger number of transplanted cells in thework by Giebel et al. In addition, an interesting study byMiller et al. [88] reports on the feasibility and antitumoreffects of an intense immunosuppressive regimen followed byadoptive transfer of haploidentical NK cells in poor prognosisacute myeloid leukemia patients. As seen in the previousreports by Ruggeri and Giebel, the use ofKIR-ligand-mismatched donors significantly improved thetherapeutic effect, achieving higher complete remission rates.
The availability of clinical-grade kits based onimmunomagnetic selection to large-scale purify NK cells incell therapy facilities will foster investigation into adoptivetransfer of activated NK cells in different clinical trial settings,including bone marrow transplantation.
8. Expert opinion and conclusion
NK cells are an immune population with inducible ability tolyse transformed and infected cells. Persistent appearance ofNK cells in studies of immunosurveillance and theirinvolvement in the antitumor activity of variousimmunotherapeutic strategies indicate a potential for therapy.The mechanisms governing NK cell function can beartificially manipulated to enhance activation and cytotoxicityby either suppressing inhibitory receptors or stimulatingactivatory receptors.
In the authors’ experiments, a transplantable set ofMCA-induced sarcomas is spontaneously rejected inimmunocompetent mice in a NK cell-dependent fashion.Interestingly, T cell depletion also permits tumorprogression. Therefore, the immune system behaves in aconcerted and integrated fashion in which NK cells play anecessary role cooperating with other leukocytes, includingT cells. NK cell functions in experimental systems, such as theauthors’, include direct cytotoxicity and pro-inflammatoryfunctions (mainly early antigen-independent secretion of
IFN-γ and TNF). Direct cytotoxicity represents a potentialsource of antigens for cross-presentation by surroundingDCs [89], and pro-inflammatory cytokines are known toinfluence both T and DCs in order to optimize the overallcellular immune response.
A potential drawback for targeted translational research isthat the molecular network of inhibitory NK receptors isevolutionarily divergent between rodents and primates. Themain receptors surveying MHC class I expression on targetcells in mice are C-type lectins, whereas the immunoglobulinsuperfamily molecules predominate in humans. Althoughthere is much functional resemblance, this implies that animalexperimentation on mouse tumors with agents such asmonoclonal antibodies blocking mouse NK inhibitoryreceptors will not find easy direct translation into the clinic.In spite of this caveat, humanized monoclonal antibodiesinterfering with inhibitory KIRs and with NKG2A areadvanced in the process of being taken to the clinic [90].
Knowledge of the receptor systems and signaling pathwaysinvolved in NK cell activation offers opportunity fortherapeutics. NK cell functions can be enhanced by usingsoluble cytokines, such as IL-15, monoclonal antibodiesblocking inhibitory receptors [90,91], manipulating theNKG2D pathway of NK cell activation, and by adoptivetransfer of ex vivo activated NK cells. In the case of adoptivetransfer, the intratumoral route of administration should beexplored for solid malignancies.
Many molecular mechanisms of NK cells are shared byNKT cells, a subset of T cells bearing and invariant antigenreceptor [92]. With regard to tumor immunology, these cellshave been implicated in therapeutic antitumor immunity aswell as in immune evasion via their production of IL-13 [93].NKT cells also participate in bidirectional interactions withDCs that have been revised elsewhere [94].
Two additional important points to be considered are thecross-talk of NK cells with other leukocytes and the broadexpression of molecules originally studied on NK cells that areshared by many T cell subtypes and DCs. Therefore,therapeutic interventions acting on these molecules provideinteresting opportunities to be integrated in combinatorialimmunotherapy strategies.
Acknowledgements
Financial support was from Ministerio de Educación yCiencia (MEC-SAF2005-03131), Departamento deEducación del Gobierno de Navarra, Redes Temáticas deInvestigación Cooperativa RETIC, FIS PI060932 and ‘UTEfor project FIMA’. OM and AA are recipients of scholarshipsfrom Ministerio de Educación (MEC) and Fondo deInvestigación Sanitaria (BEFI).
Exp
ert O
pin.
Bio
l. T
her.
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
idad
De
Nav
arra
Fo
r pe
rson
al u
se o
nly.

Arina, Murillo, Dubrot, Azpilikueta, Alfaro, Pérez-Gracia, Bendandi, Palencia, Hervás-Stubbs & Melero
Expert Opin. Biol. Ther. (2007) 7(5) 611
BibliographyPapers of special note have been highlighted as either of interest (•) or of considerable interest (••) to readers.
