submitted for the award of the degree of doctor of philosophy
TRANSCRIPT

TAXONOMY AND ECOLOGY OF NEMATODES OFAGRICULTURAL FIELDS AND
WASTELANDS
THESIS
SUBMITTED FOR THE AWARD OF THE DEGREE OF
Doctor of Philosophy IN
ZOOLOGY
BY
GAURAV KUMAR SINGH
DEPARTMENT OF ZOOLOGY ALIGARH MUSLIM UNIVERSITY
ALIGARH (INDIA) 2011

Wxw|vtàxw gÉ `ç
UxÄÉäxw ctÜxÇàá

IRFAN AHMAD Ph.D. DEPARTMENT OF ZOOLOGY Professor Aligarh Muslim University,
Aligarh – 202002, INDIA [email protected]
Date: ………………….
Certificate
This is to certify that the research work presented in the thesis entitled,
“Taxonomy and Ecology of Nematodes of Agricultural Fields and Wastelands” by
Mr. Gaurav Kumar Singh is original and was carried out under my supervision. I have
permitted Mr. Singh to submit it to the Aligarh Muslim University, Aligarh in
fulfillment of the requirements for the degree of Doctor of Philosophy in Zoology.
IRFAN AHMAD (Supervisor)

Acknowledgements
I bow in deep reverence to GOD the Almighty, who blessed me with an innumerable favour of
academic work.
It has been a great opportunity for me to work under the able guidance of Prof. Irfan Ahmad,
Chairman, Department of Zoology, Aligarh Muslim University, Aligarh. I express my sincere
gratitude for his ever-lasting important advices, valuable suggestions, stimulating discussions,
constructive criticism, sense of perfection and precision which enabled me to complete this work. Inspite
of his tight departmental commitments he kept his door open wide for me. His affectionate instinct and
constant encouragement were always a boon to me. Any error if still remains is entirely my own.
I am thankful to the Chairmen (past and present), Department of Zoology, Aligarh Muslim
University, Aligarh for providing necessary laboratory facilities for the work.
I feel more than obliged to all my respected teachers Prof. M. Shamim Jairajpuri (INSA
Senior Scientist), Prof. Wasim Ahmad, Prof. Qudsia Tahseen and Prof. Mahalaqa Choudhary for
their valuable advices and kind suggestions.
I wish to acknowledge my seniors and lab colleagues, Dr. Noorus Sabah, Dr. Md.
Mahamood, Dr. Ali Asghar Shah, Mr. Puneet Kumar, Ms. Gazala Yousuf & Ms. Nadia Sufyan.
I am also thankful to my senior Dr. Md. Baniyamuddin and colleagues Dr. Shikha Ahlawat,
Ms. Uzma Tauheed, Mrs. Tabinda Nusrat , Ms. Malka Mustaqim and Ms. Sumaiya Ahad for
their constant inspirations and support.
Special thanks are due to my friend and colleague Vijay Vikram Singh for his unconditional
and constant help during compilation of this work.
Thanks are also due to my friends Imran and Vimal for their constant encouragement during
the course of present work.
Last but not the least, I would like to thank my wife for making me believe that I can do it and
my little angel for giving me cheers of my life.
My brothers Mr. Umesh Singh and Mr. Saurabh and my bhabhi Mrs. Meenakshi deserve
special thanks for creating ideal milieu at home which helped me to complete the present work.
The financial assistance from the Ministry of Environment and Forest, Government of India,
New Delhi is also thankfully acknowledged.
Gaurav Kumar Singh

CONTENTS
PART – A
Page
INTRODUCTION 1
HISTORICAL BACKGROUND 12
MATERIALS AND METHODS 21
SYSTEMATICS 24
ORDER RHABDITIDA 24
SUBORDER CEPHALOBINA 24
SUPERFAMILY CEPHALOBOIDEA 25
FAMILY CEPHALOBIDAE 26
SUBFAMILY CEPHALOBINAE 26
Genus Pseudacrobeles 27
Pseudacrobeles ventricauda sp. n. 28
Pseudacrobeles mucronatus sp. n. 34
SUBFAMILY ACROBELINAE 41
Genus Acrobeles 42
Acrobeles mariannae 42
Genus Acrobeloides 47
Acrobeloides glandulatus sp. n. 47
Genus Cervidellus 54
Cervidellus neoalutus sp. n. 54
Cervidellus minutus sp. n. 60

Genus Chiloplacus 66
Chiloplacus aligarhensis sp. n. 66
Genus Nothacrobeles 73
Nothacrobeles punctatus sp.n. 73
Genus Stegellata 80
Stegellata ophioglossa 80
Genus Zeldia 85
Zeldia tridentata 85
SUPERFAMILY PANAGROLAIMOIDEA 90
FAMILY PANAGROLAIMIDAE 90
SUBFAMILY TRICEPHALOBINAE 91
Genus Tricephalobus 91
Tricephalobus quadripapilli sp. n. 92
FAMILY BREVIBUCCIDAE 98
Subfamily Brevibuccinae 98
Genus Brevibucca 98
Brevibucca postamphidia sp. n 99
Genus Plectonchus 105
Plectonchus coptaxii sp. n. 105
SUMMARY 112

PART – B
INTRODUCTION 115
MATERIALS AND METHODS 125
RESULTS 131
DISCUSSION 153
REFERENCES 162

Part – A Taxonomy

Introduction

The nematodes are a successful group of invertebrates placed at a low
level in the taxonomic hierarchy of the animal kingdom. They are the most
diverse phylum, and one of the most diverse of all animals. They have
successfully adapted to nearly every ecological habitat from marine to fresh
water, from the Polar regions to the tropics, as well as the highest to the lowest of
elevations. They are ubiquitous in freshwater, marine, and terrestrial
environments, where they often outnumber other animals in both individual and
species counts, and are found in the locations as diverse as Antarctica and
oceanic trenches. Some of them also can withstand complete dryness on the
surface of rocks. Their size too is extremely variable ranging from less than 100
µm (Greefiella minutum) to greater than 8 metres (Placentonema gigantissima).
The nematodes are the planets most abundant metazoa; of every five animals,
four are nematodes (Platt, 1994; Bongers & Ferris, 1999). They are particularly
abundant in marine, freshwater, and soil habitats. A square yard of woodland or
agricultural habitat may contain several million nematodes. The existence of
nematodes living in water, soil or in parasitizing the plants remained largely
unknown perhaps, because of their exceedingly small size, absence of any
colouration, mostly underground habitat and the difficulties encountered in their
isolation.
The phylum Nematoda is characterized by high species diversity. It has
been estimated that the total number of described and undescribed species might
be ranged from 0.1 to 100 million (May, 1988; Hammond, 1992;
Lambshead,1993; Coomans, 2000). The nematodes are not only numerically
1

abundant, but they are also very diverse in terms of species. Usually species
richness at a single site is high with an average of 20-60 species per soil sample.
In soil, the nematodes dominate in number as well as species over all
other soil inhabiting animals collectively and have occupied all possible habitats
representing a very wide range of biological diversity. Soil nematode
communities have the potential to provide insights into many soil processes and
functions as most nematodes are active in soil throughout the year (Ritz &
Trudgill, 1999). Nematodes can be used as bioindicators of soil health because
they are ubiquitous and have diverse feeding behaviours and life strategies
(Bongers & Bongers, 1998; Neher, 2001). They occupy several trophic grades
and a central position in the soil food web and play significant roles in biological
processes such as nitrogen cycling and plant growth patterns (Neher, 2001). Soil
nematodes stabilize soil ecosystems, promote substance cycling and energy flow
(Ingham et al., 1985). The ecological classification of terrestrial nematodes have
usually been based on feeding biology (trophic functions) and on life strategies;
colonizers versus persisters (Bongers, 1990). Yeates et al. (1993) classified the
terrestrial nematodes into eight trophic groups viz., plant feeding, hyphal feeding,
substrate ingestion, predation on animals, bacterial feeding, unicellular eukaryote
feeding, dispersal/infective stage of parasites and omnivorous. Free-living
nematodes promote decomposition of soil organic matter, mineralization of plant
nutrients and nutrient cycling, amend soil physico-chemical property and
improve soil fertility (Ferris et al., 2004). Some free living nematodes suppress
bacterial, fungal and nematode diseases (Khan & Kim, 2007).
2

All terrestrial ecosystems consist of aboveground and belowground
components that interact to influence community- and ecosystem-level processes.
Several recent studies have indicated that biotic interactions in soil can regulate
the structure and functioning of the above ground communities (Wardle et al.,
2004). Soil nematode communities can provide unique insights into many aspects
of soil processes; they can offer a holistic measure of the biotic and functional
status of soils (Ritz & Trudgill, 1999). They are good environmental indicators
because of their strong relationships with land management (Todd, 1996; Neher
and Campbell, 1994; Liang et al., 1999; Fiscus and Neher, 2002) and
aboveground vegetation (Ingham et al., 1985; Bongers and Bongers, 1998;
Bongers and Ferris, 1999; Yeates, 1999).
Measurement of meiofaunal diversity and abundance is an important but
time consuming process. Morphological identification of individual organisms to
named species is often not technically possible due to sheer abundance, small
size, and lack of expert knowledge of the groups encountered. This is especially
true of nematodes, whose diversity in soils and sediments remains essentially
unknown. Surveys of benthic sediments suggest that the total species number for
marine nematodes may exceed 1 million (Lambshead 1993; Lambshead 2001),
with only a few thousand described in the scientific literature (Malakhov 1994;
De Ley & Blaxter 2002). In terrestrial systems, nematode diversity appears to be
under-reported (Lawton et al., 1998), with, for example, only about 200 species
of soil nematodes being described from the British Isles (Boag & Yeates 1998).
The maximum number of nematode taxa described from a single soil site is 228
from a prairie in Kansas, USA (Orr & Dickerson 1966; Boag & Yeates 1998).
3

Nematodes are arguably the most numerous metazoans in the soil and aquatic
sediments (Lambshead, 2004), the extraordinary species diversity, paucity of
trained taxonomists, labour intensive work for traditional morphological
identification of soil fauna challenges the nematode taxonomy. To develop new
technology for identification, classification, genome relatedness and diversity, in
nematode genepools the technology to be developed will use molecular,
biosystematic, informatic and genetic tools. New approaches are coming in use to
aid species identifications within the context of a classical morphological system.
Currently a shift from the purely phenotypic to using a combination of both
phenotypic and molecular methods is observed (Powers et al., 1997; Powers,
2004). Also, the phylogenetic species concept has gained more support recently
(Adams, 1998, 2002) and ways to extend its theoretical appeal into practicality
have been evaluated (Nadler, 2002).
For years, morphological identification was the only method widely used
to identify nematodes. As our knowledge of nematodes of agronomical
importance increased, it became clear that morphology alone did not reveal the
complete picture of observed pathological differences between populations
within morphologically delimited species. As a result, new methods have been
looked for that can better predict observed pathological behaviors among
populations within species. Numerous molecular techniques have been developed
that are capable of identifying and quantifying nematodes at the species level and
below. Techniques such as isoenzyme pattern analysis, restriction fragment
length polymorphism (RFLP) analysis, random amplified polymorphic DNA
(RAPD) analysis, polymerase chain reaction (PCR), quantitative polymerase
4

chain reaction (qPCR), and sequencing of diagnostic rDNA regions have all been
used successfully to identify and quantify several agriculturally important plant-
parasitic nematodes. These methods have their own advantages and limitations.
Most of these methods have been widely used in the diagnostics of agriculturally
important nematodes. DNA sequences of marker genes, Denaturing Gradient Gel
Electrophoresis (DGGE) and the more recently developed method of
pyrosequencing are the three methods employed in biodiversity studies of
freeliving forms. Andre et al. (2002) highlighted the need for the development
and consistency of methods in soil faunal monitoring; commenting that molecular
techniques for community analysis are now widely used in soil microbiology and
have greatly expanded our knowledge of soil microbes. Molecular methods
provide an alternative to traditional morphological identification for routine
assessment of described species. Their application has enabled profiling of
environmental samples of soil microbial populations, overcoming the need to
culture and identify bacteria and fungi from complex mixtures (Amann et al.,
1995) and similarly may reduce the taxonomic expertise currently required to
characterise microfaunal communities.
Blaxter et al. (1998) produced the first molecular phylogenetic framework
of the phylum Nematoda. They constructed a database of small subunit (SSU)
sequences from 53 taxa, including 41 new sequences to construct a phylogenetic
tree of nematodes. Sequences were aligned with reference to a secondary-
structure model and on the basis of similarity. They recognise three major clades:
Clade I groups the vertebrate-parasitic order Trichocephalida with the insect-
parasitic Mermithida, plant-parasitic Dorylaimida and free-living Mononchida,
5

Clade II links the plant-parasitic Triplonchida with the free-living Enoplida &
Monhysterida and clade C+S groups Chromadorida and Secernentea. Within
Secernentea three major clades were identified (clades III, IV and V). Clade III
represents a grouping of vertebrate- and arthropod-parasitic taxa from the orders
Ascaridida, Spirurida, Oxyurida and Rhigonematida. Clade IV a ‘cephalobid’
clade, groups the plant-parasitic orders Tylenchida and Aphelenchida, the
vertebrate-parasitic genus Strongyloides and the entomopathogenic genus
Steinernema with free-living bacteriovores of the rhabditid families Cephalobidae
and Panagrolaimidae. Clade V groups C. elegans and other members of the
suborder Rhabditina with the vertebrate-parasitic order Strongylida, the
entomopathogenic genus Heterorhabditis and the order Diplogasterida. De Ley
and Blaxter (2002, 2004) updated the classification of the phylum Nematoda
using molecular data available from additional species, with morphological data
to assist the placement of taxa for which SSU sequences were not yet available.
They used SSU phylogenies to develop a novel classification reflecting recent
evolutionary findings and proposing the infraorders Cephalobomorpha,
Panagrolaimomorpha and Tylenchomorpha, all within a considerably expanded
suborder Tylenchina.
Nematodes of the suborder Cephalobina Andrassy, 1974 include an
ecologically and morphologically diverse array of species that range from soil
dwelling microbivores to parasites of vertebrates (Strongyloidoidea, including
Strongyloides) and invertebrates [entomopathogens used commercially for
biological control (Steinernema)]. Despite a long history of study, certain of these
microbivores (Cephaloboidea) present some of the most intractable problems in
6

nematode systematics (De Ley, 1997); the lack of an evolutionary framework for
these taxa has prevented the identification of natural groups and inhibited
understanding of soil biodiversity and nematode ecology.
The Cephaloboidea are a relatively distinctive group of widely distributed
bacterial-feeding soil nematodes, most frequently represented by the family
Cephalobidae, which includes more than 275 nominal species and 24 genera.
These nematodes, which are often striking in their labial morphology, are found
in soils worldwide and are typically the most abundant microbivores in nutrient-
poor soils such as deserts (Freckman and Mankau, 1986) and dry Antarctic
valleys (Freckman and Virginia, 1997). Despite their abundance and
cosmopolitan distribution, cephalobs have been among the most difficult
nematodes to diagnose, identify and classify. Historically these genera have
primarily been recognized based on variation in labial morphology, but molecular
phylogenies show the same general labial (probolae) morphotype often results
from recurrent similarity, a result consistent with the phenotypic plasticity of
probolae for some species in ecological time. The taxonomy, identification and
classification of cephalobs have become more difficult with time. For example,
genera that once seemed discrete based on morphological observations of
relatively few species have been blurred by discovery and description of
additional species with confounding character combinations, or overlaps between
characters previously considered diagnostic (De Ley, 1997). As a result, the
morphological characters originally proposed for distinguishing most genera are
now thought to be of questionable value, as demonstrated for Acrobeloides,
Cephalobus, Chiloplacus, Eucephalobus and Pseudacrobeles (De Ley, 1997).
7

More potentially disturbing are longstanding reports of substantial intraspecific
variation of lip structures within and between natural and in vitro cultured
populations of certain species of Cephalobidae (Allen and Noffsinger, 1972;
Anderson, 1965, 1968; Anderson and Hooper, 1970), with the range of variation
within single species sometimes exceeding differences used to discriminate
among genera. Addressing these systematic problems by developing a
phylogenetic framework for cephalobs is essential, because investigations of soil
biodiversity and ecology almost invariably require identification of cephalobs,
and taxa from this suborder are increasingly used as model organisms for
comparative developmental studies (Baldwin et al., 1997; Dolinski et al., 2001;
Félix et al., 2000; Goldstein et al., 1998; Schierenberg, 2000). Moreover, a
comprehensive phylogenetic framework for cephalobs has the potential to
provide insights into the evolution of features that may be associated with
nematode parasitism of plants, annelids, insects, and vertebrates.
Proposed phylogenetic affinities of cephalobs have been quite diverse.
For much of the early history of nematode taxonomy, cephalobs were considered
very closely related to rhabditids, the group that includes the premier nematode
model organism Caenorhabditis elegans. Other authors have proposed closer
affinities with certain predominantly parasitic nematodes, most notably with the
fungivorous or phytophagous tylenchs (Siddiqi, 1980), the annelid parasitic
drilonematids (Coomans and Goodey, 1965; De Ley and Coomans, 1990;
Lisetskaya, 1968; Spiridonov et al., 2005), and the entomopathogenic
steinernematids (Poinar, 1993). Regions of 28S rDNA subunit, including the
D2/D3 variable domains, have been used to infer relationships among certain
8

closely related species, primarily congeners (Baldwin et al., 2001; De Ley et al.,
1999; Nadler et al., 2003; Stock et al., 2001), Molecular phylogenies with
taxonomically broader representation for cephalobs have been inferred using
sequences from SSU (18S) ribosomal DNA (Blaxter et al., 1998; Félix et al.,
2000; Goldstein et al., 1998) and RNA polymerase II genes (Baldwin et al.,
1997). These studies have provided new insights into the relationships of
cephalobs to other major groups of nematodes, including the unexpected Wnding
that Tylenchida and Aphelenchida, which include the most economically
important plant parasites among nematodes, share most recent common ancestry
with cephalobs (Blaxter et al., 1998). A phylogeny of Cephaloboidea also
provides the opportunity to test the monophyly of other genera and examine their
relationships. Molecular phylogenetic trees can further serve as independent
frameworks for the investigation of morphological characters in nematodes
(Baldwin et al., 1997; Nadler & Hudspeth, 1998).
Recently Nadler et al. (2006) studied the phylogeny of Cephalobina. The
phylogenetic analyses of ribosomal (LSU) sequence data from 53 taxa revealed
strong support for monophyly of taxa representing the cephaloboidea, but do not
support the monophyly of most genera within this superfamily. Trees inferred
from LSU sequences, include a large clade containing most (but not all) genera
classically representing cephalobs plus plant parasites from the orders Tylenchida
and Aphelenchida. Published phylogenies inferred from SSU rDNA, although
including far fewer cephalob taxa, also recover this relationship (Blaxter et al.,
1998; Félix et al., 2000). Thus, both SSU and LSU sequence data indicate that
certain plant parasitic species share most recent common ancestry with
9

cephalobs, contrary to most traditional evolutionary concepts. Although the LSU
trees reveal that a large group of taxa normally classified as cephalobs are
monophyletic, the exclusion of certain genera and the close relationship of some
plant parasites indicates that even newer classification proposals for these taxa
(De Ley and Blaxter, 2002) will require some amendments to reflect these
findings. The LSU trees clearly refute the hypothesis that Macrolaimellus is more
closely related to chambersiellids (represented by Macrolaimus and Fescia in the
LSU trees), and strongly support its inclusion in Cephaloboidea.
Meldal et al. (2007) added SSU rDNA sequences for 100 un-sequenced
species of nematodes, including 46 marine taxa. Sequences for more than 200
taxa have been analysed based on Bayesian inference and logDet-transformed
distances. The phylogenetic analysis provided the support for the re-classification
of secernentea as the order Rhabditida that derived from a common ancestor of
chromadorean orders Araeolaimida, Chromadorida, Desmodorida,
Desmoscolecida and Monhysterida. Their analysis also support the the position of
Bunonema close to the Diplogasteroidea in the Rhabditina. Meldal et al. also
proved that SSU rDNA genes are very effective in the recovery of many
monophyletic group within the phylum Nematoda and provided clarification of
relationships that were uncertain or controversial. However, there were certain
limitations to the use of SSU. The SSU gene did not provide significant support
for the class Chromadoria or clear evidence for the relationship between the three
classes, Enoplia, Dorylaimia, and Chromadoria. Furthermore, across the whole
phylum, the phylogenetically informative characters of the SSU gene are not
informative in a parsimony analysis, highlighting the short-comings of the
10

parsimony method for large-scale phylogenetic modelling. Recently Abebe et al.
(2011) presented a review of the various techniques used in the taxonomy of free
living and plant parasitic nematodes and critique those methods in the context of
recent developments and trends including their implications in nematode
taxonomy, biodiversity and biogeography.
11

Historical Background

Although nematology attracted attention and recognition only in 20th
century, our knowledge of a few species of nematodes of medical importance
dates back to Papyrus Ebers (Circa 1500 BC). The intestinal round worm
(Ascaris lumbricoides), filarid (Wucheraria bancrofti) and guinea worm or fiery
serpent of Moses (Dracunculus medinensis) were already known to the ancient
man. However, marine, freshwater, soil and plant nematodes remained little
known groups mainly because of their extremely small size and the difficulties
encountered in their isolation, mounting and observation.
Knowledge of free-living nematodes dates back to 1656 when Borellus
for the first time observed Turbatrix aceti the ‘vinegar eels’. Observations and
descriptions of plant parasitic nematodes, which were less conspicuous to ancient
scientists, didn’t receive as much or as early attention as did animal parasites.
Needham (1743) solved the “riddle of cockle” when he crushed one of the
diseased wheat grains and observed “Aquatic Animals” the first plant parasitic
nematode (Anguina tritici). He found tiny serpent-like worms, which were later
named Vibrio tritici Steinbuch (1799). Muller (1783) described several species of
free-living freshwater nematodes. Nematode taxonomy further developed and
landmark progress was observed in the middle of 19th century. Around 1850,
marine biologists began to recognize nematodes; there were, studies on the
nematodes of Iceland (Leuckart, 1849), the Mediterranean (Eberth, 1863), the
English coast (Bastian, 1865), the coast of Brittany (Villot, 1875) and on
nematodes collected by various expeditions (Von Linstow, 1876). Freshwater
nematodes received further interest around 1890 with the papers of Daday (1897)
on the Hungarian fauna. Dujardin (1845), Bastian (1865), Schneider (1866), de
12

Man (1884), Daday (1905) and Maupas (1900) were the pioneers of the field.
Early work on the free-living nematodes included careful descriptions of
Enoplus, Oncholaimus, Rhabditis and Dorylaimus (Dujardin, 1845). He was first
to recognize the close relationship of free-living and plant parasitic nematodes.
Bastian (1865), made significant contributions in the field of Nematology. He
grouped the free-living nematodes into soil, fresh water and marine forms and
described 100 new species of 30 genera in which 23 were new to science, in a
single paper. de Man (1876-1927) listed eight families of free-living nematodes.
de Man’s (1884) formula for denoting measurements of nematodes is universally
used in taxonomy till date. Cobb, a contemporary of de Man, is considered, the
Father of Nematology. He placed nematodes under separate Phylum Nemata.
Significant changes in classification were proposed by Cobb (1920), De Coninck
(1965), Maggenti (1963, 70), and by Andrássy (1976, 84). In Chitwood’s (1933,
37) classification, ‘Nematoda’ was treated as a phylum with two classes,
‘Phasmidia’ and ‘Aphasmidia’, based on presence or absence of phasmids. The
terms Secernentea and Adenophorea were introduced by Chitwood (1958) who
proposed the system of classification of nematodes including free-living and
parasitic nematodes. Andrássy has contributed extensively to the taxonomy of
major groups of terrestrial and freshwater nematodes. In his productive career he
described more than 500 taxa of nematodes and at least 39 taxa were named after
him. Besides his voluminous contributions to nematode taxonomy and
systematics, he has had an enormous influence on soil and nematode ecology. He
published keys for identification, proposed and raised higher taxa, amended and
put forth classification schemes besides authoring valuable books including the
13

extremely useful compilation, ‘Klasse Nematoda' (1984) based on the diagnosis
of orders of Araeolaimida, Enoplida, Chromadorida, Monhysterida, and
Rhabditida and their subordinate taxa. This book exercised major influence on
the direction of nematode ecology in that it bridged the gap between nematode
taxonomy and soil ecology.
Order Rhabditida was erected by Chitwood (1933) for bacteriophagus
rhabditids. Dujardin (1845) first established the genus Rhabditis with Rhabditis
terricola as its type species. However it was not clearly defined until more than
one hundred year later (Dougherty, 1955). Örley (1880) proposed a family
Rhabditidae, for the genera Anguillula, Cephalobus, Oxyuris, Rhabditis and
Teratocephalus. He placed this family in the higher category “Rhabditi formae”
which formed a connecting link between free living and animal parasitic
nematodes. Micoletzky (1922) described seven species. His system was,
however, rather artificial in that he united all nematodes having a prismatic,
unarmed stoma under the family Rhabditidae, viz. the subfamilies
Cylindrolaiminae, Plectinae, Rhabditinae and Bunonematinae. The subfamily
Rhabditinae was itself heterogenous, and composed of the following genera:
Rhabditis, Diploscapter, Cephalobus, Chambersiella, Teratocephalus and
Rhodolaimus.
The record of cephalobid nematode can be traced back to 1656 when
Borellus observed “vinegar eels” for the first time. Muller (1783) named these
eels as Vibrio aceti, later on it was redescribed and shuffled to various taxa by
several authors, finally Peters (1927) proposed the genus Turbatrix and accepted
T. aceti as its type species. Though the first cephalob species “Cephalobus
14

persegnis” was formally described in 1865 by Bastian, it was not until the work
of Cobb (1924) and Thorne (1925, 1937) that basic taxonomic concepts and
terminology were established. Cobb (1924) rehabilitated the genus Acrobeles von
Linstow, 1877 and suggested the subgenera Acrobeles and Acrobeloides. Thorne
(1925) accepted the subgenera proposed by Cobb and produced a detailed
account on morphology, systematics and taxonomy of the genus Acrobeles. He
described thirty new species of Acrobeles and grouped them under two
subgenera. Thorne’s (1937) revision of the Cephalobidae has been of immense
value and importance. He (l.c) proposed subfamily Acrobelinae for genera
Placodira, Chiloplacus, Cervidellus and Zeldia and redescribed the genus
Acrobeloides. He also proposed superfamily Panagrolaimoidea with family
Panagrolaimidae and subfamily Panagrolaiminae. Later on he (1938, 39)
described the genera Panagrobelus and Panagrellus under Panagrolaiminae and
Stegellata under Acrobelinae. Simultaneously, Steiner (1934, 36, 38) proposed
the genus Procephalobus under the family Panagrolaimidae, and in Cephalobidae
the genera Eucephalobus, Tricephalobus and Pseudacrobeles were proposed.
Andrássy (1967) published detailed information on Cephalobinae
Filipjev, 1934. In 1974, he proposed the Suborder Cephalobina, to include an
incredibly diverse array of free-living microbivores. He included three
superfamilies Cephaloboidea Filipjev, 1934, Elaphonematoidea Heyns, 1962 and
Panagrolaimoidea Thorne, 1937 under Cephalobina. In Cephaloboidea he erected
the family Metacrobelidae and placed Metacrobeles Loof, 1962 under it. Further,
in 1984, he added two more superfamilies viz. Drilonematoidea Peirantoni, 1916
and Myolaimoidea Andrassy, 1958 under Cephalobina and accepted eight
15

families and ten subfamilies in these five superfamilies. He also proposed five
new genera Acrobelophis, Ypsylonellus, Stegelletina, Panagrocephalus and
Panagrobelium. Several other scientists who contributed to morphology and
taxonomy of cephalobids were Fuchs (1930), Filipjev (1934), Timm (1956, 60,
71), Brezeski (1960) and Loof (1962) who added few more genera and species to
the group.
Heyns (1962) published a series of papers on cephalobids and proposed a
superfamily Elaphonematoidea with family Elaphonematidae for the genus
Elaphonema. He also proposed family Osstellidae with subfamily Osstellinae for
the genus Osstella. In 1968 he described a genus Paracrobeles. Nesterov (1970)
proposed a genus Acromoldavicus and redescribed Acrobeloides skrjabini. Allen
and Noffsinger (1971, 72) revised the genus Zeldia and added few species
besides an identification key. They also proposed a new genus Nothacrobeles
under Acrobelinae and described four new species viz. N. sheri, N. lepidus, N.
maximus and N. subtilus and transferred Zeldia acrobeles to Nothacrobeles
acrobeles as a new combination.
Boström (1984-2000) worked extensively on the taxonomy of
cephalobids. He (1984a, b) described morphological variability of Chiloplacus
minimus and compared the morphological features of three species of
Eucephalobus viz., E. striatus, E. oxyuroides and E. mucronatus by light and
scanning electron microscopy. In 1985, he described four new species viz.
Acrobeles oosiensis, Zeldia brevicauda, Cervidellus neftasiensis, C. serratus,
redescribed Acrobeloides emarginatus (de Man, 1880) Thorne, 1937 and
proposed a new genus Acrolobus. He (1988a, b) further described Cervidellus
16

spitzbergensis, Acrobelophis minimus, Acrobeloides tricornis, Eucephalobus
articus and conducted the morphological and systematic studies to investigate the
structure and function of labial probolae of the family Cephalobidae. In 1989, he
gave description of three populations of Pangrolaimus viz. P. superbus, P.
rigidus and P. detritophagus. He (1990, `91, `92) reported Heterocephalobellus
putamiensis, Seleborca complexa, Zeldia punctata, Acrobeloides ciliatus and
Cervidellus serratus from the soil samples from Greece. Boström (1993a, b)
described Cephalobus persegnis and Eucephalobus striatus from Ireland and E.
hooperi and Acrobeloides nanus from Malaysia. He (1995) described
Panagrolaimus magnivulvatus from Antarctica and in 2000, he reported a
divergent population of Cervidellus capraeolus (De Ley, Geraert & Coomans,
1990) Boström & De Ley, 1996 from Bahamas.
Rashid, Geraert & Sharma (1984) also contributed to Cephalobina by
working on their morphology, taxonomy and systematics. They proposed two
new genera Cephalonema and Heterocephalobellus with C. longicauda and H.
brasilensis as their type respectively under the family Cephalobidae. They
described two new species viz. Heterocephalobus tabacum and Cephalobus
pseudoparvus and also proposed three synonyms: Acrobeles capensis as a junior
synonym of A. mariannae, the genus Pseudocephalobus as a synonym of
Teratolobus and the family Alirhabditidae as a synonym of the Cephalobidae.
Rashid et al. (1989) synonymized Acrobelinae with Cephalobinae mainly on the
basis of presence or absence of labial probolae and indentations of the head
border.
17

De Ley et al. (1990-97) added a good number of species besides
publishing revision of genera and also revised the terminology of stoma
components by studying the ultrastructure of the stoma in Cephalobidae,
Panagrolaimidae and Rhabditidae. Siddiqi (1993) proposed five new genera and
eight new species of Cephalobina. He (2002) also described a new genus
Catoralaimellus with C. cornutus as its type and two new species of
Macrolaimellus viz., M. crassus and M. filumicus. Velde et al. (1994) elucidated
the ultrastructure of buccal cavity and cuticle in three species of cephalobs.
Morphology, oviposition and embryogenesis of Acrobeloides nanus was studied
by Bird et al. (1994). Vinciguerra (1994) reported new genus Metacrolobus
festonatus. Vinciguerra and Clausi (1996) reported two new species of
Acrobelophis viz. A. lanceolatus and A. fuegensis from soil in Argentina. A new
genus Penjatinema was described and morphology of P. natalense was discussed
by Heyns and Swart (1998). Clausi (1998) reported Cervidellus vinciguerrae
sp.n. from the soil samples around the moss plants in Argentina. Karegar et al.
(1998) described one new and one known species of Stegelletina and three
species of Cervidellus. Holovachov et al. (2001) described a new genus
Acroukrainicus with A. sagittiferus as its type species. Further, Holovachov &
Boström (2006) proposed two new genera viz. Panagrolobus and Deleyia under
subfamily Cephalobinae and described three species P. vanmegenae, D. poinari
and D. aspiculata. Abolafia and Peña-Santiago (2002, 2003, 2006, 2009) added
several new species to various genera and revised the genera, Acrobeloides,
Chiloplacus, Pseudacrobeles, Nothacrobeles, Panagrolaimus, Cephalobus and
also provided keys to species identification.
18

Nematode problems of various kinds must have existed since time
immemorial. In India work on Nematology has been started as one of the
disciplines of agricultural sciences. Barber (1901) was the first to record the
infestation of root-knot nematode on tea in South India. During the period 1901-
1958 there had been very little progress, though there were historical breaking
discoveries (Barber, 1901; Butler, 1906, 1913, 1919; Ayyar, 1926, 1933; Dastur,
1936). Brief historical perspectives of the growth and development of
nematology in India have been reported by various authors in the past (Swarup et
al., 1967; Seshadri, 1965; Swarup and Seshadri, 1974).
Organized research on plant nematodes started only after the end of 1950.
The 1960s could be regarded as the most active phase for the growth of
nematology in general and nematode taxonomy in particular in India. Primarily,
nematological research in India has focussed more on the plant parasitic and
animal parasitic groups. Little work has been done on the free-living group-
Rhabditida, probably because they have no direct concern with agriculture and
livestock. Only a few scientists have worked on this heterogenous group during
the late 60s and 70s. One of the earliest reports was the description of Tridontus
longicaudatus Khera, 1965. Khera (1969) described Mesodiplogasteroides,
Operculorhabditis, Saprorhabditis and Praeputirhabditis and in 1970 he further
described two new genera viz. Paradoxogaster and Gobindonema. Later in 1971
he described Paradoxorhabditis in the subfamily protorhabditinae. Suryawanshi
(1971) described Tawdenema and Syedella which later on synonimized with
Acrostichus and Pareudiplogaster respectively. Jairajpuri et al. (1973)
redescribed Tridontus longicaudatus (Mononchoides longicaudatus) and
19

20
synonymised Syedella with Tridontus. Tahseen et al. (2005) and Ahmad et al.
(2007) described the new genera Metarhabditis and Sclerorhabditis respectively.
Recently, Ahmad et al. (2010), Mahamood & Ahmad (2009) added new species
under the genera Mesorhabditis, Diplogasteroides and Rhabditidoides
Similarly the studies on the taxonomy of cephalobid nematodes has not
been carried out extensively. Khera (1968), described the genus Acrobelinema (=
Chiloplacus) with A. cornis as its type species. Suryawanshi (1971) proposed the
genus Alirhabditis and erected the subfamily Alirhabditinae for this genus. Joshi
(1972) reported a new genus Pseudocephalobus (= Teratolobus) from
Marathwada. Ali et al. (1973) described two new species of Drilocephalobus and
proposed a new family Drilocephalobidae with a revised classification of the
superfamily Cephaloboidea. Rathore and Nama (1992) described two new
species viz. Acrobeloides conoidis and Chiloplacus jodhpurensis from Jodhpur..
Recently, Tahseen et al. (1999) made the morphometrical observations on the
populations belonging to subfamily Acrobelinae viz. Zeldia punctata,
Chiloplacus subtenius and Seleborca complexa through scanning electron
microscopy.
The present work has been divided into two parts. Part A deals with the
taxonomy and biodiversity of free living nematodes. The nematodes of suborder
Cephalobina have been described in this part. Cephalobids are typically the most
diverse and abundant microbivores found in the soils world wide. This group is
interesting because they were equally abundant in both type of habitats studied
(i.e. crop fields and wastelands) during present work. In India the work on the
taxonomy of this group has not been carried out extensively, so in the present

work the taxonomy is restricted to the nematodes of suborder Cephalobina. A
total of ten new and three already known species have been described and
illustrated. In Part B ecological aspects like nematode community structure and
ecological indices, in two different types of habitats (i.e. crop field and wasteland)
have been analysed and compared. Different indices have been calculated for
diversity and food web studies.
20a

Materials &
Methods

Sampling: Samples for cephalobid nematodes were collected from agricultural
fields, wastelands, soil samples rich in organic debris, decayed and decaying
plant parts and farmyard manure from different parts of the country, especially
from Aligarh district and Eastern and Western Ghats of India. Some samples
from older collections were also screened. The samples were collected at regular
period throughout the year. The soil samples were taken from a depth of 10-20
cm and kept in airtight polythene bags. All relevant information such as host,
locality and date of collection were marked on the samples and these samples
were then brought to the laboratory for further processing.
Processing of soil samples: Samples were processed by modified Cobb’s (1918)
sieving and decantation and modified Baermann’s funnel techniques. From each
large sample, a sub-sample of about 500 cc was taken and mixed thoroughly with
water in a bucket taking care to remove debris and break the large clods and soil
crumbs. The bucket was then filled with water and the suspension was stirred to
make it homogenous. The mixture was kept undisturbed for about half a minute
so as to allow heavy matter to settle down to bottom of bucket. The suspension
was then passed into another bucket through a coarse sieve (2 mm pore size)
which retained large debris, roots and leaves etc. The suspension in the second
bucket was stirred thoroughly and was kept undisturbed for 30 seconds and then
poured through a fine sieve of mesh number 300 (pore size 53 µm). Nematodes
and very fine soil particles were retained on the sieve, the residue was then
collected in a beaker. This step was repeated 2 to 3 times for good recovery of
nematodes.
21

Isolation of nematodes: The residue collected in the beaker was poured on a
small coarse sieve lined with tissue paper. This sieve was then placed on a
Baermann’s funnel containing water sufficient to touch the bottom of the sieve.
Special care was taken to avoid trapping air bubbles at the bottom of the sieve.
The stem of the funnel was fitted with a rubber tubing provided with a stopper.
The nematodes migrated from the sieve into the clear water of the funnel and
accumulated at the bottom. After 24 hours, a small amount of water was drained
into a cavity block through the rubber tubing. The nematodes thus isolated were
fixed and processed for mounting on slides.
Killing and fixation: The collected nematodes in cavity blocks were left
undisturbed for a few minutes so as to allow them to settle down at the bottom.
Excess water was removed using a fine dropper and the hot FA fixative (8 ml of
40% commercial formaldehyde + 2 ml of glycerol + 90 ml of distilled water) was
poured into the cavity block. This act simultaneously killed and fixed the
nematodes.
Mounting and sealing: 24 hours after fixation, the nematodes were transferred
to a mixture of glycerine-alcohol (5 parts glycerine + 95 parts 30% alcohol) in a
cavity block, which was then in a desiccator containing anhydrous calcium
chloride. After 3 to 4 weeks the nematodes were dehydrated and were ready to be
mounted. A small drop of anhydrous glycerine was placed on a clean glass slide
and the nematodes were transferred from the cavity block into this drop. Either a
ring of wax was made or the pieces of wax were kept around the drop and a
22

circular glass cover slip was gently placed on the ring or pieces. This slide was
then heated on a hot plate. As the wax melted it sealed the drop of glycerine with
the nematodes.
Measurements and drawings: Measurements were made on specimens
mounted in dehydrated glycerine with an ocular micrometer. De Man’s (1884)
formula was used to denote the dimensions of nematodes. All morphological
observations, drawings and photographs were made on Olympus BX 50 and
Nikon 80i DIC microscopes.
Abbreviations used in the text
L = Total body length
a = Body length / greatest body diameter
b = Body length / distance from anterior end to the oesophago-intestinal junction
c = Body length / tail length
c´ = Tail length / anal body diameter
V = Distance of vulva from anterior end x 100 / body length
ABD = Anal body diameter
VBD = Vulval body diameter
Diam. = Diameter/diameters
23

Systematics

Order Rhabditida Chitwood, 1933
Diagnosis: Cuticle usually annulated, rarely ornamented with longitudinal striae
or punctations. Labial region mostly continuous, lips separate, three or six, often
with projections. Amphids small, inconspicuous, on lateral lips, exceptionally
large and more posterior in position. Mouth cavity of two main types: tuboid or
more or less spacious; in former case unarmed or possessing minute denticles, in
latter case usually provided with well developed teeth. Pharynx with either
median or terminal valvular bulb. Excretory pore visible. Intestine with wide
lumen. Three rectal glands generally present. Female reproductive system
didelphic or mono-prodelphic, in latter case generally with posterior uterine sac.
Ovaries reflexed. Ovi- or viviparous. Spicules ocassionaly fused. Male either
with paired genital papillae or with caudal bursa possessing paired rod like
papillae or ribs. Tail often different in each sex, without caudal; glands or
spinneret, but with distinct phasmids.
Type suborder: Rhabditina Chitwood, 1933
Other suborders: Cephalobina Andrassy, 1974
Diplogastrina Micoletzky, 1922
Myolaimina Inglis, 1983
Teratocephalina Andrassy, 1974
Suborder Cephalobina Andrassy, 1974
Diagnosis: Lips three or six, mostly separate; labial region often bearing
projections (probolae) of very various appearances. Amphids minute, pore-like,
on lateral lips. Mouth cavity tuboid, composed of six rings: cheilo-, gymno-, pro-,
24

meso-, meta- and telostom. The four latter (= stegostom) surrounded by
pharyngeal collar. Dorsal wall of metastom with a minute tooth-like projection.
Corpus and isthmus of pharynx well separated, terminal bulb possessing a well
developed grinder. Excretory pore distinct. Female genital system organ always
unpaired, prevulval, but ovary reflexed beyond the vulva. Spermatheca generally
present at anterior flexure of gonad. Predominantly oviparous. Spicules simple,
never fused, gubernaculum present. Male supplements papilloid, arranged in
pairs. No bursa. Tail generally short. Phasmids well discernible.
Type Superfamily: Cephaloboidea Filipjev, 1934
Other Superfamilies: Chambersielloidea Thorne, 1937
Panagrolaimoidea Thorne, 1937
Superfamily Cephaloboidea Filipjev, 1934
Diagnosis: Lip region varies from simple amalgamated type to those having
elaborate modified structures. Stoma quite narrow with uniformly sclerotized ring
elements, divided into three distinct sections, with or without metastomal tooth.
Oesophagus with or without grinders apparatus in the basal bulb. Female gonad
mono-prodelphic, reflexed, ovary extending beyond vulva, postvulval part in
most cases showing double flexures; spermatheca invariably present at anterior
flexure of gonad. Testis single, with reflexed terminal part. Spicules ventrally
curved with velum and capitulum. Gubernaculum present. Bursa absent. Genital
papillae present or absent.
Type family: Cephalobidae Filipjev, 1934
Other families: Elaphonematidae Heyns, 1962
Osstellidae Heyns, 1962
25

Family Cephalobidae Filipjev, 1934
Diagnosis: Cuticle annulated with sharply bordered lateral fields, occassionally
divided by longitudinal lines. Three cephalic probolae and three or six labial
probolae often present. Amphids located on lateral lips. Mouth cavity tuboid,
generally very narrow, ring elements small, uniformly sclerotized except for the
mostly unsclerotized gymnostom; cheilostom wider than the other rings. Dorsal
wall of metastom with minute tooth like projection. Pharynx consisting of usual
three sections, bulb strong. Vulval opening at two-thirds of body length, ovary
reflexed far posterior to vulva, its postvulval section in almost every case with
double flexures. Post-uterine sac present, generally short, rarely absent. Males in
general nearly as frequent as females. Phasmids distinct.
Type subfamily: Cephalobinae Filipjev, 1934
Other subfamilies: Acrobelinae Thorne, 1937
Acrolobinae De Ley, Siddiqi and Boström, 1993
Metacrobelinae Andrássy, 1974
Subfamily Cephalobinae Filipjev, 1934
Diagnosis: Lip region with simple cephalic and labial probolae, mostly
hexaradiate in symmetry, no deep clefts between lips. Cheilostom a broad
chamber, gymnostom short, dorsal metastegostom with a small tooth. Pharyngeal
corpus cylindrical, basal bulb with well developed grinder. Nerve ring usually
surrounding the base of corpus anterior to the isthmus. Female gonad single,
reflexed, ovary extending beyond vulva; spermatheca present at anterior flexure
of gonad. Males without bursa. Genital papillae present.
26

