antigen dose escalation study of a vegf-based therapeutic cancer vaccine in non human primates
TRANSCRIPT

G
J
Ai
YJa
b
a
ARR1AA
KVCVV
1
oaatber
vas
0d
ARTICLE IN PRESS Model
VAC-12524; No. of Pages 10
Vaccine xxx (2011) xxx– xxx
Contents lists available at SciVerse ScienceDirect
Vaccine
jou rn al h om epa ge: www.elsev ier .com/ locate /vacc ine
ntigen dose escalation study of a VEGF-based therapeutic cancer vaccinen non human primates
anelys Moreraa,∗, Mónica Bequet-Romeroa, Marta Ayalaa, Pedro Puente Pérezb, Jorge Castrob,avier Sáncheza, José Suárez Albab, Julio Ancízarb, Karelia Cosmeb, Jorge V. Gavilondoa
Cancer Immunotherapy Laboratory, Department of Pharmaceuticals, Center for Genetic Engineering and Biotechnology, P.O. Box 6162, Playa Cubanacábn, Havana 10600, CubaAnimal Facility, Center for Genetic Engineering and Biotechnology, P.O. Box 6162, Playa Cubanacábn, Havana 10600, Cuba
r t i c l e i n f o
rticle history:eceived 30 May 2011eceived in revised form4 September 2011ccepted 28 October 2011vailable online xxx
eywords:EGFanceraccineSSP
a b s t r a c t
CIGB-247 is a cancer therapeutic, based on recombinant modified human vascular endothelial growthfactor (VEGF) as antigen, in combination with the oil free adjuvant VSSP (very small sized proteoliposomesof Neisseria meningitidis outer membrane). Our previous experimental studies in mice with CIGB-247 haveshown that the vaccine has both anti-tumoral and anti-metastatic activity, and produces both antibodiesthat block VEGF–VEGF receptor interaction, and a specific T-cell cytotoxic response against tumor cells.CIGB-247, with an antigen dose of 100 �g, has been characterized by an excellent safety profile in mice,rats, rabbits, and non human primates.
In this article we extend the immunogenicity and safety studies of CIGB-247 in non human primates,scaling the antigen dose from 100 �g to 200 and 400 �g/vaccination. Our results indicate that such doseescalation did not affect animal behavior, clinical status, and blood parameters and biochemistry. Also,vaccination did not interfere with skin deep skin wound healing. Anti-VEGF IgG antibodies and specificT-cell mediated responses were documented at all three studied doses. Antigen dose apparently did notdetermine differences in maximum antibody titer during the 8 weekly immunization induction phase,or the subsequent increase in antibodies seen for monthly boosters delivered afterwards. Higher antigen
doses had a positive influence in antibody titer maintenance, after cessation of immunizations. Boosterswere important to achieve maximum antibody VEGF blocking activity, and specific T-cell responses inall individuals. Purified IgG from CIGB-247 immunized monkey sera was able to impair proliferationand formation of capillary-like structures in Matrigel, for HMEC cells in culture. Altogether, these resultssupport the further clinical development of the CIGB-247 therapeutic cancer vaccine, and inform on thepotential mechanisms involved in its effect.. Introduction
Active immunization against vascular endothelial factor (VEGF)r its receptors, -the main growth factor system involved in tumorngiogenesis-, is an emerging strategy for controlling tumor growthnd progression [1]. Vaccination with whole endothelial or placen-al cells, and with defined molecular compositions, have proven toe effective in reducing tumor growth in some mouse tumor mod-ls [2,3], and three of these new vaccines strategies have alreadyeached human clinical trials [4–6].
Along this research line, our group has developed the cancer
Please cite this article in press as: Morera Y, et al. Antigen dose escalatioprimates. Vaccine (2011), doi:10.1016/j.vaccine.2011.10.082
accine candidate CIGB-247, a recombinant modified human VEGFntigen, combined with the VSSP bacterial adjuvant (very smallized proteoliposomes (VSSP) from the outer membrane protein
∗ Corresponding author. Tel.: +53 2716022.E-mail address: [email protected] (Y. Morera).
264-410X/$ – see front matter © 2011 Elsevier Ltd. All rights reserved.oi:10.1016/j.vaccine.2011.10.082
© 2011 Elsevier Ltd. All rights reserved.
of Neisseria meningitidis) [7]. Experiments done in C57Bl/6 [8] andBALB/c mice (manuscript in preparation), challenged with fourdifferent experimental tumors, have shown that the vaccine hasboth anti-tumoral and anti-metastatic potential. CIGB-247 inducesa transient immune response of antibodies that block VEGF–VEGFreceptor 2 (VEGFR2; KDR) interaction, and generates specific cyto-toxic T cells [8].
While our vaccine is different in its anti-tumor mechanism andphysiological response to passive immunotherapy with anti-VEGFmonoclonal antibodies like Bevacizumab [9], or synthetic anti-angiogenic drugs [10], the gathering of preclinical safety data isan important task in the further development of CIGB-247, more-over when there is an increasing recognition of the diverse adverseeffects related to the disturbance of VEGF-dependent physiological
n study of a VEGF-based therapeutic cancer vaccine in non human
functions and homeostasis by the aforementioned approved cancerdrugs [11].
Immunization experiments done by us in rats, rabbits and thenon human primate Chlorocebus aethiops sabaeus, using the defined

ING Model
J
2 ccine
a8lts
fmfiem
anseHsmca
2
2
2
Cttd
Bfatlt
2
obti4ha1
ioCwViMr
2
h
ARTICLEVAC-12524; No. of Pages 10
Y. Morera et al. / Va
nti-tumor weekly dose of 100 �g of the antigen per week, for weeks, and a booster a month later, have indicated an excel-
ent safety profile for the vaccine [12]. In this paper we expandhe immunogenicity and safety studies of CIGB-247 in monkeys bycaling up the antigen dose.
Our present study also describes the effects of IgG purifiedrom sera of immunized monkeys on VEGF–VEGFR2 interaction,
icrovascular endothelial cell (HMEC) proliferation, cell cycle, andormation of tubular cellular structures in matrigel, all of whichnforms about the potential anti-angiogenic activity of the vaccine-licited antibodies. T cell responses after immunization were alsoeasured by specific DTH and using an autologous CTL assay.Our results show that the vaccine dose increments did not affect
nimal behavior, clinical status, or blood biochemistry. Also, vacci-ation did not interfere with skin deep wound healing. Purifiederum IgGs from CIGB-247 immunized monkeys inhibit prolif-ration, blocks cell cycle progression, and impair the ability ofMEC cells to form capillary-like structures in Matrigel under VEGF
timulation. DTH was positive in immunized animals and bloodononuclear cells were cytotoxic to peripheral blood mononu-
lear autologous cells, which were previously incubated with thentigen.
. Materials and methods
.1. Animals
.1.1. MonkeysTen adult African green monkeys (Chlorocebus, -formerly
ercopithecus-, aethiops sabaeus) of either sex, weighting from 3o 7 kg, were used. Animals were caged individually in speciallyasked areas and randomized into groups of three to receive eachose. One animal was used as control.
All animals were purchased from the National Center for Animalreeding (CENPALAB, Havana, Cuba), and maintained in the animal
acility of the Center for Genetic Engineering and Biotechnology inccordance with the Cuban guidelines for the care and use of labora-ory animals. Animals were adapted to laboratory conditions for ateast two weeks, and fed with standard laboratory food, accordingo the species.
.2. Vaccine
Recombinant human modified VEGF (P64K-hVEGFKDR−) wasbtained as previously described [8,12]. This antigen is encodedy a human VEGF isoform 121 gene, mutated in three residueso substantially reduce VEGFR2 (KDR) binding, and is expressedn Escherichia coli as an N-terminus fusion protein with the first7 aminoacids of the N. meningitidis (Nm) P64K protein. P64K-VEGFKDR− was purified under GMP conditions using ion metalffinity chromatography (IMAC) and stored liquid at −20 ◦C and
mg/mL until used.For vaccination, the antigen was mixed at the moment of
njection with an adjuvant composed of very small sized prote-liposomes (VSSP) from the outer membrane protein of Nm [8].linical grade preparations (0.8 mg of protein/0.5 mL/vial) of VSSPere supplied by the Center for Molecular Immunology of Havana.SSP is a strong stimulator of both humoral and cell-mediated
mmune responses, known to induce epitope spreading, modulateDSC phenotype, increase dendritic cells maturation, and reduce
egulatory T cells [7,13].