1. WALLACE ME, SMYTH MJ: The role of natural killer cells in tumour control-effectors and regulators of adaptive immunity. Springer Semin. Immunopathol. (2005) 27(1):49-64.
2. WU J, LANIER LL: Natural killer cells and cancer. Adv. Cancer Res. (2003) 90:127-156.
3. RAULET DH: Natural killer cells. In: Fundamental Immunology. Paul WE (Ed.), Lippincott Williams & Wilkins, Philadelphia, PA, USA (2003):365-391.
4. DI SANTO JP: Natural killer cell developmental pathways: a question of balance. Annu. Rev. Immunol. (2006) 24:257-286.
5. CHAN CW, CRAFTON E, FAN HN et al.: Interferon-producing killer dendritic cells provide a link between innate and adaptive immunity. Nat. Med. (2006) 12(2):207-213.
• See [6].
6. TAIEB J, CHAPUT N, MENARD C et al.: A novel dendritic cell subset involved in tumour immunosurveillance. Nat. Med. (2006) 12(2):214-219.
• References [5] and [6] introduce IKDCs as a newly characterized immune subpopulation, different from NK cells and DCs, but sharing features of both cell types.
7. LOPEZ-BOTET M, PEREZ-VILLAR JJ, CARRETERO M et al.: Structure and function of the CD94 C-type lectin receptor complex involved in recognition of HLA class I molecules. Immunol. Rev. (1997) 155:165-174.
8. MAK TW, SAUNDERS M: Bridging innate and adaptive immunity: NK, γδ T, and NKT cells. In: The Immune Response: Basic and Clinical Principles. Mak TW, Saunders M (Eds), Academic Press, San Diego, CA, USA (2005).
9. TAKEDA K, HAYAKAWA Y, SMYTH MJ et al.: Involvement of tumour necrosis factor-related apoptosis-inducing ligand in surveillance of tumour metastasis by liver natural killer cells. Nat. Med. (2001) 7(1):94-100.
10. CRETNEY E, TAKEDA K, YAGITA H et al.: Increased susceptibility to tumour initiation and metastasis in TNF-related apoptosis-inducing ligand-deficient mice. J. Immunol. (2002) 168(3):1356-1361.
11. SMYTH MJ, TAKEDA K, HAYAKAWA Y et al.: Nature’s TRAIL-on a path to cancer immunotherapy. Immunity (2003) 18(1):1-6.
12. SCREPANTI V, WALLIN RP, LJUNGGREN HG, GRANDIEN A: A central role for death receptor-mediated apoptosis in the rejection of tumours by NK cells. J. Immunol. (2001) 167(4):2068-2073.
13. YOKOYAMA WM: Natural killer cells. In: Fundamental Immunology. Paul WE (Ed.), Lippincott Williams & Wilkins, Philadelphia, PA, USA (1999):575-603.
14. RODA JM, PARIHAR R, LEHMAN A et al.: Interleukin-21 enhances NK cell activation in response to antibody-coated targets. J. Immunol. (2006) 177(1):120-129.
15. SMYTH MJ, HAYAKAWA Y, TAKEDA K, YAGITA H: New aspects of natural-killer-cell surveillance and therapy of cancer. Nat. Rev. Cancer (2002) 2(11):850-861.
16. DUNN GP, KOEBEL CM, SCHREIBER RD: Interferons, immunity and cancer immunoediting. Nat. Rev. Immunol. (2006) 6(11):836-848.
17. KARRE K, LJUNGGREN HG, PIONTEK G, KIESSLING R: Selective rejection of H-2-deficient lymphoma variants suggests alternative immune defence strategy. Nature (1986) 319(6055):675-678.
18. MANDELBOIM O, LIEBERMAN N, LEV M et al.: Recognition of haemagglutinins on virus-infected cells by NKp46 activates lysis by human NK cells. Nature (2001) 409(6823):1055-1060.
19. LANIER LL: NK cell recognition. Annu. Rev. Immunol. (2005) 23:225-274.
20. LEE N, LLANO M, CARRETERO M et al.: HLA-E is a major ligand for the natural killer inhibitory receptor CD94/NKG2A. Proc. Natl. Acad. Sci. USA (1998) 95(9):5199-5204.
21. ITO M, MARUYAMA T, SAITO N et al.: Killer cell lectin-like receptor G1 binds three members of the classical cadherin family to inhibit NK cell cytotoxicity. J. Exp. Med. (2006) 203(2):289-295.
