angiogenesis inhibition in neuroblastoma - diva
TRANSCRIPT

Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Medicine 1254
Application of a New Logic to OldDrugs: Angiogenesis Inhibition in
Neuroblastoma
BY
ÅSA SVENSSON
ACTA UNIVERSITATIS UPSALIENSISUPPSALA 2003

Contents
Introduction.....................................................................................................1 A general introduction of childhood cancer and their treatment................1 Neuroblastoma is a common tumor in early childhood .............................2 Angiogenesis is a prerequisite for tumor growth .......................................2 Angiogenesis in neuroblastoma .................................................................5 The new logic of cancer treatment - angiogenesis inhibition.....................6 Old drugs in angiogenesis inhibition..........................................................8 The mouse as an in vivo model of neuroblastoma....................................10
Aims of the investigation..............................................................................12
Materials and methods ..................................................................................13 Test substances.........................................................................................13 The mouse model of tumor growth and angiogenesis inhibition .............13 Avian model of angiogenesis in vivo .......................................................15 In vitro model of angiogenesis .................................................................15 Monitoring angiogenesis stimulators .......................................................15 Vascular counts ........................................................................................16 Immunohistochemistry.............................................................................16 Stereological quantifications ....................................................................18 Statistical methods ...................................................................................18
Results and discussion ..................................................................................20 Confirmation of angiogenesis inhibition induced by TNP-470 and SU5416 on neuroblastoma xenografts .....................................................20
Angiogenesis can be reduced without significant reduction of tumor growth (paper I) ...................................................................................21 Importance of VEGF-A in the progression of experimental neuroblastoma (paper II) .....................................................................22
Discovery of a new use for old drugs.......................................................22 Combination treatments of neuroblastoma (paper III) .............................24
Discovery of digoxin as an angiogenesis inhibitor (paper IV) ............26
Conclusions...................................................................................................27
General discussion and future perspectives ..................................................28

Sammanfattning på svenska..........................................................................30
Acknowledgements.......................................................................................31
References.....................................................................................................33
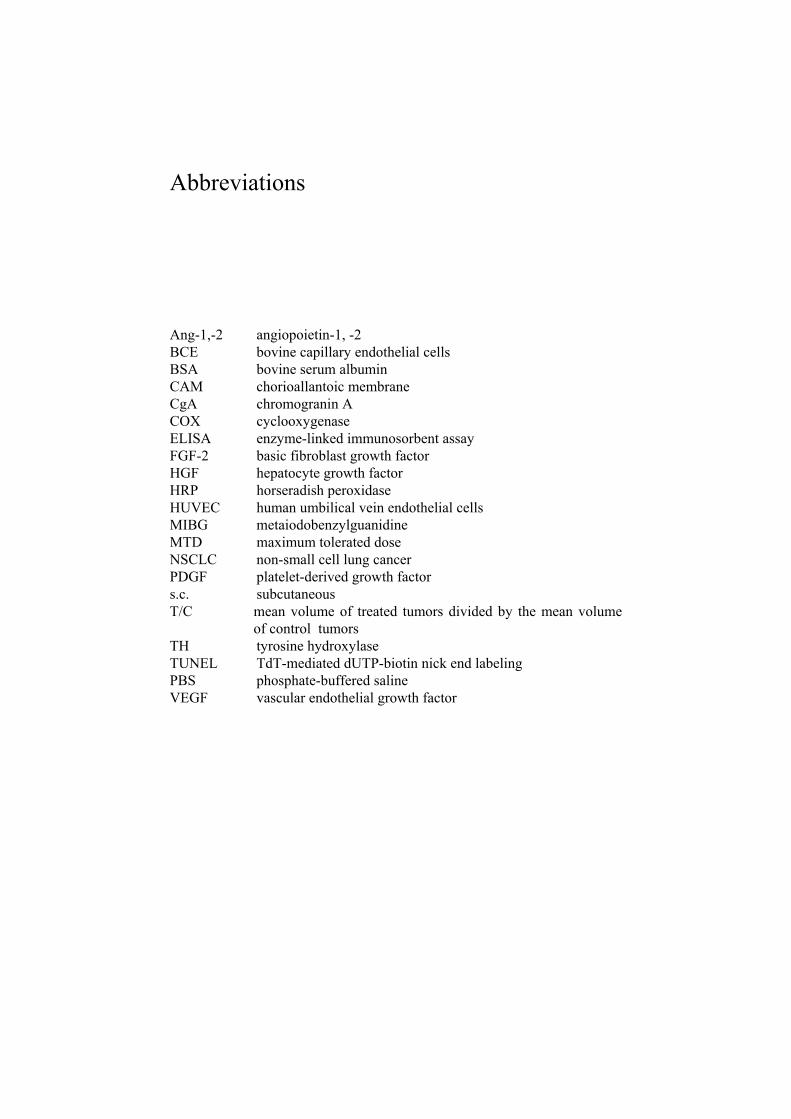
Abbreviations
Ang-1,-2 angiopoietin-1, -2 BCE bovine capillary endothelial cells BSA bovine serum albumin CAM chorioallantoic membrane CgA chromogranin A COX cyclooxygenase ELISA enzyme-linked immunosorbent assay FGF-2 basic fibroblast growth factor HGF hepatocyte growth factor HRP horseradish peroxidase HUVEC human umbilical vein endothelial cells MIBG metaiodobenzylguanidine MTD maximum tolerated dose NSCLC non-small cell lung cancer PDGF platelet-derived growth factor s.c. subcutaneous T/C mean volume of treated tumors divided by the mean volume
of control tumors TH tyrosine hydroxylase TUNEL TdT-mediated dUTP-biotin nick end labeling PBS phosphate-buffered saline VEGF vascular endothelial growth factor

List of publications
This thesis is based on the following articles, which are referred to in the text by their Roman numerals:
I Svensson Å, Bäckman U, Christofferson R; Angiogenesis can be reduced without significant reduction of tumor growth, submitted.
II Bäckman U, Svensson Å, Christofferson R; Importance of Vascular Endothelial Growth Factor A in the Progression of Experimental Neuroblastoma, Angiogenesis, in press.
III Svensson Å, Bäckman U, Jonsson E, Larsson R, and Christofferson R; CHS 828 Inhibits Neuroblastoma Growth in Mice Alone and in Combination with Antiangiogenic Drugs, Pediatric Research, 51: 607-11, 2002.
IV Svensson Å, Matsamoto T, Bäckman U, Christofferson R; Digoxin inhibits angiogenesis and tumor growth in mice, submitted.
The published papers are reprinted with permission of the copyright holders.

1
Introduction
A general introduction of childhood cancer and their treatment “It isn’t an experience anyone would opt for, it may seem bleak at the beginning, and you have to go through a lot. But you can live through it, and your kids can turn out great” Sandra, mother of leukemia survivor Erin, North America
Childhood cancers differ from adult cancers in several respects. Compared to that in children, medulloblastoma in adults differs histologically, in the location of the tumor and in the growth rate (Sarkar et al. 2002). Drug resistance profiles in adult acute lymphoblastic leukemia (ALL) differ from those in childhood ALL (Styczynski et al. 2000). Furthermore, the long expected life span and the growth of a childhood cancer survivor increase the risk of secondary malignancies, early onset cardiac failure, infertility, and growth impairment caused by chemotherapy. The annual incidence of childhood malignancies in Sweden is approximately 15 cases per 100.000 children as recorded by the Nordic Society of Pediatric Haematology and Onclology (NOPHO, Gustafsson et al. 1999). Despite its relatively low incidence, malignancy is one of the most common causes of death in pediatric patients (Mott et al. 1995) Cancer treatment in general, mostly involves chemotherapy, surgical excision of the tumor, and radiotherapy. Chemotherapeutic drugs are usually given at the maximum tolerated dose (MTD). Since the therapeutic index is low, a slight decrease in dose can impair the efficacy and increase the risk of recurrence. The treatment entails a risk of life threatening toxicity during its administration, often demanding aggressive supportive care and bone marrow transplantation. Both acute toxicity and late adverse effects are of great concern. The acute toxicity involves the hematological, gastrointestinal, pulmonary, cardiac, renal, bladder, and the nervous system, as well as causing alopecia. In addition to the hair loss, these effects are manifested as nausea, pain, diarrhea, vomiting, and fever. Late effects may appear years after completion of therapy, in the form of infertility, impairment of growth and intellectual development, organ-specific

2
toxicities and secondary malignancies (da Cunha et al. 1984, Samra et al. 1985, Hawkins et al. 1987, Kingston et al. 1987, Glauser et al., 1991, Steinherz et al. 1991). The late effects depend on the chemotherapy given, the dose, and the age of the child during therapy.
Neuroblastoma is a common tumor in early childhood
“Each school day enough children are diagnosed with cancer to empty two classrooms” National Childhood Cancer Fund, North America
Neuroblastoma is the most common extracranial solid malignant tumor in infants (Brodeur and Castleberry 1997) and accounts for approximately 6% of all childhood cancers (Gustafsson et al. 1999). It originates from cells of the sympathetic nervous system. In Sweden, approximately 10-15 children under the age of 15 years are diagnosed with this disease every year (Gustafsson et al. 1999). The prognosis varies with age at diagnosis, tumor stage, and certain biological markers. The overall 5-year survival in Europe is about 45% (Cotterill et al. 2000). In patients above one year of age with an advanced tumor stage and dismal biological markers, the outcome remains poor despite aggressive, multimodality therapy, and hence there is a clinical need for new treatment strategies (Berthold and Hero 2000, Castel et al. 2001, Ikeda et al. 2002).
Angiogenesis is a prerequisite for tumor growth
“When we try to pick anything out by itself, we find it hitched to everything else in the universe” John Muir
Three decades ago, Judah Folkman presented the hypothesis that tumors require angiogenesis i. e., the formation of new blood vessels from existing vasculature, to grow and metastasize (Folkman 1971, 1972). Angiogenesis occurs physiologically in adults during the ovarian cycle, and in wound healing, but is also necessary for growth and development in the embryo and the child. Under pathological conditions, angiogenesis is a prerequisite for tumor growth beyond a cubic millimeter (Hanahan and Folkman 1996). The physiological and pathological angiogenesis involve many similar processes. The difference between the two forms is often ultrastructural: tumor vessels often lack

3
pericytes, are dilated and leaky, and have an incomplete basement membrane, and the vessel walls may consist of both tumor cells and endothelial cells (Benjamin et al. 1999, Carmeliet et al. 1996, Chang et al. 2000). The tumor vessels may develop from pre-existing vessels by sprouting or intussusception, and also by engagement of circulating endothelial cells from the bone marrow (Fig.1). The molecules involved in tumor angiogenesis are not fully known and may differ to some extent from those in physiological angiogenesis, examples of such angiogenesis stimulators are listed in table 1.
Fig. 1. Examples of blood vessel formation. (a) sprouting of endothelial cells toward an external stimuli, for example VEGF produced by tumor cells. (b) intussusception, i. e. , the interstitial pressure forces vessel walls into proximity and the vessel divides lengthwise. (c) vasculogenesis by endothelial cell precursors. (d) surrounding tumor cells coopting an existing vessel. (e) tumor cells forming canals, mimicking blood vessels. Illustration R. Christofferson.
a
d
c
b
e