Type genus: Cephalobus Bastian, 1865
Other genera: Bunobus De Ley, Siddiqi & Boström, 1993
Eucephalobus Steiner, 1936
Heterocephalobellus Rashid, Geraert & Sharma, 1985
Heterocephalobus (Brzeski, 1960) Brzeski, 1961
Pseudacrobeles Steiner, 1938
Genus Pseudacrobeles Steiner, 1938
Diagnosis: Lateral fields with three incisures, fading out at or near the phasmids.
Lip region with hexaradiate, triradiate or bilateral symmetry and bearing 6+4
papilliform sensillae. Amphids small slits or oval pores at bases of lateral lips.
Lips separate or amalgamated; lateral lips may be reduced. Cephalic probolae
absent to short-setiform. Labial probolae absent to low knobs or ridges. Radial
ridges absent, tangential ridges present or absent. Mouth opening circular to
triangular, occasionally with small radial striae separating small liplet-like
structures but never extending deeply between the lips. Stoma with six sets of
sclerotizations. Cheilorhabdions comma-, bar- or granule- shaped in optical
section; cheilostom wide. Appearance of gymnostom in lateral view varying
from being as wide as cheilostom and having sclerotized rhabdia, to being as
narrow as stegostom and having inconspicuous rhabdia. Stegostom sections
clearly narrower than cheilostom. Females with post-uterine branch usually
developed, never surpassing ovary tip. Female tail sharp or blunt, conical, from
2.5 to 10 anal body diam. long. Male tail with or without mucro, with or without
27

extension of the body core beyond the posteriormost papillae. Gubernaculum
with cornua crurum never prominent.
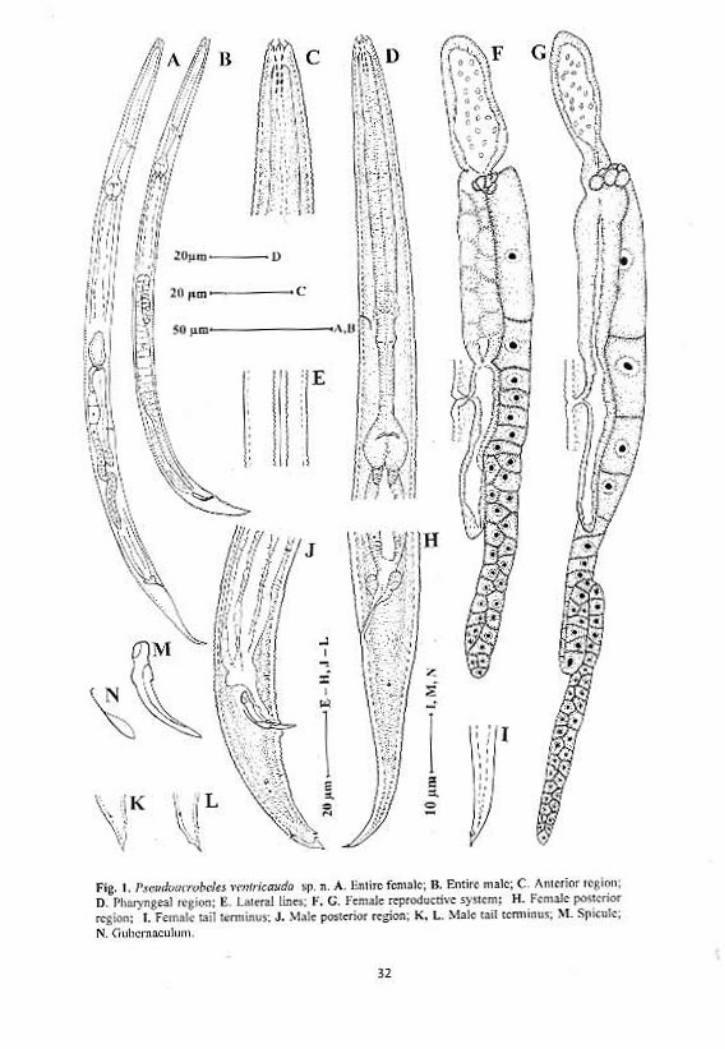
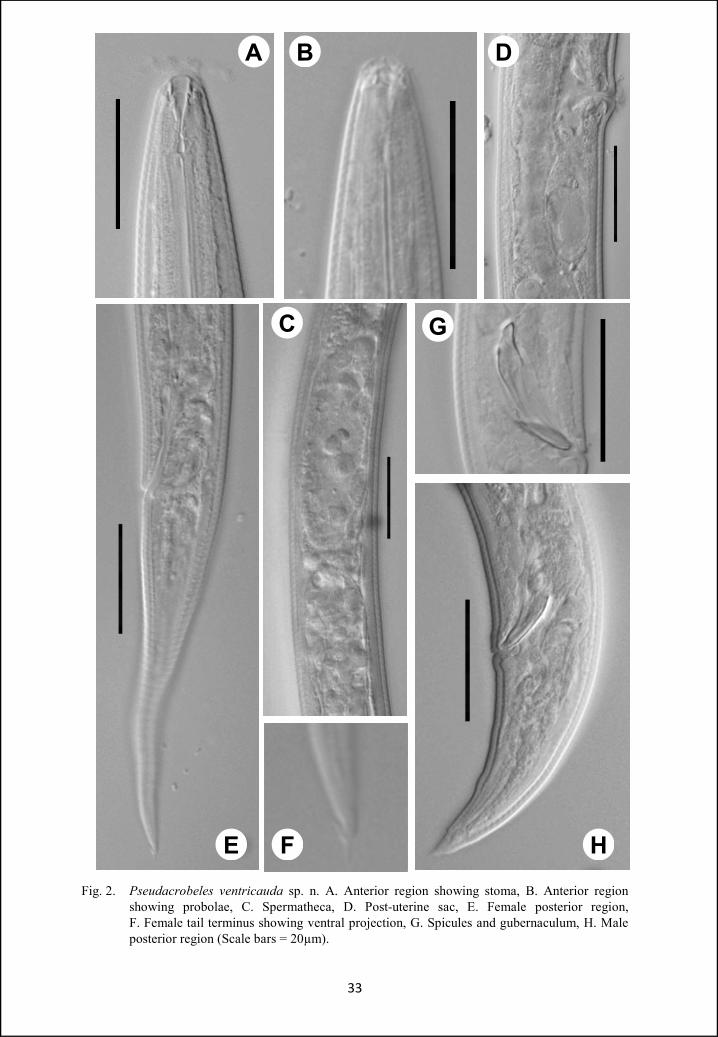
Pseudacrobeles ventricauda sp. n.
(Fig. 1, 2)
Measurements: In Table 1.
Females: Body slender, slightly ventrally curved upon fixation, tapering
gradually towards both the ends. Cuticle transversely annulated, annules 1.4-2.0
µm wide at midbody. Lateral fields with three incisures, usually indistinct. Lips
separated, lip region with 6+4 papillae. Cephalic probolae setiform. Labial
probolae present, low. Stoma cephaloboid. Cheilostom with bar-shaped rhabdia;
gymnostom intermediate between cheilostom and stegostom in width and degree
of sclerotization. Stegostom narrow with weakly sclerotized rhabdia,
metastegostom with a minute dorsal tooth . Pharyngeal corpus cylindrical, 4.5-5.5
times isthmus length. Isthmus visibly demarcated from corpus by transverse
markings. Basal bulb pyriform, with grinders. Nerve ring surrounding the
posterior part of the corpus, at 65-70% of neck length. Excretory pore opposite
nerve ring, at 66-71% of neck length. Hemizonid and excretory pore are at the
same level. Intestine with distinct wide lumen. Cardia conoid, enveloped by
intestinal tissue.
Reproductive system mono-prodelphic. Ovary reversed, on right side of
intestine, with a double flexure or sometimes without any flexure posterior to
vulva. Oocytes arranged in one or more rows in the germinal zone and in single
or double row in maturation zone. Oviduct short. Spermatheca well developed
1.3 – 2.6 times corresponding body diam. long, containing spermatozoa. Uterus
28

well developed about 2-3 times the corresponding body diam. long, sometimes
with single ova in the lumen. Post-uterine sac 1.2-1.8 times the corresponding
body diam. long. Vagina thick-walled, 0.25-0.33 times the vulval body diam. Tail
elongate-conoid gradually tapering to a sharply pointed tip. A small subterminal
ventral projection is also present near the tail tip.
Males: General appearance similar to that of females. Habitus ventrally curved,
more in the posterior region. Reproductive system monorchic. Testis with
anterior ventral flexure on right side of intestine. Tail conical bearing an acute
mucro. Genital papillae eight pairs; two pairs precloacal (subventral), one
adcloacal (subventral) and five pairs postcloacal. Of the five postcloacal pairs,
two pairs (one subventral and one lateral) are anterior to phasmid, one subdorsal
pair posterior to phasmid and two subventral pairs near the tail terminus. Spicules
with rounded manubrium, slightly arcuate lamina and rounded tip. Gubernaculum
with well developed crura.
Type habitat and locality: Sandy soil collected from a grass bed near a river close
to the Rushikonda beach, Vishakapatnam.
Type specimens
Holotype female on slide Pseudacrobeles ventricauda sp. n./1; ten
females and ten males (paratypes) on slides Pseudacrobeles ventricauda sp. n./2-
8 deposited in the nematode collection of Department of Zoology, Aligarh
Muslim University, Aligarh.
Diagnosis and relationship
Pseudacrobeles ventricauda sp. n. is characterized by setiform cephalic
probolae, Cheilostom with bar-shaped rhabdia. Post-uterine sac 1.2-1.8 times the
29

corresponding body diam. long. Vagina thick walled, 0.25-0.35 times the vulval
body diam. Female tail elongate-conoid, with a small subterminal ventral
projection and acute tail terminus. Male tail conical, with fine terminal mucro.
The new species resembles Pseudacrobeles variabilis (Steiner, 1936)
Steiner, 1938 in general morphological characters and body size but differs from
it in having relatively longer post-uterine branch (36-47 µm vs 16-35 µm),
smaller c´ value in females (3.7-4.4 vs 4.6-7.2) and in the shape of female tail
(tail with a sub-terminal ventral projection vs tail terminus straight). The new
species also resembles P. tabacum (Rashid, Geraert & Sharma, 1985) De Ley,
Siddiqi & Boström, 1993 in morphological details. However, the new species
differs from P. tabacum in having comparatively longer pharynx in males (134-
156 µm vs 115-133 µm long), slightly smaller b-value both in males and females
(3.5-4.0 vs 4.0-4.5 in males and 3.4-4.1 vs 4.2-5.0 in females), longer post-uterine
branch 36-47 µm vs 22-31 µm), in the shape of female tail (tail terminus with a
ventral projection vs tail terminus without projection) and slightly smaller
spicules (19-21 µm vs 21-25 µm).
30
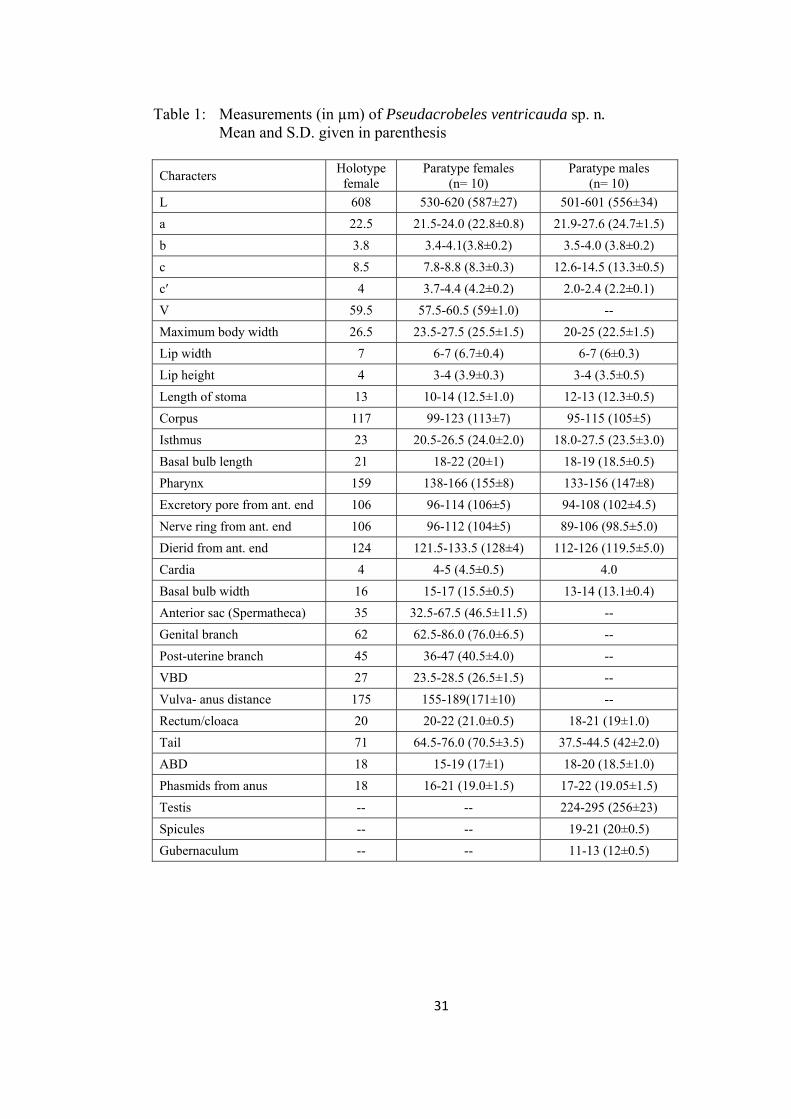
Table 1: Measurements (in µm) of Pseudacrobeles ventricauda sp. n. Mean and S.D. given in parenthesis
Characters Holotype
female Paratype females
(n= 10) Paratype males
(n= 10)
L 608 530-620 (587±27) 501-601 (556±34)
a 22.5 21.5-24.0 (22.8±0.8) 21.9-27.6 (24.7±1.5)
b 3.8 3.4-4.1(3.8±0.2) 3.5-4.0 (3.8±0.2)
c 8.5 7.8-8.8 (8.3±0.3) 12.6-14.5 (13.3±0.5)
c′ 4 3.7-4.4 (4.2±0.2) 2.0-2.4 (2.2±0.1)
V 59.5 57.5-60.5 (59±1.0) --
Maximum body width 26.5 23.5-27.5 (25.5±1.5) 20-25 (22.5±1.5)
Lip width 7 6-7 (6.7±0.4) 6-7 (6±0.3)
Lip height 4 3-4 (3.9±0.3) 3-4 (3.5±0.5)
Length of stoma 13 10-14 (12.5±1.0) 12-13 (12.3±0.5)
Corpus 117 99-123 (113±7) 95-115 (105±5)
Isthmus 23 20.5-26.5 (24.0±2.0) 18.0-27.5 (23.5±3.0)
Basal bulb length 21 18-22 (20±1) 18-19 (18.5±0.5)
Pharynx 159 138-166 (155±8) 133-156 (147±8)
Excretory pore from ant. end 106 96-114 (106±5) 94-108 (102±4.5)
Nerve ring from ant. end 106 96-112 (104±5) 89-106 (98.5±5.0)
Dierid from ant. end 124 121.5-133.5 (128±4) 112-126 (119.5±5.0)
Cardia 4 4-5 (4.5±0.5) 4.0
Basal bulb width 16 15-17 (15.5±0.5) 13-14 (13.1±0.4)
Anterior sac (Spermatheca) 35 32.5-67.5 (46.5±11.5) --
Genital branch 62 62.5-86.0 (76.0±6.5) --
Post-uterine branch 45 36-47 (40.5±4.0) --
VBD 27 23.5-28.5 (26.5±1.5) --
Vulva- anus distance 175 155-189(171±10) --
Rectum/cloaca 20 20-22 (21.0±0.5) 18-21 (19±1.0)
Tail 71 64.5-76.0 (70.5±3.5) 37.5-44.5 (42±2.0)
ABD 18 15-19 (17±1) 18-20 (18.5±1.0)
Phasmids from anus 18 16-21 (19.0±1.5) 17-22 (19.05±1.5)
Testis -- -- 224-295 (256±23)
Spicules -- -- 19-21 (20±0.5)
Gubernaculum -- -- 11-13 (12±0.5)
31
A
Fig. 2. Pseudacrobeles ventricauda sp. n. A. Anterior region showing stoma, B. Anterior region showing probolae, C. Spermatheca, D. Post-uterine sac, E. Female posterior region, F. Female tail terminus showing ventral projection, G. Spicules and gubernaculum, H. Male posterior region (Scale bars = 20µm).
33
B D
GC
E F H

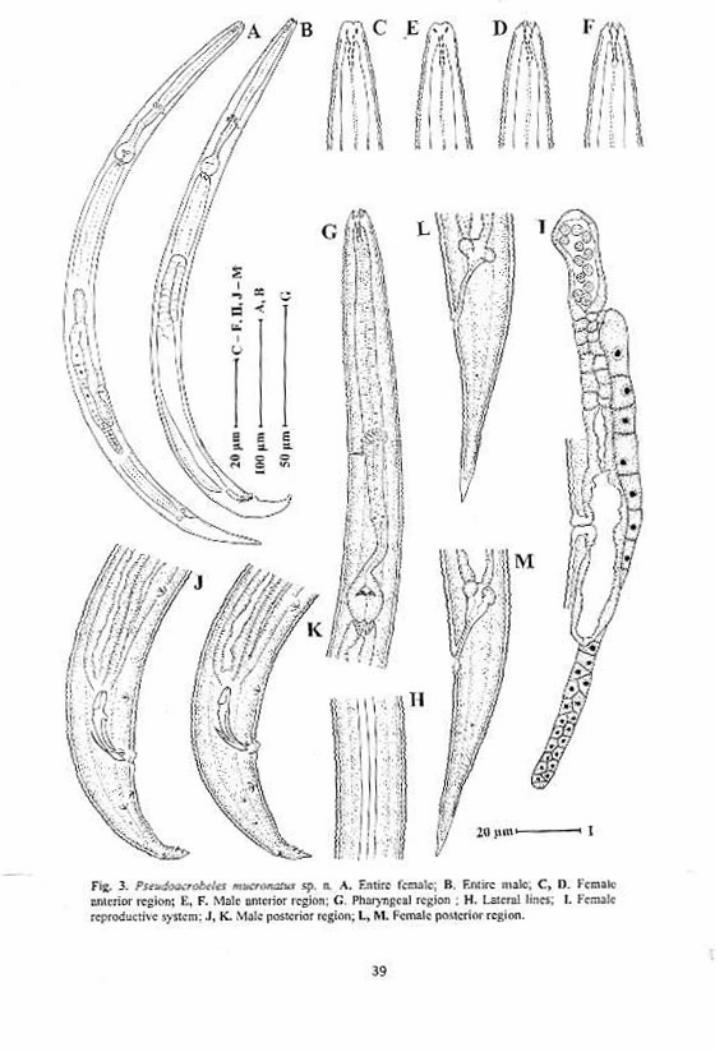
Pseudacrobeles mucronatus sp. n.
(Fig. 3, 4)
Measurements: In Table 2.
Females: Body slightly ventrally curved after fixation. Cuticle about 1.0-1.5 µm
thick at mid body, with transverse annules. Annules about 1.7-2.2 µm wide at
mid body. Lateral fields occupying 14-22% of mid body diam., with three
incisures. Lip region with triradiate symmetry. Cephalic probolae varying from
distinctly setiform to completely absent. Labial probolae varying from small but
distinct knobs, to flat ridges formed by partially fused lips. Stoma cephaloboid,
cheilorhabdia bar shaped. Gymnostom intermediate between cheilostom and
stegostom in width and degree of sclerotization. Stegostom with distinguished
rhabdia, dorsal metarhabdion with a small tooth like projection. Pharyngeal
corpus cylindrical, 4.8-7.5 times isthmus length. Isthmus short 16.0-22.5 µm
long, Basal bulb pyriform, with well developed valves. Nerve ring lying in the
corpus region, at 57-78% of pharynx length. Excretory pore at the level, or upto
five annules posterior to the trailing edge of nerve ring. Dierid 4-7 annules
posterior to excretory pore. Cardia short, conoid surrounded by intestinal tissue.
Reproductive system mono-prodelphic, ovary directed posteriorly, with
oocytes generally arranged in one or two rows in the germinal zone and in a
single row in maturation zone. Oviduct short. Spermatheca well developed 0.5-
1.5 times corresponding body diam long. Uterus tubular, differentiated into
anterior glandular part, and posterior muscular part with distinct lumen. Post-
uterine sac 0.8-1.3 vulval body diam. long. Vagina thick walled. Vulva transverse
34

slit like. Rectum 1.2-1.6 anal body diam. long. Tail elongate conoid, 3.5-4.5 anal
body diam. long, with acute terminus. Phasmid at 23-34% of tail length.
Males: Anterior region of males similar to that of females. Variability in the lip
region is also in concurrence with females. Habitus ventrally curved, more in the
posterior region giving it a ‘J’ shaped appearance. Reproductive system
monorchic. Testis ventrally reflexed anteriorly, with flexure on right side of
intestine. Tail conoid with mucronate tip. Mucro either small (with blunt tip) or
long (with pointed tip). Genital papillae eight pairs; two pairs precloacal
(subventral), one adcloacal (subventral) and five pairs postcloacal. Of the five
postcloacal pairs, two pairs (one subventral and one lateral) are anterior to
phasmid, one dorsal pair posterior to phasmid and two subventral pairs near the
tail terminus. Spicules cephaloboid, rounded manubrium, calamus slightly
narrower than manubrium, lamina ventrally curved, with acute terminus.
Gubernaculum with well developed crura.
Type habitat and locality: Decaying organic matter and leaf litter collected from
Indira Gandhi Zoological Park, Vishakapatnam, Andhra Pradesh, India.
Type specimens
Holotype female on slide Pseudacrobeles mucronatus sp. n./1; nine
females and six males (paratypes) on slides Pseudacrobeles mucronatus sp. n. /2-
6, deposited in the nematode collection of Department of Zoology, Aligarh
Muslim University, Aligarh.
Diagnosis and relationship
Pseudacrobeles mucronatus sp. n. is characterized by three incisures in
lateral fields. Cephalic probolae varying from distinctly setiform to completely
35

absent. Labial probolae varying from small but distinct knobs to flat ridges.
Cheilorhabdia bar shaped. Gymnostom intermediate between cheilostom and
metastegostom in width and degree of sclerotization. Pharyngeal corpus
cylindrical, 4.8-7.5 times isthmus length. Males with mucronate tail tip. Mucro
either small (with blunt tip) or long (with pointed tip). Gubernaculum with well
developed crura.
The new species closely resembles Pseudacrobeles variabilis (Steiner,
1936) Steiner, 1938 in general morphological characters and body size. However,
new species differs from it in having smaller c´ value in females (3.6-4.5 vs 4.8-
7.2), larger corpus to isthmus ratio (4.8-7.5 vs 3.1-4.5 in females and 5.7-6.4 vs
3.1-4.2 in males) and slightly longer gubernaculum (12-14 µm vs 10-12 µm). The
new species also resembles P. baloghi Andrassy, 1968 in body size, shape of lip
region and morphometric values. However, the present species differs from P.
baloghi in having longer body in males (0.52-0.58 mm vs 0.4-0.51), smaller c´
value (2.0-2.4 vs 2.8-3.7), larger corpus to isthmus ratio (4.8-7.5 vs 2.9-4.2 in
females and 5.7-6.4 vs 3.2-4.0 in males), more posterior position of phasmid in
males (20-24 vs 13-19), in shape of male tail (mucronate tail tip vs tail tip with
body core extending in spike for 4-11µm) and slightly longer gubernaculum (12-
14 vs 10-12).
P. mucronatus sp. n. closely resembles P. tabacum Rashid et al., 1985 in
general morphometrics and morphology. However, differences were traced in the
shape of cheilostom (never in granular form vs bar to granule form), longer
pharynx in males (150-163 vs 115-131), smaller b value (3.2-4.0 vs 4.2-5.0 in
females and 3.3-3.6 vs 4.0-4.6 in males), position of nerve ring in females (57-72
36

% vs 73-74 % of the pharynx length), larger corpus to isthmus ratio in males (5.7-
6.4 vs 4.1-4.7). The new species also resembles P. laevis Thorne, 1937 in body
size, structure of cephalic and labial probolae and other morphological details.
However, differences were found in ‘a’ value (20.7-23.5 vs 24-28 in females and
19.5-23.8 vs 25-29 in males), ‘c’ value of females (8.3-10.0 vs 11-14) and
corpus:isthmus ratio (4.8-7.5 vs 3.3-4.5 in females and 5.7-6.4 vs 3.2-4.1 in
males).
37
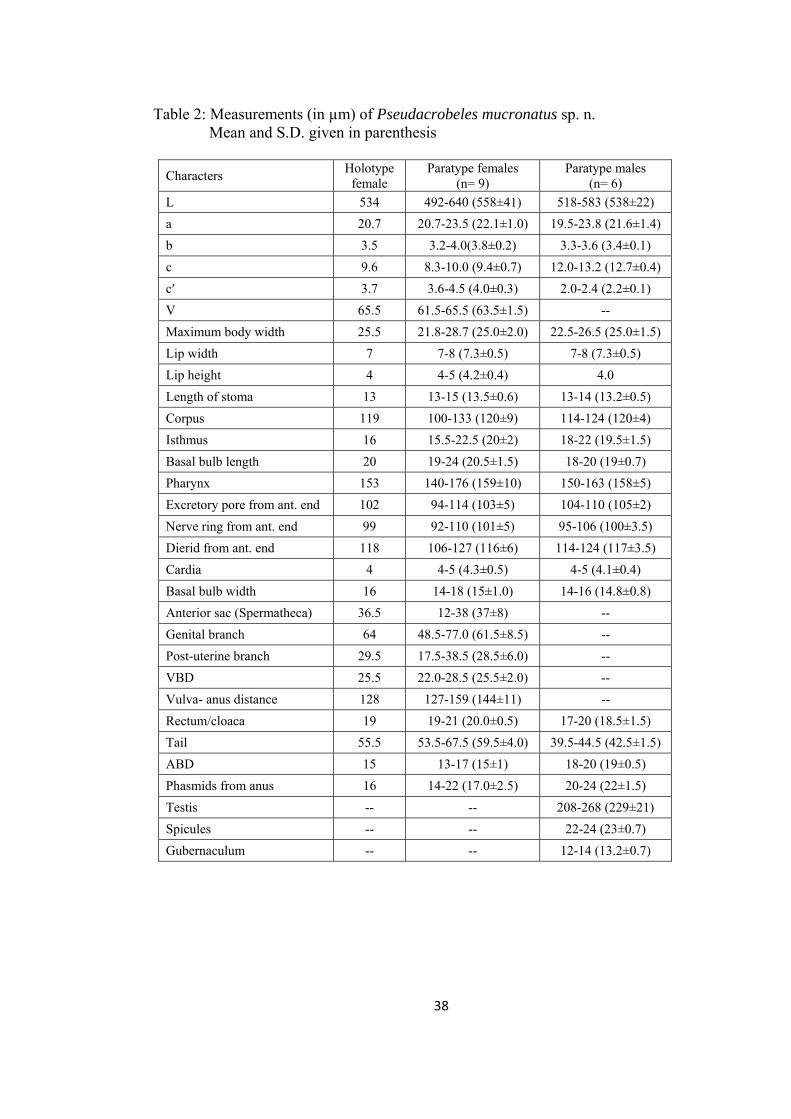
Table 2: Measurements (in µm) of Pseudacrobeles mucronatus sp. n. Mean and S.D. given in parenthesis
Characters Holotype
female Paratype females
(n= 9) Paratype males
(n= 6)
L 534 492-640 (558±41) 518-583 (538±22)
a 20.7 20.7-23.5 (22.1±1.0) 19.5-23.8 (21.6±1.4)
b 3.5 3.2-4.0(3.8±0.2) 3.3-3.6 (3.4±0.1)
c 9.6 8.3-10.0 (9.4±0.7) 12.0-13.2 (12.7±0.4)
c′ 3.7 3.6-4.5 (4.0±0.3) 2.0-2.4 (2.2±0.1)
V 65.5 61.5-65.5 (63.5±1.5) --
Maximum body width 25.5 21.8-28.7 (25.0±2.0) 22.5-26.5 (25.0±1.5)
Lip width 7 7-8 (7.3±0.5) 7-8 (7.3±0.5)
Lip height 4 4-5 (4.2±0.4) 4.0
Length of stoma 13 13-15 (13.5±0.6) 13-14 (13.2±0.5)
Corpus 119 100-133 (120±9) 114-124 (120±4)
Isthmus 16 15.5-22.5 (20±2) 18-22 (19.5±1.5)
Basal bulb length 20 19-24 (20.5±1.5) 18-20 (19±0.7)
Pharynx 153 140-176 (159±10) 150-163 (158±5)
Excretory pore from ant. end 102 94-114 (103±5) 104-110 (105±2)
Nerve ring from ant. end 99 92-110 (101±5) 95-106 (100±3.5)
Dierid from ant. end 118 106-127 (116±6) 114-124 (117±3.5)
Cardia 4 4-5 (4.3±0.5) 4-5 (4.1±0.4)
Basal bulb width 16 14-18 (15±1.0) 14-16 (14.8±0.8)
Anterior sac (Spermatheca) 36.5 12-38 (37±8) --
Genital branch 64 48.5-77.0 (61.5±8.5) --
Post-uterine branch 29.5 17.5-38.5 (28.5±6.0) --
VBD 25.5 22.0-28.5 (25.5±2.0) --
Vulva- anus distance 128 127-159 (144±11) --
Rectum/cloaca 19 19-21 (20.0±0.5) 17-20 (18.5±1.5)
Tail 55.5 53.5-67.5 (59.5±4.0) 39.5-44.5 (42.5±1.5)
ABD 15 13-17 (15±1) 18-20 (19±0.5)
Phasmids from anus 16 14-22 (17.0±2.5) 20-24 (22±1.5)
Testis -- -- 208-268 (229±21)
Spicules -- -- 22-24 (23±0.7)
Gubernaculum -- -- 12-14 (13.2±0.7)
38
A
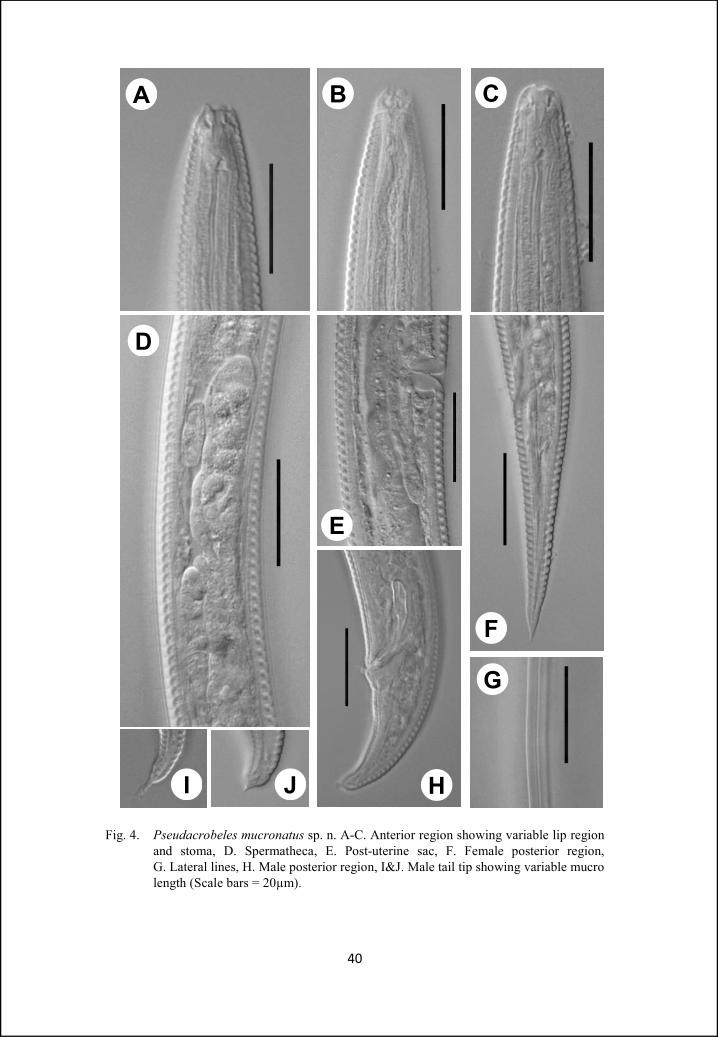
Fig. 4. Pseudacrobeles mucronatus sp. n. A-C. Anterior region showing variable lip region and stoma, D. Spermatheca, E. Post-uterine sac, F. Female posterior region, G. Lateral lines, H. Male posterior region, I&J. Male tail tip showing variable mucro length (Scale bars = 20µm).
40
B C
F
G
HJI
E
D

Subfamily Acrobelinae Thorne, 1937
Diagnosis: Lip region having cephalic probolae with complicated structures,
labial probolae large and exhibit triradiate symmetry, deep clefts present between
the lips. Cheilostom a broad chamber, gymnostom short, dorsal metastegostom
with a small tooth. Pharyngeal corpus cylindrical, basal bulb with well developed
grinder. Nerve ring usually surrounding the base of corpus or anterior half of
isthmus. Female gonad single, reflexed, ovary extending beyond vulva, straight
or with flexure posterior to vulva; spermatheca present at anterior flexure of
gonad. Males without bursa. Genital papillae present.
Type genus: Acrobeles Linstow, 1877
Other genera: Acrobeloides (Cobb, 1924) Thorne, 1937
Acrobelophis Andrassy, 1984
Acroukrainicus Holovachov, Boström & Susulovsky, 2001
Cervidellus Thorne, 1937
Chiloplacoides Heyns, 1994
Chiloplacus Thorne, 1937
Nothacrobeles Allen & Noffsinger, 1971
Paracrobeles Heyns, 1968
Pentjatinema Heyns & Swart, 1998
Placodira Thorne, 1937
Scottnema Timm, 1971
Stegelleta Thorne, 1938
Stegelletina Andrassy, 1984
Triligulla Siddiqi, 1993
Zeldia Thorne, 1937
41

Genus Acrobeles Linstow, 1877
Diagnosis: Body small to large (L=0.3-1.1 mm). Cuticle single or double, with
large annules, with or without longitudinal striae, punctations and/or pores.
Lateral field with two or three incisures, if cuticle double then often with
undulating internal pseudolines. Amphids relatively distinct, circular. Labial
probolae long, deeply bifurcated. Each prong with at least seven tines, its tip
usually with two elongate , separated apical tines.cephalic probolae high,
triangular, separate and fringed by numerous tines. Stoma cephaloboid with
distinct cheilorhabdia that are large and spherical in cross section. Pharyngea
corpus cylindrical to fusiform. Excretory pore position varying from very anterior
to opposite basal bulb. Female reproductive system cephaloboid, spermatheca
and post-uterine sac small to large. Vulva flush with body, occasionally sunken.
Males with three pairs of precloacal papillae, five pairs of postcloacal papillae
and one median papillae on the precloacal lip. Tails in both sexes conical, usually
with acute tip.
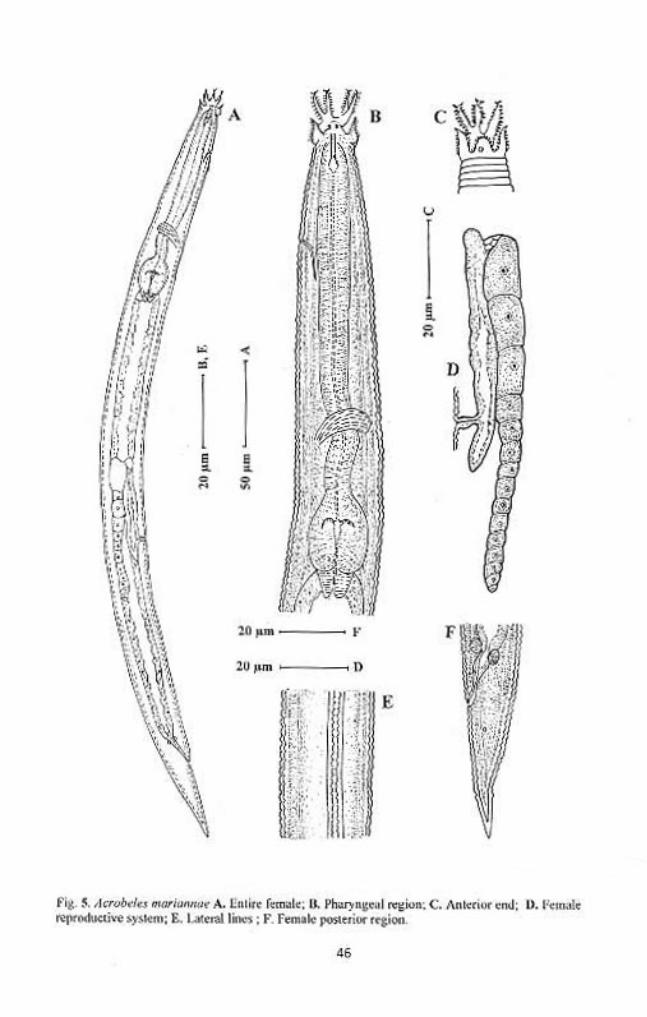
Acrobeles mariannae Andrassy, 1968
(Fig. 5)
Measurements: In Table 3.
Females: Body, straight to slightly curved ventrad, gradually tapering at both
extremities. Cuticle double, both layers similar, coarsely annulated. Annules
about 1.5-2.0 µm wide at mid body. Lateral fields with four incisures, slightly
elevated from body contour, outer incisures smooth, inner ones are crenate.
Labial probolae 7-11 µm high, bifurcated to about 50-60% of their length, each
42

arm bearing membranous tines varying in length and shape, rounded to rod-like,
and apical ones longer than the posterior ones. Inner sides with 6-8 tines and
outer sides with 7-9 tines, terminal tines ‘T’ shape. Cephalic probolae forming a
circle around the labials, each probola triangular, flap like, with 7 or 8 tines at
primary and secondary axil margins. Primary and secondary axils with similar
morphology (‘U’ shaped), with two guard processes each. Third tine of the
secondary axil is long and forwardly directed. Amphidial apertures rounded, at
the base of lateral cephalic probolae. Stoma cephaloboid, rhabdia distinctly
demarcated, cheilostom wide, with rounded rhabdia, gymnostom narrower than
cheilostom and as wide as stegostom, dorsal metarhabdion with tooth. Pharyngeal
corpus cylindrical 4-8 times longer than isthmus. Isthmus short, distinctly
separated from corpus by transverse marking. Basal bulb pyriform, with well
developed grinders. Nerve ring surrounding the corpus near corpus-isthmus
junction, at about 66-71% of pharynx length. Excretory pore far forward 26-38
µm from anterior end. Hemizonid in the posterior half of isthmus. Dierids at the
level of basal bulb. Cardia short, conoid surrounded by the intestinal tissue.
Intestine with small and granular cells, and wide lumen.
Reproductive system mono-prodelphic, ovary reversed, with or without
double flexure posterior to vulva. Oocytes arranged in one or two rows in the
germinal zone and in a single row in proliferative zone of ovary. Oviduct short.
Spermatheca reduced, less than the corresponding body diam. in length. Uterus
tubular, more than two corresponding diam. long, differentiated into a proximal
glandular and a distal muscular with thin walls. Post-uterine sac small, less than
the vulval body diam. in length. Vagina tubular, perpendicular to body axis, one
43

third of body diam. long. Vulva a transverse slit. Rectum 1.0-1.3 anal body diam.
long, anus an arcuate transverse slit. Tail conoid, gradually narrowing to an acute
terminus. Phasmids distinct, 33-39% or about one anal body diam. posterior to
anus.
Male: Not found.
Habitat and locality: Soil collected from the barren fields and road side ditches
near All India Radio station, Anoopshahr road, Aligarh.
Voucher specimen
Twelve females on slides Acrobeles mariannae/1-5 deposited in the
nematode collection of Department of Zoology, Aligarh Muslim University,
Aligarh.
Remarks
A. mariannae is a terrestrial species. It is widely distributed and is known
from The Netherlands, Hungary, Pakistan, Sudan, Kenya, Namibia, South Africa,
Brazil, Paraguay and Krakatau Islands, Hungary. This species is easily
differentiated from the other species of the genus by its small body and a very
anteriorly located excretory pore. The morphology and morphometric values of
our population of A. mariannae corresponds well with those of described
populations. A. mariannae is reported for the first time from India.
44
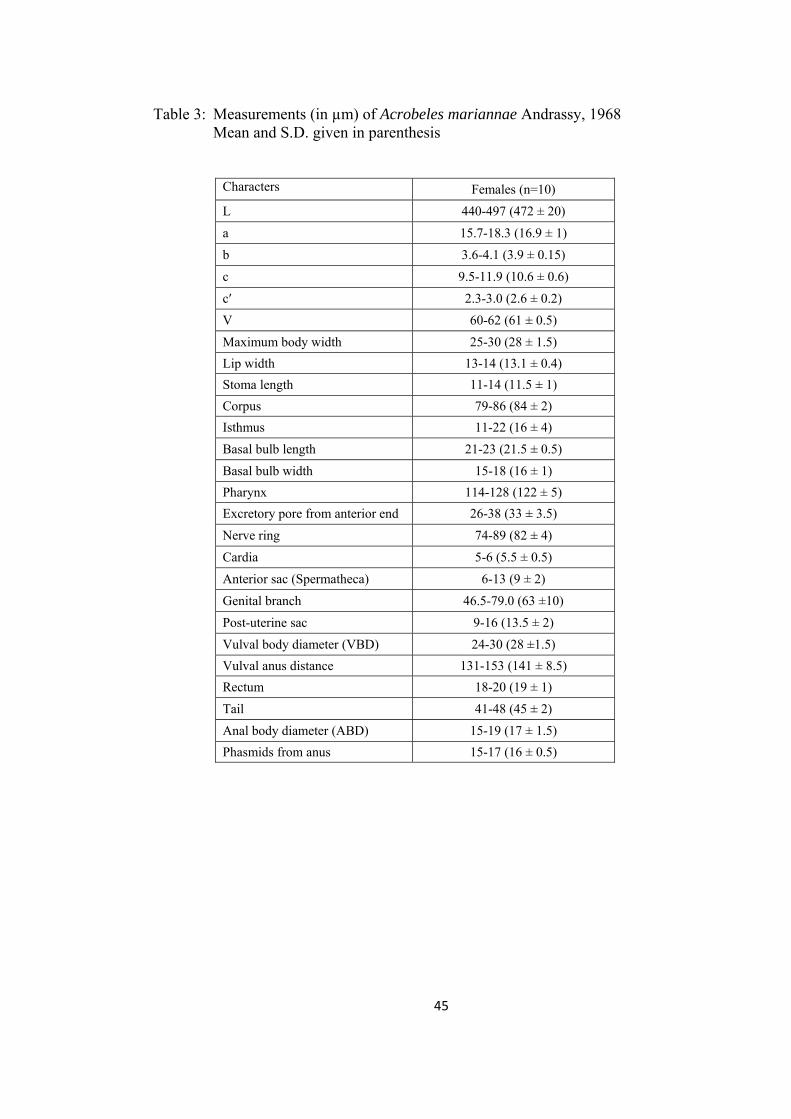
Table 3: Measurements (in µm) of Acrobeles mariannae Andrassy, 1968 Mean and S.D. given in parenthesis
Characters Females (n=10)
L 440-497 (472 ± 20)
a 15.7-18.3 (16.9 ± 1)
b 3.6-4.1 (3.9 ± 0.15)
c 9.5-11.9 (10.6 ± 0.6)
c′ 2.3-3.0 (2.6 ± 0.2)
V 60-62 (61 ± 0.5)
Maximum body width 25-30 (28 ± 1.5)
Lip width 13-14 (13.1 ± 0.4)
Stoma length 11-14 (11.5 ± 1)
Corpus 79-86 (84 ± 2)
Isthmus 11-22 (16 ± 4)
Basal bulb length 21-23 (21.5 ± 0.5)
Basal bulb width 15-18 (16 ± 1)
Pharynx 114-128 (122 ± 5)
Excretory pore from anterior end 26-38 (33 ± 3.5)
Nerve ring 74-89 (82 ± 4)
Cardia 5-6 (5.5 ± 0.5)
Anterior sac (Spermatheca) 6-13 (9 ± 2)
Genital branch 46.5-79.0 (63 ±10)
Post-uterine sac 9-16 (13.5 ± 2)
Vulval body diameter (VBD) 24-30 (28 ±1.5)
Vulval anus distance 131-153 (141 ± 8.5)
Rectum 18-20 (19 ± 1)
Tail 41-48 (45 ± 2)
Anal body diameter (ABD) 15-19 (17 ± 1.5)
Phasmids from anus 15-17 (16 ± 0.5)
45