22. ANFOSSI N, ANDRE P, GUIA S et al.: Human NK cell education by inhibitory receptors for MHC class I. Immunity (2006) 25(2):331-342.
23. BOTTINO C, MORETTA L, MORETTA A: NK cell activating receptors and tumour recognition in humans. Curr. Top. Microbiol. Immunol. (2006) 298:175-182.
24. RAULET DH: Roles of the NKG2D immunoreceptor and its ligands. Nat. Rev. Immunol. (2003) 3(10):781-790.
25. FARAG SS, CALIGIURI MA: Human natural killer cell development and biology. Blood Rev. (2006) 20(3):123-137.
26. ARNON TI, LEV M, KATZ G et al.: Recognition of viral hemagglutinins by NKp44 but not by NKp30. Eur. J. Immunol. (2001) 31(9):2680-2689.
27. ARAMBURU J, BALBOA MA, RODRIGUEZ A et al.: Stimulation of IL-2-activated natural killer cells through the Kp43 surface antigen upregulates TNF-alpha production involving the LFA-1 integrin. J. Immunol. (1993) 151(7):3420-3429.
28. OSMAN MS, BURSHTYN DN, KANE KP: Activating Ly-49 receptors regulate LFA-1-mediated adhesion by NK cells. J. Immunol. (2007) 178(3):1261-1267.
29. BARBER DF, FAURE M, LONG EO: LFA-1 contributes an early signal for NK cell cytotoxicity. J. Immunol. (2004) 173(6):3653-3659.
30. MELERO I, BALBOA MA, ALONSO JL et al.: Signalling through the LFA-1 leucocyte integrin actively regulates intercellular adhesion and tumour necrosis factor-alpha production in natural killer cells. Eur. J. Immunol. (1993) 23(8):1859-1865.
31. KELLY JM, DARCY PK, MARKBY JL et al.: Induction of tumour-specific T cell memory by NK cell-mediated tumour rejection. Nat. Immunol. (2002) 3(1):83-90.
•• One of the first experimental evidences of the ability of NK cells to mediate the establishment of adaptive immunity against tumours.
32. KELLY JM, TAKEDA K, DARCY PK, YAGITA H, SMYTH MJ: A role for IFN-gamma in primary and secondary immunity generated by NK cell-sensitive tumour-expressing CD80 in vivo. J. Immunol. (2002) 168(9):4472-4479.
33. LEIBSON PJ: Signal transduction during natural killer cell activation: inside the mind of a killer. Immunity (1997) 6(6):655-661.
Exp
ert O
pin.
Bio
l. T
her.
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
idad
De
Nav
arra
Fo
r pe
rson
al u
se o
nly.

Cellular liaisons of natural killer lymphocytes in immunology and immunotherapy of cancer
612 Expert Opin. Biol. Ther. (2007) 7(5)
34. KALINSKI P, MAILLIARD RB, GIERMASZ A et al.: Natural killer-dendritic cell cross-talk in cancer immunotherapy. Expert Opin. Biol. Ther. (2005) 5(10):1303-1315.
35. DEGLI-ESPOSTI MA, SMYTH MJ: Close encounters of different kinds: dendritic cells and NK cells take centre stage. Nat. Rev. Immunol. (2005) 5(2):112-124.
36. FERLAZZO G, PACK M, THOMAS D et al.: Distinct roles of IL-12 and IL-15 in human natural killer cell activation by dendritic cells from secondary lymphoid organs. Proc. Natl. Acad. Sci. USA (2004) 101(47):16606-16611.
37. GRANUCCI F, ZANONI I, PAVELKA N et al.: A contribution of mouse dendritic cell-derived IL-2 for NK cell activation. J. Exp. Med. (2004) 200(3):287-295.
38. BORG C, JALIL A, LADERACH D et al.: NK cell activation by dendritic cells (DCs) requires the formation of a synapse leading to IL-12 polarization in DCs. Blood (2004) 104(10):3267-3275.
39. TERME M, TOMASELLO E, MARUYAMA K et al.: IL-4 confers NK stimulatory capacity to murine dendritic cells: a signalling pathway involving KARAP/DAP12-triggering receptor expressed on myeloid cell 2 molecules. J. Immunol. (2004) 172(10):5957-5966.
40. SEMINO C, ANGELINI G, POGGI A, RUBARTELLI A: NK/iDC interaction results in IL-18 secretion by DCs at the synaptic cleft followed by NK cell activation and release of the DC maturation factor HMGB1. Blood (2005) 106(2):609-616.
