skripta biogeografija 2013
TRANSCRIPT

Biogeografija
1. Uvodni dio
Biogeografija je znanost koja se bavi proučavanjem rasprostranjenosti biljnih i životinjskih vrsta na Zemljinoj površini, iz čega proizlazi da biogeografija ima za cilj otkriti gdje pojedini organizmi žive, utvrditi njihovu brojnost, te proučiti osnovne zemljopisne i povijesne okolnosti koji su doveli do toga da se pojedina vrsta javlja u određenom području. Rasprostranjenost biljnih/životinjskih vrsta najčešće se promatra od regionalnih do kontinentskih razmjera. Uobičajeni primjeri rasprostranjenosti vrsta na svim tim razinama mogu se objasniti spletom povijesnih okolnosti odnosno ukupnim utjecajem povijesnih čimbenika na dotičnom prostorom, kao što su: (i) specijacija1 , (ii) izumiranje2, (iii) pomicanje kontinenata3, (iv) glacijacija4, i s time u vezi podizanje i spuštanje razine mora, promjene toka rijeka i promjene ekoloških čimbenika koji vladaju na nekom području (obitavalištu) s jedne strane, i izolacijom određenog kopnenog područja, povezanog s dostupnim protokom i zalihom energije s druge strane.
1 Specijacija je evolucijski proces kojim su nastale nove biološke vrste. 2 Izumiranje, u biologiji i ekologiji, predstavlja nestanak određenog organizma ili skupine organizama (svojti), odnosno određene vrste. Općenito se trenutkom izumiranja smatra ugibanje posljednje jedinke određene vrste, iako je nestanak dotične vrste uslijedio znatno ranije, prije ove točke. 3 Vidi, Teorija o jedinstvenom kopnu. 4 Glacijacija (glacijalno razdoblje) je vremenski interval (tisuće godina) unutar ledenog doba koje je obilježeno hladnijim temperaturama i širenjem ledenjaka. S druge pak strane, Interglacijal (ili, interglacijalno razdoblje) su razdoblja toplije klime. 5U biologiji refugija (množina: refugie), ponekad jednostavno nazivamo utočište odnosno mjesto izoliranih i/ili reliktnih populacija koje su nekada bile široko rasprostranjene.

Tijekom razdoblja ekoloških promjena na Zemlji, biogeografija uključuje proučavanje biljnih i životinjskih vrsta kroz njihovu prošlost i/ili sadašnjost ako žive u refugijima 5 , odnosno proučava njihova staništa ili obitavališta gdje su sve opstale. Prema tome, biogeografija općenito čini više od pukog postavljanja pitanja koja vrsta i gdje?, nego također nudi pitanje i zašto?, a što je ponekad i od presudne važnosti. Suvremena biogeografija često se koristi geografskim informacijskim sustavom (GIS-om), kako bi razjasnila sve čimbenike koji utječu na rasprostranjenost organizama, kao i predvidjela buduće trendove u rasprostranjivanju istih. GIS koristi i matematičke modele u rješavanju ekoloških problema, prije svega s prostornog aspekta
. Slika 1. Specijacije: a) Alopatrička (od grčkog Allos, "drugi" + grčki patrida, "domovina") ili zemljopisna specijacija je specijacija koja se javlja kada biološke populacije iste vrste postaju izolirane zbog neke geografske promjene/prepreke (nastajanje rasjeda, planinskih lanaca, društvene promjene, kao što je iseljavanje i slično). Izolirane populacije zatim su podvrgnute genotipskim i / ili fenotipskim razlikama kao što je (a) selektivno izlaganje različitim pritiscima; npr. prilagodba jedinki na način ishrane, (b) razvoj neovisne genetske tendencije, i (c) različite mutacije u populacije gena. b) Simpatrička specijacija je proces kroz koji su nove vrste evoluirale iz jedne prethodne vrste predaka, a pri tom su nastanjivale isto geografsko područje.
U svojim zaključcima, biogeografija se u većini slučajeva oslanja na proučavanje i promatranje na razini otoka ili skupine otoka. Otočnim područjima/staništima mnogo je lakše upravljati i istraživati, jer su prostorno jasno ograničeni i odvojeni od većih ekosustava na kopnu. Otoci