Please cite this article in press as: Morera Y, et al. Antigen dose escalatioprimates. Vaccine (2011), doi:10.1016/j.vaccine.2011.10.082
.3. Other recombinant proteins
GST-fused human VEGF isoform 121 (GST-hVEGF121) as well asuman VEGF isoform 121 (hVEGF121), were produced in E. coli as
PRESSxxx (2011) xxx– xxx
described by Morera et al. [12,14]. Commercial recombinant VEGF-A, isoform 121 (hrVEGF), was purchased from R&D.
N. meningitidis P64K protein was supplied by the DevelopmentDepartment of the Center for Genetic Engineering and Biotechnol-ogy of Havana. P64K is produced routinely to be used as a vaccinecarrier protein [15,16].
2.4. Vaccine doses and schedule
Monkeys were screened first for antibodies to P64K and VEGFproteins, and considered naive with respect to both antigens whenspecific antibodies were undetectable by ELISA (titer <1:50) (seemethods below). Animals were subsequently ranked by weightand age and then randomly assigned to three groups of three ani-mals each. One animal was left without immunization as control.Animals were anesthetized by intramuscular injection of ketaminehydrochloride (10 mg/kg) before immunization. The three groupsof monkeys were immunized with 100, 200 or 400 �g of antigen,respectively. All vaccinations were done combining the antigenwith 200 �g (protein) of VSSP, for a total volume of 0.5 mL.
In the induction phase, the monkeys were subcutaneously vac-cinated once a week, for 8 weeks. Vaccination maintenance phasestarted after the specific antibody titer drop was evident, andanimals received 3 additional immunizations, once every month.Antibody responses were evaluated during the induction and main-tenance phases, and after a final three months rest period.
Blood samples were collected before each new vaccination.Serum from clotted blood was stored at −20 ◦C until used.
2.5. ELISA for anti VEGF antibodies in serum
Antibody titers were measured through an enzyme-linkedimmunosorbent assay (ELISA) as previously described [12]. Serasamples were analyzed for anti-P64K or anti-human VEGF by ELISA.EIA 96-well plates (Costar) were coated overnight at 4 ◦C with10 �g/mL of P64K or GSThVEGF121 in PBS. After three washes with0.1% Tween 20 in PBS, the plates were blocked with 2% skim milkin PBS for 1 h at 22 ◦C, followed by new washes. PBS-diluted serawere added to wells and incubated for 1 h at 22 ◦C. Wells werethen washed three times and incubated with anti human Fc spe-cific antibody (Jackson ImmunoResearch). After incubation for 1 hat 22 ◦C, plates were washed again and incubated with substrate-chromogen solution (OPD 0.75 mg/mL, hydrogen peroxide 0.015%,in citrate-phosphate buffer, pH 5.5) for 15 min. The reaction wasstopped by adding 50 �L of 2 M sulphuric acid solution and theabsorbance was read at 492 nm in a BioRad microtiter plate reader.The 492 nm absorbance value corresponding to a PBS sample wassubtracted from all the obtained diluted serum or plasma val-ues. Non-linear regression curves were adjusted for the OD valuesobtained from the dilutions of each individual sample, and the valuecorresponding to 3 standard deviations greater than the mean ODobtained in wells that contained non-immune samples was inter-polated and considered as the titer.
2.6. ELISA test for serum and antibody blocking of VEGF–VEGFR2interaction in non human primates
Plates were coated overnight at 4 ◦C with 10 �g/mL ofGSThVEGF121 in PBS. After three washes with 0.1% Tween 20 inPBS, the plates were blocked with 2% skim milk in PBS for 1 h at22 ◦C, followed by new washes. Serial dilutions of sera or differentconcentration of purified serum antibodies were added and incu-
n study of a VEGF-based therapeutic cancer vaccine in non human
bated for 1 h at 22 ◦C. Then, 125 �g of recombinant human VEGFreceptor 2/Fc chimera (KDR-Fc; Sigma) were added to the wellsand additionally incubated for 40 min at 22 ◦C. Wells were thenwashed three times and incubated either with rabbit anti-human

ING Model
J
ccine x
FwtAspaaimoFs
2
nCqatiI(wtIis
2
(uauomswodrc
4sptgpwplwcaac“ht
ARTICLEVAC-12524; No. of Pages 10
Y. Morera et al. / Va
c HRPO-conjugated antibodies (Jackson ImmunoResearch), orith goat biotinilated anti-human VEGFR2/KDR antibody (R&D),
he latter followed by avidin peroxidase conjugate (Peprotech).fter washings, plates were incubated with substrate–chromogenolution (OPD 0.75 mg/mL, hydrogen peroxide 0.015%, in citrate-hosphate buffer, pH 5.5) for 15 min. Reaction was stopped bydding 2 M sulphuric acid and the absorbance read at 492 nm in
BioRad microtiter plate reader. The ability of sera to neutral-ze VEGF–VEGFR2 interaction was expressed as a percent of the
aximum binding of KDR-Fc to GST-hVEGF121 in the presencef pre-immune serum, by the formula: % Inhibition of VEGF/KDR-c interaction = 100 − (A492 nm immune serum/A492 nm pre-immuneerum) × 100.
.7. Purification of monkey serum antibodies after vaccination
Monkey IgG was purified from sera of pre-immune and immu-ized animals using affinity chromatography (PROSEP-G Spinolumns; Millipore), as suggested by the manufacturer. IgG wasuantified by ELISA: a 96-well plate was coated with 3 �g/mL ofnti-human kappa light chain antibodies (Sigma), and 50 �L ofhe test or control samples were added per well. After incubat-ng 16 h at 4 ◦C, the reactions were developed using anti-humangG gamma chain antibodies, conjugated with alkaline phosphataseSigma) diluted 1:5000 for 1 h to 37 ◦C. �-nitrophenyl phosphateas employed as substrate. A standard curve of serial 1:2 dilu-
ions, starting at 30 ng/mL, of a humanized anti-EGF receptorgG recombinant antibody (TheraCIM®, CIMAB S.A., Havana) wasncluded in order to quantify the amount of IgG present in theamples.
.8. Delayed type hypersensitivity (DTH) skin testing in monkeys
Animals were sedated with intramuscular ketamine chloride10 mg/kg) prior to invasive or direct manipulations. DTH was eval-ated in all monkeys 4 months after the first immunization. Testntigens included P64K-hVEGFKDR− and hrVEGF. Saline buffer wassed a control. The back of the monkeys were shaved and 60 �Lf the test antigens (100 �g) were injected intradermally in theiddle of circles marked with indelible ink, using 0.5 mL insulin
yringes fitted with 29 gauge needles. After 48 h, the injection sitesere independently assessed by two experienced readers unaware
f animal treatment. Induration diameter was measured with aigital caliper and results were expressed as the group geomet-ic mean area, by antigen [17,18]. Erythema and swelling were notonsidered in the measurement.
Four millimeter punch biopsies were made at selected sites8 h after DTH induction. Paraffin embedded sections (5 �M) weretained with hematoxylin and eosin and reviewed by a veterinaryathologist unaware of group assignment or test antigen. At leastwo sections from each biopsy were examined. For each sample, theeneral nature of the dermal infiltrate was evaluated in terms of theresence of mononuclear cells, neutrophils, or eosinophils. Sectionsere then scored in a semi-quantitative fashion for the amount oferivascular mononuclear cells in the upper dermis, a key histo-
ogical feature of a DTH reaction in monkey skin [17,18]. Scoringas made on a 4+ scale for the amount of perivascular mononu-
lear cell infiltration: 0 = none, 1+ = sparse, 2+ = some, 3+ = modest,
Please cite this article in press as: Morera Y, et al. Antigen dose escalatioprimates. Vaccine (2011), doi:10.1016/j.vaccine.2011.10.082
nd 4+ = abundant. Scores of ≥2+ were considered consistent with DTH reaction, similar to earlier reports whereby DTH reactionsonsisted of perivascular mononuclear cell infiltrate layers ≥5×thicker” than normal or negative control monkey skin [18]. Theistological scores were summarized by group as a general indica-ion of DTH reactivity.
PRESSxx (2011) xxx– xxx 3
2.9. Direct cell cytotoxicity assay with monkey peripheral bloodmononuclear cells (PBMC)
Heparinized blood samples were collected from monkeys sevendays after the last booster. PBMC were isolated by Ficoll gradient.Cells were maintained at a density of 1 × 106 cells/mL in RPMI-1640medium supplemented with 10% FBS (PHA). PBMC from each mon-key (target cells) were incubated or not with the immunizationantigen for 2 h, and later labeled with CFSE as described [19]. Sub-sequently these cells were mixed with autologous PBMC (effectorscells) at a 2:1 ratio and direct cytolysis was evaluated by FACS,expressed as the percent reduction of the CFSE labeled population[20,21].