4
When the tumor grows and the need for metabolic exchange increases, the tumor cells start producing angiogenesis stimulators in response to stress, such as hypoxia and nutrient depletion. The balance of angiogenesis stimulators and inhibitors is shifted and angiogenesis increases (Carmeliet et al. 1998, Maxwell et al. 1997, Ryan et al. 1998, Shweiki et al. 1995, Hanahan and Folkman 1996). The existing vessels dilate and become leaky in a response to angiogenic stimuli such as vascular endothelial growth factor (VEGF, Kevil et al. 1998, Kohn et. al 1992). The basement membrane is degraded to allow endothelial cell invasion into the extra cellular matrix in response to metalloproteinases (Moses 1997). The endothelial cells proliferate and migrate toward the tumor when stimulated by VEGF, and basic fibroblast growth factor (FGF-2, Connolly et al. 1989, Dimmeler et al. 2000, Gospodarowicz et al. 1987, Terranova et al. 1985). In a differentiating region behind the migrating and proliferating cells, the cells cease to proliferate and they undergo morphological changes and adhere to each other to form the lumen of a new capillary tube. To some extent, circulating bone marrow -derived endothelial cell precursors are recruited to form the endothelium of the new capillary (Lyden et al. 2001, Peichev et al. 2000). The scenario outlined above refers to tumor angiogenesis by sprouting. As proposed recently, in the absence of VEGF, tumor cells around a pre-existing vessel may stimulate endothelial cells in the vessel to express Ang-2. The expression of Ang-2 is followed by vessel degradation. Thus the tumor cells become hypoxic and start producing angiogenesis stimulators such as VEGF, and new vessels are formed by sprouting processes similar to those described above (Holash et al.1999a, Holash et al.1999b,Yancopoulos et. al. 2000). A capillary can also be split lengthwise into two vessels by the formation of endothelial pillars, which merge by mechanical forces from the interstitial tissue. This event is called intussusception (Patan et al. 1996).
5
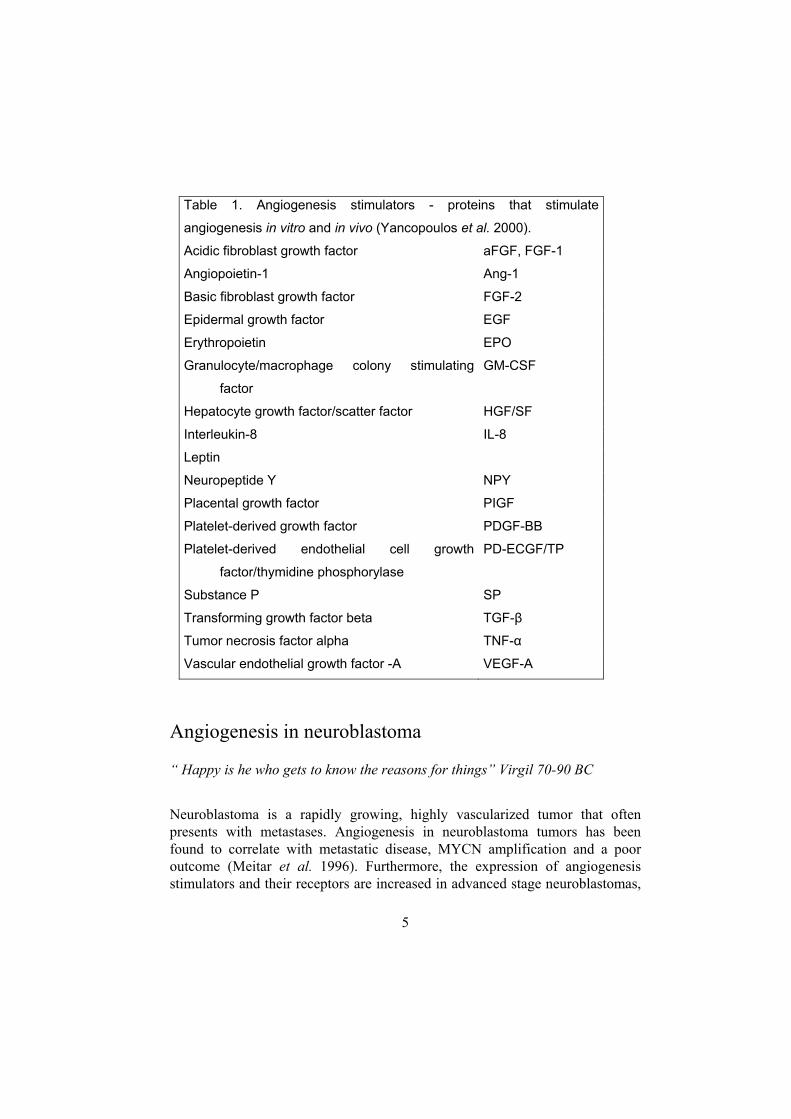
Table 1. Angiogenesis stimulators - proteins that stimulate
angiogenesis in vitro and in vivo (Yancopoulos et al. 2000).
Acidic fibroblast growth factor aFGF, FGF-1
Angiopoietin-1 Ang-1
Basic fibroblast growth factor FGF-2
Epidermal growth factor EGF
Erythropoietin EPO
Granulocyte/macrophage colony stimulating
factor
GM-CSF
Hepatocyte growth factor/scatter factor HGF/SF
Interleukin-8 IL-8
Leptin
Neuropeptide Y NPY
Placental growth factor PIGF
Platelet-derived growth factor PDGF-BB
Platelet-derived endothelial cell growth
factor/thymidine phosphorylase
PD-ECGF/TP
Substance P SP
Transforming growth factor beta TGF-β
Tumor necrosis factor alpha TNF-α
Vascular endothelial growth factor -A VEGF-A
Angiogenesis in neuroblastoma
“ Happy is he who gets to know the reasons for things” Virgil 70-90 BC
Neuroblastoma is a rapidly growing, highly vascularized tumor that often presents with metastases. Angiogenesis in neuroblastoma tumors has been found to correlate with metastatic disease, MYCN amplification and a poor outcome (Meitar et al. 1996). Furthermore, the expression of angiogenesis stimulators and their receptors are increased in advanced stage neuroblastomas,

6
for example VEGF-A, whose expression is up-regulated by hypoxia and whose mRNA becomes more stable (Eggert et al. 2000, Fakhari et al. 2002, Fukuzawa et al. 2002, Meister et al. 1999, Rössler et al. 1999, Komuro et al. 2001). TNP-470 and inhibition of VEGF signaling have shown antitumor activity in animal models of neuroblastoma (Wassberg et al. 1997, Klement et al. 2000, Nagabuchi et al. 1997, Yoshizawa et al. 2000, Davidoff et al. 2001). Together, all these observations suggest that antiangiogenic therapy may be useful in treating children with neuroblastoma.
The new logic of cancer treatment - angiogenesis inhibition “If an angel could grant me one wish, I would wish for a cure of cancer” Chiana, leukemia patient, North America
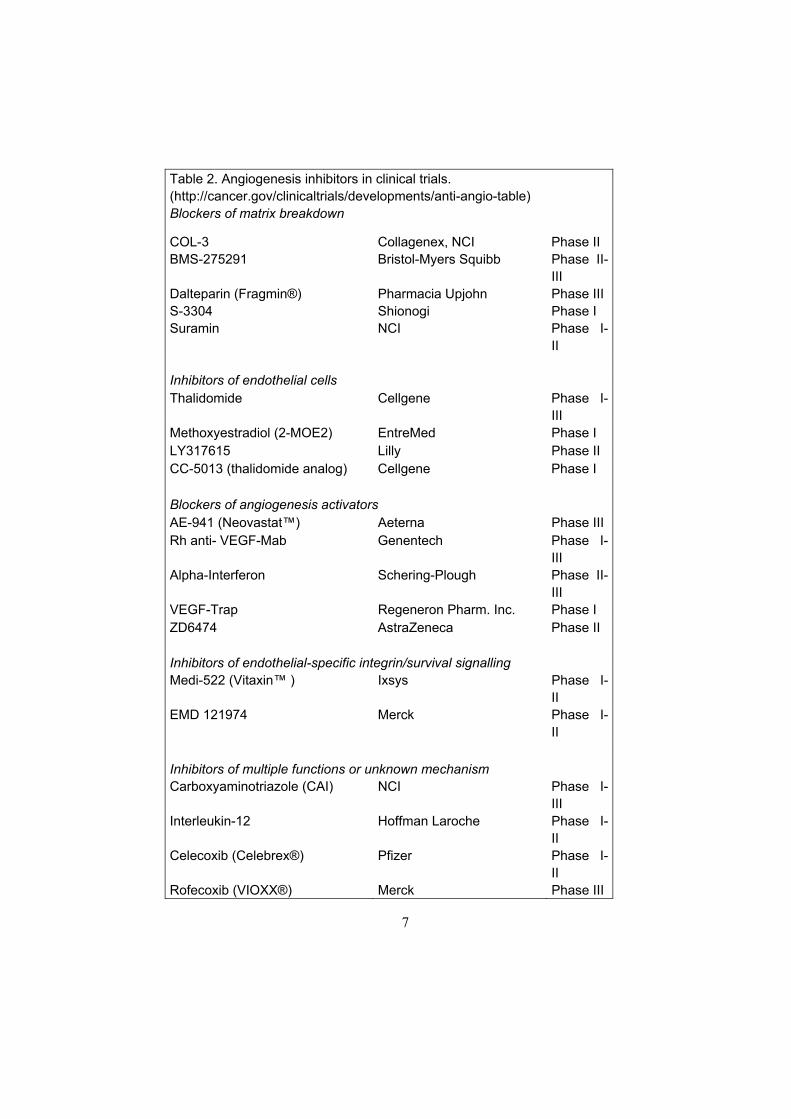
Tumor angiogenesis can be targeted by different approaches: inhibition of blood vessel growth by specific angiogenesis inhibitors; combining angiogenesis inhibitors with standard chemotherapeutic drugs; or metronomic scheduling of chemotherapeutic drugs. A number of substances with an angiogenesis inhibitory capacity have been identified during the last 10 years. Today, many of these substances have entered clinical trials against various cancer forms (Table 2). Tumor angiogenesis can be inhibited at many levels in the angiogenesis process by different inhibitors of the metalloproteinases, of angiogenic growth factors, of endothelial cells, and of the tumor vasculature (Deplanque and Harris 2000). Two angiogenesis inhibitors in clinical trials are TNP-470, which inhibits endothelial cell proliferation (Kusaka et al. 1994), and SU5416, which blocks downstream signaling from VEGF-receptor-2, an important pathway in angiogenesis (Fong et al. 1999). The side effects of angiogenesis inhibition are unclear. However, both TNP-470 and SU 5416 have been reported to delay wound healing in animals (Klein et al. 1999, Gelaw et al 2001). In pediatric patients, in whom angiogenesis is a prerequisite for organ growth and development, the physiological angiogenesis may also be affected.
7
Table 2. Angiogenesis inhibitors in clinical trials. (http://cancer.gov/clinicaltrials/developments/anti-angio-table) Blockers of matrix breakdown
COL-3 Collagenex, NCI Phase II BMS-275291 Bristol-Myers Squibb Phase II-
III Dalteparin (Fragmin®) Pharmacia Upjohn Phase III S-3304 Shionogi Phase I Suramin NCI Phase I-
II Inhibitors of endothelial cells Thalidomide Cellgene Phase I-
III Methoxyestradiol (2-MOE2) EntreMed Phase I LY317615 Lilly Phase II CC-5013 (thalidomide analog) Cellgene Phase I Blockers of angiogenesis activators AE-941 (Neovastat™) Aeterna Phase III Rh anti- VEGF-Mab Genentech Phase I-
III Alpha-Interferon Schering-Plough Phase II-
III VEGF-Trap Regeneron Pharm. Inc. Phase I ZD6474 AstraZeneca Phase II Inhibitors of endothelial-specific integrin/survival signalling Medi-522 (Vitaxin™ ) Ixsys Phase I-
II EMD 121974 Merck Phase I-
II Inhibitors of multiple functions or unknown mechanism Carboxyaminotriazole (CAI) NCI Phase I-
III Interleukin-12 Hoffman Laroche Phase I-
II Celecoxib (Celebrex®) Pfizer Phase I-
II Rofecoxib (VIOXX®) Merck Phase III

8
Old drugs in angiogenesis inhibition “ Why are things as they are and not otherwise?” Johannes Kepler 1571-1630 In this thesis the term “old drugs” refers to drugs that primarily were developed for another purpose than angiogenesis inhibition. One new way of using these old drugs, such as chemotherapeutic agents, would be to combine them with specific angiogenesis inhibitors. Pre-clinical tests of combinations of angiogenesis inhibitors and cytotoxic drugs have shown that angiogenesis inhibitors can act as effective adjuvants in improving the efficacy of the chemotherapeutic drugs. The angiogenesis inhibitory combination of TNP-470 and minocycline can potentiate the effects of DNA- targeting treatments such as cyclophosphamide, adriamycin, cisplatin, 5-fluorouracil, and radiotherapy. This combination increases growth delay in both in vitro and in vivo models, reduces Lewis lung carcinoma metastases in mice, and results in a 40-50% cure rate in the animals (Teicher et.al. 1995, Teicher et al. 1996, Kakeji and Teicher 1997). Furthermore, pre-treatment with angiogenesis inhibitors increases the uptake of the small lipophilic DNA binding dye Hoechst 33342, and decreases the tumor hypoxic fraction of tumor xenografts (Teicher et al. 1995, Teicher et al. 1996, Kakeji and Teicher 1997). The combination of the angiogenesis inhibitor TNP-470 and cyclophosphamide effectively eradicates drug resistant Lewis lung carcinoma in mice, and a VEGFR-2 antibody together with vinblastine causes regression of neuroblastomas without toxicity (Browder et al. 2000, Klement et al. 2000). Clinical phase III combination trials are taking place in the United States for AG 3340 and carboplatin/paclitaxel or gemcitabine/cisplatin; and there are phase I-II combination studies of squalamine and chemotherapeutic drugs for treatment of the most common form of lung cancer, non-small cell lung carcinoma (NSCLC). Another clinical trial of an angiogenesis inhibitor in combination with a chemotherapeutic drug is that of thalidomide plus carboplatin for treatment of glioblastoma multiforme (Deplanque and Harris 2000). When the dosing schedule is changed, some old chemotherapeutic drugs have recently been found able to act as angiogenesis inhibitors (metronomic scheduling; Bocci et al. 2002). In fact, some of the initial, rapid effects of chemotherapeutic drugs may be due to angiogenesis inhibition. Metronomic scheduling of chemotherapeutic drugs could be beneficial in reducing the toxicity of the drug and might even overcome multi-drug resistance (Browder et al. 2000, Klement et al. 2000). By combining the use of specific angiogenesis inhibitors with metronomic scheduling of chemotherapeutic agents, the duration

