the preparation of hemoglobin in a dry … · the preparation of hemoglobin in a dry and ......
TRANSCRIPT

THE PREPARATION OF HEMOGLOBIN IN A DRY AND ACTIVE STATE*
BY DEMPSIE B. MORRISON AND ALAN HISEY
(From the Department of Chemistry, University of Tennessee School of Biological Sciences, Memphis)
(Received for publication, December 7, 1936)
It is well known that crystals of oxyhemoglobin are changed by drying into a form which retains little or no activity with oxygen. Bohr (1) has called the inactive material /?-hemoglobin, and Huf- ner (2) has shown it to be a mixture of methemoglobin with vari- able small amounts of oxyhemoglobin. Heidelberger (3), describ- ing his method of crystallizing oxyhemoglobin, cautions against allowing the crystals to become dry if loss in activity is to be avoided. Van Slyke, Hastings, Heidelberger, and Neil1 (4) state, “If dried, even at high vacuum and in the cold, hemoglobin is almost completely inactivated, although in solubility and color it still resembles oxyhemoglobin.” Barcroft (5), referring to his own experience with it and that of Bayliss and Parsons, calls such dried material inactive hemoglobin.
In our initial unsuccessful attempts (which included the use of various organic solvents) to improve the activity of dried oxy- hemoglobin we noted that methemoglobin formation could be largely if not completely avoided by reducing the hemoglobin before drying. We are now able to define conditions for preparing reduced hemoglobin in a dried and stable condition without sig- nificant loss of oxygen capacity.
EXPERIMENTAL
Hemoglobin Xolutions and Crystals-Oxalated or defibrinated blood of the dog, pig, ox, or human is centrifuged, the plasma
* A preliminary report was made before the American Society of Bio- logical Chemists (Proc. Am. Sot. Biol. Chem., 8, lxxiii (1936);J. Biol. Chem., 114 (1936)).
693
by guest on August 2, 2018
http://ww
w.jbc.org/
Dow
nloaded from

694 Dried Hemoglobin
removed as closely as possible, and the cells washed in the centri- fuge four or five times with cold 0.9 per cent NaCl. The washed cells are then treated in one of the following ways: (a) chilled in an ice bath and an equal volume of ice-cold distilled water added (the mixture is then extracted, with centrifuging, three times with ice-cold C.P. anhydrous ether1 to complete laking and remove stroma) ; (b) diluted, laked with alumina cream, and centrifuged to remove stroma; (c) after addition of sufficient cold distilled water to lake completely, centrifuged at high speed to remove any debris; (d) diluted with 0.9 per cent NaCl without laking; (e) as in (c), followed by dialysis against distilled water in the cold room.
Crystals of dog hemoglobin were prepared by a procedure which we have described (6).
There is no indication that the method of preparing the hemo- globin solution has any effect upon the activity of the dried residue provided the preliminary treatment does not in itself cause in- activation.
Method of Drying-Drying is accomplished by vacuum distilla- tion at 38”. The apparatus consists of a glass drying chamber connected by pressure tubing through a s-way stop-cock to a flask containing concentrated H&30*, which, in turn, is connected through a mercury manometer to a Cenco Hyvac pump. The in- let tube to the flask extends below the surface of the acid to facili- tate absorption of water vapor. For small samples of hemoglobin, up to 40 ml. of solution, the drying chamber is a 1 liter round bottom Pyrex flask with ground-in stop-cock. For larger samples, up to 200 ml., a large flat bottom desiccator is used.
In drying small samples, the hemoglobin solution is placed in the drying chamber, the system closed, and the pump started. Initial foaming is controlled through stop-cock regulation by allowing the pressure in the drying chamber to fall slowly. As reduction proceeds foaming diminishes, and pressure in the cham- ber is then allowed to fall to that of the rest of the system. The chamber is shaken vigorously by hand to assure as complete reduc- tion of the hemoglobin as possible. The sample, which is cold
1 Anhydrous ether was used, as it contained the least amount of impuri- ties of any grade of ether we were able to purchase. Most grades of ether contain impurities which convert hemoglobin to methemoglobin.
by guest on August 2, 2018
http://ww
w.jbc.org/
Dow
nloaded from