Genus Acrobeloides (Cobb, 1924) Thorne, 1937
Diagnosis: Body length varying from 0.3-1.2 mm. cuticle annulated, lateral fields
with two to five incisures extending generally to tip of tail. Lips three, labial
probolae hemispheroid or conoid, point always uni-tipped. Cephalic probolae
present, but low, not strongly differentiated. Proximal half of oesophagous,
fusiform. Stoma cephaloboid, narrow. Vulva near two-thirds of body length,
ovary with double postvulval flexures. Males mostly unknown. Tail short and
plump, broadly rounded or conoid.
Acrobeloides glandulatus sp. n.
(Fig. 6, 7)
Measurements: In Table 4.
Females: Body cylindrical, tapering gradually towards both ends, slightly
ventrally curved after fixation. Cuticle with strong transverse annules, annuli 3-
4µm wide at midbody. Lateral fields 22-30% of the mid-body diam., with five
incisures; outer incisures crenate and irregularly aerolated. Lips six, amalgamated
in pairs, flattened. Primary axils distinct, with smooth margins. Secondary axils
scarcely demarcated. Three low and rounded labial probolae. Amphidial
apertures slit-like. Stoma cephaloboid. Cheilostom wide, with small and ovoid
rhabdia, gymnostom and stegostom slightly narrower than cheilostom,
metastegostom with a small tooth on dorsal rhabdia. Pharyngeal corpus
cylindrical, 4.7-7.5 times isthmus length. Corpus-isthmus junction distinguished
by transverse markings. Isthmus smaller than basal bulb. Basal bulb ovoid or
pyriform, with strongly developed grinder. Nerve ring surrounding the isthmus in
47

anterior half, or at 73-78% of neck length. Excretory pore at the level of basal
bulb, 77-92% of neck length. Dierids at level or slightly posterior of basal bulb
region. Cardia conoid, surrounded by intestinal tissue. Intestine with distinct wide
lumen, intestinal cells small, granular in appearance with distinct nuclei.
Reproductive system mono-prodelphic. Ovary reversed, straight or with
double flexure in its postvulvular part. Oocytes arranged in two or more rows in
the germinal zone. Oviduct short, tubular. Spermatheca 0.6-2.0 times the
corresponding body diam. long, with few sperms. Uterus long, differentiated in
an elongated proximal glandular part and a short distal muscular part with wide
and distinct lumen. Post-uterine sac 1-2 vulval body diam. long. Vagina with
thick walls, one-third of body diam. A pair of spheroid glands associated with the
vagina, each gland is arranged on either side of vagina, both glands seems to
open into the vaginal lumen through a common duct. Vulva transverse, lips
slightly protruded. Rectum less than one anal body diam. long. Tail conoid, 1.7-
2.1 anal body diam. long, with rounded terminus. Phasmid at 46 – 55 % of tail
length.
Males: General appearance similar to that of female, but with slightly smaller
body, ventrally curved in the posterior region adopting ‘J’ shape. Reproductive
system monorchic. Testis reflexed ventrally anteriorly on the right side of
intestine. Tail conoid, ventrally curved, with rounded terminus. Genital papillae
eight pairs; three pairs pre-cloacal (subventral), five pairs post-cloacal. Of the
five post-cloacal pairs two pairs (one subventral and one lateral) anterior to
phasmid, three pairs (one dorsal, one lateral and one subventral) near the tail
terminus. Spicules strong, small manubrium, calamus broad, slightly ventrally
48

curved lamina with pointed tip. Gubernaculum straight, more than half of spicule
length, narrow and pointed distally.
Type habitat and locality: Soil collected from a ploughed field at Nohati village,
Madrak, Aligarh.
Type specimens
Holotype female on slide Acrobeloides glandulatus sp. n./1, nine females
and eight males (paratypes) on slides Acrobeloides glandulatus sp. n./2-10
deposited in the nematode collection of Department of Zoology, Aligarh Muslim
University, Aligarh.
Diagnosis and relationship
Acrobeloides glandulatus sp. n. is characterized by a body length of 625-
825 µm in females and 558-717 µm in males. Lateral fields with five incisures.
Low and rounded labial probolae. Pharyngeal corpus 4.7-7.5 times isthmus
length. Well developed spermatheca (0.6-2.0 times corresponding body diam.
long) and long post-uterine sac (1-2 vulval body diam. long). Presence of one pair
of gland on either side of vagina. Spicules 41-48 µm long. Straight
gubernaculum, with narrow and pointed distal end.
The new species resembles Acrobeloides bodenheimeri (Steiner, 1936)
Thorne, 1937 in general morphological characters, morphometric values and
body size but differs from it in having longer post-uterine sac (57-72 µm vs 29-55
µm), variable shape of ovary (with or without double flexure vs with double
flexure), larger corpus-isthmus ratio (5-8 vs 3.8-4.2), longer spicules (41-48 µm
49

vs 35-39 µm) and in the shape of gubernaculum (straight and pointed distal tip vs
ventrally curved and blunt distal tip).
50
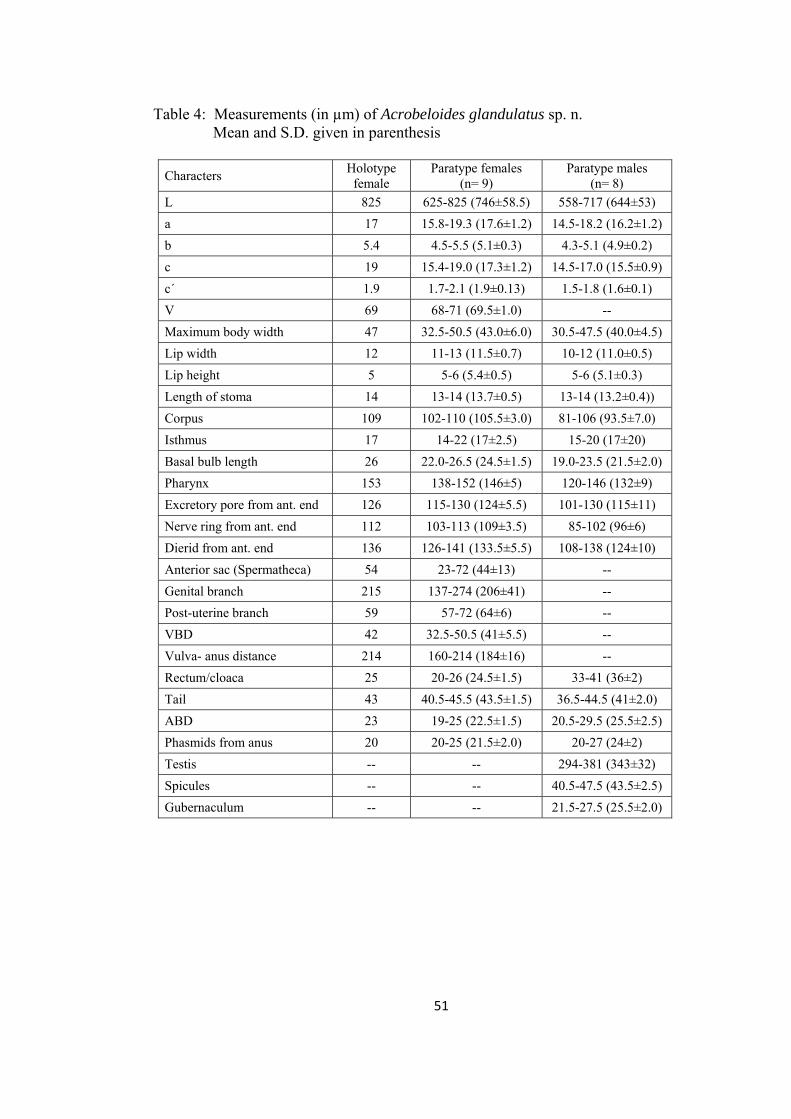
Table 4: Measurements (in µm) of Acrobeloides glandulatus sp. n. Mean and S.D. given in parenthesis
Characters Holotype
female Paratype females
(n= 9) Paratype males
(n= 8)
L 825 625-825 (746±58.5) 558-717 (644±53)
a 17 15.8-19.3 (17.6±1.2) 14.5-18.2 (16.2±1.2)
b 5.4 4.5-5.5 (5.1±0.3) 4.3-5.1 (4.9±0.2)
c 19 15.4-19.0 (17.3±1.2) 14.5-17.0 (15.5±0.9)
c´ 1.9 1.7-2.1 (1.9±0.13) 1.5-1.8 (1.6±0.1)
V 69 68-71 (69.5±1.0) --
Maximum body width 47 32.5-50.5 (43.0±6.0) 30.5-47.5 (40.0±4.5)
Lip width 12 11-13 (11.5±0.7) 10-12 (11.0±0.5)
Lip height 5 5-6 (5.4±0.5) 5-6 (5.1±0.3)
Length of stoma 14 13-14 (13.7±0.5) 13-14 (13.2±0.4))
Corpus 109 102-110 (105.5±3.0) 81-106 (93.5±7.0)
Isthmus 17 14-22 (17±2.5) 15-20 (17±20)
Basal bulb length 26 22.0-26.5 (24.5±1.5) 19.0-23.5 (21.5±2.0)
Pharynx 153 138-152 (146±5) 120-146 (132±9)
Excretory pore from ant. end 126 115-130 (124±5.5) 101-130 (115±11)
Nerve ring from ant. end 112 103-113 (109±3.5) 85-102 (96±6)
Dierid from ant. end 136 126-141 (133.5±5.5) 108-138 (124±10)
Anterior sac (Spermatheca) 54 23-72 (44±13) --
Genital branch 215 137-274 (206±41) --
Post-uterine branch 59 57-72 (64±6) --
VBD 42 32.5-50.5 (41±5.5) --
Vulva- anus distance 214 160-214 (184±16) --
Rectum/cloaca 25 20-26 (24.5±1.5) 33-41 (36±2)
Tail 43 40.5-45.5 (43.5±1.5) 36.5-44.5 (41±2.0)
ABD 23 19-25 (22.5±1.5) 20.5-29.5 (25.5±2.5)
Phasmids from anus 20 20-25 (21.5±2.0) 20-27 (24±2)
Testis -- -- 294-381 (343±32)
Spicules -- -- 40.5-47.5 (43.5±2.5)
Gubernaculum -- -- 21.5-27.5 (25.5±2.0)
51
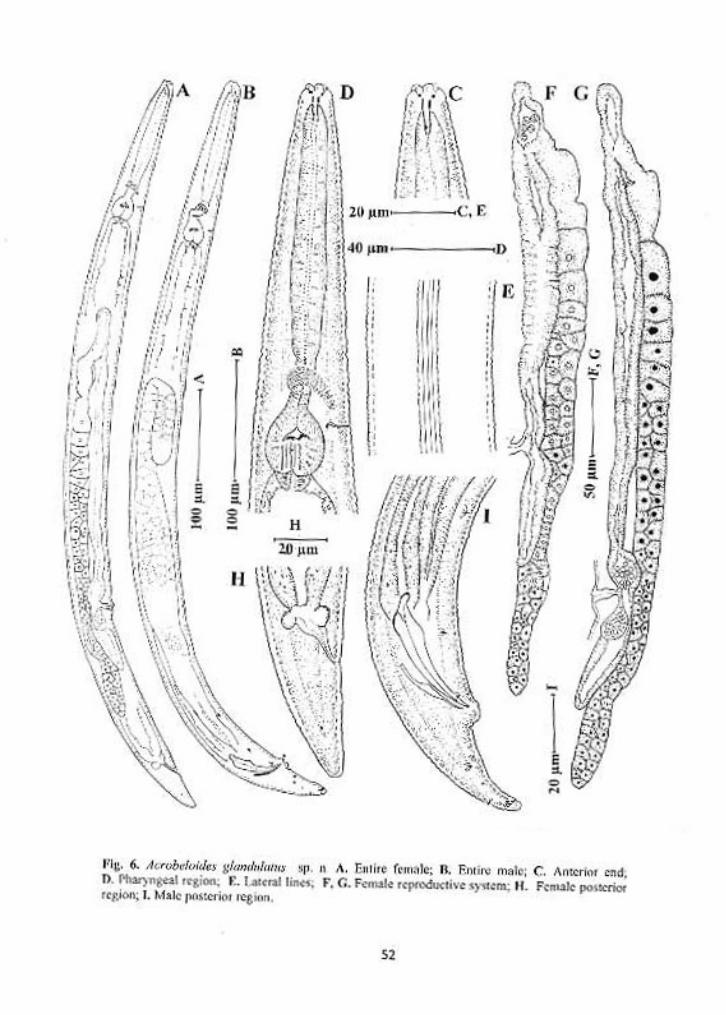
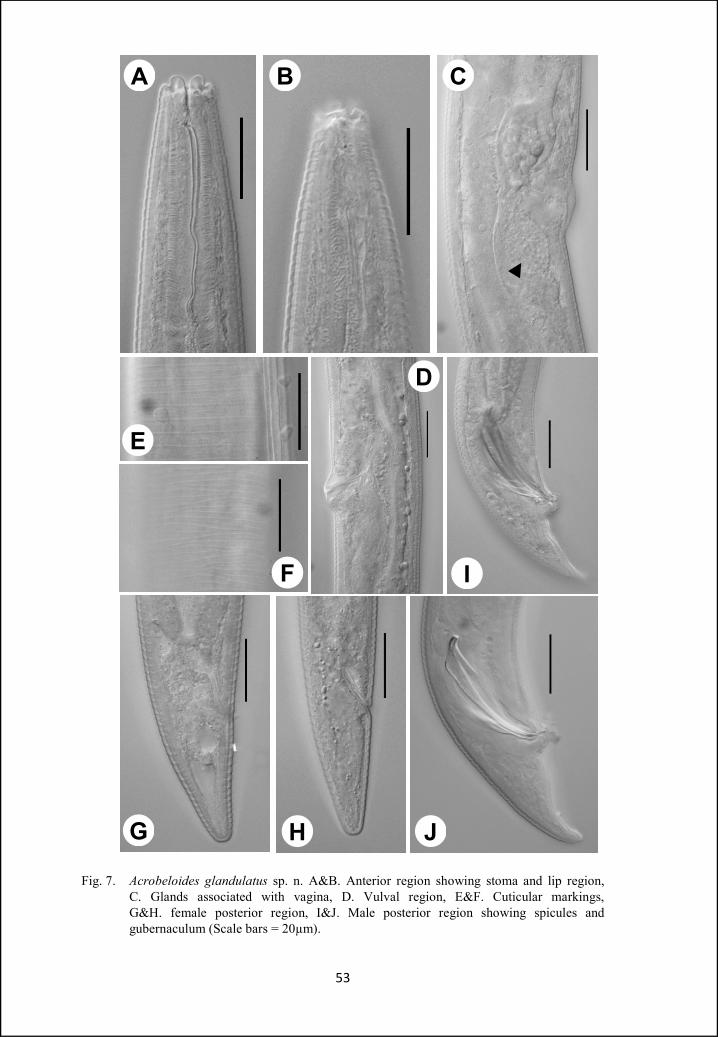
A B C
I
D
E
F
HG J
Fig. 7. Acrobeloides glandulatus sp. n. A&B. Anterior region showing stoma and lip region, C. Glands associated with vagina, D. Vulval region, E&F. Cuticular markings, G&H. female posterior region, I&J. Male posterior region showing spicules and gubernaculum (Scale bars = 20µm).
53

Genus Cervidellus Thorne, 1937
Diagnosis: Very small nematodes ranging from 0.3 to 0.5mm. Cuticle finely
annulated, lateral fields with two, three or five incisures. Lip margins with U-
shaped refractive elements, cephalic probolae six, triangular or leaf-like. Labial
probolae thin, Y-shaped, occasionally with secondary tines. Pharyngeal corpus
cylindrical. Female genital apparatus cephaloboid. Tails of both sexes conoid
with pointed tip.
Cervidellus neoalutus sp. n.
(Fig. 8, 9)
Measurements: In Table 5.
Females: Body slender, slightly ventrally curved after fixation, gradually tapering
towards both extremities. Cuticle double, transversely annulated, each annule
with two rows of punctations. Annuli 1.8-2.2 µm wide at pharyngeal region and
about 1.7-2.0 µm wide at midbody and tail region. Lateral fields with four
crenated incisures, without areolation, covering about 1/5th of body diam. at
midbody. Lateral lines start at 40-50% of pharynx length from anterior end as
two faint lines and differentiate into four lines at level of basal bulb. The two
inner incisures are more widely separated as compared to the outer ones,
appearing as two pairs of four lines. All incisures reach the tail terminus.
Cephalic region with six probolae, each consisting of five leaf-like elements: in
the centre, the longest and slightly clavate element; more outwards and on both
sides two pairs of shorter and acute elements. Labial probolae about 5µm long
with bifurcation at two levels, primary bifurcation at 2/3rd of probolae length,
54

secondary bifurcation apical. Amphidial apertures small, rounded. Stoma
cephaloboid. Cheilostom wide, with granular rhabdia. Gymnostom and stegostom
with less distinct rhabdia, narrower than cheilostom. Dorsal metastegostom with
a minute tooth like projection. Pharyngeal corpus cylindrical 2.5-3.0 times
isthmus length. Basal bulb pyriform or ovoid with well developed grinder. Nerve
ring at 63-71 % of neck length, surrounding isthmus in its anterior half. Excretory
pore at the level or slightly anterior to nerve ring. Dierids in the isthmus region, at
71-78% of neck length. Cardia rounded to conoid, surrounded by intestinal tissue.
Intestinal walls with differentiated rectangular cells, intestine with wide lumen.
Reproductive system mono-prodelphic. Ovary reversed, extending
posterior to vulva, postvulval flexure absent. Oocytes with large nuclei, arranged
in a single or double row in the germinal zone and in a single row in the
maturation zone. Oviduct short, spermatheca well developed, about 1.5 vulval
body diam. long. Uterus simple, tubular with narrow lumen and without any
differentiation. Post-uterine sac 1.0-2.5 times vulval body diam. long. Vagina
narrow, tubular, one-fourth of body diam. Vulval opening a small transverse slit.
Tail conical, with acute terminus. Phasmids distinctly visible, about 0.6-1.0 anal
body diam. posterior to anus.
Males: Anterior region similar to that of females. Reproductive system
monorchic. Testis reflexed ventrad anteriorly, on right side of intestine. Spicules
cephaloboid, manubrium round, calamus slightly narrower than manubrium,
lamina ventrally curved, with acute terminus. Gubernaculum straight, with well
developed crura. Genital papillae seven pairs, three pairs precloacal (subventral)
and four pairs postcloacal. Out of the four postcloacal papillae, two pairs (one
55

subventral & one lateral) anterior to phasmid and two pairs (one subdorsal & one
lateral) posterior to phasmid. A mid-ventral papilla on the anterior cloacal lip is
also present.
Type habitat and locality: Soil from a barren field near Central School, Bad
village, Mathura.
Type specimens
Holotype female on slide Cervidellus neoalutus sp. n./1, nine females and
three males (paratypes) on slides Cervidellus neoalutus sp. n./2-8 deposited in
the nematode collection of Department of Zoology, Aligarh Muslim University,
Aligarh.
Diagnosis and relationship
Cervidellus neoalutus sp. n. is characterized by a double cuticle, lateral
fields with four crenate incisures with inner ones more widely separated than
outer ones. Symmetrical lips, with refractive elements, labial probolae with
bifurcation at two levels and small post-uterine sac about 1.2-2 times vulval body
diam. in length.
The new species closely resembles C. alutus (Siddiqi, 1993) Shahina and
De Ley, 1997 in having double cuticle, symmetrical lips with refractive elements
but differs from it in the length of stoma (14-15 µm vs 10 µm), smaller post-
uterine sac (1.2 – 2 vs 2.3 - 2.6 times vulval body diam. long) and in the presence
of males (males absent in C. alutus).
56
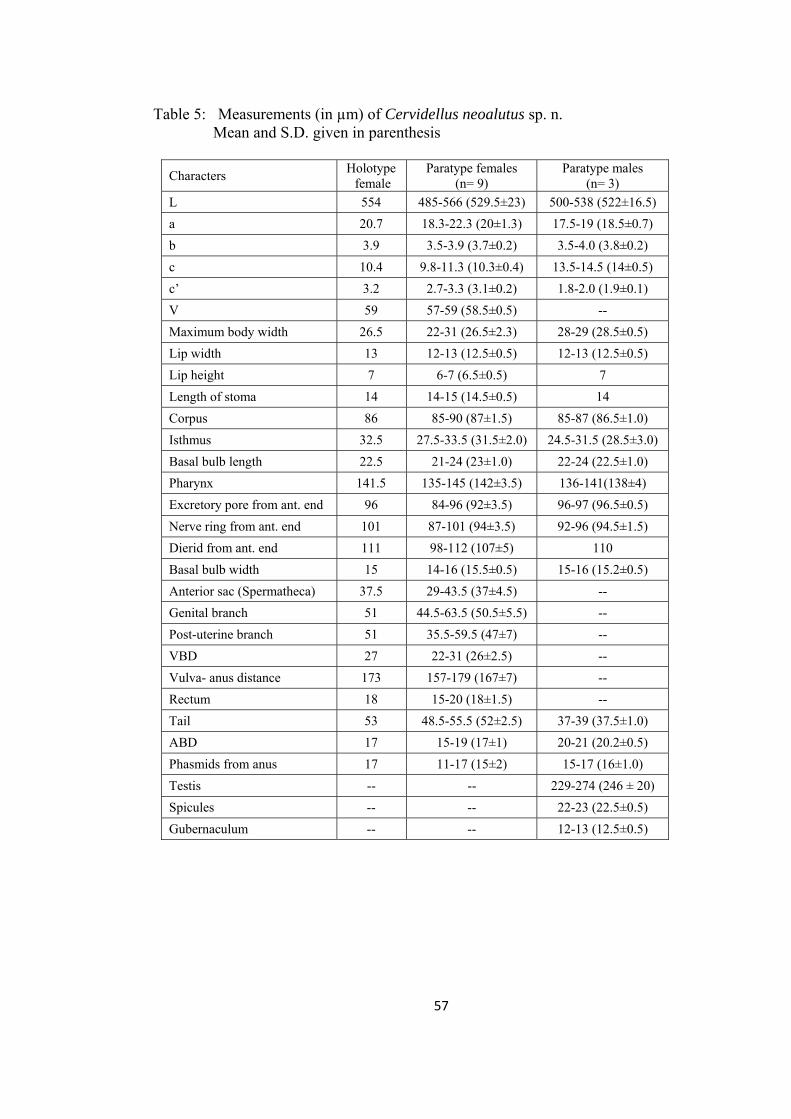
Table 5: Measurements (in µm) of Cervidellus neoalutus sp. n. Mean and S.D. given in parenthesis
Characters Holotype female
Paratype females (n= 9)
Paratype males (n= 3)
L 554 485-566 (529.5±23) 500-538 (522±16.5)
a 20.7 18.3-22.3 (20±1.3) 17.5-19 (18.5±0.7)
b 3.9 3.5-3.9 (3.7±0.2) 3.5-4.0 (3.8±0.2)
c 10.4 9.8-11.3 (10.3±0.4) 13.5-14.5 (14±0.5)
c’ 3.2 2.7-3.3 (3.1±0.2) 1.8-2.0 (1.9±0.1)
V 59 57-59 (58.5±0.5) --
Maximum body width 26.5 22-31 (26.5±2.3) 28-29 (28.5±0.5)
Lip width 13 12-13 (12.5±0.5) 12-13 (12.5±0.5)
Lip height 7 6-7 (6.5±0.5) 7
Length of stoma 14 14-15 (14.5±0.5) 14
Corpus 86 85-90 (87±1.5) 85-87 (86.5±1.0)
Isthmus 32.5 27.5-33.5 (31.5±2.0) 24.5-31.5 (28.5±3.0)
Basal bulb length 22.5 21-24 (23±1.0) 22-24 (22.5±1.0)
Pharynx 141.5 135-145 (142±3.5) 136-141(138±4)
Excretory pore from ant. end 96 84-96 (92±3.5) 96-97 (96.5±0.5)
Nerve ring from ant. end 101 87-101 (94±3.5) 92-96 (94.5±1.5)
Dierid from ant. end 111 98-112 (107±5) 110
Basal bulb width 15 14-16 (15.5±0.5) 15-16 (15.2±0.5)
Anterior sac (Spermatheca) 37.5 29-43.5 (37±4.5) --
Genital branch 51 44.5-63.5 (50.5±5.5) --
Post-uterine branch 51 35.5-59.5 (47±7) --
VBD 27 22-31 (26±2.5) --
Vulva- anus distance 173 157-179 (167±7) --
Rectum 18 15-20 (18±1.5) --
Tail 53 48.5-55.5 (52±2.5) 37-39 (37.5±1.0)
ABD 17 15-19 (17±1) 20-21 (20.2±0.5)
Phasmids from anus 17 11-17 (15±2) 15-17 (16±1.0)
Testis -- -- 229-274 (246 ± 20)
Spicules -- -- 22-23 (22.5±0.5)
Gubernaculum -- -- 12-13 (12.5±0.5)
57

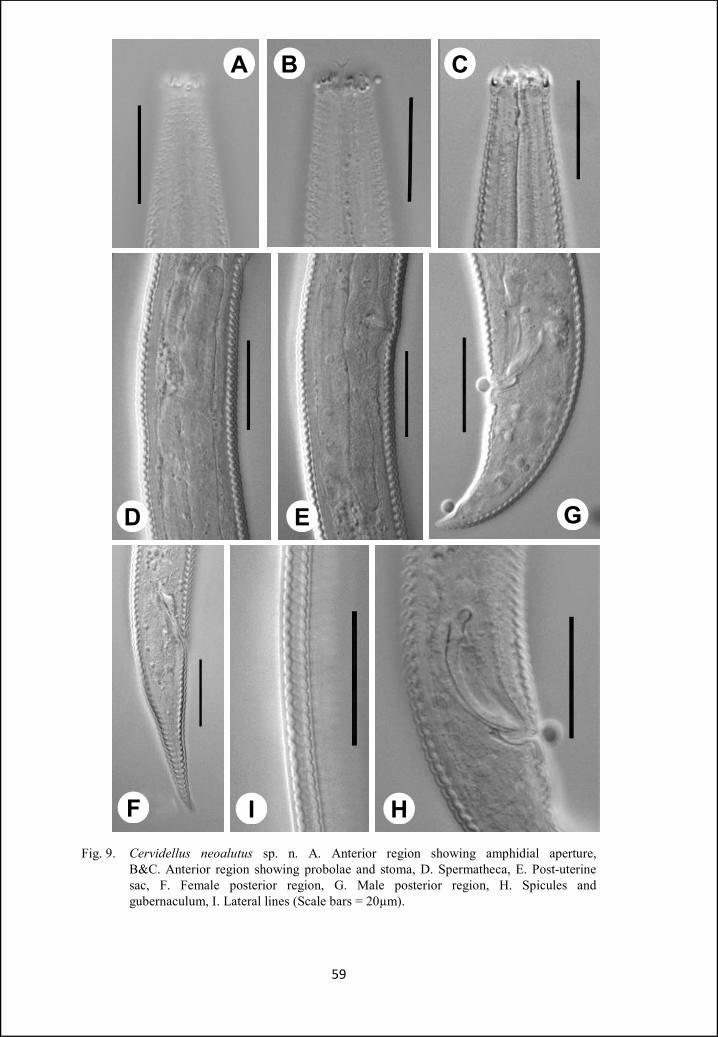
Fig. 9. Cervidellus neoalutus sp. n. A. Anterior region showing amphidial aperture, B&C. Anterior region showing probolae and stoma, D. Spermatheca, E. Post-uterine sac, F. Female posterior region, G. Male posterior region, H. Spicules and gubernaculum, I. Lateral lines (Scale bars = 20µm).
59
A B C
GED
F I H

Cervidellus minutus sp. n.
(Fig. 10, 11)
Measurements: In Table 6.
Females: Body small, slightly ventrally curved after fixation. Cuticle simple, with
transverse annulations. Lateral fields 12-14% of mid-body diam. wide, with three
incisures. Outer incisures crenate extending up to tail tip, central incisure ends at
phasmids. Cephalic probolae six, with refractive margins, each consisting of five
leaf- like elements. Primary and secondary axils similar in shape. Labial probolae
reduced, rarely visible in light microscope. Amphidial apertures small, ovoid.
Cheilostom with granular rhabdia, gymno- and stego-rhabdions poorly
developed. Pharyngeal corpus slightly fusiform, sometimes with wide lumen, 3.1-
3.7 times isthmus length. Corpus-isthmus junction distinguished by faint
transverse markings. Basal bulb spheroid, with well developed grinder. Nerve
ring at 56-61% of neck length, encircling isthmus in its anterior half. Excretory
pore at the level or just posterior to corpus-isthmus junction. Excretory duct
distinctly sclerotized. Dierids in the isthmus region or at the level of basal bulb.
Intestine with wide lumen. Cardia conoid about 2-3µm long, surrounded by
intestinal tissue.
Reproductive system mono-prodelphic, on right side of intestine. Ovary
reversed, without flexure posterior to vulva. Oocytes arranged in a single or
double row in germinal zone and in a single row in proliferative zone. Oviduct
short, not clearly demarcated. Spermatheca weakly developed, smaller than the
corresponding body diam. Uterus uniform in thickness, without any
differentiation between glandular and muscular part, lumen not visible. Post-
60

uterine sac less than one vulval body diam. long. Vagina straight, about 1/3rd of
body diam. with slightly thickened walls. Vulva transverse, slit like. Rectum
slightly more than one anal body diam. long. Rectal glands barely visible. Tail
conical, about 2-3 anal body diam. long, terminus acute, with subterminal dorsal
projection. Phasmids with distinct opening, about half anal body diam. posterior
to anus.
Male: Not found.
Type habitat and locality: Collected from the rhizosphere of Guava (Psidium
guajava) and Cashew-nut (Anacardium occidentale), in mixed plantation, from
Satpada, near Chilka lake, Orrisa, India.
Type specimens
Holotype female on slide Cervidellus minutus sp. n./1 and five females
(paratypes) on slides Cervidellus minutus sp. n./2-4 deposited in the nematode
collection of Department of Zoology, Aligarh Muslim University, Aligarh.
Diagnosis and relationship
Cervidellus minutus sp. n. is characterized by its small body (226-266
µm), lateral fields with three incisures. Lips with refractive margins. Reduced
labial probolae. Distinctly sclerotized excretory duct. Small spermatheca (5-6
µm) and post-uterine sac (less than one anal body diam.). Conoid tail, with acute
terminus and hook-like subterminal dorsal projection.
The new species differs from all other species of the genus in having a
subterminal dorsal projection in female tail. It, however, closely resembles C.
neftasiensis Boström, 1986 and C. vexilliger (de Man, 1880) Thorne, 1937 in
general morphometrics, shape of labial region, number of lateral lines. It can be
61

differentiated from C. neftasiensis in having comparatively smaller body length
(226-266 µm vs 268-330 µm), smaller spermatheca (5-6µm vs 13-28µm), and
relatively smaller genital branch (26-36µm vs 37-71µm). C. minutus sp. n. also
differs from C. vexilliger in having smaller spermatheca (5-6 µm vs 11-37 µm)
and relatively smaller post-uterine branch (7-8 µm vs 8-47 µm).
62
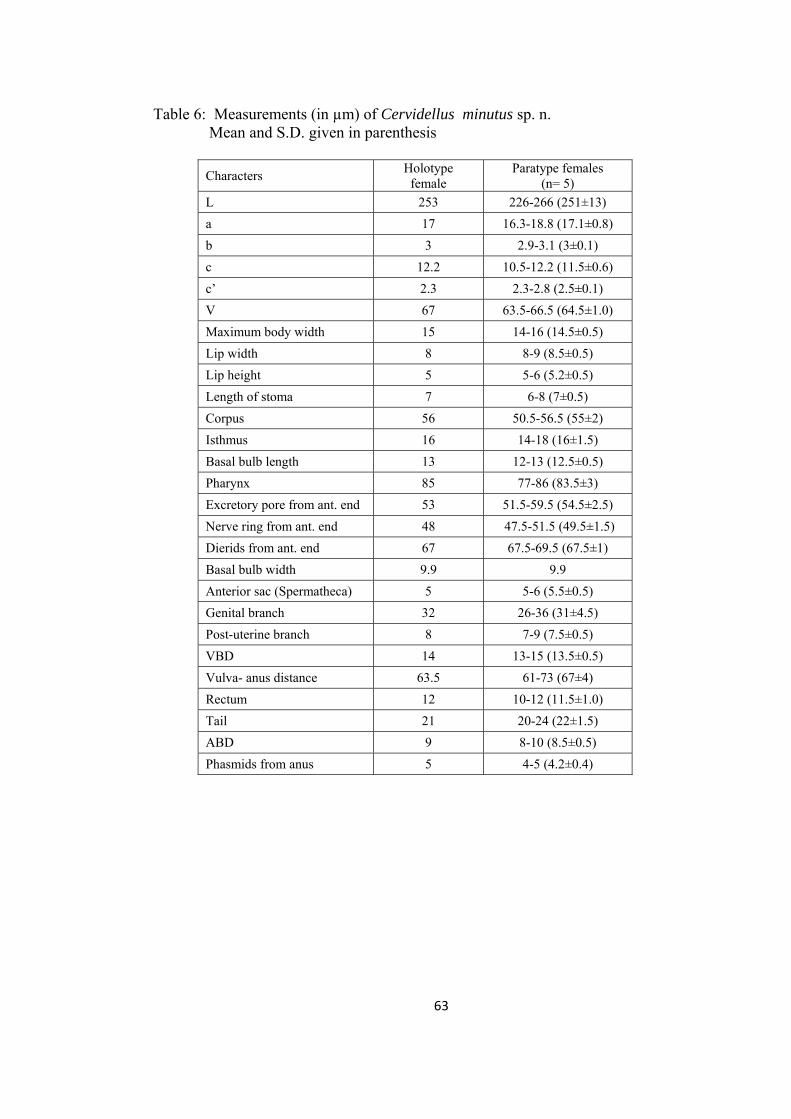
Table 6: Measurements (in µm) of Cervidellus minutus sp. n. Mean and S.D. given in parenthesis
Characters Holotype
female Paratype females
(n= 5)
L 253 226-266 (251±13)
a 17 16.3-18.8 (17.1±0.8)
b 3 2.9-3.1 (3±0.1)
c 12.2 10.5-12.2 (11.5±0.6)
c’ 2.3 2.3-2.8 (2.5±0.1)
V 67 63.5-66.5 (64.5±1.0)
Maximum body width 15 14-16 (14.5±0.5)
Lip width 8 8-9 (8.5±0.5)
Lip height 5 5-6 (5.2±0.5)
Length of stoma 7 6-8 (7±0.5)
Corpus 56 50.5-56.5 (55±2)
Isthmus 16 14-18 (16±1.5)
Basal bulb length 13 12-13 (12.5±0.5)
Pharynx 85 77-86 (83.5±3)
Excretory pore from ant. end 53 51.5-59.5 (54.5±2.5)
Nerve ring from ant. end 48 47.5-51.5 (49.5±1.5)
Dierids from ant. end 67 67.5-69.5 (67.5±1)
Basal bulb width 9.9 9.9
Anterior sac (Spermatheca) 5 5-6 (5.5±0.5)
Genital branch 32 26-36 (31±4.5)
Post-uterine branch 8 7-9 (7.5±0.5)
VBD 14 13-15 (13.5±0.5)
Vulva- anus distance 63.5 61-73 (67±4)
Rectum 12 10-12 (11.5±1.0)
Tail 21 20-24 (22±1.5)
ABD 9 8-10 (8.5±0.5)
Phasmids from anus 5 4-5 (4.2±0.4)
63
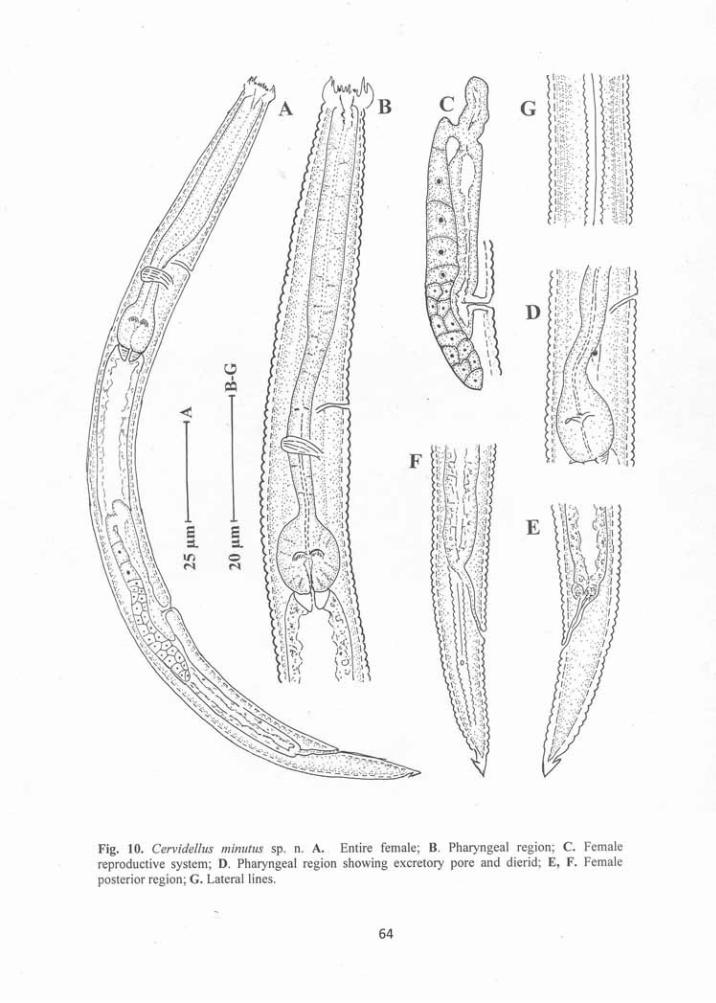

A B C
ED
F
GFig. 11. Cervidellus minutus sp. n. A. Pharyngeal region, B Anterior region showing cephalic
probolae with refractive margins, C Anterior region showing stoma, D. Lateral lines, E Female reproductive system, F. Posterior region, G. Tail tip showing dorsal projection (Scale bars = 20µm).
65

Genus Chiloplacus Thorne, 1937
Diagnosis: Body 0.3-1.0 mm long. Cuticle annulated, lateral fields with 3-6 lines.
Lips three. Labial probolae biacute or bifurcate (apically incised), symmetrical or
assymetrical, with broad “shafts”. Cephalic probolae small, generally with two
incisures. Pharyngeal corpus cylindrical. Post-vulval uterine sac of variable
lengths. Female tail straight, short, cylindroid, terminus broadly rounded. Male
tail ventrally bent, phasmids posterior to lateral caudal papilla.
Chiloplacus aligarhensis sp. n.
(Fig. 12, 13)
Measurements: In Table 7.
Females: Body slender, straight or slightly ventrally curved after fixation. Cuticle
annulated, annules 2.3-3.3 µm wide at mid-body, with irregular punctations
throughout the body length. Lateral fields about 1/4th body diam. at midbody,
with five incisures. Lateral lines arise at about two stoma lengths from anterior
end and from middle of procorpus differentiate into five lines that extends upto
the tail terminus except the central incisure that ends at phasmids. Lip region with
six lips. Cephalic probolae small, with deep primary axils and shallow secondary
axils. Labial probolae high (6-7µm), with shallow bifurcation forming finely
developed prongs. Stoma cephaloboid, rhabdions clearly differentiated;
cheilostom wide with minute oval or triangular rhabdia, gymnostom slightly
narrower than cheilostom and as wide as stegostom, metastegostom with small
tooth like projection. Pharyngeal corpus cylindrical, 8-11 times isthmus length.
Corpus-isthmus junction demarcated by faint transverse markings. Isthmus short,
66

17-22 µm long, basal bulb ovoid, with well developed grinder. Nerve ring
surrounding corpus in its posterior half, at 60-66% of pharynx length. Excretory
pore at 60-72% of pharynx length. Dierids 5-8 annules posterior to excretory
pore. Cardia short, conoid, surrounded by intestinal tissue. Intestine with wide
lumen and thin walls in its anterior region, cell differentiation not visible in
anterior and posterior regions of intestine. Cells in the mid-intestine region well
differentiated, rectangular in shape, granular in appearance with distinct nuclei.
Reproductive system mono-prodelphic, ovary reversed, without flexure
posterior to vulva, sometimes with swollen germinal portion with oocytes
arranged in one or two rows. Oocytes in a single row in the proliferative zone.
Oviduct short. Spermatheca well developed, longer than the corresponding body
diam.. Uterus tubular, more than two corresponding body diam. long,
differentiated into a proximal glandular part and a distal swollen muscular part
with thin walls. Post-uterine sac well developed, 2.5-4.0 vulval body diam. long.
Vagina muscular, anteriorly inclined, about half of vulval body diam. long. Vulva
transverse, slit-like, vulval lips may be slightly protruded. Rectum 1.0-1.3 anal
body diam. long. Tail subcylindrical with rounded terminus. Phasmids distinct,
1.0-1.5 anal body diam. posterior to anus.
Males: General morphology similar to that of females. Body J-shaped after
fixation. Reproductive system monorchic. Testis reflexed ventrally anteriorly,
with flexure on right side of intestine. Apical germinative zone with three to four
rows of spermatogonia, maturation zone with spermatocytes and differentiated
sperms at its distal end. Vas deferens highly granular in appearance without any
valve or sphincter at the junction of maturation zone. Ejaculatory duct with
67

similar appearance as of vas deferens but with distinct lumen containing sperms.
Tail conoid, ventrally curved. Genital papillae eight pairs; three pairs pre-cloacal,
ventro-lateral in position and five pairs post-cloacal. Of the post-cloacal pairs,
one ventro-lateral and one lateral pair anterior to phasmids, one pair dorso-lateral
near tail terminus and two terminal ventro-lateral pairs. A single median pre-
cloacal papilla present on the anterior cloacal lip. Spicules slightly ventrally
arcuate, manubrium round and bent ventrally, calamus with thin walls, lamina
thick, ventrally curved, bearing a longitudinal incisure from the calamus,
Gubernaculum well developed, trough-shaped, about half of spicule length, with
serrated margins at its proximal end.
Type habitat and locality: Soil around the rhizosphere of grasses from a barren
field near village Lodha on Aligarh-Khair road.
Type specimens
Holotype female on slide Chiloplacus aligarhensis sp. n./1, ten females
and eleven males (paratypes) on slides Chiloplacus aligarhensis sp. n./2-11
deposited in the nematode collection of Department of Zoology, Aligarh Muslim
University, Aligarh.
Diagnosis and relationship
Chiloplacus aligarhensis sp. n. is characterized by the long body (0.75-
0.83 mm in females and 0.72-0.82 mm in males), lateral fields with five
incisures, labial probolae with well developed prongs, pharyngeal corpus 7-10
times isthmus length, ovary without flexures posterior to vulva, well developed,
offset spermatheca, long post-uterine sac (85-109 µm), female tail subcylindrical,
68

2-3 anal body diam. long, male tail conoid 45-52 µm long, spicules 30-35µm
long and gubernaculum about half of spicules length.
The new species resembles C. tenuis Rashid & Heyns, 1990, C. subtenuis
Rashid & Heyns, 1990 and C. magnus Rashid & Heyns, 1990 in general
morphometrics, shape and body size. However, from C. tenuis it can be
differentiated in the shape of labial probolae (prongs small and straight vs prongs
long and curved towards each other), longer pharyngeal corpus in females (155-
177µm vs 105-150µm), longer female tail (42-49µm vs 25-40µm. From C.
subtenuis it differs by slightly smaller pharynx in females (197-216µm vs 219-
240µm), males of C. subtenuis differs in having longer tail (45-52µm vs 38-
43µm), wider body diam. at cloaca (22-27µm vs 18-20µm), slightly larger c′
value (1.9-2.3 vs 1.6-1.8) and in the arrangement of genital papillae (three pre-
cloacal & five post-cloacal pairs vs four pre-cloacal & four post-cloacal pairs).
From C. magnus the new species differs by its smaller body size (753-829µm vs
883-1522µm), slightly posterior position of nerve ring and excretory pore (60-
66% & 60-72% vs 35-59% & 38-56% of the pharyngeal length respectively),
smaller pharynx (197-216 µm vs 222-289 µm in females and 191-209 µm vs 211-
245 µm in males), relatively smaller spermatheca (35-47 µm vs 48-99 µm),
smaller genital branch (73-113µm vs 143-218µm), smaller vulva-anus/tail ratio
(4.2-5.0 vs 5.2-8.0) and in having smaller testis (294-412µm vs 421-741µm).
69
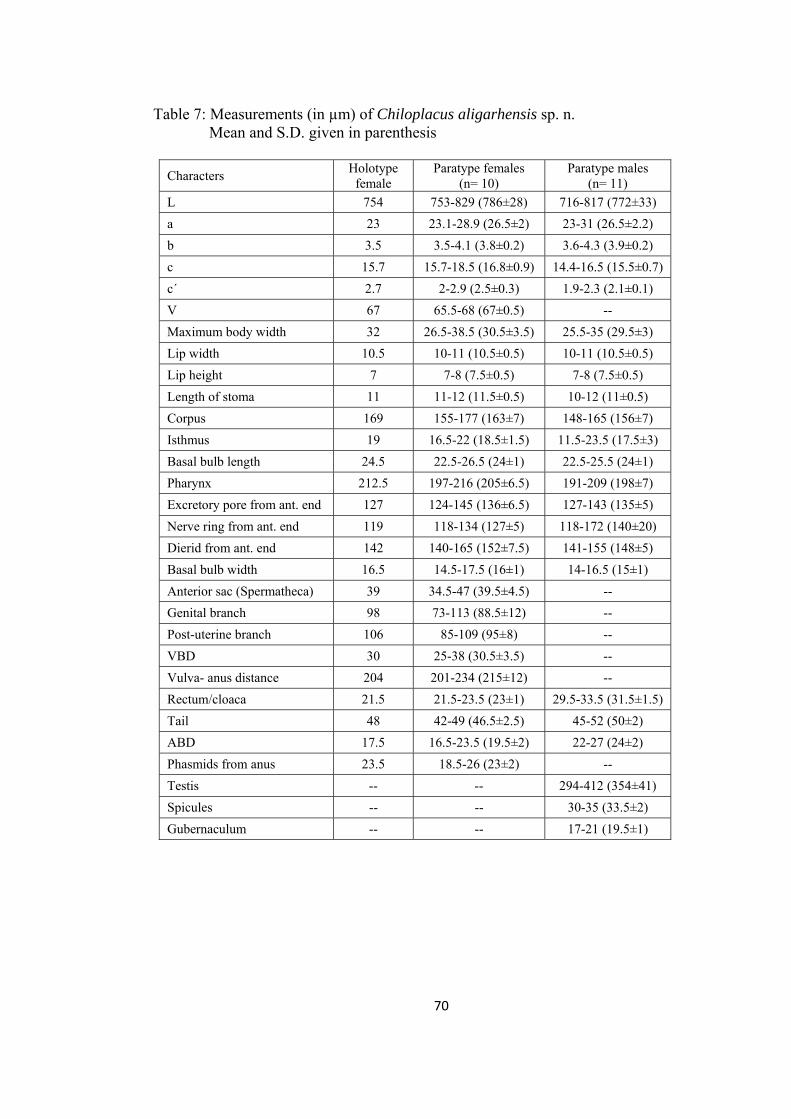
Table 7: Measurements (in µm) of Chiloplacus aligarhensis sp. n. Mean and S.D. given in parenthesis
Characters Holotype
female Paratype females
(n= 10) Paratype males
(n= 11)
L 754 753-829 (786±28) 716-817 (772±33)
a 23 23.1-28.9 (26.5±2) 23-31 (26.5±2.2)
b 3.5 3.5-4.1 (3.8±0.2) 3.6-4.3 (3.9±0.2)
c 15.7 15.7-18.5 (16.8±0.9) 14.4-16.5 (15.5±0.7)
c´ 2.7 2-2.9 (2.5±0.3) 1.9-2.3 (2.1±0.1)
V 67 65.5-68 (67±0.5) --
Maximum body width 32 26.5-38.5 (30.5±3.5) 25.5-35 (29.5±3)
Lip width 10.5 10-11 (10.5±0.5) 10-11 (10.5±0.5)
Lip height 7 7-8 (7.5±0.5) 7-8 (7.5±0.5)
Length of stoma 11 11-12 (11.5±0.5) 10-12 (11±0.5)
Corpus 169 155-177 (163±7) 148-165 (156±7)
Isthmus 19 16.5-22 (18.5±1.5) 11.5-23.5 (17.5±3)
Basal bulb length 24.5 22.5-26.5 (24±1) 22.5-25.5 (24±1)
Pharynx 212.5 197-216 (205±6.5) 191-209 (198±7)
Excretory pore from ant. end 127 124-145 (136±6.5) 127-143 (135±5)
Nerve ring from ant. end 119 118-134 (127±5) 118-172 (140±20)
Dierid from ant. end 142 140-165 (152±7.5) 141-155 (148±5)
Basal bulb width 16.5 14.5-17.5 (16±1) 14-16.5 (15±1)
Anterior sac (Spermatheca) 39 34.5-47 (39.5±4.5) --
Genital branch 98 73-113 (88.5±12) --
Post-uterine branch 106 85-109 (95±8) --
VBD 30 25-38 (30.5±3.5) --
Vulva- anus distance 204 201-234 (215±12) --
Rectum/cloaca 21.5 21.5-23.5 (23±1) 29.5-33.5 (31.5±1.5)
Tail 48 42-49 (46.5±2.5) 45-52 (50±2)
ABD 17.5 16.5-23.5 (19.5±2) 22-27 (24±2)
Phasmids from anus 23.5 18.5-26 (23±2) --
Testis -- -- 294-412 (354±41)
Spicules -- -- 30-35 (33.5±2)
Gubernaculum -- -- 17-21 (19.5±1)
70
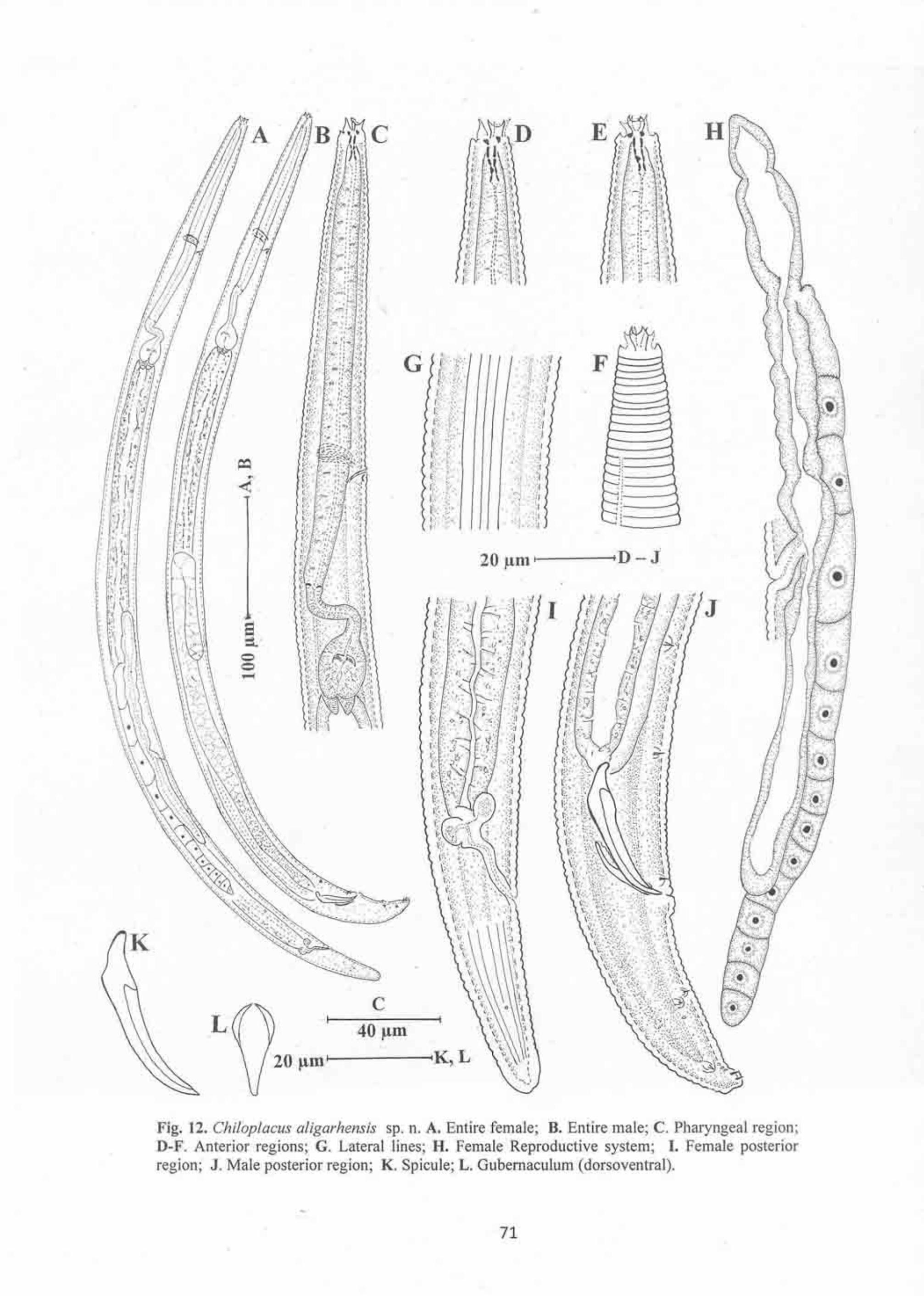
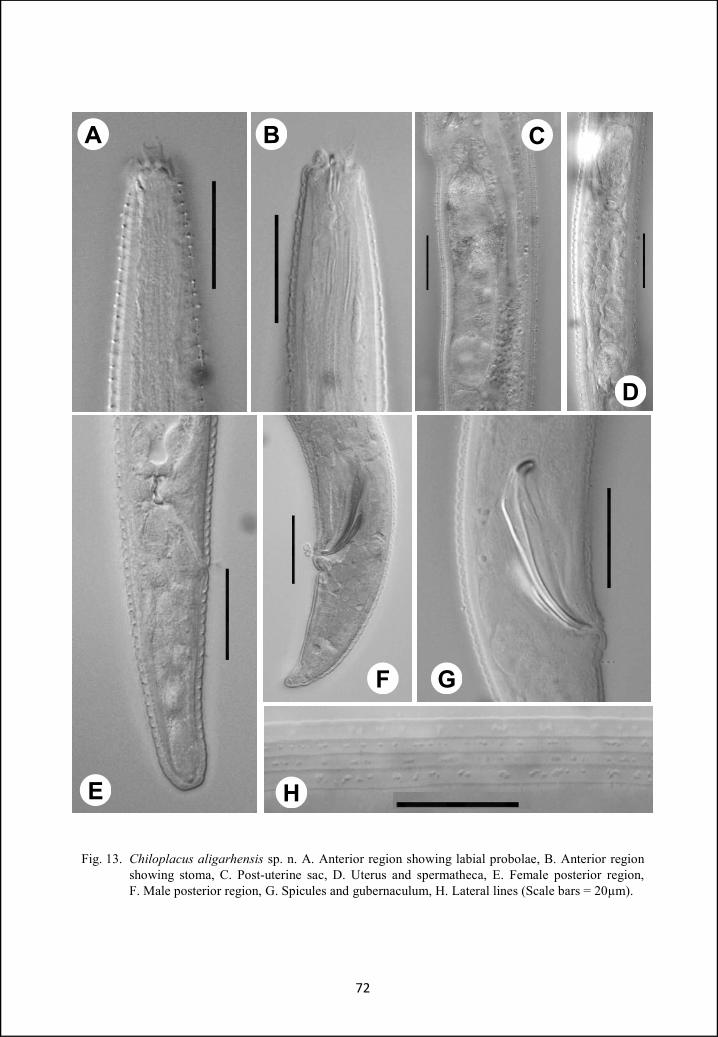
Fig. 13. Chiloplacus aligarhensis sp. n. A. Anterior region showing labial probolae, B. Anterior region showing stoma, C. Post-uterine sac, D. Uterus and spermatheca, E. Female posterior region, F. Male posterior region, G. Spicules and gubernaculum, H. Lateral lines (Scale bars = 20µm).
72
A B C
D
GF
E H

