urolithiasis in rats consuming a dl bitartrate form of...
TRANSCRIPT

Comparative MedicineCopyright 2005by the American Association for Laboratory Animal Science
Vol 55, No 4August 2005
354
Received: 12/1/04. Revision requested: 5/23/05. Accepted: 5/25/05.1Behavioral Toxicology Laboratory, Experimental Psychology, 226 Thach, Auburn University, Alabama 36849, 2Department of Psychology, Alabama State University, Montgomery, Alabama 36101, 3Department of Pathobiology, 166 Greene Hall, College of Veterinary Medicine, Auburn University, Alabama 36849, 4Laboratory Animal Health, 311 Greene Hall Annex, College of Veterinary Medicine, Auburn University, Alabama 36849, 5Department of Nutrition and Food Science, 328 Spidle Hall, Auburn University, Alabama 36849, 6Department of Medicine/Research Ser-vice, Medical College of Wisconsin/Zablocki VA Medical Center, 5000 W. National Avenue, Milwaukee, Wisconsin 53295.
*Corresponding author.
The present paper describes the natural history of an out-break of urolithiasis in a colony of Long-Evans rats maintained to study the interactions between nutrients and methylmercury on development and aging. The appearance of this disease in rats and its association with choline bitartrate in some shipments of purified diets (15) indicates that racemic bitartaric acid plays a role, previously unrecognized, in the development of urolithiasis in rats and probably in other animals or in humans.
Laboratory animal models of urolithiasis have focused on cal-cium, oxalate, phosphate, magnesium, ammonia, urinary pH, and citrate as contributors to the formation of uroliths (19). These dietary components are important determinants of the composi-tion and supersaturation potential of urine (18, 19, 27, 30, 31, 35), factors that contribute to the potential for urolith formation (19). Molar ratios of calcium to phosphorus are especially critical; a ratio that is too low can cause renal calcium in the kidney to precipitate into uroliths (31).
The American Institute of Nutrition (AIN)-76 diet was refor-mulated in 1993, in part because of its tendency to induce a uri-nary tract chemistry favorable to the formation of calculi and to produce calcium deposits in the kidneys of female rats (31). The calcium-to-phosphorus (Ca:P) ratio was increased from 0.75 in the AIN-76 formulation to 1.3:1 in the newer AIN-93 formu-
Urolithiasis in Rats Consuming a dl Bitartrate Form of Choline in a Purified Diet
M. Christopher Newland,1,* Phyllis A. Reile,2 Eva A. Sartin,3 Michael Hart,4 Margaret C. Craig-Schmidt,5
Ian Mandel,6 and Neil Mandel6
Urolithiasis appeared in rats maintained to study the effects of nutrients and methylmercury on development and aging. After a year, the mortality rate was approximately 10%, and by 2 years, it had increased to nearly 30%. Clini-cal signs and urinary tract pathology were examined as a function of diet, duration on diet, gender, methylmercury exposure, genetics, and other potential risk factors by using survival analyses and qualitative comparisons. Urolithia-sis in female rats appeared 15 weeks after beginning a purified diet and after 5 weeks for male rats. After 97 weeks, the mortality rate of female rats was 22% and for male rats was 64%. Lifetime urolithiasis-associated mortality was about 2% in a group of rats that consumed the contaminated diet for < 30 weeks. No urolithiasis occurred in siblings or cohorts of the rats described here that were maintained on a standard rodent chow containing choline chloride. Urolithiasis was traced to racemic, rather than levo-, bitartaric acid in some purified diets shipped in 2001 and 2002. It is unknown when the impurity first appeared in the diet, so estimates of exposure duration are upper limits. Chronic methylmercury exposure increased vulnerability. Some families (dam + offspring) had multiple cases of urolithiasis, but probability models constructed to evaluate familial clustering revealed no evidence for a genetic predisposition to urolithiasis apart from gender. Removing racemic tartaric acid did not decrease mortality once rats had been on the diet for 20 to 30 weeks, but it helped when exposure duration was shorter.
lation. This improvement yields renal calcium concentrations that are 26 times lower than those seen in the older diet; the decreased calcium concentrations are thought to be inconsistent with urolith formulation (31). In this respect, rats on the AIN-93 diet resemble those on standard rodent chow diets. For this reason, the appearance of urolithiasis in rats on the AIN-93 diet was unexpected.
The AIN-93 revision also made the purified diet especially suitable for long-term feeding studies (7, 8). This, and the control that purified diets offer in mineral and lipid concentrations (30, 31), led us to select AIN-93 to study interactions between dietary polyunsaturated fatty acids and methylmercury exposure. Rats were maintained on diets based on the AIN-93 formulation but that differed in their concentration of n-3 polyunsaturated fatty acids. In addition, some rats were maintained on various concen-trations of methylmercury in their drinking water, whereas oth-ers were exposed to methylmercury only during gestation. These conditions afforded examination of dietary lipids and methylmer-cury as risk factors for urolithiasis, especially relevant because the kidney accumulates mercury (12, 32).
Suspicion that choline bitartrate might be linked to urolithia-sis in our colony and others experiencing similar outbreaks arose after a vendor noted that rats fed a choline-deficient AIN-93 based diet did not develop urolithiasis. Klurfeld (15) later report-ed the presence of urolithiasis in several colonies in the United States and that “synthetic dl bitartaric acid was substituted for the previously used neutral l tartaric acid isomer” in some ship-ments of the diet. The AIN formulation specifies that choline be stabilized with chloride or the natural (levo) isomer of bitartaric acid. Klurfeld also noted that dietary concentrations of racemic (dl) tartaric acid previously associated with urolithiasis were 10 times greater than the concentration in the suspect shipments.
Pages 354-367

355
The current episode, therefore, indicates that racemic bitartaric acid is far more potent than previously appreciated.
Further direct evidence for urolith-forming properties of race-mic tartaric acid was demonstrated recently by provoking uro-lithiasis experimentally in rats fed a diet containing racemic choline bitartrate (13). Urolithiasis then was prevented by coad-ministration of a P glycoprotein inhibitor, a finding that suggests a mechanism for urolith formation (13). Finally, the racemic form of tartaric acid remains in the kidney longer than does the levo form and promotes the formation of crystals and calculi (6).
When the outbreak described here occurred, laboratories in the United States and Canada that reported a similar problem were located and contacted. The results of these contacts are described below. Projects involving aging, neuroscience, cardiovascular function, pharmacology, and toxicology were affected, but the con-sistent factor was that they maintained rats on an AIN-93 formu-lation for at least several months and over a similar span of time, roughly between 2001 and 2002. By the time the dietary cause of urolithiasis was identified, the present study had progressed far enough to favor completion while tracking its progress. The present report, therefore, is an opportunistic rather than planned investigation of urolithiasis from a dietary source. The following potential risk factors received detailed attention: gender, n-3 poly-unsaturated fatty acids, genetics, duration of exposure to choline bitartaric acid, removal of choline bitartrate, methylmercury ex-posure, and infectious or environmental processes.
Materials and MethodsAnimals. Two generations of Long-Evans rats were fed cus-
tomized diets based on the AIN-93 formulation to study interac-tions between nutrients and methylmercury on development and aging. Water was available ad libitum. A total of 364 rats maintained on the purified diet began the investigation, and 74 cases of urolithiasis were diagnosed on the basis of clinical signs and pathology. As determined through survival analyses (de-scribed later), the mortality rate due to urolithiasis is higher than implied by these numbers because many rats were euth-anized (“censored”) before they had an opportunity to develop urolithiasis.
In the F0 generation, there were 108 rats (all female) with 22 cases of urolithiasis. In the F1 generation, there were 256 rats with 52 cases of urolithiasis. Thirty of the F1 rats were male, and there were 17 cases of urolithiasis among these. An additional 64 female F1 rats, siblings of the ones described here, were placed on a chow diet (Harlan Teklad) at weaning and later were sent to
another institution for long-term behavioral testing (not listed in Table 1 and not included in survival analyses). These rats were food-restricted and maintained under similar conditions to the rats described here.
The F0 generation comprised 72 (Batch 1) and 36 (Batch 2) female Long-Evans rats obtained from Harlan (Indianapolis, Ind.) at 2.5 months of age (Table 1). Batch 2 rats were obtained 4.5 months after Batch 1 rats, because of breeding difficulties in one experimental group. Once at Auburn University, the rats were conventionally housed in shoebox cages with aspen bedding (Sani-Chips, Harlan Teklab, Indianapolis, Ind.) and wire-bar tops (no isolator tops or forced ventilation). Bedding was changed twice weekly, and rats were housed individually in an environ-mentally controlled colony room with a 12:12-h light:dark cycle (lights on, 0700). Individual housing was necessary because be-havioral testing used food reinforcers, so each rat’s food intake had to be controlled individually.