D. R. Morrison and A. Hisey 695
when placed in the chamber, is kept cold during the reduction of the hemoglobin by the rapid evaporation of the water, thus mini- mizing methemoglobin formation at this most critical stage. The chamber is now placed in a water bath at 38”, and the acid- absorbing flask in an adjoining bath at room temperature. Dis- tillation proceeds rapidly and the final pressure is too low to be measured accurately on our manometer. Both chamber and flask are rocked by a motor-driven mechanism. Thus the hemo- globin solution spreads and dries in a thin film, while agitation of the acid in the absorbing flask increases its efficiency.
If crystals are to be dried, they must be suspended in sufficient water to dissolve completely as they are reduced, before appre- ciable drying occurs.
When larger samples are to be dried, the desiccator is connected while empty, rocking is started, and the whole system is evacuated. The hemoglobin solution is then slowly admitted to the evacuated chamber through a capillary tube whose tip is drawn to a very blunt point to prevent clogging by the accumulation of dried hemoglobin on the outside of the capillary. The hemoglobin solution enters the chamber as a fine, hair-like stream which strikes the bottom with considerable velocity and spatters to the sides of the chamber. Reduction is practically instantaneous, foaming scarcely occurs, and the small droplets dry almost at once. However, the drying capacity of our system is not suffi- cient to prevent the accumulation of some liquid and a consider- ably longer time is required to dry the larger samples under these conditions. We have not been able to get quite as high activity or uniform drying as with the smaller samples.
In either procedure drying is continued until flakes of dry hemoglobin begin to pull away from the sides of the chamber. If the chamber is now removed from the bath and struck a sharp blow with the hand, a considerable amount of the hemoglobin is de- tached from the walls. The time required for drying is about 20 minutes for the smaller samples and up to 3 hours for the largest samples.
The dried hemoglobin is handled in one of two ways, with effects upon properties which will be discussed later. (a) The sample is removed from the drying chamber and ground to a powder in a mortar without protection from oxygen of the air. Portions may
by guest on August 2, 2018
http://ww
w.jbc.org/
Dow
nloaded from

696 Dried Hemoglobin
be dissolved immediately and analyzed, and other portions sealed in vacua in glass ampules. (b) An alternative procedure, which gives consistently higher activities of the dried hemoglobin, is to connect the drying chamber by pressure tubing to a tonometer containing somewhat more distilled water than will be used to dissolve the sample. The tonometer, in turn, is attached to a vacuum line and the water completely deaerated. As the tonome- ter is shaken to facilitate removal of air, evaporation of water causes it to become quite cool. During this evacuation water travels back to the closed stop-cock of the drying chamber in an unbroken column. The desired quantity of water is now admitted to the chamber and, with shaking, the sample dissolves rapidly, forming a solution of reduced hemoglobin. So little gas is present that the solution gives a metallic click and practically no foam when shaken. Air is now admitted and the hemoglobin is readily converted on shaking into oxyhemoglobin.
Analytical Methods
All gasometric analyses were made in the Van Slyke mano- metric blood apparatus. Total pigment was determined by the method of Van Slyke and Hiller (7) or spectrophotometrically by the procedure of Drabkin and Austin (8), or by both methods, and frequently by the use of undried control samples. After our samples are dissolved and oxygenated, the combined experimental error in oxygen capacity and total pigment estimations does not exceed fl per cent. Data on Samples 130-b and 135-b (Table II) are representative of repeated checks upon our experimental error. Approximately the first one-fourth of the samples was analyzed for active hemoglobin by the carbon monoxide capacity method of Van Slyke and Hiller (9), and the remainder by the oxygen capacity method of Van Slyke and Neil1 (10). A few samples were determined by both methods with excellent agree- ment in results. A number of samples analyzed by the oxygen capacity method were also analyzed with the spectrophotometer (Bausch and Lomb), with a modification of the procedure of Aus- tin and Drabkin (11).
In the spectrophotometric analysis of a mixture of oxyhemo- globin and methemoglobin as outlined by Austin and Drabkin, three values or sets of values are required: the extinction coeffi-
by guest on August 2, 2018
http://ww
w.jbc.org/
Dow
nloaded from