9
of the angiogenesis inhibition may be lengthened (Kim et al. 2002a, Soffer et al. 2001) In addition some old drugs can have a potential as an angiogenesis inhibitor themselves. A number of old drugs have re-attracted scientific attention as potential angiogenesis inhibitors. Thalidomide, a sedative and anti-emetic drug, is a potent teratogen which caused amelia or dysmelia in infants whose mothers had ingested the drug during early pregnancy. It has now regained interest as an angiogenesis inhibitor (D’Amato et al. 1994). Clinical trials of thalidomide as treatment mainly of multiple myeloma and Kaposi’s sarcoma are currently being conducted. Interferon alpha is another drug of interest; it has been used in the treatment of hepatitis and leukemia and currently is in phase III trials for cancer, partly because of its properties as an angiogenesis inhibitor. A third group of drugs of interest in cancer is the group of anti-inflammatory cyclooxogenase-2 (COX-2) inhibitors, such as celecoxib and rofecoxib. They inhibit angiogenesis by a direct effect on endothelial cells (Jones et al. 1999). COX inhibitors are able to inhibit tumor growth and angiogenesis in xenograft models and are currently being tested in phase III trials (Mohammed et al. 2002, Sawaoka et al. 1999). A number of chemotherapeutic agents have also been suggested as possible angiogenesis inhibitors. To determine whether the effect of the drug is truly antiangiogenic or, rather, cytotoxic is difficult, since there is no clear definition of an angiogenesis inhibitor. A few criteria were recently proposed (Table 3).
Table 3. Criteria for a drug to be considered as an angiogenesis inhibitor (after Miller et al. 2001)
• Endothelial cell specificity • Interference with endothelial cell
function • Interference with angiogenesis
mechanisms • Inhibits angiogenesis in vivo

10
The mouse as an in vivo model of neuroblastoma “It has recently been discovered that research causes cancer in rats” Unknown In vivo xenograft models are of great importance in the development and testing of new therapies for neuroblastoma, since the low incidence of neuroblastoma impedes clinical trials of multiple new drugs. Some criteria for an ideal murine disease model have been proposed (Table 4, modified from Beltinger and Debatin 2001, Kerbel 1999, Rosenberg et al. 1999, Killion et al.1999).
Table 4. Proposed criteria for an ideal murine disease model • The genetics of the model should represent the genetics of the
disease • The histology of the model should resemble the clinical histology • The development of the tumors should occur at the same time in
different animals, at a high frequency, and in regions easily accessible for diagnosis and therapy.
• The model should be orthotopic to the human tumor. • The metastasis pattern should resemble the clinical situation. • The therapeutic response in the model should be similar to that
of clinically well-documented treatments. To date, however, no neuroblastoma model has met all these criteria (Fig. 2). The neuroblastoma model used in the present work is based on human SH-SY5Y neuroblastoma cells xenografted to NMRI nunu mice. The SH-SY5Y cell line derives from a lethal, primary thoracic neuroblastoma tumor (Biedler et al. 1973). SH-SY5Y cells, like most neuroblastoma tumors, do not exhibit amplification of the proto-oncogene MYCN and are thus not suitable for studies of MYCN-targeted treatments. However, MYCN amplification is associated with a poor prognosis, more aggressive tumor growth, and increased angiogenesis in patients (Ikeda et. al, 2002, Ribatti et. al, 2002). Subcutaneously xenografted SH-SY5Y tumors in nude rodents are histologically human neuroblastomas, which grow aggressively, are angiogenic, and respond well to treatment with angiogenesis inhibitors (Wassberg et al. 1997, 1999). The tumors occur at approximately the same time and at a high frequency and reproducibility.

These subcutaneous tumors do not metastasize in nude mice or rats. There are a few models with a metastasizing pattern resembling the clinical disease (Khanna et al. 2002). However, to obtain a metastasizing model the tumor cells have to be injected intravenously (Turner et al.1990, Bogenmann 1996) or orthotopically, in SCID mice. It has been argued that metastasizing models resemble the late stage of the disease (Beltinger and Debatin 2001). The sites of the metastases also tend to differ from those in the clinical disease. Another problem with a metastasizing model is the difficulty in measuring the total tumor burden, which can easily be assessed with a caliper in the SH-SY5Y subcutaneous (s.c) non-metastasizing model. We conclude that our animal model, despite its discrepancies from clinical neuroblastomas, is reproducible and relevant.
MY1p s
Neuroblas nests sepstroma wiproliferatio
Tpao
Genetics CN
36 deletion
11
Histology tic cells in groups or
arated incomplete th poor schwannian n
Lybolivm
Primary tumors horacic araspinal bdominal tumorsr adrenal tumors
Metastases mphatic, nemarrow, er and skin etastases
Fig. 2 Example of proposed criteria for an ”ideal ”neuroblastoma mousemodel
12
Aims of the investigation
“A good traveler has no fixed plans and is not intent on arriving” LaoTzu 570-590 B. C.
The general aim of this study was to investigate the potential of “old drugs” as angiogenesis inhibitors in neuroblastoma. The specific aims were:
• to investigate the efficacy of angiogenesis inhibition induced by the two well-characterized angiogenesis inhibitors SU5416 and TNP-470 in the SH-SY5Y mouse model of neuroblastoma;
• to study the efficacies of the chemotherapeutic agent CHS 828 and digoxin in inhibiting tumor growth and angiogenesis in the mouse model of neuroblastoma, when given alone;
• to test the efficacy of treatment with TNP-470 and SU5416 in combination with the chemotherapeutic agent CHS 828 in the mouse model.

13
Materials and methods
“The thing about good ideas is that they soon degenerate to hard work” Unknown
Test substances The drugs investigated were the angiogenesis inhibitors TNP-470 (a kind gift from Takeda Chemical Industries Ltd., Osaka, Japan), SU5416 (a kind gift from Sugen Inc., South San Francisco, CA), the chemotherapeutic agent CHS 828 (N-(6-chlorophenoxyhexyl)-N’-cyano-N’’-4-pyridylguanidine, (a kind gift from Leo Pharmaceutical Products, Ballerup, Denmark) and the cardiac glycoside digoxin (Digoxin NM Pharma, NM Pharma AB, Stockholm, Sweden). These drugs were chosen to represent different kinds of drugs with different mechanisms of action. TNP-470 inhibits endothelial cell proliferation (Kusaka et al. 1994), and SU5416 blocks downstream signaling from VEGF receptor-2 (Fong et al. 1999). CHS 828 belongs to a new class of chemotherapeutic drugs, the pyridyl cyanoguanidines. It has potent anti-tumor activity both in vitro and in vivo (Hjarnaa et. al 1999). Cardiac glycosides such as digoxin and the structurally related compound digitoxin have been reported to inhibit growth of multiple tumor cell types in vitro and seem to have a favorable effect in breast carcinoma (Haux 1999, Stenkvist 1999, Johansson et al. 2001). Their mechanism of tumor inhibition has not been fully investigated, however.
The mouse model of tumor growth and angiogenesis
inhibition
A reproducible experimental neuroblastoma model for studies of growth and angiogenesis inhibition was first presented by Wassberg et al. 1997, using nude rats. In order to increase the number of animals in each group and to facilitate molecular analyses, we later changed to nude mice. Tumor cells are injected s.c.

14
in NMRI nunu mice, and when the tumor has grown to a size of approximately 0.3 ml, the animals are randomized into different treatment groups and control groups, and treatment begins. The tumor is allowed to grow to a maximum volume corresponding to approximately 10% of the body weight of the animals. The animals are then euthanized, and blood is drawn from the right ventricle. Thereafter the whole body is perfused with 4% paraformaldehyde in phosphate buffer through a cannula inserted into the aorta. The tumors are excised, placed in 4% paraformaldehyde, dehydrated and embedded in paraffin. This model makes it possible to study different kinds of tumors in vivo, permitting monitoring of real-time treatment effects, toxicity and systemic effects of a substance. It also allows studies of treatments with combination of angiogenesis inhibitors and chemotherapeutic drugs in vivo. There are no metastases in our model, which in this context is an advantage, since it is easy to measure the whole tumor burden without surgery or imaging techniques. We have mainly used SH-SY5Y neuroblastoma cells, but we have also used murine Lewis lung carcinoma cells and human colonic carcinoma LS 174T cells to produce the in vivo tumors. Recently, however, we have established models for MYCN amplified neuroblastoma using the human cell line IMR 32 and Kelly (Fig. 3), with a tumor formation frequency of 91% and 62% respectively. The NMRI nunu strain was chosen since xenografted tumors do not form metastases and the strain has a high success rate in the formation of tumors. The mice are T-cell -deficient but can develop immunity with maturation. We choose to use relatively young animals to prevent this from happening and to be able to study
the effects of the treatment on animal growth.
a ba ba b
Fig. 3 Staining of blood vessels by BS-1, in xenografts from two MYCN amplified neuroblastomas. (a) Kelly and (b) IMR-32. Magnification 200x and bar 50 µm