41. PICCIOLI D, SBRANA S, MELANDRI E, VALIANTE NM: Contact-dependent stimulation and inhibition of dendritic cells by natural killer cells. J. Exp. Med. (2002) 195(3):335-341.
42. FERLAZZO G, TSANG ML, MORETTA L et al.: Human dendritic cells activate resting natural killer (NK) cells and are recognized via the NKp30 receptor by activated NK cells. J. Exp. Med. (2002) 195(3):343-351.
43. GEROSA F, BALDANI-GUERRA B, NISII C et al.: Reciprocal activating interaction between natural killer cells and dendritic cells. J. Exp. Med. (2002) 195(3):327-333.
44. POGGI A, CAROSIO R, SPAGGIARI GM et al.: NK cell activation by dendritic cells is dependent on LFA-1-mediated induction of calcium-calmodulin kinase II: inhibition by HIV-1 Tat C-terminal domain. J. Immunol. (2002) 168(1):95-101.
45. HAYAKAWA Y, SCREPANTI V, YAGITA H et al.: NK cell TRAIL eliminates immature dendritic cells in vivo and limits dendritic cell vaccination efficacy. J. Immunol. (2004) 172(1):123-129.
46. DELLA CHIESA M, VITALE M, CARLOMAGNO S et al.: The natural killer cell-mediated killing of autologous dendritic cells is confined to a cell subset expressing CD94/NKG2A but lacking inhibitory killer Ig-like receptors. Eur. J. Immunol. (2003) 33(6):1657-1666.
47. PAN PY, GU P, LI Q et al.: Regulation of dendritic cell function by NK cells: mechanisms underlying the synergism in the combination therapy of IL-12 and 4-1BB activation. J. Immunol. (2004) 172(8):4779-4789.
48. MOCIKAT R, BRAUMULLER H, GUMY A et al.: Natural killer cells activated by MHC class I(low) targets prime dendritic cells to induce protective CD8 T cell responses. Immunity (2003) 19(4):561-569.
•• In this paper the mechanisms leading to the induction of protective T cell responses against MHC-Ilow targets are dissected and found to include the sequential activation of NK cells, DCs and CD8 T cells.
49. MARTIN-FONTECHA A, THOMSEN LL, BRETT S et al.: Induced recruitment of NK cells to lymph nodes provides IFN-gamma for T(H)1 priming. Nat. Immunol. (2004) 5(12):1260-1265.
•• This work highlights the importance of NK cells as an early source of IFN-γ, necessary for the induction of Th1 responses.
50. STRBO N, OIZUMI S, SOTOSEK-TOKMADZIC V, PODACK ER: Perforin is required for innate and adaptive immunity induced by heat-shock protein gp96. Immunity (2003) 18(3):381-390.
51. WESTWOOD JA, KELLY JM, TANNER JE et al.: Cutting edge: novel priming of tumour-specific immunity by NKG2D-triggered NK cell-mediated tumour rejection and Th1-independent CD4+ T cell pathway. J. Immunol. (2004) 172(2):757-761.
52. GROH V, RHINEHART R, SECRIST H et al.: Broad tumour-associated expression and recognition by tumour-derived gamma delta T cells of MICA and MICB. Proc. Natl. Acad. Sci. USA (1999) 96(12):6879-6884.
53. DIEFENBACH A, JAMIESON AM, LIU SD, SHASTRI N, RAULET DH: Ligands for the murine NKG2D receptor: expression by tumour cells and activation of NK cells and macrophages. Nat. Immunol. (2000) 1(2):119-126.
54. UPSHAW JL, LEIBSON PJ: NKG2D-mediated activation of cytotoxic lymphocytes: unique signalling pathways and distinct functional outcomes. Semin. Immunol. (2006) 18(3):167-175.
55. REGUNATHAN J, CHEN Y, WANG D, MALARKANNAN S: NKG2D receptor-mediated NK cell function is regulated by inhibitory Ly49 receptors. Blood (2005) 105(1):233-240.
56. CARBONE E, NERI P, MESURACA M et al.: HLA class I, NKG2D, and natural cytotoxicity receptors regulate multiple myeloma cell recognition by natural killer cells. Blood (2005) 105(1):251-258.
57. DIEFENBACH A, JENSEN ER, JAMIESON AM, RAULET DH: Rae1 and H60 ligands of the NKG2D receptor stimulate tumour immunity. Nature (2001) 413(6852):165-171.