Slika 2. Dodo (Raphus cucullatus), izumrla vrsta pregledaju staništa te lakše utvrde koja nova vrsta ili vrste su tek nedavno kolonizirale otok, te se nadalje može lakše pratiti kako se pojedina vrsta rasprostranjuje diljem otoka. Proučavanjem određenih otoka kao staništa za biljke i životinje postignuti su mnogi uspjesi koji su se kasnije mogli primijeniti na slična kopnena staništa. Isto tako, otoci mogu biti vrlo različiti u svojim biomima, jer ih nalazimo sve od tropske do arktičke klime. Ovakva stanišna bioraznolikost omogućuje proučavanje vrlo širokog broja i raspona biljnih i životinjskih vrsta u različitim dijelovima svijeta. Jedan od prvih znanstvenik koji je prepoznao važnost tih lokacija bio značajna evolucijska odstupanja u različitih vrsta roda zeba, zapravo su doprinijeli razvoju je Charles Darwin. Nakon bioloških istraživanja otočne skupine Galapagos,

koja su uključivala

Slika 3. Teorija o jedinstvenom kopnu Slika. 4. Glacijacijacijska i interglacijacijska razdoblja na Zemlji. Posljednje ledeno doba (glacijal) je završilo pred otprilike 15.000 godina. Holocenska epoha sada je interglacijal. (Inače, holocen je geološka epoha odnosno razdoblje kvartara koja je započela prije 11.700 godina i nastavlja se do danas).

2. Povijest biogeografije
Prije objavljivanja knjige „Teorija otočne biogeografije autora MacArthur i Wilson, (1967) (koja predstavlja proširenu radnju na istu temu iz 1963.), biogeografija kao znanstvena disciplina uglavnom je promatrana kao povijesna tema, te je kao takva uglavnom bila čisto deskriptivnog karaktera. Međutim, Mac Arthur i Wilson su svojim radovima promijenili to shvaćanje te su pokazali kako se bogatstvo nekog prostora određenim biljnim i životinjskim vrstama može predvidjeti u opsegu različitih čimbenika, kao što su povijesne okolnosti, stanište, stopa imigracije i stopa izumiranja. Sve navedeno je u novije vrijeme povećalo zanimanje za otočnu biogeografiju. Primjena teorije otočne biogeografije na pojedine dijelove staništa potaknula je razvoj novih znanstvenih područja kao što su: (i) održiva biologija i (ii) krajobrazna biologija. Razvoj klasične biogeografije posebno je uslijedio primjenom dostignuća u području molekularne sistematike. S razvojem molekularne sistematike znanstvenicima je omogućeno testiranje teorije o podrijetlu i disperziji populacija (kao npr. nekih endema). Dok je primjerice klasična biogeografija mogla samo nagađati o podrijetlu vrsta na Havajskom otočju, molekularna sistematika je omogućila znanstvenicima da testiraju teorije o srodstvenim odnosima populacija s pretpostavljenim izvorima tih populacija u Aziji i Sjevernoj Americi. Neke osnovne postavke u biogeografiji uključuju:
• evoluciju (promjena genetskog sastava populacija)
• izumiranje (nestajanje vrsta)
• disperzija (udaljavanje populacija izvan točke njihova podrijetla; disperzija je srodna migraciji).
Općenito, možemo istaknuti da je biogeografija sinoptička znanost usko povezana s:
• geografijom,
• biologijom,
• geologijom,
• klimatologijom,
• ekologijom i
• evolucijom.
3. Povezanost biogeografije s drugim prirodnim znanostima

