stress selectively increases fos protein in dopamine neurons innervating the prefrontal cortex
TRANSCRIPT

Stress Selectively Increases FosProtein in Dopamine NeuronsInnervating the Prefrontal Cortex
Ariel Y. Deutch,1 Maggie C. Lee,1 Martha H.Gillham,1 Dorothy A. Cameron,1 Menek Goldstein,2
and Michael J. Iadarola3
1 Department of Psychiatry, Yale University Schoolof Medicine, New Haven, Connecticut 06508 andDepartment of Veteran Affairs Medical Center, WestHaven, Connecticut 06516,2 Department ofPsychiatry, New York University School ofMedicine, New York, New York 10016, and3 Neurobiology and Anesthesiology Branch,National Institute of Dental Research, NationalInstitutes of Health, Bethesda, Maryland 20892
Stress-induced alterations in expression of c-fos protein(Fos) in mesencephalic dopamine (DA) neurons of therat were examined in order to discern which midbrainDA neurons are metabolically activated by stress. Re-straint stress for 30 min increased the number of DAneurons exhibiting Fos-like immunoreactivity in the ven-tral tegmental area (VTA), but not in the substantia nigraor retrorubral field. Stress elicited an increase in thenumber of DA neurons expressing Fos in specific nucleiwithin the VTA. Administration of the anxiogenic /9-car-boline FG 7142 also increased the total number of VTADA neurons expressing Fos protein, whereas pretreat-ment with an anxiolytic benzodiazepine (diazepam) par-tially prevented the stress-induced increase in Fos ex-pression. Restraint stress for 30 min increasedconcentrations of the DA metabolite 3,4-dihydroxy-phenylacetic acid (DOPAC) in the nucleus accumbensand striatum, as well as in the prefrontal cortex. Ret-rograde tracer studies revealed that stress increasedFos protein expression in a distinct subset of DA neuronsprojecting to the prefrontal cortex. In contrast, Fos ex-pression was not increased in any DA neurons projectingto the nucleus accumbens. The present data indicatethat there are at least two functionally distinct DA sys-tems embedded within the prefrontal cortex of the rat.
Acute exposure to mild footshock stress results in themetabolic activation of the dopamine (DA) innerva-tion of the medial prefrontal cortical (PFC), but notother mesotelencephalic DA terminal fields, such asthe nucleus accumbens (NAS) "or striatum (Thierry etal., 1976; Reinhard et al., 1982; Deutch et al., 1985;Iuvone and Dunn, 1986). This regionally specific pat-tern of activation of mesotelencephalic DA terminalfields is also observed after exposure to other rela-tively mild stressors, including conditioned fear (Her-man et al., 1982; Deutch et al., 1985; Claustre et al.,1986). In addition to biochemical changes indicativeof the metabolic activation of the PFC DA system, bothmild footshock stress and conditioned fear increasethe concentration of the DA metabolite 3,4-dihy-droxyphenylacetic acid (DOPAC) and enhance in vivotyrosine hydroxylation in A10 DA neurons in the ven-tral tegmental area (VTA; Deutch et al., 1985; Hiradaet al., 1989), source of DA afferents to the PFC (Swan-son, 1982). There is no apparent metabolic activationof DA neurons in the substantia nigra (SN; A9 cellgroup), source of the striatal DA innervation, or inthe retrorubral field (RRF; A8 cell group), which in-nervates mesolimbic and striatal regions (Deutch etal., 1985, 1988).
The selective activation of the PFC DA innervationis observed only under certain stress conditions. In-creases in either the intensity (e.g., increased foot-shock current) or duration of stress result in the met-abolic activation of mesolimbic DA terminal field areas,such as the NAS (Fadda et al., 1978; Dunn, 1988; Rothet al., 1988); still more intense stressors result in en-hanced DA release and the metabolic activation ofthe striatal DA innervation as well as mesocorticolim-bic DA systems (Keller et al., 1983; Antelman et al.,1988; Dunn, 1988; Roth et al., 1988; Abercrombie etal., 1989).
The mechanisms underlying the regionally specificpattern of augmented DA metabolism and release fol-lowing mild footshock stress are not known. DA re-lease can be enhanced secondary to increases in thefiring rate or changes in the firing pattern of DA neu-rons, and the attendant impulse-mediated release ofthe amine at the terminals. Alternatively, DA releasefrom nerve terminals can occur by presynaptic mech-anisms, which do not involve alterations in the firingrate of the DA neurons, through stimulation of ap-
Cerebral Cortex July/Aug 1991;l:273-292; 1047-3211/91/14.00
at York U
niversity Libraries on July 9, 2014
http://cercor.oxfordjournals.org/D
ownloaded from

propriate heteroceptors located on DA terminals(Chesselet et al., 1982; Chesselet, 1984).
Electrophysiological data indicate that noxiousstimulation of the anesthetized rat selectively increas-es the firing rate of A10 DA neurons that innervatethe PFC, but not DA neurons that project to the NAS(Mantz et al., 1989). These data suggest that the pref-erential metabolic activation of the PFC DA inner-vation by mild stress may be attributable, at least inpart, to an impulse-dependent increase in DA metab-olism and release. However, these data are discordantwith the hypothesis that a similar mechanism is op-erative in the NAS, since the firing rate of mesoac-cumbens DA neurons is not increased by noxious tailpinch. Electrophysiological studies aimed at deter-mining precisely which midbrain DA neurons are ac-tivated by stress have been hampered by the technicaldifficulties of recording from DA neurons identifiedas to projection target by antidromic activation in theawake, freely moving animal.
The expression of the immediate-early gene c-foshas been suggested to serve as a marker of metabol-ically active neurons at the cellular level (Sagar et al.,1988; Dragunowand Faull, 1989; Morgan and Curran,1989). In transcriptional regulation c-Fosis of criticalimportance (Morgan and Curran, 1989; Sheng andGreenberg, 1990). This gene encodes the nuclearphosphoprotein Fos, which, with the protein productof another early-immediate gene (Jun)> binds coop-eratively to the AP-1 DNA regulatory site (Gentz etal., 1989). The expression of Fos protein can be in-duced by a variety of physiological and pharmaco-logical stimuli (Curran and Morgan, 1985; Hunt et al.,1987; Morgan et al., 1987; Sagar et al., 1988; Robertsonet al., 1989; Herrera and Robertson, 1990), consistentwith the suggestion that Fos expression may serve asa useful method for identifying at least certain neu-rons that are metabolically activated.
In an attempt to determine which mesencephalicDA neurons are metabolically activated by stress, wehave examined stress-induced alterations in expres-sion of Fos protein in DA neurons using immunohis-tochemical methods. This approach has been usedpreviously to investigate the effects of stress on med-ullary and pontine catecholaminergic neurons (Cec-catelli et al., 1989).
Materials and Methods
AnimalsAdult male Sprague-Dawley rats (Camm Research LabAnimals, Wayne, NJ), body weight 250-275 gm, wereused as subjects. Animals were group housed on a 12:12-hr light/dark cycle with light onset at 06:00. Foodand water were available ad libitum.
Stress Procedure and Drug InjectionsAnimals were subjected to 30 min restraint stress atroom temperature. This procedure was selected onthe basis of our previous data indicating that 30 minrestraint results in a nonselective enhancement of DAmetabolism across the mesotelencephalic DA termi-
nal fields: DA metabolism is increased in mesolimbicand striatal DA innervations in addition to the PFCDA system (Roth et al., 1988). All animals werestressed in the morning, between 08:30 and 11:00.After 30 min of restraint, animals were placed in aquiet room until death 4 hr 30 min later by chloralhydrate/pentobarbital (Equithesin) overdose; a smallnumber of animals (N = 4) were killed 2 hr 15 minafter the restraint period. In all cases, both stressedand control animals were perfused on the same day,and immunohistochemical processing of tissue wasperformed using the same antibody solutions at thesame time. Similarly, in cases involving pharmaco-logical manipulations requiring parenteral adminis-tration of drugs, both drug- and vehicle-injected (con-trol) animals were treated and perfused on the sameday. The interval of time between injection with Equi-thesin and initiation of perfusion was less than 10min; this interval is short enough to preclude anychange in Fos protein attributable to the anesthetic.
In order to test the ability of an anxiolytic benzo-diazepine to prevent stress-induced changes in Fosexpression, animals were intraperitoneally injectedwith 5.0 mg/kg diazepam (DZ) 30 min prior to re-straint; animals were killed 4 hr 30 min after restrainttermination. Animals treated with DZ alone were sac-rificed 5 hr 30 min after injection, resulting in aninterval between DZ injection and death comparableto that used in animals pretreated with the benzodi-azepine and subjected to 30 min of restraint stress.
The effects of the anxiogenic benzodiazepine in-verse agonist FG 7142 (./V-methyl-j8-carboline-3-car-boxylate) were also examined. FG 7142 (20 mg/kg,i.p.; Research Biochemicals Inc.) was injected 5 hrprior to death; this survival period was selected inorder to achieve a time comparable to that of animalssubjected to 30 min restraint stress. As a control pro-cedure, vehicle (1.5% Tween 80 in water) was in-jected, and animals were killed 5 hr later.
ImmunobistocbemistryAnimals were deeply anesthetized with Equithesinand transcardially perfused with phosphate-bufferedsaline, followed by 250 ml of ice-cold 4% parafor-maldehyde in 0.1 M sodium phosphate buffer (pH7.4). The brains were postfixed at 4°C for 14-18 hrbefore being cryoprotected in 30% sucrose in 0.1 Msodium phosphate; frozen sections were cut at 40 AIMon a sliding microtome. In some cases vibratome sec-tions were prepared.
Sections were washed in 0.05 M Tris-buffered saline(TBS), pH 7.4, incubated in methanolic peroxide for10 min, washed extensively in TBS, and then incu-bated in TBS containing 4% normal goat serum and0.3% Triton X-100 (TBS+) for 30-60 min. Free-floatingsections were then incubated in a rabbit polyclonalantiserum directed against the M-peptide of Fos (1:2000) for 72 hr at 4°C under constant agitation. Thisantibody has been previously characterized and rec-ognizes Fos and Fos-related antigens (Quinn et al.,1989).
The free-floating sections were then washed in TBS+
274 PFC DA Neurons Express Fos • Deutch et al.
at York U
niversity Libraries on July 9, 2014
http://cercor.oxfordjournals.org/D
ownloaded from