The vendor’s health surveillance reports obtained upon pro-curement revealed the barrier facility of origin to be specific pathogen-free as follows: serologically negative (enzyme-linked immunosorbent assays, immunofluorescent antibodies, and hemagglutination inhibition assays were used) for Mycoplasma pulmonis, Sendai virus, sialodacryoadenitis virus/rat corona vi-rus, Killham’s rat virus, Toolan’s H-1 virus, rat parvovirus, pneu-monia virus of mice, reovirus 3, Hantaan virus, transmissible murine encephalomyelitis virus, mouse adenovirus, lymphocytic choriomeningitis virus, Clostridium piliforme, and Encephali-tozoon cuniculi; polymerase chain reaction assay (PCR)-nega-tive for cilia-associated respiratory bacillus and cytomegalovirus (some of the sentinel rats were PCR-positive for Helicobacter sp., with no clinical disease); respiratory and enteric cultures were negative for Salmonella spp., Corynebacterium kutscheri, Bordetella bronchiseptica, Streptococcus zooepidemicus, Strep-tobacillus moniliformis, Group B β-Streptococcus, Streptococcus pneumoniae, Pasteurella pneumotropica, Pseudomonas aerugi-nosa, and Mycoplasma spp.; and examinations for ectoparasites and endoparasites were negative (microscopic examination of pelt and cecal contents as well perianal cellophane tape impres-sions for endoparasites).
Batch 1 rats were allocated into two diet groups and three methylmercury exposure groups (described later) and bred with males maintained on standard rodent chow diets (Harlan Teklad, Indianapolis, Ind.) and tap water. Male breeders were eutha-nized after breeding. Batch 2 was treated identically to Batch 1 except where noted (Table 1). The rats described in Table 1 were
Table 1. Characteristics of the two generations exposed to racemic tartaric acid
F0 Generation (breeders) F1 Generation (offspring)
Batch 1 Batch 2 Batch 1 Batch 2
Number of rats 72 36 201 55 (0 male) (0 male) (29 male) (1 male)
Age (weeks) at which AIN-93 diet commenced 10–12 10 3 3
Mean exposure (weeks) to choline bitartrate diet (range) 45 24 29 10 (39–51) (18–30) (19–39) (5–16)
Mean age (weeks) at which choline chloride diet commenced (range) 56 34 32 13 (51–62) (28–40) (22–42) (8–19)
Methyl-mercury exposure Timing Chronic, began at Chronic, began at Prenatal only Prenatal only 12 weeks of age 12 weeks of age
Concentration in drinking water 0, 0.5, and 5.0 ppm Hg 0 and 5.0 ppm Hg 0, 0.5, and 5.0 ppm Hg 0, 0.5, and 5.0 ppm Hg
Urolithiasis in rats

Vol 55, No 4Comparative MedicineAugust 2005
356
retained for the duration of the project.At birth, large litters were culled to a maximum of eight pups
and weaned at 21 days of age. Food restriction was observed throughout the study in order to enhance the reinforcing effi-cacy of sucrose in behavioral experiments. After weaning, the F0 dams’ body weights were reduced gradually to 260 to 270 g. F1 rats were free-fed until they reached 240 to 250 g (female rats) or 315 to 325 g (male rats). Rats on food restriction were weighed at least three days weekly, and the amount of feed required to maintain a rat’s body weight was determined on an individual basis. Caloric restriction is consistent with good health in life-span studies (7, 9) and, in our experience with Long-Evans rats, these weights establish food as an effective reinforcer for behav-ioral investigation of drugs and toxicants while being consistent with good health (22-26, 29).
Animals were observed daily by the veterinary staff, veteri-nary technician, animal care staff, or the research team. Fre-quent communication regarding animal health issues occurred among all personnel involved in daily care. All experiments were approved by the Auburn University Institutional Animal Care and Use Committee.
Health surveillance program.The animal facility main-tains a quarterly health surveillance program using noncontact sentinel rats housed on the bottom shelf of a cage rack in each colony room. Sentinel rats were exposed to soiled bedding from the cages of rats in that room and to water bottles, stoppers, and feed from rats that died. Sentinel animals removed for termi-nal health surveillance analysis were replaced with new rats. Each quarterly sentinel evaluation involved serology and evalu-ation for external parasites and pinworms. The agents assayed varied between sampling periods but included all of the agents listed in the vendor’s health surveillance report except for En-cephalitozoon cuniculi and cytomegalovirus. In addition, one of the rats in the study that became sick was sampled serologi-cally. A comprehensive analysis was performed on the sentinel animals annually to include necropsy for gross and histopatho-logical evaluation, serology, and microbiological cultures. Before the cause of mortality was ascertained, one rat from the colony showing clinical signs of lethargy and weight loss was evaluated serologically for pathogens.
Diets. Three days after arriving at the colony at 10 to 12 weeks of age, the F0 rats were placed on one of two customized, isoca-loric AIN-93 diets purchased from Dyets, Inc. (Bethlehem, Pa.). The “coconut-oil” and “fish-oil” diets were designed to produce “low” and “high” concentrations of n-3 polyunsaturated fatty ac-ids, respectively. F0 rats consumed a “growth” diet containing 7% fat throughout pregnancy and lactation. F1 rats consumed the growth diet until they reached 170 g (female rats) or 240 g (male rats). At all other times, a “maintenance” diet containing 4% fat was used. To facilitate comparisons with a planned study on se-lenium–methylmercury interactions, selenium content was at a nominal concentration of 0.18 ppm, including contributions from casein, except during breeding, when selenium was increased to 0.4 ppm. Sixty-four F1 rats received a standard rodent chow diet (Harlan Teklad) beginning at weaning.
Both diets contained a fat mixture of 42.8% palm oil, 9.2% saf-flower oil, and 15.0% soybean oil. The coconut oil (CO) mixture included 33% coconut oil, whereas the fish-oil mixture included 33% fish oil (EPAX, Pronova Biocare, Lysaker, Norway) to pro-vide low and high concentrations of n-3 fatty acids, respectively.
The 36 female rats in Batch 2 were all fed the coconut-oil diet. All National Research Council (NRC) recommendations for rodent diets were followed except those addressing selenium concentra-tions and the composition of the fat blends.
Choline was included as choline chloride (equimolar for cho-line), and the diets were purchased from Research Diets (New Brunswick, N.J.) at 9 to 11 months into the study, 6 to 8 months after the first case of urolithiasis. This purchase was prompted by suspicion that choline bitartrate was the cause of urolithiasis (we emphasize here that there is no evidence that the feed ven-dor was at fault, and both vendors were cooperative throughout the investigation). Because it might retard urolith formation, po-tassium citrate was added to the Research Diets formulation to increase the potassium concentration to 6 g/kg, as in the AIN-76 formulation (21, 28, 38). The fat and selenium contents of these diets were as described earlier.
As of this writing, a second study has been initiated, using animals maintained on an AIN-93-based diet, but with differ-ent concentrations of selenium instead of different fat mixtures. Choline chloride is used in the diet instead of choline bitartrate. Breeders have been on the diet for approximately 58 weeks, and no case of urolithiasis has been seen.
Methylmercury exposure. Upon arrival, the F0 rats were provided tap water. After consuming the purified diet ad libitum for 18 days, they were weighed, and each diet group was fur-ther allocated into methylmercury-exposure groups matched for weight (Table 1). The 72 dams in Batch 1 received tap water con-taining 0, 0.5, or 5.0 ppm mercury (as methylmercuric chloride; Alfa Aesar, Ward Hill, Mass.). Sodium carbonate buffer (< 5 nM) was added to help dissolve the methylmercury (36), and this buffer also was added to the water of the no-mercury group. Of the 36 dams in Batch 2, 8 received tap water only, and 28 were given water containing 5.0 ppm mercury as methylmercuric chloride. Methylmercury exposure began 22 days before breed-ing commenced.
Clinical signs and pathology. Rats were observed daily. Clinical signs suggesting urolithiasis included (in various com-binations) hematuria, red-stained bedding, red-stained or wet pelage (especially over the abdomen), sensitivity to touch in the abdominal area, swollen or palpable kidney or bladder, unkempt fur, anorexia, reduced urination, reduced water intake, and un-expected weight loss or gain (due to fluid retention). Animals were permitted to recover if they did not appear in distress. How-ever, anorexia and abrupt weight loss or gain indicated a poor prognosis and prompted timely euthanasia with CO2 or pento-barbital overdose. In addition, moribund rats were euthanized immediately. The transition from onset of clinical signs to being moribund or dying ranged from hours to days. Regrettably, the occasional rapid progression to illness sometimes precluded in-tervention with euthanasia.