Genus Nothacrobeles Allen & Noffsinger, 1971
Diagnosis: Body length varying from 0.4 to 0.9 mm. Cuticle with broad annules,
with or without longitudinal striae or punctations. Lateral fields with two to four
incisures. Cephalic probolae in pairs, low, with or without serrate sculpture.
Labial probolae short to moderately long, slightly bifurcate and bordered with
small tines; outer rim or shaft with basal ridge. Amphids minute. Pharyngeal
corpus cylindrical. Female gonad cephaloboid. Tails in both sexes conical with
acute tip. Phasmids anterior to the lateral caudal papilla in males.
Nothacrobeles punctatus sp. n.
(Fig. 14, 15)
Measurements: In Table 8.
Females: Body robust, slightly ventrally curved after fixation. Cuticle simple,
transversely annulated, annuli 3-4 µm wide at midbody, with two rows of
punctuations. Lateral fields about 1/6th of the body diam. at midbody. Incisures
start about two stoma lengths from anterior end as two crenate lines and
differentiate into four lines at the level of isthmus. Outer incisures crenate, inner
ones smooth, areolation absent. Lateral fields extend up to the tail terminus.
Labial probolae 8-10 µm long, bifurcate for half their length, with a prominent
dentate basal ridge protruding outwards towards the cephalic probolae. Prongs
divergent and bifurcate, outer longer than the inner. Each prong with six
triangular tines on inner margin and eight on outer margin. Primary and
secondary axils deep with ‘U’ and ‘V’ shape respectively. Amphidial openings
minute, oval or round in shape. Stoma cephaloboid. Cheilostom wide, with
73

almost triangular rhabdia. Gymnostom narrower than cheilostom and as wide as
stegostom. Dorsal metarhabdion with a minute tooth. Pharyngeal corpus slightly
fusiform 3.5-4.0 times isthmus length. Corpus-isthmus junction with transverse
markings. Basal bulb spheroid, with well developed grinder. Nerve ring at 66-
78% of neck length, surrounding isthmus in anterior half. Excretory pore at the
level or just posterior to nerve ring. Hemizonid about two annules posterior to
excretory pore. Dierids in the isthmus region or at the level of basal bulb. Cardia
conoid, 5-7µm long, surrounded by intestinal tissue. Intestine with distinct wide
lumen throughout its length, intestinal cells well differentiated, rectangular in
shape.
Reproductive system mono-prodelphic, ovary reversed, without any
flexure. Oocytes arranged in a single row throughout. Oviduct short, tubular,
made of eight differentiated cells. Spermatheca well developed, usually longer
than the corresponding body diam., with few sperms. Uterus well developed,
about two to three times body diam. long. Uterus differentiated into a proximal
glandular part with narrow lumen and a swollen distal muscular part with wide
lumen. Distal and proximal parts of uterus separated by slight constriction. Post-
uterine sac less than one vulval body diam. in length. Vagina thick walled, about
1/3rd of vulval body diam. Vulva transverse, slit-like. Rectum slightly less than
one anal body diam. long, with three rectal glands. Tail conical, 2-3 anal-body
diam. long, with acute terminus. Phasmids located at less than one anal body
diam. posterior to anus.
Males: General morphology similar to females. Posture slightly straighter than
females but more ventrally curved posteriorly. Reproductive system monorchic,
74

testis reflexed ventrally, flexure on right side of intestine. Germinal zone with
two to three rows of spermatogonia, leading to the maturation zone with
spermatocytes and differentiated sperms at its distal end. Vas deferens with
numerous small round sperms. Ejaculatory duct wider than vas deferens with
narrow lumen. Tail conical, ventrally curved, terminating in an acute mucro.
Phasmids 1.3-1.5 anal body diam. posterior to anus. Genital papillae eight pairs;
two pairs subventral precloacal, one pair subventral adcloacal and five pairs
postcloacal. Of five postcloacal pairs, two pairs (one lateral and one subventral)
are located just posterior to phasmid and three pairs (one subdorsal, one
subventral and one lateral) near the tail tip. Spicules ventrally arcuate;
manubrium rounded; calamus cylindrical; lamina swollen near calamus.
Gubernaculum well developed, slightly bent anteriorly.
Type habitat and locality: Soil sample collected from a potato field, Shahrekord,
Iran
Type specimens
Holotype female on slide Nothacrobeles punctatus sp. n./1, three females
and six males (paratypes) on slides Nothacrobeles punctatus sp. n./1-4 deposited
in the nematode collection of Department of Zoology, Aligarh Muslim
University, Aligarh.
Diagnosis and relationships
Nothacrobeles punctatus sp. n. is characterized by having two rows of
punctations in each annule; bifurcate labial probolae, with divergent prongs;
spermatheca 28-50 µm long; post-uterine sac less than one vulval body diam.
75

long; female tail conical with acute terminus; phasmids at 21-32% of tail length;
spicules 45-49 µm long and gubernaculum 30-34 µm long.
The new species resembles Nothacrobeles subtilis Allen and Noffsinger,
1971 and N. maximus Allen and Noffsinger, 1971 in general morphometrics.
However, the new species differs from N. subtilis in cuticular punctations (with
punctations vs without punctations), more anterior position of phasmid (21-32%
vs near middle of tail) and in the presence of males. From N. maximus it can
easily be differentiated by the structure of cuticle (without longitudinal striations
vs with longitudinal striations), cuticular punctations (with punctations vs without
punctations), lateral fields (incisures crenate anterior to dierids then smooth
throughout the body length vs incisures crenate posterior to dierids) and in the
presence of males (males absent in N. maximus).
76
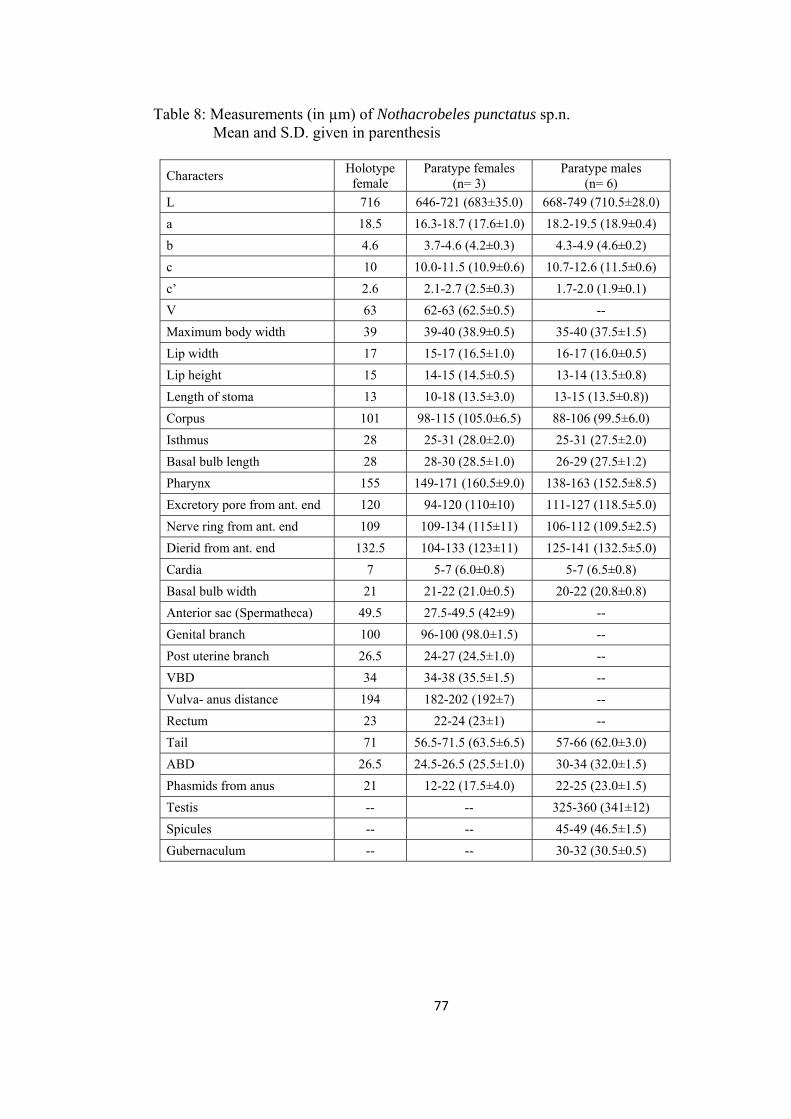
Table 8: Measurements (in µm) of Nothacrobeles punctatus sp.n. Mean and S.D. given in parenthesis
Characters Holotype
female Paratype females
(n= 3) Paratype males
(n= 6)
L 716 646-721 (683±35.0) 668-749 (710.5±28.0)
a 18.5 16.3-18.7 (17.6±1.0) 18.2-19.5 (18.9±0.4)
b 4.6 3.7-4.6 (4.2±0.3) 4.3-4.9 (4.6±0.2)
c 10 10.0-11.5 (10.9±0.6) 10.7-12.6 (11.5±0.6)
c’ 2.6 2.1-2.7 (2.5±0.3) 1.7-2.0 (1.9±0.1)
V 63 62-63 (62.5±0.5) --
Maximum body width 39 39-40 (38.9±0.5) 35-40 (37.5±1.5)
Lip width 17 15-17 (16.5±1.0) 16-17 (16.0±0.5)
Lip height 15 14-15 (14.5±0.5) 13-14 (13.5±0.8)
Length of stoma 13 10-18 (13.5±3.0) 13-15 (13.5±0.8))
Corpus 101 98-115 (105.0±6.5) 88-106 (99.5±6.0)
Isthmus 28 25-31 (28.0±2.0) 25-31 (27.5±2.0)
Basal bulb length 28 28-30 (28.5±1.0) 26-29 (27.5±1.2)
Pharynx 155 149-171 (160.5±9.0) 138-163 (152.5±8.5)
Excretory pore from ant. end 120 94-120 (110±10) 111-127 (118.5±5.0)
Nerve ring from ant. end 109 109-134 (115±11) 106-112 (109.5±2.5)
Dierid from ant. end 132.5 104-133 (123±11) 125-141 (132.5±5.0)
Cardia 7 5-7 (6.0±0.8) 5-7 (6.5±0.8)
Basal bulb width 21 21-22 (21.0±0.5) 20-22 (20.8±0.8)
Anterior sac (Spermatheca) 49.5 27.5-49.5 (42±9) --
Genital branch 100 96-100 (98.0±1.5) --
Post uterine branch 26.5 24-27 (24.5±1.0) --
VBD 34 34-38 (35.5±1.5) --
Vulva- anus distance 194 182-202 (192±7) --
Rectum 23 22-24 (23±1) --
Tail 71 56.5-71.5 (63.5±6.5) 57-66 (62.0±3.0)
ABD 26.5 24.5-26.5 (25.5±1.0) 30-34 (32.0±1.5)
Phasmids from anus 21 12-22 (17.5±4.0) 22-25 (23.0±1.5)
Testis -- -- 325-360 (341±12)
Spicules -- -- 45-49 (46.5±1.5)
Gubernaculum -- -- 30-32 (30.5±0.5)
77
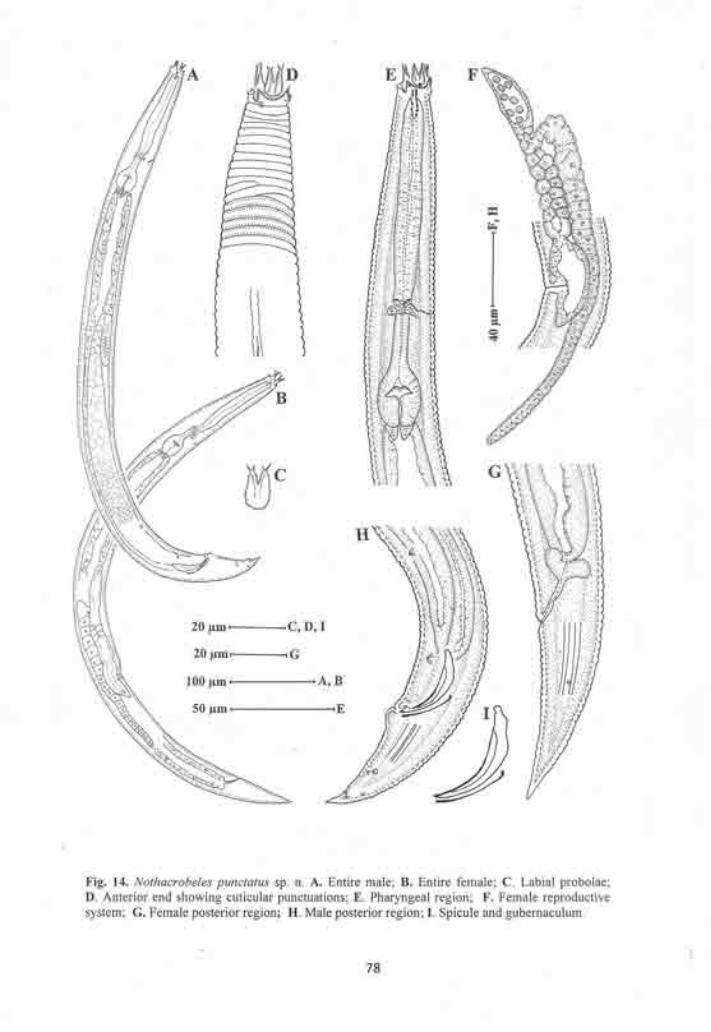

H I
E
F
G
C
B DA
Fig. 15. Nothacrobeles punctatus sp. n. A. Anterior region showing stoma, B&C. Labial probolae, D. Vulval region showing post-uterine sac, E. Glandular part of uterus and spermatheca, F. Cuticular punctations, G. Female posterior region, H. Spicules and gubernaculum, I. Lateral lines (Scale bars = 20µm).
79

Genus Stegellata Thorne, 1938
Diagnosis: Body 0.3-0.7 mm long. Cuticle with transverse as well as longitudinal
striations, dividing each other to form small quadrate blocks. Lateral fields with
three to five incisures. Cephalic probolae simple, low. Labial probolae tuning
fork-shaped with thin shaft and U-like prongs. Pharyngeal corpus cylindrical.
Female genital organs cephaloboid. Female tail broadly, male tail narrowly
rounded.
Stegellata ophioglossa Andrassy, 1967
(Fig. 16)
Measurements: In Table 9.
Females: Body small, straight to slightly ventrally curved after fixation. Cuticle
with transverse and longitudinal striations, giving it a tessellated appearance.
Lateral fields distinct, occupying about 1/5th of the midbody diam.. Incisures
three, outer ones crenate. Lip region with six separate lips having rounded
margins. Labial probolae with wide base, 4-6µm long, bifurcate to about 1/3rd of
their length. Prongs divergent and equal in size, forming a semicircular arc.
Amphidial apertures round. Stoma cephaloboid, cheilostom wide, with small
ovoid rhabdia. Gymnostom narrower than cheilostom, and as wide as stegostom.
Metastegostom with a minute tooth like projection on dorsal rhabdia. Pharyngeal
corpus cylindrical, with distinct lumen, 3.3-4.5 times isthmus length. Corpus-
isthmus junction distinctly demarcated. Basal bulb ovoid or pear-shaped with
well a developed grinder. Nerve ring surrounding the base of corpus at 60-67% of
neck length. Excretory pore opposite the trailing end of nerve ring or at 62-71%
80

of neck length. Hemizonid just posterior to the excretory pore. Dierids at about
65-71% of neck length, lying in the anterior half of isthmus. Cardia short, conoid,
surrounded by intestinal tissue. Intestine with wide lumen at its anterior region.
Reproductive system mono-prodelphic. Ovary posteriorly directed
without any flexure. Oocytes arranged in two rows in the germinal zone and in a
single row in the proliferative zone. Oviduct short, tubular. Spermatheca less than
the corresponding body diam long, without sperms. Uterus simple, tube like,
undifferentiated. Post-uterine sac less than one vulval body diam. long. Vagina,
thick walled, with sclerotization. Vulva transverse, slit like. Rectum 1.2-1.6 anal
body diam. long. Tail cylindrical, 2.6-3.5 anal body diam. long, with
approximately 20 ventral annuli. Tail terminus flat. Phasmids one anal body
diam. posterior to anus.
Males: Not found.
Habitat and locality: Soil sample collected from the root zone of Sorghum sp.
from village Nohati, Madrak and from the root zone of Trifolium alexandrinum
from village Pisava, Chandaus, Aligarh.
Voucher specimens
17 females on slides Stegellata ophioglossa (N)/1-9 and 12 females on
slides Stegellata ophioglossa (P)/1-4 deposited in the nematode collection of
Department of Zoology, Aligarh Muslim University, Aligarh.
Remarks
S. ophioglossa is a terrestrial nematode, this species have previously been
recovered from sandy soils and dune sands. It is widely distributed and has been
described from Europe (Hungary, Italy), Asia (Uzbekistan, Mongolia), Africa
81

(Senegal) and South America (Venezuela). This is the first report of S.
ophioglossa from India, where, it was recovered from sandy as well as from
loamy soil. Despite the change in type of habitat the measurements and
descriptions of our specimens agree well with that of Stegellata ophioglossa
Andrassy, 1967. However slight variations from the original description were
observed in the height of labial probolae (4-6 µm vs 8-11 µm), relatively smaller
pharynx (96-112 µm vs 110-130 µm), smaller post-vulval uterine sac (less than
corresponding body diam. vs as long as or longer than corresponding body diam.)
82
Table 9: Measurements (in µm) of Stegellata ophioglossa Andrassy, 1967 Mean and S.D. given in parenthesis
Characters Females (n= 10)
Nohati population Females (n=12)
Pisava population
L 327 – 386 (347 ± 16) 341 – 409 (347 ± 16)
a 18.3 – 23 (21 ± 1.5) 19.2 – 22 (20.2 ± 0.8)
b 3.1 – 3.7 (3.4 ± 0.2) 3.2 – 3.7 (3.4 ± 0.2)
c 10.3 – 11.9 (11.1 ± 0.5) 10 – 11.4 (10.6 ± 0.5)
c’ 2.6 – 3.4 (3 ± 0.2) 2.9 – 3.7 (3.1 ± 0.2)
V 62 – 64.5 (63 ± 0.5) 61 – 63.5 (62 ± 0.5)
Maximum body width 15 – 18 (16.5 ± 1) 15.5 – 20 (18 ± 1)
Lip width 6 – 7 (6 ± 0.3) 6 – 7 (6.9 ± 0.3)
Length of stoma 9.0 9.0 – 10 (9.5 ± 0.5)
Corpus 66.5 – 72.5 (70 ± 2) 69.5 – 77 (72 ± 2)
Isthmus 14.5 – 21.5 (18 ± 1.5) 18.5 – 24.5 (22 ± 1.5)
Basal bulb length 14 – 16 (14.5± 0.5) 13 – 17 (14.5± 1)
Pharynx 96 – 112 (102.5 ± 4.5) 104 – 114 (108 ± 3)
Excretory pore from ant. end 67.5 – 73.5 (69 ± 1.5) 69.5 – 75 (73 ± 1.5)
Nerve ring from ant. end 61.5 – 68.5 (65.5 ± 2.5) 68 – 73 (70.5 ± 1.5)
Dierid from ant. end 72.5 – 79 (74.5 ± 2.5) 75 – 82 (78.5 ± 2)
Basal bulb width 9 – 12 (10 ± 0.5) 10 – 13 (11 ± 0.5)
Anterior sac (Spermatheca) 10 – 14 (12 ± 1.5) 7 – 10 (9 ± 1)
Genital branch 36.5 – 65.5 (45.5 ± 7.5) 32.5 – 48.5 (40 ± 4)
Post uterine branch 8 – 12 (10 ± 1.2) 8 – 11 (10 ± 1)
VBD 14 – 17 (15.5 ± 1) 15 – 19 (17 ± 1)
Vulva- anus distance 89 – 113 (98 ± 6.5) 92 – 119 (105 ± 7)
Rectum 13 – 15 (14 ± 0.5) 14 – 16 (15 ± 0.5)
Tail 28.5 –34.5 (31 ± 1.5) 30.5 –37.5 (35 ± 2)
ABD 9 – 12 (10.5 ± 1) 10 – 12 (11 ± 0.5)
Phasmids from anus 9 – 10 (9.5 ± 0.5) 7 – 15 (10.5 ± 2)
83
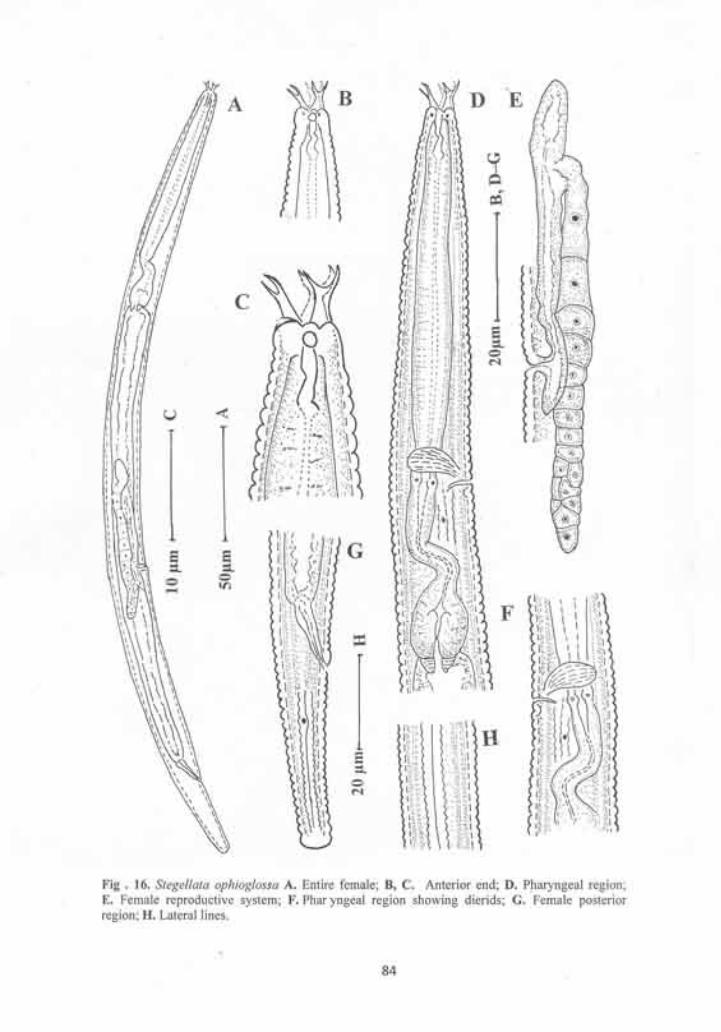

Genus Zeldia Thorne, 1937
Diagnosis: Body length between 0.6-1.0 mm. Cuticle annulated, annuli with or
without punctations or tessellation. Lateral fields with three to five incisures,
outer lines sometimes crenate. Lateral fields with or without areolations. Cephalic
probolae triangular, flap-like, low with setose projections. Labial probolae low
and rounded to elongate and shallowly to deeply bifurcate at one level; without
tines. Amphidial apertures elongate-oval. Lining of cheilostom without or with 1-
3 tooth-like processes. Pharyngeal corpus long and cylindrical. Intestine often
with prerectum. Postvulval uterine branch short or absent. Tails in both sexes
conoid, with pointed or finely rounded tip. Males rare or unknown in most
species.
Zeldia tridentata Allen and Noffsinger, 1972
(Fig.19, 20)
Measurements: In Table 10.
Females: Body robust, gradually tapering toward both the ends, straight or
slightly curved ventrally. Cuticle with transverse annules, 1.6 – 2 µm wide at the
pharynx base and midbody and 1.6 - 1.8 µm wide at tail. Each annule with two
rows of punctations. Lateral fields with three lines, outer ones crenate, areolated
in the pharyngeal region. Lip region 10-11 µm wide, 4-5µm high. Labial
probolae low, with round margins and shallow grooves. Primary axils deep, with
dentate guard processes. Amphidial apertures oval. Cheilostom with prominent
cylindrical walls, each cheilorhabdion associated with a structure having three
teeth. Gymnostom longer than cheilostom, metastegostom with a small tooth like
85

process on dorsal wall. Pharyngeal corpus cylindrical, 7.9-9.2 times isthmus
length. Isthmus shorter than basal bulb. Basal bulb ovoid, 21-26 x 18-21 µm,
with a grinder at its middle or slightly anterior. Cardia conoid, surrounded by
intestinal tissue. Intestine with wide lumen. Nerve ring at 59-66% of neck length,
surrounding the distal part of the corpus. Excretory pore just posterior to nerve
ring or one to two annules anterior to hemizonid. Deirids in the posterior region
of corpus, or at 67-78 % of neck length.
Reproductive system mono-prodelphic, ovary reversed, on right side of
intestine, with or without additional flexures posterior to vulva. Oocytes usually
arranged in a single row throughout the ovary length. Spermatheca scarcely
developed. Oviduct short. Uterus tubular, without any differentiation. Post-
uterine sac short, 0.3-0.5 times vulval body diam. long. Vagina with thick walls,
slightly anteriorly directed. Rectum 21-28 µm long. Tail elongate conoid, 4-5
anal body diam. long, with acute terminus. Phasmids 1-3 µm posterior to anus.
Male: Not found
Habitat and locality
Soil sample collected from an orchard of guava and cashew-nut near
Chilika lake, Puri, Orissa.
Voucher specimens
9 females on slides Zeldia tridentata/1-2 deposited in the nematode
collection of Department of Zoology, Aligarh Muslim University, Aligarh.
Remarks
Z. tridentata is distinguished from other species of the genus by the
presence of three teeth associated with each cheilorhabdion and the longer tail.
86

This species is widely distributed throughout the world and has been collected
from India, Jamaica, Philippine Islands, Srilanka, Taiwan, Thailand, and
Venezuela. Most of the species of the genus were collected from the soil or sand
around the root zone of different plants. The present population was also
collected from the sandy soil, around the root zone of Guava and Cashew Nut
plantations.
The measurements and descriptions of our specimens agree well with that
of Zeldia tridentata Allen & Noffsinger, 1972. However differences from
original population were found in the lateral fields (outer incisures crenated vs
smooth). Rashid et al., 1984, collected two females of this species from Itapebi,
Lombardia, Brazil, (Host: Theobroma cacao). They Illustrate and redescribed the
species and added more details to the original description. In Brazilian
population, cuticular punctations were not observed, however the punctations
have been reported in the original descriptions and are distinctly visible in our
population also. They also mentioned the ovary with a double flexure posterior to
vulva but in our population the specimens without any flexure were also found
along with the specimens having double flexure. In Brazilian population the
phasmid lies at about 5-6 annules posterior to anus however, in our population
phasmid lies just posterior to anus.
87
Table 10: Measurements (in µm) of Zeldia tridentata Allen and Noffsinger, 1972 Mean and S.D. given in parenthesis
Characters Females (n= 9)
L 635-779 (724±46)
a 19.0-23.5 (20.8±1.3)
b 3.7-4.0 (3.9±0.1)
c 8.5-9.0 (8.8±0.2)
c’ 4.1-4.9 (4.5±0.3)
V 61-62 (61.5±0.3)
Maximum body width 30.5-40.5 (35±3.0)
Lip width 10-11 (11±0.5)
Lip height 4-5 (4.5±0.5)
Length of stoma 15-17 (15.5±0.5)
Corpus 135.5-155.5 (147±6.5)
Isthmus 14-19 (16.5±1.5)
Basal bulb length 20-25.5 (23.5±1.5)
Pharynx 170-200 (187±9.0)
Excretory pore from ant. end 109-126.5 (121±5.5)
Nerve ring from ant. end 105-121.5 (116.5±5.5)
Dierid from ant. end 124-149 (137.5±9.0)
Cardia 5-6 (5±0.5)
Basal bulb width 18-21 (19±1)
Anterior sac (Spermatheca) 7-14 (10.5±2.5)
Genital branch 70-135 (9±21)
Post uterine branch 12-18 (13.5±2.0)
VBD 31.5-40.5 (35.5±3.5)
Vulva- anus distance 166-209 (194±14)
Rectum 21-28 (24.5±2.0)
Tail 73-87 (82.5±4.0)
ABD 16-21 (17.5±1.5)
Phasmids from anus 1-3 (2±0.7)
88
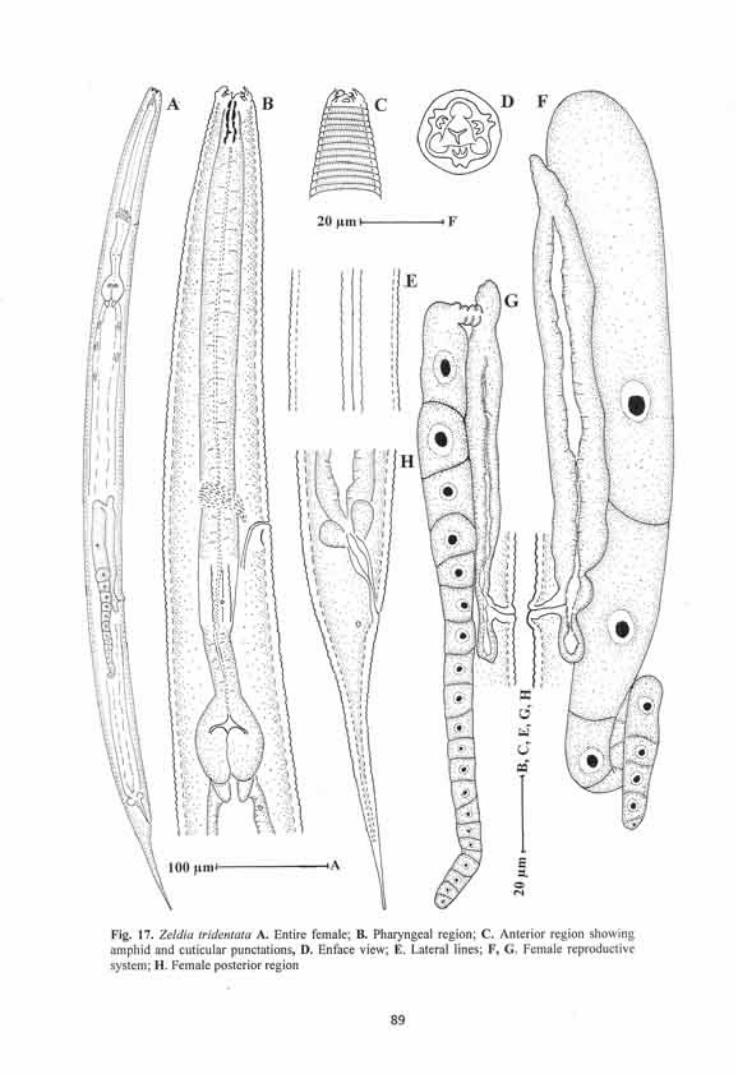

Superfamily Panagrolaimoidea Thorne, 1937
Diagnosis: Buccal cavity a single, fairly wide chamber, sometimes tending to
taper at its base. Metastom anisoglottoid and anisomorphic, narrower than
anterior part; telostom small and narrow. Pro-, meso- and telorhabdions usually
conspicuously thickened; cheilorhabdions sometimes thickened; metarhabdions
not thickened but metastom segments often bearing small teeth. Pharyngeal
corpus cylindrical or with swollen valveless median bulb; a narrow isthmus and a
basal bulb with grinders. Female genital organ prodelphic, ovary reflexed once,
usually well down into body; a short post-vulval sac sometimes present. Tip of
male gonad usually reflexed. Male supplements papilloid, arranged in pairs. No
bursa. Phasmids well discernible.
Type family: Panagrolaimidae Thorne, 1937
Other subfamilies: Alirhabditidae Suryawanshi, 1971
Brevibuccidae Paramonov, 1956
Family Panagrolaimidae Thorne, 1937
Diagnosis: Cuticle annulated, lateral fields distinct. Lip region practically without
probolae. Lips three or six, moderately developed.Amphids located on lateral
lips, small. Stoma consisting of the usual six elements, its anterior section
(cheilo- and gymnostom) spacious, stegostom tapering, metastom with a small
tooth-like projection. Pharynx consisting of corpus, isthmus and bulb. Female
genital organ prodelphic, reflexed part extending far behind vulva, rarely with a
simple flexure. Anterior separated portion of uterus serving as a spermatheca.
Postvulval uterine sac present. Males mostly abundant. Preanal genital papillae in
90

5-7 pairs. Tail conoid or elongate, in male generally shorter than in female.
Phasmids always distinct.
Type subfamily: Panagrolaiminae Thorne, 1937
Other subfamilies: Panagrellinae Andrassy, 1976
Tricephalobinae Andrassy, 1976
Turbatricinae Goodey, 1943
Baujardiinae Andrassy, 2005
Subfamily Tricephalobinae Andrassy, 1976
Diagnosis: Cuticle finely annulated, lateral fields distinct. Lip region without
probolae. Lips three or six, moderately developed. Cheliostom small and less
sclerotized, gymnostom well developed, stegostom tapering, Pharyngeal corpus
with bulb like median swelling. Terminal bulb very strong. Female genital organ
panagrolaimoid. Postvulval uterine sac present. Males mostly abundant. Genital
papillae 6-7 pairs. Tail uniformly conoid in female. Phasmids always distinct.
Type genus: Tricephalobus Steiner, 1936
Other genus: Halicephalobus timm, 1956
Genus Tricephalobus Steiner, 1936
Diagnosis: Body length between 0.5-1.0 mm. cuticle finely annulated, lateral
fields narrow. Head broad, continuous with body, lips three, separate. Cheilostom
short and not sclerotized, gymnostom well developed, stegostom funnel- shaped.
Pharyngeal corpus posteriorly swollen, bulb-like. Terminal bulb very strong.
Female genital organ panagrolaimoid, postvulval branch present. Both sexes
91

equally common. Spicules with narrowed proximal end, gubernaculum thin.
Genital papillae six or seven pairs. Tail conoid in female, and strongly narrowed
at posterior half in male, terminus sharp.
Tricephalobus quadripapilli sp.n.
(Fig.18, 19)
Measurements: In Table 11.
Females: Body slender, slightly ventrally curved after fixation. Cuticle simple,
with fine transverse annulations. Lateral fields narrow with two incisures,
occupying about 1/9th of the body diam. at midbody. Incisures arise at level of
median bulb as two smooth lines and extend up to the level of phasmids. Labial
probolae absent. Lips three, low and rounded, continuous with body contour.
Amphids indistinct. Stoma tubular, cheilostom with indistinct rhabdia.
Gymnostom slender, tube like, surrounded by a granular band like structure.
Stegostom as long as gymnostom, with slightly tapering dorsal wall. Pharyngeal
corpus posteriorly swollen into an oval median bulb, 2.7 – 3.7 times isthmus
length. Isthmus tubular. Basal bulb pyriform, with well developed grinder. Nerve
ring at 67-73% of neck length, surrounding isthmus at its middle. Excretory pore
at level of basal bulb. Dierids situated at the level of basal bulb or slightly
posterior to pharynx. Cardia short, conoid, surrounded by intestinal tissue.
Intestine with distinct wide lumen.
Reproductive system mono-prodelphic. Ovary reversed, without any
flexure, extending upto anus level. Oocytes arranged in multiple rows in germinal
zone and in a single row in maturation zone. Oviduct short, tubular, separated
92

from spermatheca by constriction. Spermatheca in continuation of uterus,
separated from it by slight constriction. Uterus well developed, differentiated into
a proximal glandular part and a distal muscular part with distinct lumen. Distal
part is twice as long as muscular part, both parts were separated by a constriction.
Post-uterine sac continuous with uterus, small, less than half vulval body diam. in
length. Vagina slightly anteriorly directed, about 1/3 - 1/2 of vulval body diam. in
depth. Vulva transverse, slit-like. Vulval lips slightly protruded. Rectum about
one anal body diam. long. Tail conical, 2.6-3.7 anal-body diam. long, with
narrowly rounded terminus. Phasmids 1.3-1.8 anal body diam. posterior to anus.
Males: Body slightly smaller than females. General morphology similar to that of
females. Reproductive system monarchic. Testis reflexed ventrally, flexure on
right side of intestine. Germinal zone with two to three rows of spermatogonia,
leading to the maturation zone with spermatocytes. Tail conical, ventrally curved,
strongly narrowed in its posterior half, terminating in a rounded tip. Phasmids
indistinct. Genital papillae four pairs; one subventral pair precloacal, situated at
level of spicule head, three pairs postcloacal. Of three postcloacal pairs, one
subventral pair is located at less than one anal body diam. posterior to cloacal
opening. Two pairs (one subventral and one subdorsal) located just anterior to the
beginning of narrower part of the tail. Spicules broad, with rounded manubrium,
broad calamus, ventrally arcuate lamina, with two longitudinal incisures from the
calamus. Spicule tip rounded. Gubernaculum small, less then half of spicules
length.
Type habitat and locality: Farm yard manure collected from a fallow field,
Poonch Jammu & Kashmir, India.
93

