cadena respiratoria fosforilacion oxidativa
TRANSCRIPT

Cadena Respiratoria(Cadena de Transporte de Electrones)
y
Fosforilación Oxidativa
MAOS-1
Unidad 3Horas 9,10 y
11

OBJETIVO ESPECÍFICO 6 CONTENIDO
Explicar las características de la CADENA RESPIRATORIA y de la FOSFORILACIÓN OXIDATIVA.
1.Características generales de la cadena
respiratoria. 2.Organización y función de los
componentes de la cadena respiratoria.
3.Acoplamiento entre la cadena respiratoria y la fosforilación oxidativa.
4.Energética de la fosforilación oxidativa. Relación P/O.
5.Inhibición de la cadena respiratoria, sitios de inhibición y ejemplos de inhibidores.
6.Inhibición de la fosforilación oxidativa, sitios de inhibición y ejemplos de inhibidores.
7.Mecanismo de acción de los desacopladores de la cadena respiratoria con la fosforilación oxidativa. Ejemplos.
3 horas
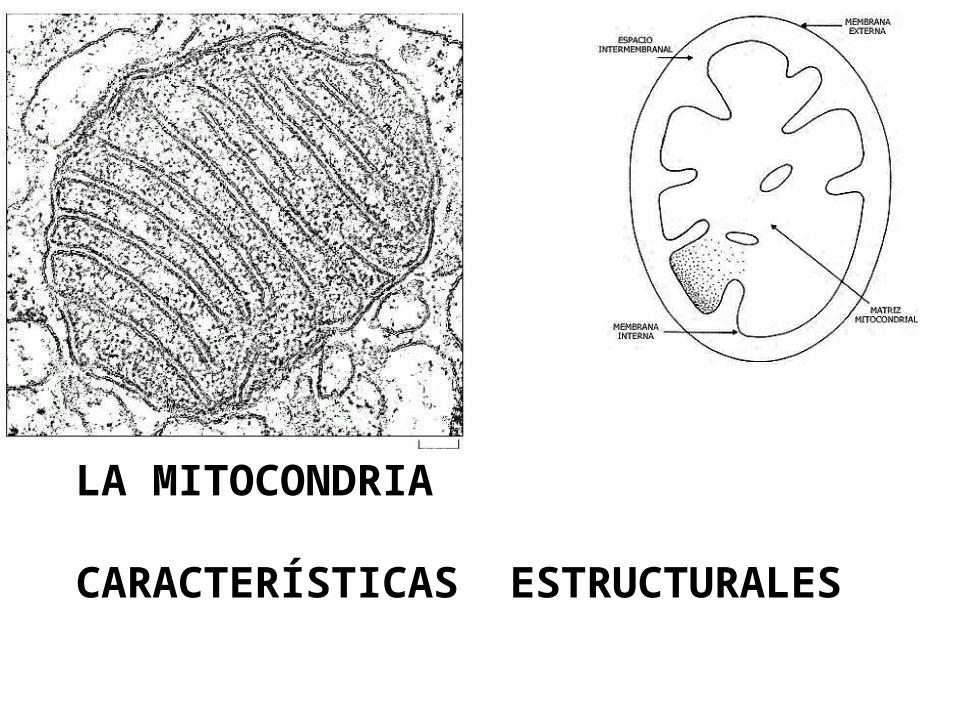
LA MITOCONDRIA
CARACTERÍSTICAS ESTRUCTURALES

Membrana externa (MME): La MME es muy permeable, a la mayor parte de las moléculas pequeñas e iones, debido a que contiene muchas copias de una proteína transportadora llamada Porina, que forma poros que permiten la libre difusión de moléculas hasta de 10kDa . Estas moléculas entran al EIM, pero la mayoría no atraviesa la impermeable MMI. Otras proteínas ubicadas en esta MME incluyen enzimas implicadas en el metabolismo de los lípidos. EIM: Es el espacio entre la MME y la MMI. El EIM es químicamente equivalente al del citosol. Matriz: este gran espacio interno contiene una mezcla altamente concentrada de cientos de enzimas, incluyendo aquellas requeridas para la oxidación del piruvato y los ácidos grasos y las del ciclo del acido cítrico. La matriz también contiene ADN mitocondrial, ribosomas mitocondriales ARNt, y varios enzimas requeridas para la expresión de los genes mitocondriales.
LA MITOCONDRIAPartes que la constituyen

Esta plegada formando invaginaciones llamadas crestas, lo que incrementa grandemente su área total. Es altamente especializada. Contiene una alta proporción de Cardiolipina un fosfolípido que ayuda a la MMI a ser especialmente impermeable a iones y moléculas polares. Es permeable sólo a moléculas pequeñas neutras, como el O2, H2O, y CO2.
Contiene proteínas con 3 tipos de funciones:
1) Las Enzimas que llevan a cabo la reacciones de oxidación de la cadena respiratoria, las cuales son esenciales para el proceso de fosforilación oxidativa, el cual genera la mayoría del ATP de la célula animal.
2) La ATP sintasa que produce ATP en la matriz (fosforilación oxidativa), y
3) Las Proteínas de Transporte (Translocasas), que permiten el pasaje de metabolitos a través de la MMI.
Membrana Mitocondrial interna (MMI)
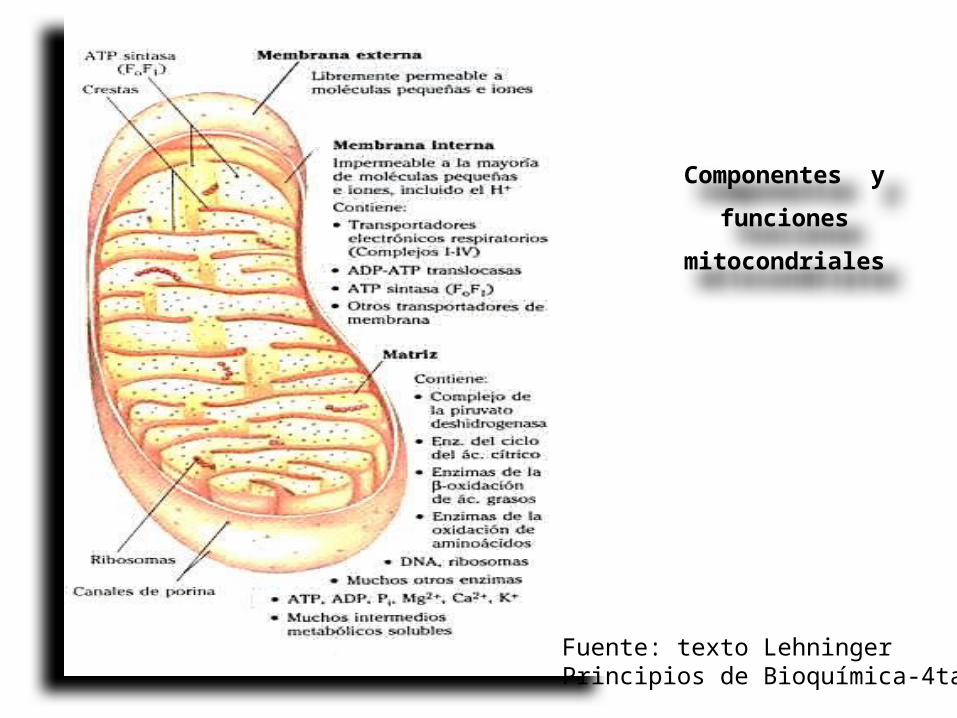
Componentes y funciones
mitocondriales
Fuente: texto Lehninger Principios de Bioquímica-4ta ed

la Respiración Celular.Se divide en 3 etapas para su estudio:
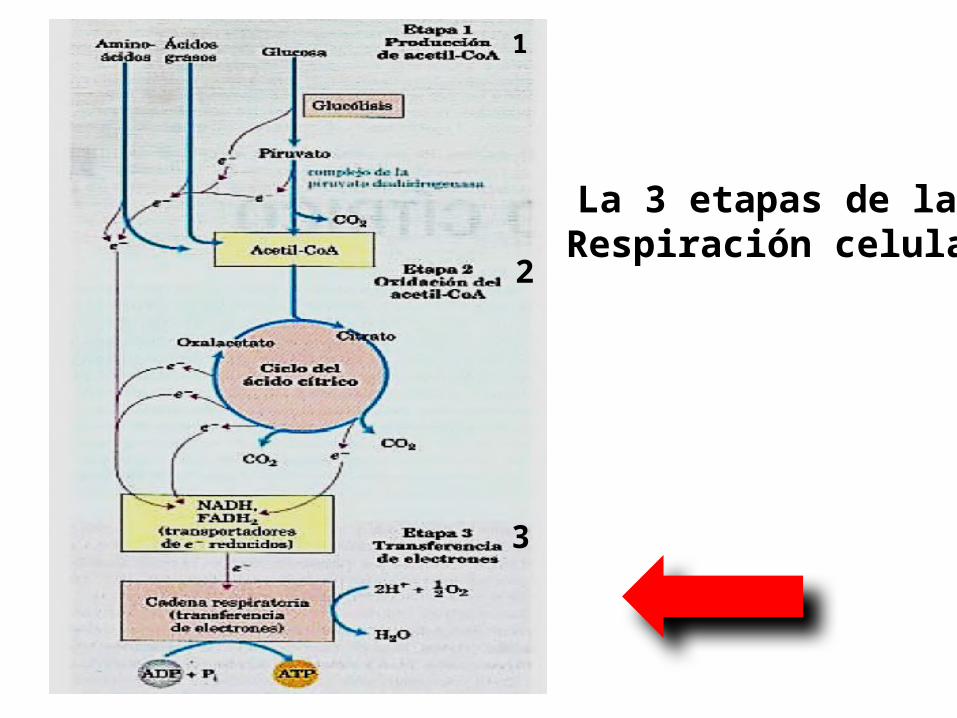
La 3 etapas de la Respiración celular
1
2
3
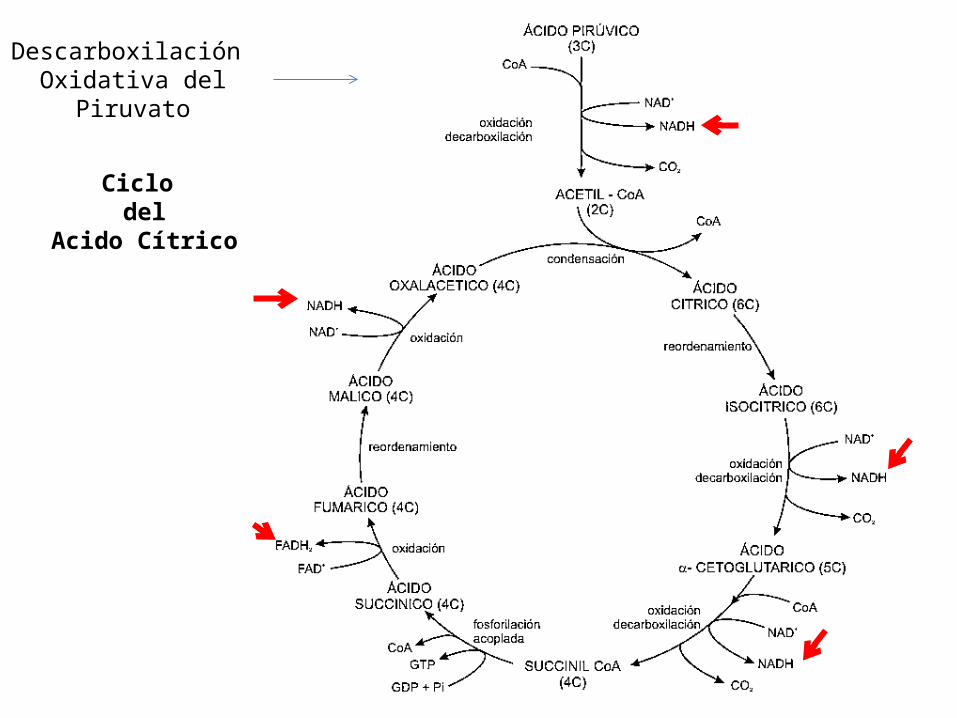
Ciclo del
Acido Cítrico
Descarboxilación Oxidativa del
Piruvato
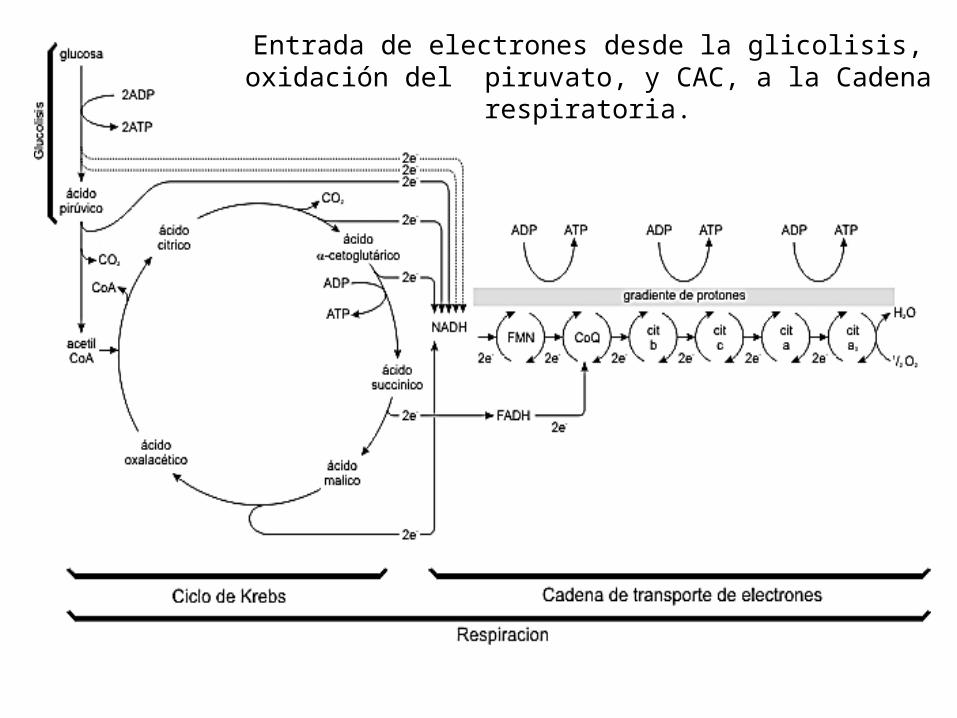
Entrada de electrones desde la glicolisis, oxidación del piruvato, y CAC, a la Cadena
respiratoria.