Necropsy reports were available on 130 animals. Necropsy consisted of gross evaluation of the respiratory, digestive, uri-nary, hematopoietic, lymphoid, genital, and musculoskeletal sys-tems. Tissues that were prepared for histopathology were fixed in 10% buffered formalin, embedded in paraffin, and processed routinely.
Investigation of other potential variables. All rats drank municipal water, so Auburn Water Works Board water-quality re-ports were examined to ensure that bacteriological, radiological, inorganic, and organic contaminants were within appropriate

357
ranges. To verify the drinking-water concentrations of methyl-mercury, one sample from each batch of the two exposure levels was sent for analysis by atomic absorption spectrophotometry by the Trace Metals and Organic Contaminants Divisions’ Hu-man Toxicology Branch (INSPQ, Quebec, Canada). This analysis was conducted when methylmercury-containing drinking water solutions were prepared, about every 1 to 3 months, depending on how many rats were consuming mercury. Water samples from three individual rats were sent to Auburn University’s Soil Test-ing Laboratory for determination of 16 minerals (Ca, K, Mg, P, Cu, Fe, Mn, Zn, B, Al, Ba, Co, Cr, Pb, and Na). Nylon chew-bones had been provided to inhibit incisor overgrowth 1 month prior to the first death. The rats’ gnawing of these caused tiny flakes to appear that, conceivably, could have irritated or obstructed the GI tract, so the chew-bones were removed after the first few deaths occurred and returned to the rats after the cause of death was ascertained.
Diet samples were tested by one of the authors (MCS) to en-sure that the fatty acid composition was as intended. Diet sam-ples from rats that died were tested by Covance Laboratories, (Madison, Wis.) and Grand Forks Human Nutrition Research Center (Grand Forks, N. Dak.) to test selenium concentrations and by Auburn University’s Soil Testing Laboratory for deter-mination of the same 16 minerals tested in the water samples. Three to ten samples were sent to each facility.
For a brief period, coincident with the first cases, the humidity in the animal facility was poorly controlled because of a malfunc-tioning humidifier in the air-handling system. This mechanical error resulted in occasional bouts of low humidity (~10%) and, consequently, elevated fluid consumption. Control over humidity soon was regained, and humidity readings remained within a nor-mal range (39 to 52%). Room temperatures were well controlled and stayed between 70 and 74°F (ca. 21 to 23°C). A careful analy-sis of animal care procedures was conducted, as was a review of all protocols in the colony rooms and laboratory for changes in soaps, floor cleaners, and other possible causes, however remote.
Composition of uroliths. Five uroliths from the Auburn colony and three from each of two additional institutions were analyzed by high-resolution x-ray diffraction (XRD) and Fourier transform infrared analysis (FTIR). For the XRD analysis, uro-liths were ground in an agate mortar and pestle and exposed to CuK alpha radiation from a high-brilliance rotating anode x-ray generator (Rigaku/MSC Inc., The Woodlands, Tex.) and a Guinier powder diffraction camera (Robert Huber Diffraktion-stechnik, Rimsting, Germany). Samples were exposed at room temperature for ≤ 1 h with constant sample motion to minimize any preferred orientation of crystals. Diffraction maxima were matched with data from the International Joint Commission on Powder Diffraction Standards (JCPDS, Philadelphia, Pa.) library and the extensive local diffraction library of > 60,000 analyses of human urinary tract calculi.
A portion of the urolith powder was mixed with KBr (Spec-traTech/ThermoNicolet, Madison, Wis.) and pressed into an opti-cally clear wafer for FTIR analysis conducted on a Nexus 670 infrared spectrometer (ThermoNicolet) at room temperature. Sixty-four spectra were collected for each analysis, computer-averaged, and subjected to fast Fourier transform calculation to maximize the signal-to-noise ratio in the spectral data. The resulting spectra were subjected to search–match algorithms us-ing infrared spectral libraries including commercially available
libraries (Sigma Chemical Company, St. Louis, Mo., and Aldrich Chemical Company, Milwaukee, Wis.) and the aforementioned local library.
Analyses of bitartaric acid from other vendors. One of the authors (NM) was contacted by a feed vendor when suspicion developed that choline bitartaric acid might be linked to uroli-thiasis in several colonies. Samples of a purified diet (AIN-93 formulation from Bio-Serv, Frenchtown, N.J.) from other institu-tions were analyzed, and the results are reported here. Choline bitartrate received special scrutiny and therefore was subjected to XRD and FTIR analyses. Multiple batches were analyzed to confirm or eliminate the possibility that the composition changed during the course of the studies.
Appearance of urolithiasis in other colonies in the Unit-ed States. An inquiry was posted on CompMed, an electronic discussion list hosted by the American Association for Laboratory Animal Science, to determine whether other investigators have had similar experiences in their laboratory. If so, then details were sought about the conditions associated with the outbreak.
Survival analyses. Using the rats described in Table 1, non-parametric Kaplan–Meier survivor functions were generated to describe mortality due to urolithiasis and mortality due to other causes. Stratified analyses then were conducted to describe the role of methylmercury exposure in mortality due to all causes other than urolithiasis in the F0 generation (chronic exposure) and F1 generation (prenatal exposure). Finally, stratified analy-ses were conducted to describe mortality due to urolithiasis in the following strata: generation (F0 versus F1), gender for the F1 generation, methylmercury exposure (each generation), dietary variables, and batch (a proxy for duration of exposure to the cho-line bitartrate, conducted separately for each generation). A sep-arate curve was generated for each stratum. A Mantel–Haenszel unweighted log-rank test was used to evaluate statistically the differences among curves. Two weighted versions of the log-rank test (Breslow–Gehan and Tarone–Ware) also were examined, but they produced similar results so they are not presented. Chi-square values and the associated P values are reported. The degrees of freedom is the number of strata – 1; i.e., df = 2 for analyses of methylmercury exposure and 1 for all other analyses. All survival analyses were conducted using SYSTAT software (Port Richman, Calif.).
To conduct survival analyses and analyses of family clusters (described later), a diagnosis of “urolithiasis” was made when clinical signs related to urinary tract disease were present and accompanied by uroliths. Deaths due to other natural causes (tumors, trichobezoars), euthanasia associated with methylmer-cury exposure (for example, long-term methylmercury exposure sometimes induced sensory–motor deficits that impaired eating), or as part of colony management were treated as “censored” data for survival analyses of deaths due to urolithiasis. That is, they were deducted from the census from subsequent, but not previ-ous, calculations of incidence (14). All healthy rats euthanized at the end of the study were treated as censored or nonurolithiasis cases. During survival analyses of “deaths due to other causes,” deaths due to natural causes were treated as cases, but deaths associated with urolithiasis or as part of colony management were treated as censored data.
The presence of uroliths is required for a definitive diagnosis of urolithiasis, but strict reliance on such a criterion for survival analyses or for analyses of litter effects could have skewed the
Urolithiasis in rats

Vol 55, No 4Comparative MedicineAugust 2005
358
outcome. Five cases were scored as “normal” even though there were specific clinical signs suggestive of urinary tract disease (e.g., hematuria and palpably enlarged kidneys) coupled with urinary tract lesions suggestive of previous urolithiasis (mod-erate to severe hydronephrosis with pyelonephritis), but there were no calculi detected at necropsy. It is strongly suspected that calculi had been successfully passed in these five animals. For 11 cases, there were multiple clinical signs, but pathology was not available, so the clinical suspicion could not be confirmed. All survival analyses were repeated using relaxed diagnostic criteria that included these cases. The relaxed criteria did not change any conclusion, and these results are not reported.
Of 236 asymptomatic rats euthanized to manage the colony, for analyses of tissue mercury, or at the termination of the study (some not listed in Table 1), necropsies were conducted on 30 ar-bitrarily selected asymptomatic individuals to determine wheth-er uroliths could be present in otherwise healthy rats. In four animals, small, 1- to 2-mm uroliths were observed at necropsy. These four cases were scored as clinically normal because the uroliths were incidental findings and not a cause of illness or death. Therefore, these incidental cases were treated identically to the rats that were apparently healthy and euthanized but that did not undergo necropsy. Because it not known how many of the uninspected rats had uroliths, it is appropriate to score all asymptomatic cases the same.
Analysis of familial clustering. There were many occasions in which two or more rats from the same litter died of urolithia-sis, suggesting a possible genetic contribution to susceptibility. Here, the definition of urolithiasis is the same as that used for survival analysis.