58. CERWENKA A, BARON JL, LANIER LL: Ectopic expression of retinoic acid early inducible-1 gene (RAE-1) permits natural killer cell-mediated rejection of a MHC class I-bearing tumour in vivo. Proc. Natl. Acad. Sci. USA (2001) 98(20):11521-11526.
• References [57] and [58] are the first papers linking expression of NKG2D ligands and tumour rejection in vivo.
59. DIEFENBACH A, HSIA JK, HSIUNG MY, RAULET DH: A novel ligand for the NKG2D receptor activates NK cells and macrophages and induces tumour immunity. Eur. J. Immunol. (2003) 33(2):381-391.
60. SMYTH MJ, SWANN J, CRETNEY E et al.: NKG2D function protects the host from tumour initiation. J. Exp. Med. (2005) 202(5):583-588.
61. CONEJO-GARCIA JR, BENENCIA F, COURREGES MC et al.: Ovarian carcinoma expresses the NKG2D ligand Letal and promotes the survival and expansion of CD28- antitumour T cells. Cancer Res. (2004) 64(6):2175-2182.
Exp
ert O
pin.
Bio
l. T
her.
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
idad
De
Nav
arra
Fo
r pe
rson
al u
se o
nly.

Arina, Murillo, Dubrot, Azpilikueta, Alfaro, Pérez-Gracia, Bendandi, Palencia, Hervás-Stubbs & Melero
Expert Opin. Biol. Ther. (2007) 7(5) 613
62. GASSER S, RAULET DH: The DNA damage response arouses the immune system. Cancer Res. (2006) 66(8):3959-3962.
63. GIRARDI M, OPPENHEIM DE, STEELE CR et al.: Regulation of cutaneous malignancy by gammadelta T cells. Science (2001) 294(5542):605-609.
64. GASSER S, ORSULIC S, BROWN EJ, RAULET DH: The DNA damage pathway regulates innate immune system ligands of the NKG2D receptor. Nature (2005) 436(7054):1186-1190.
• Milestone article that finds the link between tumorigenesis and expression of NKG2D ligands.
65. ROUTES JM, RYAN S, MORRIS K et al.: Adenovirus serotype 5 E1A sensitizes tumour cells to NKG2D-dependent NK cell lysis and tumour rejection. J. Exp. Med. (2005) 202(11):1477-1482.
66. GROH V, WU J, YEE C, SPIES T: Tumour-derived soluble MIC ligands impair expression of NKG2D and T cell activation. Nature (2002) 419(6908):734-738.
• The first of a series of observations about the NKG2D system as a target for tumour manipulation conducive to immune evasion.
67. SALIH HR, RAMMENSEE HG, STEINLE A: Cutting edge: downregulation of MICA on human tumours by proteolytic shedding. J. Immunol. (2002) 169(8):4098-4102.
68. WALDHAUER I, STEINLE A: Proteolytic release of soluble UL16-binding protein 2 from tumour cells. Cancer Res. (2006) 66(5):2520-2526.
69. OGASAWARA K, HAMERMAN JA, HSIN H et al.: Impairment of NK cell function by NKG2D modulation in NOD mice. Immunity (2003) 18(1):41-51.
70. OPPENHEIM DE, ROBERTS SJ, CLARKE SL et al.: Sustained localized expression of ligand for the activating NKG2D receptor impairs natural cytotoxicity in vivo and reduces tumour immunosurveillance. Nat. Immunol. (2005) 6(9):928-937.
71. COUDERT JD, ZIMMER J, TOMASELLO E et al.: Altered NKG2D function in NK cells induced by chronic exposure to NKG2D ligand-expressing tumour cells. Blood (2005) 106(5):1711-1717.
72. GROH V, BRUHL A, EL-GABALAWY H, NELSON JL, SPIES T: Stimulation of T cell autoreactivity by anomalous expression of NKG2D and its MIC ligands in rheumatoid arthritis. Proc. Natl. Acad. Sci. USA (2003) 100(16):9452-9457.
73. GROH V, SMYTHE K, DAI Z, SPIES T: Fas ligand-mediated paracrine T cell regulation by the receptor NKG2D in tumour immunity. Nat. Immunol. (2006) 7(7):755-762.
74. KRIEGESKORTE AK, GEBHARDT FE, PORCELLINI S et al.: NKG2D-independent suppression of T cell proliferation by H60 and MICA. Proc. Natl. Acad. Sci. USA (2005) 102(33):11805-11810.