prior to incubation in goat anti-rabbit IgG (1:50; An-tibodies, Inc.) for 120 min. Sections were washedextensively in TBS+ and incubated in rabbit PAP (1:100; Sternberger-Meyer Immunocytochemicals) for90 min. The sections were again washed in TBS, andthen reacted in 0.025% diaminobenzidine in 0.1 Msodium acetate (pH 6.5) containing 1.5% nickel am-monium sulfate and 0.15% cobalt chloride, with0.0009% H2O2. The reaction was stopped by rinsingin TBS, and the sections were immersed in 1% para-formaldehyde in 0.1 M sodium phosphate for 2 min.Control procedures included preadsorption of theprimary antibody with the synthetic M-peptide (10"6
M) against which the antibody was generated, andimmunohistochemical processing in the absence ofthe primary antibody. There was no detectable label-ing after preadsorption of the antibody with syntheticM-peptide or following deletion of the primary anti-body; in addition, preadsorption with M-peptide elim-inated the ability of the antibody to recognize Fos orFos-related antigens on Western blots. Sections werewashed in TBS before being incubated in TBS+ for 30min, followed by incubation in a 1:1000 dilution ofmouse monoclonal antibody directed against tyrosinehydroxylase (TH; Semenenko et al., 1986) overnightat room temperature. Sections were then processedusing a biotinylated secondary antibody and avidin(Vector Laboratories), and the reaction product wasrevealed with 0.05% diaminobenzidine in TBS con-taining 0.009% H2O2. Sections were mounted on slides,dehydrated, cleared, and coverslipped. The locationof Fos-like-immunoreactive (Fos-li) and TH-li neu-rons were then plotted. In order to avoid cumbersomephrasing (e.g., Fos-li-TH-li neurons), the immuno-reactive protein products being detected are referredto as Fos and TH.
Double-labeled Fos-TH neurons were plotted andcounted in each of the subnuclei of the VTA, as wellas in the substantia nigra and retrorubral field, at fourdifferent anteroposterior levels as illustrated in Figure1. The limits of the subnuclei of the VTA were definedusing the boundaries of Paxinos and Watson (1982).We deviated from the description of the borders ofthese subnuclei as delineated by Paxinos and Watsonas follows: The rostral pole of the VTA, which is notseparated by Paxinos and Watson into individual nu-clei, was divided into a dorsal sector, which we in-cluded as the rostral pole of the nucleus parabran-chialis pigmentosus, and a ventral sector, which wasconsidered as a part of the nucleus paranigralis. Inaddition, since there is a dorsal population of caudalAlO DA neurons that extends beyond the dorsal limitsof the caudal linear nucleus as defined by Paxinos andWatson, we arbitrarily expanded the boundaries ofthis nucleus to extend dorsally to the area medial tothe parvocellular region of the oculomotor nucleus.The subnuclei of the VTA as defined in the presentstudy are shown in Figure 1, adapted from Paxinosand Watson (1982).
Retrograde tracer injections [50 nlofa 2.5% solutionof fluorogold (FG) in 0.01 M sodium acetate, pH 5]were made into the medial PFC (N = 4) or into the
NAS 0V= 4) of adult male rats. Two weeks after sur-gery, animals were subjected to 30 min restraint stressand, 4 hr 30 min later were overdosed with Equithesinand processed for immunohistochemistry. Sectionswere first processed for the localization of Fos-li usingthe immunoperoxidase method described above. Sec-tions were then incubated in mouse anti-TH (1:500)for 14-16 hr at room temperature, washed in TBS+,and incubated in rhodamine-conjugated goat anti-mouse IgG (1:50; Chemicon International) for 60 minat 37°C. Sections were then extensively washed inTBS, slide mounted, and viewed with epifluores-cence. In order to determine the projection targets ofthose DA neurons in which stress elicited an in-creased expression of Fos protein, neurons that wereretrogradely labeled and exhibited both Fos and THwere charted and counted.
BiochemistryIn order to confirm that stress resulted in the meta-bolic activation of DA neurons, animals were re-strained for 30 min and killed immediately thereafter;control animals were anesthetized and perfused uponremoval from the home cage. Animals were killed atthe end of the 30 min restraint period since we hadpreviously observed that DA turnover returns to base-line values within 30 min of the termination of stress(Bannon et al., 1986). The brain was removed fromthe cranial vault, and the PFC, NAS, striatum, VTA,and SN were dissected as previously described(Deutch et al., 1985).
The samples were stored at —75°C until assayed forconcentrations of dopamine and its primary metab-olite 3,4-dihydroxyphenylacetic acid (DOPAC) byHPLC with electrochemical detection, following themethod of Kalivas and Taylor (1985). Briefly, sampleswere sonicated in 500 /il mobile phase (0.1 M trichlo-roacetic acid, 0.01 M sodium acetate, and 0.001 M EDTA,pH 3.85, with 18% methanol admixed to modulateretention time) containing 1 x 10~7 M isoproterenolas the internal standard. One hundred microliters ofthe homogenate were removed for subsequent pro-tein analysis, and the remaining sample was centri-fuged. The supernatant was then centrifuged again,and the second supernatant was injected onto a re-verse-phase C18 column; the flow rate of the mobilephase was set at 0.9 ml/min, and the amines wereoxidized at 0.7 V. Protein levels were determined bythe method of Lowry et al. (1951).
Statistical AnalysisData were analyzed by means of analyses of varianceand subsequent post-hoc tests (Bonferroni r-test) whenindicated. Initial two-way analyses of variances of theeffects of control condition (quiet normal control andvehicle-injected control) on the number of Fos-DAneurons in the VTA, SN, or RRF, or in the varioussubnuclei of the VTA, did not reveal any significantinteraction or main effect. Accordingly, the two con-trol groups were collapsed into one combined con-trol; descriptive data from the individual and com-bined control groups are shown in Tables 1 and 2.
Cerebral Cortex July/Aug 1991, VI N 4 275
at York U
niversity Libraries on July 9, 2014
http://cercor.oxfordjournals.org/D
ownloaded from

scp /' ;:\ \RMC\
276 PFC DA Neurons Express Fos • Deutch et al.
at York U
niversity Libraries on July 9, 2014
http://cercor.oxfordjournals.org/D
ownloaded from
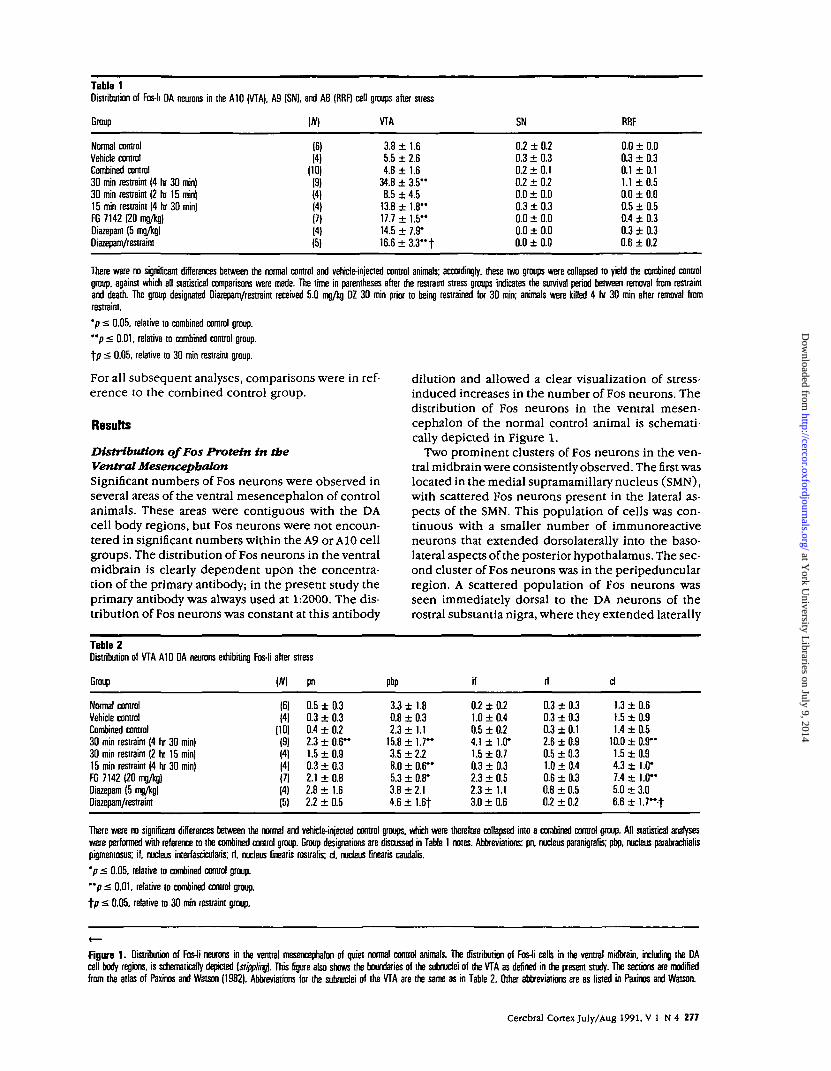
Table 1Distribution of Fos-li DA neurons in the A10 (VTA), A9 (SN), and A8 (RRF) cell groups after stress
Group (#) VTA SN RRF
Normal controlVehicle controlCombined control30 min restraint (4 hr 30 min)30 min restraint (2 hr 15 min)15 min restrain! (4 hr 30 min)FG 7142 (20 mg/kg)Diazepam (5 mg/kg)Diazepam/restraint
(6)(4)
(10)(9)(4)(4)(?)(4)(5)
3.8 ± 1.65.5 ± 2.64.8 ± 1.6
34.8 ± 3 .5 "8.5 ± 4.5
13.8 ± 1 . 8 "17.7 ± 1.5"14.5 ± 7.9*16.6 ± 3 . 3 " t
0.2 ± 0.20.3 ± 0.30.2 ± 0.10.2 ± 0.20.0 ± 0.00.3 ± 0.30.0 ± 0.00.0 ± 0.00.0 ± 0.0
0.0 ± 0.00.3 ± 0.30.1 ± 0.11.1 ± 0.50.0 ± 0.00.5 ± 0.50.4 ± 0.30.3 ± 0.30.6 ± 0.2
There were no significant differences between the normal control and vehicle-injected control animals; accordingly, these two groups were collapsed to yield the combined controlgroup, against which all statistical comparisons were made. The time in parentheses after the restraint stress groups indicates the survival period between removal from restraintand death. The group designated Diazepam/restraint received 5.0 mg/kg DZ 30 min prior to being restrained for 30 min; animals were killed 4 hr 30 min after removal fromrestraint.
'p £ 0.05. relative to combined control group.
"p < 0.01, relative to combined control group.
t/> £ 0.05. relative to 30 min restraint group.
For all subsequent analyses, comparisons were in ref-erence to the combined control group.
Results
Distribution ofFos Protein in theVentral MesencepbalonSignificant numbers of Fos neurons were observed inseveral areas of the ventral mesencephalon of controlanimals. These areas were contiguous with the DAcell body regions, but Fos neurons were not encoun-tered in significant numbers within the A9 or A1O cellgroups. The distribution of Fos neurons in the ventralmidbrain is clearly dependent upon the concentra-tion of the primary antibody; in the present study theprimary antibody was always used at 1:2000. The dis-tribution of Fos neurons was constant at this antibody
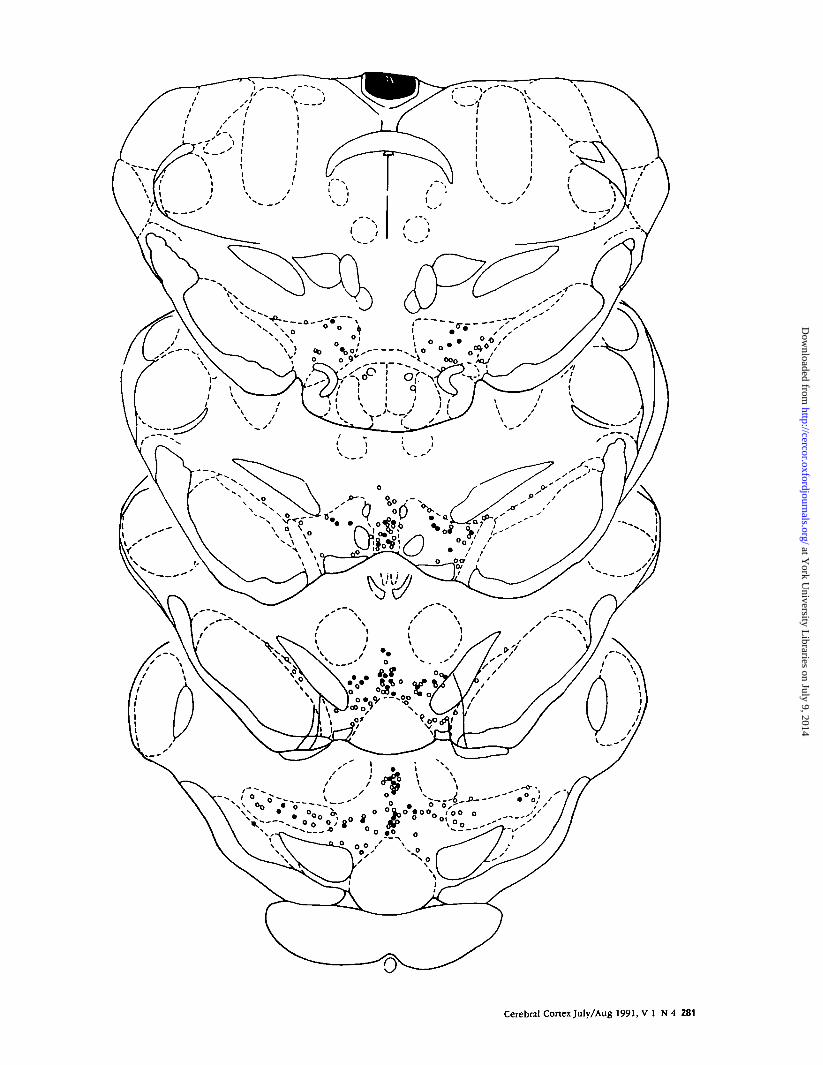
dilution and allowed a clear visualization of stress-induced increases in the number of Fos neurons. Thedistribution of Fos neurons in the ventral mesen-cephalon of the normal control animal is schemati-cally depicted in Figure 1.
Two prominent clusters of Fos neurons in the ven-tral midbrain were consistently observed. The first waslocated in the medial supramamillary nucleus (SMN),with scattered Fos neurons present in the lateral as-pects of the SMN. This population of cells was con-tinuous with a smaller number of immunoreactiveneurons that extended dorsolaterally into the baso-lateral aspects of the posterior hypothalamus. The sec-ond cluster of Fos neurons was in the peripeduncularregion. A scattered population of Fos neurons wasseen immediately dorsal to the DA neurons of therostral substantia nigra, where they extended laterally
Table 2Distribution of VTA A10 DA neurons exhibiting
Group
Normal controlVehicle controlCombined control30 min restraint (4 hr 30 min)30 min restrain! (2 hr 15 min)15 min restraint (4 hr 30 min)FG 7142 (20 mg/kg)Diazepam (5 mg/kg)Diazepam/restraint
Fos-li after stress
W
(6)(41
(10)(9)14)(4)(7)(4)(5)
pn
0.5 ± 0.30.3 ± 0.30.4 ± 0.22.3 ± 0 . 6 "1.5 ±0.90.3 ± 0.32.1 ± 0.82.8 ± 1.62.2 ± 0.5
pbp
3.3 ± 1.80.8 ± 0.32.3 ± 1.1
15.8 ± 1.7"3.5 ± 2.28.0 ± 0.6"5.3 ± 0.8*3.8 ± 2.14.6 ± 1.6t
if
0.2 ± 0.21.0 ± 0.40.5 ± 0.24.1 ± 1.0*1.5 ±0.70.3 ± 0.32.3 ± 0.52.3 ±1.13.0 ± 0.6
rl
0.3 ± 0.30.3 ± 0.30.3 ± 0.12.6 ± 0.90.5 ± 0.31.0 ± 0.40.6 ± 0.30.8 ± 0.50.2 + 0.2
cl
1.3 ± 0.61.5 ± 0.91.4 ±0.5
10.0 ± 0.9"1.5 ±0.94.3 ± 1.0*7.4 ± 1.0"5.0 ± 3.06.6 ± 1.7—-t
There were no significant differences between the normal and vehicle-injected control groups, which were therefore collapsed into a combined control group. All statistical analyseswere performed with reference to the combined control group. Group designations are discussed in Table 1 notes. Abbreviations: pn. nucleus paranigralis; pbp. nucleus parabrachialispigmentosus: if. nucleus interfascicularis; rl. nucleus linearis rostralis; cl, nucleus linearis caudatis.
'p < 0.05, relative to combined control group.
"p < 0.01. relative to combined control group.
f/7 < 0.05. relative to 30 min restraint group.
Figure 1 . Distribution of Fos-li neurons in the ventral mesencephalon of quiet normal control animals. The distribution of Fos-li cells in the ventral midbrain, including the DAcell body regions, is schematically depicted [stippling. This figure also shows the boundaries of the subnudei of the VTA as defined in the present study. The sections are modifiedfrom the atlas of Paxinos and Watson (1982). Abbreviations for the subnudei of the VTA are the same as in Table 2. Other abbreviations are as listed in Paxinos and Watson.
Cerebral Cortex July/Aug 1991, V 1 N 4 277
at York U
niversity Libraries on July 9, 2014
http://cercor.oxfordjournals.org/D
ownloaded from