3.1. Geografija Geografija (grč. γεωγραφία) je znanost koja proučava prostornu stvarnost Zemljine površine. Prema klasičnom starogrčkom nazivu, geografija nije bila znanost s određenim objektom i ciljem proučavanja, već neka vrsta opće enciklopedije o Zemlji. Riječ geografija prvi je skovao Eratosten oko 200. god. pr. Kr. kako bi označio opisno učenje o Zemlji. Uz filozofiju i povijest, geografija spada među najstarije znanosti ljudske civilizacije. Za objašnjavanje geografskog prostora, geografija se koristi metodama prirodnih i društvenih znanosti. S obzirom na raznolikost geografskog prostora, geografija je kao znanost vrlo kompleksna. Za geografiju se može reći da je dualna ili mosna znanost, koja povezuje prirodne i društvene znanosti. Još se naziva i svjetskom disciplinom. Geografija je, dakle, sintetička znanost, koja spaja mnoga dostignuća brojnih znanosti radi objašnjavanja geografskog prostora. Ona pripada u skupinu prirodnih znanosti jer joj je osnovno polazište u prirodnim značajkama geoprostora. Geografija kao znanost o geografskom prostoru (Zemljinoj površini) objašnjava raširenost, utjecaje i međuzavisnost svih najvažnijih prirodnih i društvenih čimbenika, koji sudjeluju u oblikovanju geoprostora kao cjeline ili njegovih prostornih dijelova ili regija6 .
6 Regija je bilo koje, manje-više jasno razgraničeno zemljopisno područje ili površina neke zemlje ili kontinenta, definirana svojom geografijom, kulturom i poviješću. Regija kao pojam se koristi kao upravna jedinica u političkoj geografiji.
Slika 5. Karta Svijeta
Regija je bilo koje, manje-više jasno razgraničeno zemljopisno područje ili površina neke zemlje ili kontinenta, definirana svojom geografijom, kulturom i poviješću. Regija kao pojam se koristi kao upravna jedinica u političkoj geografiji

3.2. Biologija
Biologija je nastala od grčkog bios = život i logos = znanost. To je znanstvena disciplina koja se bavi proučavanjem živoga svijeta. Biologija proučava svojstva i ponašanje organizama, njihov rast, razvoj i razmnožavanje, interakcije između različitih organizama, te organizama i njihovog okoliša. Biologija obuhvaća široki spektar znanstvenih područja koja se često smatraju različitim disciplinama. Te discipline zajedno proučavaju život na različitim razinama. Molekularna biologija, biokemija i molekularna genetika su velikim dijelom znanstvene discipline koje se preklapaju, a proučavaju životne procese na molekularnoj razini. Stanična biologija proučava život na razini pojedinačnih stanica. Anatomija, histologija i fiziologija proučavaju finu građu i funkciju organizama na razini tkiva, organa i organskih sustava. Na Zemlji postoji više od 10 milijuna različitih vrsta. Njihova veličina pokriva raspon od onih koje su mikroskopski malene pa do organizama veličine plavog kita. Svi oblici života imaju izvjesne značajke koje ih čine specifičnim. Te značajke su na primjer mogućnost reprodukcije, rasta, ali i mogućnost prilagođavanja okolini.
3.3. Geologija
Geologija je znanost koja se bavi proučavanjem Zemlje, a nastala je od grčkih riječi γη (Gea = Zemlja) i λογος (logos = znanost). Geologija pokušava objasniti kako je Zemlja formirana i kako se mijenja. Znanstvenici koji se time bave zovu se geolozi, a proučavaju tlo, stijene, planine, oceane i druge dijelove Zemlje. Također, geologija može obuhvatiti proučavanje građe drugih (terestričkih) planeta, pa se naziv može proširiti na pojam planetarne geologije. Povijest geologije. Spoznaje o vrlo složenoj problematici postanka i razvitka Zemlje javile su se postupno, a neke datiraju još iz antičkih vremena. No, tek u 15.st. dolazi do pokušaja sistematizacije znanja o Zemlji, a postupno se javljaju i novi pojmovi kao temelj geologije u nastajanju. Tako su na primjer u Srednjem vijeku ostaci izumrlih organizama najčešće smatrani "igrom prirode" ili dokazima "općeg potopa". No, već je Leonardo da Vinci (1452 - 1519) upozorio da jednim se "potopom" ne može objasniti rasprostranjenost fosilnih ostataka morskih organizama na kopnu. Osim toga, on je bio svjestan dugog trajanja