The probability of clustering was evaluated in the 72 families comprising Batch 1. Only this batch was used for this analy-sis because detailed breeding records were maintained for this batch, exposure to the contaminated diet was extensive for Batch 1 so the mortality rate was higher, and all six experimental con-ditions (Diet × MeHg) were represented equally.
A three-stage probability model was constructed to determine whether the familial clustering experienced could have occurred by chance. In stage 1, the probability of seeing urolithiasis in a rat was estimated by dividing the number of cases by the origi-nal census (excluding censored cases). After 6 months on the diet, there were 6 cases out of 72 F0 rats, yielding a probability of 0.083 that a rat was diagnosed with urolithiasis. The equivalent probability for the F1 generation was 0.078. After 12 months on diet, these numbers increased to 0.250 and 0.167, respectively. To simplify presentation of this model, the prevalence is assumed to be equal for both generations: 0.08 after 6 months on the diet and 0.18 after 12 months. These are not simple averages of F0 and F1 because the population bases (denominators) are differ-ent. The model also was evaluated assuming generation-specific
prevalence rates, but the outcome did not differ substantively, and the results are not presented here.
In stage 2, the probability of obtaining two or more cases of urolithiasis within a family of size F = L + 1 (L = litter size; L + 1 includes the dam) was estimated using Equation 1 (20), which describes a binomial distribution. Consider the rats on diet for 12 months. This calculation is similar to flipping a biased coin (Pheads = 0.18) F times and obtaining at least two heads. The re-sults are shown in Table 2. If F = 4, then the probability of ob-taining two or more cases of fatal urolithiasis in a single family is 0.16, and it increases to 0.32 for families of size 6. Equation 1 shows how this number was calculated.
where PU is the probability of urolithiasis, PU is the probability that a rat does not develop urolithiasis (= 1 – PU ) , F is the family size (litter size + 1), and U is the number of families affected.
Family size was determined as follows. At weaning, offspring were arbitrarily numbered A, B, C, X1, and X2, depending on the experiment for which they were destined. A litter of three off-spring would only have A, B, and C rats whereas a litter with five offspring would include all five designations. For statistical pur-poses, a small litter could be reconstructed from a large one by omitting the X1 and X2 rats. Therefore, only the rats designated A, B, and C were included when determining the prevalence of urolithiasis in families of size 4 (dam plus three offspring). If an X1 rat developed urolithiasis, it was not included in the analysis of four-member families, but it was included in the population of families of size 5 and 6. Families of sizes 5 and 6 included X1 and X2 rats, respectively.
In stage 3, the probability of obtaining at least U families with two or more cases of urolithiasis out of M families was modeled as a binomial distribution. Therefore, it was viewed as flipping a second biased coin M times and obtaining at least U heads. To il-lustrate, we continue the example above for a family of size 4 and set the probability of obtaining at least two cases at 0.16. Of 43 families, 6 had two or more cases of urolithiasis after 12 months on diet. The probability of this happening (i.e., of seeing at least six such families) by chance alone is 0.72. Equation 2 was used to determine this probability.
where PU,F is the probability of obtaining two or more cases of urolithiasis in a family of size F from Equation 1, U is the num-ber of families with at least two cases of urolithiasis, and M is the number of litters of at least L rats available for evaluation.
Table 2. Cases of urolithiasis within families
Probability of obtaining Probability of Probability of 2 or more Number of families Number of families at least M litters with Months on diet urolithiasis Family size (F) cases in family of size F of size F with ≥ 2 cases (M) ≥ 2 cases of renal disease
6 0.08 4 0.034 43 1 0.78 0.08 5 0.054 37 2 0.61 0.08 6 0.077 27 3 0.35 12 0.18 4 0.16 43 6 0.72 0.18 5 0.24 37 10 0.38 0.18 6 0.32 27 7 0.80
(2)
(1)

359
ResultsOverall mortality rate. Figure 1 shows survival functions
describing mortality associated with urolithiasis and mortality due to all other natural causes (other than colony management) as a function of weeks on a purified diet. Urolithiasis was de-tected within 10 to 20 weeks on the diet, when the F0 generation was 22 to 30 weeks old and the F1 rats were between 13 and 23 weeks old. Mortality was approximately 10% after 1 year and 30% after 2 years on diet. Mortality due to other causes did not begin to appear until after 70 weeks of age. More specific analy-ses are described later.
Of the 64 male rats sent to a different university and main-tained on a standard rodent chow diet for 2 years, no deaths clinically resembled those attributed to urolithiasis (not included in Fig. 1). The overall fatality rate in those rats resembled that typically seen with Long-Evans rats.
Pathology. Lesions included one or more of the following: uro-liths, lower urinary tract obstruction resulting in distension of the urinary bladder and ureters, and hydronephrosis with moderate to severe atrophy of renal parenchyma (Fig. 2). Uroliths were found in the kidneys, ureters, urinary bladder, and urethra and were hard, white to tan, and round or conformed to the structure they occupied. Renal uroliths were 0.2 to 1 cm in diameter (mode, 0.2 to 0.5 cm), whereas those in the ureter were 0.1 to 0.3 cm in diameter, and those in the bladder were 0.2 to 2.0 cm (mode, about 0.5 cm). Those in the urethra were about 0.2 cm in diam-eter. In addition, the urinary tract of some animals also contained
hard particulate matter (< 0.1 mm in diameter). Microscopically, there was evidence of pyelonephritis and dilated collecting ducts and distal and proximal tubules. Histopathology was sometimes difficult to interpret because of postmortem autolysis.
Of 79 cases in which there were clinical signs suggestive of uro-lithiasis and necropsy results were available, uroliths were identi-fied in 70. In five of the remaining nine animals, lesions suggesting previous urinary tract obstruction were seen. These lesions in-cluded variable hydronephrosis and suppurative pyelonephritis.
Four of the 30 necropsies conducted on asymptomatic ani-mals revealed small (2 to 3 mm) uroliths in the kidney or uri-nary bladder. There were no other lesions and no clinical signs in these animals.
Composition of the uroliths and diet. The XRD and FTIR spectra from all 11 uroliths analyzed from three separate insti-tutions were identical. The XRD maxima could not be matched with any International Joint Commission on Powder Diffraction Standards (JCPDS) standards or with any known urinary tract stone component in an extensive database of human urinary tract stone (urolith) diffraction standards. The diffraction pat-tern of the calculi also was compared to the diffraction patterns from the AIN-93 diet and from choline bitartrate supplied by the diet manufacturer (Fig. 3).
The FTIR analysis yielded spectral data representing chemical bonding in the sample molecule(s). FTIR spectral search–match analysis of the urolith samples also did not yield any positive matches with the Medical College of Wisconsin human urinary tract stone library and specifically did not match any calcium phosphate or calcium oxalate spectra. A match of the sample spectra with the Aldrich and Sigma libraries suggested that the uroliths contained a molecule(s) with chemistry similar to glyox-ylic acid and a choline moiety or complex, as shown in Fig. 3A.
Figure 3A shows the spectrum derived from the rat urolith su-perimposed on the spectrum derived from the vendor-supplied choline bitartrate. Figure 3B shows the spectrum of the rat urolith superimposed on the spectrum of glyoxylic acid taken from the Al-drich and Sigma libraries. As shown in Fig. 3A, the spectral maxi-ma in the choline bitartrate at 3492, 3445, 1123, and 699 cm-1 that are associated with various atomic vibrational modes in amine structures are visible in the urolith spectra. These maxima are not present in the glyoxylic acid spectra (Fig. 3B). In addition, maxima in the choline bitartrate spectra at 1719 and 955 cm-1 associated with some vibrational modes in carboxylic acids are found in the stone spectra but not in the glyoxylic acid spectra. In Fig. 3B, spectral maxima found in the glyoxylic spectra at 3204, 1608, 1395, and 616 cm-1 are present in the rat spectra but not in the choline bitartrate spectra. The glyoxylic acid spectra identified as a match in the Aldrich and Sigma spectral libraries is used in this analysis as being representative of a molecule or family of molecules probably associated with the metabolism of the dicarboxylic moiety in choline bitartrate.
The remaining nonmatched spectral maxima in the rats’ uro-lith spectra were matched with the spectral libraries described earlier and with the extensive human kidney stone FTIR spec-tral library of the National Veterans Administration Crystal Identification Center and the associated Molecular Structure Laboratory at the Medical College of Wisconsin and the Zablocki VA Medical Center. The maxima are consistent with macromo-lecular components typically seen in animal and human urine and in urinary tract stones. As the urolith contains metabolic
Figure 1. Survival functions describing mortality associated with urolithiasis (solid circles) and mortality due to all other causes (open circles). Functions are expressed as a function of weeks since the begin-ning of the purified diet. Censoring is not indicated on these graphs. All rats described in Table 1 are included in these curves. Note that urolithiasis-related mortality began after animals had been on the diet for approximately 20 to 30 weeks and continued at a constant rate, as indicated by a constant slope, throughout the duration of the study.