2.10. Skin wound repair assessment in non human primates
The effects of the vaccine administration on skin deep woundhealing were evaluated using the 4 mm punch biopsy sites done innon human primates for DTH histological assessment. Followinghaemostasis, the wound contours were traced upon transpar-ent plastic sheets for planimetric analysis, and closure dynamicsdetermined using the standard cutaneous round ulcer model [22].Wound closure dynamics was studied at days 0, 6, 12 and 21 and thepercent of wound healing was calculated as the percent of woundarea reduction as compared to the initial values.
2.11. Follow up of clinical, behavioral, and laboratory parametersof non human primates
Each animal was observed daily for general clinical signs.Body weight, rectal temperature, and clinical examination wereassessed before each vaccine administration. After the last immu-nization, the tests and measurements were carried out monthly,to complete a whole one-year observation period. Cageside obser-vations included mortality, general health and signs of toxicity.Physical examinations included evaluation of skin and fur char-acteristics, eye and mucous membranes, respiratory, circulatory,autonomic and central nervous systems, and somatic motor andbehavior patterns. Emphasis was made on detecting hepatomegaly,splenomegaly or regional lymphadenopathy, as well as abnormal-ities at the injection site. Two skill veterinarians performed theseexams.
A toxicity grading system for classifying the local reactionselicited by the vaccine was adapted from the Common Toxicity Cri-teria (CTC) of the National Cancer Institute (NCI) of USA: (a) nodamage – if the skin at the injection site was within normal limits;(b) mild damage – if apparent pain, hardening, itching or erythemawas present; (c) moderate damage – if apparent pain or swellingwith inflammation or phlebitis was present; (d) severe damage – ifsevere or prolonged ulceration or necrosis was present, requiringnot additional care measurements.
Before the third and sixth vaccine administrations, and witha monthly frequency to complete a whole one-year observationperiod, blood samples were taken from femoral veins to deter-mine: hemoglobin, hematocrit, platelets, white blood cell count,neutrophils, monocytes, eosinophils, reticulocytes, alkaline phos-phatase, aspartate aminotransferase, alanine aminotransferase,total bilirubin, albumin, total proteins, glucose and creatinine.
Animals were sedated with intramuscular ketamine chloride(10 mg/kg) prior to invasive or direct manipulations.
2.12. In vitro proliferation assay with HMEC cells
n study of a VEGF-based therapeutic cancer vaccine in non human
For the growth assay, HMEC microendothelial cells were usedbetween passages 18 and 20. One hundred microliters with4000 cells in MCDB-131 medium supplemented with 0.2% FBS

ING Model
J
4 ccine
waCtw(6va(tftr
aon
2
pims(w(wC5fiaD
2
cithpmmiunp
2
lD
pSbmof
A
ARTICLEVAC-12524; No. of Pages 10
Y. Morera et al. / Va
ere seeded per well in 96-well culture plates (Costar). Aftern overnight period, purified IgG from monkeys immunized withIGB-247, purified IgG from human serum (PAA) (negative inhibi-ion control) or Bevacizumab (Roche) (positive inhibition control)ere diluted in serum free culture medium with or without rhVEGF
7.5 ng/mL) and added to cell cultures at a concentration range of5–250 �g/mL. All samples were tested in triplicate wells to a finalolume of 200 �L. After 72 h of incubation at 37 ◦C in 5% CO2–95% airtmosphere, proliferation was measured by the Alamar Blue assayAlamarBlueTM, AbD Serotec) following the manufacturer’s instruc-ions. Briefly, Alamar blue solution (1:10) was added to the culturesor 8 h. To calculate the percent difference in reduction betweenreated and control cells in proliferation assays the absorbance wasead at wavelengths 570 nm and 630 nm.
The results were documented as mean ± SEM, and are expresseds the percentage of proliferating cells relative to the proliferationf endothelial cells, treated with the equivalent concentration ofegative IgG.
.13. Cell cycle analysis
Cells were plated at a density of 1 × 105 cells/well in 6-welllates (Costar, Cambridge, MA). Twenty-four hours later, cells were
ncubated with serum-free medium containing purified IgG fromonkeys immunized with CIGB-247, purified IgG from human
erum (PAA) (negative inhibition control) or Bevacizumab (Roche)positive inhibition control), all at 200 �g/mL. This experimentas carried out with or without the addition of external rhVEGF
7.5 ng/mL). After 72 h incubation, the cells were trypsinized,ashed and permeabilized with cold 70% ethanol for 2 h at 4 ◦C.ellular DNA was stained for 30 min with a solution containing0 �g/mL of propidium iodide and 400 �g/mL RNAse A. DNA pro-le and cell cycle analysis were performed by flow cytometry on
FACS instrument (PAS-III, PARTEC) by collecting 25,000 events.ata were analyzed using the FLOWMAX software (PARTEC).
.14. Capillary-like structures formation by HMEC cells
The effects of the IgGs purified from immunized animals onapillary like structure formation by HMEC cells was analyzedn matrigel. HMEC cells were harvested after non-enzymaticreatment in MCDB-131 medium supplemented with 0.2% FBS;rVEGF 7.5 ng/mL, and immediately added to matrigel-coatedlates (GeltrexTM reduced growth factor basement membraneatrix, Gibco). Purified IgGs were diluted in serum free cultureedium and added to cell cultures at final concentrations rang-
ng the doses: 65–0.250 �g/mL. Control human purified IgG wassed at similar concentration and volumes. Sixteen hour later, theumber of capillary-like structures was counted in each well in ahase contrast microscope (Nikon), at a magnification of 200×.
.15. Statistical analysis
Homogeneity of variances for anti-VEGF titers was ana-yzed with Bartlett’s test and data normality was verified using’Agostino & Pearson omnibus normality test.
Titer values for each point of the antibody kinetics were com-ared for all the animals with the pre-immune values using thetudent paired t-test. In the particular case of titers after theooster, these were compared to the observed before the start of theaintenance phase. In all cases titer increments with dose amount
Please cite this article in press as: Morera Y, et al. Antigen dose escalatioprimates. Vaccine (2011), doi:10.1016/j.vaccine.2011.10.082
r number were analyzed using one-way ANOVA and a post-testor linear trend.
Differences among the dose groups were analyzed by one-wayNOVA, followed by Tukey post-test for multiple comparisons.
PRESSxxx (2011) xxx– xxx
When appropriated student unpaired t-test was used and themeans ± SEM were reported.
Values of p < 0.05 were considered significant at a 95% confi-dence interval. All statistical tests were two-sided, and conductedusing the GraphPad Prism v5.0 statistic package (GraphPad Soft-ware, Inc.).
3. Results
3.1. Antibody response
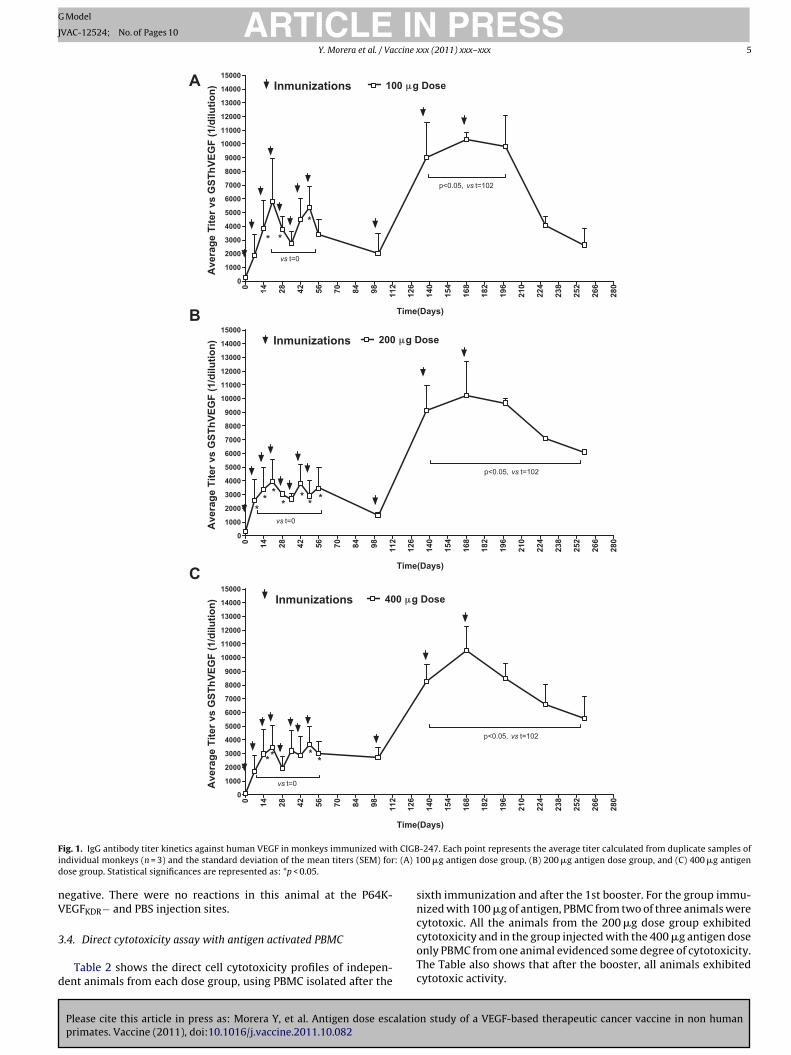
The average anti-human VEGF IgG antibody titers correspond-ing to each blood extraction during the monkey vaccinationexperiments are depicted in Fig. 1. Titers during the inductionphase were compared to pre-immune values using the unpairedt-test. In general, anti human VEGF antibody titers rapidly aug-ment to 1:4000–1:6000 (p < 0.05, paired t-test), following the firstthree immunizations. Further vaccinations maintain titer levels,with some fluctuations between animals for a same dose level.