Slika 6. Geološka karta EuropeSlika 6. Geološka karta Europe.
geološke prošlosti, a opisao je i prvi geo-kemijski ciklus (voda ispire sol iz tla i odnosi je u more koje se tako zaslanjuje, a zbog izdizanja morskog dna stvaraju se lagune gdje se voda isparuje i taloži novi sloj, koji opet može biti potopljen...). Leonardo da Vinci je shvatio i odnos erozije tla i izdizanja

kopna (erozija narušava ravnotežu u litosferi, a ona se ponovno uspostavlja izdizanjem). Širi interes za geološku problematiku izazvale su rasprave između tzv. neptunista i plutonista. Neptunisti, na čelu s A. G. Wernerom su oživili staru ideju Talesa iz Mileta (7/6.st.pr.Kr.), pripisujući postanak stijena litosfere - vodi. Zbog toga su i dobili naziv prema antičkom bogu oceana Neptunu. Plutonisti, na čelu s J. Hutton su oživili zapažanje Strabona (1.st.pr.Kr.) držeći da su pojedine stijene nastale u vezi s vulkanskim erupcijama. Nazvani su po bogu podzemlja, Plutonu. H. B. de Saussure, (18.st.) prvi je shvatio da su nagnuti slojevi zemljine kore posljedica kretanja litosfere i prodora starijih stijena kroz mlađe. E. de Beaumont (19.st.) prvi spoznaje ulogu rasjeda u postanku doline Rajne, a tvrdi da tektonske sile nastaju zbog hlađenja Zemlje i stezanja njezina obujma. Georges Cuvier (18/19.st.) postavlja temelje znanstvenog proučavanja fosilnih ostataka organizama, a William Smith (1769-1839) ih sustavno primjenjuje za određivanje relativne starosti stijena Zemljine kore te za crtanje prve geološke karte na kojoj je prikazana površina Velike Britanije. Uočava se i lateralna varijabilnost istodobnih stijena, pa tako nastaje pojam facijesa (A. Gressly, 1838). Za geologiju je od iznimne važnosti pronalazak radioaktivnosti, tako su geolozi napokon dobili moćan alat za određivanje starosti stijena. Naime, prije toga geološka vremenska skala nije imala brojke i nije se znalo kada je koje razdoblje Zemljine prošlosti počelo ili završilo, nego su se samo određivali relativni odnosi. Starost Zemlje prvi je odredio američki geokemičar Clair Cameron Patterson, polazeći od pretpostavke da je postanak svih tijela Sunčevog sustava bio istovremen, pa je određivao starost meteorita. Njegova brojka od 4,5 milijarde godina nije se previše mijenjala niti razvojem drugih metoda. Najpoznatiji i jedini geolog koji je uzorkovao stijene izvan našega matičnog planeta jeste dr. Harrison Schmitt (rođ. 1935.). On je, uz to što je geolog, bio i astronaut, senator te do sada zadnja osobe koja je hodala Mjesecom. Pojam geosinklinale Zemljine kore dobiva na značenju 1900. g. kada ga je istakao E. Haug pri postanku lančanih planinskih sustava. Ta je hipoteza, kao previše komplicirana te manjkava dokazima i mogućnostima predviđanja, bila osuđena na propast. Geosinklinala ostaje u središtu interesa geologa sve sredine 1960-ih, kada se pokazalo da je pogrešna pa je zamijenjena puno elegantnijom teorijom tektonike ploča. Ta smjena geoloških paradigmi i prihvaćanje koncepta tektonike ploča smatra se jednim od najvećih napredaka u povijesti geologije. Napokon su geolozi dobili jednostavnu i sveobuhvatnu teoriju kojom mogu objasniti 7kao labilnog sedimentacijskog prostora, nastalog lomljenjem i savijanjem

7 Geosinklinala je Izdužen, ulegnut i labilan sedimentacijski prostor litosfere između kontinentalnih masa. U geološkoj prošlosti u geosinklinali su se nakupljali sedimenti sa susjednih kopnenih blokova, koji su se kasnije pod utjecajem orogenetskih sila nabrali u velika gorja.