15
Avian model of angiogenesis in vivo By opening a window in the shell of fertilized chicken eggs, the vascularized chorioallantoic membrane (CAM) is exposed (Friedlander et al. 1995). A filter paper, soaked in the drug of interest, is placed on the membrane and incubated for three days. The membrane is then cut out, and the numbers of vessels or vessel branching points are quantified under a dissection microscope. The CAM assay was used for testing the angiogenesis inhibitory effect of digoxin (paper IV).
In vitro model of angiogenesis Endothelial growth inhibition was studied in growth-stimulated bovine capillary endothelial cells (BCE) or human umbilical vein endothelial cells (HUVEC), plated on flat-bottomed 96-well microtiter plates containing the drugs of interest. The survival index (SI) of the cells in each well was measured after 72 h. The number of viable cells is directly proportional to the fluorescence after hydrolysis of fluorescein diacetate (Larsson et al., 1992).
Monitoring angiogenesis stimulators Blood samples from treated animals were analyzed for VEGF, FGF-2 and hepatocyte growth factor. These samples were compared with samples from tumor-free and tumor-bearing control animals. Human VEGF-A165, FGF-2 and HGF concentrations in plasma were measured with enzyme-linked immunosorbent assays (ELISA) according to the manufacturer’s instructions (Quantikine®; R&D Systems, Minneapolis, MN). To ensure detection of intact VEGF-A, western blots from sera were performed as described by Amaral et al. (2001), using the same antibody as the ELISA kit. The in vivo expression of VEGF-A in neuroblastoma tumors was also measured by in-situ hybridization on tissue sections. The template used for hybridization was a 740-bp BamHI, HindIII VEGFA165 cDNA fragment (kindly provided by Dr.Arne Östman, Ludwig Institute for Cancer Research, Uppsala, Sweden). β-actin was used as a positive control and hybridization with the corresponding sense probe was used as a negative control. VEGFA165 protein was detected by the ABC/HRP method (K0355, Dako A/S, Glostrup, Denmark). The primary antibody (goat anti-human VEGFA165, R&D AB-293-NA) was applied. As a positive control, human fetal kidney was used, and omission of the primary antibody served as a negative control.
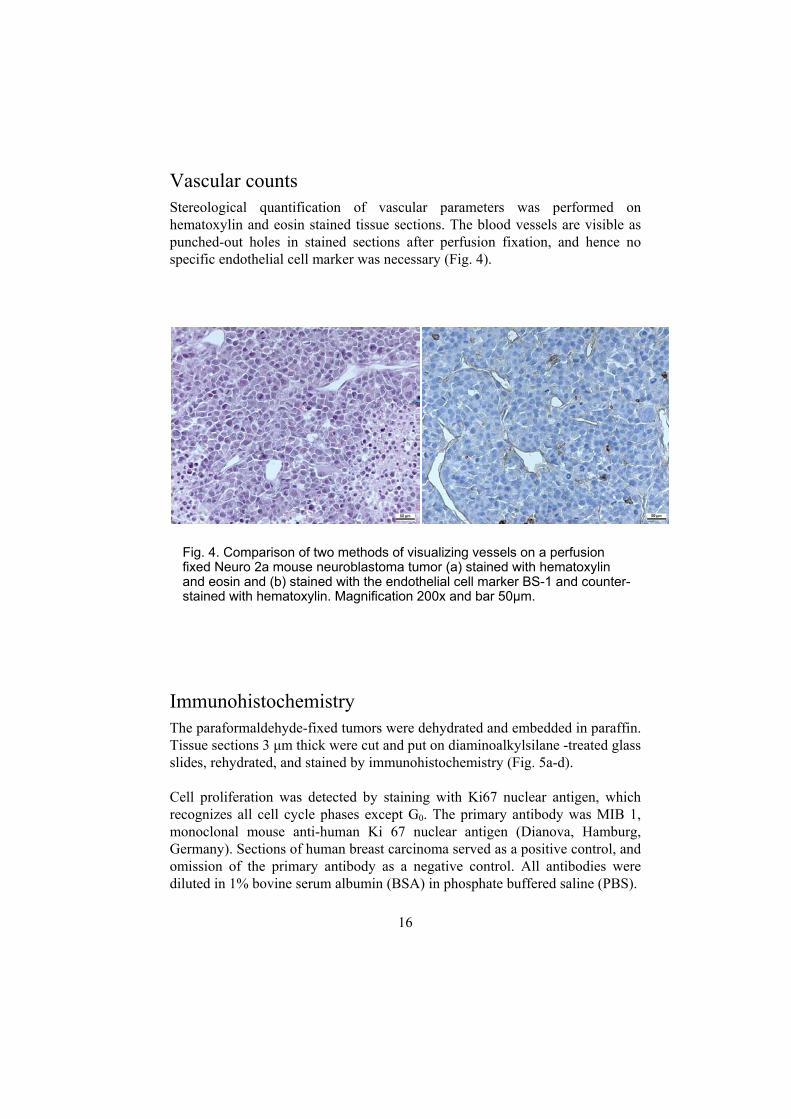
Vascular counts Stereological quantification of vascular parameters was performed on hematoxylin and eosin stained tissue sections. The blood vessels are visible as punched-out holes in stained sections after perfusion fixation, and hence no specific endothelial cell marker was necessary (Fig. 4).
ImmunohThe paraforTissue sectislides, rehyd Cell proliferecognizes amonoclonalGermany). Somission ofdiluted in 1%
Fig. 4. Cofixed Neuand eosinstained w
a
16
istochemistry maldehyde-fixed tumors were dons 3 µm thick were cut and purated, and stained by immunoh
ration was detected by staininll cell cycle phases except G0
mouse anti-human Ki 67 nuections of human breast carcin the primary antibody as a ne
bovine serum albumin (BSA)
mparison of two methods of visro 2a mouse neuroblastoma tum and (b) stained with the endothith hematoxylin. Magnification 2
b
ehydrated and embedded in paraffin. t on diaminoalkylsilane -treated glass istochemistry (Fig. 5a-d).
g with Ki67 nuclear antigen, which . The primary antibody was MIB 1, clear antigen (Dianova, Hamburg,
oma served as a positive control, and gative control. All antibodies were in phosphate buffered saline (PBS).
ualizing vessels on a perfusion or (a) stained with hematoxylin
elial cell marker BS-1 and counter-00x and bar 50µm.
17
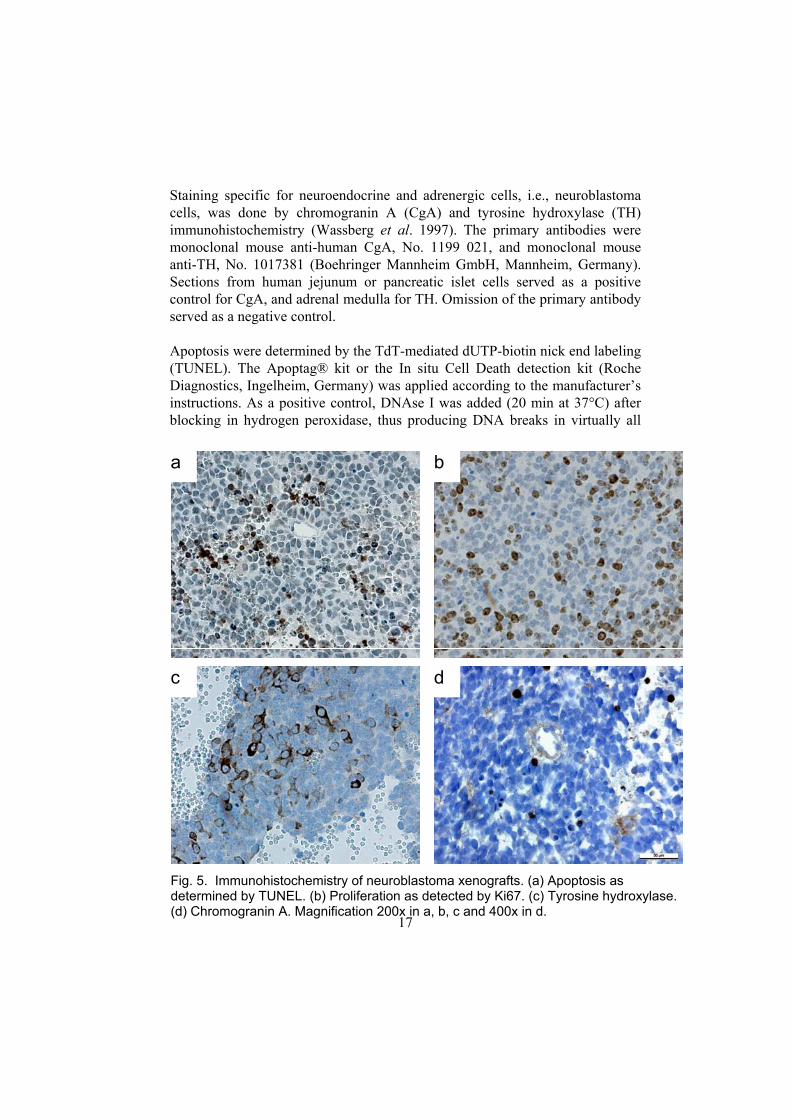
Staining specific for neuroendocrine and adrenergic cells, i.e., neuroblastoma cells, was done by chromogranin A (CgA) and tyrosine hydroxylase (TH) immunohistochemistry (Wassberg et al. 1997). The primary antibodies were monoclonal mouse anti-human CgA, No. 1199 021, and monoclonal mouse anti-TH, No. 1017381 (Boehringer Mannheim GmbH, Mannheim, Germany). Sections from human jejunum or pancreatic islet cells served as a positive control for CgA, and adrenal medulla for TH. Omission of the primary antibody served as a negative control. Apoptosis were determined by the TdT-mediated dUTP-biotin nick end labeling (TUNEL). The Apoptag® kit or the In situ Cell Death detection kit (Roche Diagnostics, Ingelheim, Germany) was applied according to the manufacturer’s instructions. As a positive control, DNAse I was added (20 min at 37°C) after blocking in hydrogen peroxidase, thus producing DNA breaks in virtually all
Fig. 5. Immunohistochemistry of neuroblastoma xenografts. (a) Apoptosis as determined by TUNEL. (b) Proliferation as detected by Ki67. (c) Tyrosine hydroxylase. (d) Chromogranin A. Magnification 200x in a, b, c and 400x in d.
a b
c d

18
cells.
Stereological quantifications
One observer quantified the tissue sections in a blinded fashion. Structures were counted at × 400 with an eyepiece grid (506800, Leica, Singapore) of 10 × 10 squares (0.25 × 0.25 mm). The grid was placed at random at the upper left corner of a section, and then systematically advanced every 1 to 3 mm (depending on the tumor size) in both directions by use of the microscope’s goniometer stage. Morphological parameters from at least 25 grids were quantified for each tumor (Table 5). Areas with hemorrhage and apoptotic or necrotic cells were designated as non-viable and were excluded from analyses of other parameters (Wassberg et al. 1997), but were used for calculation of the viable tissue fraction.
The fractions of CgA-, Ki 67-, and TUNEL-positive tumor cells were determined among 2,000 cells in each section.
Statistical methods Data were processed in Statistica 5.0 for Windows (StatSoft Inc, Tulsa, OK) or GraphPad Prism® for Windows (GraphPad Software Inc, San Diego, CA). Differences between two groups were analyzed with the Mann-Whitney U test, and differences between several groups with the Kruskal-Wallis test followed by Dunn’s post test. Differences for which P<0.05 were considered statistically significant.

19
Table 5. Definitions of vascular parameters (Wassberg et al. 1999).
Symbol Dimension Equation Definition
Lv (ves) mm-2
(frame)vc
(ves)(ves) v
x AnQΣ
x2 = L Length of vessels per
tumor volume - length
density
Vv (ves) 1
)pcg(vc
)ves( (ves) v
P xnPΣ
=V Volume of vessels
per tumor volume -
volumetric density
Sv (ves) mm-1
)lig(vc
)ves( (ves)
xLnIΣ
x2 =Sv Surface area of
vessels per tumor
volume - surface
density
đ (ves) mm
πa
x2 = d(ves)
(ves) Mean section
diameter of vessels
The eyepiece grid was used as an unbiased counting frame, a point counting
grid and a line intercept grid. Q (ves)= number of vessel profiles in one counting
frame; nvc= number of grids with viable tissue in the uppermost square of the
right; A (frame)= area of one counting frame; P (ves)= number of test points
hitting vessels (any layer or lumen) in one point counting grid; P (pcg)= number
of test points in one point counting grid; l (ves)= number of intersections with
vessels in one line intercept grid; L (lig)= length of test lines in one line intercept
grid; a(ves)= mean section area of vessels.
20
Results and discussion
“The end of all our exploring will be to arrive where we started and know the place for the first time” T. S. Eliot
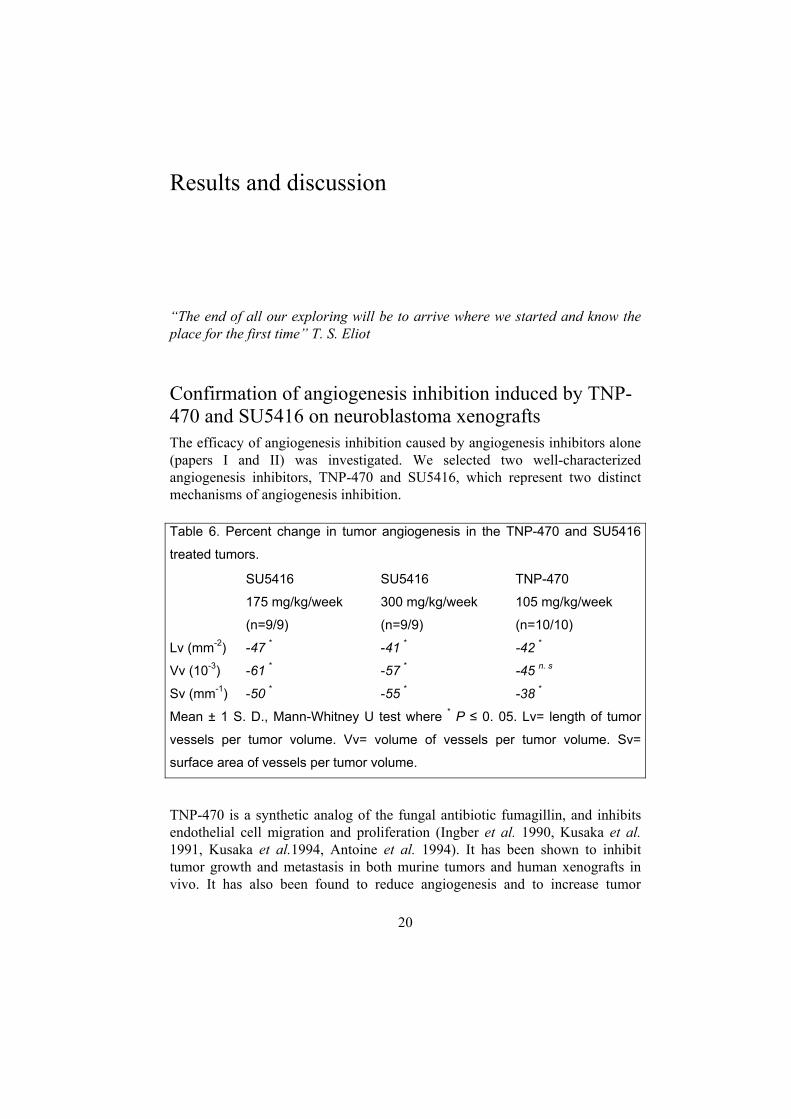
Confirmation of angiogenesis inhibition induced by TNP-470 and SU5416 on neuroblastoma xenografts The efficacy of angiogenesis inhibition caused by angiogenesis inhibitors alone (papers I and II) was investigated. We selected two well-characterized angiogenesis inhibitors, TNP-470 and SU5416, which represent two distinct mechanisms of angiogenesis inhibition.
TNP-470 is a synthetic analog of the fungal antibiotic fumagillin, and inhibits endothelial cell migration and proliferation (Ingber et al. 1990, Kusaka et al. 1991, Kusaka et al.1994, Antoine et al. 1994). It has been shown to inhibit tumor growth and metastasis in both murine tumors and human xenografts in vivo. It has also been found to reduce angiogenesis and to increase tumor
Table 6. Percent change in tumor angiogenesis in the TNP-470 and SU5416
treated tumors.
SU5416
175 mg/kg/week
(n=9/9)
SU5416
300 mg/kg/week
(n=9/9)
TNP-470
105 mg/kg/week
(n=10/10)
Lv (mm-2)
Vv (10-3)
Sv (mm-1)
-47 *
-61 *
-50 *
-41 *
-57 *
-55 *
-42 *
-45 n. s
-38 *
Mean ± 1 S. D., Mann-Whitney U test where * P ≤ 0. 05. Lv= length of tumor
vessels per tumor volume. Vv= volume of vessels per tumor volume. Sv=
surface area of vessels per tumor volume.