to merge with a larger number of Fos cells extendingfrom the pars posteropolaris of the zona incerta to theperipeduncular area. The peripeduncular neurons thatexpressed Fos were present throughout the antero-posterior extent of the region.
Effects of Restraint Stress on Fos Expression inMidbrain DA NeuronsRats restrained for 30 min and killed 4 hr 30 min laterexhibited an increased number of midbrain DA neu-rons expressing Fos protein (Figs. 2-4). The stress-elicited increase in dual-labeled Fos-TH neurons wasrestricted to the VTA (A10 cell group). No significantincrease in the number of A9 or A8 DA neurons ex-pressing Fos protein after stress exposure was ob-served (Table 1). Since a two-factor (treatment x re-gion) ANOVA revealed that the number of double-labeled VTA neurons in quiet control animals and inanimals that received a vehicle injection did not sig-nificantly differ [interaction F(l,4) = 1.33, NS], for allsubsequent statistical comparisons the two types ofcontrols (normal controls and vehicle-injected con-trols) were pooled (Tables 1, 2).
The stress-induced increase in double-labeled (Fos-TH) neurons was regionally specific within the VTA(see Table 1, Figs. 3, 4). Stress resulted in a largeincrease in the number of Fos-DA neurons in thecaudal linear nucleus; the mean number of DA neu-rons expressing Fos protein in this nucleus was seventimes greater than observed in control animals (seeTable 2). The other subdivision of the VTA in whichstress elicited a large increase in the number of dou-ble-labeled cells was the nucleus parabrachialis, par-ticularly within the dorsal aspects (Table 2). Stressincreased the number of interfascicular nucleus A10DA neurons expressing Fos protein; however, the to-tal number of double-labeled cells observed after stressin this nucleus was relatively small. Similarly, stresselicited a small but significant increase in the numberof nucleus paranigralis DA cells expressing Fos pro-tein. The number of rostral linear DA neurons thatwere Fos positive was not significantly increased after30 min restraint. The distribution of double-labeledcells across the VTA, as reflected by the percentageof the total number of double-labeled cells observedin each of the subnuclei comprising the VTA, was
virtually identical in control and restraint-stressed an-imals (see Fig. 5).
The total number of VTA DA neurons expressingFos after 15 min of restraint was significantly greaterthan in control rats, but significantly less than thatobserved after 30 min restraint (Table 2). However,the regional distribution of the Fos-DA cells after 15min restraint was very similar to that observed afterthe longer-duration stressor, with an increased num-ber of cells in the parabrachialis and caudal linearnuclei, but not in the other VTA areas or in the SN orRRF.
The number and distribution of VTA DA cells ex-pressing Fos protein in animals restrained for 30 minbut killed 2 hr 15 min (rather than 4 hr 30 min) afterremoval from restraint were not significantly differentfrom the number of A10 DA neurons expressing Fosprotein at the shorter survival interval (see Tables 1,2).
Effects ofFG 7142 on Fos Expression inMidbrain DA NeuronsTreatment of rats with the anxiogenic 0-carboline FG7142 increased the number of VTA DA neurons ex-pressing Fos protein (see Fig. 6, Table 1). The totalnumber of A10 DA neurons that were Fos positive waslower than that observed after 30 min restraint stress(with 4 hr 30 min survival), but significantly higherthan that observed in animals injected with vehicle(Table 1). Injection of the vehicle did not significantlyincrease the number of DA neurons expressing Fosprotein relative to quiet normal control animals (seeTables 1, 2).
Although FG 7142 administration increased thenumber of A10 DA neurons that expressed Fos, albeitto a lesser degree than did restraint stress, the distri-bution of these double-labeled cells differed from thatobserved after 30 min restraint stress (Table 2, Fig.5). The percentage of double-labeled neurons in thenucleus parabrachialis (of the total population of Fos-TH neurons in the VTA) was lower than observed afterrestraint, whereas the percentage of neurons in thecaudal linear nucleus was greater than in restrainedanimals (see Figs. 5, 6); there was a tendency towarda greater percentage increase in the nucleus parani-gralis, as well. Thus, the distribution of Fos-DA neu-
Figura 2 . Examples of double-labeled Fos-li—TH-li neurons in the ventral tegmemal area of the rat. A A DA neuron in the nucleus parabrachialis pigmentosus exhibiting Fos-li(a/roiv). The Fos-li product is confined to the nucleus and is clearly visible against the more diffuse cytoplasmic TH-li-reactive product. The arrowheads point to Fos-li material inthe nucleus of an adjacent DA neuron. In this neuron, the density of the reaction product is considerably less than seen in the first neuron, but is again confined to the nucleus:when viewed in color, this nuclear staining was gray and contrasted against the brown TH product. B. Two Fos-li neurons in the nucleus paranigralis of the VTA. A double-labeledFos-li-TH-ii neuron [arrow] can be seen adjacem to a neuron that exhibits Fos-li but is TH negative [arrowhead^. The single-labeled Fos-li neuron is slightly out of the plane offocus and is overlying TH-li fibers. C. Double-labeled Fos-li DA neurons in the caudal linear nucleus. Two double-labeled cells (arrows) can be seen near a norMiopaminergic Fos-licell [arrmheao). D, A double-labeled Fos-li-TH-h' neuron [mow] in the dorsal extreme of the caudal linear nucleus. To the right of this neuron are two large dopaminergic neuronsthat are also Fos-li, but the nuclear Fos-li reaction product is not in the plane of focus.
Fignra 3 . Distribution of DA neurons expressing Fos protein {solid circles) and non-dopaminergic neurons exhibiting Fos-li [open circles] in the ventral mesencephakm of a quietnormal comrol animal (case CFS-107). Only a few dopamine neurons are immunoreactive foe Fos protein, whereas there is a somewhat greater number of Fos-li neurons that arenot dopaminergic within the general borders of the mesencephalic DA cell groups. Fos-li neurons outside the limits of the AB, A9, and A10 DA cell groups are not plotted.
Figure 4 . The distribution of Fos-li Da neurons [solid circles) and mvdopaminergic neurons immunoreactive for Fes protein in the ventral midbrain pi a rat subjected to 30min restraint stress and lulled 4 hr 30 mm later (case CFS-109). There is a relatively large number of double-labeled Fos-li-TH-li neurons in the VTA. in contrast to the normalcontrol animal (Fig. 4). Particularly striking is the concentration of the Fos-li DA neurons in the nucleus parabrachialis and the caudal linear nucleus of the VTA. In contrast, thereare virtually no Fos-li DA neurons in the SN or the RRF.
278 PFC DA Neurons Express Fos • Deutch et al.
at York U
niversity Libraries on July 9, 2014
http://cercor.oxfordjournals.org/D
ownloaded from

' # •«* ,
B
M IP
Cerebral Cortex July/Aug 1991, V 1 N 4 279
at York U
niversity Libraries on July 9, 2014
http://cercor.oxfordjournals.org/D
ownloaded from