Es la fuente principal de energía para la síntesis de ATP en los animales y
otros organismos eucariontes.
Cadena Respiratoria mitocondrial
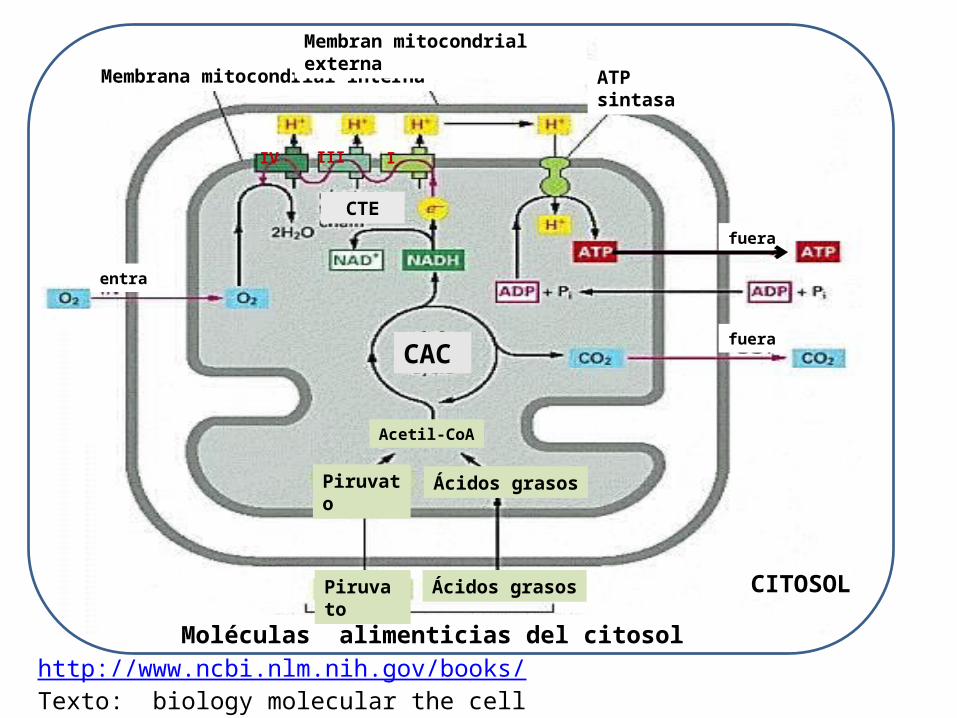
http://www.ncbi.nlm.nih.gov/books/Texto: biology molecular the cell
Moléculas alimenticias del citosol
Piruvato
Ácidos grasos
Acetil-CoA
ATP sintasa
Membrana mitocondrial internaMembran mitocondrial externa
fuera
fuera
entra
CTE
Piruvato
Ácidos grasos CITOSOL
CAC
IV III I
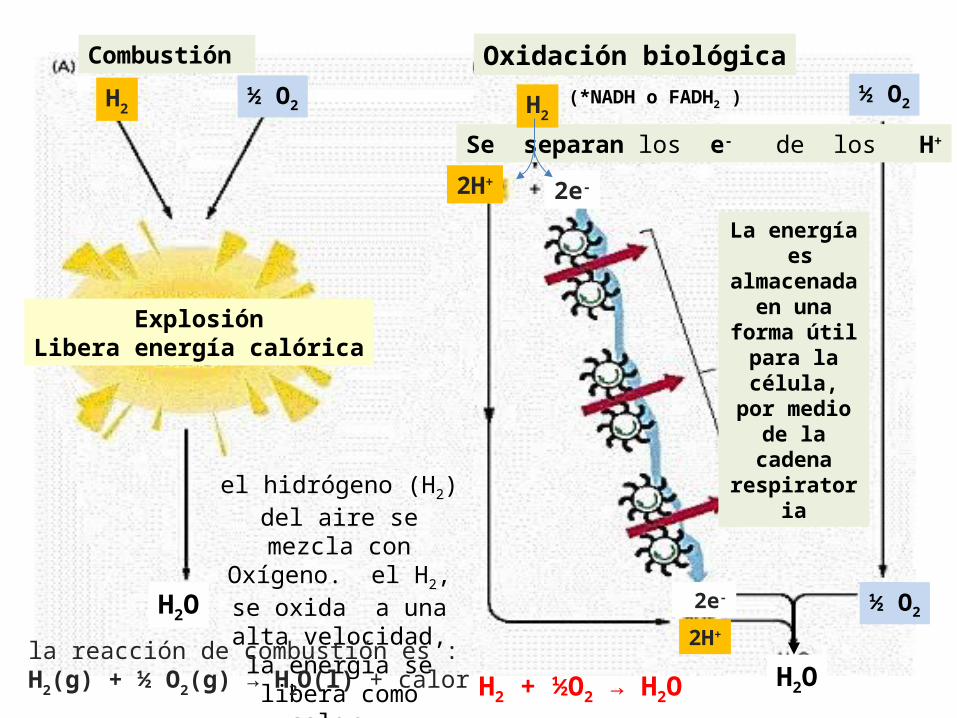
Oxidación biológica
Se separan los e- de los H+
La energía es
almacenada en una
forma útil para la célula,
por medio de la cadena
respiratoria
Combustión
el hidrógeno (H2) del aire se mezcla con
Oxígeno. el H2, se oxida a una alta velocidad, la energía se libera como
calor.
ExplosiónLibera energía calórica
la reacción de combustión es :H2(g) + ½ O2(g) → H2O(l) + calor H2 + ½O2 → H2O
H2
2H+ 2e-
2e-
2H+
½ O2
½ O2
H2O
H2 ½ O2
H2O
(*NADH o FADH2 )

1.Características generales de la Cadena respiratoria.
2.Organización y función de los componentes de la Cadena respiratoria.
3.Acoplamiento entre la Cadena respiratoria y la fosforilación oxidativa

Es un conjunto de cuatro (4) complejos enzimáticos firmemente unidos dentro de la MMI, además de Ubiquinona y Citocromo c, actuando todos en estrecha asociación, como transportadores electrónicos.
El nombre de Cadena Transportadora de Electrones(CTE) hace énfasis en el hecho de que el sistema es una cadena de reacciones redox en la que hay interacción sucesiva de transportadores que transfieren electrones.
La denominación de CADENA RESPIRATORIA (CR) hace referencia a que el transportador final de la cadena transfiere los electrones al oxígeno, es decir el sistema consume oxígeno.
Cadena Respiratoria (CR)Características de la

La CR REOXIDA las coenzimas reducidas NADH y FADH2, y los electrones que estas transportan , son conducidos al Oxígeno (O2), el ultimo aceptor de electrones.
Este proceso es esencial para el metabolismo ya que proporciona NAD+ y FAD que se puede dirigir a las vías metabólicas que los requieren para seguir funcionando, como por el ejemplo, el Ciclo del Ácido Cítrico.
Las Reacciones de Oxidorreducción que se dan en la CR, liberan energía la cual se utiliza para producir ATP.
Cadena Respiratoria (CR)Características de la

Existen 2 puntos de entrada a la CTE:Un Punto de entrada para la coenzima NADH (el complejo I) yUn Punto de entrada para la coenzima FADH2 (la CoQ)
Las reacciones de la CTE se ordenan según el valor de sus potenciales de reducción , encontrándose el menos positivo en el extremo izquierdo ( - 0,32 V), y el mas positivo a la derecha (+0,82 V) , por tanto los electrones fluyen sucesivamente a lo largo de la cadena hasta que finalmente son transferidos al oxigeno (O2).
Por tanto los electrones pasan a través de los 4 complejos desde los potenciales de reducción estándar mas bajos hacia los mas altos.
Contienen unas clases especiales de proteínas :1. FLAVOPROTEÍNAS2. PROTEÍNAS FE-S2. CITOCROMOS .
Características de la Cadena Respiratoria

Componentes
de la
Cadena Respiratoria
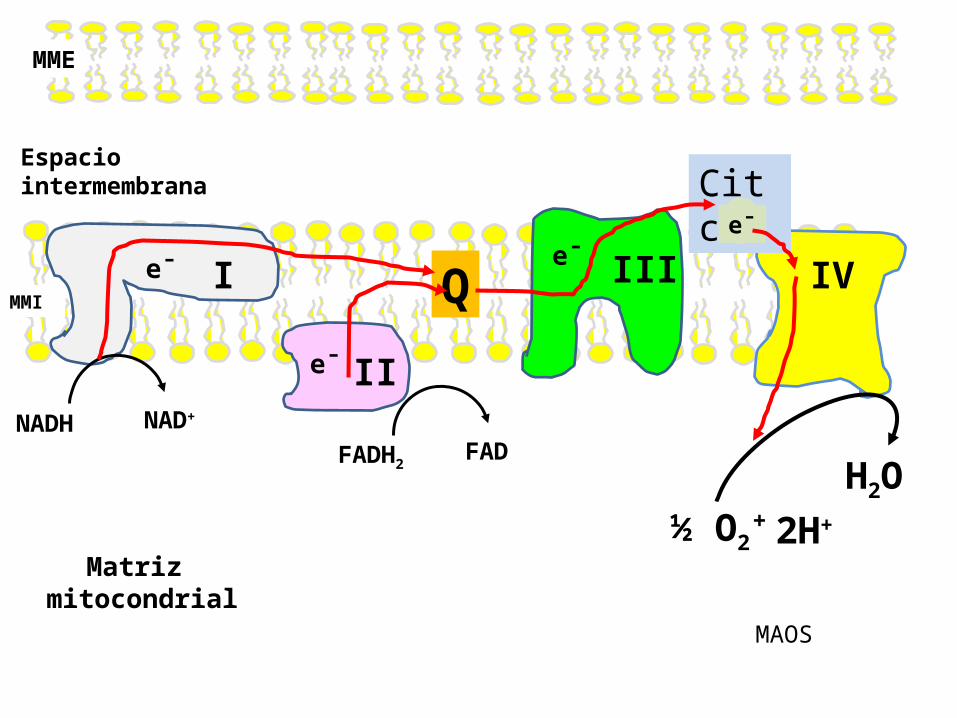
I
II
III IV
Cit c
Espaciointermembrana
Q
MME
MMI
NADH NAD+
FADH2 FAD
e¯
e¯
e¯e¯
Matriz mitocondrial
2H+
H2O½ O2
+
MAOS

(I) Flavoproteínas (fp)En la CR hay 2 flavoproteínas, estas contienen FMN o FAD fuertemente unidos, las cuales no se disocian de la enzima durante su oxidación y reducción, por lo que se comportan como grupos prostéticos. Estas son:
1. La enzima: NADH deshidrogenasa (también llamada NADH-Q oxidorreductasa o Complejo I) , contiene FMN que recibe los hidrógenos del NADH.
2. La enzima: Succinato deshidrogenasa(también llamada Complejo II), contiene FAD que recibe los hidrógenos del Succinato.
Las dos enzimas están fijadas a la MMI.
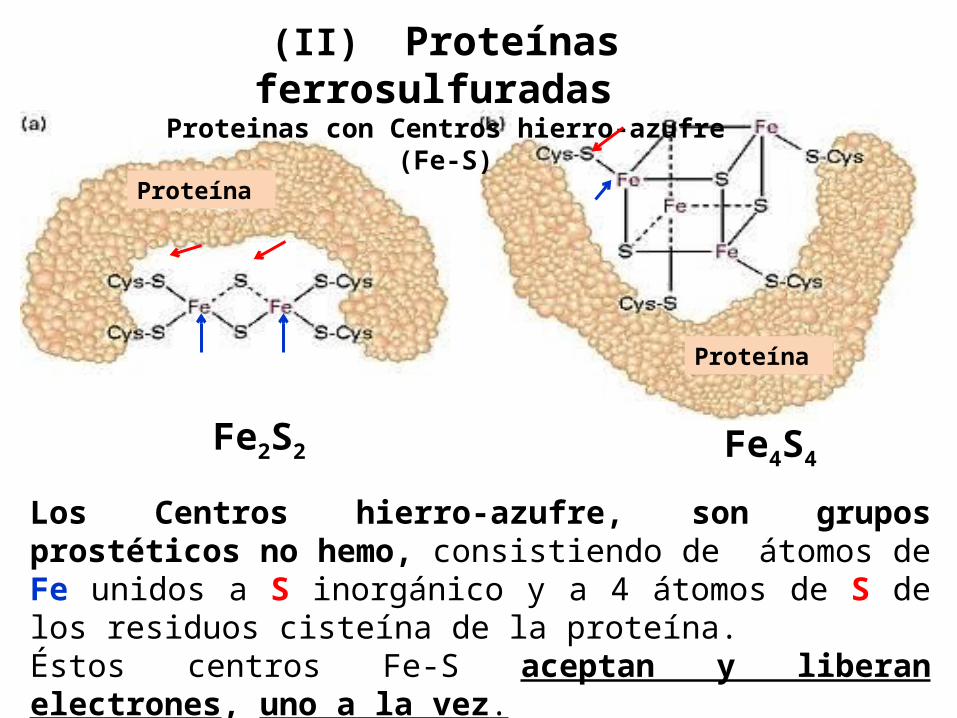
(II) Proteínas ferrosulfuradas
Proteinas con Centros hierro-azufre (Fe-S)
Los Centros hierro-azufre, son grupos prostéticos no hemo, consistiendo de átomos de Fe unidos a S inorgánico y a 4 átomos de S de los residuos cisteína de la proteína.Éstos centros Fe-S aceptan y liberan electrones, uno a la vez.
Fe4S4Fe2S2
Proteína
Proteína

Coenzima Qtambién nombrada como, Ubiquinona, y abreviada:
Q , CoQ10
La estructura de la Ubiquinona consiste en una 1,4 benzoquinona que contiene en los mamíferos una cadena lateral hidrófoba de 10 unidades de isopreno (Q10) , que la hace Soluble en los fosfolípidos en la MMI y, es capaz de difundir y moverse libremente a través de ésta.
Se le llama Ubiquinona ( por su ubicuidad , es una quinona que se encuentra en todas partes )
Los animales pueden sintetizarla Es el único transportador de electrones de la CR que
no es una proteína. Es un transportador de átomos de hidrógeno (protones
+ electrones), desde las flavoproteínas Complejo I y II, hacia los citocromos.
(III)
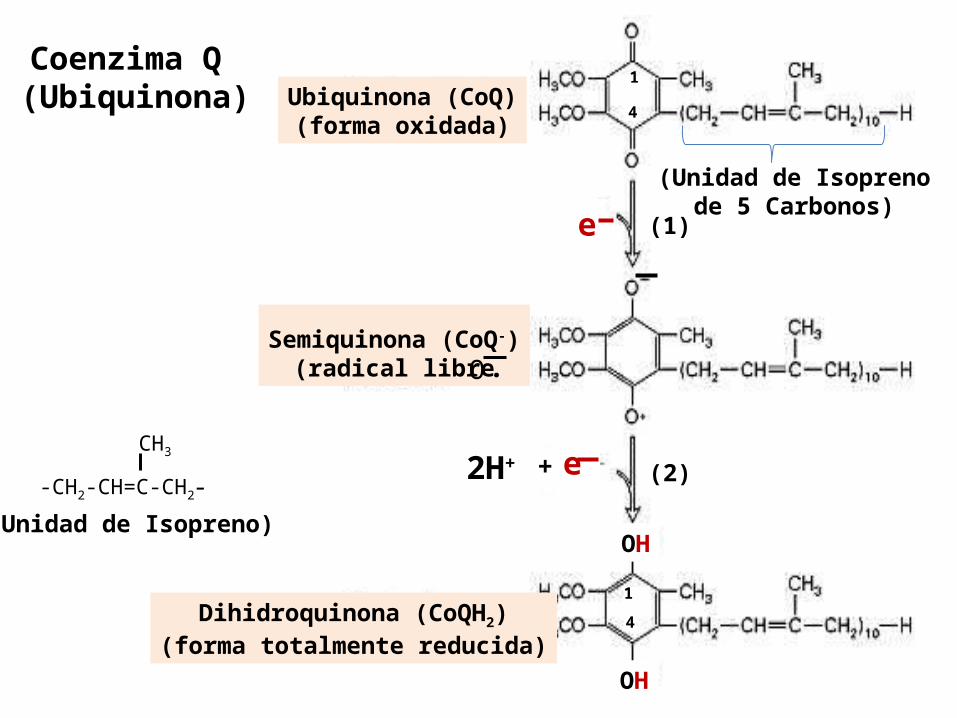
Coenzima Q (Ubiquinona) Ubiquinona (CoQ)
(forma oxidada)
Semiquinona (CoQ-)(radical libre
Dihidroquinona (CoQH2)(forma totalmente reducida)
e
e2H+ +
OH
OH
O.
(Unidad de Isoprenode 5 Carbonos)
-CH2-CH=C-CH2
CH3
(Unidad de Isopreno)-
1
4
(2)
(1)
14