21
necrosis in experimental neuroblastoma by increasing differentiation and apoptosis (“agonal differentiation”;Wassberg et al.1999). SU5416 is an angiogenesis inhibitor currently in phase III clinical trials. SU5416 inhibits the vascular endothelial growth factor signaling by acting as a VEGF receptor 2 tyrosine kinase inhibitor. It is a selective ATP-competitive inhibitor of the kinase (Fong et al. 1999). Both drugs significantly inhibited the blood vessel growth (Table 6).
Angiogenesis can be reduced without significant reduction of tumor growth (paper I) TNP-470 has been used successfully in studies of angiogenesis inhibition in neuroblastoma xenografts in our rat model. During an attempt to replicate these studies in our mouse model, no significant reduction of tumor growth was found (mean tumor volume in treated animals divided by mean of control tumors, T/C, day 18 of therapy = 0.69, P =0.20). Quantification of vascular parameters surprisingly showed significant inhibition of blood vessel growth. TNP-470 reduced angiogenesis, in consistency with our results in rats (Wassberg et al. 1997), but it had no significant impact on net tumor growth. Nor did it have any significant effect on tumor necrosis, apoptosis, or proliferation. Further, the serum levels of FGF-2 and VEGF did not differ between the controls and treated animals. How can there be a substantial reduction in blood vessel density without the tumor growth being affected?
Our proposed answer is that if the angiogenesis is greater than the metabolic demand, a tumor will exhibit a rapid decrease in vessel density until equilibrium is reached (Hlatky et al. 2002). This concept may explain why the TNP-470 treated tumors showed significantly reduced vessel parameters while the tumor growth was only moderately affected. It has also been shown that VEGF can upregulate bcl-2 expression in neuroblastoma cells and protects them from apoptosis (Beierle et al. 2002). It is possible that VEGF protects the tumor cells from apoptosis when a non-VEGF -targeting treatment approach is used, in contrast to SU5416 therapy. This concept would imply that clinicians should be cautious in using microvessel density as a surrogate marker of the response to angiogenesis inhibitors in patients, as tumor regression may not be seen until after further therapy (Hlatky et al. 2002).

22
Importance of VEGF-A in the progression of experimental neuroblastoma (paper II) A variety of human tumors express VEGF and its receptors. VEGF is considered to be one of the key mediators of angiogenesis (Ferrara and Alitalo 1991, Dvorak et al. 1995, Klagsbrun and Soker 1993). Our SH-SY5Y neuroblastoma xenograft model expresses VEGF-A on both the mRNA (Fig. 6) and protein levels. The plasma concentrations of VEGF-A are elevated in animals with neuroblastoma tumors with a volume of more than 1.4 ml, and there is a correlation between VEGF-A levels in plasma and neuroblastoma tumor size. Treatment with SU5416 reduced the growth of neuroblastoma tumors by 65% without apparent systemic toxicity. SU5416 treatment also suppressed tumor angiogenesis. An increase in plasma VEGF-A levels per milliliter tumor volume was observed during therapy. The increase in VEGF indicates that the angiogenesis inhibition was successful, since up-regulation of VEGF often occurs with increasing tumor hypoxia (Rössler et al. 1999). The persistence in tumor growth despite successful inhibition of angiogenesis may be explained by host vessel cooption (Holash et al. 1999a, 1999b). It has been suggested that xenograft models of neuroblastoma are dependent on such cooption in the early stages of tumor growth, and that if VEGF is only partially inhibited the tumor may evade the effects of angiogenesis inhibition treatment (Rowe et al. 2000, Kim et al. 2002b).
Discovery of a new use for old drugs CHS 828 is a potent inhibitor of neuroblastoma growth (paper III)
Fig. 6. In situ VEGF expression in SH-SY5Y neuroblastoma xenografts treated with
SU5416. (a) Light field, (b) dark field. Magnification x 250
a b
Fig. 6. In situ VEGF expression in SH-SY5Y neuroblastoma xenografts treated with
SU5416. (a) Light field, (b) dark field. Magnification x 250
a b

23
Recently, it has been suggested that chemotherapeutic drugs can inhibit angiogenesis when given at a non-toxic dose at a frequency sufficient to keep the blood concentration of the drug fairly constant. These drugs may still be toxic to tumor cells, in addition to their inhibitory effects on angiogenesis and endothelial cell proliferation. The tumor cell toxicity is probably also an important effect when the drugs are given at high doses. If the drugs are given at low doses and at short intervals, the tumor vasculature - with its proliferating endothelial cells - may be targeted directly. Endothelial cells have a stable genome, and thus multi-drug resistance will be avoided (Browder et al. 2000). We therefore decided to test a new chemotherapeutic drug, CHS 828, to see whether it could reduce the growth of neuroblastoma tumors and angiogenesis in our neuroblastoma xenograft model when given at a low dose on a daily basis (paper III).
CHS 828 belongs to a new class of chemotherapeutic drugs, the pyridyl cyanoguanidines. It has potent anti-tumor activity both in vitro and in vivo. It shows low correlation with the activity patterns of known chemotherapeutic drugs, and with known patterns of multi-drug resistance (Hjarnaa et al. 1999). Interestingly, CHS 828 has structural similarities to the catecholamine precursor metaiodobenzylguanidine (MIBG), which is known to accumulate in neuroblastomas and is used for imaging and radiotherapy (Montaldo et al. 1991, Troncone and Rufini 1997). Furthermore, CHS 828 and MIBG seem to have some comparable potency in inhibition of mitochondrial respiration, with an increase in glycolysis and the acidification rate after treatment of tumor cells in vitro (Ekelund et al. 2000).
In our neuroblastoma model, CHS 828 at a sub-toxic dose caused a total tumor regression in 4/9 animals, with a T/C ratio of 0.18 at day 18 of treatment. Thus CHS 828 proved to be highly effective in treatment of neuroblastoma xenograft tumors. The issue of angiogenesis inhibition proved to be more difficult to assay, on account of the limited access to evaluable tumor samples because of tumor regression. However, although not significant, there was a tendency to a decrease in vascular parameters in the CHS 828 -treated tumors.
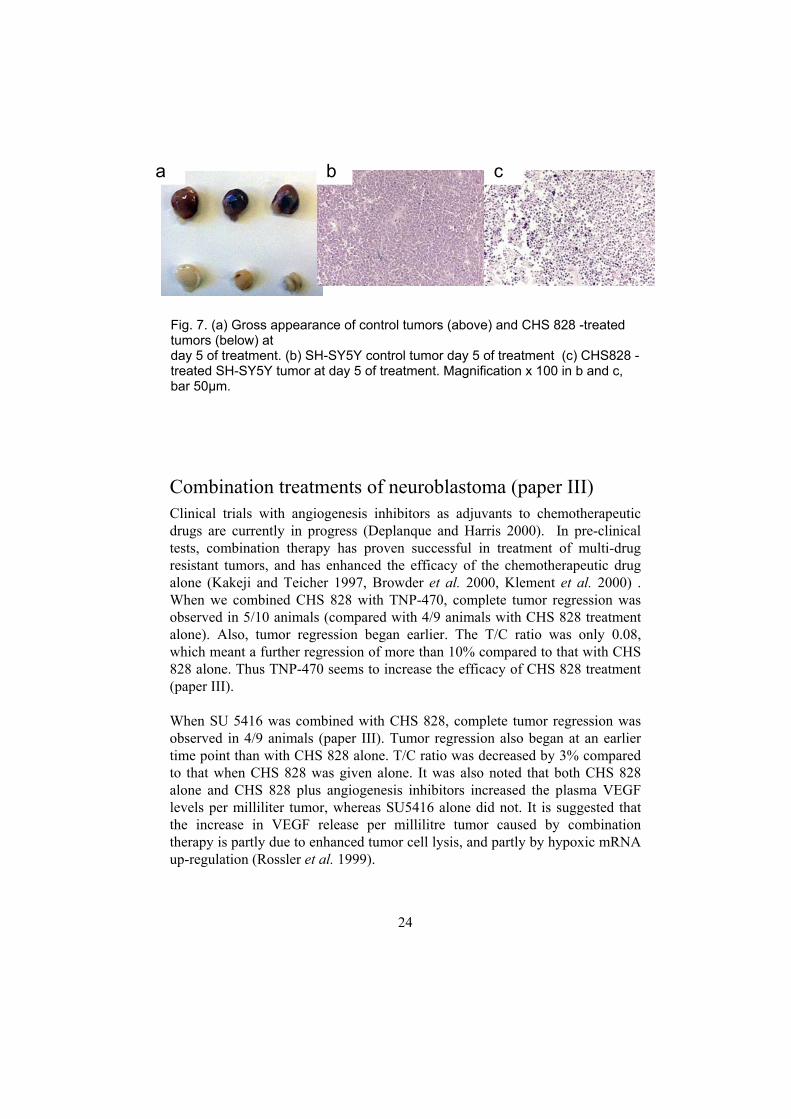
We also tested CHS 828 treatment at a dose of 20 mg/kg q.o.d. for five days only. We found that five days’ treatment caused a 29% reduction in tumor size compared to the initial value. The tumors were small and pale in appearance (Fig. 7a). Histologically a massive cell death was observed in treated tumors (Fig. 7 b,c, in preparation).
24
Combination treatments of neuroblastoma (paper III) Clinical trials with angiogenesis inhibitors as adjuvants to chemotherapeutic drugs are currently in progress (Deplanque and Harris 2000). In pre-clinical tests, combination therapy has proven successful in treatment of multi-drug resistant tumors, and has enhanced the efficacy of the chemotherapeutic drug alone (Kakeji and Teicher 1997, Browder et al. 2000, Klement et al. 2000) . When we combined CHS 828 with TNP-470, complete tumor regression was observed in 5/10 animals (compared with 4/9 animals with CHS 828 treatment alone). Also, tumor regression began earlier. The T/C ratio was only 0.08, which meant a further regression of more than 10% compared to that with CHS 828 alone. Thus TNP-470 seems to increase the efficacy of CHS 828 treatment (paper III). When SU 5416 was combined with CHS 828, complete tumor regression was observed in 4/9 animals (paper III). Tumor regression also began at an earlier time point than with CHS 828 alone. T/C ratio was decreased by 3% compared to that when CHS 828 was given alone. It was also noted that both CHS 828 alone and CHS 828 plus angiogenesis inhibitors increased the plasma VEGF levels per milliliter tumor, whereas SU5416 alone did not. It is suggested that the increase in VEGF release per millilitre tumor caused by combination therapy is partly due to enhanced tumor cell lysis, and partly by hypoxic mRNA up-regulation (Rossler et al. 1999).
Fig. 7. (a) Gross appearance of control tumors (above) and CHS 828 -treated tumors (below) at day 5 of treatment. (b) SH-SY5Y control tumor day 5 of treatment (c) CHS828 -treated SH-SY5Y tumor at day 5 of treatment. Magnification x 100 in b and c, bar 50µm.
a b c