A decrease in specific IgG antibody titers was detected in thesamples taken 46 days after the eighth dose, being this more abruptfor the animals vaccinated with the 100 �g and 200 �g antigendoses. Monthly immunizations were re-initiated on day 102 for thethree antigen dose groups. Anti-VEGF antibody titers increased toan average of 1:9000 (p < 0.05, paired t-test) after the first boosterfor all groups, and these levels were sustained for the length of themaintenance phase. Titer values had dropped one month after thethird and last maintenance dose, with lower values for the animalsreceiving the dose of 100 �g of antigen (1:3000 vs. ca. 1:6000 forthe 200 and 400 �g of antigen doses).
3.2. Ability of monkey elicited antibodies to block VEGF/KDRinteraction
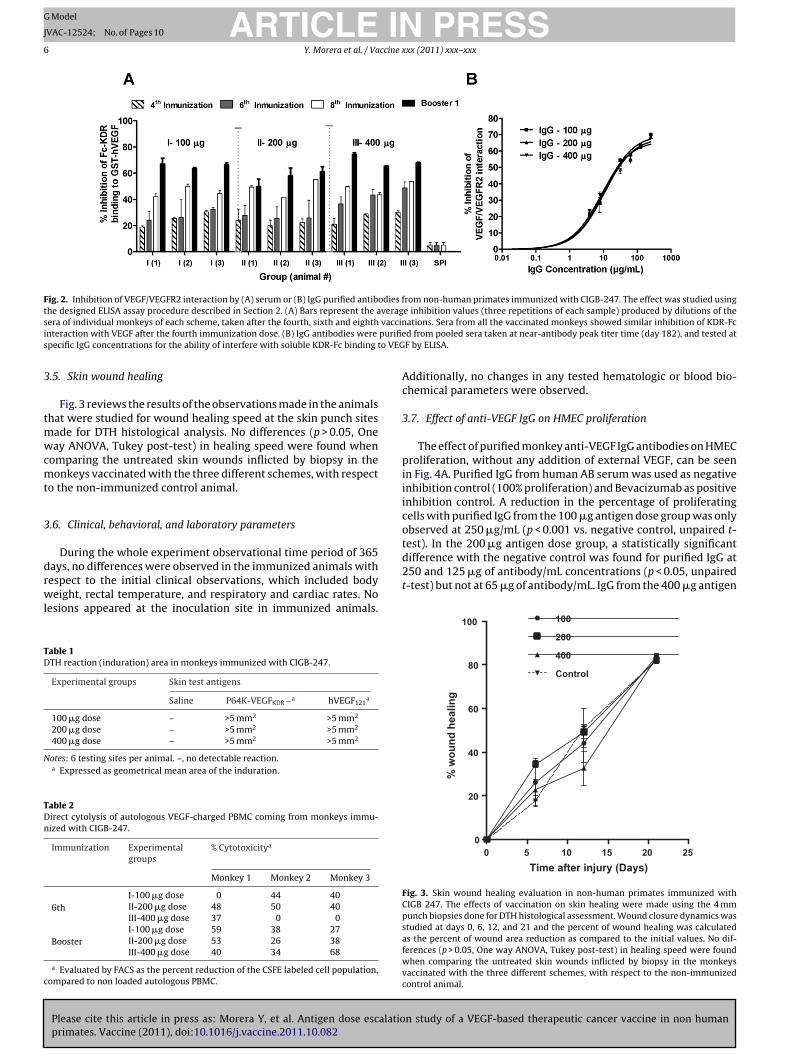
The ability of monkey serum to block KDR-Fc interaction withhuman VEGF was estimated using an ELISA assay. Fig. 2A depictsthe average inhibition values (three repetitions of each sample)produced by 1:500 dilutions of the sera of individual monkeys ofeach scheme, taken after the fourth, sixth, and eighth vaccinations,and after the first booster. The three dose groups exhibited sim-ilar immunization-increasing inhibition values, already differentfrom the pre-immune values in the first sample tested (4th immu-nization; Student paired t-test p < 0.05). Maximum inhibition ofVEGF/KDR-Fc interaction was produced by the sample taken afterthe first booster dose, for all animals, exception made of monkeylabeled number 2 of the 200 �g antigen dose.
IgG antibodies were purified from sera of individual monkeysat peak antibody titer time (day 182), and tested at different con-centrations in the same ELISA inhibition system. Fig. 2B shows thatpurified antibodies from the three experimental groups have sim-ilar specific inhibition activity.
3.3. DTH skin testing
The DTH test was safe and well tolerated, with no adverseevents such as blistering or ulceration. Monkeys from all groupsreacted against hVEGF121 and the P64K-VEGFKDR− vaccine antigen.All reactions at the two VEGF injection sites were histologically con-sistent with a positive DTH. Table 1 depicts the geometrical meanarea of the induration, per dose group, per tested substance. Atthe saline control sites, no reactions (indurations) were reported
n study of a VEGF-based therapeutic cancer vaccine in non human
in any of the immunization groups. The non-immunized controlmonkey used in this experiment developed a 4 mm2 indurationin the hVEGF121 injection sites, but the biopsies showed allergic-like reactions (abundant eosinophils) and were considered DTH
ARTICLE IN PRESSG Model
JVAC-12524; No. of Pages 10
Y. Morera et al. / Vaccine xxx (2011) xxx– xxx 5
0 14 28 42 56 70 84 98 112
126
140
154
168
182
196
210
224
238
252
266
2800
1000
2000
3000
4000
5000
6000
7000
8000
9000
10000
11000
12000
13000
14000
15000 Inmunizations
Time(Days)
100 μg Dose
**
*
vs t=0
p<0.05, vs t=102
Ave
rage
Tite
r vs
GST
hVEG
F (1
/dilu
tion)
0 14 28 42 56 70 84 98 112
126
140
154
168
182
196
210
224
238
252
266
2800
1000
2000
3000
4000
5000
6000
7000
8000
9000
10000
11000
12000
13000
14000
15000 Inmunizations
Time(Days)
400 μg Dose
** **
vs t=0
p<0.05, vs t=102
Ave
rage
Tite
r vs
GST
hVEG
F (1
/dilu
tion)
0 14 28 42 56 70 84 98 112
126
140
154
168
182
196
210
224
238
252
266
2800
1000
2000
3000
4000
5000
6000
7000
8000
9000
10000
11000
12000
13000
14000
15000 Inmunizations
Time(Days)
200 μg Dose
**
*** * *
vs t=0
p<0.05, vs t=102
Ave
rage
Tite
r vs
GST
hVEG
F (1
/dilu
tion)
A
B
C
F h CIGBi : (A) 1d
nV
3
d
ig. 1. IgG antibody titer kinetics against human VEGF in monkeys immunized witndividual monkeys (n = 3) and the standard deviation of the mean titers (SEM) forose group. Statistical significances are represented as: *p < 0.05.
egative. There were no reactions in this animal at the P64K-EGFKDR− and PBS injection sites.
Please cite this article in press as: Morera Y, et al. Antigen dose escalatioprimates. Vaccine (2011), doi:10.1016/j.vaccine.2011.10.082
.4. Direct cytotoxicity assay with antigen activated PBMC
Table 2 shows the direct cell cytotoxicity profiles of indepen-ent animals from each dose group, using PBMC isolated after the
-247. Each point represents the average titer calculated from duplicate samples of00 �g antigen dose group, (B) 200 �g antigen dose group, and (C) 400 �g antigen
sixth immunization and after the 1st booster. For the group immu-nized with 100 �g of antigen, PBMC from two of three animals werecytotoxic. All the animals from the 200 �g dose group exhibited
n study of a VEGF-based therapeutic cancer vaccine in non human
cytotoxicity and in the group injected with the 400 �g antigen doseonly PBMC from one animal evidenced some degree of cytotoxicity.The Table also shows that after the booster, all animals exhibitedcytotoxic activity.