gotovo sve geološke procese - od rasporeda potresa i vulkana, preko izdizanja planina sve do metamorfnih facijesa i tipova magmatizma. Najzaslužniji za razvoj teorije tektonike ploča bili su Alfred Wegener, njemački geofizičar koji je popularizirao hipotezu o pomicanju kontinenata, te Harry Hess, američki marinski geolog koji je uočio proces tzv. širenja morskog dna. Međutim, ne treba zaboraviti da su razvoju ove teorije doprinijeli mnogi znanstvenici, posebno grupa geofizičara sa sveučilišta Columbia, koja je spojila ova dva koncepta u jedinstvenu teoriju kakvu danas poznajemo. Od naših istraživača koji su u ranim fazama razvitka znanosti o Zemlji ističu se Ruđer Bošković (1711. - 1787.) koji je pisao o kompenzaciji masa unutar planina i tako postavio temelje kasnijem razvoju teorije izostazije Đuro Pilar(1846. - 1893.) raspravlja o uzrocima ledenih doba i povezuje ih s ekscentričnošću Zemljine putanje, a kasnije se bavi uzrocima potresa za koje kaže da su najuže povezni sa stvaranjem pukotina i rasjeda u Zemljinoj kori. Andrija Mohorovičić (1857. - 1936.) na osnovi potresa sa žarištem u Pokuplju (25 km južno od Zagreba) dokazao (1909. god.) da u dubini od 54 km postoji jak diskontinuitet u širenju potresnih valova, koji je po njemu dobio naziv Mohorovičićev diskontinuitet. Za razvitak geologije u nacionalnim okvirima zaslužan je Lj. Vukotinović(1813 - 1893.) potičući pisanje znanstvenih djela na narodnom jeziku što je zahtijevalo i stvaranje domaće terminologije. Profesionalne temelje nacionalnom geološkom školstvu u Hrvatskoj, ne samo u okviru visokoškolskog obrazovanja, nego i u istraživačkom pogledu, postavljaju Đuro Pilar i Dragutin Gorjanović Kramberger (1856. - 1936.).
izostazija je ravnoteža između susjedinih dijelova (blokova) kore i dijelova plašta ispod njih. Budući da su stijene kore lakše od stijena plašta, može se reći da kora 'pluta' na plaštu. Dijelovi kore se dižu ili tonu sve dok se ne uspostavi izostatska ravnoteža tj. težina potisnutog dijela plašta ne bude jednaka težini bloka kore. Takva vertikalna kretanja kore se još zove izostatsko podešavanje (engl. isostatic adjustment).
Važniji geološki principi: Nekada općepoznati princip uniformitarianizma je ključ za rješavanje nekih pitanja geološke prošlosti. Glasi: "Sadašnjost je ključ za prošlost" što bi značilo da procesi koji vladaju danas su vladali i u prošlosti te promatranjem današnjih zbivanja možemo zaključivati o geološkoj prošlosti. Međutim, taj princip treba uzeti s krajnjom opreznošću s obzirom na činjenicu da se neki procesi u nekim razdobljima geološke prošlosti nisu zbivali istim intenzitetom kao danas, te da neki procesi koji su djelovali u geološkoj prošlosti ne djeluju danas

(npr. diferencijacija Zemlje). Zbog toga je danas u geološkoj zajednici prihvaćen blaži oblik uniformitarianizma - aktualizam, čiji se princip može sažeti u: "Prirodni zakoni koji vladaju danas, vladali su i u geološkoj prošlosti."