Type specimens
Holotype female on slide Tricephalobus quadripapilli sp. n./1; nine
females and ten males (paratypes) on slides Tricephalobus quadripapilli sp. n./2-
6, deposited in the nematode collection of Department of Zoology, Aligarh
Muslim University, Aligarh.
Diagnosis and relationship
Tricephalobus quadripapilli sp. n. is characterized by very fine transverse
annulations, a granular band around the gymnostom region, lateral fields with
two incisures, a well developed median bulb, ovary extending upto rectum
region, a constriction between glandular and muscular parts of uterus, sett off
spermatheca, broad spicules about 18-24 µm long, genital papillae four pairs and
tail with narrowed posterior half region.
The new species closely resembles T. steineri (Andrassy, 1952), Rühm,
1956 in general morphology and morphometrics. However, it can be
differentiated from T. steineri in having smaller post-uterine sac (less than half vs
one corresponding body diam. long), smaller tail (2.6-3.7 vs 4 ABD long in
females and 2.2-2.5 vs 3.5 ABD long in males), number of precloacal genital
papillae (one vs three).
94
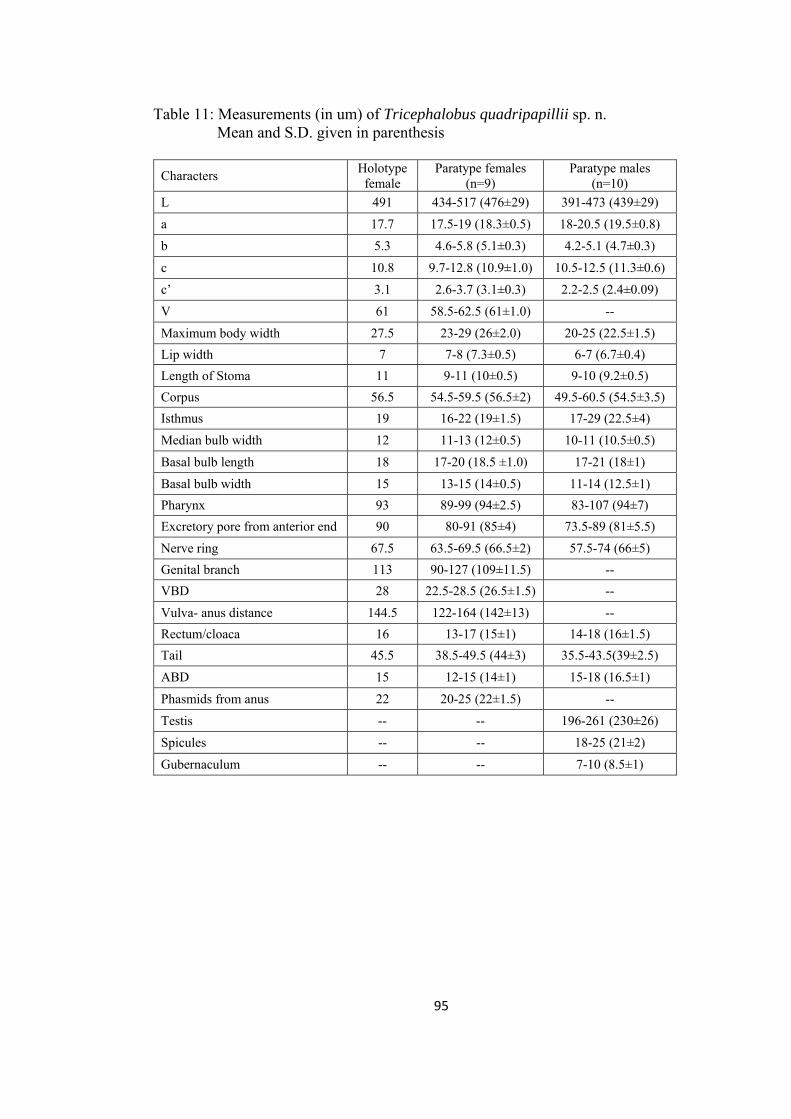
Table 11: Measurements (in um) of Tricephalobus quadripapillii sp. n. Mean and S.D. given in parenthesis
Characters Holotype
female Paratype females
(n=9) Paratype males
(n=10)
L 491 434-517 (476±29) 391-473 (439±29)
a 17.7 17.5-19 (18.3±0.5) 18-20.5 (19.5±0.8)
b 5.3 4.6-5.8 (5.1±0.3) 4.2-5.1 (4.7±0.3)
c 10.8 9.7-12.8 (10.9±1.0) 10.5-12.5 (11.3±0.6)
c’ 3.1 2.6-3.7 (3.1±0.3) 2.2-2.5 (2.4±0.09)
V 61 58.5-62.5 (61±1.0) --
Maximum body width 27.5 23-29 (26±2.0) 20-25 (22.5±1.5)
Lip width 7 7-8 (7.3±0.5) 6-7 (6.7±0.4)
Length of Stoma 11 9-11 (10±0.5) 9-10 (9.2±0.5)
Corpus 56.5 54.5-59.5 (56.5±2) 49.5-60.5 (54.5±3.5)
Isthmus 19 16-22 (19±1.5) 17-29 (22.5±4)
Median bulb width 12 11-13 (12±0.5) 10-11 (10.5±0.5)
Basal bulb length 18 17-20 (18.5 ±1.0) 17-21 (18±1)
Basal bulb width 15 13-15 (14±0.5) 11-14 (12.5±1)
Pharynx 93 89-99 (94±2.5) 83-107 (94±7)
Excretory pore from anterior end 90 80-91 (85±4) 73.5-89 (81±5.5)
Nerve ring 67.5 63.5-69.5 (66.5±2) 57.5-74 (66±5)
Genital branch 113 90-127 (109±11.5) --
VBD 28 22.5-28.5 (26.5±1.5) --
Vulva- anus distance 144.5 122-164 (142±13) --
Rectum/cloaca 16 13-17 (15±1) 14-18 (16±1.5)
Tail 45.5 38.5-49.5 (44±3) 35.5-43.5(39±2.5)
ABD 15 12-15 (14±1) 15-18 (16.5±1)
Phasmids from anus 22 20-25 (22±1.5) --
Testis -- -- 196-261 (230±26)
Spicules -- -- 18-25 (21±2)
Gubernaculum -- -- 7-10 (8.5±1)
95
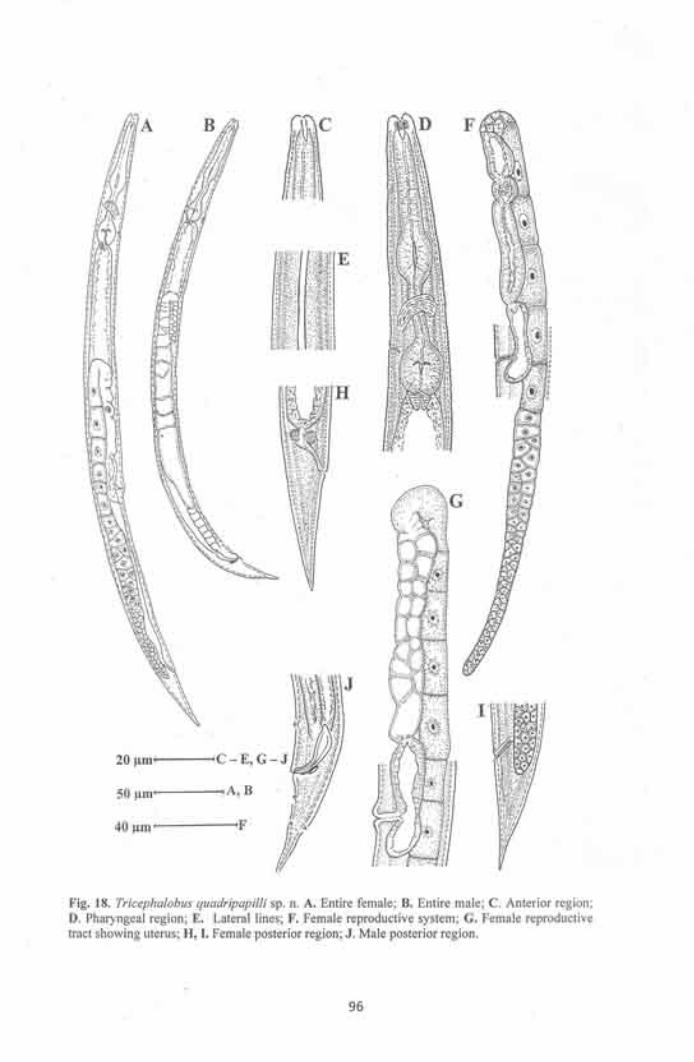
Fig. 19. Tricephalobus quadripapilli sp. n. A. Pharyngeal region, B. Anterior region showing stoma, C. Anterior region showing granular band around gymnstom, D. Lateral lines, E. Vulval region showing post-uterine sac, F. Female reproductive tract showing uterus, G. Female posterior region, H. Spicules and gubernaculum, I. Male posterior region (Scale bars = 20µm).
97
H G
F
I
A B
C
D
E

Family Brevibuccidae Paramonov, 1956
Diagnosis: Lips six, stoma relatively small, nearly twice as long as wide.
Pharyngeal corpus cylindrical or posteriorly swollen into a strong terminal bulb.
Female genital organ panagrolaimoid. Reflexed part of ovary not reaching to
vulva. Post-vulval uterine sac absent.
Type and only subfamily: Brevibuccinnae Paramonov, 1956.
Subfamily Brevibuccinae Paramonov, 1956
Diagnosis: Lips six, stoma relatively small, nearly twice as long as wide or
smaller or equal to lip region width. Pharyngeal corpus cylindrical or posteriorly
swollen. Reflexed part of ovary not reaching to vulva. Post-vulval uterine sac
absent. Both arms of spicules are equal or unequal, species often with unusually
long spicules.
Type genus: Brevibucca Goodey, 1935
Other genera: Cuticonema Sanwal, 1959
Plectonchus Fuchs, 1930
Genus Brevibucca Goodey, 1935
Diagnosis: Stoma short, expanding slightly at base, cheilostom and gymnostom
thick walled, forming half stoma depth; stegostom with an inwardly projecting
short tooth. No pharyngeal collar. Pharyngeal corpus cylindrical, isthmus short,
terminal bulb with grinder. Vulva posterior, gonad mono-prodelphic. Ovary
98

reversed not reaching vulval opening. A uninucleate gland cell on each side of
the uterus close to vagina. Testis outstretched. Spicules paired but of unequal
size. Gubernaculum present. Bursa absent. Genital papillae eight pairs.
Brevibucca postamphidia sp. n.
(Fig. 20, 21)
Measurements: In Table 12.
Females: Body slender, slightly arcuate upon fixation, tapering towards both
ends. Cuticle with fine transverse and longitudinal striations. Punctations fine,
sub-cuticular, transversely arranged. Lips separate, with minute hair like papillae.
Amphidial openings large, oval, post-labial at about 1/3rd of stoma length from
anterior end. Amphidial canal and fovea prominent. Cheilostom well developed,
slightly longer than wide, with thick cuticularised walls. Gymnostom short and
cuticularised. Stegostom anisomorphic. Dorsal wall provided with a tooth, each
subventrals with a smaller tooth. Pharyngeal corpus cylindrical, muscular, about
2.5 times isthmus length. Corpus isthmus junction distinct. Isthmus tubular. Basal
bulb ovoid, with well developed grinder and single haustrulum. Excretory pore
at level of corpus region, at about 38-43% of the pharyngeal length. Paired
excretory cells present just above basal bulb. Nerve ring encircling isthmus just
below corpus-isthmus junction, approximately at 62-66% of pharyngeal length.
Pharyngo-intestinal junction with well developed cardia consisting of three flaps.
Intestinal cells large, granular. Intestinal lumen variable in width.
99

Female reproductive system mono-prodelphic, ovary reflexed. Oviduct
narrow, joining ovary at a point slightly posterior to anterior tip. Spermatheca set
off by a slight dilation. Uterus undifferentiated, filled with embryonated eggs
and /or sperms. Up to ten eggs may be present at a time in the uterus. Vagina
strongly muscular, vaginal lumen unusually curved. Vulva a small transverse slit,
situated about 2.5 anal body diam. anterior to the anus. A uninucleate gland cell
on each side of the uterus close to vagina, connected by a duct to the uterus.
Post-uterine sac absent. Rectum 1.5-1.9 anal body diam. long, usually with wide
lumen. Tail extremely long filiform, 11-24 anal body diam., with fine tip.
Phasmidial openings distinct, 2-3 anal body diam. posterior to anus. Phasmidial
glands extending to just below the rectum.
Males: Body smaller than females, strongly curved ventrally in the posterior
region. Anterior end similar to that of females. Testis single, straight,
outstretched. Spicules dissimilar and of unequal size, larger one 27-30µm long
and smaller one 20-22μm long. Gubernaculum about half of larger spicule in
length, slightly curved proximally and with a sleeve distally. Genital papillae
eight pairs; three pre-cloacal (two subventrals and one lateral adcloacal), five
post-cloacal pairs. Of five postcloacal pairs, one subventral pair is located just
posterior to cloacal opening while three subventrals and one subdorsal pair are
closely grouped. This group is present just beyond phasmid, at the end of conoid
part of tail. Male tail with conoid part and a long filiform part.
Type habitat and locality: Decaying banana rhizome collected from Haldwani,
Uttarakhand.
100

Type specimens
Holotype female on slide Brevibucca postamphidia sp. n./1; twenty one
females and twelve males (paratypes) on slides Brevibucca postamphidia sp.
n./2-14, deposited in the nematode collection of Department of Zoology, Aligarh
Muslim University, Aligarh.
Diagnosis and relationship
Brevibucca postamphidia sp. n. is characterized by post-labial amphids
and large amphidial openings. Pharyngeal corpus about 2.5 times isthmus length.
Strongly muscular vagina with unusually curved vaginal lumen. Vulval slit about
2.5 anal body diam. to anus. A uninucleate gland cell on either side of uterus.
Long filiform female tail, about 11-24 anal body diam. in length (c=2.6-3.9).
Males with unequal and dissimilar spicules and eight genital papillae.
The new species closely resembles Brevibucca punctata Timm, 1960 in
general morphology and morphometric values in females but differs from it in
having smaller ‘c’ value in males (5.9-7.7 vs 8-10), structure of testis (outstreched
vs reflexed), size of gubernaculum (12-13 µm vs 18-22 µm) and in the position of
genital papillae (subdorsal pair anterior or almost at level of subventral group vs
subdorsal pair located posterior to subventral group).
101
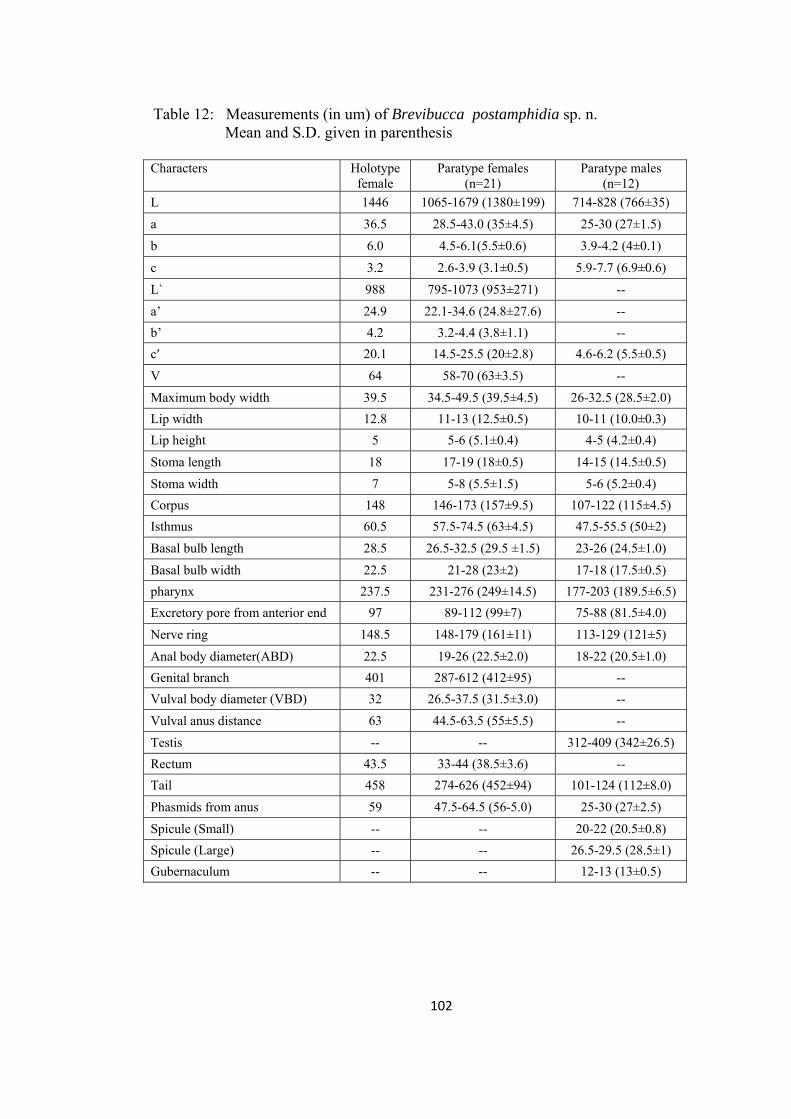
Table 12: Measurements (in um) of Brevibucca postamphidia sp. n. Mean and S.D. given in parenthesis Characters Holotype
female Paratype females
(n=21) Paratype males
(n=12)
L 1446 1065-1679 (1380±199) 714-828 (766±35)
a 36.5 28.5-43.0 (35±4.5) 25-30 (27±1.5)
b 6.0 4.5-6.1(5.5±0.6) 3.9-4.2 (4±0.1)
c 3.2 2.6-3.9 (3.1±0.5) 5.9-7.7 (6.9±0.6)
L` 988 795-1073 (953±271) --
a’ 24.9 22.1-34.6 (24.8±27.6) --
b’ 4.2 3.2-4.4 (3.8±1.1) --
c′ 20.1 14.5-25.5 (20±2.8) 4.6-6.2 (5.5±0.5)
V 64 58-70 (63±3.5) --
Maximum body width 39.5 34.5-49.5 (39.5±4.5) 26-32.5 (28.5±2.0)
Lip width 12.8 11-13 (12.5±0.5) 10-11 (10.0±0.3)
Lip height 5 5-6 (5.1±0.4) 4-5 (4.2±0.4)
Stoma length 18 17-19 (18±0.5) 14-15 (14.5±0.5)
Stoma width 7 5-8 (5.5±1.5) 5-6 (5.2±0.4)
Corpus 148 146-173 (157±9.5) 107-122 (115±4.5)
Isthmus 60.5 57.5-74.5 (63±4.5) 47.5-55.5 (50±2)
Basal bulb length 28.5 26.5-32.5 (29.5 ±1.5) 23-26 (24.5±1.0)
Basal bulb width 22.5 21-28 (23±2) 17-18 (17.5±0.5)
pharynx 237.5 231-276 (249±14.5) 177-203 (189.5±6.5)
Excretory pore from anterior end 97 89-112 (99±7) 75-88 (81.5±4.0)
Nerve ring 148.5 148-179 (161±11) 113-129 (121±5)
Anal body diameter(ABD) 22.5 19-26 (22.5±2.0) 18-22 (20.5±1.0)
Genital branch 401 287-612 (412±95) --
Vulval body diameter (VBD) 32 26.5-37.5 (31.5±3.0) --
Vulval anus distance 63 44.5-63.5 (55±5.5) --
Testis -- -- 312-409 (342±26.5)
Rectum 43.5 33-44 (38.5±3.6) --
Tail 458 274-626 (452±94) 101-124 (112±8.0)
Phasmids from anus 59 47.5-64.5 (56-5.0) 25-30 (27±2.5)
Spicule (Small) -- -- 20-22 (20.5±0.8)
Spicule (Large) -- -- 26.5-29.5 (28.5±1)
Gubernaculum -- -- 12-13 (13±0.5)
102
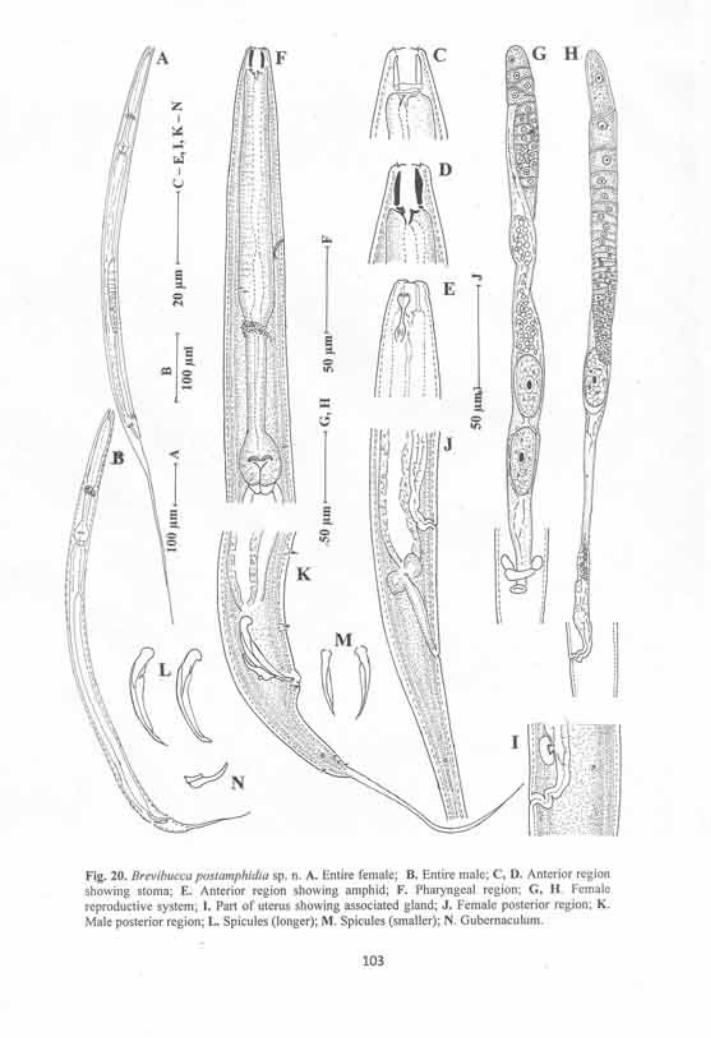
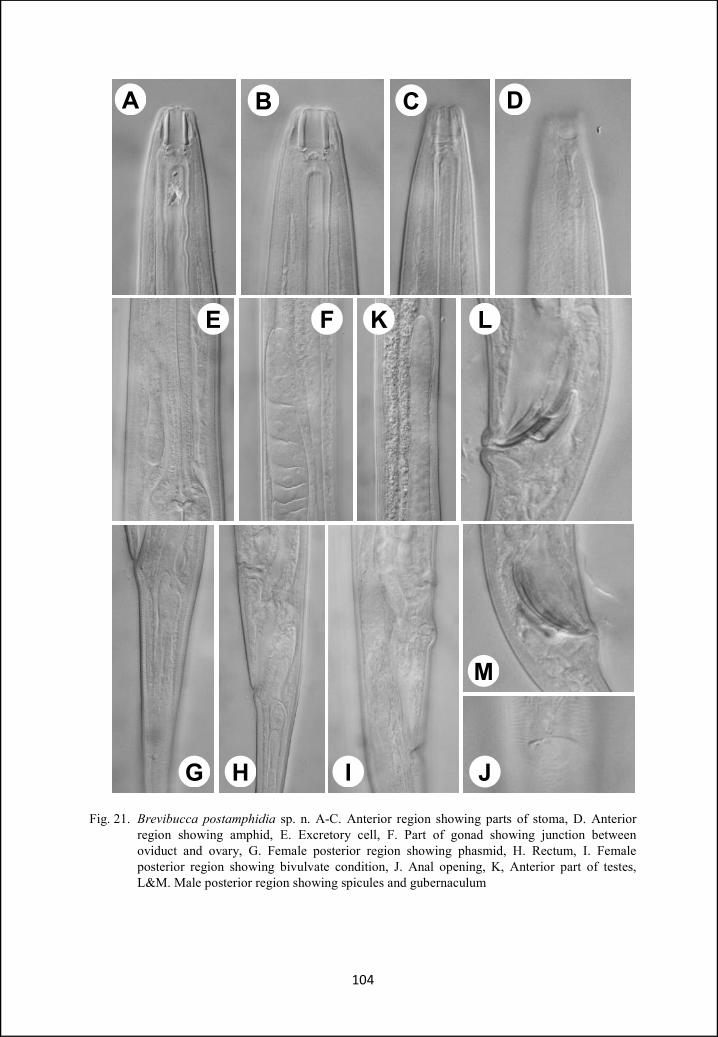
Fig. 21. Brevibucca postamphidia sp. n. A-C. Anterior region showing parts of stoma, D. Anterior region showing amphid, E. Excretory cell, F. Part of gonad showing junction between oviduct and ovary, G. Female posterior region showing phasmid, H. Rectum, I. Female posterior region showing bivulvate condition, J. Anal opening, K, Anterior part of testes, L&M. Male posterior region showing spicules and gubernaculum
104
A B C D
LKFE
G H I J
M

Genus Plectonchus Fuchs, 1930
Diagnosis: Body slightly curved ventrally when heat-killed. Cuticle with delicate
annulations and fine transverse striae. Lateral fields with two ridges. Lip region
with six low lips slightly separated. Amphids circular. Stoma with a wide and
short cheilostom and gymnostom. Corpus cylindrical, without a distinction
between pro- and metacarpus. Isthmus long and narrow. Basal bulb rounded or
pyriform with distinct grinders. Nerve ring encircling the isthmus. Female
reproductive system monodelphic-prodelphic.
Plectonchus coptaxii sp. n.
(Fig. 22, 23)
Measurements: In Table 13.
Females: Body slender, straight to slightly ventrally curved after fixation,
tapering abruptly and curved strongly beyond vulva. Cuticle 2-3 µm thick,
hyaline, with very fine transverse striations. Lateral fields indistinct. Lips round
with short hair-like papillae. Amphidial apertures oval, post-labial in position.
Stoma very shallow, rhabdions fused and poorly developed, without any
armature. Pharynx panagrolaimoid. Pharyngeal corpus cylindrical, longer than
the combined length of isthmus and basal bulb. Corpus-isthmus junction very
gradual sometimes indistinct. Isthmus tubular, 0.45-0.6 times corpus length.
Basal bulb pyriform, with distinct grinder. Nerve ring encircling isthmus in its
anterior half, at 58-66% of the neck length. Excretory pore in the corpus region,
at 43-47 % of neck length. Dierids and hemizonid indistinct. Cardia conoid,
surrounded by intestinal tissue. Intestine with wide lumen.
105

Reproductive system monodelphic-prodelphic. Ovary on right side of
intestine, reversed, without flexure, never extending beyond vulva. Oocytes
arranged in two rows in the germinal zone and in single row in the maturation
zone. Oviduct short. Spermatheca small, less than the corresponding body diam.
in length, slightly offset and without sperms. Uterus tubular differentiated into a
proximal and a distal part with thin walls and distinct lumen. The lumen of distal
part usually filled with a single row of granular structures. Vagina short,
anteriorly directed. Vulval lips depressed to form a sunken vulva. Post-uterine
branch absent. A ventral body pore is present slightly posterior to vulva. Rectum
straight, 1.1-1.3 anal body diam. long. Tail elongate conoid, 3.1-4.2 anal body
diam. long, terminus pointed. Phasmids about one anal body diam. posterior to
anus, at 27-35% of tail length.
Males: Anterior end similar to that of females, habitus ventrally curved, more in
the posterior region. Cuticle thick and hyaline like that of females. Testis reflexed
ventrally, on right side of intestine. Spicules dissimilar, unequal and separate.
Right spicule slightly longer than left, straight, slender, with a bifid tip. Left
spicule more robust, arcuate, with fusiform lamina and without a bifid tip.
Gubernaculum heavily sclerotized, more than half of spicule length, strongly
curved at its distal end. Genital papillae eight pairs. Two subventral pairs
precloacal and six pairs postcloacal. Of six postcloacal pairs, two pairs (one
subventral and one lateral) are anterior to phasmid. Two subdorsal pairs and two
subventral pairs are located posterior to phasmid. Tail conoid with acute
terminus.
106

Type habitat and locality: Organic manure collected from an agricultural field,
Mendhar, Jammu and Kashmir, India.
Type specimens
Holotype female on slide Plectonchus coptaxii sp. n./1; nine females and
eight males (paratypes) on slides Plectonchus coptaxii sp. n./2-5 deposited in the
nematode collection of Department of Zoology, Aligarh Muslim University,
Aligarh.
Diagnosis and relationship
Plectonchus coptaxii sp. n. is characterized by a slender body; thick
cuticle with hyaline portion and fine transverse striations. Lips with hair like
papillae. Amphids oval, post labial in position. Stoma shallow without any
armature, rabdions fused. Ovary reversed, without flexure. Distal part of uterus
with a single row of granular structures. Ventral body pore posterior to vulva.
Female tail elongate conoid, with narrowly rounded terminus. Male tail conoid,
with acute terminus. Spicules symmetric or asymmetric. Gubernaculum strongly
sclerotized, more than half spicule length. Genital papillae eight pairs.
The new species closely resembles P. molgos Massey, 1974 but can be
differentiated by the structure of cuticle (with transverse striae vs without
transverse striae), larger ‘a’ value in females (24.5-31.9 vs 20.4-24.4), smaller ‘b’
value (4.0-4.4 vs 5.1-5.6 in females and 3.9-4.1 vs 4.2-4.9 in males), more
posteriorly located vulva (V= 78-81% vs V=75%), shape of vulval lips
(depressed vs protuberant), tail shape (without constriction vs constricted) and
number and arrangement of genital papillae (eight pairs vs six pairs).
107

P. coptaxii sp. n. differs from P. wyganti Massey, 1964, in having smaller
body (485-585 µm vs 700 µm in females and 472-517 µm vs 600 µm in males),
smaller ‘b’ value (4.0-4.4 vs 4.5 in females and 3.9-4.2 vs 4.6 in males), greater
‘c’ value in males (12-13 vs 10.8), number and arrangement of genital papillae
(eight pairs vs seven pairs).
108
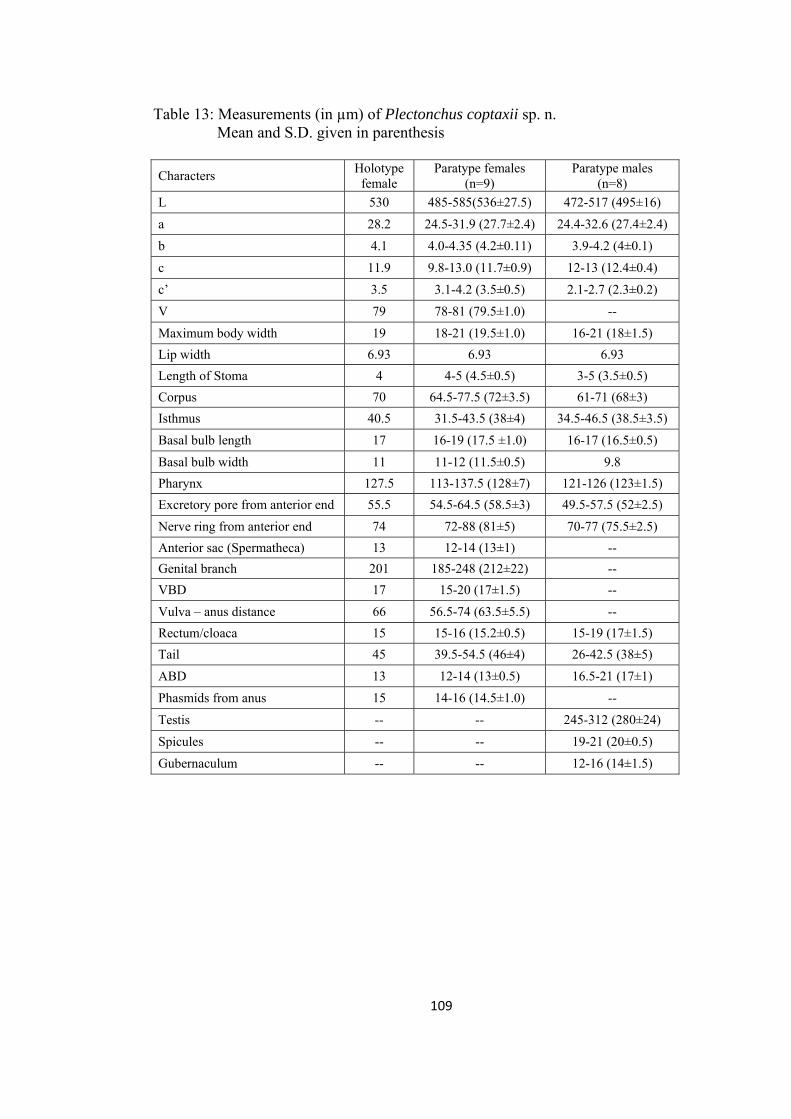
Table 13: Measurements (in µm) of Plectonchus coptaxii sp. n. Mean and S.D. given in parenthesis
Characters Holotype
female Paratype females
(n=9) Paratype males
(n=8)
L 530 485-585(536±27.5) 472-517 (495±16)
a 28.2 24.5-31.9 (27.7±2.4) 24.4-32.6 (27.4±2.4)
b 4.1 4.0-4.35 (4.2±0.11) 3.9-4.2 (4±0.1)
c 11.9 9.8-13.0 (11.7±0.9) 12-13 (12.4±0.4)
c’ 3.5 3.1-4.2 (3.5±0.5) 2.1-2.7 (2.3±0.2)
V 79 78-81 (79.5±1.0) --
Maximum body width 19 18-21 (19.5±1.0) 16-21 (18±1.5)
Lip width 6.93 6.93 6.93
Length of Stoma 4 4-5 (4.5±0.5) 3-5 (3.5±0.5)
Corpus 70 64.5-77.5 (72±3.5) 61-71 (68±3)
Isthmus 40.5 31.5-43.5 (38±4) 34.5-46.5 (38.5±3.5)
Basal bulb length 17 16-19 (17.5 ±1.0) 16-17 (16.5±0.5)
Basal bulb width 11 11-12 (11.5±0.5) 9.8
Pharynx 127.5 113-137.5 (128±7) 121-126 (123±1.5)
Excretory pore from anterior end 55.5 54.5-64.5 (58.5±3) 49.5-57.5 (52±2.5)
Nerve ring from anterior end 74 72-88 (81±5) 70-77 (75.5±2.5)
Anterior sac (Spermatheca) 13 12-14 (13±1) --
Genital branch 201 185-248 (212±22) --
VBD 17 15-20 (17±1.5) --
Vulva – anus distance 66 56.5-74 (63.5±5.5) --
Rectum/cloaca 15 15-16 (15.2±0.5) 15-19 (17±1.5)
Tail 45 39.5-54.5 (46±4) 26-42.5 (38±5)
ABD 13 12-14 (13±0.5) 16.5-21 (17±1)
Phasmids from anus 15 14-16 (14.5±1.0) --
Testis -- -- 245-312 (280±24)
Spicules -- -- 19-21 (20±0.5)
Gubernaculum -- -- 12-16 (14±1.5)
109
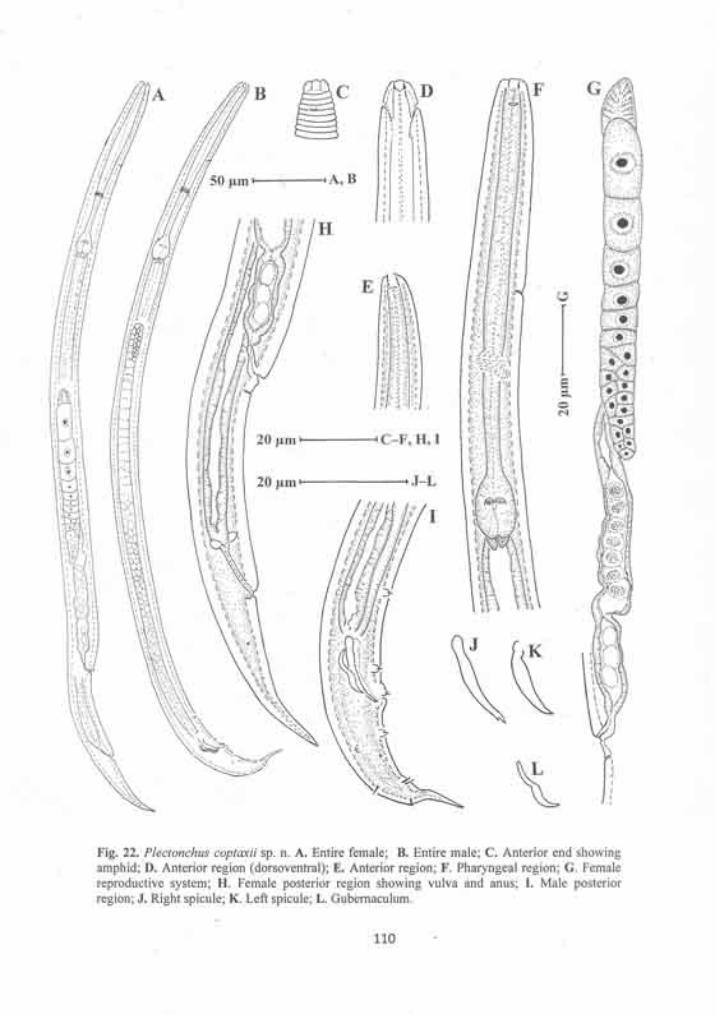
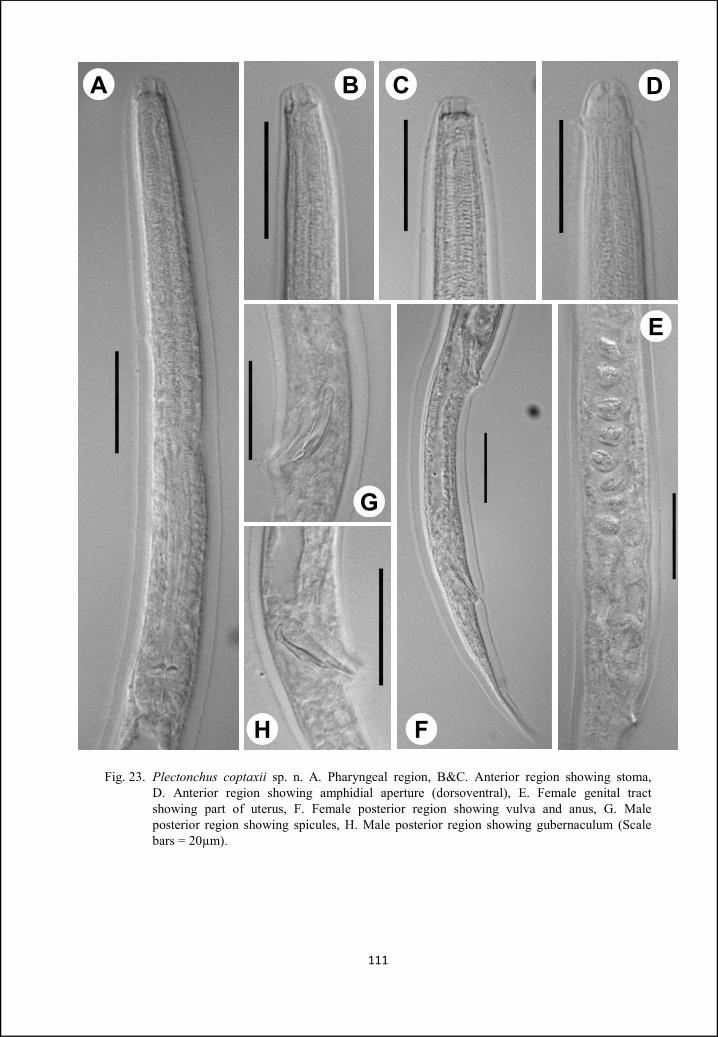
Fig. 23. Plectonchus coptaxii sp. n. A. Pharyngeal region, B&C. Anterior region showing stoma, D. Anterior region showing amphidial aperture (dorsoventral), E. Female genital tract showing part of uterus, F. Female posterior region showing vulva and anus, G. Male posterior region showing spicules, H. Male posterior region showing gubernaculum (Scale bars = 20µm).
111
A B C D
G
H F
E

Summary

The present work represents a taxonomic study of the nematodes of suborder
Cephalobina. Samples of soil and organic manure were collected from various parts
of the country in addition some samples from old collections were also studied. The
nematodes were isolated by Cobb’s sieving and decantation and modified
Baermann’s funnel techniques. The extracted nematode samples were examined
under stereoscopic microscope. Nematodes were simultaneously killed and fixed in
hot FA (4:1). Later, the nematodes were transferred into glycerine-alcohol (5:95) and
kept in a desiccator for dehydration. Dehydrated nematodes were mounted in
anhydrous glycerine on glass slides using wax as sealing material. All Measurements
were made on specimens mounted in dehydrated glycerine with an ocular
micrometer. De Man’s (1884) formula was used to denote the dimensions of
nematodes. All morphological observations and drawings were made on Nikon 80i
DIC microscope and photographs were taken by ProgRes C3 camera mounted on
Olympus BX 50 DIC microscope.
In all, thirteen species belonging to eleven genera, falling under two
superfamilies, three families and four subfamilies has been described. Of these ten
species which are new to science have been described and illustrated in addition
three known species have also been described. Two known species are being
reported for the first time from India. The terminology used in the text to describe
the parts of stoma is of De Ley et al. (1995).
112

113
The systematic position of genera and species, described in the present study are
given below
I. Order
Rhabditida
II. Suborder
Cephalobina
III. Superfamilies
1. Cephaloboidea 2. Panagrolaimoidea
IV. Families
1. Cephalobidae
2. Panagrolaimidae
3. Brevibuccidae
V. Subfamilies
1. Cephalobinae
2. Acrobelinae
3. Tricephalobinae
4. Brevibuccinae
VI. Genera
1. Pseudacrobeles
2. Acrobeles
3. Acrobeloides
4. Cervidellus
5. Chiloplacus
6. Nothacrobeles
7. Stegellata
8. Zeldia
9. Tricephalobus
10. Brevibucca
11. Plectonchus

VII. Species
1. Pseudacrobeles ventricauda sp. n.
2. Pseudacrobeles mucronata sp. n.
3. Acrobeles mariannae
4. Acrobeloides glandulatus sp. n.
5. Cervidellus neoalutus sp. n.
6. Cervidellus minutus sp. n.
7. Chiloplacus aligarhensis sp. n.
8. Nothacrobeles punctatus sp. n.
9. Stegellata ophioglossa
10. Zeldia tridentata
11. Tricephalobus quadripapilli sp. n.
12. Brevibucca postamphidia sp. n.
13. Plectonchus coptaxii sp. n.
114

Part – B Ecology

Introduction

Soil is home to many vertebrate and invertebrate species. Invertebrate
species largely outnumber the vertebrate species. The functional characteristics of
these invertebrates generally depend upon the soil characteristics and thus any
changes in soil condition effect the population dynamics of these invertebrates
and are easily reflected in various forms.
Soil microfauna, such as protozoa and nematodes, are important
constituents of soil foodwebs (Bonkowski, 2004). Their activities regulate the
size and function of fungal and bacterial populations in the soil (Ingham et al.,
1985; Poll et al., 2007), plant community composition (De Deyn et al., 2003) and
rates of carbon (Bradford et al., 2007) and nitrogen (Standing et al., 2006)
turnover. Nematodes are of particular interest because they are the most
numerous soil mesofauna and occupy all consumer trophic levels within the soil
foodweb. Therefore, their community structure can provide important insights
regarding many aspects of ecosystem function (Ritz & Trudgill, 1999; De Ruiter
et al., 2005).
The assemblage of plant and soil nematode species occurring in a natural
or a managed ecosystem constitutes the nematode community. These
communities are sensitive to changes in food supply (Yeates, 1987) and
environment (Bongers et al., 1991; Ettema & Bongers, 1993; Freckman &
Ettema, 1993; Samoiloff, 1987; Wasilewska, 1989). Thus, communities also have
a significant role in regulating decomposition and nutrient cycling (Anderson et
al., 1983; Ingham et al., 1985) and occupy a central position in the soil food web
(Moore & de Ruiter, 1991). When attributes of soil nematode communities are
quantified through measures such as diversity index (Shannon & Weaver, 1949)
115

or maturity index (Bongers, 1990; Yeates, 1994), an indication of relative soil
biological or ecological health is obtained, which can be used as one measure to
address issues of change in ecological condition of soils in agricultural systems.
Since, nematodes are so abundant and omnipresent in ecosystems, they
serve as elegant indicators of environmental disturbance (Bongers 1990; Ferris et
al., 2001; Yeates, 2003; Höss et al., 2004; Schratzberger et al., 2006; Heininger
et al., 2007). Nematodes possess the most important attributes of any prospective
bioindicator (Cairns et al., 1993): abundance in virtually all environments,
diversity of life strategies and feeding habits (Freckman, 1988; Yeates et al.,
1993), short life cycles, and relatively well-defined sampling procedures. Several
attempt have been made by researchers to develop relationships between
nematode community structure and succession of natural ecosystems or
environmental disturbance (Ettema & Bongers, 1993; Freckman & Ettema, 1993;
Freckman & Virginia, 1997; De Goede & Dekker, 1993; Wasilewska, 1994;
Yeates & Bird, 1994).
Soil microbes play an important role in plant nutrient cycling in organic
farming (Allison, 1973). Additions of organic matter to soil are expected to
increase numbers of bacteriovores and fungivorous nematodes and decrease
numbers of plant-parasitic nematodes (Bohlen & Edwards, 1994; Freckman,
1988; Griffiths et al., 1994). Applications of manure add both organic matter and
microbes, a source of food for the nematodes (Andrén & Lagerlöf, 1983; Weiss
& Larink, 1991). When bacteria are plentiful in soil, bacteriovorous nematodes
may discharge amino acids in substantial amounts. However, as bacterial
population decrease, nematodes begin to starve, and protein catabolism for
116

maintenance energy requirements leads to increased ammonium excretion by
nematodes (Anderson et al., 1983). Nitrogen content appears to be an important
measure of potential microbial activity and, subsequently, the rate of
decomposition (Neely et al., 1991).
Prior to the initiation of research on C. elegans, other than the activities of
a few taxonomists, most studies on soil nematodes centered on the biology and
management of those that cause damage to higher plants. A milestone in the
ecology of free-living soil nematodes was the seven-year study in Denmark by
Overgaard Nielsen (1949) on nematode faunae of different soils, their
physiological ecology and even their ecosystem services. Further notable
ecological contributions emerged in the 1970s and 1980s. Centres of ecological
study on nematodes developed in Sweden (Sohlenius, 1973), Poland (Prejs, 1970;
Wasilewska, 1970), Italy (Zullini, 1976), and Russia (Tsalolikhin, 1976). In the
USA, there was a surge of activity in soil ecology at the National Resource
Ecology Laboratory in Colorado Springs, led by Coleman and others (Yeates &
Coleman, 1982), and similar activity at the Institute of Ecology of the University
of Georgia, led by Crossley and colleagues (Stinner & Crossley, 1982). In the
same time period, Yeates was developing a very productive programme on the
ecology of soil nematodes in New Zealand (Yeates, 1979). A significant
contribution was the publication of the PhD research of Ingham, with its
accompanying review of preceding studies, in which the functional significance
of bacterivore and fungivore nematodes was established by the demonstration
that their excretion of nitrogen in excess of structural and metabolic needs
stimulated plant growth (Ingham et al., 1985).
117

The ecological classification of terrestrial nematodes has usually been
based on their feeding biology (trophic functions) and on the life strategies viz.
colonizers vs persiters (Bongers, 1990). Yeates et al., (1993) categorized
terrestrial nematodes into eight feeding groups, but most ecologists classify soil
nematodes into five feeding groups as discussed below (Yeates, 1998, Yeates &
Bongers, 1999).
1. Plant feeding/herbivores: - Nematodes feeding on vascular plants use a
tylenchid stylet/ dorylaimoid stylet or onchiostyle. They may be further
categorised on the habit of feeding an area of plant on which they feed.
2. Hyphal feeding/Fungivores:- Nematodes in this category puncture fungal
hyphae by a stylet with narrow lumen.
3. Bacterial feeding/bacteriovores:- This category includes species that feed
on any prokaryotic food source, through a narrow or broad stoma. They may
also ingest other types of food. Soil stages of parasites of vertebrates and
invertebrates are also included in this category.
4. Predators:- Feed on other nematodes or small invertebrates such as rotifers
and enchytraieds. They may be either ingestors or piercers.
5. Omnivores:- They appear to feed on a range of food source. It is usual to
restrict the term omnivore to some dorylaimids.
Within the five groups, strong relationship are found between herbivores
and fungal feeders, between herbivores and predators and between fungal feeders
and predators (Yeates et al.,1993 and Gomes et al., 2003). Degree of correlation
between different trophic groups is calculated by using the Karl Pearson’s
coefficient of correlation. It is used to measure the degree of relationship between
118

two or more variables, which is based on arithmetic mean and/or standard
deviation.
The idea of using ecological indices as indicators of ecosystem quality
(e.g. diversity, stability, and resilience) has received increased attention over the
last decade. Indices may be useful tools because they not only provide
quantitative means to characterize an ecosystem, but also to compare different
ecosystems (Ferris et al., 1996; Porazinska et al.,1998; Yeates & Bird, 1994; and
Yeates et al.,1997). Based on the densities of genera and trophic groups,
ecological indices of the nematode community are derived. The Shannon-Weaver
diversity index (Shannon & Weaver, 1949) is used to compare diversity of either
genera or trophic groups, and Simpson index (1949) is used to compare either
generic or trophic dominance. Maturity index Bongers (1990) and Yeates (1994)
is a semi-quantitative measure since it takes into consideration biological and
ecological characteristics of individual nematode species comprising a particular
community and these indices seems to offer better prospects for detecting and
sufficiently illustrating changes in the soil environment. The maturity index is
calculated as a weighted mean of the c-p values of nematodes in the sample.
Bongers (1990) defined two types of maturity indices: Maturity index (MI) which
includes nematodes belonging to all feeding types except herbivores, and Plant
parasitic index (PPI) which includes herbivores only. In general, the higher a
maturity index value, the more mature and stable the ecosystem. Based on effects
of the geographic distribution of nematodes, nematode feeding types, soils and
succession. Bongers et al., (1995) demonstrated that under certain conditions the
PPI and MI behave in opposite manners and suggested that an increase in the
119