ARTICLE IN PRESSG Model
JVAC-12524; No. of Pages 10
6 Y. Morera et al. / Vaccine xxx (2011) xxx– xxx
Fig. 2. Inhibition of VEGF/VEGFR2 interaction by (A) serum or (B) IgG purified antibodies from non-human primates immunized with CIGB-247. The effect was studied usingthe designed ELISA assay procedure described in Section 2. (A) Bars represent the average inhibition values (three repetitions of each sample) produced by dilutions of thes vaccini purifies o VEG
3
tmwcmt
3
drwl
TD
N
TDn
c
test). In the 200 �g antigen dose group, a statistically significantdifference with the negative control was found for purified IgG at250 and 125 �g of antibody/mL concentrations (p < 0.05, unpaired
era of individual monkeys of each scheme, taken after the fourth, sixth and eighthnteraction with VEGF after the fourth immunization dose. (B) IgG antibodies were
pecific IgG concentrations for the ability of interfere with soluble KDR-Fc binding t
.5. Skin wound healing
Fig. 3 reviews the results of the observations made in the animalshat were studied for wound healing speed at the skin punch sites
ade for DTH histological analysis. No differences (p > 0.05, Oneay ANOVA, Tukey post-test) in healing speed were found when
omparing the untreated skin wounds inflicted by biopsy in theonkeys vaccinated with the three different schemes, with respect
o the non-immunized control animal.
.6. Clinical, behavioral, and laboratory parameters
During the whole experiment observational time period of 365ays, no differences were observed in the immunized animals with
Please cite this article in press as: Morera Y, et al. Antigen dose escalatioprimates. Vaccine (2011), doi:10.1016/j.vaccine.2011.10.082
espect to the initial clinical observations, which included bodyeight, rectal temperature, and respiratory and cardiac rates. No
esions appeared at the inoculation site in immunized animals.
able 1TH reaction (induration) area in monkeys immunized with CIGB-247.
Experimental groups Skin test antigens
Saline P64K-VEGFKDR−a hVEGF121a
100 �g dose – >5 mm2 >5 mm2
200 �g dose – >5 mm2 >5 mm2
400 �g dose – >5 mm2 >5 mm2
otes: 6 testing sites per animal. –, no detectable reaction.a Expressed as geometrical mean area of the induration.
able 2irect cytolysis of autologous VEGF-charged PBMC coming from monkeys immu-ized with CIGB-247.
Immunization Experimentalgroups
% Cytotoxicitya
Monkey 1 Monkey 2 Monkey 3
6thI-100 �g dose 0 44 40II-200 �g dose 48 50 40III-400 �g dose 37 0 0
BoosterI-100 �g dose 59 38 27II-200 �g dose 53 26 38III-400 �g dose 40 34 68
a Evaluated by FACS as the percent reduction of the CSFE labeled cell population,ompared to non loaded autologous PBMC.
ations. Sera from all the vaccinated monkeys showed similar inhibition of KDR-Fcd from pooled sera taken at near-antibody peak titer time (day 182), and tested atF by ELISA.
Additionally, no changes in any tested hematologic or blood bio-chemical parameters were observed.
3.7. Effect of anti-VEGF IgG on HMEC proliferation
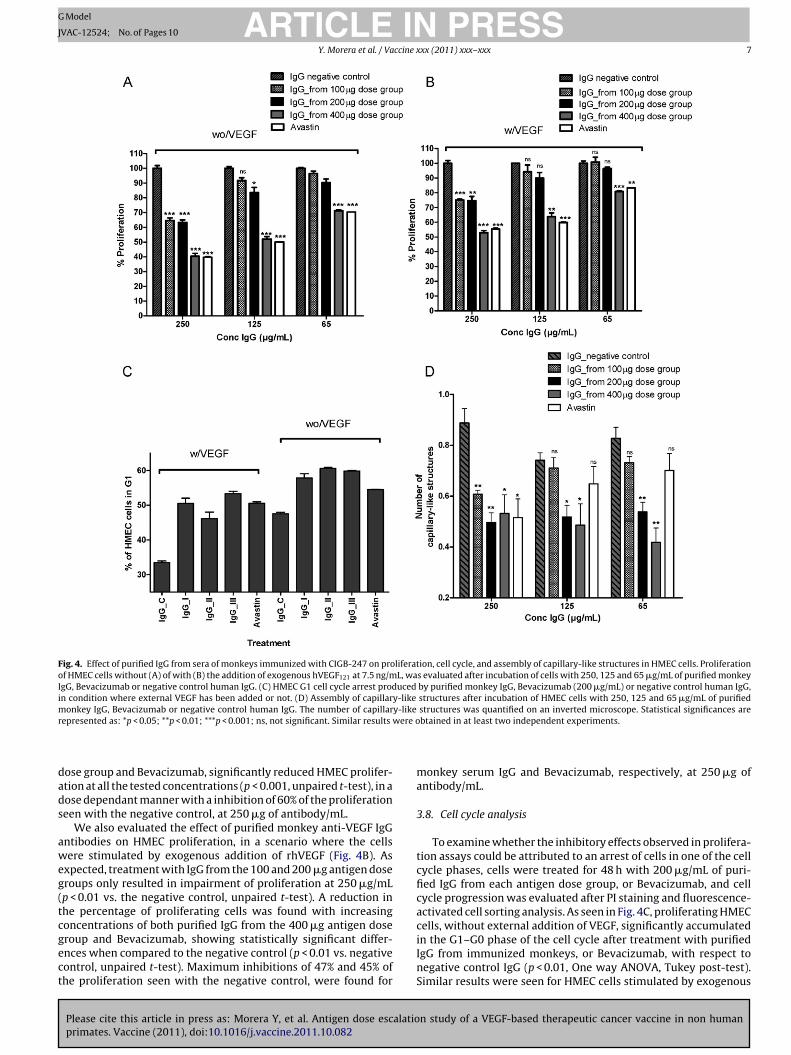
The effect of purified monkey anti-VEGF IgG antibodies on HMECproliferation, without any addition of external VEGF, can be seenin Fig. 4A. Purified IgG from human AB serum was used as negativeinhibition control (100% proliferation) and Bevacizumab as positiveinhibition control. A reduction in the percentage of proliferatingcells with purified IgG from the 100 �g antigen dose group was onlyobserved at 250 �g/mL (p < 0.001 vs. negative control, unpaired t-
n study of a VEGF-based therapeutic cancer vaccine in non human
t-test) but not at 65 �g of antibody/mL. IgG from the 400 �g antigen
0 5 10 15 20 250
20
40
60
80
100 100
200
400
Control
Time after injury (Days)
% w
ound
hea
ling
Fig. 3. Skin wound healing evaluation in non-human primates immunized withCIGB 247. The effects of vaccination on skin healing were made using the 4 mmpunch biopsies done for DTH histological assessment. Wound closure dynamics wasstudied at days 0, 6, 12, and 21 and the percent of wound healing was calculatedas the percent of wound area reduction as compared to the initial values. No dif-ferences (p > 0.05, One way ANOVA, Tukey post-test) in healing speed were foundwhen comparing the untreated skin wounds inflicted by biopsy in the monkeysvaccinated with the three different schemes, with respect to the non-immunizedcontrol animal.
ARTICLE IN PRESSG Model
JVAC-12524; No. of Pages 10
Y. Morera et al. / Vaccine xxx (2011) xxx– xxx 7
Fig. 4. Effect of purified IgG from sera of monkeys immunized with CIGB-247 on proliferation, cell cycle, and assembly of capillary-like structures in HMEC cells. Proliferationof HMEC cells without (A) of with (B) the addition of exogenous hVEGF121 at 7.5 ng/mL, was evaluated after incubation of cells with 250, 125 and 65 �g/mL of purified monkeyIgG, Bevacizumab or negative control human IgG. (C) HMEC G1 cell cycle arrest produced by purified monkey IgG, Bevacizumab (200 �g/mL) or negative control human IgG,in condition where external VEGF has been added or not. (D) Assembly of capillary-like structures after incubation of HMEC cells with 250, 125 and 65 �g/mL of purifiedmonkey IgG, Bevacizumab or negative control human IgG. The number of capillary-like structures was quantified on an inverted microscope. Statistical significances arer were o
dads
aweg(tcgect
epresented as: *p < 0.05; **p < 0.01; ***p < 0.001; ns, not significant. Similar results
ose group and Bevacizumab, significantly reduced HMEC prolifer-tion at all the tested concentrations (p < 0.001, unpaired t-test), in aose dependant manner with a inhibition of 60% of the proliferationeen with the negative control, at 250 �g of antibody/mL.