75. HAYAKAWA Y, SMYTH MJ: NKG2D and cytotoxic effector function in tumour immune surveillance. Semin. Immunol. (2006) 18(3):176-185.
76. WHITESIDE TL, HERBERMAN RB: Role of human natural killer cells in health and disease. Clin. Diagn. Lab. Immunol. (1994) 1(2):125-133.
77. MARGOLIN KA: Interleukin-2 in the treatment of renal cancer. Semin. Oncol. (2000) 27(2):194-203.
78. PHAN GQ, ATTIA P, STEINBERG SM, WHITE DE, ROSENBERG SA: Factors associated with response to high-dose interleukin-2 in patients with metastatic melanoma. J. Clin. Oncol. (2001) 19(15):3477-3482.
79. ROSENBERG SA, LOTZE MT, MUUL LM et al.: A progress report on the treatment of 157 patients with advanced cancer using lymphokine-activated killer cells and interleukin-2 or high-dose interleukin-2 alone. N. Engl. J. Med. (1987) 316(15):889-897.
80. BUZIO C, ANDRULLI S, SANTI R et al.: Long-term immunotherapy with low-dose interleukin-2 and interferon-alpha in the treatment of patients with advanced renal cell carcinoma. Cancer (2001) 92(9):2286-2296.
81. SILLA LM, WHITESIDE TL, BALL ED: The role of natural killer cells in the treatment of chronic myeloid leukaemia. J. Hematother. (1995) 4(4):269-279.
82. HARTMANN F, RENNER C, JUNG W et al.: Anti-CD16/CD30 bispecific antibody treatment for Hodgkin’s disease: role of infusion schedule and costimulation with cytokines. Clin. Cancer Res. (2001) 7(7):1873-1881.
83. YASUMURA S, LIN WC, HIRABAYASHI H et al.: Immunotherapy of liver metastases of human gastric carcinoma with IL-2-activated natural killer cells. Cancer Res. (1994) 54(14):3808-3816.
84. RUGGERI L, CAPANNI M, URBANI E et al.: Effectiveness of donor natural killer cell alloreactivity in mismatched hematopoietic transplants. Science (2002) 295(5562):2097-2100.
• This work supports haploidentical tranplantation in leukemia patients due to its high efficacy and clear advantages in the search for donors.
85. SHLOMCHIK WD, COUZENS MS, TANG CB et al.: Prevention of graft versus host disease by inactivation of host antigen-presenting cells. Science (1999) 285(5426):412-415.
86. DAVIES SM, RUGGIERI L, DEFOR T et al.: Evaluation of KIR ligand incompatibility in mismatched unrelated donor hematopoietic transplants. Killer immunoglobulin-like receptor. Blood (2002) 100(10):3825-3827.
87. GIEBEL S, LOCATELLI F, LAMPARELLI T et al.: Survival advantage with KIR ligand incompatibility in hematopoietic stem cell transplantation from unrelated donors. Blood (2003) 102(3):814-819.
88. MILLER JS, SOIGNIER Y, PANOSKALTSIS-MORTARI A et al.: Successful adoptive transfer and in vivo expansion of human haploidentical NK cells in patients with cancer. Blood (2005) 105(8):3051-3057.
• This study shows that haploidentical NK cells can persist and expand in vivo when they are infused following high-intensity immunosuppression regimes, and may have a role in the treatment of patients with hematological malignancies.
89. MELERO I, ARINA A, MURILLO O et al.: Immunogenic cell death and cross-priming are reaching the clinical immunotherapy arena. Clin. Cancer Res. (2006) 12(8):2385-2389.
90. MELERO I, HERVAS-STUBBS S, GLENNIE M, PARDOLL DM, CHEN L: Immunostimulatory monoclonal antibodies for cancer therapy. Nat. Rev. Cancer. (2007) 7(2):95-106.
Exp
ert O
pin.
Bio
l. T
her.
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
idad
De
Nav
arra
Fo
r pe
rson
al u
se o
nly.

Cellular liaisons of natural killer lymphocytes in immunology and immunotherapy of cancer
614 Expert Opin. Biol. Ther. (2007) 7(5)
91. MURILLO O, ARINA A, TIRAPU I et al.: Potentiation of therapeutic immune responses against malignancies with monoclonal antibodies. Clin. Cancer Res. (2003) 9(15):5454-5464.
92. GODFREY DI, KRONENBERG M: Going both ways: immune regulation via CD1d-dependent NKT cells. J. Clin. Invest. (2004) 114(10):1379-1388.