Urolithiasis in rats
Vol 55, No 4Comparative MedicineAugust 2005
360
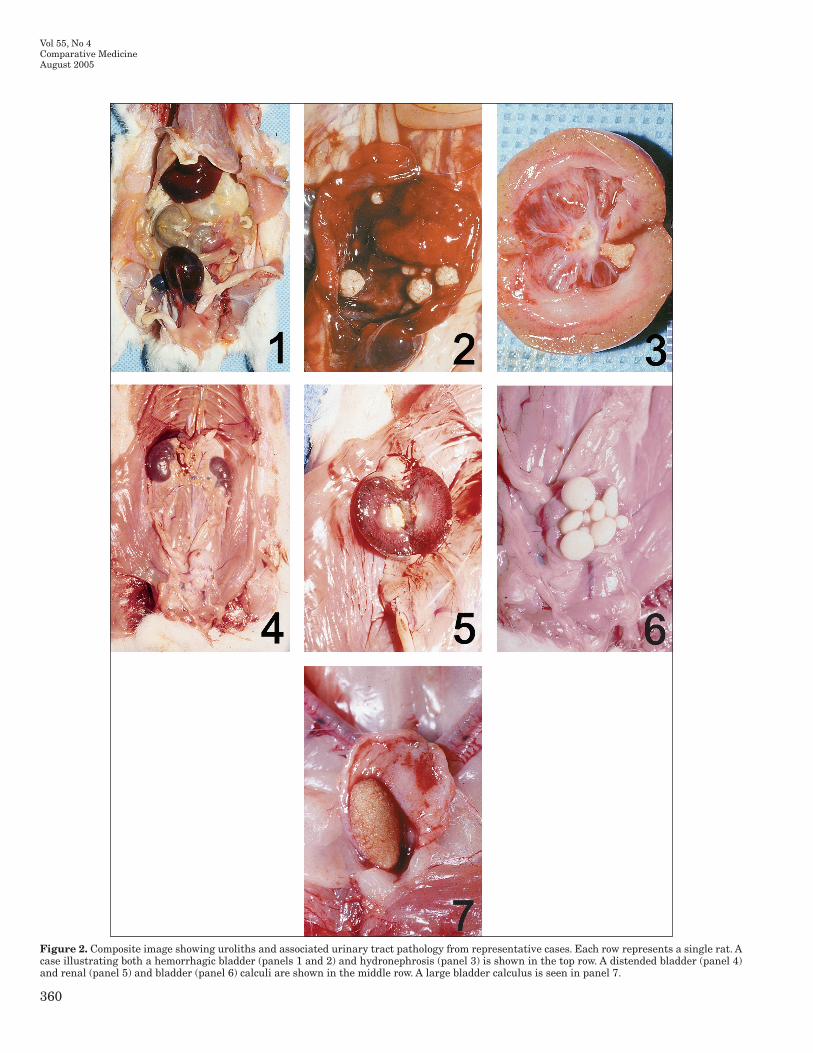
Figure 2. Composite image showing uroliths and associated urinary tract pathology from representative cases. Each row represents a single rat. A case illustrating both a hemorrhagic bladder (panels 1 and 2) and hydronephrosis (panel 3) is shown in the top row. A distended bladder (panel 4) and renal (panel 5) and bladder (panel 6) calculi are shown in the middle row. A large bladder calculus is seen in panel 7.

361
Figure 3. Fourier transform infrared analysis of a rat urolith compared with choline bitartrate (A) and glyoxylic acid (B).
A
B
Urolithiasis in rats

Vol 55, No 4Comparative MedicineAugust 2005
362
products of the originally ingested material, and as the AIN diet contained choline bitartrate, the combined spectral maxima from choline bitartrate and glyoxylic acid indicates that the urolith contains dicarboxylic acid structure(s) and molecule(s) contain-ing a choline moiety. Together, these results demonstrate that the rats’ uroliths were composed of metabolic products of choline bitartrate contained in their AIN-93 diet.
Health surveillance and other potential ingestive or environmental variables. The results of the rodent sentinel animal health surveillance program indicated that the colony had remained free of all pathogens screened, even when sentinel rats were provided food, water, and bedding from deceased rats. Likewise, a rat from the colony showing clinical signs of uroli-thiasis was found to be negative for all infectious agents when analyzed by serology.
Reports issued by the municipal water-works board indicated that water quality was in compliance with federal and state re-quirements. Bacteriological, radiological, inorganic, and organic contaminants were screened, and none was found to exceed mini-mal contaminant levels. No protozoan parasites were found in treated water. Separate analyses of tap water revealed no substan-tive deviations from nominal levels of the minerals tested. Simi-larly, analysis of the diet revealed no noteworthy deviations from nominal concentrations of minerals, mercury, and fatty acids.
Pellets taken from the cages of rats that died and livers from some that were euthanized were analyzed for selenium concen-trations, and no abnormal values were reported.
Humidity was eliminated as a contributor because deaths continued after the humidity system in the animal facility was repaired. Furthermore, no other labs in the affected building reported urolithiasis. Reviews of staffing, laboratory protocols in the colony rooms, and changes in soaps, floor cleaners, and other possible environmental causes failed to reveal a contributing fac-tor. The removal of the nylon chew-bones did not help, and after it became clear that the cause of death lay in the urinary tract, the chew-bones were returned to the rats.
Appearance in other colonies in the U.S. The inquiry on CompMed was especially valuable in eliminating several possi-ble contributing factors and in pointing to the diet as a determi-nant. Responses to this inquiry, which ranged from brief written replies to long and detailed telephone conversations, revealed that urolithiasis occurred in Sprague-Dawley, Long-Evans, and Wistar rats. Furthermore, where sentinel rats were used in these colonies, no infectious processes were identified, as in ours. No vendor was consistently identified as providing the diets; instead, several commercial vendors were represented. Questions about experimental protocols, housing conditions, temperature, gender and humidity revealed no consistent pattern across these colo-nies. Some colonies were on caloric restriction, and others were free-fed. No consistent environmental variable could be identified, except that all used the AIN-93 formulation for at least a month and that the outbreaks had occurred in the previous year or so, suggestive of a recent change in the ingredients of the diet.
Survival analyses. Figure 4 shows the mortality associated with causes other than urolithiasis. As seen in the top panel, mortality was associated with methylmercury concentration for the animals exposed chronically (χ2 = 9.2, P < 0.01). Mortality accelerated considerably after 70 weeks on diet (81 weeks of age) for those animals with 5 ppm of methylmercury in their drink-ing water. Mortality was low and unrelated to methylmercury
Figure 4. Stratified survival analyses of non-urolith-associated deaths in animals exposed chronically (A) to methylmercury (0 ppm, solid circles; 0.5 ppm, open circles; 5.0 ppm, solid triangles) and those exposed prenatally only (B). A high mortality rate was associated with chronic exposure to 5 ppm mercury (as methylmercury) in the drinking water (P < 0.01).
exposure (χ2 = 0.56, P < 0.76) for the animals exposed to methyl-mercury during gestation (bottom panel).
The survival function describing mortality associated with uro-
A
B

363
lithiasis for Batch 2 (Fig. 5A) was significantly (χ2 = 9.2, P < 0.002) shallower, indicating a lower mortality rate, than that for Batch 1. Only two rats in Batch 2 died of urolithiasis. One of these deaths was of an F0 rat that had been on the choline bitartrate diet for 30 weeks. The other was of an F1 rat that had been on the choline bitartrate diet for 16 weeks. Subsequent analyses (not shown) implied that the effect was primarily in the F1 generation. Batch 2’s survival function was shallower than Batch 1’s for each genera-tion, but the P value for the F0 rats alone was only 0.24, whereas that for the F1 generation alone was 0.005 (data not shown).
The survival function for male rats was significantly (χ2 = 24.3, P < 0.0001) steeper than that for female rats (Fig. 5B). Male rats began to die after a shorter time on diet. After nearly 100 days on diet, about 60% of F1 males had died of urolithiasis whereas only slightly more than 20% of females had died.