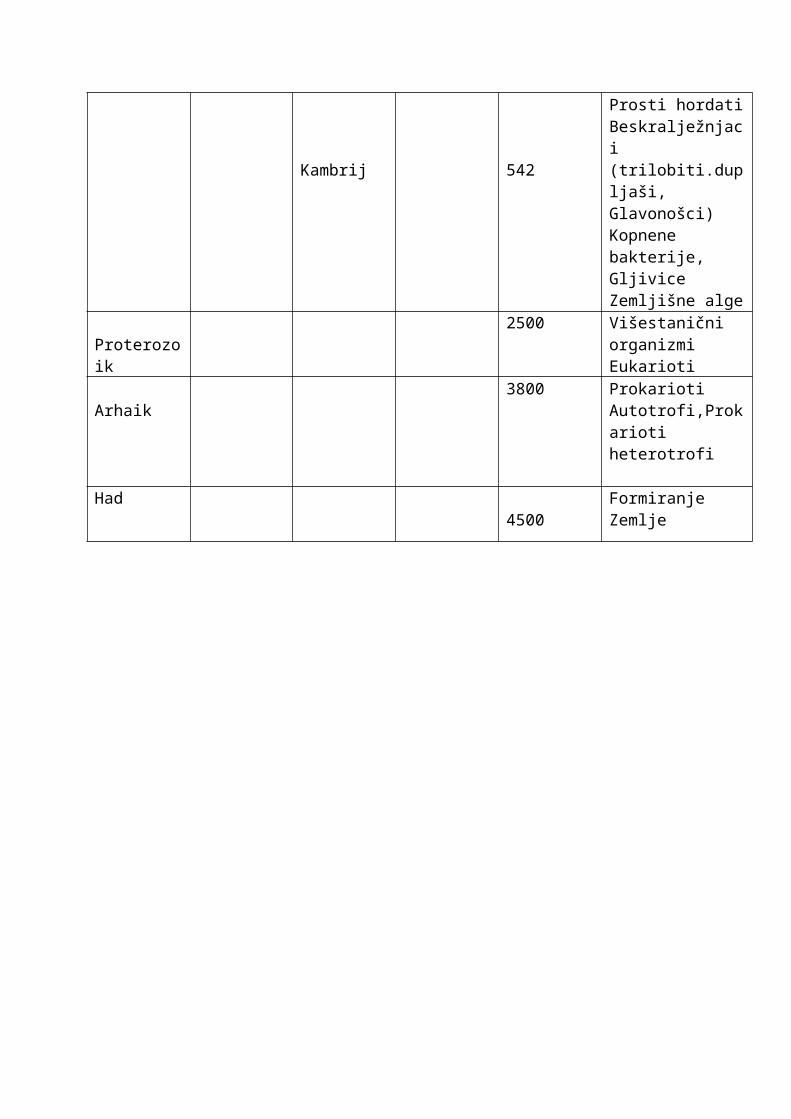
Eon era razdoblje Ep0ha P0četak D0gađaji
Fanerozoik Kenozoik kvartar holocen 0,01 Kulturna revolucijapleistocen 1,87 Čovjek
Tercijar
Pliocen 5HominidiHominoidiMastodonti
Miocen 24 MoropusOligocen 38 Ptice trkačice
EocenDivergencija sisavaca
paleogen Placentalni sisavci
Mezozoik
kreda 145 Nestanak dinosauraSkrivenosjemenjače
Jura 200 PticeDinosauri
Trijas 251 Aplacentalni sisavciGolosjemenjače
Paleozoik
Perm 299 Nestanaktrilobita
Karbon 318 GmazoviCikade
devon 359
InsektiVodozemciRibe i košljoribepaprati
Silur 418Hrskavičave ribePrve kopnene vaskularne biljke
Ordovicij 488 Mahovine
Kambrij 542
Prosti hordatiBeskralježnjaci(trilobiti.dupljaši,Glavonošci)Kopnene bakterije,GljiviceZemljišne alge
Proterozoik2500 Višestanični
organizmiEukarioti
Arhaik3800 Prokarioti
Autotrofi,Prokariotiheterotrofi
Had4500
Formiranje Zemlje

3.4. Klimatologija Klimatologija je znanost o klimi. Bavi se proučavanjem opći značajki klime raznih dijelova Svijeta, u prvom redu klasifikacijama klime i njezinim rasprostranjenjem. Klimatologija se dijeli na: (1.) opću (fizičku) klimatologiju i (2) klimatografiju. Opća klimatologija proučava fizičke osnove klime kao prosječnog stanja atmosfere. Klimatografija je prikaz klima pojedinih regija ili meteoroloških postaja. Suvremena klimatologija je kompleksna znanost jer osim pojedinačnih klimatskih elemenata istražuje i njihove međusobne odnose kako bi se došlo do što pouzdanijih podataka. Kako je klima osim u prostoru promjenljiva i u vremenu, unutar klimatologije se razvila i paleoklimatologija koja proučava klimu prošlosti.