PPI/MI ratio might reflect ecosystem enrichment. It has been found that PPI/MI
is an effective indicator of enrichment in agro-ecosystems (Ferris et al., 2001).
The combination of nematode feeding groups and cp scaling into
functional guilds have developed nematode faunal analysis into a powerful tool
which can be used as an indicator of soil health and soil food web conditions.
Various functional guilds of nematodes have been described to compute
Enrichment index (EI) and Structure index (SI). Enrichment index is based on
expected responsiveness of the opportunistic guilds (Ba1) to the food resource
enrichment. Thus, EI describes whether a soil ecosystem is nutrient enriched
(high EI) or depleted (low EI). The SI represents an aggregation of functional
guilds with cp values ranging from 3-5. SI describes whether soil ecosystem is
structured/ matured (high SI) or disturbed/ degraded (low SI). In addition to these
indices, Channel index (CI) is also calculated, which is the percentage of
fungivores among the total of fungivores and opportunistic bacteriovores to
describe the dominant decomposition channels in the food web. The CI also
provides a mean of tracking succession between fungivore and bacteriovore
nematodes as organic resources are supplied and depleted in agricultural systems
(Ruess & Ferris, 2002). Decomposition rates of readily degraded materials in
bacterial pathways are expected to be faster than that in fungal pathways where
materials may be more complex. Due to the similarity of C/N of fungi and
fungivore nematodes, mineralization rates in fungal channels is slower than those
in bacterial channels where there is greater difference between C/N ratios of
predators and prey (Ingham et al., 1985; Ferris et al., 1997; Chen & Ferris, 1999;
Okada & Ferris, 2001). Such analysis provides slightly different information than
120

other analysis based on biomass or diversity and take into consideration both
descriptive and quantitative information on the soil ecosystem. Further, the
redistribution of bacteria to new food sources by survival of passage through the
nematode intestine is an important accelerator of decomposition process
(Wasilewska et al., 1981; Freckman, 1988) for which fungivore nematodes
probably do not provide an equivalent function in the fungal channel.
Nematode faunal analysis is evolving as a powerful bioindicator of the
soil condition and of structural and functional attributes of the soil food web
(Bongers & Ferris, 1999; Neher, 2001). Recent developments of such analyses
include recognition of an enrichment trajectory and a structure trajectory. The
latter measures the abundance of trophic linkages in the food web and the
probability of regulatory effects on opportunist populations through exploitation
and competition. The enrichment trajectory reflects supply-side characteristics of
the food web and the increase in the primary consumers of incoming organic
material (Ferris et al., 2001). Food webs become enriched when disturbance
occurs and resources become available due to external input, organism mortality,
turnover, or favorable shifts in the environment (Odum, 1985; Van Veen &
Kuikeman, 1990). The enrichment-opportunist bacteriovore nematode guild
includes species in the families Rhabditidae, Panagrolaimidae and Diplogastridae
(Bongers & Ferris, 1999; Ferris et al., 2001). They are classified as cp-1
organisms, characterized by short generation time, small eggs and high fecundity,
in the colonizer-persister scale of Bongers (1990). They appear to feed
continuously in the enriched media and then form metabolically suppressed
dauerlarvae as resources are diminished. As microbial blooms fade, enrichment-
121

oppurtunist microbivores may be replaced by general-opportunist with
specialized morphological, physiological and behavioral adaptations for more
deliberate feeding on less-available resources. The general-opportunist
nematodes, classified as cp-2, are predominantly bacterial scavengers in the
Cephalobidae and fungal-feeders in the Aphelenchidae, Aphelenchoididae and
Anguinidae. Increased abundance of fungal-feeding opportunists occurs when the
available organic pool is conductive to fungal decomposition as, for example,
when complex organic material becomes available in the soil or when fungal
activity is enhanced under conditions less favorable for bacterial decomposition
(Eitminaviĕūté et al., 1976; Wasilewska et al., 1981). In fact, nematodes in the
general-opportunist guild commence to increase with the initial enrichment, but
at slower rate than the enrichment opportunists so that they become
successionally predominant as the latter guild is declining (Bongers, 1990; Ferris
et al., 1996b, 2001; Bongers & Bongers, 1998; Bongers & Ferris, 1999; Chen &
Ferris, 2000).
Studies are being carried out throughout the world on various aspects of
nematode ecology. Most of the studies are concentrating on the impact of heavy
metals on nematode assembleges and secondary succession of nematode guilds.
Zhang et al. (2007) studied responses of nematode assemblages to Cu and Zn
pollution on corn fields near a copper smelter in Northeast China. Dechang et al.
(2009) & Tomar et al. (2009) studied effects of heavy metals on nematode
functional guilds in agro-ecosystems of Shenyang, Liaoning province, China and
reported that the concentrations of lead were most significant at distance of 20 m
from the highway, and than concentration gradually decreased to further
122

distances. Cheng & Grewal (2009) studied dynamics of soil food web under
urban landscape and reported that anthropogenic activities resulting in the loss of
top soil can have profound effect on the structure of soil food web, which may
severely limit its capacity to support optimal nutrient cycling. Mahran et al.
(2009) studied response of nematode communities to hog manuring and reported
that hog manuring is effective in killing plant parasitic nematodes. Hánèl (2010)
studied succession of nematodes in agro-ecosystems and compared it with
abandoned fields in South Bohemia. Biodiversity and trophic structure of soil
nematode communities in subtropical ecosystems was studied by Bierdermann &
Boutton (2010). Their study mainly focused on nematode responses to woody
plant encroachment in urban areas. They reported that energy flow through
nematode food webs in grasslands appear relatively diversified and SI indicated
that nematode communities within old clusters were more simplified than those
in grassland due to reduction in densities of omnivores and predators. Soil
nematode communities of a banana planted agro-ecosystem were studied by
Tabarant et al. (2011). They concluded that plant residue and bagasse which are
mainly composed of cellulose and lignins which are difficult to decompose.
These chemicals mainly favor fungal decomposition pathways and permitted
development of carnivorous nematodes and thus increasing channel index. Park
et al. (2011) studied effects of heavy metal contamination in abandoned mines.
They examined two areas one with heavy metal infection from mines and another
control area which had no infection and found that diversity of nematodes was
less in contaminated area as compared with non contaminated area. Liu et al.,
(2011) studied effects of biological crusts on soil nematode communities and
123

found that nematode abundances, generic richness and food web based indices as
EI and SI positively correlated with different crust ages.
The nematodes have been widely studied in India in contrast of plant
parasites and control of nematode menace in agro-ecosystems. Various
researchers have used nematodes for studying effects of organic amendments in
field and micro-plot experiments. Singh (1970) studied control of plant parasitic
nematodes with organic soil amendments. Akhtar (1993, 1999 & 2000) studied
possible relation of organic soil amendments and nematodes. Devi & Das (1998)
studied effects of organic amendments of root knot nematodes. Hassan et al.
(2008 & 2009) studied nematodes of Zea mays and their possible control through
organic amendments. There are some sporadic reports of study of nematode
diversity in different ecosystems. Tomar et al. (2006) reported diversity of
nematodes at a Mango orchard in Aligarh. Baniyamuddin et al. (2007) studied
functional diversity of nematodes of natural forests in Arunachal Pradesh. Tomar
& Ahmad (2009) studied nematode community in a natural woodland of Aligarh
region, and reported it to be a stable ecosystem on the basis of maturity and plant
parasitic indices.
The aim of the present study was to investigate nematode communities
and temporal changes in the nematode fauna of two different habitats (crop field
and natural wasteland), for a period of one year. These two habitats were located
at a distance of 6 Kms from each other. Specifically, population dynamics, faunal
analysis, and community composition of soil nematodes have been studied to
assess the role of nematodes as indicators of soil condition.
124

Materials &
Methods

Site description: - The experimental sites a) Crop field (CF) and b) Wasteland
(WL) were selected on the basis of differences in their vegetation and soil
disturbance. Both the sampling sites were located in Aligarh district. The climate
of the region, in general the blocks in the west are drier as compared to those in
the east. The temperature rises as high as 44.6˚C during summer and drops down
to as low as 4.8˚C during winters. The mean maximum and mean minimum
temperature of district Aligarh is 26.7˚C and 15.5˚C respectively. Monsoon starts
in July and runs to September. While in winter season, the region receives very
few showers. The mean annual rainfall of the district is 434 mm, mean relative
humidity 65%. Thus the Climate of the district Aligarh is Semi-Arid. Served by
numerous rivers, rivulets and canals of the Ganga. Few rivers like Karwan,
Rutba and Kali pass through the district but remain almost dry except during
rainy season.
Site a):- The crop field situated about 14 Kms from Aligarh city, on Aligarh –
Moradabad highway (27° 57′ N, 78° 10′ E) was selected. The field was cultivated
under annual conventional cropping system whereby wheat was the only crop
sown during the sampling time with fallow periods in between. During sampling
crop was at different stages of development. Organic amendments were added to
the crop field in form of manures during September-October. Chemical fertilizer
mainly NPK were added to field during December-January
Site b) :- This site was natural wasteland, more than two decades old, situated
about 8 Kms from Aligarh city on Aligarh – Moradabad highway (27° 55′ N, 78°
54′ E) was selected for sampling. The wasteland covers more than two sq. Km
125

area on either side of highway. Wasteland usually remained dry and bare during
most of the time except monsoon. During monsoon most parts of the area were
covered by grasses.
Sampling
Soil samples from crop field were collected following the growth pattern
of the crop, simultaneously samples were also collected from wasteland. In
Aligarh district monsoon starts in July and runs to September. Wheat was sown
during the month of October/November and harvested during February/March.
Sampling was undertaken on eight occasions from June 2008 to April 2009 at
different stages of development of wheat crop including the fallow periods before
and after the cropping period. Each soil sample consists of five cores (1 cm2
cross sectional area) from a depth of 0-10 cm. All the samples were collected
windward in N-W direction. Samples were tagged, stored in sealed plastic bags
and brought to laboratory for further processing. For sampling from both the
areas a diagonal transect was selected and samples were collected from the same.
Processing of soil samples
About 100 cc of soil from each sample was processed by Cobb (1915)
sieving and decantation and modified Baermann’s funnel technique as discussed
earlier in Part A.
Isolation and Killing & Fixation of Nematodes
The isolation, killing and fixation of nematodes were done by the same
process as it was done in Part A.
126

Counting of Nematodes
Population count of nematodes was made using Syracuse counting dish.
The suspension was made homogenous by bubbling with pipette thoroughly
before taking 2 ml of nematode suspension in the dish for counting. Counting of
each sample was done three times and mean was obtained. The final population
was obtained by multiplying final quantity of nematode suspension (50 ml) with
mean number of nematodes counted and dividing by the quantity of suspension
used for counting (2 ml).
Identification
Mass slides containing about two hundred nematodes per sample were
prepared for identification. Identification up to generic level was done mainly
using Goodey (1963); Jairajpuri & Khan (1982); Andrássy (1984, 2005), Siddiqi
(1986), Jairajpuri & Ahmad (1992); Ahmad (1996). Trophic group were allocated
according to Yeates et al. (1993) and cp groups were assigned after Bongers
(1990).
Data Analysis
Nematode diversity was described using the univariate measures of the
Shanon index calculated at genus level (H’) and multivariate analysis was
performed by ANOVA using statistical programme SPSS. Shanon’s diversity
(H’) was calculated by SPECDIVE. Nematodes were assigned to five main
trophic groups (bacteriovores, fungivores, herbivores, omnivores and predators)
after Yeates et al. (1993). Maturity index (MI) was calculated to estimate the
relative state of two ecosystems studied. Trophic diversity was calculated by the
127

trophic diversity index, (TDI) (Heip et al., 1988). Structure index (SI) and
enrichment index (EI) were calculated to determine the relative stability of the
ecosystem studied. Nematode channel ratio was calculated to reflect differences
in the mineralization of dead and live plant tissues. (Wasilewska, 1994). In all the
above mentioned indices, nematode families were allocated cp scale according to
their perceived life history strategy.
Detailed description of the formulae used are given below
Shannon’s diversity (H′) = −Σ (pi ln pi)
Maturity Index (MI)
n
iifiVMI
1.
Where Vi= cp value of the ith taxon.
f(i) the frequency of that taxon in a sample
* Maturity index (MI) is calculated as the weighted mean of the individual
cp value.
Plant Parasitic index (PPI)
XiPPiXiPPI /
Where, Ppi = PP value assigned to taxon i according to Bongers (1990).
Xi = abundance of taxon i in the sample.
128

Nematode Channel Ratio (NCR)
FB
BNCR
Where, B = Total abundance of Bacterial feeding nematodes
F = Total abundance of Fungal feeding nematodes.
Enrichment index (EI) = (e/e+b) x100
Structure index (SI) = (s/s+b) x100
where e, b & s are sum products of assigned weights and number of
individuals of all genera (Table 1,2).
Trophic Diversity index (TDI) = 1 ⁄ ∑pi²
where pi² is the proportional contribution of ith trophic group.
129

Table 1:- Nematode functional guilds in different Food web conditions.
Basal (b) Structured (s) Enriched (e)
Ba2- Cephalobidae Ba3- Prismatolaimidae Ba1- Rhabditidae,
Panagrolaimidae
Fu2- Aphelenchidae,
Aphelenchoididae
Anguinidae.
Fu3- Diptherophoridae
Fu4- Leptonchidae
Pr2- Aphelenchid Carnivores
Pr3- Tripylidae
Pr4- Mononchidae
Pr5- Discolaimidae
Om4- Dorylaimidae
Om5- Thornenematidae,
Qudsianematidae
The numbers used in the table denotes the assigned c-p value to the trophic groups as follows:
Ba- Bacteriovores
Fu- Fungivores
Pr- Predators
Om- Omnivores
130

Results

Nematode Diversity
During the present course of study a total of 50 genera belonging to 10
orders and 30 families were encountered from the crop field (CF) while 46 genera
belonging to 8 orders and 25 families occurred in the wasteland (WL). The
number of genera varied from 13 to 23 per sample in crop field while in
wasteland it ranged from 7 to 21 per sample. In terms of abundance the number
varied from 105 to 1178 individuals in crop field and 106 to 568 in wasteland per
100 cc of soil. Acrobeloides was the most abundant genus in crop field while
Dorylaimellus was the most abundant genus in the wasteland.
In the crop field, in terms of number of genera (Fig. 1, A) the Order
Dorylaimida was most frequent (30%) with 15 genera under 8 families, followed
by Rhabditida (22%) with 11 genera under 4 families, Tylenchida (20%) with 10
genera under 6 families, Araeolaimida (8%) with 4 genera under 3 families,
Aphelenchida (6%) with 3 genera under 3 families, Enoplida (4%) with 2 genera
under 2 families & Monhysterida (4%) represented by 2 genera under 1 family,
while Alaimida (2%), Diptherophorida (2%) and Chromadorida (2%) were
represented by 1 genus each. However, in wasteland, (Fig. 1, C) Order
Dorylaimida (32%) was most prevalent with 7 families representing 15 genera
followed by Tylenchida (22%) with 10 genera under 6 families, Rhabditida
(22%) with 4 families representing 10 genera, Aphelenchida (7%) and
Chromadorida (7%) were represented by 3 genera each under 3 families and & 1
family respectively, while Araeolaimida (4%) and Monhysterida (4%) were
represented by 2 genera each under 2 families and 1 family respectively and
Enoplida (2%) was the least prevalent with single genus.
131

In terms of number of individuals (Fig. 1, B), Rhabditida (44%) was most
abundant, followed by Tylenchida (19%), Dorylaimida (12%), Aphelenchida
(11%), Araeolaimida (8%), Enoplida (2%), Alaimida, Chromadorida,
Monhysterida and Diptherophorida (1% each) in crop field. While in wasteland
(Fig. 1, D) Rhabditida (33%) was most abundant, followed by Dorylaimida
(27%), Aphelenchida (11%), Tylenchida (10%), Chromadorida (9%),
Araeolaimida (5%), Enoplida (4%) and Monhysterida (1%).
Trophic diversity
In the crop field (Fig. 2, A), bacteriovores constituted the most dominant
group in terms of number of genera and abundance and were represented by 20
genera. This was followed by herbivores, fungivores, omnivores and predators,
which were represented by 9, 8, 7 & 6 genera respectively. In terms of number of
individuals (Fig. 2, B) bacteriovores (58%) was the most abundant group,
followed by fungivores (21%), herbivores (13%), omnivores (5%) and predators
(3%). The trophic diversity index (TDI) of the area ranged from 1.03-1.25
(1.1±0.07). Among bacteriovores the genus Acrobeloides was most dominant
while the genera Hoplolaimus, Aphelenchus, Moshajia and Discolaimus were
most dominant among herbivores, fungivores, omnivores and predators
respectively. Least dominant genera among bacteriovores, herbivores, omnivores,
fungivores and predators were Drilocephalobus, Hemicriconemoides,
Thornenema, Leptonchus and Tripyla respectively (Table 2). Bacteriovores,
fungivores and herbivores were present in all samples while omnivores and
predators were absent from two samples each. In the crop field (Fig. 3) all
132

nematode population remained relatively low from June to December. Thereafter
the bacteriovores increased in numbers with a peak population in March followed
by a sharp decline. The fungivores followed a somewhat similar pattern but the
population remained comparatively low. All other groups showed peak
population in January but their number was very low.
In the wasteland (Fig. 2, C) also bacteriovores constituted the most
dominant group in terms of number of genera and abundance and were
represented by 18 genera. This was followed by herbivores with 10 genera,
omnivores with 7 genera, fungivores with 6 genera and predators with 5 genera.
The bacteriovores (51%) was the most abundant group in terms of number
followed by fungivores (29%), herbivores (11%), omnivores (6%) and predators
(3%) (Fig. 2, D). The trophic diversity index (TDI) of the area ranged from 1.05-
1.23 (1.12±0.06). Among bacteriovores, the genus Acrobeles was dominant,
while the Ditylenchus, Moshajia, Dorylaimellus and Seinura were most dominant
among herbivores, omnivores, fungivores and predators respectively. Least
dominant genera among various trophic groups were Stegellata, Psilenchus,
Tylencholaimus, Epidorylaimus and Discolaimoides (Table 3). All the samples
contained bacteriovores and fungivores, while herbivores were absent in one
sample and omnivores and predators were not present in four and five sample
respectively. In the wasteland (Fig. 3), population of all groups except herbivores
remained uniform from June to October, dipped in December and peaked of in
January followed by a rapid decline thereafter. At the peak population
bacteriovores were in maximum number followed by fungivores and omnivores.
The herbivore population remained relatively low and showed a peak in October.
133

Nematode Community Dynamics
The diversity of nematode fauna in agroecosystems and their relationships
to soil processes suggests that they are potential bioindicators. However, the
effects of plants, soil types and nematode biogeography mean a ‘functional
group’ may be a better indicator than particular nematodes. Permanent grassland
may be regarded as providing a baseline for nematode diversity in a given soil.
The relative abundance of fungal-feeding and bacterial-feeding nematodes serve
as sensitive indicator of management changes (Yeates & Bongers, 1999). For
assessing the community dynamics and role of nematodes in the agroecosystems,
various indices such as Shannon’s diversity index (H’), Maturity index (MI)
including plant parasitic families, Maturity index 2-5 (MI25, excluding Ba1
functional guild), Plant parasitic index (PPI), Nematode channel ratio (NCR) and
PPI/MI were calculated. The percent abundance of cephalobids, tylenchids,
dorylaims and other nematodes were also obtained.
Food web diagnostics of the ecosystem was studied in terms of
Enrichment index (EI), Structure index (SI) and Basal index (BI) following the
weighted faunal analysis concept of Ferris et al. (2001).
Shannon’s diversity index (H’) in crop field was found to be 2.21–2.73
(2.45±0.19) and in wasteland it ranged from 1.88–2.54 (2.17±0.22). The maturity
index (MI) ranged from 2.1–2.59 (2.35±0.18) in crop field and 2.36–3.64
(2.9±0.41) in wasteland. MI25 ranged from 2.13–2.59 (2.4±0.18) and 2.37–3.66
(2.91±0.41) in crop field and wasteland respectively. The plant parasitic index
(PPI) for the crop field varied from 2.21–3.19 (2.73±0.33) while it was 2.0–3.0
134

(2.65±0.41) for wasteland. Nematode channel ratio (NCR) was found to be 0.61–
0.94 (0.76±0.11) and 0.49–0.8 (0.63±0.1) and the values for PPI/MI were 1.01–
1.38 (1.16±0.13) and 0.72–1.28 (0.99±0.21) for crop field and wasteland
respectively (Table 4 & 5). Comparison of crop field samples for indices revealed
that MI was almost constant for all sampling times, while PPI was highest during
June which gradually decreased till December (i.e. growth phase) and than it
increased gradually. NCR also was almost constant throughout. In wasteland MI
showed a gradual increase till December and then gradually decreased, while PPI
and NCR were almost constant for the sampling times (Table 4 & 5, Fig. 4).
The enrichment index (EI) ranged from 13.5–41.1 (26.5±9.8) and 5.2–
38.9 (17.9±10.6) and the structure index (SI) was found to be 22.7–64.2
(47.7±15.8) and 43.3–90 (70.5±15) in the crop field and wasteland respectively.
Basal index (BI) varied from 31.47–68.9 (43.8±13.08) in the crop field and 9.76–
49.3 (26.3±12.15) in the wasteland (Table 4 & 5). Comparison of food web
indices for crop field indicated that BI was highest during December (i.e. growth
phase) showing a linear spline, while the values for SI gradually increased with a
sudden decrease in December. The EI values also followed same pattern as of SI.
In wasteland SI gradually increased till December and then remains constant
while BI gradually decreased till December and increased constantly for the next
sampling times. EI shows a sudden dip in the month of October and then it
gradually increased till the end of sampling (Table 4 & 5, Fig. 5).
The abundance of cephalobids per sample ranged from 32.2-58.6%
(42.5±9.4) in crop field and 14.4-44.6% (31.8±9.2) in wasteland while of
tylenchids it ranged from 8.9-39.8% (19.4±9.8) in crop field and 1.4-22.3%
135

(10.3±6.6) in wasteland. The dorylaims abundance varied from 1.9-19.5%
(11.7±6.2) and 12.6-44.9% (27.2±10.1) and others groups from 14.7-38.2%
(26.4±8) and 12.3-36.3% (30.7±8.3) in crop field and wasteland respectively
(Table 4 & 5, Fig. 6).
In crop field (Table 6), population of dorylaims show very high degree of
positive correlation with MI and SI (Fig. 7, A & B), and very significant negative
correlation with BI (Fig. 7, C). With all other indices dorylaims show either
negative correlation or almost no correlation. Population of tylenchids show low
level of positive correlation with PPI/MI ratio and to certain level with PPI (Fig.
7, D), while some degree of negative correlation with MI and SI. Population of
cephalobids show a low level of positive correlation with PPI and low level of
negative correlation with MI while almost no correlation was found with EI ans
SI. Almost no correlation was found with MI while it was negatively correlated
with PPI, EI and SI.
In Wasteland (Table 6), population of dorylaims showed very high degree
of positive correlation with MI and SI (Fig. 8, A & B) while significant negative
correlation was found with PPI/MI ratio and BI. Population of tylenchids show
some degree of positive correlation with MI, PPI and SI while they are negatively
correlated to some degree with EI, BI and PPI/MI ratio. Population of
cephalobids showed high degree of negative correlation with MI (Fig. 8, C) and
SI while significant positive correlation was found with H′ and PPI/MI ratio and
upto certain level with PPI, however, there was almost no correlation was found
with EI. Other nematodes show some degree of positive correlation with EI while
certain level of negative correlation with MI, PPI and SI was observed.
136

Significant negative correlation was also observed between population of
dorylaims and cephalobids (Fig. 8, D).
137
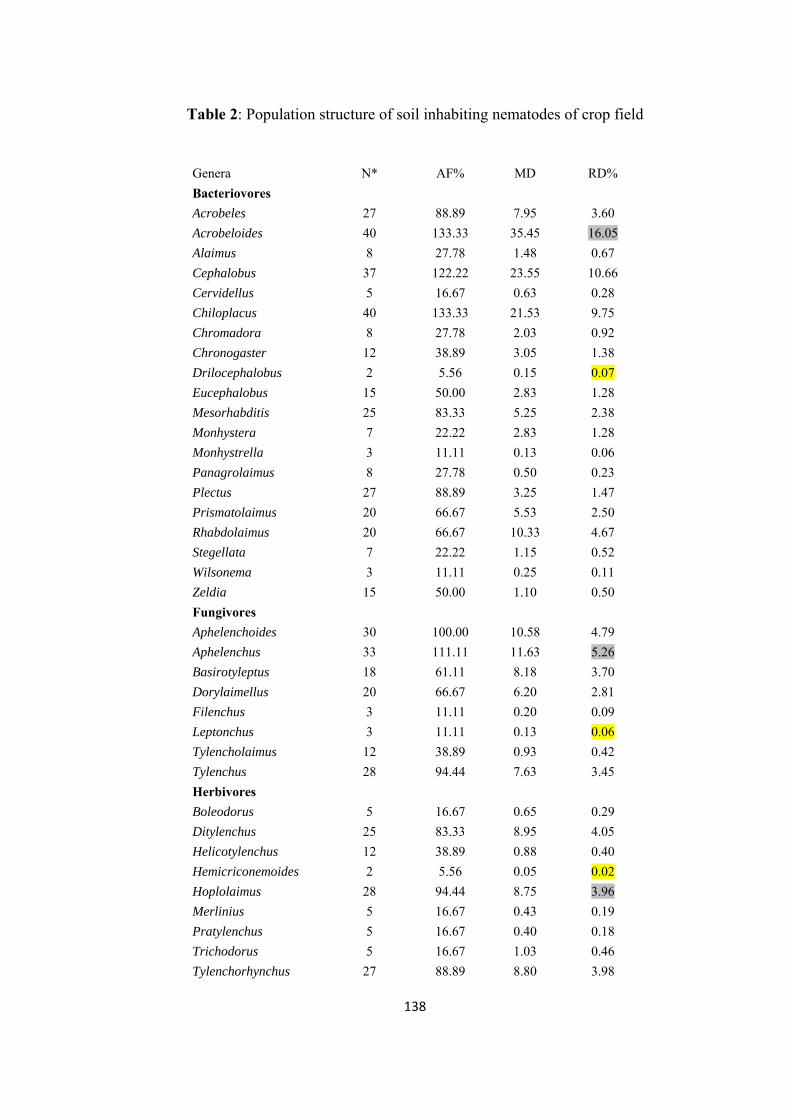
Table 2: Population structure of soil inhabiting nematodes of crop field
Genera N* AF% MD RD%
Bacteriovores
Acrobeles 27 88.89 7.95 3.60
Acrobeloides 40 133.33 35.45 16.05
Alaimus 8 27.78 1.48 0.67
Cephalobus 37 122.22 23.55 10.66
Cervidellus 5 16.67 0.63 0.28
Chiloplacus 40 133.33 21.53 9.75
Chromadora 8 27.78 2.03 0.92
Chronogaster 12 38.89 3.05 1.38
Drilocephalobus 2 5.56 0.15 0.07
Eucephalobus 15 50.00 2.83 1.28
Mesorhabditis 25 83.33 5.25 2.38
Monhystera 7 22.22 2.83 1.28
Monhystrella 3 11.11 0.13 0.06
Panagrolaimus 8 27.78 0.50 0.23
Plectus 27 88.89 3.25 1.47
Prismatolaimus 20 66.67 5.53 2.50
Rhabdolaimus 20 66.67 10.33 4.67
Stegellata 7 22.22 1.15 0.52
Wilsonema 3 11.11 0.25 0.11
Zeldia 15 50.00 1.10 0.50
Fungivores
Aphelenchoides 30 100.00 10.58 4.79
Aphelenchus 33 111.11 11.63 5.26
Basirotyleptus 18 61.11 8.18 3.70
Dorylaimellus 20 66.67 6.20 2.81
Filenchus 3 11.11 0.20 0.09
Leptonchus 3 11.11 0.13 0.06
Tylencholaimus 12 38.89 0.93 0.42
Tylenchus 28 94.44 7.63 3.45
Herbivores
Boleodorus 5 16.67 0.65 0.29
Ditylenchus 25 83.33 8.95 4.05
Helicotylenchus 12 38.89 0.88 0.40
Hemicriconemoides 2 5.56 0.05 0.02
Hoplolaimus 28 94.44 8.75 3.96
Merlinius 5 16.67 0.43 0.19
Pratylenchus 5 16.67 0.40 0.18
Trichodorus 5 16.67 1.03 0.46
Tylenchorhynchus 27 88.89 8.80 3.98
138

Omnivores
Epidorylaimus 3 11.11 0.08 0.03
Eudorylaimus 8 27.78 1.10 0.50
Latocephalus 3 11.11 0.05 0.02
Moshajia 32 105.56 8.30 3.76
Oriverutus 2 5.56 0.08 0.03
Thonus 8 27.78 0.63 0.28
Thornenema 2 5.56 0.05 0.02
Predators
Aporcelaimellus 10 33.33 0.93 0.42
Aquatides 5 16.67 0.70 0.32
Discolaimus 20 66.67 2.00 0.91
Discolaimoides 8 27.78 0.93 0.42
Seinura 10 33.33 1.73 0.78
Tripyla 2 5.56 0.03 0.01
*Mean of five replicates
139
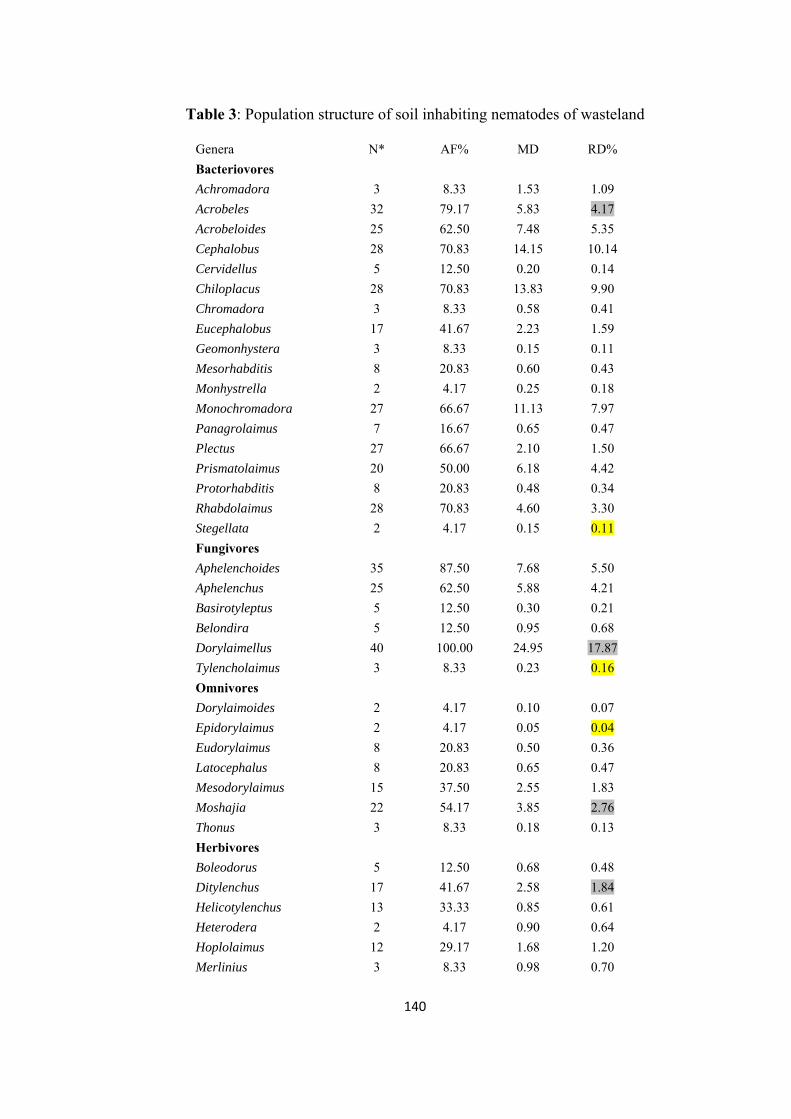
Table 3: Population structure of soil inhabiting nematodes of wasteland
Genera N* AF% MD RD%
Bacteriovores
Achromadora 3 8.33 1.53 1.09
Acrobeles 32 79.17 5.83 4.17
Acrobeloides 25 62.50 7.48 5.35
Cephalobus 28 70.83 14.15 10.14
Cervidellus 5 12.50 0.20 0.14
Chiloplacus 28 70.83 13.83 9.90
Chromadora 3 8.33 0.58 0.41
Eucephalobus 17 41.67 2.23 1.59
Geomonhystera 3 8.33 0.15 0.11
Mesorhabditis 8 20.83 0.60 0.43
Monhystrella 2 4.17 0.25 0.18
Monochromadora 27 66.67 11.13 7.97
Panagrolaimus 7 16.67 0.65 0.47
Plectus 27 66.67 2.10 1.50
Prismatolaimus 20 50.00 6.18 4.42
Protorhabditis 8 20.83 0.48 0.34
Rhabdolaimus 28 70.83 4.60 3.30
Stegellata 2 4.17 0.15 0.11
Fungivores
Aphelenchoides 35 87.50 7.68 5.50
Aphelenchus 25 62.50 5.88 4.21
Basirotyleptus 5 12.50 0.30 0.21
Belondira 5 12.50 0.95 0.68
Dorylaimellus 40 100.00 24.95 17.87
Tylencholaimus 3 8.33 0.23 0.16
Omnivores
Dorylaimoides 2 4.17 0.10 0.07
Epidorylaimus 2 4.17 0.05 0.04
Eudorylaimus 8 20.83 0.50 0.36
Latocephalus 8 20.83 0.65 0.47
Mesodorylaimus 15 37.50 2.55 1.83
Moshajia 22 54.17 3.85 2.76
Thonus 3 8.33 0.18 0.13
Herbivores
Boleodorus 5 12.50 0.68 0.48
Ditylenchus 17 41.67 2.58 1.84
Helicotylenchus 13 33.33 0.85 0.61
Heterodera 2 4.17 0.90 0.64
Hoplolaimus 12 29.17 1.68 1.20
Merlinius 3 8.33 0.98 0.70
140
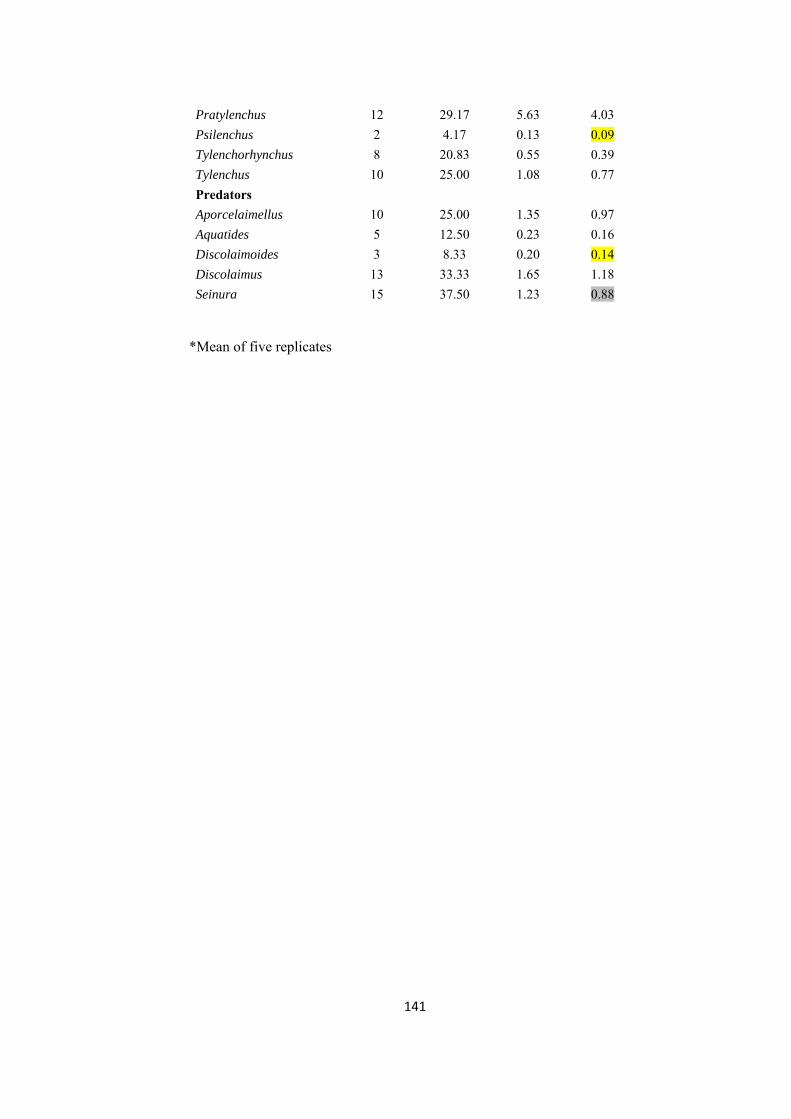
Pratylenchus 12 29.17 5.63 4.03
Psilenchus 2 4.17 0.13 0.09
Tylenchorhynchus 8 20.83 0.55 0.39
Tylenchus 10 25.00 1.08 0.77
Predators
Aporcelaimellus 10 25.00 1.35 0.97
Aquatides 5 12.50 0.23 0.16
Discolaimoides 3 8.33 0.20 0.14
Discolaimus 13 33.33 1.65 1.18
Seinura 15 37.50 1.23 0.88
*Mean of five replicates
141

Tab
le 4
: Cha
ract
eris
tics o
f cro
p fie
ld
Ju
ne
Sept
embe
rO
ctob
er
Dec
embe
r Ja
nuar
y Fe
brua
ry
Mar
ch
Apr
il M
ean±
SD
H'
2.24
2.
21
2.63
2.
33
2.73
2.
53
2.34
2.
56
2.45
±0.1
9
MI
2.32
2.
31
2.38
2.
10
2.50
2.
12
2.49
2.
59
2.35
±0.1
8
MI2
5 2.
33
2.42
2.
50
2.13
2.
58
2.16
2.
50
2.59
2.
4±0.
18
PPI
3.19
3.
00
2.83
2.
45
2.56
2.
21
3.00
2.
60
2.73
±0.3
3
PPI/M
I 1.
38
1.30
1.
19
1.16
1.
03
1.04
1.
21
1.01
1.
16±0
.13
EI
26.7
5 26
.19
41.1
4 13
.51
38.2
4 29
.33
15.1
9 22
.04
26.5
±9.8
SI
43.6
2 50
.77
56.5
6 22
.70
60.4
7 25
.59
57.4
4 64
.25
47.7
±15.
8
BI
46.9
2 41
.01
33.9
0 68
.91
31.4
7 56
.11
39.5
6 32
.41
43.8
±13.
08
TDI
1.26
1.
05
1.04
1.
06
1.12
1.
06
1.05
1.
10
1.1±
0.07
NC
R
0.74
0.
81
0.83
0.
94
0.61
0.
74
0.76
0.
64
0.76
±0.1
1
% C
epha
lobi
ds
32.2
6 58
.63
51.1
3 45
.28
36.3
6 37
.06
46.3
2 33
.02
42.5
±9.4
% T
ylen
chid
s 39
.78
13.4
0 13
.15
24.6
7 22
.64
18.9
5 8.
93
13.6
2 19
.4±9
.8
% D
oryl
aim
s 6.
89
13.2
7 12
.33
1.94
19
.51
5.83
17
.00
16.9
2 11
.7±6
.2
% O
ther
s 21
.07
14.6
9 23
.39
28.1
1 21
.48
38.1
6 27
.74
36.4
4 26
.4±8
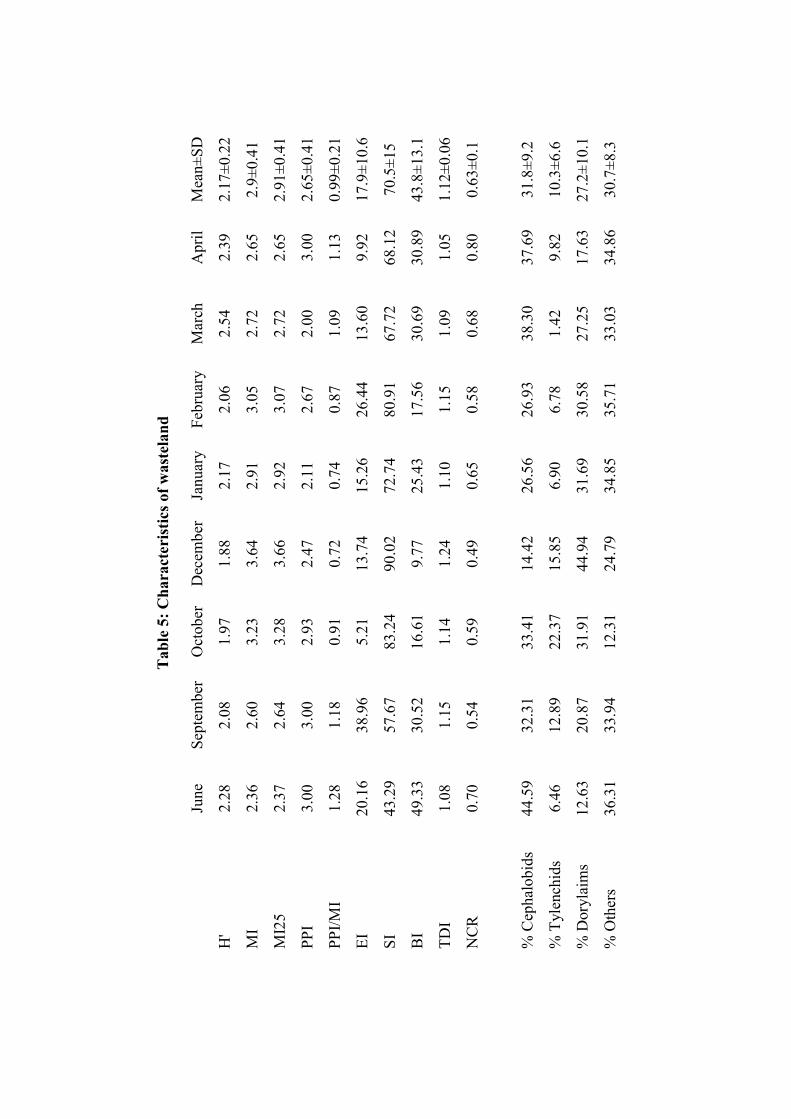
Tab
le 5
: Cha
ract
eris
tics o
f was
tela
nd
Ju
ne
Sept
embe
r O
ctob
er
Dec
embe
rJa
nuar
y Fe
brua
ry
Mar
ch
Apr
il M
ean±
SD
H'
2.28
2.
08
1.97
1.
88
2.17
2.
06
2.54
2.
39
2.17
±0.2
2
MI
2.36
2.
60
3.23
3.
64
2.91
3.
05
2.72
2.
65
2.9±
0.41
MI2
5 2.
37
2.64
3.
28
3.66
2.
92
3.07
2.
72
2.65
2.
91±0
.41
PPI
3.00
3.
00
2.93
2.
47
2.11
2.
67
2.00
3.
00
2.65
±0.4
1
PPI/M
I 1.
28
1.18
0.
91
0.72
0.
74
0.87
1.
09
1.13
0.
99±0
.21
EI
20.1
6 38
.96
5.21
13
.74
15.2
6 26
.44
13.6
0 9.
92
17.9
±10.
6
SI
43.2
9 57
.67
83.2
4 90
.02
72.7
4 80
.91
67.7
2 68
.12
70.5
±15
BI
49.3
3 30
.52
16.6
1 9.
77
25.4
3 17
.56
30.6
9 30
.89
43.8
±13.
1
TDI
1.08
1.
15
1.14
1.
24
1.10
1.
15
1.09
1.
05
1.12
±0.0
6
NC
R
0.70
0.
54
0.59
0.
49
0.65
0.
58
0.68
0.
80
0.63
±0.1
% C
epha
lobi
ds
44.5
9 32
.31
33.4
1 14
.42
26.5
6 26
.93
38.3
0 37
.69
31.8
±9.2
% T
ylen
chid
s 6.
46
12.8
9 22
.37
15.8
5 6.
90
6.78
1.
42
9.82
10
.3±6
.6
% D
oryl
aim
s 12
.63
20.8
7 31
.91
44.9
4 31
.69
30.5
8 27
.25
17.6
3 27
.2±1
0.1
% O
ther
s 36
.31
33.9
4 12
.31
24.7
9 34
.85
35.7
1 33
.03
34.8
6 30
.7±8
.3
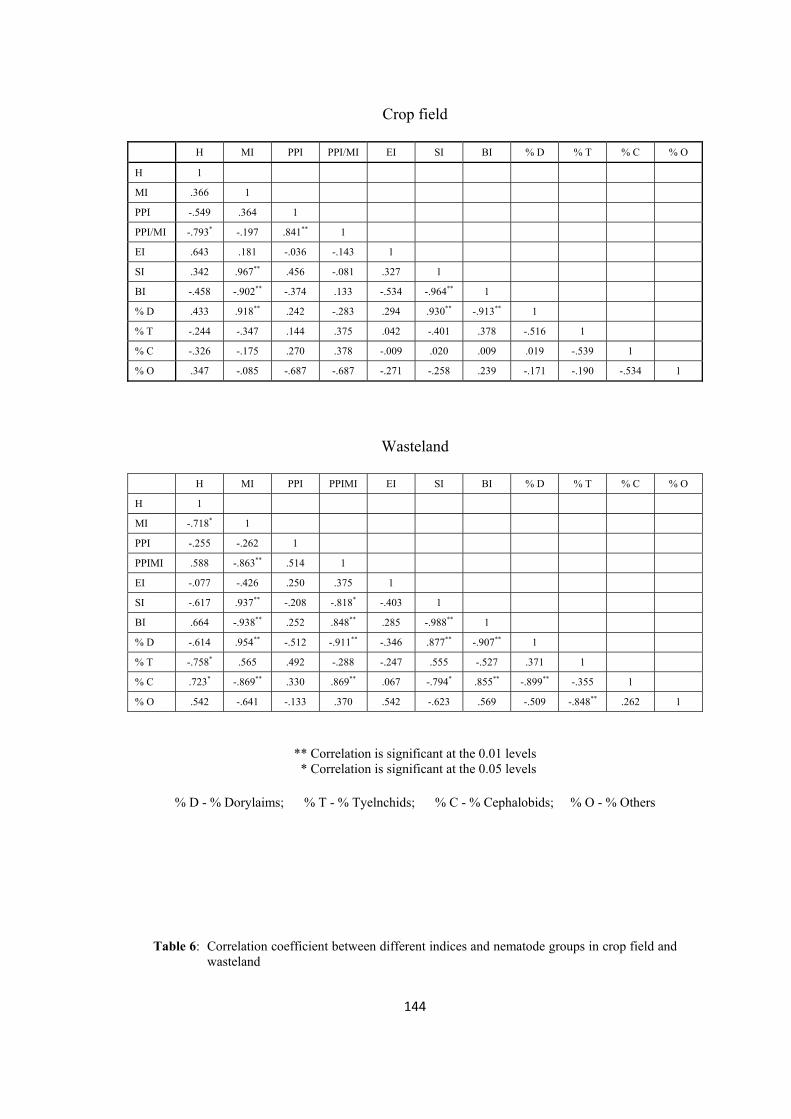
Crop field
H MI PPI PPI/MI EI SI BI % D % T % C % O
H 1
MI .366 1
PPI -.549 .364 1
PPI/MI -.793* -.197 .841** 1
EI .643 .181 -.036 -.143 1
SI .342 .967** .456 -.081 .327 1
BI -.458 -.902** -.374 .133 -.534 -.964** 1
% D .433 .918** .242 -.283 .294 .930** -.913** 1
% T -.244 -.347 .144 .375 .042 -.401 .378 -.516 1
% C -.326 -.175 .270 .378 -.009 .020 .009 .019 -.539 1
% O .347 -.085 -.687 -.687 -.271 -.258 .239 -.171 -.190 -.534 1
Wasteland
H MI PPI PPIMI EI SI BI % D % T % C % O
H 1
MI -.718* 1
PPI -.255 -.262 1
PPIMI .588 -.863** .514 1
EI -.077 -.426 .250 .375 1
SI -.617 .937** -.208 -.818* -.403 1
BI .664 -.938** .252 .848** .285 -.988** 1
% D -.614 .954** -.512 -.911** -.346 .877** -.907** 1
% T -.758* .565 .492 -.288 -.247 .555 -.527 .371 1
% C .723* -.869** .330 .869** .067 -.794* .855** -.899** -.355 1
% O .542 -.641 -.133 .370 .542 -.623 .569 -.509 -.848** .262 1
** Correlation is significant at the 0.01 levels * Correlation is significant at the 0.05 levels
% D - % Dorylaims; % T - % Tyelnchids; % C - % Cephalobids; % O - % Others
Table 6: Correlation coefficient between different indices and nematode groups in crop field and wasteland
144
Fig.1: Ordinal diversity and abundance of nematodes in a Crop field (A & B) and wasteland (C & D)
145
A
B
C
D
Fig. 2: Trophic diversity and abundance of nematodes in a crop field (A & B) and wasteland (C & D)
146
C
D
B
A
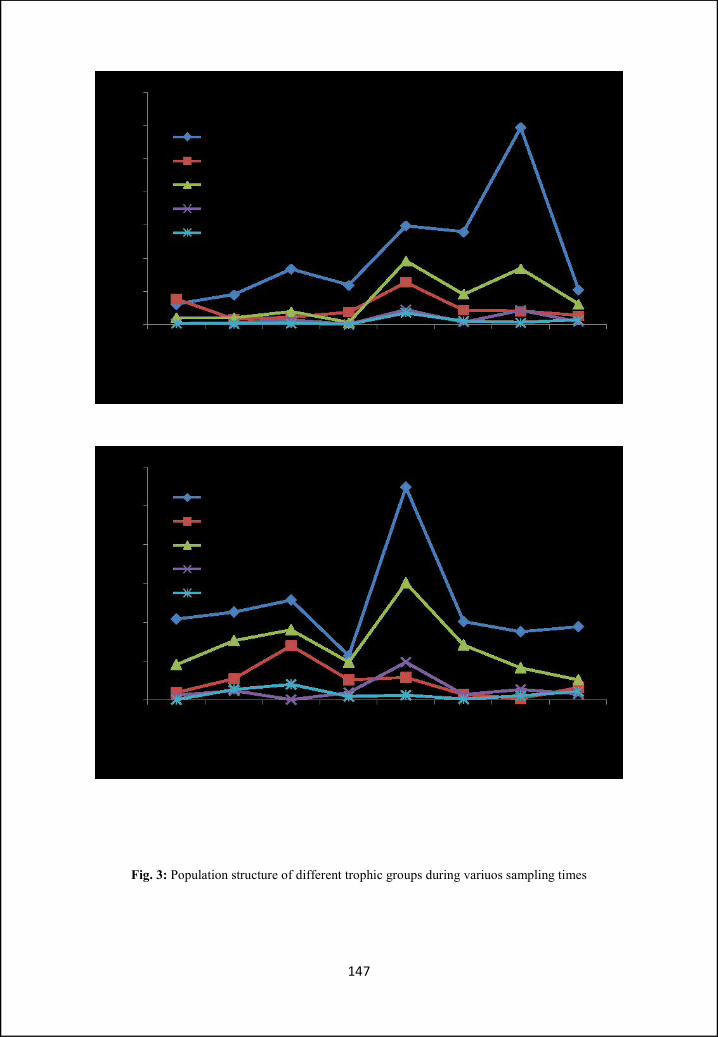
Fig. 3: Population structure of different trophic groups during variuos sampling times
147
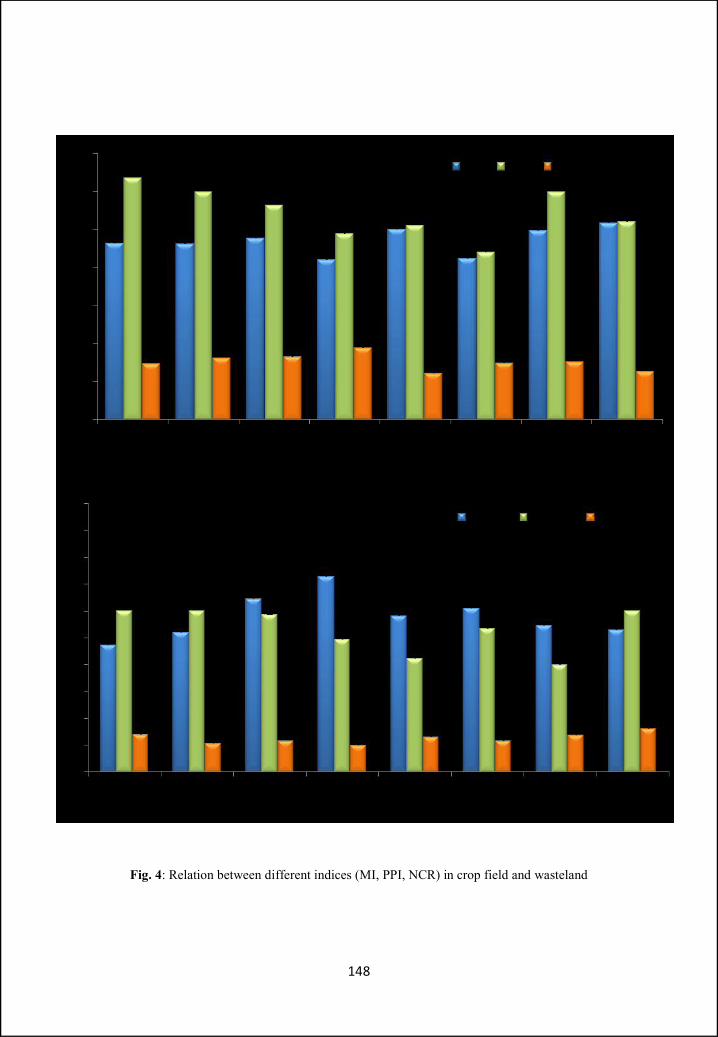
Fig. 4: Relation between different indices (MI, PPI, NCR) in crop field and wasteland
148