(IV) Los Citocromos Un citocromo es una proteína transportadora de
electrones que contienen un grupo prostético hemo, con un átomo de hierro.
El átomo de hierro del hemo sufre una oxidorreducción reversible:
La unidireccionalidad del flujo de electrones en la CR esta dictada por la propiedad de los citocromos a tener diferentes E’o
Los electrones se mueven a través de los 5 tipos de citocromos de la CR en el siguiente orden:
b c1 c a a3
Fe+3 Fe+2
Hierro oxidado(férrico)
Hierro reducido(ferroso)
+ 1e- 1e
Reducción
Oxidación
Hay 5 tipos de citocromos en la CR.
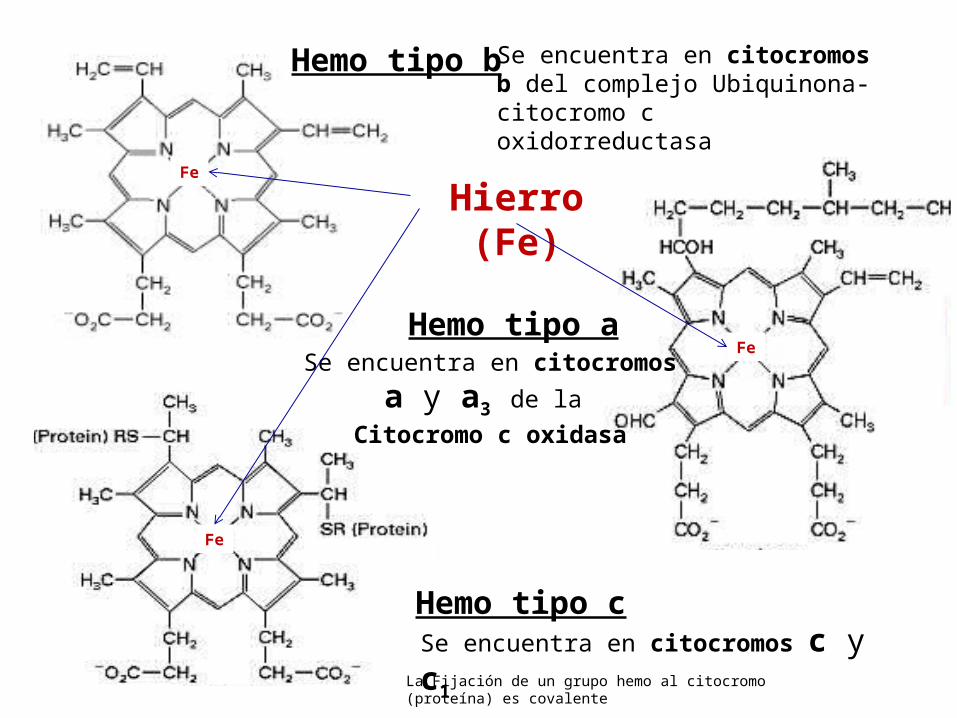
Hemo tipo bSe encuentra en citocromos b del complejo Ubiquinona-citocromo c oxidorreductasa
Hemo tipo a
Se encuentra en citocromos c y c1
Se encuentra en citocromosa y a3 de la
Citocromo c oxidasa
Hemo tipo c
Fe
Fe
Fe
Hierro (Fe)
La Fijación de un grupo hemo al citocromo (proteína) es covalente

A excepción de la CoQ y el Citocromo c el resto de los transportadores electrónicos se asocian en forma de grandes
Complejos enzimáticos
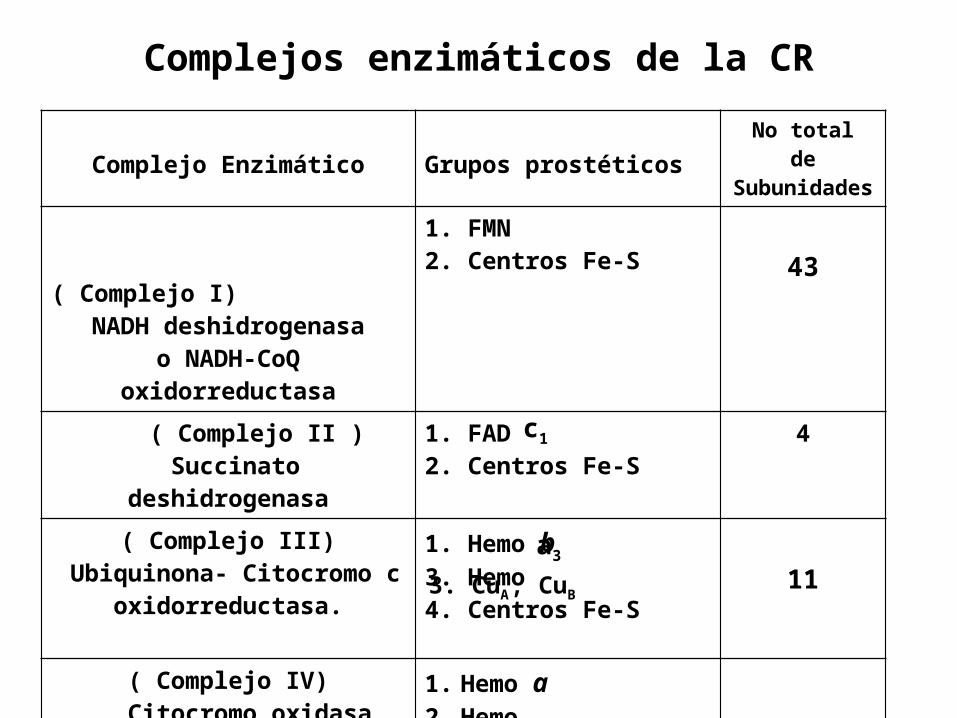
Complejo Enzimático Grupos prostéticosNo total
de Subunidades
( Complejo I)NADH deshidrogenasa
o NADH-CoQ oxidorreductasa
1. FMN2. Centros Fe-S 43
( Complejo II ) Succinato
deshidrogenasa
1. FAD2. Centros Fe-S
4
( Complejo III) Ubiquinona- Citocromo c
oxidorreductasa.
1. Hemo b3. Hemo 4. Centros Fe-S
11
( Complejo IV) Citocromo oxidasa
1. Hemo a2. Hemo
8
Complejos enzimáticos de la CR
a3
c1
, CuB3. CuA

Los complejos de proteínas guían a los e- a lo largo de la Cadena respiratoria , de manera que estos se mueven secuencialmente desde uno a otro complejo enzimático.

Ordenamientode la
Cadena Respiratoria

En las mitocondrias que respiran, cada molécula de NADH libera 2 electrones a la Cadena Respiratoria; estos electrones en última instancia, reducen a 1 átomo de oxígeno (media molécula de O2), formando una molécula de agua (H2O).
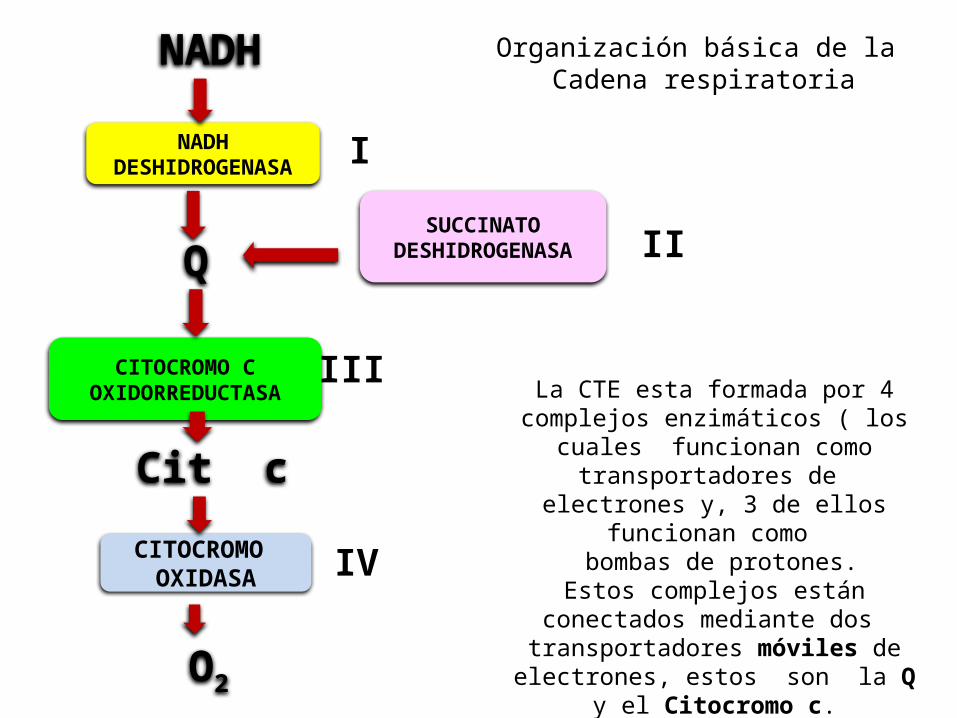
La CTE esta formada por 4 complejos enzimáticos ( los
cuales funcionan como transportadores de
electrones y, 3 de ellos funcionan como
bombas de protones. Estos complejos están conectados mediante dos transportadores móviles de
electrones, estos son la Q y el Citocromo c.
SUCCINATODESHIDROGENASA
O2
NADHDESHIDROGENASA
CITOCROMO C OXIDORREDUCTASA
CITOCROMO OXIDASA
NADH
Q
Cit c
I
II
III
IV
Organización básica de la Cadena respiratoria
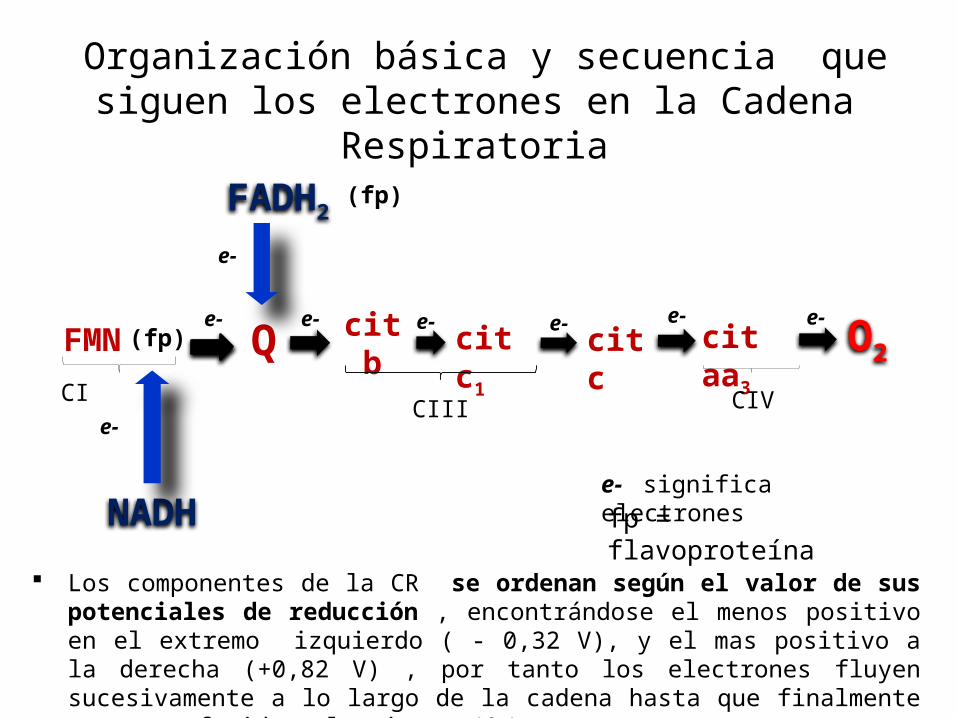
Organización básica y secuencia que siguen los electrones en la Cadena
Respiratoria
O2FMN cit b
NADH
Q cit c1
FADH2
cit c
cit aa3
e-
e-
e- e- e- e-e-e-
(fp)
e- significa electrones
(fp)
fp = flavoproteína
CI CIII CIV
Los componentes de la CR se ordenan según el valor de sus potenciales de reducción , encontrándose el menos positivo en el extremo izquierdo ( - 0,32 V), y el mas positivo a la derecha (+0,82 V) , por tanto los electrones fluyen sucesivamente a lo largo de la cadena hasta que finalmente son transferidos al oxigeno (O2).

Reacciones de oxidorreducción de los complejos enzimáticos
de la Cadena Respiratoria

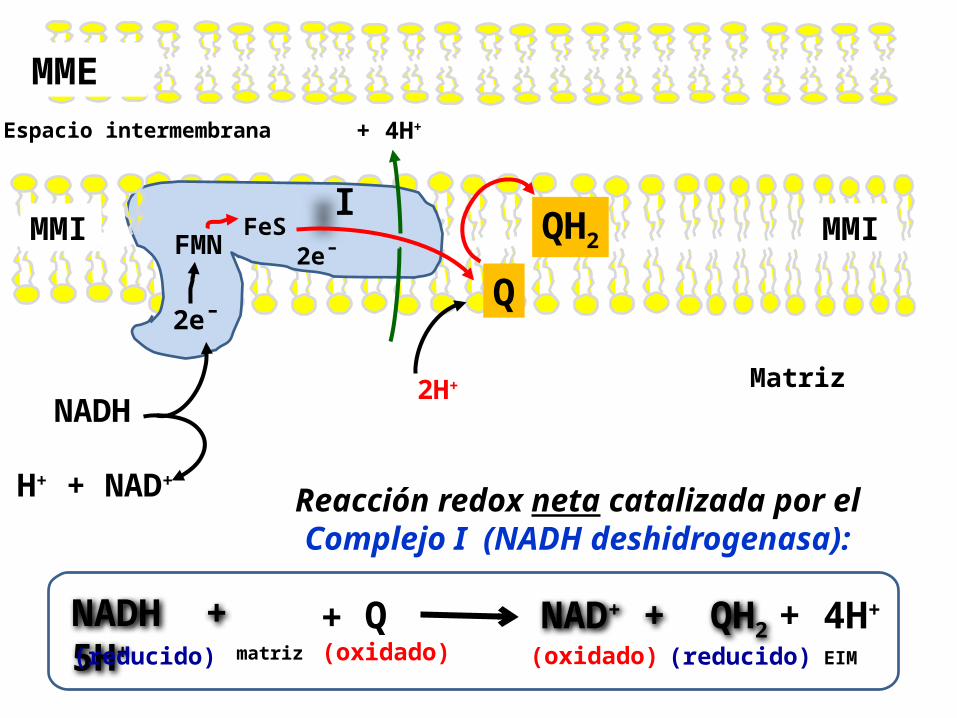
Reacción catalizada por el Complejo I, NADH deshidrogenasa:
Cataliza simultáneamente dos procesos acoplados:1. La transferencia exergónica de 2e- y 1H+
(un ion hidruro) del NADH y 1 H+ (un protón) de la matriz, hasta la Ubiquinona (Q).
los 2 electrones fluyen desde el NADH al FMN y luego a FeS que puede aceptar los electrones , pero uno cada vez, y de aquí hasta la Q (Ubiquinona).
2. Ocurre la transferencia endergónica de 4 protones de la matriz hacia el EIM. Por tanto , el complejo I es una bomba de protones impulsada por la energía de la transferencia de los 2 electrones.
MME
Reacción redox neta catalizada por el Complejo I (NADH deshidrogenasa):
NADH + 5H+
(reducido) (oxidado)
NAD+ + QH2 (reducido) (oxidado)
Q+ + 4H+matriz EIM
Espacio intermembrana
Q
Matriz
IFMN 2e¯
FeS QH2
2H+
+ 4H+
NADH
2e¯
H+ + NAD+
MMI MMI