The F0 rats experienced chronic exposure to methylmercury. For this group, the survival function appears steeper for those exposed to 0.5 ppm of methylmercury than for the other groups (Fig. 5C). The P value was close to traditional levels of signifi-cance (χ2 = 5.4, P = 0.07). An elevated mortality rate in the 5.0-ppm exposure group (Fig. 2) due to high mercury exposure may have masked the appearance of urolithiasis. Prenatal exposure to methylmercury did not influence urolithiasis mortality (Fig. 5D, χ2 = 0.11, P = 0.95). There was no effect attributable to poly-unsaturated fatty acid composition of the diet (χ2 = 0.40, P = 0.53, graphs not shown).
The independent variable “weeks on purified diet” includes weeks on both the choline bitartrate and choline chloride diets. A separate analysis was conducted to examine whether the change in diet diminished the incidence of urolithiasis. To do this, a time variable, “weeks on chloride diet,” was constructed and compared with “weeks on bitartrate diet” by using stratified survival analy-sis. The change in the diet did not decrease the incidence of uro-lithiasis. Graphs are not shown.
Analysis of familial clustering. The probability of obtaining multiple cases of urolithiasis in a single family ranged from 0.07 to 0.32 depending on family size and age (Table 2). Equation 1 was used to derive this number. The probability of seeing two or more cases increased with family size and duration on diet. After 12 months on diet, as many as 25% of litters had experi-enced two or more cases of urolithiasis, but this proportion was well within what is expected by chance. In no analysis did the number of litters with multiple cases exceed that expected by chance. The model that generated these numbers was modified to examine assumptions that entered into simplifying equation 1. Accordingly, the model was re-evaluated using separate preva-lence rates for dams and offspring. It was also re-evaluated in a way that assumed that the probability that an offspring develop-ing urolithiasis was not independent of the probability that the dam was affected. The results are not shown here because these modifications did not substantively change the conclusions in Table 2, although the exact probabilities varied.
The number of litters with zero or one case of urolithiasis was large, and inspection of the distribution suggested that a Poisson distribution might provide a better descriptor than a binomial distribution. Therefore, the probability model also was re-evalu-ated three additional times, once for each family size, by using a Poisson rather than a binomial distribution. The distributions shown in Table 2 fit a Poisson distribution with means of 0.814, 0.97, and 1.19 cases per family for families of size 4, 5, and 6,
respectively. The resulting Poisson distribution fit the obtained values with R2 > 0.99 for each family size, and the P value for χ2 goodness-of-fit tests ranged from 0.82 to 0.96. Therefore, the number of cases of urolithiasis per litter was fit with a simple counting distribution very precisely. The use of a Poisson rather than a binomial process also did not alter the conclusion that the rate of familial clustering experienced in this study could occur by chance.
Discussion Urolithiasis is a significant clinical problem in human and
veterinary medicine (2, 3, 19, 27, 28, 37), but the role of dietary racemic bitartaric acid in this disease is only now becoming ap-preciated. Levo tartaric acid is a natural and soluble form found in many foods and certain wines. This l isomer is excreted by the kidney and appears to be biologically inert, but the racemic version (dl) accumulates in the kidney and induces structural changes consistent with crystalluria, a precursor of urolithiasis (4, 6, 10). As evidenced in the outbreak described here, racemic bitartaric acid is a potent inducer of urolithiasis. By the time breeder (F0) rats in the present study were 28 months old and their offspring (F1 rats) were 16 to 19 months old, the mortality rate was nearly 30%.
Pathology. The most important clinical signs of urolithia-sis were hematuria and the presence of urine on the abdominal fur, usually associated with listlessness. Urinary tract lesions and the presence of calculi seen at necropsy usually confirmed a tentative diagnosis of urolithiasis based on clinical presenta-tion. The number and size of uroliths in the present study were highly variable. In some animals, numerous small uroliths filled the bladder, ureters, or renal pelvises. In other animals, severe hydronephrosis was associated with extensive destruction of the renal pelvis, medulla, and cortex, sometimes without obvious uro-liths. These changes could have been due to previous obstruction in the lower urinary tract, ascending pyelonephritis, or both.
Partial to complete obstruction of urinary outflow can cause mild to severe pressure necrosis of the renal pelvis, medulla, and eventually the cortex. In addition, urinary calculi can inflame and cause degeneration and necrosis of the epithelial lining of the urinary tract. Incomplete emptying of the urinary system due to obstruction, coupled with the loss of epithelial integrity, allows bacterial overgrowth and subsequently an ascending uri-nary tract infection. In the case of a severe infection, bacteria can gain access to systemic circulation and cause sepsis. These fac-tors, taken together, make it clear that once potentially obstruc-tive uroliths form, the future health of the rat is at considerable risk, perhaps irreversibly, because the calculi are highly persis-tent. This situation poses a special problem in long-term studies because an asymptomatic rat can become ill suddenly and die, thereby jeopardizing the study.
Composition of the uroliths. The x-ray diffraction pat-terns did not reveal a match between the molecular structure of uroliths in this study and documented XRD patterns, but the FTIR analysis was more revealing. This finding is not surprising because crystallographic XRD analysis is based on the diffrac-tion of x-rays by the atoms in a crystal structure. Any change in a molecule’s chemical composition, as might occur due to the metabolism of the molecule, would result in a molecular crystal structure different from that of the parent molecule. Commercial diffraction libraries do not often contain patterns for metabolites
Urolithiasis in rats

Vol 55, No 4Comparative MedicineAugust 2005
364
Figure 5. Stratified survival analyses of urolithiasis associated with batch (a proxy for duration of exposure to the contaminated diet; Batch 1, solid circles; Batch 2, open circles), gender (female rats, solid circles; male rats, open circles), and methylmercury exposure. There was a significant effect of batch (P < 0.002) and of gender (P < 0.0001) and a marginally significant effect of chronic methylmercury exposure (P = 0.07).
because there are often many metabolic products for any biologi-cally active compound. Therefore, while a match between the dif-fraction pattern of a urolith and that of choline bitartrate would
be compelling evidence of a connection between urolithiasis and the ingestion of choline bitartrate, a mismatch does not rule one out. Instead, it shows that choline bitartrate does not maintain
A
C
B
D

365
its unique chemistry as it is processed in vivo.The infrared pattern from the FTIR was especially informative
because it provides data on the major chemistry in the sample; the process of metabolism usually maintains large portions of the chemical structure of the parent molecule. A failure to find a positive match between the FTIR spectra from the rats’ uroliths and spectra of any compound contained in the human urinary tract stone library at the Medical College of Wisconsin was es-pecially interesting. In particular, the spectra did not match any calcium phosphate or calcium oxalate standard spectra, eliminat-ing these common forms of urolithiasis as playing a role here.
A search–match analysis of the sample spectra from the rats’ uroliths with commercially available Aldrich and Sigma infra-red spectral libraries, however, did implicate molecule(s) with chemistry similar to that of glyoxylic acid as well as molecule(s) containing a choline moiety. In particular, the spectral absorption maxima from the uroliths matched both the spectrum for choline bitartrate supplied by a feed vendor and glyoxylic acid. The pres-ence of spectral maxima indicative of choline bitartrate and gly-oxylic acid indicates a dicarboxylic acid structure with spectral maxima representing the choline moiety as well. These results are consistent with the hypothesis that urolithiasis resulted from racemic choline bitartrate. Especially interesting is that these results also raise the possibility that bitartrate-induced urolithiasis could be misdiagnosed as being calcium acid-based.
The maxima in the remaining nonmatched portion of the spec-tra also were consistent with spectra from macromolecular com-ponents normally seen in diets and in animal and human urine and uroliths. Therefore, the calculi contained metabolic products of the originally ingested material as well as the choline bitar-taric acid that had been added to purified diets.
Spectra taken from uroliths contributed from several institu-tions matched each other and matched the choline bitartaric acid provided by a different source vendor than the one in the present study. These results offer strong evidence that the calculi were composed of metabolic products of choline bitartaric acid and suggest that a common source of this racemic bitartaric acid was used by multiple vendors.
The role of diet and irreversibility of urolithiasis. Uro-lithiasis-associated mortality first appeared in a female breeder 20 weeks after starting the AIN-93 based diet containing cho-line bitartrate, but it is not known when the racemic form first appeared or how long it was present. Mortality continued at a constant rate even after the diet was changed 6 to 8 months later to one containing choline chloride. Although this observation might imply that the form of choline (bitartrate versus chloride) was irrelevant to the formation of uroliths, it is more likely that fatalities after the change to choline chloride were due to previ-ously formed uroliths. Therefore, once formed, calculi continue to reside in the urinary tract where they can remain benign or eventually become pathogenic.
Evidence that a rat must remain exposed to the impurity for a minimal duration derives from rats in the second batch. Only two deaths due to urolithiasis occurred in this batch, and these occurred in rats born that had been on the bitartrate diet longer than others in that cohort.