25
The issue of why a chemotherapeutic drug is more effective when given in combination with an angiogenesis inhibitor is poorly understood. Since drug resistance is associated with tumor hypoxia , it seems reasonable to assume that an inhibitor of tumor angiogenesis will further increase tumor hypoxia and thereby reduce the efficacy of a chemotherapeutic drug. The published data , however, suggest that an angiogenesis inhibitor decreases the interstitial tumor pressure and increase the efficacy of a chemotherapeutic drug (Lee et al. 2000, Drevs 2000, Pietras et al. 2001, Jain 2001). The question of when the angiogenesis inhibitor should be included in combination therapy also remains to be investigated. Furthermore, what drugs should be combined? There are a vast number of angiogenesis inhibitors under development and an equally vast number of chemotherapeutic drugs on the market. Tumors are less oxygenized than normal tissue and endogenous angiogenesis inhibitors such as endostatin are down-regulated by hypoxia, whereas angiogenesis stimulators such as VEGF are up-regulated (Rossler et al. 1999, Wu et al. 2001). We hypothesize that a chemotherapeutic agent that is equally or more potent when the oxygen levels are decreased should be a good choice for combination studies. This led us to establish a fast and easy in vitro model for studying the toxicity of drugs during changes of oxygen levels (in preparation). We found that the neuroblastoma cell sensitivity to most standard chemotherapeutics drugs as well as to CHS 828 was unaffected by a reduced oxygen tension (Table 7).
Table 7. Percent changes in IC50 values of cytotoxic drugs when tested on SH-
SY5Y and SK-N-AS neuroblastoma cells on changing from a condition of high
oxygen (20%) to low oxygen (1%).
SH-SY5Y SK-N-AS
Vincristine 26 -70
Topotecan 68 71
Etoposide 58 91
Melphalan 78* 20
CHS 828 20 50
Digoxin 21 -7.1
SR 4233 (positive control) -49* -77*
* P<0.05 Mann-Whitney U test

26
Discovery of digoxin as an angiogenesis inhibitor (paper IV) We have screened a number of drugs with various uses for inhibition of growth stimulated bovine capillary endothelial cells. Digoxin showed potency as an angiogenesis inhibitor in this assay (Olausson et al., in preparation). We also found that digoxin could inhibit angiogenesis in the CAM assay. Digoxin inhibited neuroblastoma growth in our xenograft model, with a T/C of 0.56. The in vivo growth of colon cancer and Lewis lung carcinoma was moderately, but not significantly affected. In vitro experiments showed that human cells were much more sensitive to digoxin than murine cells, and neuroblastoma cells more sensitive than endothelial cells. Thus, digoxin is an unspecific inhibitor of angiogenesis but a specific inhibitor of neuroblastoma growth. The reason for this selectivity remains to be investigated. In conclusion, cardiac glycosides make an interesting basis for development of new anticancer drugs.

27
Conclusions
1. Our in vivo neuroblastoma SH-SY5Y/NMRI nunu xenograft model is relevant and reproducible, permitting analysis of tumor growth, tumor cell dynamics, and tumor angiogenesis.
2. Our experimental neuroblastomas express the angiogenesis stimulator VEGF on both the mRNA and protein levels. Hence it is a useful model for interference with the VEGF signalling system.
3. The angiogenesis inhibitor SU5416 acts by inhibiting VEGF signalling and after two weeks of treatment with this agent experimental neuroblastoma growth is reduced by 65% of therapy.
4. The angiogenesis inhibitor TNP-470 can significantly reduce tumor angiogenesis without significantly reducing the tumor growth rate, indicating that there is a threshold limit of angiogenesis reduction in our neuroblastoma model before tumor growth reduction is apparent.
5. The new chemotherapeutic CHS 828 can induce total tumor regression in experimental neuroblastoma at a subtoxic dose given orally on a daily basis. As early as after five days, CHS 828 had caused a reduction of the initial tumor size by 29 %. The effect of CHS 828 is potentiated by addition of the angiogenesis inhibitor TNP-470 or SU5416.
6. Two new mechanisms of action of digoxin were discovered, namely an unspecific reduction of angiogenesis and a specific reduction of neuroblastoma growth.

28
General discussion and future perspectives
“Today we have to use tomorrow’s solutions to today’s problems” Richard Norman
There is a wealth of evidence to indicate that tumor growth and spread are angiogenesis dependent. The angiogenic growth factors responsible for progression of clinical neuroblastoma still remain to be identified. Several observations point to VEGF-A as a key mediator. VEGF-A is present in primary neuroblastomas and in neuroblastoma tumor cell lines. It is not clear, however, whether VEGF-A correlates or not with the clinical stage or with MYCN amplification and other prognostic factors (Meitar et al. 1996, Rossler et al. 1999, Eggert et al. 2000, Komuro et al. 2001). Only a few other angiogenic growth factors and their role in neuroblastoma progression have been investigated. For example VEGF-C, and FGF-2 seem to be expressed by neuroblastomas, but do not correlate with neuroblastoma stage (Eggert et al. 2000, Komuro et al. 2001). Thus, with more than 18 known angiogenic growth factors, it is important that their role in the progression of clinical neuroblastoma is investigated. In our model we have mainly investigated the expression of VEGF-A in tumor tissue and plasma samples. We have looked for, but not found, correlations of tumor progression withexpression of FGF-2 and HGF in tumor tissue and plasma. It is possible that other angiogenic growth factors are secreted in neuroblastoma, and inhibition of their pathways would be even more effective than interference with the VEGF pathway. The most relevant angiogenic pathways in experimental neuroblastoma also need to be better characterized. Amplification of the transcription factor MYCN, which is considered to be a proto-oncogene, occurs in 20-25% of clinical neuroblastomas (Maris and Matthay 1999). MYCN overexpression correlates strongly with a poor prognosis. Our experimental model intentionally lacks MYCN expression, but we have just established an experimental model based on the human MYCN-amplified neuroblastoma cell line IMR32, which induces reproducible tumor

29
take in nude mice. Such a model is also suitable for future studies of new, experimental therapies. In vivo models of metastatic neuroblastoma have been described (Khanna et al. 2002). Such models are elegant, but a reproducible, intentionally non-metastatic subcutaneous model such as ours may be used first to determine the efficacy of a new therapy before a more expensive and cumbersome metastatic model is applied. Nevertheless, metastatic models are of considerable value in characterizing biological properties of new therapies. Another problem that needs further attention is how to decide which drugs should be combined in combination therapies. There are quite a few chemotherapeutic drugs, and all of them may not be suitable for combination studies. We suggest screening of chemotherapeutic drugs in vitro under hypoxic conditions to in order to identify candidate drugs. We hypothesize that if the tumor blood flow is restricted by angiogenesis inhibition, a chemotherapeutic is needed that is at least equally potent under hypoxic conditions will be required. However, the questions of what changes take place in the cellular microenvironment, oxygenization, and blood flow of tumors during angiogenesis inhibition needs further investigation.

30
Sammanfattning på svenska
Neuroblastom är en barncancer som drabbar 10-15 barn per år i Sverige. Trots framgångar med intensiv cellgiftsbehandling och kirurgi, kvarstår en relativt hög dödlighet. Denna avhandling fokuserar på en helt ny typ av läkemedel, s.k. angiogeneshämmare. Dessa läkemedel hindrar tumörer från att bilda nya blodkärl och därmed stryps syre- och näringstillförsel till tumören. Dessa hämmare kan fungera på olika sätt. De kan dels direkt döda de celler (endotelceller) som bildar de finaste blodkärlen, kapillärerna. Dels kan de hindra tumörcellerna från att skicka ut tillväxtfaktorer. De kan också blockera mottagarmolekylerna för dessa tillväxtfaktorer på endotelcellerna och därmed förhindra nybildning av blodkärl. Det finns i dag ett 20-tal angiogeneshämmare som prövas på patienter både i Europa och i USA. I denna avhandling undersöktes effekterna av två av dessa angiogeneshämmare, TNP-470 och SU 5416, i en djurexperimentell modell för neuroblastom. Vi fann att neuroblastomen uttryckte den tillväxtfaktor (VEGF) vars mottagare på endotelceller SU 5416 hämmar. Vi fann också att SU5416 hämmade neuroblastomtillväxten med 65 % utan biverkningar. TNP-470 förhindrar tillväxt av endotelceller, men när vi testade denna substans fann vi överraskande nog att blodkärlnybildningen hämmades med ca 40% utan att någon hämning av tumörtillväxten skedde. Detta tyder på att det finns ett tröskelvärde för hur mycket blodkärlnybildningen måste hämmas innan tumörtillväxten bromsas. Vi fann också att dessa substanser med fördel kunde kombineras med det nya cellgiftet CHS 828, som i sig kunde bota bortåt hälften av neuroblastomen. Ett gammalt beprövat läkemedel mot hjärtsvikt, digoxin, visade sig också kunna hämma blodkärlsnybildning och tillväxt av neuroblastom med 44%. Detta nya fynd öppnar en möjlighet för att använda digoxin som en modell för utveckling av nya angiogeneshämmare.

31
Acknowledgements
These studies were performed at the Department of Medical Cell Biology, Uppsala University, Sweden. This work was supported by grants from the Swedish Cancer Society, the Children’s Cancer Foundation of Sweden, H R H Crown Princess Lovisa’s Association for Child Medical Care, the Gunnar, Arvid and Elisabeth Nilsson Foundation, the Dagmar Ferb Memorial Foundation, the Mary Béve Foundation, and grants from the Faculty of Medicine at Uppsala University. I wish to express my sincere gratitude to: Associate Professor Rolf Christofferson, my supervisor, for his enthusiasm, for sharing his vast knowledge about angiogenesis research, and for teaching me how to make colorful presentations of my work. Without his help and patience, this thesis would never have been written. Professor Godfried Roomans, Professor Erik Persson and Professor Emeritus Ove Nilsson for all their kind help and for sharing their knowledge, especially during my first year as a PhD student. Barbro Einarsson, Ulrika Bäckman, Erik Sköldenberg, Mats Hjortberg and all the other members of the Angiogenesis Research group, for interesting discussions about science, wine, and other essential facts of life. The X-group and all the members of the Unit of Human Anatomy, for creating a warm and friendly working athmosphere and for all their valuable help whenever needed. All the members of the Department of Medical Cell Biology, for all their help and support, especially Agneta Bäfwe, Karin Öberg och Gun-Britt for excellent administrative assistance.