93. TERABE M, PARK JM, BERZOFSKY JA: Role of IL-13 in regulation of antitumour immunity and tumour growth. Cancer Immunol. Immunother. (2004) 53(2):79-85.
94. BENDELAC A, SAVAGE PB, TEYTON L: The biology of NKT cells. Annu. Rev. Immunol. (2006) 6:6.
95. SENTMAN CL, BARBER MA, BARBER A, ZHANG T: NK cell receptors as tools in cancer immunotherapy. Adv. Cancer Res. (2006) 95:249-292.
96. SMYTH MJ, CROWE NY, GODFREY DI: NK cells and NKT cells collaborate in host protection from methylcholanthrene-induced fibrosarcoma. Int. Immunol. (2001) 13(4):459-463.
97. ARGOV S, COCHRAN AJ, KARRE K, KLEIN GO, KLEIN G: Incidence and type of tumours induced in C57BL bg/bg mice and +/bg littermates by oral administration of DMBA. Int. J. Cancer (1981) 28(6):739-746.
98. GALLO-HENDRIKX E, COPPS J, PERCY D, CROY BA, WILDEMAN AG: Enhancement of pancreatic tumour metastasis in transgenic immunodeficient mice. Oncogene (1994) 9(10):2983-2990.
99. SHULTZ LD, SCHWEITZER PA, CHRISTIANSON SW et al.: Multiple defects in innate and adaptive immunologic function in NOD/LtSz-scid mice. J. Immunol. (1995) 154(1):180-191.
100. KIM S, IIZUKA K, AGUILA HL, WEISSMAN IL, YOKOYAMA WM: In vivo natural killer cell activities revealed by natural killer cell-deficient mice. Proc. Natl. Acad. Sci. USA (2000) 97(6):2731-2736.
101. VAN DEN BROEK ME, KAGI D, OSSENDORP F et al.: Decreased tumour surveillance in perforin-deficient mice. J. Exp. Med. (1996) 184(5):1781-1790.
102. KAPLAN DH, SHANKARAN V, DIGHE AS et al.: Demonstration of an interferon gamma-dependent tumour surveillance system in immunocompetent mice. Proc. Natl. Acad. Sci. USA (1998) 95(13):7556-7561.
103. OWEN-SCHAUB LB, VAN GOLEN KL, HILL LL, PRICE JE: Fas and Fas ligand interactions suppress melanoma lung metastasis. J. Exp. Med. (1998) 188(9):1717-1723.
104. STREET SE, CRETNEY E, SMYTH MJ: Perforin and interferon-gamma activities independently control tumour initiation, growth, and metastasis. Blood (2001) 97(1):192-197.
105. SHANKARAN V, IKEDA H, BRUCE AT et al.: IFN-gamma and lymphocytes prevent primary tumour development and shape tumour immunogenicity. Nature (2001) 410(6832):1107-1111.
106. SMYTH MJ, CRETNEY E, TAKEDA K et al.: Tumour necrosis factor-related apoptosis-inducing ligand (TRAIL) contributes to interferon gamma-dependent natural killer cell protection from tumour metastasis. J. Exp. Med. (2001) 193(6):661-670.
107. LJUNGGREN HG, KARRE K: Host resistance directed selectively against H-2-deficient lymphoma variants. Analysis of the mechanism. J. Exp. Med. (1985) 162(6):1745-1759.
108. GAO JX, LIU X, WEN J et al.: Two-signal requirement for activation and effector function of natural killer cell response to allogeneic tumour cells. Blood (2003) 102(13):4456-4463.
109. BRADLEY M, ZEYTUN A, RAFI-JANAJREH A, NAGARKATTI PS, NAGARKATTI M: Role of spontaneous and interleukin-2-induced natural killer cell activity in the cytotoxicity and rejection of Fas+ and Fas- tumour cells. Blood (1998) 92(11):4248-4255.
110. KARP SE, FARBER A, SALO JC et al.: Cytokine secretion by genetically modified nonimmunogenic murine fibrosarcoma. Tumour inhibition by IL-2 but not tumour necrosis factor. J. Immunol. (1993) 150(3):896-908.
111. ORTALDO JR, WINKLER-PICKETT RT, BERE EW JR et al.: In vivo hydrodynamic delivery of cDNA encoding IL-2: rapid, sustained redistribution, activation of mouse NK cells, and therapeutic potential in the absence of NKT cells. J. Immunol. (2005) 175(2):693-699.