A third group of rats, comprising littermates of the rats de-scribed here that were sent to a colleague at another university for a related study, was maintained on a standard rodent chow diet. A fourth group of rats comprised littermates used in other
experiments and housed in the same rooms as rats described here but maintained on a standard rodent chow diet. No animal on the standard chow diet died of urolithiasis. Postmortem ex-amination of some of these rats failed to show any evidence of urinary tract disease. Yet another group of rats has been main-tained on an AIN-93-based diet with choline chloride for about 58 weeks as of this writing, and none has shown signs of urinary tract disease. Finally, no other colonies on campus (all fed chow diets) experienced this problem. These different groups and other reports (13, 15) offer evidence that an impurity, racemic bitar-taric acid, in the AIN-93 purified diet provided during late 2001 and early 2002 was the cause of the urolithiasis.
Chronic methylmercury exposure. Stratified survival analyses indicated that chronic methylmercury exposure in-creases risk of urolithiasis, but the marginal P value of 0.07 and the absence of a dose–effect relationship suggest that this con-clusion should be offered cautiously. Nevertheless, there is good evidence that this effect is not a statistical artifact but a real phenomenon. The effect of methylmercury was seen only in the rats exposed chronically to methylmercury, and it is those rats that would be expected to have a compromised renal system. During methylmercury exposure, the kidney accumulates meth-ylmercury more so than do other organs (12, 32), and this pro-pensity affects renal function (5). Although tubular necrosis has been linked to developmental methylmercury exposure (33, 34), this abnormality occurred at much higher exposure levels than those used here, and it is unclear whether this condition would predispose an animal to urolithiasis. Such predisposition did not occur with the dosing regimen used in the present study.
The animals exposed chronically to high (5.0 ppm) methylmer-cury may not have shown the highest prevalence of urolithiasis because they had less opportunity to develop it. That is, elevated mortality due to other causes may have precluded the appear-ance of urolithiasis in these animals. These conditions should be replicated before a firm conclusion is drawn, but the evidence remains good that chronic methylmercury exposure elevates the risk of urolithiasis when the contributing factor such as racemic tartaric acid is also present.
The stratified survival analysis tests the assumption that the survival functions are proportional to one another, and in at-tributing a role to methylmercury exposure this assumption was rejected. Therefore, the elevation in risk due to chronic meth-ylmercury exposure cannot be described by a single odds ratio. Instead, the longer the animals were exposed to the methylmer-cury, the greater were the odds of dying of urolithiasis.
Selenium and other dietary components. In a study sponsored by the National Toxicology Program (NTP), high concentrations of selenium in the drinking water of Fisher rats increased the prevalence of papillary degeneration, decreased urine volume and increased hematocrit (1). Some clinical signs resembling those associated with urolithiasis, including poor grooming, ruffled fur, and weight loss also were described. The illness, however, was associated with extremely high selenium concentrations, more than 10 times those used in the present study and high enough to suppress fluid intake; the renal dam-age was attributed to dehydration.
The NTP study also reported mineralization in the kidney and a small number of cases of urolithiasis, but neither occurred in a dose-related fashion. Nevertheless, this report raised suspicion that selenium might be a contributor. For example, the feed may
Urolithiasis in rats

Vol 55, No 4Comparative MedicineAugust 2005
366
have contained selenium “hot spots” due to poor mixing, result-ing in excessive exposure of some animals. However, neither the livers of rats that died nor food pellets taken from some deceased animals’ cages revealed abnormal concentrations of selenium. Therefore, selenium is unlikely to be a contributing factor to uro-lithiasis in this colony.
Other dietary factors also do not seem to be applicable. No renal disease was reported in Sprague-Dawley rats consuming an AIN 93 diet for 114 weeks (17). Some nephropathy appeared in Sprague-Dawley rats fed a cereal-based NIH-31 diet in 110-week-old rats, but caloric restriction, as was practiced in the present study, reduced its incidence (8).
Gender differences. Male rats were considerably more at risk for urolithiasis than were females. Urolithiasis appeared sooner in male offspring than in female offspring, beginning after only 5 to 10 weeks on the diet. As the rats aged, the discrepancy in risk between males and females grew larger; by 100 weeks, nearly 60% of male rats died of urolithiasis, three times the prevalence seen in female rats. Postmortem analyses suggested that males were more likely to have bladder calculi than were fe-males, who usually formed calculi in the kidney, but the number of cases here is small.
In other reports, male cats (37), dogs (2), and rats (16) showed higher rates of urolithiasis than did female animals. Gender dif-ferences in risk of urolithiasis have been described in humans, depending on the urolith composition and the underlying etiol-ogy of urolithiasis. For example, men are 2.6 times more likely to develop calcium oxalate uroliths than are women (11). It is generally observed that male animals have a higher incidence of urolithiasis than do female animals because the long male ure-thra has a very narrow lumen. Therefore, calculi become lodged there more easily. Female animals show a higher potential for infection associated with uroliths containing magnesium am-monium phosphate, struvite, or ammonium-splitting bacteria in the urinary tract. Other reported forms of urolithiasis do not show gender differences.
Family and strain differences. Multiple cases of urolithiasis sometimes occurred in a family comprising a dam and offspring. In order to determine the likelihood that clustering occurred by chance, a three-stage probability model was constructed based on the observed frequency of urolithiasis in the population of F1 and F0 rats and on family size. The number of affected families was well within the range of what would be expected by chance.
Urolithiasis associated with the contaminated AIN-93 diet was reported in Long-Evans, Wistar, and Sprague-Dawley rats. The prevalence could not be determined in these other studies, so it is not clear at this point whether these strains are differentially sus-ceptible. In all cases, however, the prevalence was high enough to alarm investigators who, in some cases, began studies anew with new rats. Therefore, all risk factors identified (apart from gender) were environmental, but it is unclear why some rats sharing risk factors developed urolithiasis and others did not.
Other environmental variables or infectious processes. The possibility of an infectious process from several sources was examined. One possible source was the water bottle. To minimize the handling of methylmercury by staff and the amount of haz-ardous waste produced, water bottles were emptied and washed in the high-temperature cage washer less frequently than they would have been otherwise. To examine potential infectious pro-cesses, sentinel rats were exposed to water bottles, including
unwashed stoppers, and soiled bedding from rats residing in the colony. No evidence of infectious or noninfectious disease was seen in these rats.
Temperature, humidity, staffing, municipal water, minerals in food, methylmercury, fatty acid composition of the diets, infec-tious processes, nylon chew-bones, and several other potential contributors were unlikely to have been related to mortality. The posting on the CompMed listserver also provided no evidence that the various environmental measures listed were of concern. In addition, strain, feed vendor, food restriction, and age were not associated with urolithiasis. The single consistent element among all of the reports was the use of a purified diet based on the AIN-93 formulation that was obtained within a fairly narrow time window. Only in retrospect, knowing that urolithiasis due to racemic tartaric acid in the diet is the probable cause of lethality, would some of these possibilities seem unlikely a priori.
The analyses described in this report are rather complex, but the conclusion is simple: an impurity in purified diets supplied to many laboratories during 2001 and 2002 resulted in an alarm-ingly high rate of fatal urolithiasis in rats. This impurity is likely racemic bitartaric acid used to stabilize choline. It is not known how widespread this problem is in human or animal veterinary medicine, but the potency of racemic bitartaric acid appears high. The difficulty in matching the spectra from the stones with spectra from any known library indicates that if such a problem has existed before, it might have gone undocumented or even misdiagnosed as due to glyoxylic acid.
Symptomatic urolithiasis appeared in female rats that were on the diet for 20 to 30 weeks. Males were far more sensitive and required less time on the diet to show effects. Although it is likely that some calculi successfully exited the urinary tract, many persisted and continued to threaten the life of the animal. In fact, the probability of suffering fatal urolithiasis remained constant from 20–30 weeks to 100 weeks in female rats and from 10 weeks to 90 weeks in male rats. There was evidence that a re-nal system compromised by chronic exposure to methylmercury is more susceptible to the development of urolithiasis, but there was no evidence that susceptibility ran in families.
AcknowledgmentsThis research was supported by ES 10865 from the National Institute
of Environmental Health Sciences awarded to M. C. Newland. Analysis of the urolith composition was supported in part by a Veterans Admin-istration Research Career Scientist award to NM as well as National Institutes of Health awards DK030579, DK062739, and DK064616. XRD and FTIR analyses were conducted within the National Veterans Administration Crystal Identification Laboratories.