1. La Succinato deshidrogenasa, que comprende el Complejo II de la CR, es la enzima que oxida una molécula de Succinato a Fumarato en el Ciclo del Ácido Cítrico, y se encuentra unida a la membrana interna mitocondrial.
2. Los 2 electrones liberados en la conversión de Succinato a Fumarato se transfieren primero al FAD, luego a FeS, y finalmente a la CoQ, formando CoQH2 .
3. Aunque el G’°, para esta reacción es negativa, la energía liberada NO es suficiente para el bombeo de protones. Por lo tanto NO hay translocación de los protones a través de la membrana por el Complejo II.
Reacción catalizada por el Complejo II, o Succinato deshidrogenasa:
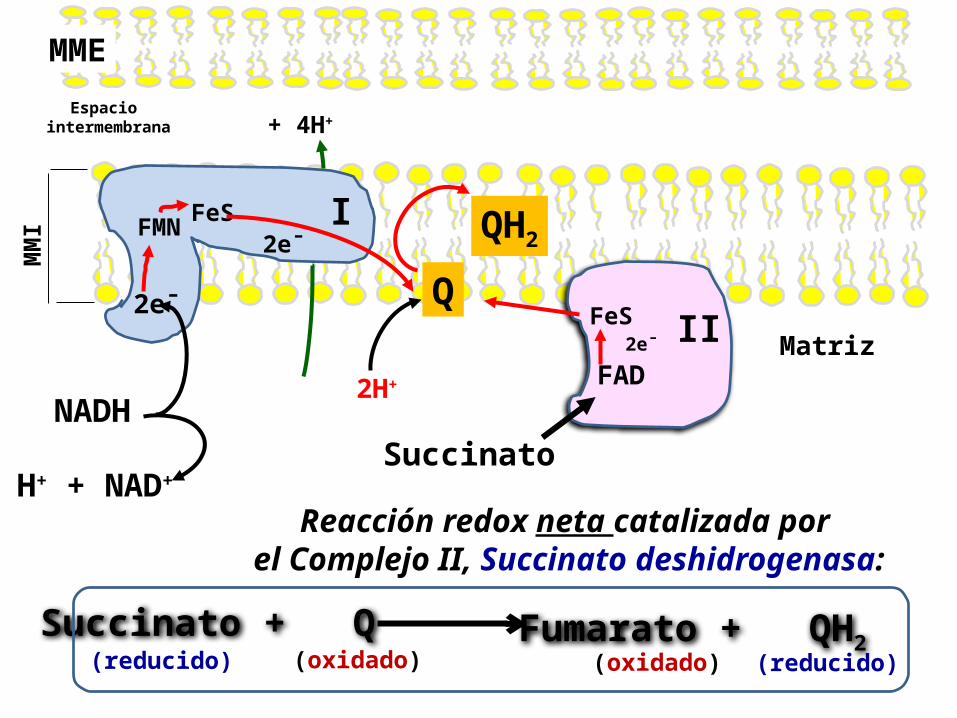
Reacción redox neta catalizada por el Complejo II, Succinato deshidrogenasa:
Succinato + Q (reducido) (oxidado)
Fumarato + QH2 (reducido) (oxidado)
Espacio intermembrana
MMEMM
I
2e¯Matriz
IFMN 2e¯FeS
2e¯ IIFAD
FeS
Succinato
QQH2
2H+
NADH
H+ + NAD+
+ 4H+
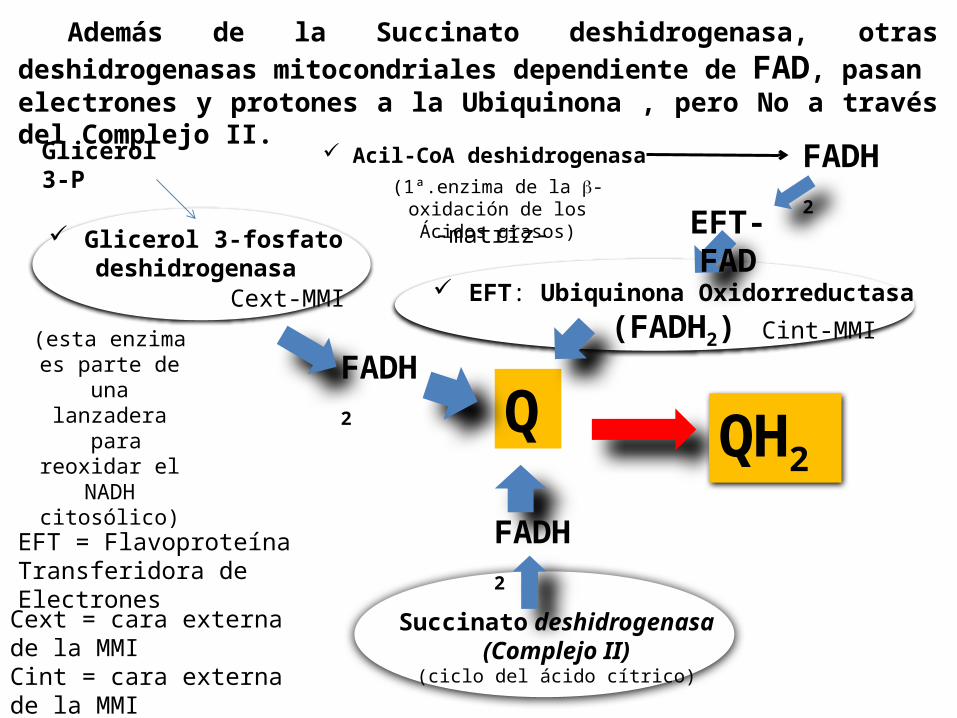
Q
Succinato deshidrogenasa (Complejo II)
(ciclo del ácido cítrico)
Glicerol 3-fosfatodeshidrogenasa
EFT: Ubiquinona Oxidorreductasa(FADH2)
QH2
EFT = Flavoproteína Transferidora de Electrones
FADH2
FADH2
Además de la Succinato deshidrogenasa, otras deshidrogenasas mitocondriales dependiente de FAD, pasan electrones y protones a la Ubiquinona , pero No a través del Complejo II. FADH
2(1ª.enzima de la -oxidación de los Ácidos grasos) EFT-
FAD
Acil-CoA deshidrogenasa
(esta enzima es parte de
una lanzadera
para reoxidar el
NADH citosólico)
Cint-MMICext-MMI
Cext = cara externa de la MMICint = cara externa de la MMI
-matriz-
Glicerol 3-P
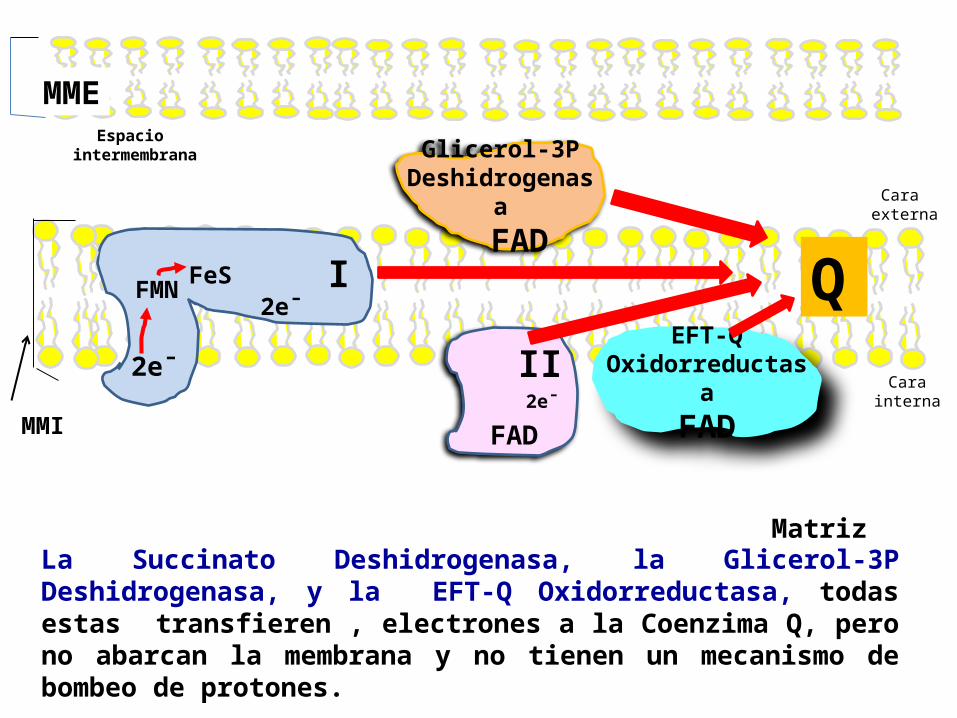
Espacio intermembrana
MME
MMI
2e¯
Matriz
IFMN 2e¯FeS
2e¯II
FAD
EFT-QOxidorreductas
aFAD
Glicerol-3PDeshidrogenas
a FAD
Q
La Succinato Deshidrogenasa, la Glicerol-3P Deshidrogenasa, y la EFT-Q Oxidorreductasa, todas estas transfieren , electrones a la Coenzima Q, pero no abarcan la membrana y no tienen un mecanismo de bombeo de protones.
Cara externa
Carainterna

1. QH2 dona 2 e- al Complejo III. Dentro de este complejo los electrones se transfieren a una proteína FeS , de aquí a 2 Citocromos tipo b, y luego son transferidos al Citocromo c1.
2. Por último, los 2 electrones se transfieren desde el Citocromo c1 a dos moléculas de la forma oxidada del Citocromo c, formando Citocromo c reducido .
3. La vía de los electrones por este tramo de la CR es complejo. Pero el efecto neto es simple: 1 QH2 se oxida a Q, y a la vez 2 Cit c se reducen.
El G’° para esta reacción es lo suficientemente negativo para que 4 protones sean translocados desde la matriz mitocondrial a través de la membrana interna para cada par de electrones transferidos.
Reacción catalizada por el Complejo III, Ubiquinona-Cit c-oxidorreductasa:
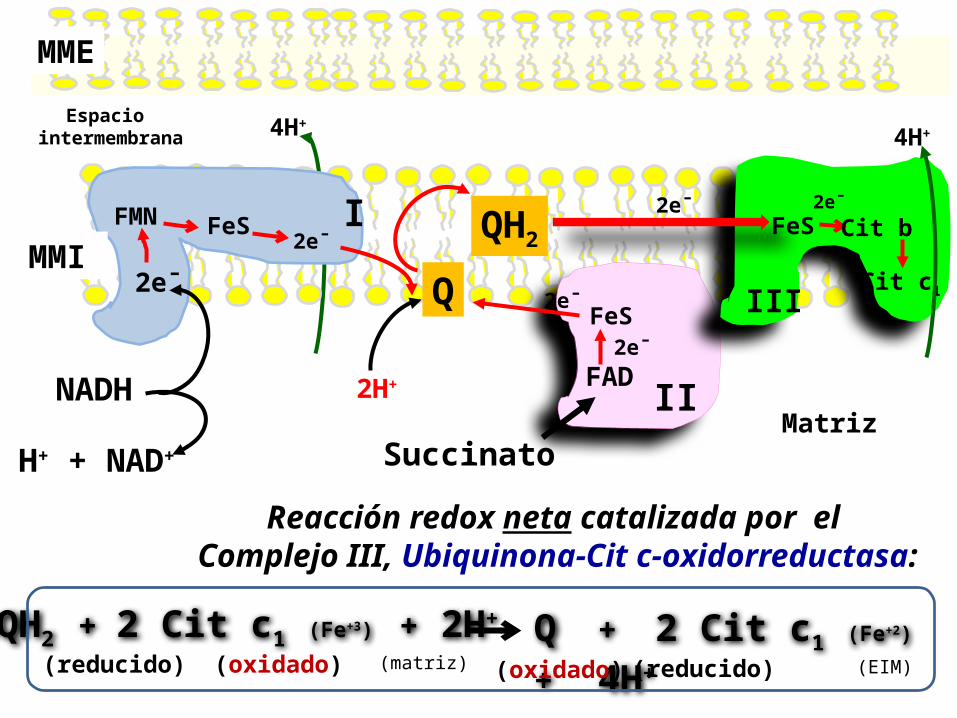
Espacio intermembrana
MME
MMI2e¯
Matriz
IFMN2e¯FeS
2e¯
IIFAD
FeS
Succinato
QQH2
2H+
III
2e¯FeS Cit b
Cit c1
Reacción redox neta catalizada por el Complejo III, Ubiquinona-Cit c-oxidorreductasa:
QH2 + 2 Cit c1 (Fe+3) + 2H+ (reducido) (oxidado)
Q + 2 Cit c1 (Fe+2)
+ 4H+ (reducido) (oxidado)(matriz) (EIM)
4H+
NADH
H+ + NAD+
4H+
2e¯
2e¯

1. Este Complejo IV transporta electrones desde el Citocromo c al Oxígeno molecular, reduciéndolo a H2O.
2. En este Complejo se trasporta 1 único electrón a la vez.
3. La transferencia de electrones a través del complejo IV , es como sigue:
primero los electrones pasan del Cit c a un par de iones de cobre (CuA), a continuación, al Citocromo a, luego a un centro Citocromo a3-CuB, y finalmente a O2, el último aceptor de electrones de la CR.
Por cada 2 electrones que pasan por el complejo, este utiliza 2 H+
como sustrato y produce1 molécula de agua (H2O).
4. También utiliza la energía de esta reacción redox para bombear 1H+ por cada electrón que pasa.5. La transferencia de cada electrón debe ocurrir sin la liberación de especies reactivas dañinas para la célula.
Reacción catalizada por el Complejo IV, Citocromo oxidasa:
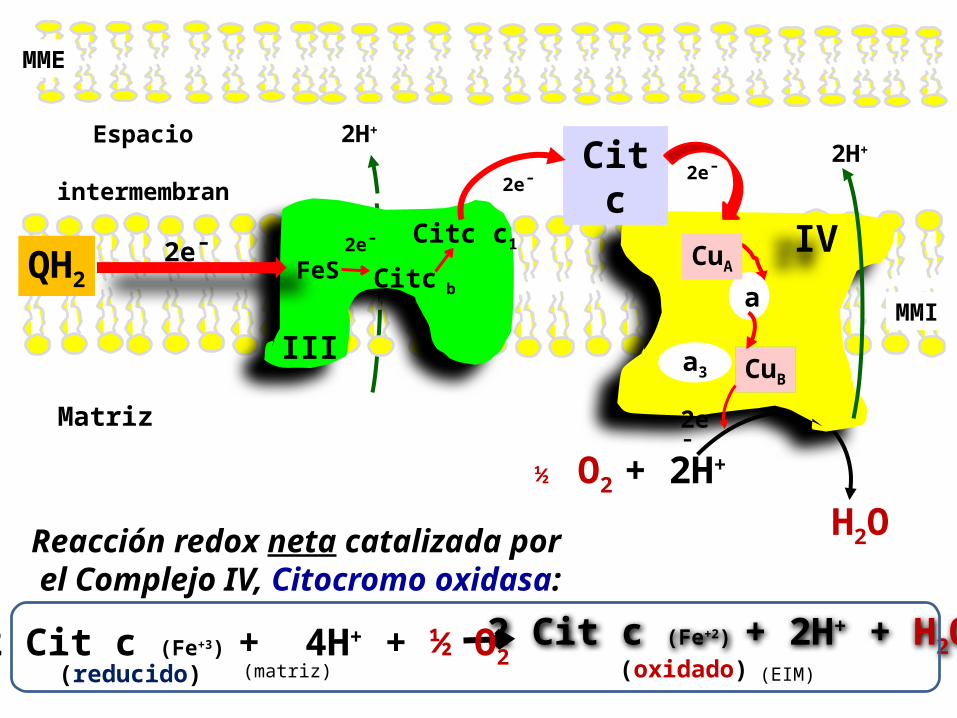
Espacio
intermembrana
Matriz
H2O + 2H+ ½ O2
IV
Cit c
III
2e¯FeS
Citc c1
a
a3
CuA
CuB
2e¯
QH2
MME
Citc b
(reducido)2 Cit c (Fe+2) + 2H+ + H2O
(oxidado)2 Cit c (Fe+3) + 4H+ + ½ O2
Reacción redox neta catalizada por el Complejo IV, Citocromo oxidasa:
(EIM)(matriz)
2H+
2e¯
2e¯2H+
MMI
2e¯