112. DIVINO CM, CHEN SH, YANG W et al.: Antitumour immunity induced by interleukin-12 gene therapy in a metastatic model of breast cancer is mediated by natural killer cells. Breast Cancer Res. Treat. (2000) 60(2):129-134.
113. RODRIGUEZ-CALVILLO M, DUARTE M, TIRAPU I et al.: Upregulation of natural killer cells functions underlies the efficacy of intratumourally injected dendritic cells engineered to produce interleukin-12. Exp. Hematol. (2002) 30(3):195-204.
114. CHAPOVAL AI, FULLER JA, KREMLEV SG, KAMDAR SJ, EVANS R: Combination chemotherapy and IL-15 administration induce permanent tumour regression in a mouse lung tumour model: NK and T cell-mediated effects antagonized by B cells. J. Immunol. (1998) 161(12):6977-6984.
115. MEAZZA R, LOLLINI PL, NANNI P et al.: Gene transfer of a secretable form of IL-15 in murine adenocarcinoma cells: effects on tumorigenicity, metastatic potential and immune response. Int. J. Cancer (2000) 87(4):574-581.
116. VERA M, RAZQUIN N, PRIETO J et al.: Intratumoural injection of dendritic cells transduced by an SV40-based vector expressing interleukin-15 induces curative immunity mediated by CD8+ T lymphocytes and NK cells. Mol. Ther. (2005) 12(5):950-959.
117. WANG G, TSCHOI M, SPOLSKI R et al.: In vivo antitumour activity of IL-21 mediated by natural killer cells. Cancer Res. (2003) 63(24):9016-9022.
118. TAKAKI R, HAYAKAWA Y, NELSON A et al.: IL-21 enhances tumour rejection through a NKG2D-dependent mechanism. J. Immunol. (2005) 175(4):2167-2173.
119. SMYTH MJ, WALLACE ME, NUTT SL et al.: Sequential activation of NKT cells and NK cells provides effective innate immunotherapy of cancer. J. Exp. Med. (2005) 201(12):1973-1985.
Exp
ert O
pin.
Bio
l. T
her.
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
idad
De
Nav
arra
Fo
r pe
rson
al u
se o
nly.

Arina, Murillo, Dubrot, Azpilikueta, Alfaro, Pérez-Gracia, Bendandi, Palencia, Hervás-Stubbs & Melero
Expert Opin. Biol. Ther. (2007) 7(5) 615
120. SMYTH MJ, SWANN J, KELLY JM et al.: NKG2D recognition and perforin effector function mediate effective cytokine immunotherapy of cancer. J. Exp. Med. (2004) 200(10):1325-1335.
121. BRAUN SE, CHEN K, FOSTER RG et al.: The CC chemokine CK beta-11/MIP-3 beta/ELC/Exodus 3 mediates tumour rejection of murine breast cancer cells through NK cells. J. Immunol. (2000) 164(8):4025-4031.
122. MELERO I, JOHNSTON JV, SHUFFORD WW, MITTLER RS, CHEN L: NK1.1 cells express 4-1BB (CDw137) costimulatory molecule and are required for tumour immunity elicited by anti4-1BB monoclonal antibodies. Cell. Immunol. (1998) 190(2):167-172.
123. WILCOX RA, TAMADA K, STROME SE, CHEN L: Signalling through NK cell-associated CD137 promotes both helper function for CD8+ cytolytic T cells and responsiveness to IL-2 but not cytolytic activity. J. Immunol. (2002) 169(8):4230-4236.
124. ESPLUGUES E, VEGA-RAMOS J, CARTOIXA D et al.: Induction of tumour NK-cell immunity by anti-CD69 antibody therapy. Blood (2005) 105(11):4399-4406.
AffiliationAinhoa Arina, Oihana Murillo, Juan Dubrot, Arantza Azpilikueta, Carlos Alfaro, Jose L Pérez-Gracia, Maurizio Bendandi, Belen Palencia, Sandra Hervás-Stubbs & Ignacio Melero†1 MD, PhD†Author for correspondence1University of Navarra, Centro de Investigación Médica Aplicada (CIMA) and Clinica Universitaria, Gene Therapy Unit, Avda. Pio XII 55, 31008, Pamplona, SpainFax: +34 948194717;E-mail: [email protected]
Exp
ert O
pin.
Bio
l. T
her.
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Uni
vers
idad
De
Nav
arra
Fo
r pe
rson
al u
se o
nly.