32
All the past and present members of the Larsson group at the Division of Clinical Pharmacology, for all their help and for being the first to show me how much fun it is to work with science in a friendly environment. Fredrik Hedborg, Helena Hermelin, Anders Larsson and Lena Fredriksson, for sharing their scientific and technical expertise. Maud Marsden, for sharing her expertise of the English language. My parents Mona and Jan-Erik, my sisters Annette and Maria and my brother Peter, Birgith and Åke, Anna and Jonas for always supporting me. I love you guys! The Bertilsson-Pfuhl family, for being my surrogate family. The kids Anna, Anton, William, Adam, Alexander, Elvira, Tea, Nathalie, Elis and Filip, for being able to completely take my thoughts off work. My friends, for being my friends even though our contacts seem to be more and more sparse. Göran, thank you for your support, encouragement, sharing your scientific knowledge and for loving me. Simon, you are the sunshine of my life! And finally, I would like to thank my cats that….frankly were no help at all……

33
References
Amaral, S. L., Roman, R. J., and Greene, A. S. (2001). Renin gene transfer restores angiogenesis and vascular endothelial growth factor expression in dahl s rats. Hypertension 37, 386-390.
Antoine, N., Greimers, R., De Roanne, C., Kusaka, M., Heinen, E., Simar, L. J., and Castronovo, V. (1994). AGM-1470, a potent angiogenesis inhibitor, prevents the entry of normal but not transformed endothelial cells into the G1 phase of the cell cycle. Cancer Res. 54, 2073-2076.
Beierle E. A., Strande L. F., and Chen M. K. (2002). VEGF upregulates BCL-2 expression and is associated with decreased apoptosis in neuroblastoma cells. J. Ped. Surg. 37, 467-471.
Beltinger C., and Debatin K.-M. (2001). Murine models for experimental therapy of pediatric solid tumors with poor prognosis. Int. J. Cancer 92, 313-318.
Benjamin L. E., Golijanin D., Itin A., Pode D., and Keshet E. (1999). Selective ablation of immature blood vessels in established human tumors follows vascular endothelial growth factor withdrawal. J. Clin. Invest. 103,159-165.
Berthold, F. and Hero, B. (2000). Neuroblastoma: current drug therapy recommendations as part of the total treatment approach. Drugs 59, 1261-1277.
Biedler J. L., Helson L., and Spengler B. A. (1973). Morphology and growth, tumorigenity, and cytogenetics of human neuroblastoma cells in continuous culture. Cancer Res. 33, 2643-2652.
Bocci G., Nicolau K. C., and Kerbel R. S. (2002). Protracted low-dose effects on human endothelial cell proliferation and survival in vitro reveal a selective antiangiogenic window for various chemotherapeutic drugs. Cancer Res. 62, 6938-6943.
Bogenmann E. (1996). A metastatic neuroblastoma model in SCID mice. Int. J. Cancer 67, 379-385.
Brodeur GM and Castleberry RP. Neuroblastoma. Pizzo PA and Poplack DG. 3rd, 761-797. 1997. Philadelphia, Lippincott-Raven Publishers. Principles and Practice of Pediatric Oncology.
Browder, T., Butterfield, C. E., Kraling, B. M., Shi, B., Marshall, B., O'Reilly, M. S., and Folkman, J. (2000). Antiangiogenic scheduling of chemotherapy improves efficacy against experimental drug-resistant cancer. Cancer Res. 60, 1878-1886.
Carmeliet P., Mackman N., Moons L., Luther T., Gressens P., Van Vlaenderen I., Demunck H., Kasper M., Breier G., Evrard P., Muller M., Risau W., Edgington T., and Collen D. (1996). Role of tissue factor in embryonic blood vessel development. Nature 383, 73-75.
Carmeliet P., Dor Y., Herbert J. M., Fukumura D., Brusselmans K., Dewerchin M., Neeman M., Bono F., Abramovitch R., Maxwell P., Koch C. J., Ratcliffe P., Moons l., Jain R. K., Collen D. and Keshet E. (1998). Role of HIF 1α in hypoxia-mediated apoptosis, cell proliferation and tumor angiogenesis. Nature 394, 485-490.

34
Castel V., Canete A., Navarro S., Garcia-Miguel P., Melero C., Acha T., Navajas A. and Badal M. D. (2001). Outcome of high-risk neuroblastoma using a dose intensity approach: improvement in initial but not in long-term results. Med. Pediatr. Oncol. 37, 537-542.
Chang Y. S., di Tommaso E., McDonald D. M., Jones R., Jain R. K., and Munn L. L. (2000). Mosaic blood vessels in tumors: frequency of cancer cells in contact with flowing blood. Proc. Natl. Acad. Sci. USA 97, 14608-14613.
Connolly D. T., Heuvelman D. M., Nelson R., Olander J. V., Eppley B. L., Delfino J. J., Siegel N. R., Leimbgruber R. M. and Feder J. (1989). Tumor vascular permeability factor stimulates endothelial cell growth and angiogenesis. J. Clin. Invest. 84, 1470-1478.
Cotterill S. J., Pearson A. D., Pritchard J., Foot A. B., Roald B., Kohler J. A., and Imeson J. (2000). Clinical prognostic factors in 1277 patients with neuroblastoma: results of The European Neuroblastoma Study Group 'Survey' 1982-1992. Eur. J. Cancer 36, 901-908.
DaCunha M. F., Meistrich M. L., Fuller L. M., Cundiff J. H., Hagemeister F. B., Velasques W. S., McLaughlin P., Riggs S. A., Cabanillas F. F. and Salvador P. G. (1984). Recovery of spermatogenesis after treatment for Hodgkin’s disease: limiting dose of MOPP chemotherapy. J. Clin. Oncol. 6, 571-577.
D’Amato R. J., Loughnan M. S., Flynn E., and Folkman J. (1994). Thalidomide is an inhibitor of angiogenesis. Proc. Natl. Acad. Sci. USA 91, 4082-4085.
Davidoff A. M., Leary M. A., Ng C. Y., and Vanin E. F. (2001). Gene therapy-mediated expression by tumor cells of the angiogenesis inhibitor flk-1 results in inhibition of neuroblastoma growth in vivo. J. Pediatr. Surg. 36, 30-36.
Deplanque G., and Harris A. L. (2000). Anti-angiogenic agents. clinical trial design and therapies in development. Eur. J. Cancer 36, 1713-1724.
Dimmeler S., Dernbach E and Zeiher A. M. (2000). Phosphorylation of the endothelial nitric oxide synthase at Ser-1177 is required for VEGF-induced endothelial cell migration. FEBS Lett. 477, 258-262.
Drevs J., Hofmann, Hugenschmidt H., Wittig C., Madjar H., Mullar M., Wood J., Martiny-Baron G., Unger C., and Marme D. (2000). Effects of PTK/ZK 222584, a specific inhibitor of vascular endothelial growth factor receptor tyrosine kinases, on primary tumor, metastasis, vessel density, and blood flow in a murine renal cell carcinoma model. Cancer Res. 60, 4819-4824.
Dvorak H. F., Brown L. F., Detmar M., and Dvorak A. M. (1995). Vascular permeability factor/vascular endothelial growth factor, microvascular hyperpermeability, and angiogenesis. Am. J. Pathol. 146, 1029-1039.
Eggert A., Ikegaki N., Kwiatkowski J., Zhao H., Brodeur G. M., and Himelstein B. P. (2000). High-level expression of angiogenic factors is associated with advanced tumor stage in human neuroblastomas. Clin. Cancer Res. 6, 1900-1908.
Ekelund S., Liminga G., Bjorkling F., Ottosen E., Schou C., Binderup L., and Larsson R. (2000). Early stimulation of acidification rate by novel cytotoxic pyridyl cyanoguanidines in human tumor cells: comparison with m- iodobenzylguanidine. Biochem. Pharmacol. 60, 839-849.
Fakhari M., Pullirsch D., Paya K., Abraham D., Hofbauer R., and Aharinejad S. (2002). Upregulation of vascular endothelial growth factor receptors is associated with advanced neuroblastoma. J. Pediatr. Surg. 37, 582-587.
Ferrara N. and Alitalo K. (1999). Clinical applications of angiogenic growth factors and their inhibitors. Nat. Med. 5, 1359-1364.

35
Folkman J. (1971). Tumor angiogenesis: therapeutic implications. N. Engl.J. Med. 285, 1182-1186.
Folkman J. (1972). Anti-angiogenesis: new concept for therapy of solid tumors. Ann. Surg. 175, 409-416.
Fong T. A., Shawver L. K., Sun L., Tang C., App H., Powell T. J., Kim Y. H., Schreck R., Wang X., Risau W., Ullrich A., Hirth K. P., and McMahon G. (1999). SU5416 is a potent and selective inhibitor of the vascular endothelial growth factor receptor (Flk-1/KDR) that inhibits tyrosine kinase catalysis, tumor vascularization, and growth of multiple tumor types. Cancer Res. 59, 99-106.
Friedlander M., Brooks P. C., Shaffer R. W., Kincaid C. M., Varner J. A., and Cheresh D. A. (1995). Definition of two angiogenic pathways by distinct alpha v integrins. Science 270, 1500-1502
Fukuzawa M., Sugiura H., Koshinaga T., Ikeda T., Hagiwara N., and Sawada T. (2002). Expression of vascular endothelial growth factor and its receptor Flk-1 in human neuroblastoma using in situ hybridization. J. Pediatr. Surg. 37, 1747-1750.
Glauser T. A., and Packer R. J. (1991). Cognitive deficits in long-term survivors of childhood brain tumors. Childs. Nerv. Syst. 7, 2-12.
Gelaw B. and Levin S. (2001). Wound-induced angiogenesis and its pharmacologic inhibition in a murine model. Surgery 130, 497-501.
Gospodarowicz D., Ferrara N., Schweigerer L. and Neufeld G. (1987). Structural characterization and biological functions of fibroblast growth factor. Endocr. Rev. 8, 95-114.
Gustafsson G., Langmark F., Pihkala U., Heyman M., de Verdier B., and Lilleaas I. J. (1999). Childhood cancer in the nordic countries, Nordic Society of Pediatric Haematology and Oncology, Gothenburg.
Hanahan, D. and Folkman, J. (1996). Patterns and emerging mechanisms of the angiogenic switch during tumorigenesis. Cell 86, 353-364.
Haux J. (1999). Digitoxin is a potential anticancer agent for several types of cancer. Med. Hypotheses 53, 543-548.
Hawkins M. M., Draper G. J. and Kingston J. E. (1987). Incidence of second primary tumors among childhood cancer survivors. Br. J. Cancer 56, 339-347.
Hjarnaa P. J., Jonsson E., Latini S., Dhar S., Larsson R., Bramm E., Skov T., and Binderup L. (1999). CHS 828, a novel pyridyl cyanoguanidine with potent antitumor activity in vitro and in vivo. Cancer Res. 59, 5751-5757.
Hlatky L., Hahnfeldt P., and Folkman J. (2002). Clinical application of antiangiogenic therapy: microvessel density, what it does and doesn’t tell us. J. Natl. Cancer Inst. 94, 883-893.
Holash J., Wiegand S. J., and Yancopoulos G. D. (1999a). New model of tumor angiogenesis: dynamic balance between vessel regression and growth mediated by angiopoietins and VEGF. Oncogene 18, 5356-5362.
Holash J., Maisonpierre P. C., Compton D., Boland P., Alexander C. R., Zagzag D., Yancopoulos G. D., and Wiegand S. J. (1999b). Vessel cooption, regression, and growth in tumors mediated by angiopoietins and VEGF. Science 284, 1994-1998.
Ikeda H., Iehara T., Tsuchida Y., Kaneko M., Hata J., Naito H., Iwafuchi M., Ohnuma N., Mugishima H., Toyoda Y., Hamazaki M., Mimaya J., Kondo S., Kawa K., Okada A., Hiyama E., Suita S., and Takamatsu H. (2002). Experience with international neuroblastoma staging system and pathology classification. Br. J. Cancer 86, 1110-1116.

36
Ingber D., Fujita T., Kishimoto S., Sudo K., Kanamaru T., Brem H., and Folkman J. (1990). Synthetic analogues of fumagillin that inhibit angiogenesis and suppress tumour growth. Nature 348, 555-557.
Jain R. K. (2001). Normalizing tumor vasculature with anti-angiogenic therapy: A new paradigm for combination therapy. Nature Med. 7, 987-989.
Johansson S., Lindholm P., Gullbo J., Larsson R., Bohlin L., and Claeson P. (2001) Cytotoxicity of digitoxin and related cardiac glycosides in human tumor cells. Anticancer Drugs 12, 475-483.
Jones M. K., Wang H., Peskar B. M., Levin E., Itani R. M., Sarfeh I. J. and Tarnawski A. S. (1999). Inhibition of angiogenesis by nonsteroidal anti-inflammatory drugs insight into mechanisms and implications for cancer growth and ulcer healing. Nat. Med. 5, 1418-1423.
Jonsson E., Friberg L. E., Karlsson M. O., Hassan S. B., Freijs A., Hansen K., and Larsson R. (2000). Determination of drug effect on tumour cells, host animal toxicity and drug pharmacokinetics in a hollow-fibre model in rats. Cancer Chemother. Pharmacol. 46, 493-500.
Kakeji Y. and Teicher B. A. (1997). Preclinical studies of the combination of angiogenic inhibitors with cytotoxic agents. Invest. New Drugs 15, 39-48.
Kerbel R. S. (1998-1999). What is the optimal rodent model for anti-tumor drug testing? Cancer Metastasis Rev. 17, 301-304.
Kevil C. G., Payne D. K., Mire E. and Alexander J. S. (1998). Vascular permeability factor/vascular endothelial growth factor-mediated permeability occurs through disorganization of endothelial junctional proteins. J. Biol. Chem. 273, 15099-15103.
Khanna C., Jaboin J. J., Drakos E., Tsokos M., and Thiele C. J. (2002). Biologically relevant orthotopic neuroblastoma xenograft models: primary adrenal tumor growth and spontaneous distant metastasis. In Vivo 16, 77-86.
Killion J. J., Radinsky R., and Fidler I. J. (1999). Orthotopic models are necessary to predict therapy of transplantable tumors in mice. Cancer Meastasis Rev. 17, 279-284.
Kim E. S., Soffer S. Z., Huang J., McCrudden K., Yokoi A., Manley C. A., Middlesworth W., Kandel J. J., and Yamashiro D. J. (2002a). Distinct response of experimental neuroblastoma to combination antiangiogenic strategies. J Pediatr. Surg. 37, 518-522.
Kim E. S., Serur A., Huang J., Manley C. A., McCrudden K. W., Frischer J. S., Soffer S. Z., Ring L., New T., Zabski S., Rudge J. S., Holash J., Yancopoulos G. D., Kandel J. J., and Yamashiro D. J. (2002b). Potent VEGF blockade causes regression of coopted vessels in a model of neuroblastoma. Proc. Natl. Acad. Sci. USA , 11399-11404.
Kingston J. E., Hawkins M. M., Draper G. J., Marsden H. B., and Kinnier Wilson L. M. (1987). Patterns of multiple primary tumors in patients treated for cancer during childhood. Br. J. Cancer 56, 331-338.
Klagsbrun M., and Soker S. (1993). VEGF/VEPF: the angiogenic factor found? Curr. Biol. 3, 699-702.
Klein S. A., Bond S. J., Gupta S. C., Yacoub O. A. and Anderson G. L. (1999). Angiogenesis inhibitor TNP-470 inhibits murine cutaneous wound healing. J. Surg. Res. 82, 268-274.
Klement G., Baruchel S., Rak J., Man S., Clark K., Hicklin D. J. , Bohlen P., and Kerbel R. S. (2000). Continuous low-dose therapy with vinblastine and VEGF receptor-2 antibody induces sustained tumor regression without overt toxicity [see comments]. J. Clin. Invest 105, R15-R24.