La reacción final en la cadena respiratoria, la reducción de O2 a H2O tiene un potencial de reducción estándar (E’o) de + 0,816 V, el más positivo en toda la cadena respiratoria, y por lo tanto esta reacción , también tiende a ocurrir hacia la derecha.½ O2 + 2H+ + 2e- H2O
Como se discutió anteriormente, el oxígeno molecular es un aceptor terminal de electrones ideal, debido a su alta afinidad por los electrones, proporciona una gran fuerza impulsora termodinámica.
E’o
+ 0,816 V
“ 2 protones son tomados de la matriz y 2 electrones que son cedidos por el Cit a3,
se unen a un átomo de oxigeno para reducirlo hasta agua”.
¿Cuánta energía se libera?
¿Se libera energía en la Cadena respiratoria?
¿Cómo se conserva de forma eficiente esta energía?
¿En que se utiliza esta energía liberada?
NADH NAD+ + H+
Complejo IE’o = 0,360 V
G´o = - 69,5 kJ/mol
Complejo IIIE’o = 0,190 V
G´o = - 36,7 kJ/mol
QSuccinato
fumarato
(+0,031V)FADH2 Complejo
II(+0,045 V)
Complejo IVE’o = 0,580 V
G´o = - 112 kJ/mol
(- 0,320 V)
½ O2 + 2H+ H2O2e-
(+ 0,816 V)
Citocromo c(+ 0,235 V)
- 0,4
- 0,2
0
+ 0,4
+ 0,6
+ 0,8
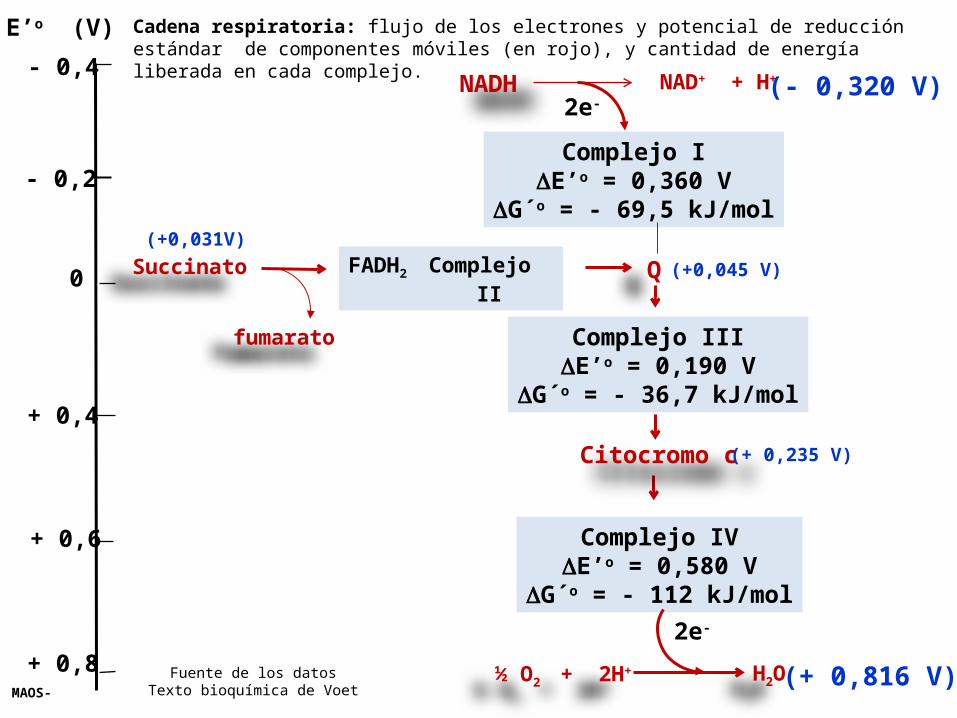
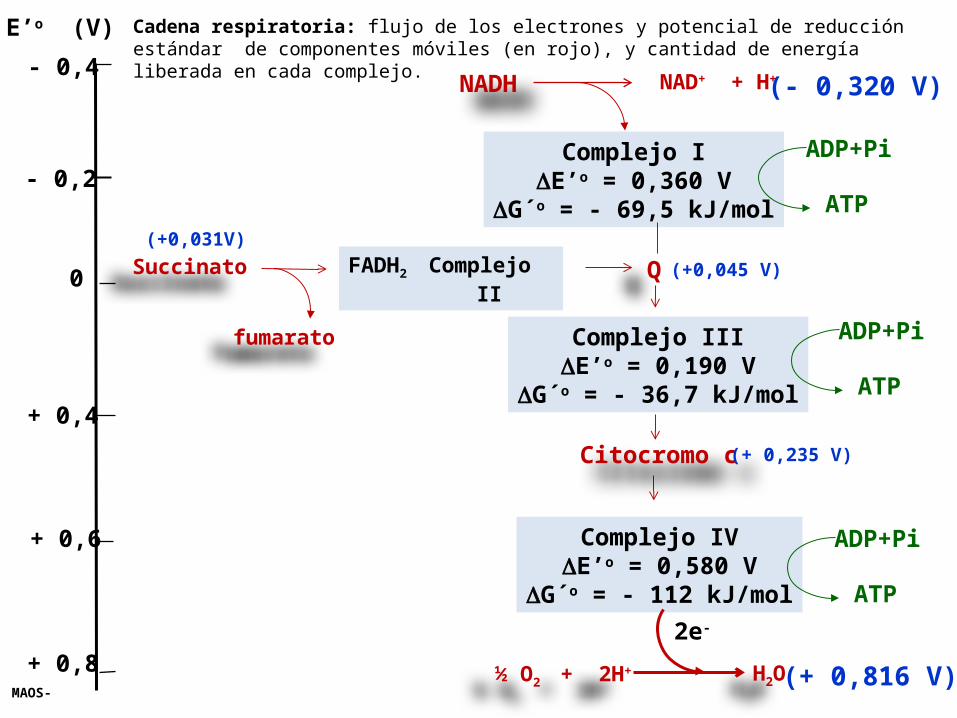
E’o (V) Cadena respiratoria: flujo de los electrones y potencial de reducción estándar de componentes móviles (en rojo), y cantidad de energía liberada en cada complejo.
MAOS-
2e-
Fuente de los datosTexto bioquímica de Voet

“ Esta energía producto de la transferencia de electrones se conserva eficientemente en un Gradiente de Protones”.
El flujo de electrones desde el NADH al O2 es un proceso exergónico que libera la cantidad de 220 kJ/mol (52 Kcal/mol)NAD+/ NADH……. E’o = - 0,320 VO2 / H2O………….. E’o = + 0,816 V
NADH + H+ + ½ O2 NAD+ + H2OREACCIÓN DE OXIDORREDUCCIÓN NETA de la Cadena Respiratoria:
G’o = - 220 kJ/mol (de NADH)
G’o = - n F E’o
G’o = - 2 (96,5 kJ / V. mol) (1,14 V)

Esta energía producto de la transferencia de electrones se conserva eficientemente en un Gradiente de protones.
El flujo de electrones desde el FADH2 (de la oxidación del succinato a fumarato) al O2 es un proceso exergónico que libera la cantidad de 150 kJ/mol
G’o = - 150 kJ/mol
G’o = - n F E’o
Fumarato / Succinato…. = + 0,031 VO2 / H2O………….. = + 0,816 V
G’o = - 2 (96,5 kJ/V. mol) ( 0,847 V)

Cuando los electrones pasan a través de 3 Complejos proteicos, que contienen afinidad cada vez mayor por lo electrones (potenciales de reducción estándar en aumento) en vez de pasar en forma directa al O2, esto permite que el gran cambio de energía libre total se divida en 3 paquetes mas pequeños, cada uno de los cuales se ACOPLA con la síntesis de ATP en un proceso llamado Fosforilación oxidativa.
Resumen:
NADH NAD+ + H+
Complejo IE’o = 0,360 V
G´o = - 69,5 kJ/mol
Complejo IIIE’o = 0,190 V
G´o = - 36,7 kJ/mol
QSuccinato
fumarato
(+0,031V)FADH2 Complejo
II(+0,045 V)
Complejo IVE’o = 0,580 V
G´o = - 112 kJ/mol
(- 0,320 V)
½ O2 + 2H+ H2O2e-
(+ 0,816 V)
Citocromo c(+ 0,235 V)
- 0,4
- 0,2
0
+ 0,4
+ 0,6
+ 0,8
E’o (V) Cadena respiratoria: flujo de los electrones y potencial de reducción estándar de componentes móviles (en rojo), y cantidad de energía liberada en cada complejo.
ADP+Pi
ATP
ADP+Pi
ATP
ADP+Pi
ATP
MAOS-

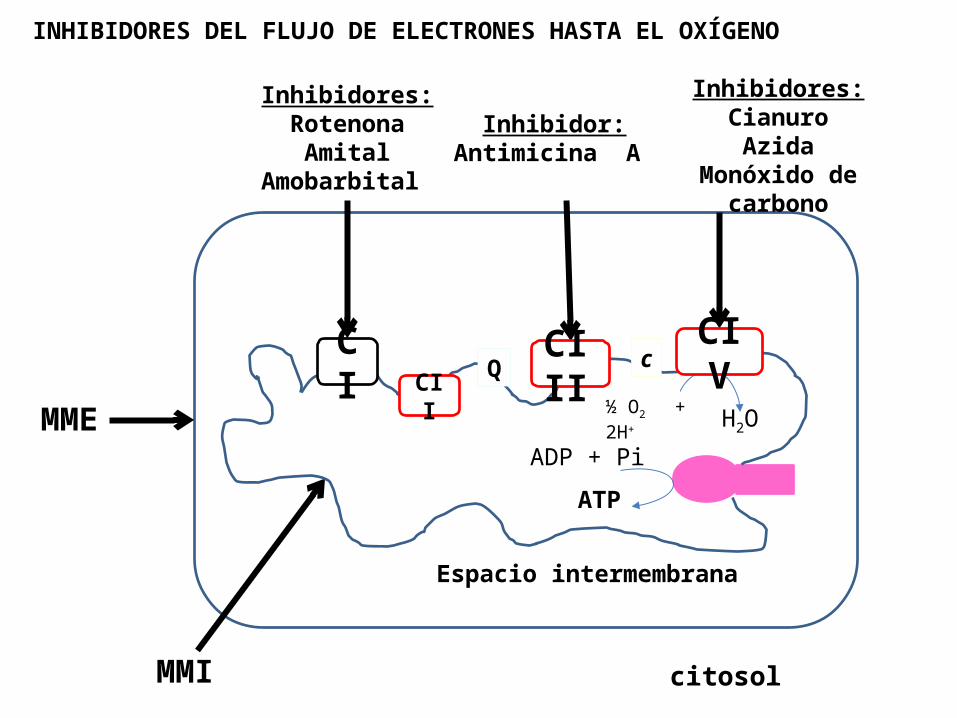
Antimicina A: un antibiótico de hongos, bloquea la Cadena respiratoria en el paso entre el citocromo b y el citocromo c1
I) Inhibidores del complejo I
II) Inhibidores del complejo III
Rotenona: veneno de peces , e insecticida
extraído de plantas Amital (barbitúrico) Amobarbital (barbitúrico) Piericidina A (antibiotico)
Inhiben la transferencia de electrones y, como consecuencia de ello, también interrumpen la
fosforilación oxidativa. Estas sustancias pueden producir rápidamente la
muerte
Inhibidores de la Cadena respiratoria

Inhibidores de la Cadena respiratoria
Cianuro (CN-)
N-3 : Azida
Monóxido de carbono (CO)
H2S
se unen al Fe+3 de la Citocromo oxidasa , por lo que evita que el oxigeno pueda unirse a la Citocromo oxidasa .
III) Inhibidores del complejo IV
ADP + PiATP
MMI
MME
Inhibidores:RotenonaAmital
Amobarbital
Inhibidor:Antimicina A
Inhibidores:CianuroAzida
Monóxido de carbono
Espacio intermembrana
citosol
INHIBIDORES DEL FLUJO DE ELECTRONES HASTA EL OXÍGENO
½ O2 + 2H+ H2O
CI CI
I
CIII
CIVQ c

Formación de especies reactivas de oxígeno
(radicales libres) durante la reacción del complejo enzimático Citocromo
oxidasa
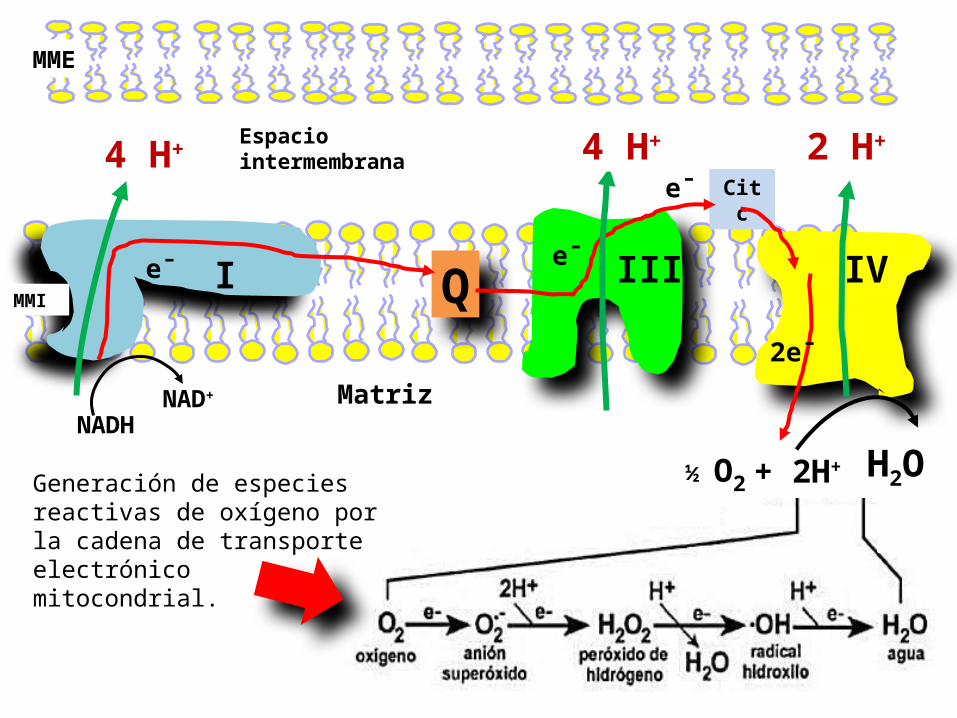
MME
I III IV
Cit c
Espacio intermembrana
QMMI
NADHNAD+
e¯ e¯
Matriz
e¯
2e¯
+ 2H+ ½ O2 H2O
4 H+ 4 H+ 2 H+
Generación de especies reactivas de oxígeno por la cadena de transporte electrónico mitocondrial.