280 PFC DA Neurons Express Fos • Deutch et al.
at York U
niversity Libraries on July 9, 2014
http://cercor.oxfordjournals.org/D
ownloaded from
Cerebral Cortex July/Aug 1991, V 1 N 4 281
at York U
niversity Libraries on July 9, 2014
http://cercor.oxfordjournals.org/D
ownloaded from

NORMAL CONTROL
1 so -u
j . H£ 50-
30-RESTRANTW30" )
FG7142
pn pbp if rl d pn pbp if r l cl pn pbp if rl el
Figure 5 . The distribution of Fos-li DA neurons in the different subnudei of theVTA after 30 min restraint stress or FG 7142 treatment. The data are expressed asthe percentage of the total number of VTA double-labeled neurons in each of the fivesubnudei that comprise the VTA. While the total number ol Fos-li DA neurons in theVTA is seven times greater in animals restrained for 30 min than in control animals,the relative distribution of those double-labeled cells observed in the subnudei isvirtually the same for the two groups. In contrast, the regional distribution of Fos-liDA neurons observed in animals treated with the anxiogenic /J-carboline FG 7142 isquite different and is marked by a higher percentage of caudal linear nucleus (e/| DAneurons expressing Fos protein but a lower percentage of Fos-li DA neurons in thenucleus parabrachialis (pip)- The number of paranigral nucleus (pn) DA neuronsexhibiting Fos-li is also somewhat higher in the animals treated with FG 7142. //,nudeus interfascicularis; rl, nudeus linearis rostralis.
rons across the subnudei of the VTA was more wide-spread after FG 7142 than seen after 30 min restraint,but the total number of double-labeled A10 neuronswas less.
Effects ofDZ Pretreatment on the Stress-inducedChanges in Fos NeuronsPretreatment of rats with DZ 30 min prior to restraintsignificantly reduced the total number of double-la-beled VTA neurons compared to animals restrainedbut not receiving the anxiolytic drug (Table 1). How-ever, the number of VTA double-labeled cells in DZ-pretreated rats was also significantly greater than thenumber of Fos-TH neurons in control animals (Table1).
DZ pretreatment decreased the number of DA cellsimmunoreactive for Fos in the caudal linear and para-brachial nuclei (see Table 2). The number of nucleusinterfascicularis Fos-DA neurons was not significantlyreduced from the number observed in animals sub-jected to 30 min restraint.
There were also differences between animals pre-treated with DZ but not restrained and normal controlanimals. The total number of A10 DA neurons ex-pressing Fos in animals injected with DZ but notstressed was comparable to that observed in DZ-treat-ed animals that were subsequently stressed (Tables1,2). However, in the animals pretreated with DZ butnot stressed, there was no significant increase in anysingle subdivision of the VTA (e.g., the nucleus para-nigralis), although the total number of VTA double-
labeled neurons was significantly greater than in con-trol animals.
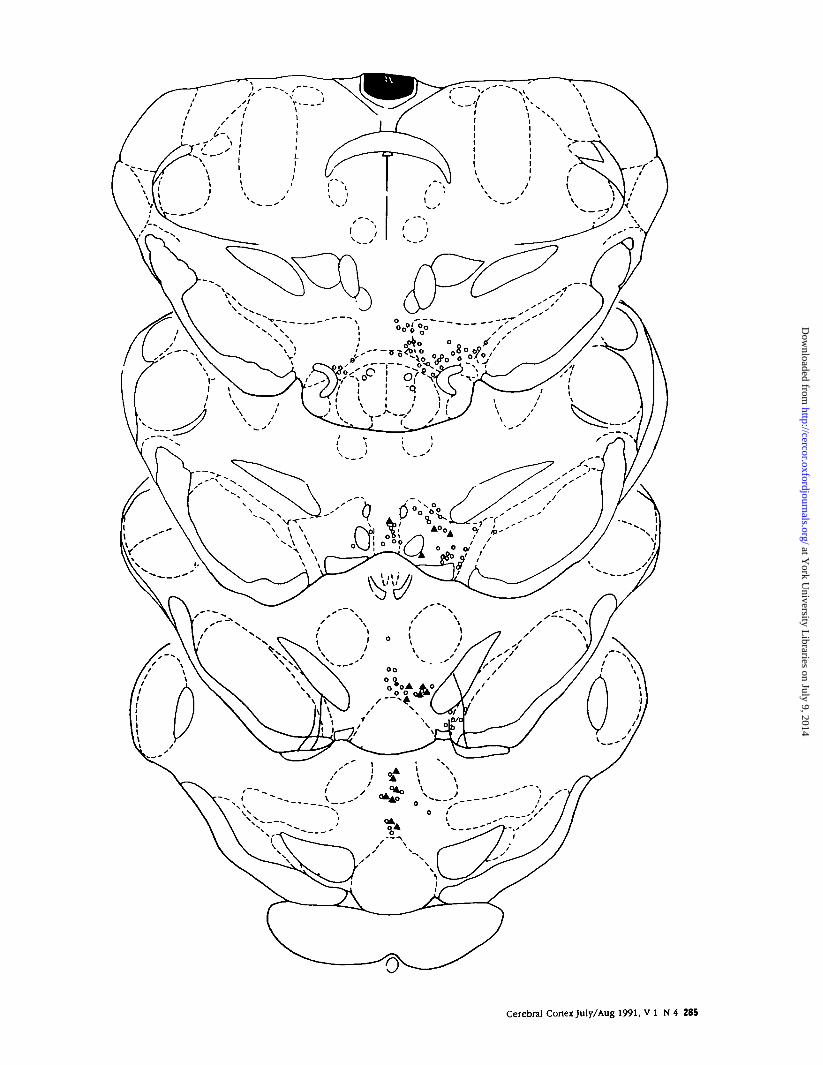
Stress-elicited Changes in Fos in DA NeuronsInnervating the PFC or NASStress increased Fos protein expression in DNA neu-rons projecting to the PFC, but not in DA neuronsinnervating the NAS. In animals with injections of FGin the PFC, retrogradely labeled cells were observedin the VTA; only a few cells were seen in the medialSN and in the transition zone between the A10 andA8 DA cell groups (see Swanson, 1982; Deutch et al.,1988). Fos was expressed both in cells retrogradelylabeled from the PFC as well as in FG-negative cells(see Fig. 7). The cells that were both Fos and FGpositive were mainly seen in the caudal linear nucle-us, with fewer observed in the nucleus parabrachialis(see Fig. 8). Cells that were retrogradely labeled andwere Fos positive virtually always were also TH im-munoreactive, indicating that the DA (but not non-dopaminergic) mesocortical neurons were activatedby stress.
Very few cells that were retrogradely labeled fromthe NAS expressed Fos. A large number of FG-positive cells were seen present in the nucleus para-nigralis and the medial SN; fewer retrogradely labeledneurons were present in the nucleus parabrachialis.Fos-positive neurons that were retrogradely labeledfrom the NAS and were unambiguously TH immu-noreactive were not seen.
Effects of Stress on Fos Expression in Non-DAMidbrain NeuronsRestraint stress increased Fos expression in DA neu-rons of the ventral midbrain. While we did not quan-titate the number of non-DA neurons expressing Fos,two populations of neurons outside of the DA cellgroups appeared to increase Fos expression after re-straint stress. These were the neurons of the peripe-duncular area and adjacent posterolateral thalamusand neurons in the SMN.
Stress appeared to markedly increase the numberof Fos neurons in the peripeduncular region. The Foscells extended ventromedially at the rostral pole (tomerge and form a continuous population with theneurons of the zona incerta), as well as dorsally (con-tinuous with the posterior intralaminar thalamic nu-cleus). An apparent stress-induced increase in thenumber of Fos neurons in this region is consistentwith the findings of LeDoux et al. (1986), who notedthat lesions specific for this area disrupt classical con-ditioning of emotional responses to auditory stimuli.Stress also appeared to increase the number of non-DA Fos neurons in the medial SMN. Finally, it ap-peared that there may be an increase in the numberof non-DA cells in the VTA that exhibited Fos afterrestraint stress (see Figs. 3, 4).
Figure 6 . Distribution of Fos-li DA neurons {solid aides/; and rnHJopaminergic neurons exhibiting Fos-li {open dries) in the ventral midbrain after FG 7142 administration(case CFS-129). While the number of dual-labeled Fos-li-TH-li neurons in the VTA is less titan that observed following restraint stress (but significantly greater than observed invernde-injected control subjects), the panem of distribution of Fos-fi DA cells across the VTA is similar to that observed following restraint stress (see Fig. 5). An increased numberof Fos-li DA neurons in the caudal linear nucleus and nudeus parabrachialis can be seen.
282 PFC DA Neurons Express Fos • Deutch et al.
at York U
niversity Libraries on July 9, 2014
http://cercor.oxfordjournals.org/D
ownloaded from

Cerebral Cortex July/Aug 1991, VI N 4 283
at York U
niversity Libraries on July 9, 2014
http://cercor.oxfordjournals.org/D
ownloaded from
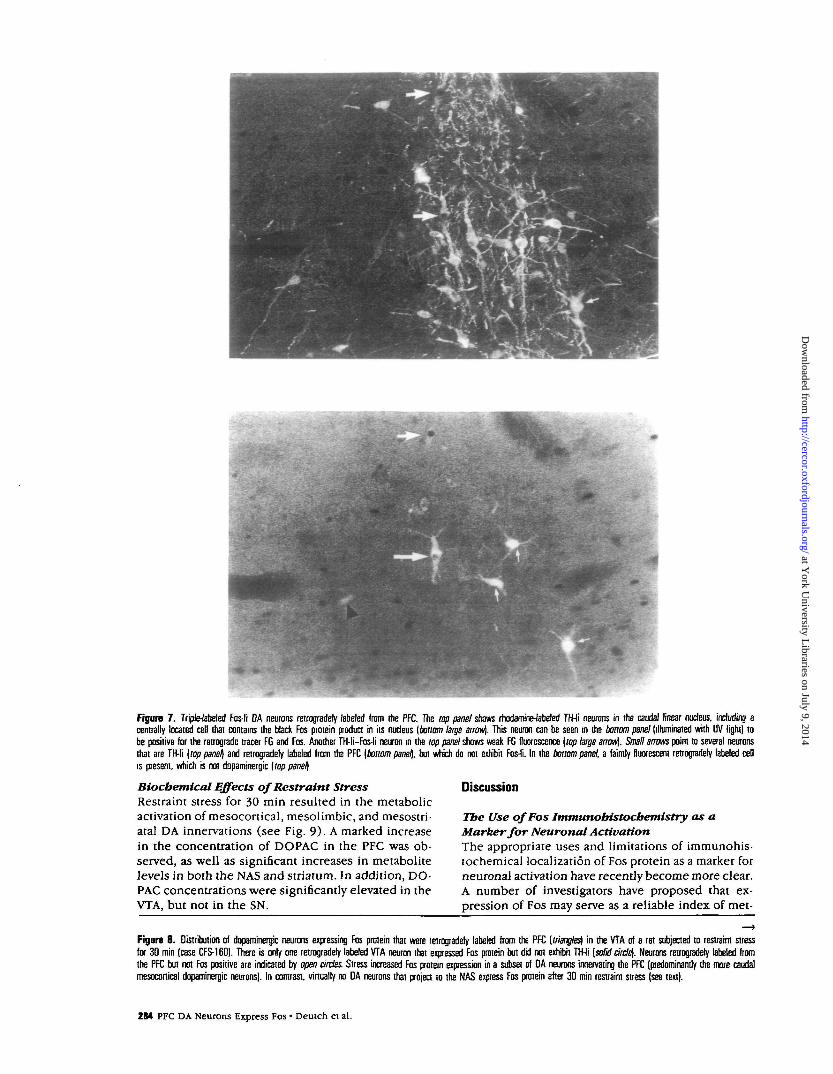
Figure 7 . Triple-labeled Fos-li DA neurons retrogradely labeled from the PFC. The top panel shows rhodamine-fabeled TH-li neurons in the caudal linear nucleus, including acentrally located cell that contains the black Fos protein product in its nucleus [bottom large arrow). This neuron can be seen in the bonom panel (illuminated with UV light) tobe positive for the retrograde tracer FG and Fos. Another TH-li—Fos-li neuron in the top panel shows weak FG fluorescence [top large arrow). Small arrows point to several neuronsthat are TH-li [top panel) and retrogradely labeled from the PFC [bottom panel\. but which do not exhibit Fos-li. In the bottom panel, a faintly fluorescent retrogradely labeled cellis present, which is not dopaminergic [top panel]
Biochemical Effects of Restraint StressRestraint stress for 30 min resulted in the metabolicactivation of mesocortical, mesolimbic, and mesostri-atal DA innervations (see Fig. 9). A marked increasein the concentration of DOPAC in the PFC was ob-served, as well as significant increases in metabolitelevels in both the NAS and striatum. In addition, DO-PAC concentrations were significantly elevated in theVTA, but not in the SN.
Discussion
Tbe Use of Fos Immunobistocbemistry as aMarker for Neuronal ActivationThe appropriate uses and limitations of immunohis-tochemical localization of Fos protein as a marker forneuronal activation have recently become more clear.A number of investigators have proposed that ex-pression of Fos may serve as a reliable index of met-
Figure 8 . Distribution of dopaminergic neurons expressing Fos protein that were retrogradery labeled from the PFC [triangles] in the VTA of a rat subjected to restraint stressfor 30 min (case CFS-160). There is only one retrogradely labeled VTA neuron that expressed Fas protein but did not exhibit TH-li [solid circle]. Neurons retrogradery labeled fromthe PFC but not Fas positive are indicated by open circles. Stress increased Fos protein expression in a subset of DA neurons innervating the PFC (predominantly the more caudalmesocortical dopaminergic neurons). In contrast, virtually no DA neurons that project to the NAS express Fos protein after 30 min restrain! stress (see text).
284 PFC DA Neurons Express Fos • Deutch et al.
at York U
niversity Libraries on July 9, 2014
http://cercor.oxfordjournals.org/D
ownloaded from
Cerebral Cortex July/Aug 1991, V 1 N 4 285
at York U
niversity Libraries on July 9, 2014
http://cercor.oxfordjournals.org/D
ownloaded from