Slika 7. Osnovni tipovi klime na Zemlji
3.5. Ekologija
Ekologija je prirodna znanost, koja proučava odnose među živim organizmima, kao i njihov utjecaj na okoliš u kojem obitavaju, te utjecaj tog okoliša na njih. Iako se razvila kao grana biologije, ekologija se, osim što se koristi saznanjima iz biologije, dodatno koristi i saznanjima iz kemije, fizike, matematike, te brojnih drugih prirodnih znanosti. Pojam ekologija često se nepravilno koristi pri opisivanju aktivnosti vezanih uz zaštitu prirode. Pojam ekologije - termin ekologija prvi put je uporabio njemački zoolog Ernst Haeckel koji je pod pojmom ekologije smatrao "odnos živih organizama u dva pravca: (i) prema njihovom organskom i (ii) neorganskom okolišu". Sama riječ ekologija dolazi od grčke riječi oikos što znači: okućnica, dom, mjesto za život.

Područja ekologije - Ekologija je široko znanstveno područje koje obuhvaća mnogo poddisciplina. Uobičajena, opća klasifikacija, koja se kreće od najnižih do najviših razina složenosti, gdje se složenost definirana kao broj članova (entiteta) i procesa unutar sustava koji se proučavaju:
• Bihevioralna ekologija proučava uloge ponašanja životinja vezanim za potrebe prilagodbe na prirodni okoliš;
• Populacijska ekologija proučava dinamiku populacija određene vrste.
• Biocenologija (ili sinekologija) usredotočuje se na interakcije između vrsta, unutar neke biocenoze (životne zajednice).
• Ekologija ekosustava proučava tokove tvari i energije kroz biotičke i abiotičke komponente ekosustava.
• Sistemska ekologija je interdisciplinarno polje usredotočeno na proučavanje, razvoj i organizaciju ekoloških sustava iz holističke (sveobuhvatne) perspektive.
• Krajobrazna ekologija proučava procese i vezu između mnogo različitih ekosustava nekog područja ili geografski veće površine.
• Evolucijska ekologija proučava ekologiju na način koji se eksplicitno fokusira na evolucijsku povijest vrsta i njihovih interakcija.
Ekologija se također može podijeliti prema području proučavanja, npr. na (i) ekologiju životinja, (ii) ekologiju bilja, (iii) ekologiju kukaca itd. Još jedna česta metoda podjele je prema proučavanom biomu, npr. na (i) arktičku ekologiju (ili, polarnu ekologiju), (ii) tropsku ekologiju, (iii) pustinjsku ekologiju, itd. Primarna tehnika koja se koristi u proučavanju, često se koristi za podjelu ekologije u podskupine (discipline), kao što su: (i) kemijska ekologija, (ii) genetička ekologija, (iii) statistička ekologija, (iv) teoretska ekologija i dr. Ova se polja međusobno ne isključuju.
3.6. Evolucija Evolucija (lat. evolutio: razvoj, razvitak). Općenito se evolucija može definirati kao razvoj iz niže razine u višu razinu, iz jednostavnoga u složeno, razvoj nekoga prirodnog ili društvenog procesa (npr. umnih sposobnosti, sredstava za proizvodnju); manifestacija neke pojave, događaja ili ideje sukcesivno, u jednom slijedu (npr. slikarstva, pjesništva,