Fig. 5: Relation between different indices (EI, SI, BI) in crop field and wasteland
149

Fig. 6: Population structure of different nematode groups during various sampling times
150
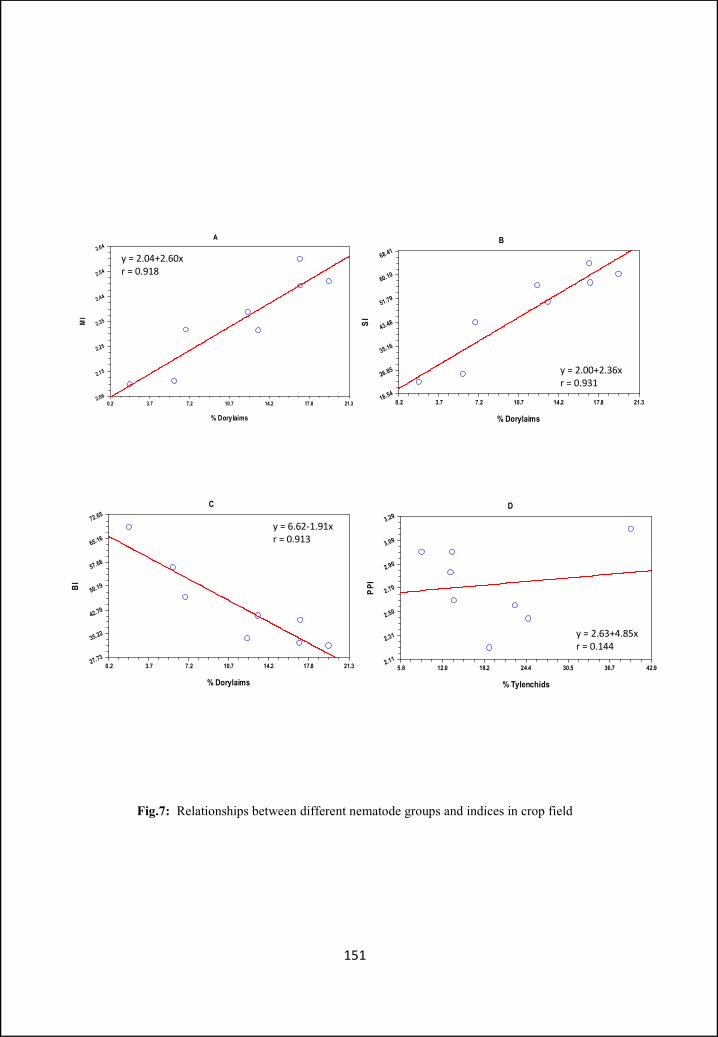
Fig.7: Relationships between different nematode groups and indices in crop field
151
A
% Dorylaims
MI
0.2 3.7 7.2 10.7 14.2 17.8 21.32.06
2.15
2.25
2.35
2.44
2.54
2.64B
% Dorylaims
SI
0.2 3.7 7.2 10.7 14.2 17.8 21.318.54
26.85
35.16
43.48
51.79
60.10
68.41
D
% Tylenchids
PPI
5.8 12.0 18.2 24.4 30.5 36.7 42.92.11
2.31
2.50
2.70
2.90
3.09
3.29
C
% Dorylaims
BI
0.2 3.7 7.2 10.7 14.2 17.8 21.327.73
35.22
42.70
50.19
57.68
65.16
72.65
y = 2.04+2.60xr = 0.918
y = 2.00+2.36xr = 0.931
y = 6.62-1.91xr = 0.913
y = 2.63+4.85xr = 0.144
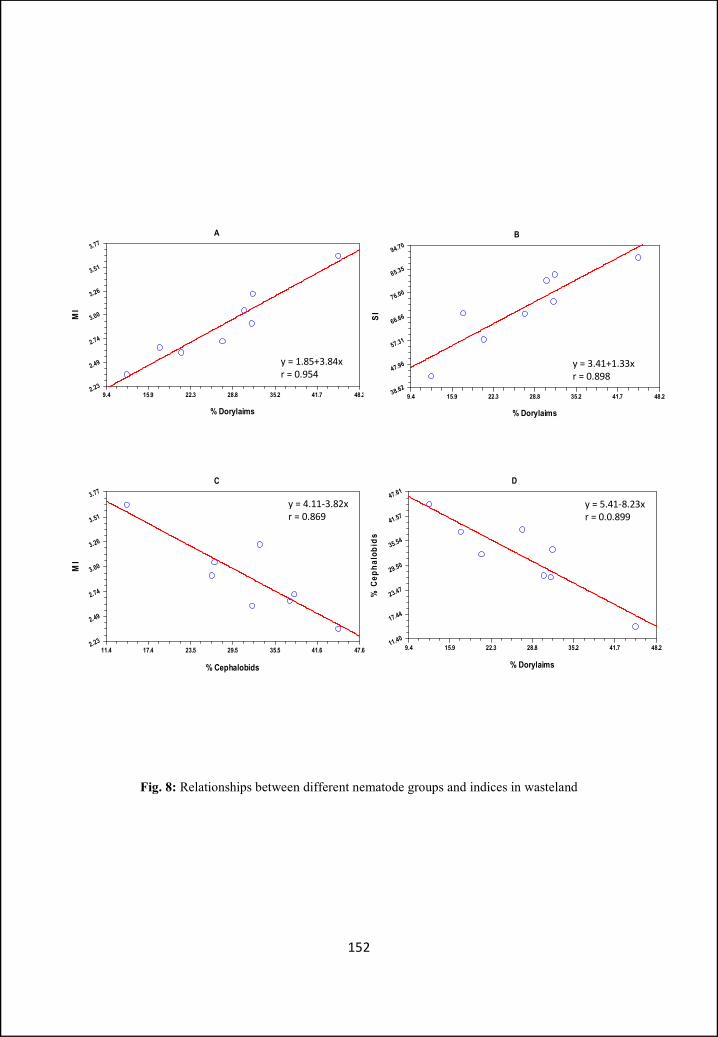
A
% Dorylaims
MI
9.4 15.9 22.3 28.8 35.2 41.7 48.22.23
2.49
2.74
3.00
3.26
3.51
3.77B
% Dorylaims
SI
9.4 15.9 22.3 28.8 35.2 41.7 48.238.62
47.96
57.31
66.66
76.00
85.35
94.70
D
% Dorylaims
% C
epha
lobi
ds
9.4 15.9 22.3 28.8 35.2 41.7 48.211.40
17.44
23.47
29.50
35.54
41.57
47.61C
% Cephalobids
MI
11.4 17.4 23.5 29.5 35.5 41.6 47.62.23
2.49
2.74
3.00
3.26
3.51
3.77
y = 4.11-3.82xr = 0.869
y = 5.41-8.23xr = 0.0.899
y = 3.41+1.33xr = 0.898
y = 1.85+3.84xr = 0.954
Fig. 8: Relationships between different nematode groups and indices in wasteland
152

Discussion

Soil fauna have strong relationships with fundamental ecosystem
processes such as decomposition and nutrient cycling, and interactions with the
microbial community, plant growth and pedogenesis and thus can serve as useful
indicators of ecosystem conditions (Parmellee, 1994; Auerswald et al., 1996;
Reddy et al., 1996; Anderson & Sparling, 1997). The nematode diversity of agro-
ecosystems offers many possibilities for use as biological indicators of
agricultural practices, soil characteristics and the degree of conservation of soils.
Whole nematode populations, represented by the structure of functional groups
probably represent, in many cases, better indicators than those derived from any
species in particular (Yeates & Bongers, 1999).
Soil nematodes have been found to regulate the bacterial and fungal
populations and thus are intimately associated with the cycling of major nutrients
in soils (Ingham et al., 1985) and a more positive view of the role of nematodes in
soil processes has been adopted (Yeates, 1987). There are thus apparently
significant possibilities for the use of nematode populations and diversity as
indicators of overall soil condition. Nematode communities are sensitive to
chemical and physical disturbances in ecosystem. These disturbances can alter
nematode communities in qualitatively different ways (Fiscus & Neher, 2002). A
higher percentage of dorylaims in the wasteland (27%) as compared to the crop
field (12%) clearly indicates that the wasteland soil is less disturbed as may be
expected. Cropping on the other hand always involves ploughing and/or tilling
together with addition of fertilizers, organic matter and pesticides/weedicides.
The dorylaims appear to be susceptible to these activities as also shown by
Thomas (1978) and Sohlenius & Wasilewska (1984). Hence, the sensitivity of the
153

dorylaims is a good indicator of soil disturbance (Neher, 2001).
In this study Acrobeloides was the most abundant genus in crop field and
confirms the work of Yeates & Bird (1994) and Gomes et al. (2002) where it was
found that cephalobids were the most abundant bacterial feeders present in
cropping systems.
Agro-ecosystems are generally characterized by periodic disturbances and
tillage, cultivation, use of pesticides and fertilizers impede natural succession.
Although each of these disturbances have specific effects but mainly results in a
decrease of diversity (Yeates & Bongers, 1999). Ploughing stimulates
mineralization and results in increase in the number and dominance of
opportunistic taxa. Pesticides influence soil biota directly or indirectly via plants.
It is generally accepted that undisturbed systems have more diverse communities
of soil organisms (Kandji et al., 2001). Plots with more ground cover vegetation
often support the most diverse assemblages of nematodes, possibly as a result of
the greater heterogeneity of resources added through the return of residues and
root exudates (Ou et al., 2005).
Bacteriovores in the crop fields showed two phases of population increase.
A small increase during December-January and a big increase during February-
March. One factor that may be responsible for the increase in the population of
these nematodes in the early part is farmyard manure that was added to the fields
before sowing. However the greater increase in the later (post-harvest) part of the
study may also be due to organic matter. This becomes available because of
decaying roots and rotting stubble after the harvest and could be the likely cause
of this second peak. Organic amendments are known to increase the population of
154

the Ba1 group of nematodes and maintain them till the material is exhausted
(Bongers & Ferris, 1999; Porazinska et al., 1999). Manuring is also known to
bring about a sudden increase in the population of bacterial feeders (Dmowska &
Kozlowska, 1988). The sharp decline of bacteriovorous nematodes during March-
April may also be due the organic matter; although this time acting perhaps
differently. Depleted organic matter, of the decaying roots and stubble, could
result in depleted bacterial populations and hence a reduced food source for the
bacteriovores resulting in their decline. Bulluck et al., (2002) have shown that
when food supply is exhausted bacteriovores populations also decline. Although
temperature records were not maintained, there is a relatively big increase in
maximum temperatures from February to April, and it may not be improper to
speculate that this could lead to drying of the top soil and also a decline in
nematode populations.
The herbivore population of the crop field varied little during the period of
study except for a small peak in January. This coincides with the maturing of the
crops and as such represents a period of an abundant food source brought about
by the proliferated root system and seems to be the most likely cause of the peak.
Increase nutrient availability is known to increase herbivore populations through
an enhanced food resource (Pattinson et al., 2004). Fertilizer application after
sowing in October did not seem to affect nematodes as all trophic group
populations remained relatively unchanged till December. Further, the peak of
the herbivores coincided with peak or rising populations of the other groups. This
contrasts with findings of Yeates (1982), Sohlenius & Bostrom (1986), Edwards
(1989) and Hoyvönen & Hühta (1989) who observed an increase in the plant
155

parasites and bacterial feeders after fertilizer application in cultivated soil or a
decrease of fungal feeders and omnivores (Sohlenius & Wasilewska, 1984;
Sohlenius & Bostrom, 1986; Sohlenius, 1990).
In the wasteland, populations fluctuated little over most part of the year
except from December to February when a small decline was followed by a peak
in January in the case of bacteriovores, fungivores and omnivores. Predator
populations remained at low levels throughout the year while the herbivores
showed a peak in October. The increase of herbivores seems to be related to the
grasses that grow in the wastelands after the rains. As winter sets in, this
vegetation begins to wither and die and enriches the soil with organic matter that
may be responsible for the peak populations of the bacteriovores, fungivores and
the omnivores. In grasslands used by grazing animals, high populations of
bacterial feeders were attributed to high level of food resource made available by
addition of organic matter in the form of dung (Freckman et al., 1979).
Shannon’s diversity index (H’) reflects diversity of nematodes in an
ecosystem. Higher values of H’ show highly diverse ecosystem while low values
show the contrary. Háněl (1995) found H’ in crop fields to vary between 2.66-
2.83. In present work, the value of H’ was higher in crop field as compared to
wastelands. This is in perfect agreement to earlier records where crop fields are
found to be highly diverse in comparison to other ecosystems (Ferris et al., 2001,
Tomar et al., 2006). Nitrogenous fertilizers increases or decreases nematode
abundance, as a consequence of the biomass production of plant root systems or
increase in microbial activity (Berger et al., 1986; Sohlenius & Boström, 1986;
Sohlenius, 1990). Nematodes in crop field had higher generic and individual
156

abundance as compared to the nematodes in wasteland which is evident from the
study of Shannon’s diversity index. The lower H′ values for wasteland which
contrasts with the idea of Kandji et al. (2001) in all likelihood was because of
lack of diversity in the food resources particularly of the herbivores.
The maturity index (MI) provides useful information on the direction of
change within a particular habit. MI values for soil subjected to varying levels of
disturbances range from less than 2.0 in nutrient enriched disturbed systems to
±4.0 in undisturbed, pristine environments (Bongers & Ferris, 1999). Agricultural
practices, such as incorporating organic material (manure) into the soil stimulate
microbial activity and provide resources for opportunistic nematode species,
consequently, there is a rapid decrease in the MI followed by a gradual increase
during subsequent succession (Bongers & Ferris, 1999). In the present study MI
ranges from 2.1-2.59 in crop field and 2.36-3.66 in wasteland and is consistent
with earlier observations (Háněl, 1994, 1998; Bongers, 2001; Tomar & Ahmad,
2009, Dechang et al., 2009). Higher MI values for wasteland could be expected as
in undisturbed soil the ratio of higher functional guild species (dorylaims) to low
functional guild species (rhabditidae, panagrolaimidae etc.) is much greater than
in the crop field (27.2 - 31.8 vs 11.7 - 42.5). The highest value of MI in the crop
field was observed in April, several weeks after harvesting. One key factor that
may be responsible is the rapid decline of bacteriovores and fungivores
accompanied by little or no change in the omnivore and predator populations. The
MI values for wasteland indicate it to be a more stable habitat, presumably
because it is free from human intervention and with limited grazing. Here also the
maximum MI value of 3.66 is most probably brought about by a decline in the
157

bacteriovore and fungivore population rather than in increase in the omnivore –
predator population.
The PPI is very good indicator of plant parasitic nematode resources. Pate
et al. (2000) estimated PPI values for crop fields at 2.3 while Neher & Campbell
(1994) recorded PPI as 2.82 and 2.51 in soybean plantations. The PPI values for
present study in crop field agree with earlier records. The lower PPI values for
wasteland is probably because of limited food resource and a lack of its variety.
The vegetation and the nematode population may have attained a state of
equilibrium as can be expected in undisturbed areas. On the other hand, cropping
provides varied and alternate sources of food to the plant parasites. PPI/MI is also
very good indicator of enrichment (Bonger’s et al., 1997). In present work low
PPI/MI values for wasteland and high values for crop field indicates that more
mature soil are less enriched as compared to manipulated soils. This is also borne
out by the greater EI in cultivated soil as compared to the undisturbed wasteland.
Similar observations have also been made by Háněl (2004) and Tomar & Ahmad
(2009). The Nematode Channel Ratio (NCR) indicates a dominance of bacterial
decomposition in both habitats but to varying degrees. Bacterial decomposition in
the wasteland is of smaller order as compared to the crop field. However this
contrasts with the observations of Bardgett and McAlister (1999) who argued that
fungal pathways of decomposition dominate in natural ecosystem.
Food web indices like EI, SI, and BI may provide an excellent means for
studying the stability of ecosystem, whether it is stressed, enriched or structured.
And provide information on the dynamics of the soil food web (Ferris et al.,
2001). Nematodes and other soil biota play an important role in releasing
158

nutrients from bacterial biomass for the uptake by the plant roots. These
nematodes that respond first are enrichment opportunist and their biomass uptake
continues as long as the bacterial activity is high (Bongers & Bongers, 1998).
The values for SI in crop field during present study were lower as
compared to wastelands. The increase followed by a somewhat constant SI
probably indicates a structured soil in a state of equilibrium where nematode
populations fluctuate depending on the biotic and abiotic factors but not
significantly affecting one another. It has been reported in earlier studies that
generally in fallow soil and woodland the values of SI are higher which may due
to high abundance of omnivores and predators suggesting a food web with more
trophic linkages (Ferris & Matute, 2003). The higher the values of SI the more
complex is the community structure. Fallow lands and forests have been reported
to be more complex communities with reference to nematode in many studies
(Tomar & Ahmad, 2009). The values of SI for crop field fluctuated more
erratically and seemed to be governed more by the population of omnivores,
predators and possibly herbivores than by bacteriovores and fungivores. EI values
were higher for crop field and lower in wastelands. The values for EI were lowest
in October and highest being during September in wasteland, while for crop field
the highest value of EI was recorded during October while the lowest value was
observed at December, which incidently is the reverse for wasteland. Low values
of EI reflect low abundance of Rhabditidae and high abundance of Cephalobidae,
which can be well explained as the population of cephalobids is high compared to
rhabditids in October for wasteland and December for crop field. EI is generally
known to reflect availability of resources to the soil food web and response of
159

primary decomposers to the resources (Ferris et al., 2004). The basal index (BI)
which is also a measure of enrichment show no significant changes over the
sampling time, except for the month of December in crop field and June in
wastelands.
Both in crop field and wasteland population of dorylaims show positive
correlation with MI and SI (P<0.01). As dorylaims have cp values in range of 4-5
and their abundance plays major role in higher values of these indices. The
incorporation of higher functional guilds for calculation of these indices results in
high degree of positive correlation. BI shows a high degree of negative correlation
(P<0.01) with dorylaims in both crop field and wasteland. This clearly indicates
the inverse relationship between dorylaimid and lower c-p value nematode guilds.
Cephalobids also show negative correlation with MI in wasteland (P<0.01) and
crop field (NS). An unusual fact of almost no correlation between PPI and
Tylechids in both areas is perplexing and is strikingly different from the
significant correlation reported by Háněl (1998) and Tomar & Ahmad (2009).
It may be concluded that nematode faunal analysis provides a good tool
for diagnosis of the complexity and status of soil food webs (Ritz and Trudgill,
1999). The presence and abundance of specific taxa is an indicator of the
complexity of the food web. Since related taxa, with similar morphological,
anatomical and physiological attributed, have similar feeding habits, useful faunal
analyses can be obtained by identification (Bongers & Ferris, 1999). As reported
earlier the present study also confirms that nematodes are very good indicators of
soil conditions. Although some contradictions have been found mainly the
absence of correlation between PPI and tylenchids is highly surprising as most
160

studies in other parts of worlds as listed earlier, show significant correlation
between these parameters. Decomposition pathways were predominantly bacterial
in both the cropfield as well as wasteland, while crop field observations confirm
to the studies of earlier workers, the wasteland observation is in contrast to this.
These contradictions are difficult to explain as various abiotic factors have not
been taken into account, which can be a matter for further studies. Some factors
like anthropogenic activities resulting in the loss of top soil, can have profound
effect on the structure of the soil food web, which may severely limit its capacity
to support optimal nutrient cycling, plant growth and other enriched functions. It
is important to understand that indices developed from nematode faunal analysis
are based on proportions of the fauna in various functional guilds. They provide
an indication of the relative proportions of services and functions, but not of their
magnitude. The biomass and abundance of organisms in various functional guilds
is important in determining the magnitude of services. If biomass is also taken
into account we can have better understanding of the resources available to soil
food web organisms. The use of food web indices give better understanding of
wastelands where human intervention and other activities are very limited
throughout the year, but in case of crop field many factors play role in defining
the condition of soil and population dynamics of nematodes, therefore a
generalized study is not possible for food webs of crop fields.
161

References

ABEBE, E., MEKETE, T. & THOMAS, W.K. (2011). A critique of current methods in nematode taxonomy. African Journal of Biotechnology 10: 312–323.
ABOLAFIA, J. & PEÑA-SANTIAGO, R. (2002). Nematodos del orden Rhabditida de Andalucía Oriental. El género Eucephalobus Steiner, 1936. Graellsia 58: 59–78.
ABOLAFIA, J. & PEÑA-SANTIAGO, R. (2003a). Nematodes of the order Rhabditida from Andalucía Oriental, Spain. The genus Acrobeloides (Cobb, 1924) Thorne, 1937, with description of A. arenicola sp. n. and a key to species. Journal of Nematode Morphology and Systematics 5: 107–130.
ABOLAFIA, J. & PEÑA-SANTIAGO, R. (2003b). Nematodes of the order Rhabditida from Andalucía Oriental, Spain. The genus Chiloplacus Thorne, 1937, with description of C. maginensis sp. n. and a key to species. Nematology 5: 243–274.
ABOLAFIA, J. & PEÑA-SANTIAGO, R. (2003c). Nematodes of the order Rhabditida from Andalucía Oriental, Spain. The genera Nothacrobeles Allen & Noffsinger, 1971 and Zeldia Thorne, 1925. Journal of Nematology 35: 233–243.
ABOLAFIA, J. & PEÑA-SANTIAGO, R. (2003d). Nematodes of the order Rhabditida from Andalucía Oriental, Spain. The genus Stegelletina Andrássy, 1984, with description of two new species. Russian Journal of Nematology 11: 37–53.
ABOLAFIA, J. & PEÑA -SANTIAGO, R. (2006). Nematodes of the order Rhabditida from Andalucia Oriental, Spain. The family Panagrolaimidae, with a compendium of species of Panagrolaimus and a key to their identification. Journal of Nematode Morphology and Systematics 8: 133–160.
ABOLAFIA, J. & PEÑA-SANTIAGO, R. (2009a). Nematodes of the order Rhabditida from Andalucia Oriental, Spain. The genus Ablechroiulus Andrássy, 1966 with description of three new species and key to species. Nematology 11: 77–94.
ABOLAFIA, J., PEÑA-SANTIAGO, R. (2009b). Nematodes of the order Rhabditida from Andalucía Oriental, Spain. The genus Cephalobus Bastian, 1865 with description of C. harpagonis sp. n. and key to species. Nematology 11: 485–508.
162

ADAMS, B. (1998). Species concepts and the evolutionary paradigm in modern nematology. Journal of Nematology 30: 1–21.
ADAMS, B. (2002). The species delimitation uncertainty principle. Journal of Nematology 33: 153–160.
AHMAD, I., SHAH, A.A. & MAHAMOOD, MD. (2007). Nematodes of the Order Rhabditida from India. Description of Sclerorhabditis tridentatus gen. n., sp.n. (Rhabditidae). Nematology 9: 43–47.
AHMAD, I., SHAH, A.A. & MAHAMOOD, MD. (2010). Nematodes of the order Rhabditida from India. Description of a new species of Mesorhabditis (Rhabditidae) and comments on M. cranganorensis. International Journal of Nematology 20: 63–68.
AHMAD, W. (1996): Plant parasitic nematodes of India: An identification Manual. Aligarh Muslim University, Aligarh, India: Department of Zoology, 347 pp.
AKHTAR, M. (1993). Utilisation of plant–origin waste materials for the control of parasitic nematodes. Bioresource Technology 46: 255–257.
AKHTAR, M. (1999). Plant growth and nematode dynamics in response to soil amendments with neem–products, urea and compost. Bioresource Technology 69: 181–183.
AKHTAR, M. (2000). Effect of organic and urea amendments in soil on nematode communities and plant growth. Soil Biology & Biochemistry 32: 573–575.
ALEXANDER, M. (1977). Introduction to soil microbiology. 2nd ed. New York, Wiley, 467p.
ALI, S.M., SURYAWANSHI, M.V. & CHISTY, Z. (1973). Two new species of Drilocephalobus Coomans & Goodey, 1965 (Nematoda : Drilocephalobidae n. fam.) from Marathwada, India, with a revised classification of the superfamily Cephaloboidea (Paramonov, 1956) Paramonov, 1962. Nematologica 19: 308–317.
ALLEN, M.W. & NOFFSINGER, E.M. (1971). Nothacrobeles n. gen., with description of four new species (Nematoda: Cephalobidae). Journal of Nematology 3: 145–153.
ALLEN, M.W. & NOFFSINGER, E.M. (1972). A review of the genus Zeldia Thorne, 1937 (Nematoda: Cephalobidae) with descriptions of seven new
163

species. Proceedings of the Helminthological Society of Washington 39: 206–223.
ALLISON, F. E. (1973). Organic farming. In Allison F. E. (ed.), Developments in soil science. III. Soil organic matter and its role in crop production. Amsterdam: Elsevier Science Publishing. pp. 558–582.
AMANN, R.I., LUDWIG, W. & SCHLEIFER, K.H. (1995). Phylogenetic identification and in-situ detection of individual microbial cells without cultivation. Microbiological Reviews 59: 143–169.
ANDERSON, A.N. & SPARLING, G.P. (1997). Ants as indicators of restoration success: microbial biomass in the Australian seasonal tropics. Restoration Ecology 5: 109–114.
ANDERSON, R. V. (1965). Acrobeloides uberrinus n. sp., with a note on morphologic variation within soil and bacteria-reared populations. Proceedings of the Helminthological Society of Washington 32: 232–235.
ANDERSON, R. V. (1968). Variation in taxonomic characters of a species of Acrobeloides (Cobb, 1924) Steiner and Buhrer, 1933. Canadian Journal of Zoology 46: 309–320.
ANDERSON, R. V., GOULD, W. D., WOODS, L. E., CAMBARDELLA, C., INGHAM, R. E. & COLEMAN, D. C. (1983). Organic and inorganic nitrogenous losses by microbivorous nematodes in soil. Oikos 40: 75–80.
ANDERSON, R. V. & HOOPER, D.J. (1970). A neotype for Cephalobus persegnis Bastian, 1865, redescription of the species,and observations on variability in taxonomic characters. Canadian Journal of Zoology 48: 457–469.
ANDRÁSSY, I. (1952). Freilebende Nematoden aus dem Bükk-Gebirge. Annales Historico-naturelles Musei Nationalis Hungarici 2: 13– 65.
ANDRÁSSY, I. (1958). Erd- und süßwassernematoden aus Bulgarien. Acta Zoologica Academiae Scientiarum Hungaricae 4: 1–88.
ANDRÁSSY, I. (1958). Szabadonélö fonálférgek Nematoda libera. Magyarország állatvilága fauna Hungariae 3: 1–362.
ANDRÁSSY, I. (1967). Die unterfamilie Cephalobinae (Nemaoda. Cephalobidae) und ihre Arten. Acta Zoologica Hungarica 13: 1–37.
164

ANDRÁSSY, I. (1968). Fauna Paraguayensis. 2. Nematoden aus den Galeriewäldern des Acaray–Flusses. Opuscula Zoologica Budapest 8: 167–315.
ANDRÁSSY, I. (1974). Über vier homonyme Nematodengattungen. Nematologica 19: 403–404.
ANDRÁSSY, I. (1974). A Nematodák evolúciója és rendszerezése. (The evolution and systematization of Nematoda). Magyar Tudományos Akadémia Biológiai Osztálya Közlem 17: 13–58.
ANDRÁSSY, I. (1976). Evolution as a basis for the systematization of nematodes. Pitman Publishing , Budapest, London, 288 p.
ANDRÁSSY, I. (1984). Klasse Nematoda (Ordungen Monhysterida, Desmoscolecida, Araeolaimida, Chromadorida, Rhabditida). Gustav Fischer Verlag. Stuttgart, 509 p.
ANDRÁSSY, I. (2005). Free-living nematodes of Hungary, I. (Nematoda, errantia). In. Csuzdi, Cs & Mahunka, S. (Eds.). Pedozoologica Hungarica, 3. Hungarian Natural History Museum & Systematic Zoology Research Group of the Hungarian Academy of Sciences, Budapest, pp. 518.
ANDRE, H.M., DUCARME, X. & LEBRUN, P. (2002). Soil biodiversity: myth, reality or conning? Oikos 96: 3–24.
ANDRÉN, O. & LAGERLÖF, J. (1983). Soil fauna (microarthropods, enchytraeids, nematodes) in Swedish agricultural cropping systems. Acta Agriculturae Scandinavica 33: 33–52.
AUERSWALD, K.,WEIGAND, S., KAINZ, M. & PHILIPP, C. (1996). Influence of soil properties on the population and activity of geophagous earthworms after five years of bare fallow. Biology and Fertility of Soils 23: 382–387.
AYYAR P.N.K. (1926). A preliminary note on the root gall nematode, Heterodera radicicola and its economic importance in South India. The Madras Agricultural Journal, 14: 113–118.
AYYAR P.N.K. (1933). Some experiments on the control of root–gall nematode (Heterodera radicicola Greeff) in South India. The Madras Agricultural Journal 21: 97–107.
BALDWIN, J.G., DE LEY, I.T., MUNDO-OCAMPO, M., DE LEY, P., NADLER, S.A. &
GEBRE, M. (2001). Acromoldavicus mojavicus n sp (Nematoda:
165

Cephaloboidea) from the Mojave Desert, California. Nematology 3: 343–353.
BALDWIN, J.G., GIBLIN DAVIS, R.M., EDDLEMAN, C.D., WILLIAMS, D.S., VIDA, J.T. & THOMAS, W.K. (1997). The buccal capsule of Aduncospiculum halicti (Nemata: Diplogasterina): an ultrastructural and molecular phylogenetic study. Canadian Journal of Zoology 75: 407–423.
BANIYAMUDDIN, MD., TOMAR, V. V. S. & AHMAD, W. (2007). Functional diversity of soil inhabiting nematodes in natural forests of Arunachal Pradesh, India. Nematologia Mediterranea 35: 109–121.
BARBER, C.A. (1901). A tea-eelworm disease in South India. Bulletin of the Department of Land Use and Agriculture, Madras 2: 227–234.
BARDGETT, R.D. & MC ALISTER, E. (1999). The measurement of soil fungal. Bacterial biomass ratios as an indicator of ecosystem self–regulation in temperate meadow grasslands. Biology and Fertility of soils 29: 282–290.
BASTIAN, H.C. (1865). Monograph on the Anguillulidae, or free Nematoids, marine, land and freshwater; with descriptions of 100 new species. Transactions of the Linnaean Society of London 25: 73–184.
BERGER, H., FOISSNER, W. & ADAM, H. (1986). Field experiments on the effects of fertilizers and lime on the soil microfauna of an alpine pasture. Pedobiologia 29: 261–272.
BIEDERMAN, L. A. & BOUTTON, T. W. (2010). Spatial variation in biodiversity and trophic structure of soil nematode communities in a subtropical savanna parkland: Responses to woody plant encroachment. Applied Soil Ecology 46: 168–176.
BIRD, A. F., DE LEY, P. & BIRD, J. (1994) Morphology, oviposition and embryogenesis in an Australian population of Acrobeloides nanus. Journal of Nematology 25: 607– 615.
BLAXTER, M.L., DE LEY, P., GAREY, J.R., LIU, L.X., SCHELDEMAN, P., VIERSTRAETE, A., VANFLETEREN, J.R., MACKEY, L.Y., DORRIS, M., FRISSE, L.M., VIDA, J.T. & THOMAS, W.K. (1998). A molecular evolutionary framework for the phylum Nematoda. Nature 392: 71–75.
BOAG, B. & YEATES, G.W. (1998). Soil nematode biodiversity in terrestrial ecosystems. Biodiversity and Conservation 7: 617–630.
166

BOHLEN, P. J. & EDWARDS, C. A. (1994). The response of nematode trophic groups to organic and inorganic nutrient inputs in agroecosystems. In J. W. Doran, D. C. Coleman, D. F. Bezdicek, and B. A. Stewart (eds.). Defining soil quality for a sustainable environment. Madison, WI: Agronomy Society of America. pp. 235–244.
BONGERS, T. (1990). The maturity index: An ecological measure of an environmental disturbance based on nematode species composition. Oecologia 83: 14–19.
BONGERS, T., ALKEMADE, R. & YEATES, G.W. (1991). Interpretation of disturbance–induced maturity decrease in marine nematode assemblages by means of the Maturity Index. Marine Ecology Progress Series 76: 135–142.
BONGERS, T. & BONGERS, M. (1998). Functional diversity of nematodes. Applied Soil Ecology 10: 239–251.
BONGERS, T., DE GOEDE, R.G.M., KORTHALS, G. & YEATES, G.W. (1995). Proposed changes of c–p classification for nematodes. Russian Journal of Nematology 3: 61–62.
BONGERS, T. & FERRIS, H. (1999). Nematode community structure as a bioindicator in environmental monitoring. Trends in Ecology and Evolution. 14: 224–228.
BONGERS, T., VAN DER MEULEN, H. & KORTHALS, G. (1997). Inverse relationship between nematode maturity index and plant parasitic index under enriched conditions. Applied Soil Ecology 6: 195–199.
BONKOWSKI, M. (2004). Protozoa and plant growth: the microbial loop in soil revisited. New Phytologist, 162: 617–631.
BORELLUS, P. (1656). 'Observatiouum Micioscopicarum Cenluria,' Hagoe coniil.is.384, Paris.
BOSTRÖM, S. (1984a). A scanning electron microscope study of three species of Eucephalobus Steiner, 1936 (Nematoda: Cephalobidae). Nematologica 30: 131–139.
BOSTRÖM, S. (1984b). Description and morphological variability of Chiloplacus minimus (Thorne, 1925) Andrássy, 1959 (Nematoda: Cephalobidae). Nematologica 30: 151– 160.
167

BOSTRÖM, S. (1985a). A new species of Zeldia Thorne (Nematoda: Cephalobidae) from Tunisia. Nematologia Mediterranea 13: 67–71.
BOSTRÖM, S. (1985b). Acrobeles oosiensis sp. n. (Nematoda: Cephalobidae) from Tunisia. Nematologia Mediterranea 13: 123–126.
BOSTRÖM, S. (1985c). Aspects on the ultrastructure of the cephalic region in two species of Acrobelinae (Nematoda: Cephalobidae). Acta Zoologica 66: 211– 216.
BOSTRÖM, S. (1985d). Description of Acrobeloides emarginatus (de Man, 1880) Thorne, 1937 and proposal of Acrolobus n. gen. (Nematoda: Cephalobidae). Revue de Nématologie 8: 335–340.
BOSTRÖM, S. (1986). Descriptions of Cervidellus serratus (Thorne, 1925) Thorne, 1937 from Sweden and C. neftasiensis n.sp. from Tunisia, with a note on some recently proposed genera (Nematoda: Cephalobidae). Nematologica 31: 365– 374.
BOSTRÖM, S. (1986). Evolution of lip structure in the Cephalobidae (Rhabditida). Revue de Nématologie 9: 290.
BOSTRÖM, S. (1987). A new terrestrial nematode species (Rhabditida: Cephalobidae) from Spitzbergen. Polar Biology 7: 375–378.
BOSTRÖM, S. (1988a). Morphological and Systematic studies of the family Cephalobidae (Nematoda: Rhabtida). Thesis, University of Stolkholm, Sweden 34 p.
BOSTRÖM, S. (1988b). A scanning electron microscope study of some species of terrestrial nematodes from Spitzbergen. Nematologica 33: 366–374.
BOSTRÖM, S. (1989). Descriptions and morphological variability of three populations of Panagrolaimus Fuchs, 1930 (Nematoda: Panagrolaimidae). Nematologica 34: 144–155.
BOSTRÖM, S. (1990). Some species of Cephalobidae (Nematoda: Rhabditida) from highland Kenya. Journal of African Zoology 104: 127–134.
BOSTRÖM, S. (1991). Some Cephalobidae (Nematoda: Rhabditida) from sand on the Island of Samos, Greece. Nematologica 37: 367–375.
BOSTRÖM, S. (1992). Some Cephalobidae (Nematoda: Rhabditida) from Crete, Greece. Fundamental and Applied Nematology 15: 289– 295.
168

BOSTRÖM, S. (1993a). Some cephalobids from Ireland and Malaysia (Nematoda: Rhabditida). Afro Asian journal of Nematology 3: 128– 134.
BOSTRÖM, S. (1993b). Some cephalobids from Turkey (Nematoda. Rhabditida). Nematologia Mediterranea, 21: 295–300.
BOSTRÖM, S. (1998). A scanning electron microscope study of some species of terrestrial nematodes from Spitzbergen. Nematologica 33: 366–374.
BOSTRÖM, S. (2000). A divergent population of Cervidellus capraeolus (De Ley, Geraert & Coomans, 1990) Boström & De Ley, 1996 (Nematoda: Cephalobidae) from the Bahamas. Journal of Nematode Morphology and Systematics 3: 95–99.
BOSTRÖM, S. & DE LEY, P. (1996). Redescription of Cervidellus vexilliger (de Man, 1880) Thorne, 1937 (Nematoda: Cephalobidae) and taxonomical consequences. Fundamental and Applied Nematology 19(4): 329– 340.
BRADFORD, M.A., TORDOFF, G.M., BLACK, H.I.J., COOK, R., EGGERS, T., GARNETT, M.H., GRAYSTON, S.J., HUTCHESON, K.A., INESON, P., NEWINGTON, J.E., OSTLE, N., SLEEP, D., STOTT, A. & HEFIN JONES, T. (2007). Carbon dynamics in a model grassland with functionally different soil communities. Functional Ecology 21: 690–697.
BRZESKI, M.W. (1960). Cephalobus (Heterocephalobus) kaezanowskii subgen. nov., sp. nov. (Nematoda: Cephalobidae). Bulletin de l’Académie Polonaise des Sciences. ClassII, Série des Sciences Biologique 8: 153–165.
BULLUCK, L.R. III, BARKER, K.R & RISTAINO, J.B. (2002), Influences of organic and synthetic soil fertility amendments on nematode trophic groups and community dynamics under tomatoes. Applied Soil Ecology 21: 233–250.
BUTLER, E.J. (1906). The wilt disease of pigeonpea and pepper. Agricultural Journal of India 1: 25–36.
BUTLER, E.J. (1913) Disease of rice. Bulletin 34. Agricultural Research Institute, Pusa, India.
BUTLER, E.J. (1919) The rice worm (Tylenchus angustus) and its control. Memoirs of the Department of Agriculture in India, Botanical Series 10: 1–37.
169