D NORMAL CONTROL
0 30" RESTRAINT
T
T -r-
PFC NAS CP VTA SN
Figure 9 . DOPAC concentrations in selected telencephalic DA terminal field regionsand in the VTA and SN in animals killed immediately after 30 min restraint stress.In addition to stress-induced increases in PFC DA metabolism, stress elicited anincrease in DOPAC concentrations mesolimbic sites such as the NAS. and the striatum(CP). Moreover, stress augmented DA metabolism in the VTA but not SN\; 'P £0.05. "P == 0.01.
abolic activation at the cellular level (Hunt et al.,1987; Sagar et al., 1988; Dragunow and Faull, 1989;Morgan and Curran, 1989; Bullitt, 1990). In the nor-mal brain, Fos-li is seen only in neurons, and not innon-neuronal elements such as glia and endothelialcells (Mugnaini et al., 1989); in contrast, Fos or Fos-related antigens (Fra) are present in glia maintainedin cell culture (Hisanaga et al., 1990) and in glia insitu following trauma (Dragunow and Robertson,1988).
While a number of physiological and pharmaco-logical stimuli that affect depolarization increase Fosexpression (Curran and Morgan, 1985; Hunt et al.,1987; Morgan et al., 1987; Sagar et al., 1988; Robertsonet al., 1989), depolarization per se does not appearto be a necessary condition for the induction of Fos(see Dragunow and Faull, 1989). For example, im-munohistochemical studies have revealed that Fos-lineurons may be absent from brain areas in whichphysiological data indicate that certain neurons aredepolarized (Bullitt, 1990). While the utility of Fosas an index of depolarization is doubtful, there is agrowing consensus that in neurons which express Fosprotein there is a functional increase in at least certainkey intracellular metabolic events in the somatoden-dritic region (Sagar et al., 1988; Ceccatelli et al., 1989;Dragunow and Faull, 1989; Bullitt, 1990; Lee et al.,1990). Thus, alterations in ion flux or, alternatively,an increase in second messengers that lead to theactivation of protein kinases but which are not nec-essarily accompanied by depolarization (e.g., in re-sponse to a neuromodulator), can increase Fos ex-pression (Curran and Morgan, 1985; Montminy et al.,1990; Sheng et al., 1990). Thus, a metabolic alterationin the somatodendritic area may be correlatedwith Fos expression. However, negative results instudies of Fos expression, particularly in the absenceof data illustrating that an enhancement of c-fos orFos can be observed in a given neuronal population,cannot be considered conclusive.
Stress-induced Alterations in Fos ProteinExpression in A1O DA NeuronsAlthough restraint stress augmented DA metabolismin the PFC, NAS, and striatum, the number of Fos-DA neurons was increased by the same stressor onlyin the VTA, and not in the SN or RRF. The increasednumber of VTA Fos-DA neurons appeared to be re-lated to stress duration, since 15 min restraint resultedin a smaller number of double-labeled cells than 30min restraint.
The increased number of Fos-TH neurons in theVTA following stress may reflect an increase in theamount of Fos protein in these neurons above basallevels too low to detect using immunohistochemicaltechniques or, alternatively, may indicate that a newpopulation of cells which under basal conditions doesnot express Fos protein is being activated by stress.The distribution of neurons across the VTA that areboth Fos and TH positive favors the first possibility.The total number of A10 DA neurons that were Fos-li was significantly increased by stress, and the relativedistribution of these neurons across the VTA was thesame in both restraint-stressed and control animals(see Fig. 5). For example, 46.9% of the double-la-beled (Fos-TH) neurons present in the VTA of con-trol animals were in the nucleus parabrachialis; 45.4%of the double-labeled cells in stressed animals werepresent in this same nucleus. The regional distribu-tion across the VTA is virtually identical for both con-trol and restraint stressed animals (Fig. 5). This ob-servation suggests that stress simply increases Fosprotein expression to levels above the threshold fordetection by immunohistochemistry, and that in quietcontrol animals the same cells express Fos proteinbut at very low levels. The low level of expression ofFos protein in the awake animal maintained in a quietenvironment is consistent with these neurons beingrelatively sensitive to environmental stimuli.
Mesoprefrontal Cortical DA Neurons andFos ExpressionElectrophysiological data suggest that stress increasesthe firing rate of mesoprefrontal cortical, but not me-solimbic or nigrostriatal, DA neurons (Trulson andPreussler, 1984; Strecker and Jacobs, 1985; Jacobs,1987; Mantz et al., 1989). VTA DA neurons that ex-pressed Fos after 30 min stress were mainly observedin the dorsal aspects of the nucleus parabrachialispigmentosus and in the interfascicular and caudallinear nuclei. This distribution is similar to the dis-tribution of VTA DA cells that project to the PFC(Swanson, 1982). The retrograde tracer studies con-firmed that stress increased Fos expression in meso-prefrontal cortical, but not mesoaccumbens, DA neu-rons.
A large number of VTA neurons projecting to thePFC were not Fos-li. While more than half of the VTAneurons that project to the PFC do not contain DA(Swanson, 1982; Gillham et al., 1990), the number oftriple-labeled cells seen in the VTA clearly suggeststhe existence of a population of DA neurons that pro-jects to the PFC and is not activated by stress. As seen
286 PFC DA Neurons Express Fos • Deutch et al.
at York U
niversity Libraries on July 9, 2014
http://cercor.oxfordjournals.org/D
ownloaded from

in Figures 7 and 8, there are indeed such mesocorticalDA neurons that do not increase Fos expression inresponse to stress. This observation indicates the pres-ence of at least two functionally distinct populationsof mesoprefrontal cortical DA neurons: those that in-crease Fos expression after stress and those that donot. This is consistent with our previous suggestionthat the DA innervation of the PFC is functionallyheterogeneous as reflected by biochemical respon-siveness to stress (see Deutch and Roth, 1990). Fur-ther examination will be required to determine theprecise characteristics of these two populations ofmesoprefrontal cortical DA neurons.
The total number of triple-labeled cells (DA neu-rons expressing Fos retrogradely labeled from the PFC)observed did not equal the number of the double-labeled (Fos-TH) neurons in the VTA after the same30 min restraint stress. While there are a number ofpossible explanations for this observation, the mostparsimonious explanation is that stress increases Fosexpression in DA neurons that innervate both the PFCand other targets. The distribution of double-labeledFos-TH neurons observed after stress resembles thedistribution of A10 DA neurons that innervate thePFC, but also resembles somewhat the distribution ofA10 neurons that project to the amygdala and ento-rhinal cortex (Swanson, 1982). The response charac-teristics of the DA innervation of the amygdala tostress have varied across studies (Herman et al., 1982;Roth et al., 1988), perhaps as a result of the difficultiesin accurate dissection of the nuclei that comprise thisstructure (Kilts et al., 1988). The effects of stress onDA release in the entorhinal cortex remain to be de-termined. Our data are consistent with the metabolicactivation of DA neurons that project to the PFC andnon-PFC sites; the amygdala and entorhinal cortexare potential targets of those DA neurons in whichFos is expressed but that do not project to the PFC.
The distribution of Fos-TH neurons after stress isstrikingly similar to the distribution of ventral mes-encephalic cells that express high levels of prepro-neurotensin mRNA (Jayaraman et al., 1990). The ma-jority, if not all, of the DA neurons that project to thePFC contain neurotensin (Seroogy et al., 1987, 1988;Studler et al., 1988). It is possible that the stress-elicited increase in Fos protein expression may berestricted to the colocalized neurotensin/DA neuronsinnervating the PFC. Since the release of neurotensinfrom coexistent neurotensin/DA fibers in the PFC doesnot occur at basal firing frequencies of these meso-cortical neurons, but only at higher firing rates (Beanand Roth, 1991), a stress-elicited increase in the firingrate of mesoprefrontal cortical DA neurons may beassociated with an increase in peptide release in thePFC.
It is of interest to note that the distribution of VTAFos-DA neurons is also quite similar to the localiza-tion of DA neurons that are densely immunoreactivefor the glucocorticoid receptor (Harfstrand et al.,1986). In light of the long-known association betweenadrenal glucocorticoids and stress, and since recentdata indicate that the glucocorticoid response ele-
ment confers positive or negative glucocorticoid reg-ulation, depending on the relative amounts of Fosand Jun present (Diamond et al., 1990), it will be ofinterest to determine if the medial A10 neuronsstrongly reactive for the glucocorticoid receptor arethose that respond to stress by increasing Fos ex-pression.
Presynaptic Regulation of Stress-induced DARelease from Mesolimbic DA NeuronsRestraint stress for 30 min significantly increasedDOPAC concentrations in the NAS. However, retro-grade tracer studies revealed that stress did notincrease Fos protein expression in midbrain DA neu-rons projecting to the NAS. The present data are there-fore consistent with a lack of somatodendritic meta-bolic activation of the mesoaccumbens andmesostriatal DA neurons, such that Fos expression isnot enhanced in NAS DA neurons. In these neurons,terminal DA release may occur through presynapticmechanisms. Fos studies do not allow conclusions tobe drawn from negative results alone. The presentdata do suggest, however, that the stress-evoked re-lease of DA in the NAS may occur via presynapticmechanisms; this hypothesis is consistent with otherdata indicating that the stress-induced release of DAfrom NAS terminals occurs through presynaptic mech-anisms.
The firing rates of VTA DA neurons that project tothe NAS and striatum are not altered by stress (Streck-er and Jacobs, 1985; Jacobs, 1987; Mantz et al., 1989).However, stress clearly evokes DA release in subcor-tical sites provided that the stressor is of sufficientintensity or duration Antelman et al., 1988; Dunn,1988; Roth et al., 1988; Abercrombie et al., 1989). Ithas therefore been suggested that stress may evokeDA release in the subcortical sites through presyn-aptic mechanisms (Deutch and Roth, 1990). Gluta-mate (Glu) increases striatal DA metabolism and re-lease via a presynaptic mechanism (Chesselet et al.,1982; Cheramy et al., 1986; Jhamandas and Marien,1987; Barbieto et al., 1990; Lonart and Zigmond,1991a,b). Corticofugal projections are the primarysource of this Glu influence. We have recently re-ported that lesions of the PFC DA innervation increasethe sensitivity of mesolimbic DA neurons to stress,such that exposure to a mild stressor that does notaugment DA metabolism in the NAS of control ani-mals will increase accumbens DA metabolism in le-sioned rats (Deutch et al., 1990). This presumablyoccurs in response to removal of cortico-accumbensneurons from tonic DA inhibition and the resultantincreased release of NAS glutamate or related excit-atory amino acid.
Benzodiazepine Modulation of Stress-inducedAlterations in Fos Protein ExpressionThe stress-induced metabolic activation of mesotel-encephalic DA neurons can be prevented by pretreat-ment with anxiolytic benzodiazepines, such as DZ(Fadda et al., 1978; Lavielle et al., 1978; Roth et al.,1988; Scatton et al., 1988). Conversely, administration
Cerebral Cortex July/Aug 1991, V I N 4 287
at York U
niversity Libraries on July 9, 2014
http://cercor.oxfordjournals.org/D
ownloaded from