37
Kohn S., Nagy J. A. Dvorak H. F. and Dvorak A. M. (1992). Pathways of macromolecular tracer transport across venules and small veins. Structural basis for the hyperpermeability of tumor blood vessels. Lab. Invest. 67, 596-607.
Komuro H., Kaneko S., Kaneko M., and Nakanishi Y. (2001). Expression of angiogenic factors and tumor progression in human neuroblastoma. J. Cancer. Res. Clin. Oncol. 12, 739-743.
Kusaka M., Sudo K., Fujita T., Marui S., Itoh F., Ingber D., and Folkman J. (1991). Potent anti-angiogenic action of AGM-1470: comparison to the fumagillin parent. Biochem. Biophys. Res. Commun. 174, 1070-1076.
Kusaka M., Sudo K., Matsutani E., Kozai Y., Marui S., Fujita T. , Ingber D., and Folkman J. (1994). Cytostatic inhibition of endothelial cell growth by the angiogenesis inhibitor TNP-470 (AGM-1470). Br. J. Cancer 69, 212-216.
Larsson R., Kristensen J., Sandberg C., Nygren P. (1992). Laboratory determination of chemotherapeutic drug resistance in tumor cells from patients with leukemia, using a fluorometric microculture cytotoxicity assay (FMCA). Int. J. Cancer 50, 177-185.
Lee C.-G. Heijn M., Tomaso E. D., Griffon-Etienne G., Ancukiewicz M., Kolke C., Park K. R., Ferrara N., Jain R. K., Suit H. D., and Boucher Y. (2000). Anti-vascular endothelial growth factor treatment auguments tumor radiation response under normoxic or hypoxic conditions. Cancer Res. 60, 5565-5570.
Lyden D., Hattori K., Dias S., Costa C., Blaikie P., Butros L., Chadburn A., Heissig B., Marks W., Witte L., Wu Y., Hicklin D., Zhu Z., Hackett N., Crystal R. G, .Moore M. A. S., Hajjar K. A., Manova K., Benezra R., and Rafii S. (2001). Impaired recruitment of bone-marrow-derived endothelial and hematopoietic precursor cells blocks tumor angiogenesis and growth. Nature 7, 1194-1201.
Maris J. M., and Matthay K. K. (1999). Molecular biology of neuroblastoma. J. Clin. Oncol. 17, 2264-2279.
Maxwell P. H., Dachs G. U., Gleadle J. M., Nicholls L. G., Harris A. L., Stratford I. J., Hankinson O., Pugh C. W. and Ratcliffe P. J. (1997). Hypoxia-inducible factor-1 modulates gene expression in solid tumors and influences both angiogenesis and tumor growth. Proc. Natl. Acad. Sci. USA 94, 8104-9109.
Meister B., Grunebach F., Bautz F., Brugger W., Fink F. M., Kanz L., and Mohle R. (1999). Expression of vascular endothelial growth factor (VEGF) and its receptors in human neuroblastoma. Eur. J. Cancer 35, 445-449.
Meitar D., Crawford S. E., Rademaker A. W., and Cohn S. L. (1996). Tumor angiogenesis correlates with metastatic disease, N-myc amplification, and poor outcome in human neuroblastoma. J. Clin. Oncol. 14 , 405-414.
Miller K. D., Sweeney C. J., and Sledge G. W. (2001). Redefining the targets: chemotherapeutics as antiangiogenics. J. Clin. Oncol. 19, 1195-1206.
Mohammed S. I., Bennett P. F., Craig B. A., Glickman N. W., Mutsaers A. J., Snyder P. W., Widmer W. R., DeGortari A. E., Bonney P. L., and Knapp D. W. (2002). Effects of the cyclooxygenase inhibitor, piroxicam, on tumor response, apoptosis, and angiogenesis in a canine model of human invasive urinary bladder cancer. Cancer Res. 62, 356-358.
Montaldo P. G., Lanciotti M., Casalaro A., Cornaglia-Ferraris P., and Ponzoni M. (1991). Accumulation of m-iodobenzylguanidine by neuroblastoma cells results from independent uptake and storage mechanisms. Cancer Res. 51, 4342-4346.
Moses M. A. (1997). The regulation of neovascularization of matrix metalloproteinases and their inhibitors. Stem Cells. 15, 180-189.
Mott M. G., Foreman N. K., and Chambers E. J. Cancer in childhood. Price, P. Sikora K. 3rd, 909-936. 1995. London, Chapman & Hall. Treatment of cancer.

38
Nagabuchi E., VanderKolk W. E., Une Y., and Ziegler M. M. (1997). TNP-470 antiangiogenic therapy for advanced murine neuroblastoma. J. Pediatr. Surg. 32, 287-293.
Patan S., Munn L.L. and Jain R. K. (1996). Intussusceptive microvascular growth in a human colon adenocarcinoma xenografts: a novel mechanism of tumor angiogenesis. Microvasc. Res. 51, 260-272.
Peichev M., Naiyer A. J., Pereira D., Zhu Z., Lane W. J., Williams M., Oz M. C., Hicklin D. J., Witte L., Moore M. A. S., and Rafi S. (2000). Expression of VEGFR-2 and AC133 by circulating human CD34+ cells identifies a population of functional endothelial precursors. Blood 95, 952-958.
Pietras K., Östman A., Sjökvist M., Buchdunger E., Reed R: K., Heldin C.-H., and Rubin K. (2001). Inhibition of platelet-derived growth factor receptors reduces interstitial hypertension and increases transcapillary transport in tumors. Cancer Res. 61, 2929-2934.
Ribatti D., Raffaghello L., Pastorino F., Nico B., Brignole C., Vacca A., and Ponzoni M. (2002). In Vivo angiogenic activity of neuroblastoma correlates with MYCN oncogene over expression. Int. J. Cancer 102, 351-354.
Rosenberg M. P., and Bortner D. (1998-1999) Why transgenic and knockout animal studies should be used (for drug efficacy studies in cancer). Cancer Metastasis Rev. 17, 295-299.
Rossler J., Breit S., Havers W., and Schweigerer L. (1999). Vascular endothelial growth factor expression in human neuroblastoma: up-regulation by hypoxia. Int. J. Cancer 81, 113-117.
Rowe D. H., Huang J., Li J., Manley C., O’Toole K. M., Stolar C. J., Yamashiro D. J., and Kandel J. J. (2000). Supression of primary tumor growth in a mouse model of human neuroblastoma. J. Pediatr. Surg. 35, 977-981.
Ryan H. E., Lo J., and Johnson R. S. (1998). HIF-1 alpha is required for solid tumor formation and embryonic vascularization. EMBO 17, 3005-3015.
Samra Y., Hertz M. and Lindner A. (1985). Urinary bladder tumors following cyclophosphamide therapy: a report of two cases with a review of the literature. Med. Pediatr. Oncol. 13, 86-91.
Sarkar C., Pramanik P., Karak A. K., Mukhopadhyuy P., Sharma M. C., Singh V. P., and Mehta V. S. (2002). Are childhood and adult medulloblastomas different? A comparative study of clinicophatological features, proliferation index and apoptotic index. J. Neurooncol., 59, 49-61.
Sawaoka H., Tsuji S., Tsujii M., Gunawan E. S., Sasaki Y., Kawano S., and Hori M. (1999). Cyclooxygenase inhibitors suppress angiogenesis and reduce tumor growth in vivo. Lab. Invest. 79, 1469-1477.
Shweiki D., Neeman M., Itin A. and Keshet E. (1995). Induction of vascular endothelial growth factor expression by hypoxia and by glucose deficiency in multicell spheroids: implications for tumor angiogenesis. Proc. Natl. Acad. Sci. USA 92, 768-772.
Soffer S. Z., Moore J. T., Kim E. Huang J., Yokoi A., Manley C., O’Toole K., Stolar C., Middlesworth W., Yamashiro D. J, and.Kandel J. J. (2001). Combination anti-angiogenic therapy: increased efficacy in a murine model of Wilms tumor. J Pediatr. Surg. 36, 1781-1784.
Steinherz L. J., Steinherz P. G., Tan C. T., Heller G. and Murphy M. L. (1991). Cardiac toxicity 4 to 20 years after completing anthracycline therapy. JAMA 266, 1672-1677.
Stenkvist B. (1999). Is digitalis a therapy for breast carcinoma? Oncol. Rep. 6, 493-6.

39
Styczynski J., Pieters R., Husimans D. R., Shuurhuis G. J., Wysocki M., and Veerman A. J. (2000). In vitro drug resistance profiles of adult versus childhood acute lymphoblastic leukemia. Br. J. Haematol. 110, 813-818.
Teicher B. A., Emi Y., Kakeji Y., and Northey D. (1996). TNP-470/minocycline/cytotoxic therapy: a systems approach to cancer therapy. Eur. J. Cancer 32A, 2461-2466.
Teicher B. A., Holden S. A., Dupuis N. P., Kakeji Y., Ikebe M., Emi Y., and Goff D. (1995). Potentiation of cytotoxic therapies by TNP-470 and minocycline in mice bearing EMT-6 mammary carcinoma. Breast Cancer Res. Treat. 36, 227-236.
Terranova V. P., DiFlorio R., Lyall R. M., Hic S., Friesel R. and Maciag T. (1985). Human endothelial cells are chemotactic to endothelial cell growth factor and heparin. J. Cell. Biol. 101, 2330-2334.
Troncone L. and Rufini V. (1997). 131I-MIBG therapy of neural crest tumours (review). Anticancer Res. 17, 1823-1831.
Turner W. J., Chatten J., and Lampson L. A. (1990). Human neuroblastoma cell growth in xenogenic hosts: comparison of T cell-deficient and NK-deficient hosts, and subcutaneous of intravenous injection routes. J. Neurooncol. 8, 121-132.
Wassberg E., Hedborg F., Sköldenberg E. G., Stridsberg M., and Christofferson R. H. (1999). Inhibition of angiogenesis induces chromaffin differentiation and apoptosis in neuroblastoma. Am. J. Pathol. 154, 395-403.
Wassberg E., Pahlman S., Westlin J. E., and Christofferson R. (1997). The angiogenesis inhibitor TNP-470 reduces the growth rate of human neuroblastoma in nude rats. Pediatr. Res. 41, 327-333.
Wu P., Yonekura H., Li H., Nozaki I., Tomono Y., Naito I., Ninomiya Y., Yamamoto H. (2001). Hypoxia down-regulates endostatin production by human microvascular endothelial cells and pericytes. Biochem. Biophys. Res. Commun. 288,1149-54.
Yancopoulos G. D., Davis S., Gale N. W., Rudge J. S., Wiegand S. J., and Holash J. (2000). Vascular-specific growth factors and blood vessel formation. Nature 407, 242-248.
Yoshizawa J., Mizuno R., Yoshida T., Hara A., Ashizuka S., Kanai M., Kuwashima N., Kurobe M. , and Yamazaki Y. (2000). Inhibitory effect of TNP-470 on hepatic metastasis of mouse neuroblastoma. J. Surg. Res. 93, 82-87.