We also evaluated the effect of purified monkey anti-VEGF IgGntibodies on HMEC proliferation, in a scenario where the cellsere stimulated by exogenous addition of rhVEGF (Fig. 4B). As
xpected, treatment with IgG from the 100 and 200 �g antigen doseroups only resulted in impairment of proliferation at 250 �g/mLp < 0.01 vs. the negative control, unpaired t-test). A reduction inhe percentage of proliferating cells was found with increasingoncentrations of both purified IgG from the 400 �g antigen dose
Please cite this article in press as: Morera Y, et al. Antigen dose escalatioprimates. Vaccine (2011), doi:10.1016/j.vaccine.2011.10.082
roup and Bevacizumab, showing statistically significant differ-nces when compared to the negative control (p < 0.01 vs. negativeontrol, unpaired t-test). Maximum inhibitions of 47% and 45% ofhe proliferation seen with the negative control, were found for
btained in at least two independent experiments.
monkey serum IgG and Bevacizumab, respectively, at 250 �g ofantibody/mL.
3.8. Cell cycle analysis
To examine whether the inhibitory effects observed in prolifera-tion assays could be attributed to an arrest of cells in one of the cellcycle phases, cells were treated for 48 h with 200 �g/mL of puri-fied IgG from each antigen dose group, or Bevacizumab, and cellcycle progression was evaluated after PI staining and fluorescence-activated cell sorting analysis. As seen in Fig. 4C, proliferating HMECcells, without external addition of VEGF, significantly accumulated
n study of a VEGF-based therapeutic cancer vaccine in non human
in the G1–G0 phase of the cell cycle after treatment with purifiedIgG from immunized monkeys, or Bevacizumab, with respect tonegative control IgG (p < 0.01, One way ANOVA, Tukey post-test).Similar results were seen for HMEC cells stimulated by exogenous

ING Model
J
8 ccine
had
3
oMootio2itw2
4
tsHdddtc[b[
msintedcov
rdorhiaTcs
otatesot
ARTICLEVAC-12524; No. of Pages 10
Y. Morera et al. / Va
rVEGF at 7.5 ng/mL. The highest number of cells in G1 cell cyclerrest (20%) was detected with purified IgG from the 400 �g antigenose group.
.9. Assembly into capillary-like structures by HMEC cells
To address the effect of the purified monkey IgG in the assemblyf capillary-like structures by HMEC cells, these were cultured inatrigel coated wells and incubated with different concentrations
f immunoglobulins from each antigen dose group, Bevacizumabr negative control human IgG. The number of capillary-like struc-ures formed was quantified on an inverted microscope 16 h afterncubation. Fig. 4D shows that a significant reduction in the numberf structures was found after incubation with purified IgG from the00 and 400 �g antigen dose groups at all tested concentrations,
n comparison to negative control human IgG (p < 0.01, unpaired t-est). IgG from the 100 �g antigen dose group and Bevacizumabere only able to impair capillary-like structures formation at
50 �g/mL.
. Discussion
There is already a substantial amount of clinical informationhat validates as promising to target the VEGF–VEGF receptorsystem for the treatment of different human tumors [23–28].owever, in cancer patients, probably because of the large drugoses needed to be applied systemically, synthetic anti-angiogenicrugs (like Sorafenib and Sunitinib), and Bevacizumab, have pro-uced serious collateral side effects such as hypertension, arterialhromboembolic events, proteinuria or renal dysfunction, woundomplications, and gastrointestinal hemorrhage and perforation11], that may be associated with the undesired simultaneous inhi-ition of some normal physiological properties of circulating VEGF29].
It is noteworthy to point out that our vaccine exhibits effectorechanisms that differ from those associated to anti-angiogenic
ynthetic drugs, and anti-VEGF therapeutic antibodies now usedn cancer treatment. Firstly, CIGB-247 is designed to affect tumoreo-vascularization and tumor cell viability by eliciting antibodieshat block the interaction of VEGF and its receptors in activatedndothelial cells, as well as specific T cytotoxic cells, that can killirectly tumor and tumor stroma cells that produce VEGF. Also,ancer cell elimination via tumor epitope spreading cannot be ruledut within the chronic vaccination scheme envisioned by us for theaccine, due to the properties of the employed adjuvant [30,31].
Secondly, being our antigen a self molecule, the immuneesponse will be physiologically controlled and its maintenanceepends on chronic vaccination, as it has been shown already forther similar vaccines [32]. In the case of the anti-VEGF humoralesponse to the vaccine, this means that, instead of relying on veryigh amounts of externally infused blocking monoclonal antibod-
es, the CIGB-247 vaccine will act via smaller (metronomic-like)mounts of effector antibodies, over an extended period of time.he vaccine also will produce a T cytotoxic cell response, acting inombination with the specific antibodies, and under a potentiallyimilar well controlled magnitude regulation.
In a previous paper [12] we commented that the amountf VEGF-blocking polyclonal antibodies produced in monkeyshrough our vaccination scheme, including the first booster, isround between two and three orders of magnitude lower thanhe maximum physiological concentration transiently achieved, for
Please cite this article in press as: Morera Y, et al. Antigen dose escalatioprimates. Vaccine (2011), doi:10.1016/j.vaccine.2011.10.082
xample, by Bevacizumab infusion, in preclinical pharmacologicaltudies [33]. Vaccination as we propose thus may allow the creationf a discrete but quasi-continuous pharmacological effect windowhat controls tumor growth, but at the same time spares normal
PRESSxxx (2011) xxx– xxx
tissues where physiological blood vessel repair and maintenanceby VEGF are needed.
Being the CIGB-247 vaccine candidate on the verge of enteringPhase I Clinical Trials in cancer patients, the present paper pro-vides not only additional support on safety, absence of effects onskin wound healing while escalating the antigen dose, but alsoimmune response kinetic information that supports our generaleffect mechanisms hypothesis, described above.
The article also provides a deeper look on the potential of theinduced anti-VEGF polyclonal antibodies to produce experimentalanti-angiogenic effects, where antigen dose in the vaccine seemsto be a parameter to take into account.
The immunization scheme used by us – an induction phasebased on weekly subcutaneous immunizations, followed bymonthly boosters using the same route –, has been previously val-idated by a number of studies conducted in mice, rats, rabbits andnon-human primates [5,6]. Weekly immunizations have producedanti-tumoral and anti-metastatic effects in different mouse strainsand tumors [34]. As compared to bi-weekly schedules, weeklyimmunization produces an increasing humoral anti-VEGF responsewith faster kinetics, as well as a higher percent of effective lysisof autologous PBMC loaded with the immunization antigen [12].Tolerance to the self VEGF molecule, in terms of producing specificand increasing humoral and cellular responses, is effectively brokenusing the used immunization scheme.
In the study reported in this paper we have maintainedthe weekly immunization schemes in non-human primates, butincreasing the antigen doses in CIGB-247 from the previously tested100 �g [12], to 200 and 400 �g. In terms of the IgG specific anti-body titer levels, our results suggest that monkey behavior toimmunization, during the induction and maintenance phases, issimilar in all studied doses. Induction titers plateau after the 3rdor 4th vaccination, and double their magnitude after a resting timeand subsequent booster. The ability of elicited antibodies to blockVEGF–VEGFR2 interaction in an ELISA assay seem also non anti-gen dose dependent, but nevertheless this property climbs for allantigen doses with each subsequent injection during the inductionphase, and peaks after the first booster. Our blocking assays usingpurified monkey IgG are also consistent with the fact that, at leastwithin the limits of the ELISA assay, the neutralizing ability of IgGantibodies produced by different antigen doses seem to be undis-tinguishable. From our experiments, it is evident that boosteringthe immune system after the induction phase and a short restingtime is relevant to achieve maximum antibody titer, antibody VEGFblocking ability and T-cell mediated responses. Boosters also eraseindividual initial differences in response to immunization, whichcan be seen during the induction phase.
It has been recognized that the relationship between vaccineantigen dose and the quantity and quality of the elicited immuneresponse, and of the desired final effect can vary depending on theantigen type, adjuvant, scheme, and species. In these scenarios, theso called “window” of immunogenicity can be broad, with too muchor too little antigen inducing tolerance, suppression or immunepolarization towards a cellular or a humoral response. While formany foreign antigens (viral, bacterial, synthetic), a linear relation-ship of immune response with dose increments has been described,this is hardly the case of auto-antigens or self molecules, due to theregulatory pressures of the immune system [35]. For some self anti-gens, it has been described that increasing antigen dose does notnecessarily produce a statistically significant difference in specificIgG titers [32,36].