D. B. Morrison and A. Hisey 697
cients of the mixture at X 630,575,560, and 540 rnp; the pH of the mixture; the concentration of total pigment. The modification of their procedure which we have employed eliminates determina- tion of the pH and is, we believe, slightly more accurate. Three aliquots of the mixture to be analyzed are pipetted into volu- metric flasks of the same capacity. The sample in Flask 1 is diluted, 5 ml. of phosphate buffer of pH 7.38 are added, and the mixture made to volume with distilled water. The other two samples are diluted and quantitatively converted to methemo- globin by potassium ferricyanide. To the mixture in Flask 2 are now added 5 ml. of the buffer, pH 7.38, and sufficient distilled water to bring to volume. Potassium cyanide is added to Flask 3 to form cyanmethemoglobin, and the mixture is diluted to volume with water. Concentrations in all cases are those given by Austin and Drabkin (11). Total pigment is determined in Solution 3. Extinction coefficients at X 630, 575, 560, and 540 rnp are measured for Solutions 1 and 2, which have the same pH and the same ionic strength, except for the effect of the small amount of ferricyanide in Solution 2. Calculations are made by the method of Austin and Drabkin (11). Agreement between values for active pigment by the oxygen capacity method and the spectrophotometric method was good. Obviously, the experi- mental error in the spectrophotometric method increases rapidly when one component of a mixture is present in disproportionately small amount.
Residual moisture of the samples was obtained by drying to constant weight in air at 105”.
Properties of Dried Hemoglobin
Dried hemoglobin appears bright red in color when thin films are viewed by transmitted light. It has a darker red color when powdered. It is freely soluble in distilled water, physiological saline, and phosphate buffers. While no attempt has been made to determine the maximum solubility of our preparations, we have frequently made solutions which had an oxygen capacity up to 14 volumes per cent.
In solutions of the dried hemoglobin there is a small amount of difficultly soluble material which, however, is present only in traces when crystalline hemoglobin is dried. That this material is not
by guest on August 2, 2018
http://ww
w.jbc.org/
Dow
nloaded from

698 Dried Hemoglobin
hemoglobin or a hemoglobin derivative is indicated by the follow- ing facts. Its removal from the solution by centrifuging does not affect the values for total pigment. When separated and sus- pended in 0.9 per cent NaCl, it dissolves slowly, forming a solution which has the appearance of plasma from slightly hemolyzed blood. When plasma is dried by our procedure, the proteins
=lmt#!L
620 600 560 560 540 520 50 A in mp
FIG. 1. Spectrophotometric curves of dried hemoglobin dissolved In the presence of air. Curve 1, pig hemoglobin, 91.2 per cent active; Curve 2, ox hemoglobin, 79.2 per cent active; Curve 3, ox hemoglobin, 96.0 per cent active; Curve 4, pig hemoglobin, 92.1 per cent active.
redissolve rather slowly, behaving as does the residue from our hemoglobin samples. We conclude, therefore, that this residue is plasma proteins which are not completely removed even by five washings of the erythrocytes with 0.9 per cent NaCl.
When the dried samples are dissolved without exposure to air, the solutions have the characteristic color of reduced hemoglobin; when air is admitted, the color changes to the bright red of oxy-
by guest on August 2, 2018
http://ww
w.jbc.org/
Dow
nloaded from