Los derivados tóxicos del oxígeno molecular, tales como superóxido radicales son rescatados por enzimas protectoras
Aunque la citocromo c oxidasa y otras proteínas que reducen el O2 son un éxito notable en no liberar intermedios, se forman inevitablemente pequeñas cantidades de anión superóxido y peróxido de hidrógeno. Superóxido, peróxido de hidrógeno, y las especies que se pueden generar a partir de ellos, tales como OH · se denominan colectivamente como especies de oxígeno reactivas o ROS.
¿Cuáles son las estrategias de defensa celular contra el daño oxidativo por ROS? El principal de ellos es la enzima superóxido dismutasa. Esta enzima elimina los radicales superóxido por catalizar la conversión de dos de estos radicales en peróxido de hidrógeno y oxígeno molecular.

El oxígeno molecular (O2) es un birradical que tras la adición secuencial de 4 electrones y 4 protones en la cadena de transporte electrónico mitocondrial, genera agua (H2O). En este proceso, en el que se produce gran cantidad de energía (ATP), pueden también generarse, como subproductos, moléculas parcialmente reducidas con 1, 2 y 3 electrones. Estas son, el radical superóxido, el peróxido de hidrógeno y el radical hidroxilo, respectivamente.Estos tres , se denominan especies reactivas de oxígeno y causan lesiones oxidativas a las macromoléculas celulares (lípidos, proteínas y DNA).
El principal productor de radicales de oxígeno en los organismos aerobios es la mitocondriautiliza aproximadamente el 98% del O2 que ingresa en la célula reduciéndolo a agua a nivel del citocromo a3 de la cadena de transporte de electronesuna pequeña parte del O2 incorporado por la mitocondria, sufre una reducción monovalente y genera especies reactivas de oxígeno, tales como el radical superóxido (O2 ), el peróxido de hidrógeno (H2O2), y radical hidroxilo (•OH)Por tanto, al ser las mitocondrias la fuente más importante de especies reactivas de oxígeno, es obvio que el DNA mitocondrial se encuentra más expuesto que el DNA nuclear a sufrir lesiones oxidativas severas que pueden conducir a mutaciones y supresiones en su propia cadena
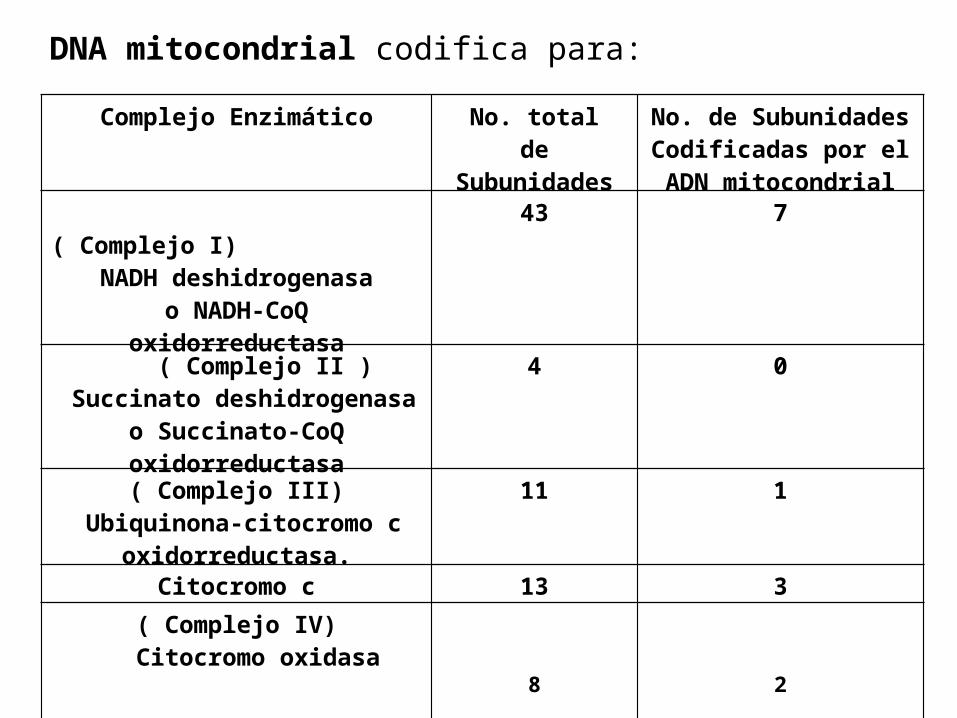
DNA mitocondrial codifica para:
Complejo Enzimático No. totalde
Subunidades
No. de SubunidadesCodificadas por el ADN mitocondrial
( Complejo I)
NADH deshidrogenasao NADH-CoQ
oxidorreductasa
43 7
( Complejo II ) Succinato deshidrogenasa
o Succinato-CoQ oxidorreductasa
4 0
( Complejo III) Ubiquinona-citocromo c
oxidorreductasa.
11 1
Citocromo c 13 3( Complejo IV)
Citocromo oxidasa8 2

Nota: las laminas desde el 54 al 59 no serán evaluados en el examen parcial de esta unidad 3

Fosforilación
OxidativaEs el proceso por el cual el NADH y FADH2 producidos por la oxidación de nutrientes SE OXIDAN y simultáneamente ocurre la SÍNTESIS de ATP .

Ecuación básica de la Fosforilación oxidativa
ADP + Pi ATP + H2OATP sintasa
G’o = + 30,5 kJ/mol
La energía libre requerida para sintetizar 1mol de ATP a partir de ADP + Pi es de 30,5kJ (*), en condiciones bioquímicas estándares. Si la energía libre estándar producto de la oxidación del NADH al O2, esta acoplada a la síntesis de ATP, es por lo tanto suficiente para conducir la formación de varios moles de ATP. (*) en condiciones intracelulares reales es de 50kJ/mol)

Mecanismo que explica el Acoplamiento energético del
Transporte de electrones a la Síntesis de ATP
quimiosmótico: este término describe las reacciones enzimáticas en las que interviene simultáneamente una reacción química y un proceso de transporte.
Teoría quimiosmótica del Dr. Peter Mitchell (1961) Premio Nobel de
química en 1978

1. La energía libre de la transferencia de electrones, se utiliza para un bombeo de Protones ( H+ ) desde la matriz de la mitocondria hacia el EIM.
2. Este movimiento de protones genera: un Gradiente de concentración de protones (es decir, una diferencia de pH) a través de la MMI.
Un Gradiente de carga electrica (es decir, una diferencia de cargas) a través de la MMI. Siendo positivo (+) el lado externo de la MMI y negativo (-) el lado interno.
Mecanismo que explica el Acoplamiento energético del transporte de electrones a
la Síntesis de ATP:
3. Juntos estos dos gradientes (de concentración de H+ y de cargas) ALMACENAN la energía libre resultante de la transferencia de electrones, y es llamada fuerza protón motriz.

4. El regreso de los protones hacia la matriz a través de Complejo de la ATP sintasa , esta ACOPLADO a la Síntesis de ATP. Proporcionando este flujo, la energía necesaria para unir un Pi al ADP .
5. La membrana interna mitocondrial es impermeable a los Iones H+ . De otro modo se disiparía el gradiente de concentración de protones.¿ Como se acopla la energía libre de la
transferencia de protones a la síntesis de ATP?
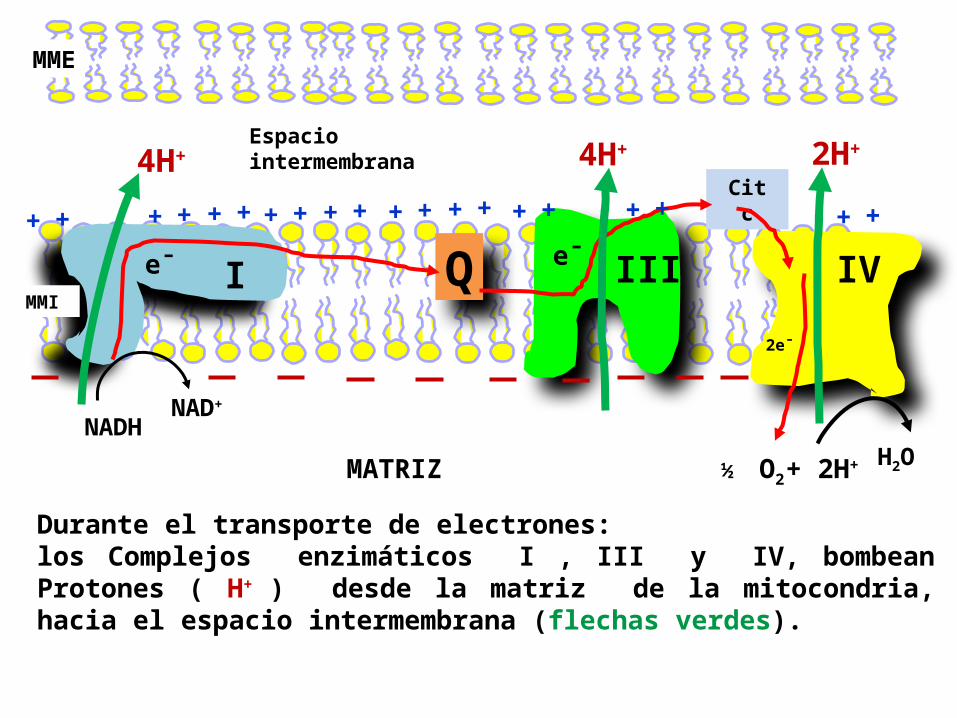
MME
I III IV
Cit c
Espacio intermembrana
QMMI
NADHNAD+
e¯ e¯
MATRIZ
4H+4H+ 2H+
2e¯
H2O + 2H+ ½ O2
+ + + + + ++ ++ + + ++ + + ++ + + +
Durante el transporte de electrones: los Complejos enzimáticos I , III y IV, bombean Protones ( H+ ) desde la matriz de la mitocondria, hacia el espacio intermembrana (flechas verdes).

La moléculas NADH y FADH2 transfieren electrones a la CR, y se libera energía durante el proceso , parte de ésta es utilizada para bombear protones desde la matriz al EIM.
El bombeo de H+ es un proceso endergónico.
Este bombeo de H+ tiene dos consecuencias:1.Se genera un Gradiente de concentración de protones (ΔpH) a través de la MMI, con el pH más alto (mas alcalino) en la matriz, que en el EIM.
2. Se genera un Gradiente de carga (Δ) (= diferencia de potencial) a través de la MMI, con el interior negativo y el exterior positivo (como resultado de la salida neta de iones positivos).
.
En conjunto la pH y la constituyen un
gradiente electroquímico , el cual almacena la energía del transporte de electrones.
Letra del alfabeto griego (Psi)
3. A esta energía almacenada en este gradiente electroquímico se le denomina FUERZA PROTON MOTRIZ.
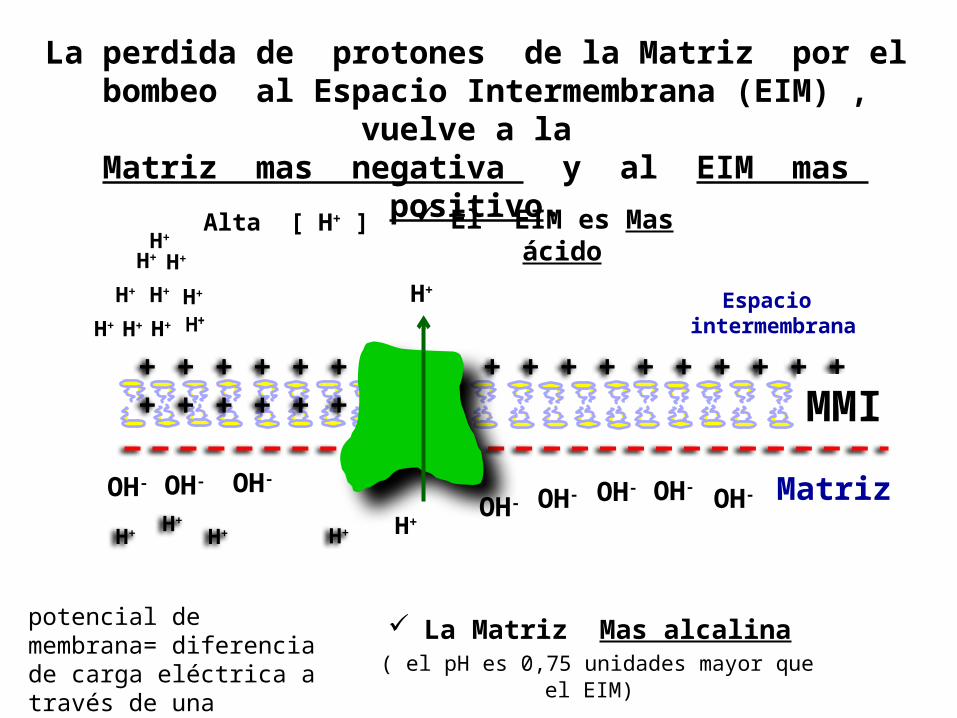
La perdida de protones de la Matriz por el bombeo al Espacio Intermembrana (EIM) ,
vuelve a la Matriz mas negativa y al EIM mas
positivo. El EIM es Mas ácido
La Matriz Mas alcalina ( el pH es 0,75 unidades mayor que
el EIM)
+ + + + + + + + + + + + + + + + + + + + + + + + + + +
Alta [ H+ ]
H+
H+ H+ H+ OH-OH-OH- OH-
OH-
MMI
Espacio intermembrana
Matriz OH- OH- OH-
H+ H+ H+ H+ H+
H+ H+ H+
H+
H+ H+
potencial de membrana= diferencia de carga eléctrica a través de una membrana

Para que se pueda mantener este gradiente electroquímico a través de la membrana interna , se deben dar las siguientes condiciones.
1.Que la membrana mitocondrial interna se mantenga intacta y no se haya hecho permeable con agentes físicos o químicos.
2.Que la transferencia de electrones sea continua, es decir que sustratos como el NADH, el FADH2 y el oxigeno se hallen disponibles.