We thank the many people, including feed vendors, individual inves-tigators, and laboratory animal veterinarians, who provided detailed information regarding environmental conditions and dietary formula-tions. Appreciation also goes to Philip Reeves (Grand Forks Human Nutrition Center, N. Dak.) for helpful conversations and for making us aware of the early literature on tartaric acid.
References 1. Abdo, K. M. 1994. Sodium selenate and sodium selenite: admin-
istered in drinking water to F344/N rats and B6C3F1 mice. NIH publication 94-3387. The National Institutes of Health, Bethesda, Md.
2. Brown, N. O., J. L. Parks, and R. W. Greene. 1977. Canine urolithiasis: retrospective analysis of 438 cases. J. Am. Vet. Med. Assoc. 110:414-418.

367
3. Brown, N. O., J. L. Parks, and R. W. Greene. 1977. Recurrence of canine urolithiasis. J. Am. Vet. Med. Assoc. 170:419-422.
4. Chasseaud, L. F., W. H. Down, and D. Kirkpatrick. 1977. Absorption and biotransformation of L(+)-tartaric acid in rats. Experientia 33:998-999.
5. Clarkson, T. W. 1972. The pharmacology of mercury compounds. Ann. Rev. Pharmacol. 12:375-406.
6. Down, W. H., R. M. Sacharin, L. F. Chasseaud, D. Kirkpatrick, and E. R. Franklin. 1977. Renal and bone uptake of tartaric acid in rats: comparison of l(+) and dl-forms. Toxicology 8:333-346.
7. Duffy, P. H., S. M. Lewis, M. A. Mayhugh, A. Mccracken, B. T. Thorn, P. G. Reeves, S. A. Blakely, D. A. Casciano, and R. J. Feuers. 2002. Effect of the AIN-93M purified diet and dietary restriction on survival in Sprague-Dawley rats: implications for chronic studies. J. Nutr. 132:101-107.
8. Duffy, P. H., S. M. Lewis, M. A. Mayhugh, R. W. Trotter, B. T. Thorn, R. J. Feuers, and A. Turturro. 2004. The effects of differ-ent levels of dietary restriction on non-neoplastic diseases in male Sprague-Dawley rats. Aging Clin. Experiment. Res. 16:68-78.
9. Duffy, P. H., J. E. Seng, S. M. Lewis, M. A. Mayhugh, A. Aidoo, D. G. Hattan, D. A. Casciano, and R. J. Feuers. 2001. The ef-fects of different levels of dietary restriction on aging and survival in the Sprague-Dawley rat: implications for chronic studies [see comment]. Aging Clin. Experiment. Res. 13:263-272.
10. Finkle, P. 1933. The fate of tartaric acid in the body. J. Biol. Chem. 6:349-355.
11. Gault, M. H. and L. Chafe. 2000. Relationship of frequency, age, sex, stone weight and composition in 15,624 stones: comparison of results for 1980 to 1983 and 1995 to 1998. J. Urol. 164:302-307.
12. Kakita, A., C. Inenaga, M. Sakamoto, and H. Takahashi. 2003. Disruption of postnatal progenitor migration and consequent abnormal pattern of glial distribution in the cerebrum following administration of methylmercury. J. Neuropathol. Experiment. Neurol. 62:835-847.
13. Kankesan, J., R. Vanama, R. Renlund, J. J. Thiessen, V. Ling, P. Rao, S. Rajalakshmi, and D. S. R. Sarma. 2003. Source of a micro-nutrient in a semi-synthetic basal diet as a causative factor in inducing urinary calculi in rats and its inhibition by PSC 833, a potent inhibitor of p-glycoprotein. Comp. Med. 53:444-447.
14. Kleinbaum, D. K. and M. Klein. 1996. Survival analysis. SpringerSpringer Verlag, New York.
15. Klurfeld, D. M. 2002. Kidney and bladder stones in rodents fed purified diets. J. Nutr. 132:3784.
16. Lee, Y. H., W. C. Huang, H. Chiang, M. T. Chen, J. K. Huang, and L. S. Chang. 1992. Determinant role of testosterone in the pathogenesis of urolithiasis in rats. J. Urol. 147:1134-1138.
17. Lewis, S. M., Z. J. Johnson, M. A. Mayhugh, and P. H. Duffy. 2003. Nutrient intake and growth characteristics of male Sprague-Dawley rats fed AIN-93M purified diet or NIH-31 natural-ingredi-ent diet in a chronic two-year study. Aging Clin. Experiment. Res. 15:460-468.
18. Lulich, J. P., C. A. Osborne, C. Lekcharoensuk, T. A. Allen, and Y. Nakagawa. 1999. Canine calcium oxalate urolithiasis. Case-based applications of therapeutic principles. Vet. Clin. North Am. Small Anim. Pract. 29:123-139.
19. Mandel, N. 1996. Mechanism of stone formation. Semin. Nephrol. 16:364-374.
20. Meyer, P. L. 1970. Introductory probability and statistical ap-plications. Addison-Wesley, Reading, Mass.
21. National Research Council. 1995. Nutrient requirements of laboratory animals. National Academy Press, Washington, D.C.
22. Newland, M. C. 1997. Quantifying the molecular structure of behavior: separate effects of caffeine, cocaine, and adenosine agonists on interresponse times and lever-press durations. Behav. Pharmacol. 8:1-16.
23. Newland, M. C. and K. W. Brown. 1998. Behavioral characte-rization of adenosine agonists after chronic caffeine exopsure: separate effects on response rates and lever-press durations. Behav. Pharmacol. 8:17-30.
24. Newland, M. C., W. W. Ng, R. B. Baggs, G. D. Gentry, B. Weiss, and R. K. Miller. 1986. Operant behavior in transition reflects neonatal exposure to cadmium. Teratology 34:231-242.
25. Newland, M. C. and E. B. Rasmussen. 2000. Aging unmasks adverse effects of gestational exposure to methylmercury in rats. Neurotoxicol. Teratol. 22:819-828.
26. Newland, M. C. and P. A. Reile. 1999. Blood and brain mercury levels after chronic gestational exposure to methylmercury in rats. Toxicol. Sci. 50:106-116.
27. Pak, C. Y. 1991. Etiology and treatment of urolithiasis. Am. J. Kidney Dis. 18:624-637.
28. Pak, C. Y. and M. I. Resnick. 2000. Medical therapy and new approaches to management of urolithiasis. Urolog. Clin. North Am. 27:243-253.
29. Rasmussen, E. B. and M. C. Newland. 2001. Developmental exposure to methylmercury alters behavioral sensitivity to d amp-hetamine and pentobarbital in adult rats. Neurotoxicol. Teratol. 23:45-55.
30. Reeves, P. G. 1997. Components of the AIN-93 diets as improve-ments in the AIN-76A diet. J. Nutr. 127:838S-841S.
31. Reeves, P. G., F. H. Nielsen, and G. C. Fahey, Jr. 1993. AIN-93 purified diets for laboratory rodents: final report of the American Institute of Nutrition ad hoc writing committee on the reformula-tion of the AIN-76A rodent diet. J. Nutr. 123:1939-1951.
32. Sakamoto, M., N. Ikegami, and A. Nakano. 1996. Protective effects of Ca2+ channel blockers against methyl mercury toxicity. Pharmacol. Toxicol. 78:193-199.
33. Sakamoto, M., A. Kakita, K. Wakabayashi, H. Takahashi, A. Nakano, and H. Akagi. 2002. Evaluation of changes in methyl-mercury accumulation in the developing rat brain and its effects: a study with consecutive and moderate dose exposure throughout gestation and lactation periods. Brain Res. 949:51-59.
34. Sakamoto, M. and A. Nakano. 1995. Comparison of mercury accumulation among the brain, liver, kidney, and the brain regions of rats administered methylmercury in various phases of postnatal development. Bull. Environ. Contam. Toxicol. 55:588-596.
35. Smith, L. H. 1990. The pathophysiology and medical treatment of urolithiasis. Semin. Nephrol. 10:31-52.
36. Stern, S., C. Cox, E. Cernichiari, M. Balys, and B. Weiss. 2001. Perinatal and lifetime exposure to methylmercury in the mouse: blood and brain concentrations of mercury to 26 months of age. Neurotoxicology 22:467-477.
37. Willeberg, P. and W. A. Priester. 1976. Feline urological syn-drome: associations with some time, space, and individual patient factors. Am. J. Vet. Res. 37:975-978.
38. Yasui, T., M. Sato, K. Fujita, K. Tozawa, S. Nomura, and K. Kohri. 2001. Effects of citrate on renal stone formation and os-teopontin expression in a rat urolithiasis model. Urol. Res. 29:50-56.
Urolithiasis in rats