D. B. Morrison and A. Hisey 699
hemoglobin. The solutions may be reduced and oxygenated repeatedly by alternate use of tacuum and admission of air. In solution the pigment’ is readily converted by ferricyanide to methemoglobin and the latter, by cyanide, to cyanmethemoglobin.
That the hemoglobin has not been appreciably altered by drying is indicated by the following facts: (a) In solution it combines re-
03 620 580 540 501
h in mp
FIG. 2. Spectrophotometric curves of Sample 93. Curve 1 from Ampule 93-c, activity 82.1 per cent; Curve 2, Ampule 93-e, activity 80.1 per cent; Curve 3, Ampule 93-f, activity 85.9 per cent; Curves 4, 5, and 6, samples from these same ampules exposed to air for varying periods of time before analysis.
versibly with oxygen and with carbon monoxide; (b) crystals of oxyhemoglobin which appear to be identical with those obtained from fresh dog blood may be obtained by usual methods from solutions of dried dog hemoglobin; (c) Figs. 1 and 2, which show spectrophotometric curves of solutions of dried hemoglobin, are typical of mixtures of oxyhemoglobin and methemoglobin.
by guest on August 2, 2018
http://ww
w.jbc.org/
Dow
nloaded from
700 Dried Hemoglobin
Dried hemoglobin in powdered form takes up oxygen rapidly from the air and more rapidly at*higher oxygen tensions. When samples are thus prepared for sealing in ampules, as the pressure in the ampules is reduced to a few mm., the powder begins to seethe and sputter from the trapped and possibly adsorbed air. To
620 580 540 h in mp
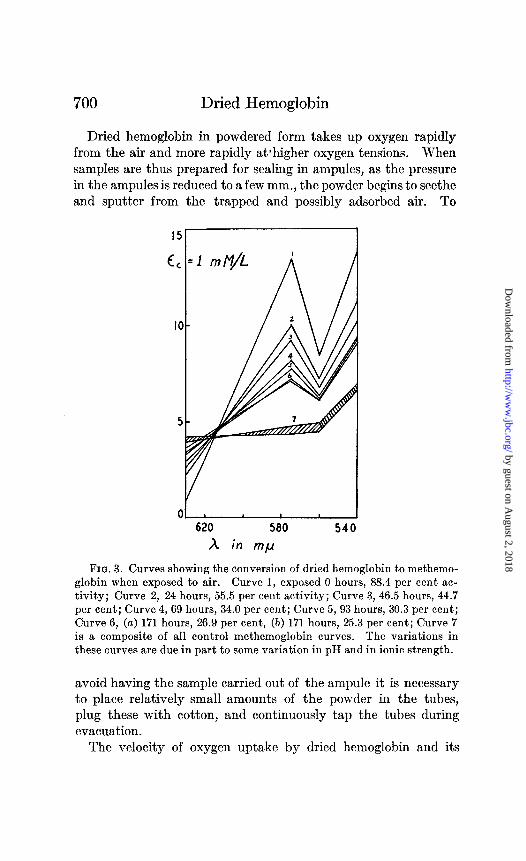
FIG. 3. Curves showing the conversion of dried hemoglobin to methemo- globin when exposed to air. Curve 1, exposed 0 hours, 88.4 per cent ac- tivity; Curve 2, 24 hours, 55.5 per cent activity; Curve 3, 46.5 hours, 44.7 per cent; Curve 4,69 hours, 34.0 per cent; Curve 5, 93 hours, 30.3 per cent; Curve 6, (a) 171 hours, 26.9 per cent, (5) 171 hours, 25.3 per cent; Curve 7 is a composite of all control methemoglobin curves. The variations in these curves are due in part to some variation in pH and in ionic strength.
avoid having the sample carried out of the ampule it is necessary to place relatively small amounts of the powder in the tubes, plug these with cotton, and continuously tap the tubes during evacuation.
The velocity of oxygen uptake by dried hemoglobin and its
by guest on August 2, 2018
http://ww
w.jbc.org/
Dow
nloaded from

D. B. Morrison and A. Hisey 701
relation to methemoglobin formation are being studied, and we expect to report later in more detail upon these behaviors. That methemoglobin is formed under these conditions is shown in Figs. 2 and 3. The activity data and spectrophotometric curves were obtained upon powdered samples whose oxygen uptake had been followed in Warburg respirometers. The gas phase in the respirometers was air at a temperature of 38”, and the time inter- vals were as noted in Fig. 3. Extinction coefficients were meas- ured at four wave-lengths.
Agreement of data for activity of our dried preparations by both gasometric and spectrophotometric analyses suggests that methemoglobin is the principal, if not sole, pigment contaminant, the spectrophotometric data being calculated on the assumption that this is so.
No further methemoglobin formation and loss of activity occur in dried and powdered samples when they are sealed in evacuated glass ampules over periods of many months.
DISCUSSION
In developing a method for drying reduced hemoglobin with retention of its reversible activity towards oxygen we have pre- pared and analyzed 135 individual samples. Some of these, e.g. Samples 93 and 110 (Table III), have been anlayzed as many as six times at varying intervals after drying. Methods used in determining active and total pigment have been checked repeat- edly against each other, both on dried samples and on undried controls, with excellent agreement.
Data for the first 129 samples are summarized in Table I in the order of activity. All of these samples were dissolved in contact with air. Under such conditions activities approaching 100 per cent have been obtained only with very small samples which in drying were uniformly distributed over the sides of the chamber in a very thin film; these dissolved within 1 or 2 minutes to form rather dilute solutions. With larger samples, delayed solution, and higher concentration, exposure to air causes appreciable loss in activity.
We have had more difficulty in obtaining high activities with dog hemoglobin than withox, pig, or human when the oxygen- inactivating factor was not excluded at critical stages of prepara-
by guest on August 2, 2018
http://ww
w.jbc.org/
Dow
nloaded from