Es una ATPasa de tipo F . El tipo “F” proviene de la identificación de esta ATPasa como un Factor de acoplamiento de energía.Es un complejo enzimático de la membrana interna mitocondrial que cataliza la síntesis de ATP a partir de ADP y Pi, acompañado por el flujo de protones desde el EIM a la matriz.
Esta compuesta por 2 componentes funcionales Fo y F1.
Fo : Es una proteína integral ubicada en la MMI, que forma un poro (canal) transmembrana para protones. La letra del subíndice “o” indica su inhibición por la OLIGOMICINA .
F1 : Es una proteína periférica de membrana, que mira hacia el interior de la matriz, contiene el sitio catalítico para la síntesis de ATP . El subíndice 1 indica que fue el primer factor identificado como necesario para la fosforilación oxidativa.
ATP sintasa
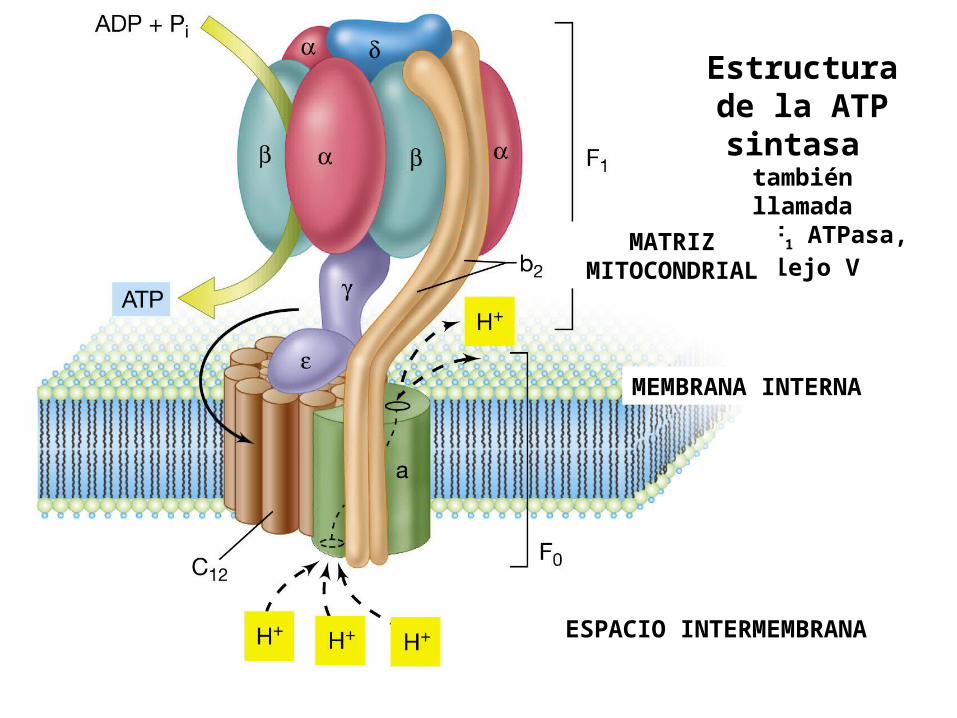
Estructura de la ATP sintasa también llamada
Fo - F1 ATPasa,o Complejo V
ESPACIO INTERMEMBRANA
MATRIZ MITOCONDRIAL
MEMBRANA INTERNA
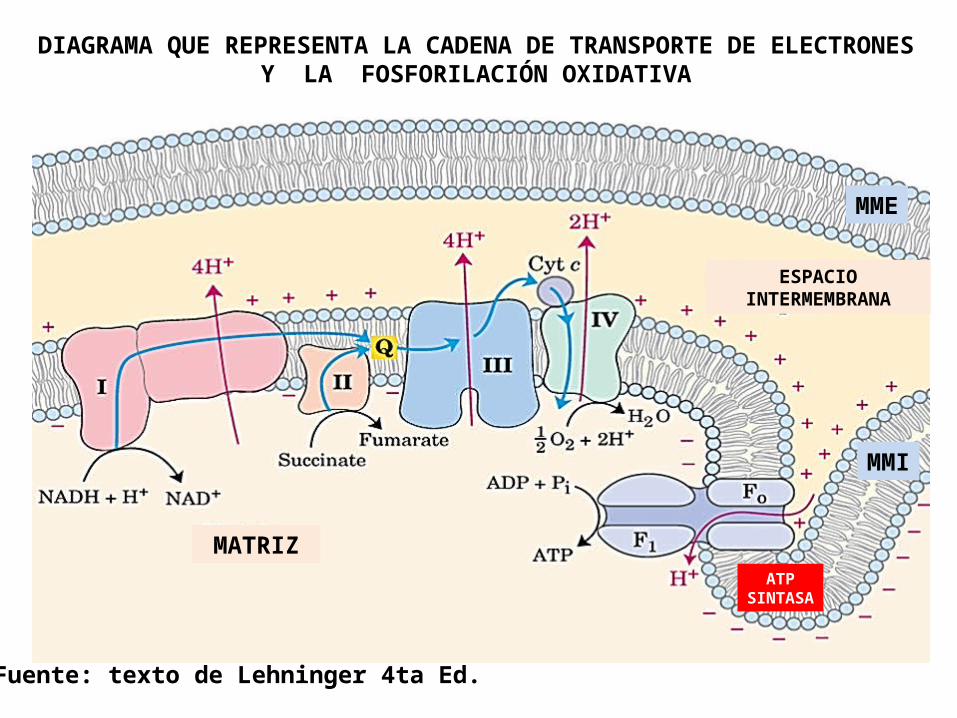
ATPSINTASA
DIAGRAMA QUE REPRESENTA LA CADENA DE TRANSPORTE DE ELECTRONES Y LA FOSFORILACIÓN OXIDATIVA
MATRIZ
ESPACIO INTERMEMBRANA
MMI
MME
Fuente: texto de Lehninger 4ta Ed.

1. Adenina- Nucleótido Translocasa. Es una proteína transportadora. El ADP es cotransportado con el ATP en direcciones opuestas por esta proteína . (El ADP hacia dentro y el ATP hacia fuera de la matriz). Este translocador es inhibido por un glucósido tóxico llamado Atractilósido (de la planta Atractylis gummifera) y el Ácido bongkrékico (antibiótico producido por un hongo que descompone la leche de coco).
2. Transportador de Fosfato
Sistemas de transporte a través de la Membrana mitocondrial interna
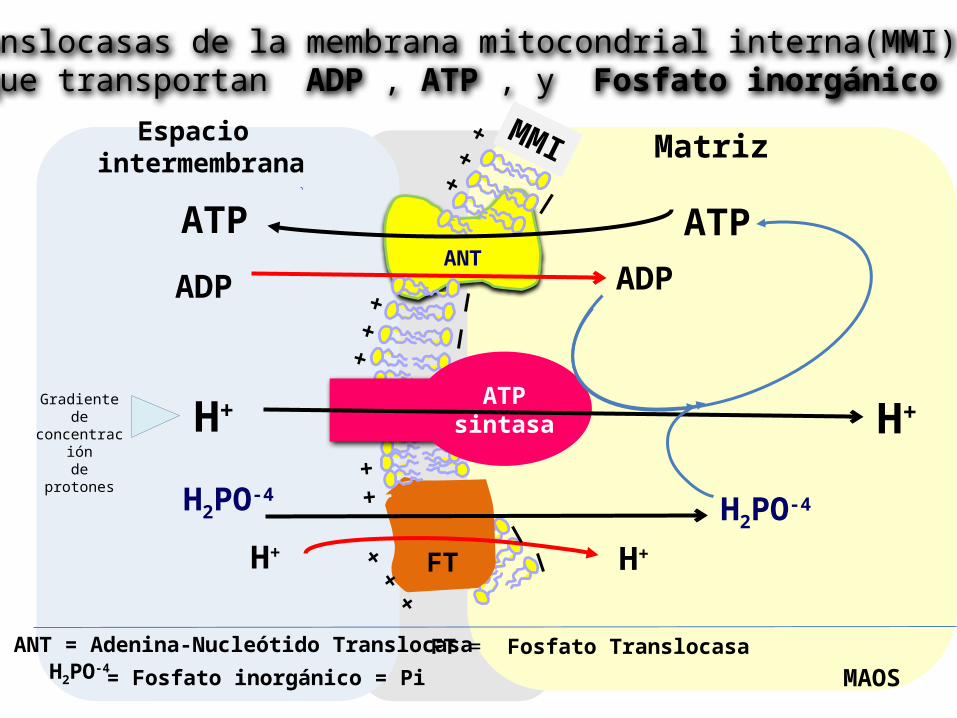
H2PO-4= Fosfato inorgánico = Pi
Translocasas de la membrana mitocondrial interna(MMI) que transportan ADP , ATP , y Fosfato inorgánico
ANT = Adenina-Nucleótido TranslocasaFT = Fosfato Translocasa
ANT
H+
ATPADP
H2PO-4 H2PO-4
H+
ADPATP
ATPsintasaH+ H+
FT
MMIEspacio intermembrana Matriz
Gradiente de
concentraciónde
protones
+++
++
+++
+++
MAOS

Importancia del transporte mitocondrial
En los tejidos aeróbicos casi todo el ATP se sintetiza dentro de la mitocondria. Sin embargo este se utiliza fuera de ella.

Relación P/O
Es la proporción de ATP sintetizado por átomo de Oxigeno reducido a H2O .
Esta relaciones P/O están sujetas a revisión

El valor consenso de los protones que se bombean hacia afuera por cada 2 e- es: de 10 para el NADH y de 6 para el Succinato.
El número de protones necesarios para impulsar la síntesis de una molécula de ATP es de 4 (de estos 1 se gasta en transportar Pi y ADP a través de la MMI).
La razón P/O cuando un NADH es el dador de electrones, es de 2,5 (10/4) y 1,5 (6/4) cuando lo es el Succinato.
Los valores 2,5 y 1,5, y los valores 3 y 2 para la razón P/O, se utilizan en los textos de bioquímica.
¿Cuántos protones han de entrar a través del Complejo Fo F1 (ATP sintasa) para
impulsar la síntesis de ATP?
Fuente: texto de Lehninger 4ta Ed.

La oxidación de un NADH se acopla a la síntesis de ATP, y genera la síntesis aprox. de 3 ATP. En consecuencia la eficiencia termodinámica de la fosforilación oxidativa es 3 x 30,5 kJ/mol x 100/220 kJ/mol = 42% bajo condiciones bioquímicas estándares.
Sin embargo bajo condiciones fisiológicas en las mitocondrias activa (donde las concentraciones de los reactivos y los productos así como el pH se alejan de las condiciones estándares), se cree que esta eficiencia termodinámica es de 70%. En comparación la eficiencia de un motor de automóvil típico es de < 30 %.
Texto de BioquímicaAutor. Voet-Voet 3ra. Edición. cap 22.pag.835

Si solo se utiliza aproximadamente el 42 % de la energía de la oxidación del NADH o FADH2 para La síntesis de ATP, ¿Qué sucede
con el resto de la energía?Parte de esta energía se pierde en forma
de calor.Por consiguiente la cadena respiratoria es
nuestra principal fuente de calor

Nota:Si la eficiencia en el proceso de síntesis de ATP
acoplada con la oxidación de NADH , fuera de 100% , seria en teoría posible sintetizar 7,51 moles de ATP
por cada mol de NADH oxidado a NAD+ , bajo condiciones estándares.

LA ENERGÍA SE GASTA EN :
1) Bombeo de protones desde la matriz a EIM (por c/ 2 electrones transferidos al O2 se bombean 4 protones por el CI; 4 por el CII, y 2 por el CIV).
2) La Síntesis de ATP.
3) El transporte de Piruvato, ADP y Pi desde el EIM hacia la matriz
4) El transporte de ATP desde la matriz hacia el EIM.

Regulación de la fosforilación oxidativaEl control de la velocidad de la Fosforilación oxidativa esta ligada a la demanda de energía celular.El consumo de O2 (tasa respiratoria) esta controlado por la concentración de ADP, la cual es una medida del estado energético de la célula.
La velocidad de la respiración celular aumenta si la concentración de ADP es alto, como sucede cuando los requerimientos de energía por la celular son altos, hay un aumento de la velocidad en la degradación ATP a ADP +Pi, por tanto hay mayor disponibilidad de ADP para la Fosforilación oxidativa y aumenta la velocidad de la Cadena respiratoria, y la fosforilación de ADP, lo que da lugar a la regeneración de ATP.El ATP de forma a la misma velocidad en que es requerido por la actividades celulares, ,
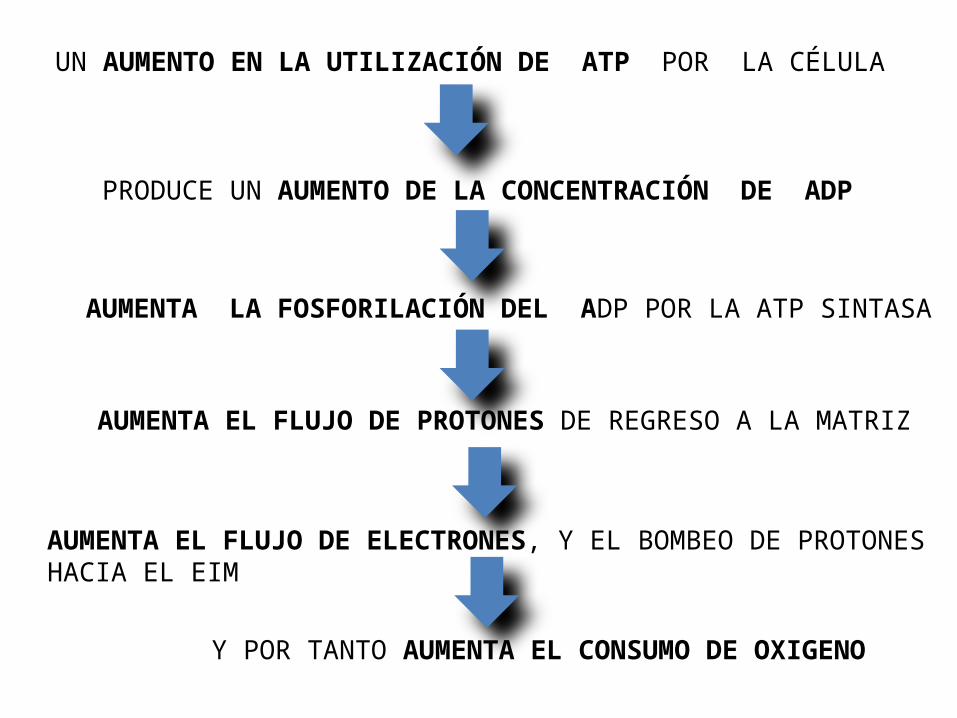
UN AUMENTO EN LA UTILIZACIÓN DE ATP POR LA CÉLULA
PRODUCE UN AUMENTO DE LA CONCENTRACIÓN DE ADP
AUMENTA LA FOSFORILACIÓN DEL ADP POR LA ATP SINTASA
AUMENTA EL FLUJO DE PROTONES DE REGRESO A LA MATRIZ
AUMENTA EL FLUJO DE ELECTRONES, Y EL BOMBEO DE PROTONES HACIA EL EIM
Y POR TANTO AUMENTA EL CONSUMO DE OXIGENO

La oligomicina un antibiótico producido por un hongo se une a la proteína F0 de la ATP sintasa, impidiendo el normal movimiento de protones hacia la matriz.Bloque completamente la oxidación y la fosforilación en las mitocondrias Intactas.
INHIBIDORES DE LA FOSFORILACIÓN OXIDATIVA

1.Atractilósido : glucósido vegetal
2. Ácido bongkrekico: antibiótico producido por un hongo.
INHIBIDORES DEL TRANSPORTE DE LOS NUCLEÓTIDOS DE ADENINA EN LA MEMBRANA MITOCONDRIAL INTERNA

Desacoplamiento entre la Cadena respiratoria y la Fosforilación
Oxidativa por químicos desacoplantes
( desacoplamiento de tipo no fisiológico)
Los desacopladores son compuestos que aumentan la permeabilidad de la membrana a
los protones.