For CIGB-247, while antigen dose produces no major detectable
n study of a VEGF-based therapeutic cancer vaccine in non human
effect in achievable maximum antibody titers using the weekly vac-cination scheme, there are some aspects that, while may not besusceptible of a profound statistical analysis due to the low num-ber of animals used, nevertheless merit some comment. The first of

ING Model
J
ccine x
tstbftthti
mtau
imtPVaTtaIiecaVcaaaes
defhavAptf
[hAand
smIbadih
t
[
[
[
[
[
ARTICLEVAC-12524; No. of Pages 10
Y. Morera et al. / Va
hese issues deals with the drop in titer after induction vaccinationtops, which appear to be slightly different for 100 �g, with respecto the other two doses. Also, the titer drop kinetics after the lastooster is different, with a longer retention of higher titer valuesor the 400 �g antigen dose. These findings may prove important forhe practical application of the CIGB-247 vaccine, as could indicatehat a higher antigen dose could prolong the period of time withigher circulating antibody titers, while lower doses could openhe way to a tighter regulation of the antibody immune response,f desired.
Finally, it does appear that the relative contribution of differentechanisms to the overall quality of the immune response is likely
o vary between individuals, as we found no correlation betweenntibody titer, blocking ability of antibodies, and CTL activity val-es, per animal.
In the present study we demonstrate for the first time thatmmunoglobulins purified from sera of CIGB-247 vaccinated ani-
als, produce biological effects in culture cells of endothelial origin,hat are strongly related to their possible anti-angiogenic potential.urified immune monkey IgG was able to prevent the autocrineEGF proliferation stimulation loop of HMEC culture cells [37], byrresting the cells in the cell-cycle G1 stage, similar to Bevacizumab.he anti-VEGF antibodies contained in the IgG monkey prepara-ions, at least under these experimental conditions; act mainly as
cytostatic rather than as a cytotoxic agent in vitro. The monkeygG antibodies also inhibited the ability HMEC cells to assemblento tubular vessel-like structures in the Matrigel assay. Thesexperiments indicate that, at similar total immunoglobulin con-entrations, the monkey polyclonal IgG antibodies are as efficients Bevacizumab. Considering that the concentration of specific anti-EGF IgG antibodies in the hyperimmune animal serum could belose to 10% of the total IgG concentration value [38], these resultsre encouraging. According to these evidences, while the totalmount of anti-VEGF IgG antibodies in the blood of immunizednimals could be much less than the concentration achieved byxternally infused monoclonal antibodies, their quality could beimilar or even superior.
With respect to the above discussion of the impact of antigenose in the quantity and quality of the immune response, the in vitroxperiments also provided us with evidence that the IgG samplesrom the animals receiving the highest vaccination antigen doseave an increased effect in the HMEC proliferation and Matrigelssays. This result was unexpected, as we had noticed no rele-ant differences in the VEGF–VEGFR2 ELISA blocking ability test.t present we could only speculate that the in vitro tests are moreowerful and add a different dimension to the testing of the poten-ial biological effect of the antibodies. This matter will be testedurther in other experiments.
VEGF and VEGF receptor experimental vaccine studies in mice8,39–43] and dogs [44], conducted by us and several other authorsave already indicated the high safety profile of such procedures.fter treating many hundreds of mice in our anti-tumor protocols,s well as rats, rabbits, and non human primates [8,12], we haveot found a single case where substance-related toxicity could beocumented.
This new study supports the aforementioned, with the out-tanding feature of the relatively longer period over which theonkeys were evaluated, and the testing of larger antigen doses.
n brief, we found no evidence of clinical, histological, hematology,lood clotting during venipuncture or blood biochemistry alter-tions in the non human primates, even with the highest antigenose employed for vaccination. Additionally, we did not observe
Please cite this article in press as: Morera Y, et al. Antigen dose escalatioprimates. Vaccine (2011), doi:10.1016/j.vaccine.2011.10.082
nterference of any vaccination schedule with skin deep woundealing.
Altogether, these preclinical results have reassuring effect onhe further clinical development of the CIGB-247 therapeutic
[
PRESSxx (2011) xxx– xxx 9
cancer vaccine. As mentioned in the Introduction, only a handful ofactive immunotherapy procedures that target the VEGF-receptorssystem have arrived to the clinical trial setting. A recent pilot studyshowed that a vaccine based on fixed whole endothelium cellswas safe in patients with a progressive malignancy [6]. Miyazawaet al. vaccinated patients of advanced pancreatic cancer with a HLA-A*2402 restricted peptide against VEGFR2. This protocol was welltolerated [4]. Finally, Nagayama et al. [5] conducted a clinical phaseI/II trial immunizing with a HLA-A*2402/A*0201-restricted vascu-lar endothelial growth factor receptor type 1 (VEGFR1)-derivedpeptide, in combination with gemcitabine in advanced pancre-atic cancer. This trial was terminated prematurely because of theadvanced stage of the disease in the enrolled patients, some ofwhich suffered gastrointestinal bleeding episodes.
These mixed results have to be looked at with caution, and prob-ably indicate that vaccines that target the ligand or the receptorshave to be planned and examined under a different lens. In any case,careful patient selection and close monitoring will be extremelyrelevant in future clinical studies with the CIGB-247 vaccine.
Acknowledgements
This work was supported by the Center for Genetic Engineeringand Biotechnology, and Biorec.
References
[1] Matejuk A, Leng Q, Chou ST, Mixson AJ. Vaccines targeting the neovasculatureof tumors. Vasc Cell 2011;3(March (1)):7.
[2] Okaji Y, Tsuno NH, Saito S, Yoneyama S, Tanaka M, Nagawa H, et al. Vaccinestargeting tumour angiogenesis – a novel strategy for cancer immunotherapy.Eur J Surg Oncol 2006;32(May (4)):363–70.
[3] Wei YQ. Immunotherapy of tumors with vaccines based on xenogeneic homol-ogous molecules. Anticancer Drugs 2002;13(March (3)):229–35.
[4] Miyazawa M, Ohsawa R, Tsunoda T, Hirono S, Kawai M, Tani M, et al. PhaseI clinical trial using peptide vaccine for human vascular endothelial growthfactor receptor 2 in combination with gemcitabine for patients with advancedpancreatic cancer. Cancer Sci 2010;101(February (2)):433–9.
[5] Nagayama H, Kazufumi M, Naoyuki I, Hideki O, Naoyuki T, Takashi N, et al.Gastrointestinal bleeding during anti-angiogenic peptide vaccination in com-bination with gemcitabine for advanced pancreatic cancer. Clin J Gastroenterol2010, doi:10.1007/s12328-010-0178-5.
[6] Okaji Y, Tsuno NH, Tanaka M, Yoneyama S, Matsuhashi M, Kitayama J, et al. Pilotstudy of anti-angiogenic vaccine using fixed whole endothelium in patientswith progressive malignancy after failure of conventional therapy. Eur J Cancer2008;44(February (3)):383–90.
[7] Mesa C, de LJ, Fernandez LE. Very small size proteoliposomes derived fromNeisseria meningitidis: an effective adjuvant for generation of CTL responses topeptide and protein antigens. Vaccine 2006;24(March (14)):2692–9.
[8] Morera Y, Bequet-Romero M, Ayala M, Lamdan H, Agger EM, Andersen P, et al.Anti-tumoral effect of active immunotherapy in C57BL/6 mice using a recombi-nant human VEGF protein as antigen and three chemically unrelated adjuvants.Angiogenesis 2008;11(4):381–93.
[9] Ferrara N, Hillan KJ, Novotny W. Bevacizumab (Avastin), a humanized anti-VEGF monoclonal antibody for cancer therapy. Biochem Biophys Res Commun2005;333(July (2)):328–35.
10] Hersey P, Bastholt L, Chiarion-Sileni V, Cinat G, Dummer R, Eggermont AM,et al. Small molecules and targeted therapies in distant metastatic disease. AnnOncol 2009;20(August (Suppl. 6)):vi35–40.
11] Chen HX, Cleck JN. Adverse effects of anticancer agents that target the VEGFpathway. Nat Rev Clin Oncol 2009;6(August (8)):465–77.
12] Morera Y, Bequet-Romero M, Ayala M, Velazco JC, Perez PP, Alba JS, et al.Immunogenicity and some safety features of a VEGF-based cancer therapeu-tic vaccine in rats, rabbits and non-human primates. Vaccine 2010;28(April(19)):3453–61.