702 Dried Hemoglobin
tion and solution of the samples. Dog hemoglobin, of the four varieties, is the most readily crystallized and, likewise, is slower to dissolve after drying; more methemoglobin is thus formed. In earlier attempts to dry dog hemoglobin crystals very thick suspen- sions of the crystals were used in expectation of shortening the drying time. However, drying occurred very quickly only on the surface and the underlying crystals were not reduced. Such preparations were invariably low in or devoid of activity. Other samples of the same crystals when suspended in sufficient distilled water to dissolve completely upon reduction, but before appre- ciable drying had taken place, gave activities up to 93 per cent. It is apparent that rapid drying of a mixture of reduced hemoglo- bin and oxyhemoglobin greatly accelerates methemoglobin forma-
TABLE I
Dried Samples Dissolved in Presence of Air
No. of samples I
Activity
2 8
23 33 28 35
per cent
98-100 95- 98 90- 95 85- 90 75- 85 lo- 75
tion. Indeed, conversion to methemoglobin is so rapid, and under proper conditions so complete, as to furnish a method for preparing methemoglobin without the use of an oxidizing agent other than oxyhemoglobin.
Such considerations, we believe, explain the failure of Van Slyke and his coworkers to obtain dry, active hemoglobin preparations. Likewise, the necessity of dissolving the dried reduced hemoglobin samples in an oxygen-free atmosphere to avoid loss of activity is explained. With oxygen present the first hemoglobin to dissolve is converted to oxyhemoglobin and this in contact with the as yet undissolved reduced hemoglobin forms methemoglobin. These conditions are the reverse of those in the process of drying thick, crystalline suspensions, where reduced hemoglobin in solution is in contact with solid oxyhemoglobin; but the result is the same.
by guest on August 2, 2018
http://ww
w.jbc.org/
Dow
nloaded from

D. B. Morrison and A. Hisey 703
Not as much methemoglobin is formed in dissolving samples as in drying them because of the lower temperature in the first in- stance and the shorter time required for complete solution than for complete drying.
In Table II are seen the greatly improved activities of samples dissolved in the reduced state and oxygenated only after solution was complete. Sample 130 is of especial interest, since data are given for the activity of the dried sample dissolved in vucuo, an undried control, and a portion of the same hemoglobin dried ex- actly as the in vacua sample but dissolved in the presence of air.
TABLE II
Dried Samples Dissolved in Distilled Water in Vacua
Sample No. Species
130-a
130-b* 130-ct 131 132 133 134 135-a 135-b*
.-
-
Pig “ “ “
Dog Human
“
ox ‘I
Active pigment Total pigment
vol. per cent vol. per cent 11.82 12.32 16.80 16.97 13.89 15.59 14.01 14.31
6.56 6.71 7.66 7.85 9.08 9.28 9.56 9.76
13.84 13.73
Activity
per cent
96.0
99.0
89.1 97.9 97.8 97.6 97.7 97.9
100.8
* Undried control samples as a check upon the accuracy of the methods used.
t A sample of a pig blood, No. 130, dried in exactly the same way as Sample 130-a but dissolved in the presence of air instead of in vacua.
A 10 per cent loss in activity due to methemoglobin formation when the sample is dissolved in contact with air is to be compared with a 3 per cent loss in activity when air was excluded during solution. Similar comparisons are shown for Samples 110 and 111 (Table III).
While our method of drying yields preparations which appear completely dry, which can be easily powdered to a fine dust, and which gain weight when exposed to air at room temperature, on heating in air at 105” there is a loss in weight of 3 to 6 per cent. We have taken this to represent residual water content of the dried samples and in the earlier part of our work we believed that
by guest on August 2, 2018
http://ww
w.jbc.org/
Dow
nloaded from