of the anxiogenic benzodiazepine inverse agonist FG7142 (Petersen and Jensen, 1984; Corda et al., 1985;File et al., 1985; Dorow et al., 1988; McGregor andAtrens, 1990) results in the metabolic activation ofthe PFC DA innervation without concurrent activationin the NAS (Tam and Roth, 1990; Knorr et al., 1989).Thus, FG 7142 elicits changes in DA systems similarto those observed following mild stress (Roth et al.,1988).
FG 7142 increased the number of VTA, but not SNor RRF, DA neurons that were Fos-li. The number ofVTA DA neurons that were Fos positive after FG 7142treatment was less than that observed in response to30 min restraint. The distribution of double-labeledFos-TH neurons in the VTA of animals receiving FG7142 differed somewhat from that observed after re-straint stress (see Fig. 5). There were significantlyfewer DA neurons in the nucleus parabrachialis ex-pressing Fos (29.9% vs. 45.4% in restrained animals),and more double-labeled cells in the caudal linearnucleus (41.8% vs. 28.7%). Thus, while there werefewer AlO DA neurons that were Fos-li after FG 7142treatment than seen after 30 min restraint (Table 1),the distribution of these neurons was less tightly re-stricted across the VTA subnuclei.
Administration of anxiolytic benzodiazepines hasbeen reported to prevent completely the stress-in-duced increase in DA metabolism in the PFC, NAS,and VTA (Fadda et al., 1978; Lavielle et al., 1978; Rothet al., 1988; Hirada et al., 1989). Pretreatment of ratswith 5.0 mg/kg DZ 30 min prior to restraint stresssignificantly decreased the number of VTA DA neu-rons that were Fos-li compared to stressed animals.However, DZ did not completely prevent a stress-elicited increase in the number of VTA DA neuronsthat expressed Fos. It is not clear if the partial ratherthan complete prevention of the stress effect is attrib-utable to the relatively severe stressor used. Previousstudies documenting the effectiveness of DZ have beenconducted using milder stressors, as reflected by thelack of an increase in striatal DOPAC concentrations.
The precise loci of action where benzodiazepinesact to prevent the stress-induced activation of themesocortical DA system are not clear (Claustre et al.,1986; Deutch and Roth, 1990). We have observed thatdirect injection of the benzodiazepine chlordiaze-poxide into the VTA partially, but not completely,prevents the stress-induced activation of the PFC DAinnervation (Deutch and Roth, 1990) and have sug-gested that benzodiazepines may act at both the cellbody and the terminal field level to prevent the in-crease in PFC DA metabolism (Knorr et al., 1989;Deutch and Roth, 1990). The partial prevention byDZ of the stress-induced increase in the number ofAlO DA neurons expressing Fos is consistent with thishypothesis. Alternatively, the ability of DZ to preventonly partially the stress-induced increase in the num-ber of AlO DA neurons expressing Fos may simplyreflect that DZ treatment alone had significant effectson Fos protein expression.
DZ significantly increased the total number of AlODA neurons expressing Fos; the total number of AlO
DA neurons that were Fos-li did not differ significantlybetween animals pretreated with DZ and exposed tostress and animals that received DZ but were notstressed. However, DZ treatment did not significantlyincrease the number of double-labeled cells in anysingle VTA subnucleus; this may be attributable tothe DZ treatment increasing variability. The reasonfor the increased number of DA neurons expressingFos protein after DZ administration is not clear. Cur-ran and Morgan (1985) noted that the induction ofc-/osin PC12 cells by NGF is significantly enhancedby benzodiazepines; however, benzodiazepines alonedid not increase c-fos expression.
One intriguing possibility to account for the DZ-induced increase in the number of DA neurons ex-pressing Fos is that the novelty of the interoceptivestate that results from the administration of DZ mayitself be a stressor or, alternatively, that novelty is atone end of a continuum extending to stress; previousreports indicate that novelty can increase PFC DAmetabolism (Tassin et al., 1980).
Time Course for Stress-induced Increase in FosExpression in AlO DA NeuronsA significant increase in the number of AlO DA neu-rons expressing Fos protein was observed in animalskilled at 4 hr 30 min, but not 2 hr 15 min, followingremoval from restraint. At the shorter time point, themean number of double-labeled VTA neurons wastwice that observed in control animals, but the vari-ability was such that it precluded statistical signifi-cance. This may simply mean that expression of de-tectable amounts of Fos protein in VTA DA neuronsbegins about 2 hr 15 min after stress termination, andthat significant changes can be observed shortly there-after.
Alternatively, our inability to detect a significantincrease in the number of AlO Fos-DA neurons at theshorter time point may suggest that the activation ofDA neurons by stress is a transsynaptic phenomenon,occurring in response to the activation of certain ex-citatory afferents to the VTA. Intracerebral transplan-tation studies of DA tissue suggest that intact afferentsto the DA neurons are required for the stress-inducedenhancement of DA metabolism (Herman etal., 1988).A number of studies have indicated that specific trans-mitter systems may subserve the stress-induced acti-vation of the PFC DA system (see Deutch and Roth,1990). Animals killed at 2 hr 15 min after stress ter-mination, while not exhibiting an increased numberof VTA DA-Fos neurons, did have an increased num-ber of Fos neurons in certain medullary and pontinenuclei that project to the VTA (Gillham et al., 1991),suggesting that the stress-elicited increase in Fos pro-tein in AlO DA neurons may be transsynaptic. Thisopens the possibility that the use of Fos immunohis-tochemistry combined with retrograde tracing mayilluminate the precise pathways through which vari-ous stressors, including those directly involving nox-ious stimuli (e.g., footshock) and those that do not(conditioned fear), gain access to midbrain DA neu-rons.
288 PFC DA Neurons Express Fos • Deutch et al.
at York U
niversity Libraries on July 9, 2014
http://cercor.oxfordjournals.org/D
ownloaded from

Finally, the increased number of Fos-DA neuronsin animals that survived for the longer period of timemay also reflect the detection of Fra by our antibody.Fra exhibit a high degree of sequence homology withFos in the leucine zipper region (Cohen and Curran,1988) through which Fos and Jun dimerize (Curranand Franza, 1988; Landschulz et a!., 1988). The timecourse of expression of Fra following various treat-ments appears to lag behind the expression of Fos(Iadarola et al., 1989; Morgan and Curran, 1989; Son-nenberg et al., 1989; Young et al., 1991). Since ourantibody recognizes both Fos and Fra, it is possiblethat the detection of Fos-li in DA neurons in thoseanimals that were killed after the longer duration re-flects the recognition of both Fos and Fra, leading toan increased ability to reach immunohistochemicaldetection threshold. Further studies utilizing eitherantibodies specific for Fos (such as those directed atthe N-terminus) or immunoblot assays will be re-quired to determine the relative involvement of Fosand Fra.
Functional ConsiderationsThe functional significance of enhanced c-fos expres-sion in certain A10 DA neurons, including those thatinnervate the PFC, is not yet known. Cellular imme-diate-early genes, including c-fos, have been ad-vanced as transduction mechanisms between stimu-lation of the neuron and adaptive neuronal responsesrequiring alterations in gene expression (Morgan andCurran, 1989). It remains to be determined whichgenes are being transcriptionally regulated by the in-crease in c-fos expression following stress.
Responses to stress involve both short- and long-term adaptive changes. The stress-elicited increase inPFC DA metabolism is of relatively short duration(Bannon et al., 1986). It is not known if there aremore enduring changes in catecholamine biosynthe-sis that allow a different DA response to subsequentstress exposure. Stress-induced alterations in TH geneexpression might be such a long-term response. Un-der normal circumstances, TH is the rate-limiting stepin DA synthesis. While short-term alterations in THare mediated by phosphorylation of the protein, lon-ger changes may involve alterations in TH gene ex-pression. Since TH transcription is induced by NGFthrough a c-ybs-dependent mechanism (Gizang-Gins-berg and Ziff, 1990), it is not unreasonable to suggestthat the stress-induced enhancement of Fos expres-sion in DA neurons may in some manner involve theinduction of TH. Alternatively, other transmitter can-didates (e.g., neurotensin or cholecystokinin), ortransmitter receptors (such as the DA D2 autorecep-tor), may be transcriptionally regulated by the in-crease in c-fos expression. At the present time, theprecise nature of the molecular adaptations occurringin DA neurons in response to stress remains unknown.Future studies will be required to define what long-term changes stress induces in DA neurons througha c-ybs-dependent mechanism.
One of the first reports concerning a functional at-tribute of the PFC dopaminergic innervation was the
observation that this system is sensitive to mild stress(Thierry et al., 1976). Since that report, numerousinvestigations have focused on the significance of themesoprefrontal cortical DA system. In particular, stud-ies in primates (including humans) have led to thespeculation that there may be functional alterationsin the PFC DA innervation that underlie certain psy-chiatric disorders, including schizophrenia (Brownet al., 1979; Weinberger, 1987). Studies in nonhumanand human primates have in particular focused on thedorsolateral convexity of the PFC and clearly indi-cated that this region is functionally distinct fromother frontal cortical areas (Goldman-Rakic, 1987).However, it has proven difficult to define a singlefunctional or anatomical homolog in the rodent tothe primate dorsolateral PFC. This is attributable inpart to the fact that the region defined as the medialPFC in the rodent is comprised of a number of distinctcytoarchitectonic regions, which have correspondingfunctional heterogeneities (Krettek and Price, 1977;Rosenkilde, 1983; Kolb, 1984; Passingham et al., 1988;Sesack et al., 1989). Biochemical and pharmacolog-ical studies have lagged behind behavioral investi-gations and have in general considered the DA in-nervation of the rodent medial PFC to be functionallyhomogeneous. However, Berger and colleagues havenoted that the DA innervation of the rodent PFC isanatomically heterogeneous and hypothesized thepresence of two mesocortical DA systems with distinctproperties (see Berger and Verney, 1984; Berger etal., 1991). There appear to be at least two distinct DAinnervations of the anteromedial PFC as defined onthe basis of the presence of impulse-modulating au-toreceptors (Shepard and German, 1988). Similarly,the DA innervations of the primate cerebral cortexappear to differ on the basis of susceptibility to theneurotoxic actions of l-methyl-4-phenyl-l,2,3,6-tetra-hydropyridine (MPTP; Elsworth et al., 1990). Thepresent data indicate that the DA innervation of thePFC is also functionally heterogeneous in respon-siveness to stress.
Over the past several years there has been a re-evaluation of the nature of information processing incorticostriatal circuits. In particular, the presence ofa parallel but segregated organization of cortico-stri-atal-thalamocortical circuits has emerged (see Gold-man-Rakic and Selemon, 1990). Both anatomical andphysiological data in the primate have supported thisview (Selemon and Goldman-Rakic, 1985; Alexanderet al., 1986; Alexander and Crutcher, 1990). It will beof importance to determine the extent of distinct DAsystems in the medial PFC of the rat, and to determineif multiple dopamine systems embedded within therat PFC form parallel functional circuits with distinctcorticofugal systems.
NotesWe wish to acknowledge the anonymous reviewer who sug-gested that the interoceptive state associated with diazepamadministration might contribute to the diazepam-inducedincrease in Fos expression in A10 DA neurons. This workwas supported by grant MH-45124 and a grant from the Scot-tish Rite Schizophrenia Research Program to A.Y.D., by the
Cerebral Cortex July/Aug 1991, V 1 N 4 289
at York U
niversity Libraries on July 9, 2014
http://cercor.oxfordjournals.org/D
ownloaded from