DESACOPLAMIENTO DE LA FOSFORILACION OXIDATIVA
Mecanismo de acción:Los desacopladores tienen como efecto desconectar la fosforilación oxidativa de la transferencia de electrones.Por tanto puede funcionar la cadena respiratoria sin tener lugar la síntesis de ATP , en esta situación la energía que normalmente se hubiera conservado en la formación del ATP, se pierde ahora en forma de calor.ejemplo de dos desacopladores químicos:
2,4 - dinitrofenol (DNP) Carbonil-cianuro-p-trifluorometoxifenilhidrazona (FCCP)
DicumarolAmbos son ionóforos, ácidos liposolubles débiles que se unen a los H+ en el EIM, difunden a través de la membrana, y los liberan en la matriz.
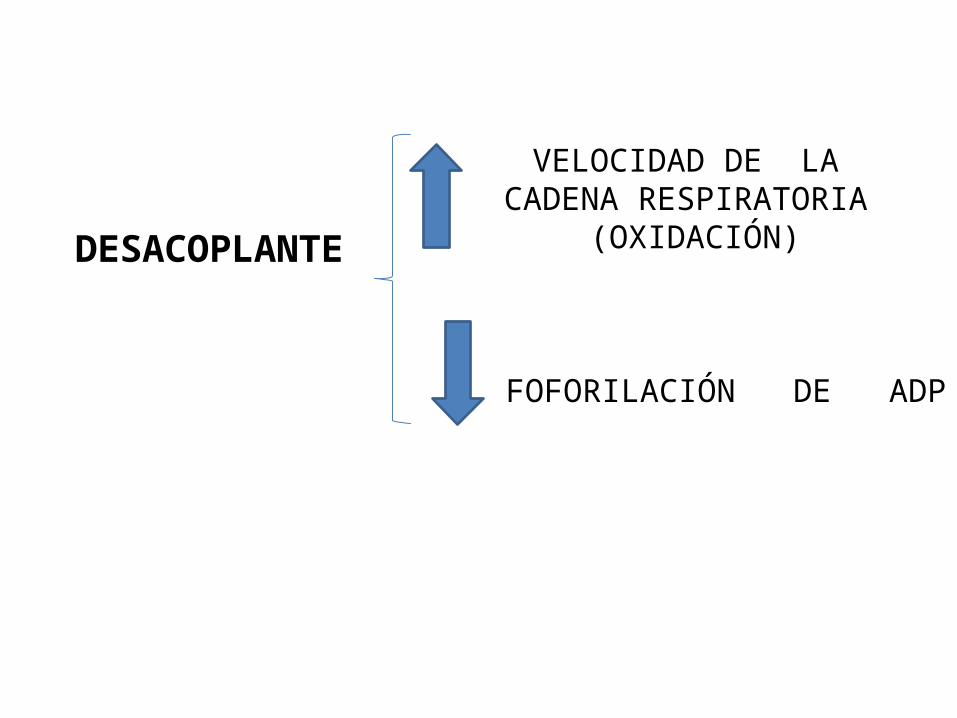
DESACOPLANTE
VELOCIDAD DE LACADENA RESPIRATORIA
(OXIDACIÓN)
FOFORILACIÓN DE ADP

Efectos fisiológicos de los desacopladores :
1. Aumenta la Tasa metabólica,
2. Aumenta el Consumo de oxigeno
3. Aumenta la Producción de calor
(fiebre).
4. Causa Perdida de Peso
“Estos dan por resultado una respiración no controlada, La concentración de ADP ya no limita la velocidad de la respiración”

Desacoplamiento fisiológico entre la Cadena respiratoria y la
Fosforilación Oxidativa
Termogénesis adaptativa
Condiciones en las que ocurre:En respuesta al frioEn periodos de HibernaciónIngestión de alimentos
Lugar : exclusivamente en tejido adiposo marrón

1. Animales Recién nacidos de algunas especies, que nacen sin pelaje (incluyendo al humano)
2. Animales hibernantes ( distribución de la grasa marrón: en su cuello y parte superior de la espalda)
3. Animales de hábitat frio4. Seres humanos adultos, en poca cantidad
(distribución de la grasa marrón: parte dorsal del tórax, parte inferior de los omóplatos, alrededor de las adrenales, riñones, corazón, región perianal e inguinal).
Generación de calor (termogénesis) en el Tejido adiposo marrón. Las Mitocondrias desacopladas del tejido adiposo marrón producen calor, bajo control hormonal,Para mantener la temperatura corporal en los :
calor calor

“ La función fisiológica del TEJIDO ADIPOSO MARRÓN (grasa parda), es generar calor ” .
es un tejido con una gran cantidad de triacilglicéridos, es rico en mitocondrias cuyos citocromos le confieren el color marrón. Contiene proteínas de desacoplamiento (UCP).
La TERMOGENINA también llamada Proteína desacopladora 1 (UCP-1) esta ubicada en la MMI del adipocito marrón , forma una canal por donde los protones son conducidos de regreso a la matriz , sin pasar a través de la ATP sintasa, desacoplando la CTE de la Fosforilación Oxidativa, esto permite la oxidación continua de combustible, sin síntesis de ATP , y disipando la energía en forma de CALOR. UCP = Uncoupling protein
M
M
LV= gota de grasaM= mitocondriaN= núcleo Adipocito
de grasa pardaAdipocito
de grasa blanca
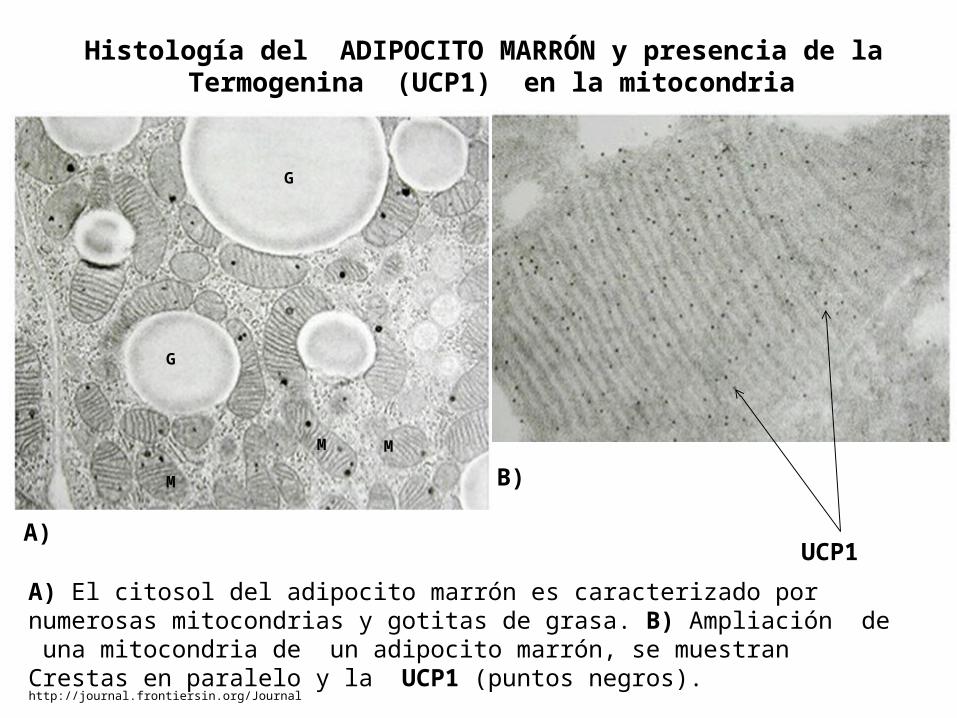
http://journal.frontiersin.org/Journal
Histología del ADIPOCITO MARRÓN y presencia de la Termogenina (UCP1) en la mitocondria
A) El citosol del adipocito marrón es caracterizado por numerosas mitocondrias y gotitas de grasa. B) Ampliación de una mitocondria de un adipocito marrón, se muestran Crestas en paralelo y la UCP1 (puntos negros).
A)
B) M M
M
G
G
UCP1
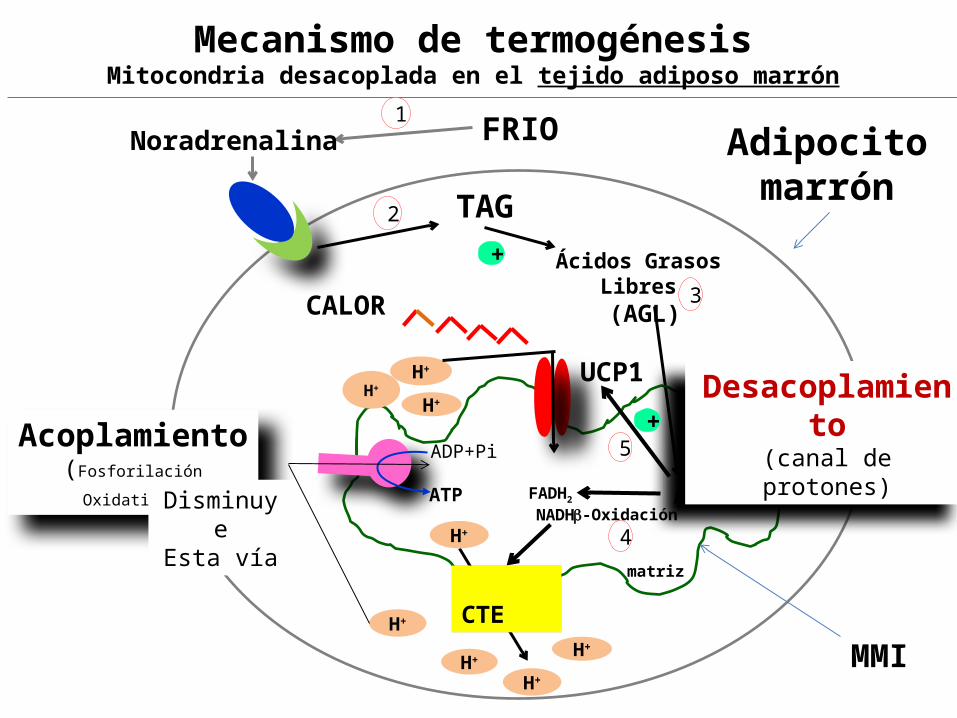
FRIONoradrenalina Adipocito marrón TAG
Ácidos Grasos Libres (AGL)
-OxidaciónFADH2 NADH
UCP1
AGL
matriz
H+
H+
H+
H+
ADP+Pi
ATP
H+
H+
CTE
Desacoplamiento
(canal de protones)
+
+
Mecanismo de termogénesisMitocondria desacoplada en el tejido adiposo marrón
CALOR
Acoplamiento(Fosforilación
Oxidativa)
1
2
3
4
5
Disminuye
Esta vía
H+H+
MMI
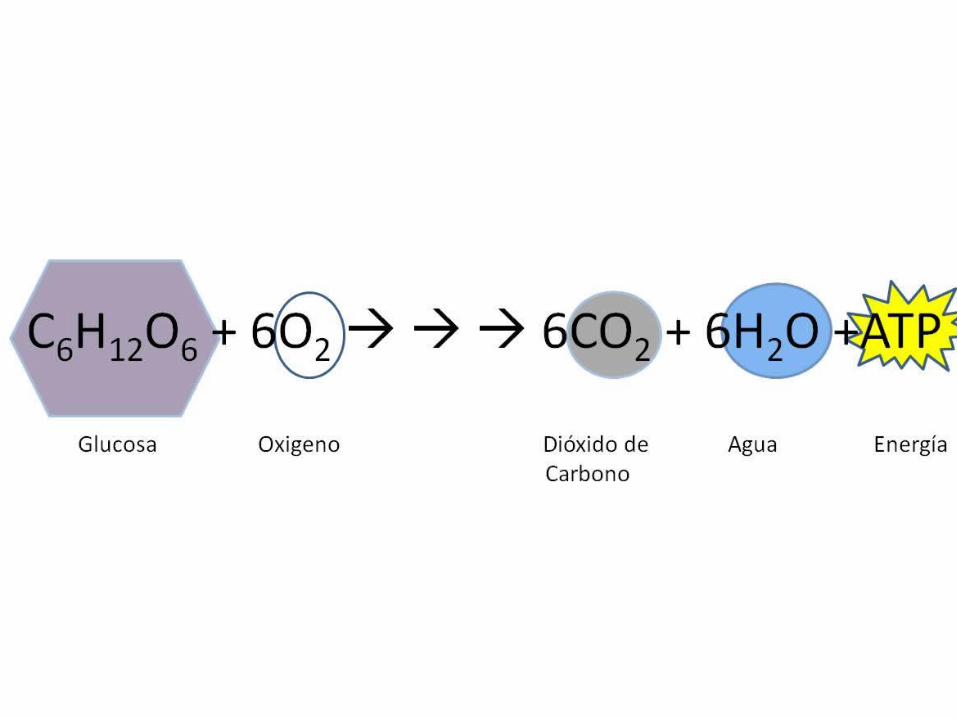
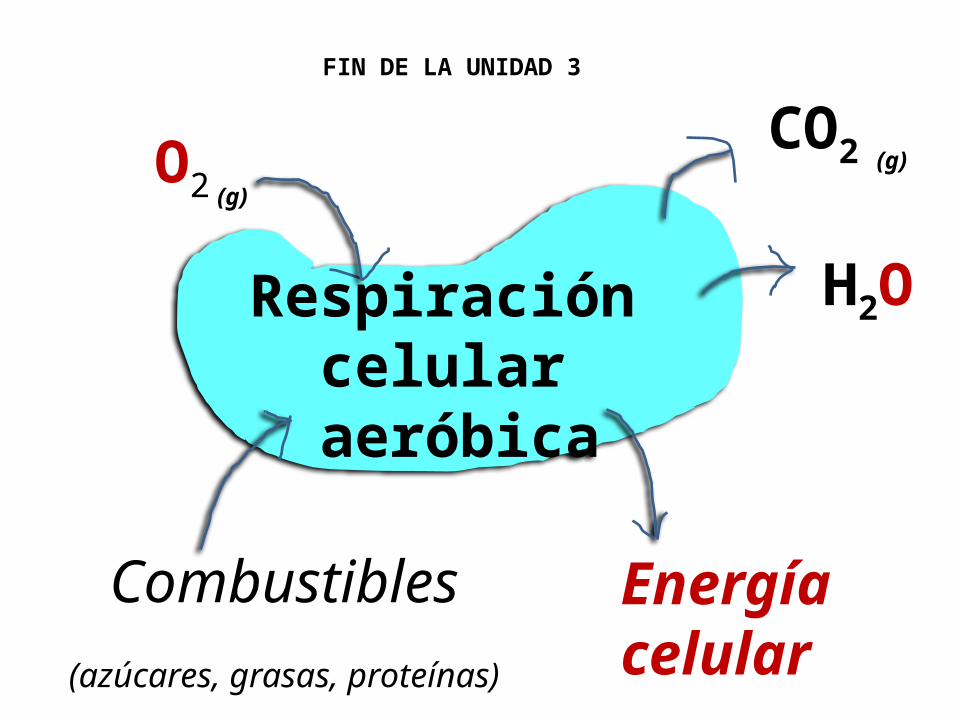
Respiración celular aeróbica
O2CO2
H2O
Energíacelular
Combustibles(azúcares, grasas, proteínas)
(g)(g)
FIN DE LA UNIDAD 3