CAIRNS, J., MCCORMICK, P. V. & NIEDERLEHNER, B. R. (1993). A proposed framework for developing indicators of ecosystem health. Hydrobiologica 236: 1–44.
CHEN, J. & FERRIS, H. (1999). The effects of nematode grazing on nitrogen mineralization during fungal decomposition of organic matter. Soil Biology and Biochemistry 31: 1265–1279.
CHEN, J. & FERRIS, H. (2000). Growth and nitrogen mineralization of selected fungi and fungal–feeding nematodes on sand amended with organic matter. Plant and Soil 218: 91–101.
CHENG, Z. & GREWAL, P.S. (2009). Dynamics of the soil nematode food web and nutrient pools under tall fescue lawns established on soil matrices resulting from common urban development activities. Applied Soil Ecology 42: 107–117.
CHITWOOD, B.G. (1933). A revised classification of the Nematoda. Journal of Parasitology 20: 131.
CHITWOOD, B.G. (1937). A revised classification of the Nematoda. Papers on Helminthology in Commemoration of the 30-Year Jubileum of K.I. Skrjabin and of the All-Union Institute of Helminthology, Moscow. 69–80.
CHITWOOD, B.G. (1958). The designation of official names for higher taxa of invertebrates. Bulletin of Zoological Nomenclature 15: 660–695.
CLAUSI, M. (1998). Cervidellus vinciguerrae sp. nov., (Nematoda: Cephalobidae) from Tierra del Fuego and notes on the genus Cervidellus Thorne, 1937. Fundamental and Applied Nematology 21: 273– 279.
COBB, N.A. (1918). Estimating the nema population of the soil. U.S. Department of Agriculture. Agricultural Technical Circular of US Department of Agriculture 1: 48p.
COBB, N.A. (1920) One hundred new nemas (type species of 100 new genera). Contributions to a Science of Nematology 9: 217–343.
COBB, N.A. (1924). Amended characterization of the nemic genera Cephalobus and Acrobeles. Journal of Parasitology 11: 108.
COOMANS, A. (2000). Nematode Systematics: past, present and future. Nematology 2: 3–7.
170

COOMANS, A. & GOODEY, J.B. (1965). Drilocephalobus congoensis n. g., n. sp. Nematologica 11: 116–120.
DADAY, E. (1897). Die freilebenden Susswasser-Nematoden Ungarns. Zoologisches Jahrbucher (Systematik) 10: 91–134.
DADAY, E. (1905). Untersuchungen über die Süsserwasser-Mikrofauna Paraguays. Zoologica, Stuttgart 18: 1–349.
DASTUR, J. F. (1936). A nematode disease of rice in the Central Provinces. Proceedings of Indian Academy of Sciences Section B, 4: 108–121.
DECHANG, H., ZHANG, X.K., TOMAR, V.V.S., LI, Q., DAZHONG, W., LIANG, W.J. (2009). Effects of heavy metal pollution of highway origin on soil nematode guilds in North Shenyang, China. Journal of Environmental Sciences 21: 193–198.
DE CONINCK, L. (1965). Structures cephalique and Appareil digestif, cavite buccale. In Grasse P. P. (Ed.). Traite de zoologie. Vol. IV. Masson et Cie, Paris, pp. 127–132,146–162.
DE DEYN, G.B., RAAIJMAKERS, C.E., ZOOMER, H.R., BERG, M.P., DE RUITER, P.C., VERHOEF, H.A., BEZEMER, T.M. & VAN DER PUTTEN, W.H. (2003). Soil invertebrate fauna enhances grassland succession and diversity. Nature 422: 711–713.
DE GOEDE, R. G. M. & DEKKER, H. H. (1993). Effects of liming and fertilization on nematode communities in coniferous forest soils. Pedobiologia 37: 193–209.
DE GOEDE, R.G.M., GEORGIEVA, B.C., VERSCHOOR, B.C. & KAMERMAN, J. (1993). Changes in nematode community structure in a primary succession of blown-out areas in a drift sand landscape. Fund. Appl. Nematol., 16: 501–513.
DE LEY, P. (1997). The current state of affairs in identification and diagnosis of the genera of the family Cephalobidae (Nematoda: Rhabditida). Mededelingen Faculteit Landbouwkundige en Toegepaste Biologische Wetenschappen Universiteit Gent 62: 657–673.
DE LEY, P. & BLAXTER, M. L. (2002). Systematic position and phylogeny. In: Lee, D. L. (Ed.), The Biology of Nematodes. Taylor and Francis, London, pp.1–30.
171

DE LEY, P. & BLAXTER, M. L. (2004) A new system for Nematoda: combining morphological characters with molecular trees, and translating clades into ranks and taxa. Nematology Monographs and Perspectives 2: 633–653.
DE LEY, P. & COOMANS, A. (1990). Drilocephalobus moldavicus Lisetskaya, 1968 from Senegal: an odd nematode adds to its reputation under the scanning electron microscope (Nematoda: Rhabditida). Revue de Nematologie 13: 37–43.
DE LEY, P., COOMANS, A. & GERAERT, E. (1990a). Metacrobeles tessellatus sp. n., second species of a rare genus (Nematoda: Rhabditida). Nematologica 35: 25–36.
DE LEY, P., FELIX, M.A., FRISSE, L.M., NADLER, S.A., STERNBERG, P.W. &
THOMAS, W.K. (1999). Molecular and morphological characterisation of two reproductively isolated species with mirror–image anatomy (Nematoda: Cephalobidae). Nematology 1: 591–612.
DE LEY, P., GERAERT, E. & COOMANS, A. (1990b). Seven cephalobids from Senegal (Nematoda: Rhabditida). Journal of African Zoology 104: 287–304.
DE LEY, P., HERNANDEZ, M.A. & AGUDO, J. (1994). A redescription of Ypsylonellus similis (Thorne, 1925) Andrássy, 1984, with descriptions of some related species (Nematoda: Cephalobidae). Fundamental and Applied Nematology 17: 1–16.
DE LEY, P., & SIDDIQI, M.R. (1991). Description of Pseudacrobeles macrocystis sp. n., with some new observations on the morphology of Cephalobidae (Nematoda). Afro–Asian Journal of Nematology 1: 31– 40.
DE LEY, P., SIDDIQI, M.R. & BOSTRÖM, S. (1993a). A revision of the genus Pseudacrobeles Steiner, 1938 (Nematoda: Cephalobidae). Part 1. Subgenus Pseudacrobeles grad. n. Fundamental and Applied Nematology 16: 219–238.
DE LEY, P., SIDDIQI, M.R. & BOSTRÖM, S. (1993b). A revision of the genus Pseudacrobeles Steiner, 1938 (Nematoda: Cephalobidae). Part 2. Subgenus Bunobus subg. n. problematical species, discussion and key. Fundamental and Applied Nematology 16: 289–308.
DE LEY, P., & VANDEBROEK, I. (1992). Stegelletina laticollaris n. sp., a second new species of the genus from Senegal (Nematoda: Cephalobidae). Belgian Journal of Zoology 122: 139–146.
172

DE LEY, P., VAN DE VELDE, M. C., MOUNPORT, D., BAUJARD, P. & COOMANS, A. (1995). Ultrastructure of the stoma in Cephalobidae, Panagrolaimidae and Rhabditidae, with a proposal for a revised stoma terminology in Rhabditida (Nematoda). Nematologica 41: 153–182.
DE MAN, J.G. (1876). Onderzoekingen over vrij in de aarde levende Nematoden. Tijdschrift der Nederlansche Dierkundige Vereeniging 2: 78–196.
DE MAN, J.G. (1880). Die einheimischen, frei in der reinen Erde und im süssen Wasser lebenden Nmatoden. Vorläufiger Bericht und descriptivsystematischer Theil. Tijdschrift der Nederlansche Dierkundige Vereeniging 5: 1–104.
DE MAN, J.G. (1884). Die frei in der reinen Erde und im süssen Wasser lebenden Nematoden der niederländis chen Fauna. Eine systematisch-faunistische Monographie, Leiden, 1–206.
DE MAN, J.G. (1895). Description of three species of Anguillulidae observed in diseased pseudo-bulbs of tropical orchids. Proceedings and Transactions of the Liverpool Biological Society 9: 76–94.
DE MAN, J.G. (1912). Odontopharynx longicaudata n. g., n. sp., eine neueForm von Anguilluliden. Zoologische Jahrbücher (Systematik) 33: 637–642.
DE MAN, J.G. (1920). Description d´une espèce nouvelle du genre Diplogaster Max Schultze: Diplogaster consobrinus, nov. sp. Annales de la Societé Royale Zoologique et Malacologique de Belgique 51: 47–54.
DE MAN, J.G. (1927). Das Männchen der Rhabditis monhystera Bütschli. Zoologischer Anzeiger 70: 51–57.
DE RUITER, P.C., NUETEL, A.M. & MOORE, J. (2005). The balance between productivity and food web structure in soil ecosystems. In: Bardgett, R.D., Usher, M.B., Hopkins, D.W. (Eds.), Biological Diversity and Function in Soils. Cambridge University Press, Cambridge, U.K.
DEVI, G. & DAS, P. (1998). Effect of different organic amendments for the management of root–knot nematode Meloidogyne incognita on carrot. Indian Journal of Nematology 28: 203–207.
DMOWSKA, E. & KOZLOWSKA, J. (1988). Communities of nematode treated with semi–liquid manure. Pedobiologia 32: 323–330.
173

DOLINSKI, C., BALDWIN, J.G. & THOMAS, W.K. (2001). Comparative survey of early embryogenesis of Secernentea (Nematoda), with phylogenetic implications. Canadian Journal of Zoology 79: 82–94.
DOUGHERTY, E.C. (1955). The genera and species of the subfamily Rhabditina Micoletzky, 1922 (Nematoda). A nomenclatorial analysis- including an addendum on the composition of the family Rhabditidae Örley, 1880. Journal of Helminthology 29: 105–152.
DUJARDIN, F. (1845). Histoire naturelle des helminthes ou vers intestinaux. Paris. 654 p.
EBERTH, C.I. (1863). Untersuchungen uber Nematoden. W. Engelman, Leipzig.
EDWARDS, C.A. (1989). Impact of herbicides on soil ecosystems. Critical Review in Plant Sciences 8: 221–257.
EITMINAVIĔŪTÉ, I., BAGDANAVIČIONĖ, Z., KADYTE, B., LAZAUSKIENĖ, L., SUKACKIENĖ, I. (1976). Characteristic successions of microorganisms and soil invertebrates in the decomposition process of straw and lupine. Pedobiologia 16: 106–115.
ETTEMA, C. & BONGERS, T. (1993). Characterization of nematode colonization and succession in disturbed soil using the maturity index. Biology and Fertility of Soils 16: 79–85.
FÉLIX, M.A., DE LEY, P., SOMMER, R.J, FRISSE, L.M., NADLER, S.A., THOMAS, W.K., VANFLETEREN, J.R., & STERNBERG, P.W. (2000). Evolution of vulva development in the Cephalobina (Nematoda). Developmental Biology 221: 68–86.
FERRIS, H., BONGERS, T. & DE GOEDE, R.G.M. (2001). A framework for soil food web diagnostics: extension of nematode faunal analysis concept. Applied Soil Ecology 18: 13–29.
FERRIS, H., BONGERS, T. & DE GOEDE, R.G.M. (2004). Nematode faunal analysis to assess food web enrichment and connectance. In Cook R.C., and Hunt D.J., (eds) Proceedings of the Fourth International Congress of Nematology. Nematology Monographs and Perspectives 2. Brill, The Netherlands, pp. 503–510
FERRIS, H. & MATUTE, M.M. (2003). Structural and functional succession in the nematode fauna of a soil food web. Applied Soil Ecology 23: 93–110.
174

FERRIS, H., VENETTE, R.C. & LAU, S.S. (1996). Dynamics of nematode communities in tomatoes grown in conventional and organic farming systems, and their impact on soil fertility. Applied Soil Ecology 3: 161–175.
FERRIS, H., VENETTE, R.C. & LAU, S.S. (1997). Population energetics of bacterial–feeding nematodes: carbon and nitrogen budgets. Soil Biology and Biochemistry 29: 1183–1194.
FERRIS, H., VENETTE, R.C. & SCOW, K.M. (2004). Soil management to enhance bacterivore and fungivore nematode populations and their nitrogen mineralisation function. Applied Soil Ecology 25: 19–35.
FILIPJEV, I. (1934). The classification of the free-living nematodes and their relation to the parasitic nematodes. Smithsonian Miscellaneous Collections (Washington) 89: 1–63.
FISCUS, D.A. & NEHER, D.A. (2002). Distinguishing sensitivity of free-living soil nematode genera to physical and chemical disturbances. Ecological Applications 12: 565–575.
FRECKMAN, D.W. (1988). Bacterivorous nematodes and organic–matter decomposition. Agriculture, Ecosystems and Environment 24: 195–217.
FRECKMAN, D. W., DUNCAN, D. A. & LARSON, J. R. (1979). Nematode density and biomass in an annual grassland ecosystem. J. Range Manage. 32: 418–421.
FRECKMAN, D.W. & ETTEMA, C.H. (1993). Assessing nematode communities in agroecosystem of varying human intervention. Agriculture, Ecosystem and Environment 45: 239–261.
FRECKMAN, D.W. & MANKAU, R. (1986). Abundance, distribution, biomass and energetics of soil nematodes in a northern Mojave desert ecosystem. Pedobiologia 29: 129–142.
FRECKMAN, D.W. & VIRGINIA, R.A. (1997). Low-diversity Antarctic soil nematode communities: distribution and response to disturbance. Ecology 72: 363–369.
FUCHS, A.G. (1930). Neue an Borken-und Rüsselkäfer gebundene Nematoden, halbparasitische und Wohnungseinmieter. Zoologische Jahrbücher (Systematik) 59: 505–646.
175

GOLDSTEIN, B., FRISSE, L.M. & THOMAS, W.K. (1998). Embryonic axis specification in nematodes evolution of the first step in development. Current Biology 8: 157–160.
GOMES, G.S., HUANG, S.P. & CARES, J.E. (2003). Nematode community, trophic structure and population fluctuation in Soybean fields. Fitopatology 28: 258–266.
GOODEY, T. (1963). Soil and freshwater nematodes (Revised by Goodey, J.B.). 2nd edition. London: Methuen, 544p.
GRIFFITHS, B. S., RITZ, K. & WHEATLEY, R. E. (1994). Nematodes as indicators of enhanced microbiological activity in a Scottish organic farming system. Soil Use and Management 10: 20–24.
GRIFFITHS, R.P., ENTRY, J.A., INGHAM, E.R. & EMMINGHAM, W.H. (1997). Chemistry and microbial activity of forest and pasture riparian–zone soils along three Pacific Northwest streams. Plant and Soil 190: 169–178.
HAMMOND, P.M. (1992). Species inventory. In: Global Diversity, Status of the Earth’s Living Resources. Chapman & Hall, London, pp. 17–39.
HÁNĚL, L. (1998). Distribution of nematodes in soil, mycorrhizài s6$k mycorrhizae and roots of spruce forests at the Boubin Mount, Czech Republic. Biologia 53: 593–603.
HÁNĚL, L. (2004). Colonization of chemical factory wastes by soil nematodes. Pedobiologia 48: 373–381.
HÁNĚL, L. (2010). An outline of soil nematode succession on abandoned fields in South Bohemia. Applied Soil Ecology. 46: 355–371.
HASSAN, J., CHISHTI, M.Z., RASHEED, M. & AHMAD, F. (2008). Nematodes Associated with Brassica compestris and Their Control. World Applied Sciences Journal 5: 543–545.
HASSAN, J., CHISHTI, M.Z., RASHEED, M., AHMAD, I., AHMAD, F. & LONE, B.A. (2009). Nematodes associated with Zea mays and their control through organic soil amendments. International Journal of Plant Production 3: 71–76.
HEININGER, P., HÖSS, S., CLAUS, E., PELZER, J. & TRAUNSPURGER, W. (2007). Nematode communities in contaminated river sediments. Environmental Pollution 146: 64–76.
176

HEIP, C., WARWICK, R.M., CARR, R.M., HERMAN, P.M.J., HUYS, R., SMOL, N. &
VAN HOLSBEKE, K. (1988). Analysis of community attributes of benthic meiofauna of Frierfjord/Langesundfjord. Marine Ecology Progress Series 46: 171–180.
HEYNS, J. (1962). Elaphonema mirabile n. gen., n. sp. (Rhabditida), a remarkable new nematode from South Africa. Proceedings of the Helminthological Society of Washington 29: 128–130.
HEYNS, J. (1962). Osstella hamata n. gen., n. sp., Zeldia serrata n. sp., and Acrobeles thornei n. sp., three new nematodes from South Africa (Rhabditida: Cephalobidae). Nematologica 8: 301–306.
HEYNS, J. (1968). Paracrobeles laterellus n. gen., n. sp. from South Africa (Nematoda: Cephalobidae). Nematologica 14: 511–514.
HEYNS, J. & SWART, A. (1998). On the morphology of Penjatinema natalense n. gen., n. sp., with proposal of a new subfamily, Penjatinematinae (Nematoda: Cephalobidae). Nematologica 44: 381– 390.
HOLOVACHOV, O. & BOSTRÖM, S. (2006). Panagrolobus vanmegenae, a new genus and species from the Netherlands and Ukraine, with taxonomic revision and two new species of the genus Teratolobus Andrássy, 1968 (Nematoda: Cephalobidae). Nematology 8: 377–395.
HOLOVACHOV, O. & BOSTRÖM, S. (2006). Description of Deleyia gen. n. with a discussion of its phylogenetic relationships to the genera Daubaylia Chitwood & Chitwood, 1934 and Myolaimus Cobb, 1920 (Nematoda: Rhabditida). Nematology 8: 223–233.
HOLOVACHOV, O., BOSTRÖM, S. & SUSULOVSKY, A. (2001). Description of Acroukrainicus sagittiferus gen. et sp. n. (Nematoda: Cephalobidae) from the area of an old sulphur industry in Ukraine. Russian Journal of Nematology 9: 51–56.
HÖSS, S., TRAUNSPURGER, W., SEVERIN, G.W., JUTTNER, I., PFISTER, G. &
SCHRAMM, K.W. (2004). Influence of 4–nonylphenol on the structure of nematode communities in freshwater microcosms. Environmental Toxicology and Chemistry 23: 1268–1275.
HOYVÖNEN, R. & HUHTA, V. (1989). Effects of lime, ash and nitrogen fertilizers on nematode populations in scots pine forest soils. Pedobiologia 33: 129–143.
177

INGHAM, R. E., TROFYMOW, J. A., INGHAM, E. R. & COLEMAN, D. C. (1985). Interactions of bacteria, fungi, and their nematode grazers: Effects on nutrient cycling and plant growth. Ecological Monographs 55: 119–140.
JAIRAJPURI, M. S. & AHMAD, W. (1992): Dorylaimida: Free living, Predacious and plant parasitic nematodes. E. J. Brill, Leiden, The Netherlands, 458 pp.
JAIRAJPURI, M.S., BAQRI, Q.H. & AHMAD, S. (1973). Nematodes of high altitudes in India. II. Studies on the genus Tridontus Khera, 1965 with notes on the synonymy of the genus Syedella Suryawanshi, 1971 (Diplogastridae). Nematologica 19: 69–74.
JAIRAJPURI, M. S. & KHAN, W. U. (1982). Predatory nematodes (Mononchida) with special reference to India. Associated Publishing Co., New Delhi, 131 pp.
JOSHI, P.M. (1972). Pseudocephalobus [Pseudocephalobus] indicus n. gen.; n. sp. (Rhabditida: Cephalobidae) from Marathwada, India. Marathwada University Journal of Science, Section B, (Biology Science) 11: 155–158.
KANDJI, S., CALLISTUS, K., OGOL, P. & ALBRECHT, A, (2001). Diversity of plant parasitic nematodes and their relationships with some soil physicochemical characteristics in improved fallows in western Kenya. Applied Soil Ecology 18: 143–157.
KAREGAR, A., DE LEY, P. & GERAERT, E. (1998). The genera Cervidellus Thorne, 1937 and Stegelletina Andrássy, 1984 from Iran, with description of S. kheirii sp. n. Russian journal of Nematology 6: 23– 36.
KHAN, Z. & KIM., Y.H. (2007). A review on the role of predatory soil nematodes in the biological control of plant parasitic nematodes. Applied Soil Ecology 35: 370–379.
KHERA, S. (1965). Nematodes from the banks of still and running waters, II. Tridontus longicaudatus n. gen., n. sp., subfamily Diplogasterinae Micoletzky, 1922 from India. Nematologica 11: 249–254.
KHERA, S. (1968). Acrobelinema cornis n.g., n.sp., subfamily Acrobelinae Thorne from rhizosphere of millets from India. Indian Journal of Helminthology 19: 159–163.
KHERA, S. (1969). Nematodes from the banks of still and running waters. VI. Rhabditida from sewer. Journal of Helminthology 43: 347–363.
178

KHERA, S. (1970). Nematodes from the banks of still and running waters. IX. Two new genera belonging to subfamily Diplogasterinae Micoletzky from India. Revista Brasileria de Biologia (Rio de Janeiro) 30: 405–409.
KHERA, S. (1971). Nematodes from the banks of still and running waters. XI. Subfamily Rhabditinae. Indian Journal of Nematology 1: 237–243.
LAMBSHEAD, P.J.D. (1993). Recent developments in marine benthic biodiversity research. Oceanis 19: 5–24.
LAMBSHEAD, P.J.D. (2001). Marine nematode biodiversity. In: Chen, Z.X., Chen, S.Y. & Dickson, D.W. (eds). Nematology, advances and perspectives. ACSE–TUP Book Series, San Francisco,USA 636 p.
LAMBSHEAD, P.J.D. (2004). Marine nematode biodiversity. In: Chen, Z.X., Chen, S.Y. and Dickson, D.W. (eds) Nematology Advances and Perspectives. CABI, Wallingford. pp. 438–468.
LAWTON, J.H., BIGNELL, D.E., BOLTON, B., BLOEMERS, G.F., EGGLETON, P., HAMMOND, P.M., HODDA, M., HOLT, R.D., LARSEN, T.B., MAWDSLEY, N.A., STORK, N.E., SRIVASTAVA, D.S. & WATT, A.D. (1998). Biodiversity inventories, indicator taxa and effects of habitat modification in tropical forest. Nature 391: 72–76.
LEUCKART, R. (1849). Zur Kenntnis der Fauna von Island. Erster Beitrag (Wurmer). Archiv Naturgeschichte 15, 148–206.
LIANG W.J., LAVIAN I., & STEINBERGER Y. (1999). Dynamics of nematode community composition in a potato field. Pedobiologia 43: 459–469.
LIANG, W., LI, Q., JIANG, Y. & NEHER, D.A. (2005). Nematode faunal analysis in an aquic brown soil fertilised with slow release urea, Northeast China. Applied Soil Ecology 29: 185–192.
LINSTOW, O. VON (1877). Helminthologische Beobachtungen. Archiv Naturgeschichte 42: 1–18.
LISETSKAYA, L.F. (1968). Novyi vid Drilocephalobus moldavicus n. sp. (Nematoda: Drilonematidae). Parazity Zhivotnykh i Rastenii 3: 94–95.
LIU, Y., LI, X., JIA, R., HUANG, L., ZHOU, Y. & GAO, Y. (2011). Effects of biological soil crusts on soil nematode communities following dune stabilization in the Tengger Desert, Northern China Original Research Article. Applied Soil Ecology 49: 118–124.
179

LOOF, P.A.A. (1962). Metacrobeles togoensis n.gen., sp.n. a remarkable acrobelin nematode from diseased coconut tree. Nematologica 7: 114–118.
MAGGENTI, A. (1963). Comparative morphology in nemic phylogeny. In: Dougherty, E.C. (Ed.). The lower Metazoa, comparative biology and phylogeny. Berkeley, USA. University of California Press, pp. 273–282.
MAGGENTI, A. (1970). System analysis and nematode phylogeny. Journal of Nematology 2: 7–15.
MAHAMOOD, MD. & AHMAD, I. (2009). Nematodes of the order Rhabditida from India. Description of Diplogasteroides uriceus sp. n. and Rhabditidoides papillatus sp. n. (Diplogastrina). Nematology 11: 541–550.
MAHRAN, A., TENUTA, M., LUMACTUD, R.A. & DAAYF, F. (2009). Response of a soil nematode community to liquid hog manure and its acidification. Applied Soil Ecology 43: 75–82.
MALAKHOV, V.V. (1994). Nematodes: Structure, Development, Classification and Phylogeny. Smithsonian Institution Press, Washington. 286 p.
MASSEY, C.L. (1964).The nematode parasites and associates of the fir engraver beetle, Scolytus ventralis LE CONTE in New Mexico. Journal of Insect Pathology 6: 133–155.
MASSEY, C.L. (1974). Biology and taxonomy of nematode parasites and associates of bark beetles in the United States. Agricultural Forestry Service (Washington), Agricultural Handbook, 446: 1–233.
MAUPAS, E.F. (1900). Modes et formea de reproduction des nématodes. Archives de Zoologie Expérimentale et Générale 7: 563–628.
MAY, R.M. (1988). How many species are there on Earth? Science 241, 1441–1449.
MC–SORLEY, R. & FREDERICK, J.J. (2000). Short term effects of cattle grazing on nematode communities in Florida pastures. Nematropica 30: 211–221.
MELDAL, B.H.M., DEBENHAM, N.J., DE LEY, P., DE LEY I.T., VANFLETEREN, J.R., VIERSTRAETE, A.R., BERT, W., BORGONIE, G., MOENS, T., TYLER, P.A., AUSTEN, M.C., BLAXTER, M.L., ROGERS, A.D. & LAMBSHEAD, P.J.D. (2007). An improved molecular phylogeny of the Nematoda with special
180

emphasis on marine taxa. Molecular Phylogenetics and Evolution 42: 622–636.
MICOLETZKY, H. (1922). Die freilebenden Erd-Nematoden mit besonderer Berücksichtigung der steiermark und der Bukowina, Zugleich mit einer revision sämtlicher nichtmariner, freilebender Nematoden in Form von Genus-Beschreibungen und Bestimmungsschlüsse. Archiv für Naturgeschichte Abteilung 87: 1–650.
MOORE, J.C. & DE RUITER, P.C. (1991). Temporal and spatial heterogeneity of trophic interactions within belowground food webs. Agriculture, Ecosystems and Environment, 34: 371–397.
MÜLLER, O. F. (1783). Anmerkungen beim Durchlesen einiger Aufsätze in den 10 ersten Stücken des Naturforschers. Naturforscher, 19: 150–176.
NADLER, S.A. (2002). Species delimitation and nematode biodiversity: phylogenies rule. Nematology 4: 615–625.
NADLER, S.A., CARRENO, R.A., ADAMS, B.J., KINDE, H., BALDWIN, J.G. &
MUNDO-OCAMPO, M. (2003). Molecular phylogenetics and diagnosis of soil and clinical isolates of Halicephalobus gingivalis (Nematoda: Cephalobina: Panagrolaimoidea), an opportunistic pathogen of horses. International Journal of Parasitology 33: 1115–1125.
NADLER, S.A., DE LEY, P., MUNDO-OCAMPO, M., SMYTHE, A.B., STOCK, S.P., BUMBARGER, D., ADAMS, B.J., DE LEY, I.T., HOLOVACHOV, O. &
BALDWIN, J.G. (2006). Phylogeny of Cephalobina (Nematoda): Molecular evidence for recurrent evolution of probolae and incongruence with traditional classifications. Molecular Phylogenetics and Evolution 40: 696–711.
NADLER, S.A. & HUDSPETH, D.S.S. (1998). Ribosomal DNA and phylogeny of the Ascaridoidea (Nemata: Secernentea): Implications for morphological evolution and classification. Molecular Phylogenetics and Evolution 10: 221–236.
NEEDHAM, F. (1743). An account of some new microscopical discoveries. London, 126 p.
NEELY, C. L., BEARE, M. H., HARGROVE, W. L. & COLEMAN, D. C. (1991). Relationships between fungal and bacterial substrate–induced respiration, biomass and plant residue composition. Soil Biology and biochemistry 23: 947–954.
181

NEHER, D. A. (2001). Role of nematodes in soil health and their use as indicators. Journal of Nematology 33: 161–168.
NEHER, D.A., & CAMPBELL, C.L. (1994). Nematode communities and microbial biomass in soils with annual and perennial crops. Applied Soil Ecology 1: 17–28.
NESTEROV, P.I. (1970). Acromoldavicus n. gen. I pereopisanie vida Acrobeloides skrjabini Nesterov et Lisethkaja, 1965 (Nematoda, Cephalobidae). RIO Akademii Nauk Moldavskoi SSR 5: 134–138.
ODUM, E.P. (1985). Trends expected in stressed ecosystems. Bioscience 35 (7): 419–422.
OKADA, H. & FERRIS, H. (2001). Temperature effects on growth and nitrogen mineralization of fungi and fungal–feeding nematodes. Plant and Soil 234: 253–262.
ÖRLEY, L. (1880). Az anguillulidák magànrajza. (Monographie der Anguilluliden). Természetrajzi Füzetek (Budapest) 4: 16–150.
ORR, C.C. & DICKERSON, O.J. (1966). Nematodes in true prairie soils of Kansas. Kansas Academy of Sciences Transcripts 69: 317–334.
OU W, LIANG W, JIANG Y, LI Q, WEN D (2005). Vertical distribution of soil nematodes under different land use types in aquic brown soil. Pedobiology, 49: 139–148.
OVERGAARD NIELSEN, C. (1949). Studies on the soil microfauna. II. The soil inhabiting nematodes. Natura Jutlandica 2: 1–131.
PARK, B.Y., LEE, J.K., RO, H.M. & KIM, Y.H. (2011). Effects of heavy metal contamination from an abandoned mine on nematode community structure as an indicator of soil ecosystem health. Applied Soil Ecology 51: 17–24.
PARMELEE, R.W. (1994). Soil fauna: linking different level of the ecological hierarchy. In: Jones, C.G., Lawton, J.H. (Eds.), Linking Species and Ecosystems. Chapman and Hall, New York, pp. 107–116.
PATE, E., NDIAYE-FAYE, N., THIOULOUSE, J., VILLENAVE, C., BONGERS, T., CADET, P. & DEBOUZIE, D. (2000). Successional trend in characteristics of soil nematode communities in cropped and fallow lands in senegal (Sonkorong). Applied Soil Ecology 14: 5–15.
182

PATTISON, T., BADLOCK, K., ARMOUR, J., PHILMOODY., RASIAH, V., COBBON, J., STEWARD, L., GUILINO, L & LINDA, S. (2004). “Super Soil.” Proceedings of the International soil science conference 5–9 December.
PETERS, B.G. (1927). The Nomenclature of the Vinegar Eelworm. Journal of Helminthology 5: 133–142.
PLATT, H.M. (1994). The phylogenetic systematics of free living nematodes (ant. Seivert Lorenzen). Ray society London. 363 p.
POINAR, G.O. (1993). Origins and phylogenetic relationships of the entomophilic rhabditids, Heterorhabditis and Steinernema. Fundamental and Applied Nematology 16: 333–338.
POLL, J., MARHAN, S., HAASE, S., HALLMANN, J., KANDELER, E. & RUESS, L. (2007). Low amounts of herbivory by root knot nematodes affect microbial community dynamics and carbon allocation in the rhizosphere. FEMS Microbial Ecology 62: 268–279.
PORAZINSKA, D.L., DUNCAN, L.W., MCSORLEY R. & GRAHAM, J.H. (1999). Nematode communities as indicators of status and processes of a soil ecosystem influenced by agricultural management practices. Applied Soil Ecology 13: 69–86.
PORAZINSKA, D.L., MCSORLEY, R., DUNCAN, L.W., GRAHAM, J.H., WHEATON, T.A. & PARSONS, L.R. (1998). Nematode community composition under various irrigation schemes in the citrus soil ecosystem. Journal of Nematology 30: 170–178.
POWERS, T. (2004). Nematode molecular diagnostics: From Bands to Barcodes. Annual Review of Phytopathology 42: 367–383.
POWERS, T., TODD, T., BURNELL, A., MURRAY, P., FLEMING, C., SZALANSKI, A., ADAMS, B. & HARRIS, T. (I997). The rDNA internal transcribed spacer region as a taxonomic marker for nematodes. Journal of Nematology 29: 441–450.
PREJS, K. (1970). Some problems of the ecology of benthic nematodes (Nematoda) of Mikolajskie Lake. Ekologia Polska 25 : 225–242.
RASHID, F. & HEYNS, J. (1990a). Chiloplacus and Macrolaimellus species from South West Africa/Namibia (Nematoda: Cephalobidae). Phytophylactica 22: 189–199.
183

RASHID, F., GERAERT, E., COOMANS, A. & SUATMADJI, W. (1989). Cephalobidae from the Krakatau region (Nematoda: Rhabditida). Nematologica 34: 125–143.
RASHID, F., GERAERT, E. & SHARMA, R.D. (1984). Morphology, taxonomy and morphometry of some Cephalobidae (Nematoda: Rhabditida) from Brazil, with descriptions of two new genera and four new species. Nematologica 30: 251–299.
RATHORE, S.S. & NAMA, H.S. (1992). On two nematodes Acrobeloides conoidis n. sp. and Chiloplacus jodhpurensis n. sp. from soil around roots of Brassica juncea. Proceedings of the National Academy of Science, India (Section B, part 4) 62: 509–513.
REDDY, V.S., REDDY, M.V., LEE, K.K., RAO, K.P.C. & SINIVASAN, S.T. (1996). Response of some soil meso- and macro-faunal populations to soil management during crop and fallow periods on a semi-arid tropical alfisol (India). European Journal of Soil Biology 32: 123–129.
RITZ, K. & TRUDGILL, D.L. (1999). Utility of nematode community analysis as an integrated measure of the functional state of soils: perspectives and challenges. Plant and Soil 212: 1–11.
RUESS, L. (2003). Nematode soil faunal analysis of decomposition pathways in different ecosystems. Nematology 5: 179–181.
RUESS, L. & FERRIS, H. (2002). Decomposition pathways and successional changes. Nematology 4: 159.
RÜHM, W. (1956a). Die Nematoden der Ipiden. Parasitologische Schriftenreihe (Jena). 6: 1–437.
SAMOILOFF, M.R. (1987). Nematodes as Indicators of Toxic Environmental Contaminants. In: Veech, J.A. and Dickson, D.W. (eds) Vistas on Nematology. E.O. Painter, DeLeon Springs,Florida, pp. 433–438.
SCHIERENBERG, E. (2000). Early development of nematode embryos: differences and similarities. Nematology 2: 57–64.
SCHNEIDER, A.F. (1866). Monographie der Nematoden. Berlin, 357 p.
SCHRATZBERGER, M., BOLAM, S., WHOMERSLEY, P. & WARR, K. (2006). Differential response of nematode colonist communities to the intertidal
184

placement of dredged material. Journal of Experimental Marine Biology and Ecology 334: 244–255.
SESHADRI, A.R. (1965). Investigations on the biology and life cycle of Criconemoides xenoplax Raski, 1952 (Nematoda: Criconematidae). Nematologica 10: 540–562.
SHANNON, C.E. & WEAVERS, W. (1949). The mathematical theory of communication. Urbana, IL: University of Illinois.
SIDDIQI, M.R. (1980). The origin and phylogeny of the nematode orders Tylenchida Thorne, 1949 and Aphelenchida, n. ord. Helminthological Abstracts – Series B 49: 143–170.
SIDDIQI, M.R. (1986). Tylenchida: Parasites of plants and insects. Wallingford, UK, CAB International. 645 pp.
SIDDIQI, M.R. (1993). Nematodes of Rain forests: 2. Five new genera and eight new species of cephalobs. Afro-Asian Journal of Nematology 3: 212–225.
SIDDIQI, M.R. (2002). Catoralaimellus cornutus gen. n., sp. n., and two new species of Macrolaimellus (Nematoda: Cephalobidae). International Journal of Nematology 12: 89–97.
SIMPSON, E.H. (1949). Measurement of diversity. Nature 163: 688.
SINGH, R.S. & SITARAMAIAH, K. (1970). Control of plant–parasitic nematodes with organic soil amendments. PANS 16: 287–297.
SOHLENIUS, B. (1973). Structure and dynamics of populations of Rhabditis (Nematodes: Rhabditidae) from forest soil. Pedobiologia 13: 368–375.
SOHLENIUS, B. (1990). Influence of cropping system and nitrogen input on soil fauna and microorganisms in a Swedish arable soil. Biology and Fertility of Soils 9: 168–173.
SOHLENIUS, B. & BOSTRÖM, S. (1986). Short-term dynamics of nematode communities in arable soil. Influence of nitrogen fertilization in barley crops. Pedobiologia 29: 183–191.
SOHLENIUS, B. & WASILEWSKA, L. (1984). Influence of irrigation and fertilization on the nematode community in a Swedish pine forest soil. Journal of Applied Ecology 21: 327–342.
185

SPIRIDONOV, S.E., IVANOVA, E.S. & WILSON, M.J. (2005). The nematodes of the genus Dicelis Dujardin, 1845 parasitic in earthworms: the interrelationships of four Eurasian populations. Russian Journal of Nematology 13: 61–81.
STANDING, D., KNOX, O.G.G., MULLINS, C.E., KILLHAM, K.K. & WILSON, M.J. (2006). Influence of nematodes on resource utilization by bacteria – an in vitro study. Microbial Ecology 52: 444–450.
STEINBUCH, J.G. (1799) Das Grasalchen, Vibrio agrostis. Naturforscher 28: 233–259.
STEINER, G. (1934). Procephalobus mycophilus n. g., n. sp. (Cephalobina), a nematode living in the sclerotia of the fungus Balansia claviceps. Proceedings of the Helminthological Society of Washington 1: 56–58.
STEINER, G. (1936). Opuscula miscellanea nematologica. III. Proceedings of the Helminthological Society of Washington 3: 16–22.
STEINER, G. (1936). Opuscula miscellanea nematologica. IV. Proceedings of the Helminthological Society of Washington 3: 74–80.
STEINER, G. (1938). Opuscula miscellanea nematologica VII. (I). Observations on nematodes associated with Irish potatoes grown in South Carolina. Proceedings of the Helminthological Society of Washington 5: 35–40.
STINNER, B.R. & CROSSLEY, D.A., JR. (1982). Nematodes in no–tillage agroecosystems. In: Freckman, D.W. (ed.) Nematodes in Soil Ecosystems. University of Texas Press, Austin, pp. 14–28.
STOCK, P., CAMPBELL, J.F. & NADLER, S.A. (2001). Phylogeny of Steinernema Travassos, 1927 (Cephalobina: Steinernematidae) inferred from ribosomal DNA sequences and morphological characters. Journal of Parasitology 87: 877–889.
SURYAWANSHI, M.V. (1971). Alirhabditis indica n. gen., n. sp. (Rhabditida: Alirhabditidae n. fam.). Tawdenema indicum n. gen., n. sp. and Syedella aurangabadensis n. gen., n. sp. (Diplogastridae) from Marathwada, India. Nematologica 17: 542–552.
SWARUP, G., NATH, R.P. & SETHI, C.L. (1967). The plant parasitic nematode genus in India. Indian Phytopathology 20: 118–123.
186

SWARUP, G. & SESHADRI, A. R. (1974). Nematology in India-problems and progress. In Raychaudhuri, S. P. & Verma, J. P. (eds.), Current trends in plant pathology, pp. 303–311.
TABARANT, P., VILLENAVE, C., RISÈDE, J–M., ROGER–ESTRADE, J., DOREL, M. (2011). Effects of organic amendments on plant–parasitic nematode populations, root damage, and banana plant growth. Biology and Fertility of Soils 47: 341–347.
TAHSEEN, Q., AHMAD, I. & JAIRAJPURI, M.S. (1999). Observations on three species of the sub family Acrobelinae (Nematoda: Cephalobidae) from India. Nematology 1: 527–537.
TAHSEEN, Q., HUSSAIN, A., TOMAR, V., SHAH, A.A. & JAIRAJPURI, M.S. (2004). Description of Metarhabditis andrassyana gen. n., sp. n. (Nematoda: Rhabditidae) from India. International Journal of Nematology 14: 163–168.
THOMAS, S.H. (1978). Population densities of nematodes under seven tillage regimes. Journal of Nematology 10: 24–27.
THORNE, G. (1925). The genus Acrobeles von Linstow, 1887. Transactions of the American Microscopical Society 44: 171–210.
THORNE, G. (1937). A revision of the nematode family Cephalobidae Chitwood and Chitwood, 1934. Proceedings of the Helminthological Society of Washington 4: 1–16.
THORNE, G. (1938). Notes on free-living and plant–parasitic nematodes. IV. Proceedings of the Helminthological Society of Washington 5: 64–65.
THORNE, G. (1939). Notes on free-living and plant-parasitic nematodes. V. Proceedings of the Helminthological Society of Washington 6: 30–32.
TIMM, R.W. (1956). Marine nematodes from the Bay of Bengal.1. Phasmidea. Journal of Bombay Natural History Society 54: 87–90.
TIMM, R.W. (1960). Brevibucca punctata n. sp. and Macrolaimus natator n. sp., new soil nematodes from East Pakistan. Biologia 6: 252–256.
TIMM, R.W. (1971). Antarctic soil and freshwater nematodes from the McMurdo Sound Region. Proceedings of the Helminthological Society of Washington 38: 42–52.
187

TODD, T.C. (1996). Effects of management practices on nematode community structure in tallgrass prairie. Applied Soil Ecology 3: 235–246.
TOMAR, V. V. S. & AHMAD, W. (2009). Food web diagnostics and functional diversity of soil inhabiting nematodes in a natural woodland. Helminthologia 46: 183–189.
TOMAR, V.V.S., BANIYAMUDDIN, M. & AHMAD, W. (2006). Community structure of soil inhabiting nematodes in a mango orchard at Aligarh, India. International Journal of Nematology 16: 89–101.
TOMAR, V.V.S, ZHANG X. K., LI, Q., JIANG, Y., LIANG, W.J. (2009). Distribution of soil nematode communities along a section of Shen–Ha Highway. Helminthologia 46: 241–246.
TSALOLIKHIN, S.J. (1976). Free–living nematodes as indicators of polluted freshwaters. In: Skarlato, O.A. (ed.) Methods of Biological Analysis of Freshwaters. Akademia Nauk, Leningrad, pp. 118–122.
VAN DE VELDE, M.C., DE LEY P., MOUNPORT, D., BAUJARD, P. & COOMANS, A. (1994). Ultrastructure of the buccal cavity and the cuticle of three Cephalobidae (Nematoda: Rhabditida). Nematologica 40: 541–563.
VAN VEEN, J.A. & KUIKMAN, P.J. (1990). Soil structural aspects of decomposition of organic matter by microbes. Biogeochemistry, 11: 213–233.
VILLOT, A. (1875). Recherches sur les Helminthes libres ou parasites des côtes de la Bretagne. Archives de Zoologie Expérimental et Générale 4: 451–482.
VINCIGUERRA, M.T. (1994). Metacrolobus festonatus gen. n. sp. n. and Scottnema lindsayae Timm, 1971 (Nematoda: Cephalobidae) from Sub Antarctic and Antarctic regions with proposal of the new subfamily Metacrolobinae. Fundamental and applied Nematology 17: 175– 180.
VINCIGUERRA, M.T. & CLAUSI, M. (1996). Two new species of Acrobelophis (Nematoda: Cephalobidae) from Sub Antarctica and notes on the genus. Afro Asian journal of Nematology 6: 104– 109.
WARDLE, D.A., BARDGETT, R.D., KLIRONOMOS, J.N., SETALA, H., VAN DER
PUTTEN, W.H. & WALL, D.H. (2004). Ecological linkages between aboveground and belowground biota. Science 304: 1629–1633.
188

WASILEWSKA, L. (1970). Nematodes of the sand dunes in the Kampinos Forest. I. Species structure. Ekologia Polska 18: 429–443.
WASILEWSKA, L. (1989). Impact of human activities on nematodes. In M. Clarholm and L. Bergstrom, editors. Ecology of arable land. Kluwer Academic, Dordrecht, The Netherlands. pp. 123–132.
WASILEWSKA, L. (1994). The effect of age of meadows on succession and diversity in soil nematode communities. Pedobiologia 38: 1–11.
WASILEWSKA, L. (1998). Changes in the proportions of groups of bacterivorous soil nematodes with different life strategies in relation to environmental conditions. Applied Soil Ecology 9: 215–220.
WASILEWSKA, L., PALIŃSKA, E. & ZIELIŃSKI, J. (1981). The role of nematodes in decomposition of plant material in a rye field. Pedobiologia 21: 182–191.
WEISS, B. & LARINK, O. (1991). Influence of sewage sludge and heavy metals on nematodes in an arable soil. Biology and Fertility of Soils 12: 5–9.
YEATES, G.W. (1979). Soil nematodes in terrestrial ecosystems. Journal of Nematology 11: 213–229.
YEATES, G.W. (1982). Variation of pasture nematode populations over 36 months in summer dry silt loam. Pedobiologia 24: 329–346.
YEATES, G.W. (1987). How plants affect nematodes. Advances in Ecological Research 17: 61–113.
YEATES, G.W. (1994). Modification and quantification of the nematode maturity index. Pedobiologia 38: 97–101.
YEATES, G.W. (1998). Feeding in free–living soil nematodes: a functional approach. In: Perry RN, Wright D.J. (eds.) Physiology and biochemistry of free–living and plant parasitic nematodes. CAB International, Wallingford. pp 245–269.
YEATES, G.W. (1999). Effects of plants on nematode community structure. Annual Review of Phytopathology 37: 127–149.
YEATES, G.W. (2003). Nematodes as soil indicators: functional and biodiversity aspects Biology and Fertility of Soils 37: 199–210.
189

190
YEATES, G.W., BARDGETT, R.D., COOK, R., HOBBS, P.J., BOWLING, P.J. &
POTTER, J.F. (1997). Faunal and microbial diversity in three Welsh grassland soils under conventional and organic management regimes. Journal of Applied Ecology 34: 453–470.
YEATES, G.W. & BIRD, A.F. (1994). Some observations on the influence of the agricultural practices on the nematode faunae of some South Australian soils. Fundamental and Applied Nematology 17: 133–145.
YEATES, G.W & BONGERS, T. (1999). Nematode diversity in agro–ecosystems. Agriculture, Ecosystems and Environment 74: 113–135.
YEATES, G.W, BONGERS, T., DE GOEDE, R.G.M., FRECKMAN, D.W. &
GEORGIEVA, S.S. (1993). Feeding habits in soil nematode families and genera– an outline for soil Ecologists. Journal of Nematology 25: 315–331.
YEATES, G.W. & COLEMAN, D.C. (1982). Role of nematodes in decomposition. In: Freckman, D.W. (ed.) Nematodes in Soil Ecosystems. University of Texas Press, Austin, pp. 55–81.
ZHANG, W.D., WANG, X.F., LI, Q., JIANG, Y. & LIANG, W.J. (2007). Soil nematode responses to heavy metal stress. Helminthologia 44: 87–91.
ZULLINI, A. (1976). Nematodes as indicators of river pollution. Nematologia Mediterranea 4: 13–22.