National Parkinson Foundation Research Laboratories at YaleUniversity, and by the Post-Traumatic Stress Disorder Centerand the Schizophrenia Research Center of the West HavenVeteran's Administration Medical Center.
Correspondence should be addressed to Ariel Y. Deutch,Department of Psychiatry, Yale University School of Medi-cine, CMHC, 34 Park Street, New Haven, CT 06508.
ReferencesAbercrombie ED, Keefe KA, DiFrischia DS, Zigmond MJ
(1989) Differential effects of stress on in wuo dopaminerelease in striatum, nucleus accumbens, and medial fron-tal cortex. J Neurochem 52:1655-1658.
Alexander GE, Crutcher MD (1990) Functional architec-ture of basal ganglia circuits: neural substrates of parallelprocessing. Trends Neurosci 13:266-271.
Alexander GE, DeLong MR, Strick PL (1986) Parallel or-ganization of functionally segregated circuits linking bas-al ganglia and cortex. Annu Rev Neurosci 9:357-381.
Antelman SM, Knopf S, Caggiula AR, Kocan D, Lysle DT,Edwards DJ (1988) Stress and enhanced dopamine uti-lization in the frontal cortex: the myth and the reality.Ann NY Acad Sci 537:262-272.
Bannon MJ, Deutch AY, Tarn S-Y, Zamir N, Eskay RL, LeeJ-M, Maggio JE, Roth RH (1986) Mild footshock stressdissociates substance P from substance K and dynorphinfrom Met- and Leu-enkephalin. Brain Res 381:393-396.
Barbieto L, Cheramy A, Godeheu G, Desce JM, Glowinski J(1990) Glutamate receptors of a quisqualate-kainatesubtype are involved in the presynaptic regulation ofdopamine release in the cat caudate nucleus in vivo. EurJ Neurosci 2:304-311.
Bean AJ, Roth RH (1991) Extracellular dopamine and neu-rotensin in rat prefrontal cortex in vivo-, effects of medianforebrain bundle stimulation frequency, pattern, and do-pamine autoreceptors. J Neurosci, in press.
Berger B, Verney C (1984) Development of the catechol-amine innervation in rat neocortex: morphological fea-tures. In: Monoamine innervation of cerebral cortex(Descarries L, Reader TR, and Jasper HH, eds), pp 95-121. New York: Liss.
Berger B, Gaspar P, Verney C (1991) Dopaminergic in-nervation of the cerebral cortex: unexpected differencesbetween rodents and primates. Trends Neurosci 14:21-27.
Brown R, Crane A, Goldman P (1979) Regional distributionof monamines in the cerebral cortex and subcorticalstructures of the rhesus monkey: concentrations and invivo synthesis rates. Brain Res 168:133-150.
BullittE (1990) Expression ofc-/os-like protein as a markerfor neuronal activity following noxious stimulation in therat. J Comp Neurol 296:517-530.
Ceccatelli S, Villar MJ, Goldstein M, Hokfelt T (1989) Ex-pression of c-Fos immunoreactiviry in transmitter-char-acterized neurons after stress. Proc Natl Acad Sci USA 86:9569-9573.
Cheramy A, Romo R, Godehu G, Baruch P, Glowinski J(1986) In vivo presynaptic control of dopamine releasein the cat caudate nucleus—II. Facilitatory or inhibitoryinfluence of L-glutamate. Neuroscience 19:1081-1090.
Chesselet MF (1984) Presynaptic regulation of neuro-transmitter release in the brain: facts and hypotheses.Neuroscience 12:347-375.
Chesselet M-F, Cheramy A, Reisine T, Lubetski L, GlowinskiJ (1982) Presynaptic regulation of striatal dopaminerelease: in vivo and in vitro studies. J Physiol (Paris) 78:420.
Claustre Y, Rivy JP, Dennis T, Scatton B (1986) Pharma-cological studies on stress-induced increase in frontalcortical dopamine metabolism in the rat. J PharmacolExp Ther 238:693-700.
Cohen DR, Curran T (1988) fra-l: a serum-inducible, cel-lular immediate-early gene that encodes a Fos-relatedantigen. Mol Cell Biol 8:2063-2069.
CordaMG,GiorgiO,GattaF, BiggioG (1985) Long-lastingproconflict effect induced by chronic administration ofthe 0-carboline derivative FG 7142. Neurosci Lett 62:237-240.
Curran T, Franza BR Jr (1988) Fos and Jun: the API con-nection. Cell 55:395-397.
Curran T, Morgan JI (1985) Superinduction of cfos bynerve growth factor in the presence of peripherally activebenzodiazepines. Science 229:1265-1268.
Deutch AY, Roth RH (1990) The determinants of stress-induced activation of the prefrontal conical dopaminesystem. In: Progress in brain research, Vol 85, The pre-frontal cortex: its structure, function, and pathology (Uyl-ings HBM, Van Eden CG, De Bruin JPC, Corner MA, andFeenstra MPG, eds), pp 357-393, Elsevier. Amsterdam.
Deutch AY, Tarn S-Y, Roth RH (1985) Footshock and con-ditioned stress increase 3,4-dihydroxyphenylacetic acid(DO PAC) in the ventral tegmental area but not substantianigra. Brain Res 333:143-146.
Deutch AY, Goldstein M, Baldino F Jr, Roth RH (1988) Therelencephalic projections of the A8 dopamine cell group.Ann NY Acad Sci 537:27-50.
Deutch AY, Clark WA, Roth RH (1990) Prefrontal conicaldopamine depletion enhances the responsiveness of me-solimbic dopamine neurons to stress. Brain Res 521:311-315.
Diamond MI, Miner JN, Yoshinaga SK, Yamamoto KR (1990)Transcription factor interactions: selectors of positive ornegative regulation from a single DNA element. Science249:1266-1272.
Dorow R, Horowski R, Paschelke, Amin M, Braestrup C(1988) Severe anxiety induced by FG 7142, a |8-carbolineligand for benzodiazepine receptors. Lancet 2:98-99-
DragunowM.Faull R (1989) The use ofc-/osas a metabolicmarker in neuronal pathway tracing. J Neurosci Methods29:261-265.
Dragunow M, Robertson HA (1988) Brain injury inducescfos protein(s) in nerve and glia-like cells in adult mam-malian brain. Brain Res 455:295-299.
Dunn AJ (1988) Stress-related activation of cerebral do-paminergic systems. Ann NY Acad Sci 537:188-205.
ElsworthJD, Deutch AY, Redmond DEJr, SladekJRJr, RothRH (1990) MPTP reduces dopamine and norepineph-rine concentrations in the supplementary motor area andcingulate conex of the primate. Neurosci Lett 114:316-322.
Fadda F, Argiolis A, Melis MR, Tissari AH, Onali PL, GessaGL (1978) Stress-induced increase in 3,4-dihydroxy-phenylacetic acid (DOPAC) levels in the cerebral cortexand in n. accumbens: reversal by diazepam. Life Sci 23:2219-2224.
File SE, Pellow S, Braestrup C (1985) Effects of the/3-carboline FG 7142 in the social interaction test of anx-iety and the holeboard: correlations between behaviorand plasma concentrations. Pharmacol Biochem Behav22:941-944.
Gentz R, Rauscher RJ III, Abate C, Curran T (1989) Fosand Jun dimerize through a parallel leucine zipper thatjuxtaposes DNA-binding domains. Science 243:1695-1699.
Gillham MH, Jennes L, Deutch AY (1990) GABA neuronsin the ventral tegmental area project to the medial pre-frontal cortex: a non-dopaminergic mesoconical system.Soc Neurosci Abstr 16:238.
Gillham MH, Iadarola MJ, Deutch AY (1991) Stress-in-duced alterations in Fos expression in neurons inner-vating the ventral tegmental area: an approach to thedetermination of the stress circuit. Soc Neurosci Abstr,in press.
Girault JA, Barbieto L, Spampinato U, Gozlan H, GlowinskiJ, Besson MJ (1986) In vivo release of endogenousamino acids from the rat striatum: further evidence for arole of glutamate and aspartate in corticostriatal neuro-transmission. J Neurochem 47:98-106.
Gizang-Ginsberg E, Ziff EB (1990) Nerve growth factor
290 PFC DA Neurons Express Fos • Deutch et al.
at York U
niversity Libraries on July 9, 2014
http://cercor.oxfordjournals.org/D
ownloaded from