13] Fernandez A, Mesa C, Marigo I, Dolcetti L, Clavell M, Oliver L, et al. Inhibitionof tumor-induced myeloid-derived suppressor cell function by a nanoparticu-lated adjuvant. J Immunol 2011;186(January (1)):264–74.
14] Morera Y, Lamdan H, Bequet M, Ayala M, Rojas G, Munoz Y, et al. Biologicallyactive vascular endothelial growth factor as a bacterial recombinant glu-tathione S-transferase fusion protein. Biotechnol Appl Biochem 2006;44(April
n study of a VEGF-based therapeutic cancer vaccine in non human
(Pt 1)):45–53.15] Gonzalez G, Crombet T, Catala M, Mirabal V, Hernandez JC, Gonzalez Y, et al.
A novel cancer vaccine composed of human-recombinant epidermal growthfactor linked to a carrier protein: report of a pilot clinical trial. Ann Oncol1998;9(April (4)):431–5.

ING Model
J
1 ccine
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
growth factor induces an anti-tumor response in C57Bl/6 mice. Angiogenesis
ARTICLEVAC-12524; No. of Pages 10
0 Y. Morera et al. / Va
16] Rodriguez PC, Rodriguez G, Gonzalez G, Lage A. Clinical development andperspectives of CIMAvax EGF, Cuban vaccine for non-small-cell lung cancertherapy. MEDICC Rev 2010;12(1):17–23.
17] Walsh DS, Gettayacamin M, Leitner WW, Lyon JA, Stewart VA, Marit G,et al. Heterologous prime-boost immunization in rhesus macaques by two,optimally spaced particle-mediated epidermal deliveries of Plasmodium fal-ciparum circumsporozoite protein-encoding DNA, followed by intramuscularRTS, S/AS02A. Vaccine 2006;24(May (19)):4167–78.
18] Yamashita S, Hirano K, Kuwasako T, Janabi M, Toyama Y, Ishigami M,et al. Physiological and pathological roles of a multi-ligand receptor CD36in atherogenesis; insights from CD36-deficient patients. Mol Cell Biochem2007;299(May (1–2)):19–22.
19] Wang XQ, Duan XM, Liu LH, Fang YQ, Tan Y. Carboxyfluorescein diacetatesuccinimidyl ester fluorescent dye for cell labeling. Acta Biochim Biophys Sin(Shanghai) 2005;37(June (6)):379–85.
20] Livingston PG, Kurane I, Ennis FA. Use of Epstein-Barr virus-transformed, autol-ogous B-lymphoblastoid cells as antigen-presenting cells for establishment andmaintenance of dengue virus-specific, human cytotoxic T lymphocyte clones.J Virol Methods 1997;67(August (1)):77–84.
21] Godoy-Ramirez K, Makitalo B, Thorstensson R, Sandstrom E, Biberfeld G, GainesH. A novel assay for assessment of HIV-specific cytotoxicity by multiparameterflow cytometry. Cytometry A 2005;68(December (2)):71–80.
22] Berlanga J, Cibrian D, Guillen I, Freyre F, Alba JS, Lopez-Saura P, et al. Methylgly-oxal administration induces diabetes-like microvascular changes and perturbsthe healing process of cutaneous wounds. Clin Sci (Lond) 2005;109(July(1)):83–95.
23] Bukowski RM. Targeted therapies bevacizumab and interferon-alpha inmetastatic renal-cell carcinoma. Nat Rev Clin Oncol 2009;6(May (5)):253–4.
24] Cohen MH. FDA drug approval summary: bevacizumab (Avastin) as treat-ment of recurrent glioblastoma multiforme. Oncologist 2009;14(November(11)):1131–8.
25] Hurwitz H. Bevacizumab plus irinotecan, fluorouracil, and leucovorin formetastatic colorectal cancer. N Engl J Med 2004;350(June (23)):2335–42.
26] Miller K. Paclitaxel plus bevacizumab versus paclitaxel alone for metastaticbreast cancer. N Engl J Med 2007;357(December (26)):2666–76.
27] Sandler A. Paclitaxel-carboplatin alone or with bevacizumab for non-small-celllung cancer. N Engl J Med 2006;355(December (24)):2542–50.
28] Subramanian ML, Abedi G, Ness S, Ahmed E, Fenberg M, Daly MK, et al.Bevacizumab vs ranibizumab for age-related macular degeneration: 1-yearoutcomes of a prospective, double-masked randomised clinical trial. Eye (Lond)2010;24(November (11)):1708–15.
29] Ferrara N. The role of VEGF in the regulation of physiological and pathological
Please cite this article in press as: Morera Y, et al. Antigen dose escalatioprimates. Vaccine (2011), doi:10.1016/j.vaccine.2011.10.082
angiogenesis. EXS 2005;20(94):9–31.30] Guthmann MD, Bitton RJ, Carnero AJ, Gabri MR, Cinat G, Koliren L, et al. Active
specific immunotherapy of melanoma with a GM3 ganglioside-based vaccine:a report on safety and immunogenicity. J Immunother 2004;27(November(6)):442–51.
[
PRESSxxx (2011) xxx– xxx
31] Vanderlugt CL, Miller SD. Epitope spreading in immune-mediated dis-eases: implications for immunotherapy. Nat Rev Immunol 2002;2(February(2)):85–95.
32] Gonzalez G, Crombet T, Neninger E, Viada C, Lage A. Therapeutic vaccinationwith epidermal growth factor (EGF) in advanced lung cancer: analysis of pooleddata from three clinical trials. Hum Vaccin 2007;3(January (1)):8–13.
33] Lin YS, Nguyen C, Mendoza JL, Escandon E, Fei D, Meng YG, et al. Preclinicalpharmacokinetics, interspecies scaling, and tissue distribution of a humanizedmonoclonal antibody against vascular endothelial growth factor. J PharmacolExp Ther 1999;288(January (1)):371–8.
34] Bequet-Romero M, Morera Y, Ayala-Ávila M, AJ, Soria Y, Blanco A, Suárez-AlbaJ, et al. CIGB-247. A VEGF-based therapeutic vaccine that reduces experimen-tal and spontaneous lung metastasis in C57Bl/6 and BALB/c mice. Vaccine,submitted for publication.
35] Cohen IR, Young DB. Autoimmunity, microbial immunity and the immunolog-ical homunculus. Immunol Today 1991;12(April (4)):105–10.
36] Ullenhag GJ, Frodin JE, Jeddi-Tehrani M, Strigard K, Eriksson E, Samanci A,et al. Durable carcinoembryonic antigen (CEA)-specific humoral and cellularimmune responses in colorectal carcinoma patients vaccinated with recombi-nant CEA and granulocyte/macrophage colony-stimulating factor. Clin CancerRes 2004;10(May (10)):3273–81.
37] Costa R, Carneiro A, Rocha A, Pirraco A, Falcao M, Vasques L, et al. Bevacizumaband ranibizumab on microvascular endothelial cells: a comparative study. J CellBiochem 2009;108(December (6)):1410–7.
38] Harlow E, Lane D. Using antibodies: a laboratory manual. Cold Spring HarborLaboratory Press; 1999, ISBN 0879695447. Ref Type: Generic.
39] Li Y, Wang MN, Li H, King KD, Bassi R, Sun H, et al. Active immunization againstthe vascular endothelial growth factor receptor flk1 inhibits tumor angiogen-esis and metastasis. J Exp Med 2002;195(June (12)):1575–84.
40] Liu JY, Wei YQ, Yang L, Zhao X, Tian L, Hou JM, et al. Immunotherapy of tumorswith vaccine based on quail homologous vascular endothelial growth factorreceptor-2. Blood 2003;102(September (5)):1815–23.
41] Wei YQ, Huang MJ, Yang L, Zhao X, Tian L, Lu Y, et al. Immunogene therapyof tumors with vaccine based on Xenopus homologous vascular endothelialgrowth factor as a model antigen. Proc Natl Acad Sci U S A 2001;98(September(20)):11545–50.
42] Rad FH, Le BH, Paturance S, Larcier P, Genne P, Ryffel B, et al. VEGF kinoidvaccine, a therapeutic approach against tumor angiogenesis and metastases.Proc Natl Acad Sci U S A 2007;104(February (8)):2837–42.
43] Bequet-Romero M, Ayala M, Acevedo BE, Rodriguez EG, Ocejo OL, Torrens I,et al. Prophylactic naked DNA vaccination with the human vascular endothelial
n study of a VEGF-based therapeutic cancer vaccine in non human
2007;10(1):23–34.44] Kamstock D, Elmslie R, Thamm D, Dow S. Evaluation of a xenogeneic VEGF
vaccine in dogs with soft tissue sarcoma. Cancer Immunol Immunother2007;56(August (8)):1299–309.