704 Dried Hemoglobin
this value might have some relation to activity. Inspection of Fig. 4 shows that no such relationship is apparent. We are still under the impression, however, that residual moisture content of the dried samples may affect the rate of methemoglobin formation upon their exposure to air; our data on this point are as yet too scanty to justify a definite conclusion.
It was observed early that dried samples become completely in- active when exposed to air for a few days. The data of Table III
14
I
.
.
.
c iTi .
-i3-
: * .
. .
96 . .
ii
. * . : .
. .
4- * . . . .
i
.
2
01
0 20 I
40 60 60 101 ACTIVITY PER CENT
-1 0
FIG. 4. The moisture content of these samples was varied intentionally to determine whether there is any relation of moisture to activity.
demonstrate the stability of the same or similar preparations when kept for long periods in evacuated ampules. These data confirm the supposition that methemoglobin is not formed unless oxygen is present. In Table III the letter following the sample number designates the ampule of that particular sample. Varia- tions in activity of the hemoglobin from different ampules of the same sample are probably due to inequalities in sampling and to differences in methemoglobin formation during solution of the several portions. Certainly four of the samples dissolved in vacua show a higher activity than portions of the same samples dissolved
by guest on August 2, 2018
http://ww
w.jbc.org/
Dow
nloaded from

D. B. Morrison and A. Hisey 705
in contact with air. The generally lower activities of the sealed samples represented in Table III are to be attributed to the rela-
TABLE III
Activity of Samples Sealed in Ampules
Sample No.
20 21 22 54-a 54-b 93-a 93-b 93-c 93-d 93-e 93-f
103-a 103-b 103-c 103-d 108-a 108-b 108-c 108-d 108-e 109-a 109-b 110-a 110-b 110-c 110-d* 110-e* 110-f* 111-a Ill-b 111-c Ill-d*
Species
Dog ox “ Pig
ox
“
Pig
ox
“
Age of sample Activity
days per cent
370 63.1 29 78.6 28 61.5 42 83.5
185 87.5 3 87.1
18 83.8 55 82.1 73 76.4
105 80.7 133 83.4
0 70.7 196 60.3 196 70.2 196 59.4
14 79.8 161 84.0 193 80.0 236 80.7 236 80.3
51 73.3 51 78.5
161 76.1 161 80.4 233 80.8 233 89.1 234 87.7 234 85.6 161 61.7 161 65.7 234 71.1 234 73.6
* Samples dissolved in vacua instead of in the presence of air.
tively large amounts of hemoglobin which were dried in one lot to provide a sufficient quantity for repeated analyses. Further-
by guest on August 2, 2018
http://ww
w.jbc.org/
Dow
nloaded from

706 Dried Hemoglobin
more, some of these lots were prepared before we fully appreciated some of the factors which affect activity.
We are indebted to Dr. T. P. Nash, Jr., for suggesting this prob- lem and for his kind encouragement.
SUMMARY
1. A method is described for drying reduced hemoglobin with little or no loss in activity.
2. Some properties of the dry hemoglobin and solutions thereof are described.
3. In the dried state reduced hemoglobin may be preserved apparently indefinitely in vacua without change in activity.
BIBLIOGRAPHY
1. Bohr, C., Skand. Arch. Physiol., 3, 85 (1892). 2. Hufner, G., Arch. Anat. u. Physiol., Physiol. AU., 130 (1894). 3. Heidelberger, M., J. Biol. Chem, 63, 31 (1922). 4. Van Slyke, D. D., Hastings, A. B., Heidelberger, M., and Neill, J. M.,
J. Biol. Chem., 64, 481 (1922). 5. Barcroft, J., The respiratory function of the blood, Cambridge, 2nd
edition, pt. 2 (1928). 6. Morrison, D. B., and Hisey, A., J. Biol. Chem., 169,233 (1935). 7. Van Slyke, D. D., and Hiller, A., J. Biol. Chem., 84, 205 (1929). 8. Drabkin, D. L., and Austin, J. H., J. Biol. Chem., 112, 51 (1935-36). 9. Van Slyke, D. D., and Hiller, A., J. Biol. Chem., 78,807 (1928).
10. Van Slyke, D. D., and Neill, J. M., J. BioZ. Chem., 61, 523 (1924). 11. Austin, J. H., and Drabkin, D. L., J. BioZ. Chem., 112, 67 (193536).
by guest on August 2, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Dempsie B. Morrison and Alan HiseyIN A DRY AND ACTIVE STATE
THE PREPARATION OF HEMOGLOBIN
1937, 117:693-706.J. Biol. Chem.
http://www.jbc.org/content/117/2/693.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
tml#ref-list-1
http://www.jbc.org/content/117/2/693.citation.full.haccessed free atThis article cites 0 references, 0 of which can be
by guest on August 2, 2018
http://ww
w.jbc.org/
Dow
nloaded from