regulates styrosine hydroxylase gene transcription througha nucleoprotein complex that contains c-Fos. Genes Dev.4A77-491.
Goldman-Rakic PS (1987) Circuitry of primate prefrontalcortex and the regulation of behavior by representationalmemory. In: Handbook of physiology, Vol 5, Higher cor-tical function (Plum F, Mountcastle VB, eds), pp 373-417. Washington, DC: American Physiological Society.
Goldman-Rakic PS, Selemon LD (1990) New frontiers inbasal ganglia research. Trends Neurosci 13:241-244.
Gonzalez GA, Yamamoto KK, Fischer WH, Karr D, MenzelP, Biggs Will, Vale WW.Montminy MR (1989) Aclusterof phosphorylation sites on the cyclic AMP-regulated nu-clear factor CREB predicted by its sequence. Nature 337:749-752.
Harfstrand A, Fuxe K, Cintra A, Agnati LF, Zini I, WikstromAC, Okret S, Yu Z-Y, Goldstein M, Steinbusch H, Ver-hofstadA, GustafssonJ-A (1986) Glucocorticoid recep-tor immunoreactivity in monoaminergic neurons of ratbrain. Proc Natl Acad Sci USA 83:9779-9783.
Herman JP, Guilloneau D, Dantzer R, Scatton B, Semerdjian-Rouquier L, Le Moal M (1982) Differential effects ofinescapable footshocks on dopamine turnover in corticaland limbic areas of the rat. Life Sci 30:2207-2214.
Herman JP, Rivet JM, Abrous N, Le Moal M (1988) Intra-cerebral dopaminergic transplants are not activated byelectrical footshock stress activating in situ mesocorti-colimbic neurons. Neurosci Lett 90:83-88.
Herrera DG, Robertson HA (1990) Application of potas-sium chloride to the brain surface induces the c-fos proto-oncogene: reversal by MK-801. Brain Res 510:166-170.
Hirada K, Deutch AY, Goldstein M (1989) The effects ofa benzodiazepine partial agonist, R0 16-6028, on basaland stress-induced in vivo tyrosine hydroxylation in themesotelencephalic dopamine system. Soc Neurosci Abstr15:1310.
Hisanaga K, Sagar SM, Hicks KJ, Swanson RA, Sharp FR(1990) c-Fos proto-oncogene expression in astrocytesassociated with differentiation or proliferation but notdepolarization. Mol Brain Res 8:69-75.
Hunt SP, Pini A, Evan G (1987) Induction of c-fos-Hkeprotein in spinal cord neurons following sensory stim-ulation. Nature 328:632-634.
Iadarola MJ, Yeung CL, Draisci G, Ruda MA (1989) Pro-longed elevation of multiple Fos immunoreactive pro-teins in rat spinal cord: Western blot and immunocyto-chemical analysis. Soc Neurosci Abstr 15:468.
Iuvone PM, Dunn AJ (1986) Tyrosine hydroxylase activa-tion in mesocortical 3,4-dihydroxy-phenylethylamineneurons following footshock. J Neurochem 47:837-844.
Jacobs BL (1987) Central monoaminergic neurons: single-unit studies in behaving animals. In: Psychopharmacol-ogy: the third generation of progress (Meltzer HY, ed),pp 159-169. New York: Raven.
Jayaraman A, Nishimori T, Dobner P, Uhl GR (1990) Cho-lecystokinin and neurotensin mRNAs are differentiallyexpressed in subnuclei of the ventral tegmental area. JComp Neurol 296:291-302.
Jhamandas K, Marien M (1987) Glutamate-evoked releaseof endogenous brain dopamine: inhibition by an excit-atory amino acid antagonist and an enkephalin analogue.Br J Pharmacol 90:641-650.
KalivasPW, Taylor A (1985) Behavioral and neurochemicaleffect of daily injection with neurotensin into the ventraltegmental area. Brain Res 358:70-76.
Keller RW Jr, Strieker EM, Zigmond MJ (1983) Environ-mental stimuli but not homeostatic challenges produceapparent increases in dopaminergic activity in the stria-turn: an analysis by in vivo voltammetry. Brain Res 279;159-170.
Kilts CD, Anderson CM, Ely TD, Mailman RB (1988) Thebiochemistry and pharmacology of mesoamygdaloid do-pamine neurons. Ann N Y Acad Sci 537:173-187.
Knorr AM, Deutch AY, Roth RH (1989) The anxiogenic/3-carboline FG 7142 increases in vivo and in vitro ty-
rosine hydroxylation in the prefrontal cortex. Brain Res495:355-361.
Kolb B (1984) Functions of the frontal cortex of the rat: acomparative review. Brain Res Rev 8:65-98.
KrettekJE, Price JL (1977) The cortical projections of themediodorsal nucleus and adjacent thalamic nuclei in therat. J Comp Neurol 171:157-192.
Landschulz WM, Johnson PF, Adashi EY, Graves BJ, Mc-Knight S (1988) The leucine zipper: a hypotheticalstructure common to a new class of DNA-binding pro-teins. Science 240:1759-1764.
Lavielle S, Tassin JP, Thierry AM, Blanc G, Herve D, Bar-thelemy C, Glowinski J (1978) Blockade by benzodi-azepines of the selective high increase in dopamine turn-over induced by stress in mesocortical dopaminergicneurons of the rat. Brain Res 168:585-594.
LeDoux JE, Sakaguchi A, IwataJ, Reis DJ (1986) Interrup-tion of projections from the medial geniculate body toan archistriatal field disrupts the classical conditioningof emotional responses to acoustic stimuli. Neuroscience17:615-627.
Lee W-S, Smith S, Hoffman GW (1990) Luteinizing hor-mone-releasing hormone neurons express Fos proteinduring the proestrous surge of luteinizing hormone. ProcNatl Acad Sci USA 87:5163-5167.
Lonart G, Zigmond MJ (1991a) Dopamine release can beevoked from striatal slices via reverse transport by highglutamate concentrations. J Pharmacol Exp Ther 256:1132-1138.
Lonart G, Zigmond MJ (1991b) L-Glutamic acid evokesCa2+independent release of dopamine from rat striatumvia dopamine uptake system. Ann NY Acad Sci, in press.
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ 1951) Pro-tein measurement with the Folin phenol reagent. J BiolChem 193:265-275.
MantzJ, Thierry AM, Glowinski J (1989) Effect of noxioustail pinch on the discharge rate of mesocortical and me-solimbic dopamine neurons: selective activation of themesocortical system. Brain Res 476:377-381.
McGregor IS, Atrens DM (1990) Stressor-like effects of FG-7142 on medial prefrontal cortex self-stimulation. BrainRes 516:170-174.
Montminy MR, Gonzalez GA, Yamamoto KK (1990) Reg-ulation of cAMP-inducible genes by CREB. Trends Neu-rosci 13:184-188.
Morgan JI, Curran T (1986) The role of ion flux in thecontrol of c-fos. Nature 322:552-555.
Morgan JI, Curran T (1989) Stimulus-transcription cou-pling in neurons: role of cellular immediate-early genes.Trends Neurosci 12:459-462.
Morgan JI, Cohen DR, HempsteadJL, Curran T (1987) Map-ping patterns of c-fos expression in the central nervoussystem after seizure. Science 237:192-197.
Mugnaini E, Berrebi AS, Morgan JI, Curran T (1989) Fos-like immunoreactivity induced by seizure in mice spe-cifically associated with euchromatin in neurons. Eur JNeurosci 1:46-52.
Passingham RE, Myers C, Rawlins N, Lightfoot V, Fearn S(1988) Premotor cortex in the rat. Behav Neurosci 102:101-109.
Paxinos G, Watson C (1982) The rat brain in stereotaxiccoordinates. Sydney: Academic.
Petersen EN, Jensen LH (1984) Proconflict effect of ben-zodiazepine receptor inverse agonists and other inhibi-tors of GABA function. EurJ Pharmacol 103:91-97.
Quinn JP, Takimoto M, Iadarola MJ, Holbrook N, Levens D(1989) Distinct factors bind the API consensus sites ingibbon ape leukemia virus and simian virus 40 enhanc-ers. J Virol 63:1737-1742.
ReinhardJF, Bannon MJ, Roth RH (1982) Acceleration bystress of dopamine synthesis and metabolism in prefron-tal cortex: antagonism by diazepam. Naunyn Schmiede-bergs Arch Pharmacol 318:374-377.
Robertson GS, Herrera DG, Dragunow M, Robertson HA(1989) L-DOPA activates c-fos in the striatum ipsilateral
Cerebral Cortex July/Aug 1991, VI N 4 291
at York U
niversity Libraries on July 9, 2014
http://cercor.oxfordjournals.org/D
ownloaded from

to a 6-hydroxydopamine lesion of the substantia nigra.EurJ Pharmacol 159:99-100.
Rosenkilde CE (1983) Functions of the prefrontal cortex:behavioral investigations using ablation and electro-physiological techniques in rats, cats, dogs, and mon-keys. Acta Physiol Scand [Suppl] 514:4-58.
Roth RH, Tam S-Y, Ida Y, Yang J-X, Deutch AY (1988)Stress and the mesocorticolimbic dopamine systems. AnnNYAcad Sci 537:138-147.
Sagar SM, Sharp FR, Curran T (1988) Expression of c/osprotein in brain: metabolic mapping at the cellular level.Science 240:1328-1331.
Scatton B, D'Angio M, Driscoll P, Serrano A (1988) An invivo voltammetric study of the response of mesocorticaland mesoaccumbens dopaminergic neurons to environ-mental stimuli in strains of rats with differing levels ofemotionality. Ann NY Acad Sci 537:124-137.
Selemon LD, Goldman-Rakic PS (1985) Longitudinal to-pography and interdigitation of corticostriatal projec-tions in the rhesus monkey. J Neurosci 5:776-794.
Semenenko FM, Cuello AC, Goldstein M, Lee KY, Sidebot-tom E (1986) A monoclonal antibody against ryrosinehydroxylase: application in light and electron micros-copy. J Histochem Cytochem 4:817-821.
Seroogy K, Mehta A, Fallon JH (1987) Neurotensin andcholecystokinin coexistence within neurons of the ven-tral mesencephalon: projections to forebrain. Exp BrainRes 68:277-289.
Seroogy K, Ceccatelli S, Schalling M, Hokfelt T, Frey P, WalshJ, Dockray G, Brown J, Buchan A, Goldstein M (1988)A subpopulation of dopaminergic neurons in rat ventralmesencephalon contains both neurotensin and chole-cystokinin. Brain Res 455:88-98.
Sesack SR, Deutch AY, Roth RH, Bunney BS (1989) To-pographical organization of efferent projections of themedial prefrontal cortex in the rat: an anterograde tract-tracing study with Phaseolus vulgaris leucoagglutinin. JComp Neurol 290:213-242.
Sheng M, Greenberg ME (1990) The regulation and func-tion of c-fos and other immediate early genes in the ner-vous system. Neuron 4:477-485.
Sheng M, McFadden G, Greenberg ME (1990) Membranedepolarization and calcium induce c-fos transcription viaphosphorylation of transcription factor CREB. Neuron 4:571-582.
Shepard PD, German DC (1988) Electrophysiological andpharmacological evidence for the existence of distinctpopulations of nigrostriatal dopaminergic neuron in therat. Neuroscience 27:537-546.
Sonnenberg JL, Macgregor-Leon P, Curran T, Morgan JI(1989) Dynamic alterations occur in the levels and com-position of transcription factor API complexes after sei-zure. Neuron 3:359-365.
Strecker RE, Jacobs BJ (1985) Substantia nigra dopaminer-gic unit activity in behaving cats: effect of arousal onspontaneous discharge and sensory evoked activity. BrainRes 361:339-350.
Studler JM, Kitabgi P, Tramu G, Herve D, Glowinski J, TassinJP (1988) Extensive colocalization of neurotensin withdopamine in rat meso-cortico-frontal dopaminergic neu-rons. Neuropeptides 11:95-100.
Swanson LW (1982) The projections of the ventral teg-mental area and adjacent regions: a combined retrogradetracer and immunofluorescence study in the rat. BrainRes Bull 9:321-353.
Tam S-Y, Roth RH (1990) Modulation of mesoprefrontaldopamine neurons by central benzodiazepine receptors.I. Pharmacological characterization. J Pharmacol ExpTher252:989-996.
Tassin JP, Herve D, Blanc G, Glowinski J (1980) Differ-ential effects of a two minute open field session on do-pamine utilization in the frontal cortices of BALB/C andC57 BL/6 mice. Neurosci Lett 17:67-71.
Thierry AM, Tassin JP, Blanc G, Glowinski J (1976) Selec-
tive activation of the mesocortical DA system by stress.Nature 263:242-244.
Trulson ME, Preussler DW (1984) Dopamine containingventral tegmental area neurons in freely moving cats:activity during the sleep-waking cycle and effects of stress.Exp Neurol 83:367-377.
Young ST, Porrino LJ, Iadorola MJ (1991) Cocaine inducesstriatal Fos immunoreactive proteins via dopaminergicD, receptors. Proc Natl Acad Sci USA 88:1291-1295.
Weinberger DR (1987) Implications of normal brain de-velopment for the pathogenesis of schizophrenia. ArchGen Psychiatry 44:660-669.
292 PFC DA Neurons Express Fos • Deutch et al.
at York U
niversity Libraries on July 9, 2014
http://cercor.oxfordjournals.org/D
ownloaded from