drame , tragedije itd.); u povijesnom razvoju društva, kvantitativna postupna promjena kao suprotnost revoluciji; svaki razvoj koji je sličan rastu žive tvari, nasuprot kreativnom shvaćanju (npr. evolucija nekog izuma, evolucija običajnoga prava); kretanje koje se sastoji od niza povezanih i uzajamno uvjetovanih radnji; proces oblikovanja nekoga finalnog proizvoda, isto tako i sam proizvod iz tog procesa, kao i svaka oblikovana etapa, jedinica iz jedne razvojne serije (npr. cvijeta iz pupoljka) itd.
3. 6.1. Brzina Evolucije
Na osnovu vremenskog trajanja evolucije razlikuju se tri tipa evolucije: 1.Braditelična - sporija od prosječne brzine evolucije, 2. Horotelična ili mezotelična prosječna brzina evolucije, i 3.Tahitelična brža od prosječne brzine evolucije. 3.6.2. Evolucija u biologiji Evolucija u biologiji je skup znanstvenih teorija i hipoteza o nastanku života, o zakonima i putovima postupnoga razvoja živoga svijeta; razvoj organizama od jednostavnih prema složenima; razvoj vrste, populacije, ili koje druge više sistematske skupine. Filogenija je znanost koja proučava evolucijski odnos među vrstama. U najširem smislu, biološka evolucija je proces u kojem nizom promjena ili razvojnih stupnjeva živi organizam ili skupina organizama stječe karakteristične morfološke i fiziološke značajke; teorija po kojoj različiti tipovi životinja i biljaka potječu od drugih preegzistentnih tipova. Upravo prikazana teorija prema kojoj se viši oblici života izvode iz nižih, temelji se na opsežnim promatranjima, koja se nadalje oslanjaju na discipline kao što su paleontologija, embriologija, poredbena anatomija, genetika, molekularna biologija (molekularna evolucija), poredbena fiziologija i znanost o životinjskom ponašanju. Sve navedene discipline znanstveno su se oblikovale već početkom XIX. stoljeća.
3.6.3. Preteče teorije o evoluciji
Ideje o evoluciji u antičkoj Grčkoj Prve ideje o evoluciji rodile su se u doba stare Grčke. Tales9 tvar uzeo vodu, a Anaksimen zrak Prvi grčki koji je konstatirao da se sve oko nas mijenja bio je HeraklitKr. ). Empedoklo (490–430, pr. Kr.) smatra da se svijet sastoji od četiri osnovna elementa: vode, zraka, vatre i zemlje. Po njemu su one prauzrok sveg stvaranja na zemlji. Demokrit jedan od osnivača teorije atoma smatra da se sve stvara slučajnim omjerom atoma. Aristotel je prikupio sve do tada poznate teorije i ideje te saznanja o evoluciji i dao svoj opis živog svijeta. On ga je podijelio na minerale, biljke, zoofite (spužve i koralje za koje smatra da su prijelazni oblici), beskrvne i krvne životinje.
3.6.4. Kasnije teorije 11(540-480, pr.
Kao što je spomenuto u prethodnom poglavlju, evolucijske misli javile su se već u staroj Grčkoj (Heraklit, Empedoklo, Aristotel). Sredinom XVIII. stoljeća Carl Linné, provodeći u svojem djelu Sustav prirode (Systema naturae, 1735.) kategorizaciju svih dotad poznatih biljaka i životinja, strogo je zastupao načelo: "Vrsta ima onoliko koliko ih je od početka stvoreno" (Tot numeramus species, quot ab initio sunt creatae). Suprotno tomu kreacionističkomComte de Buffon u djelu "Povijest prirode" (Histoire naturelle, 1749.–88.) izražava misao o promjeni
vrsta. Na temelju proučavanja velikog broja suvremenih biljaka i životinja, kao i opsežnoga paleontološkog materijala, on zaključuje da su "sve životinje proizišle od jednoga pretka, koji se tijekom vremena mijenjao i usavršavao te proizveo sve životinjske rodove". Zoolog Lacepède, autor poglavlja o ribama u Buffonovoj "Povijesti prirode", tumači sličnost organizama podrijetlom od zajedničkoga pretka i smatra da su te promjene u svezi s promjenljivim utjecajem okoliša, križanjem itd. On govori i o preživljavanju bolje prilagođenih vrsta i u tome se približava temeljima darvinizma, tj. teoriji o prirodnom odabiru, selekciji. Na prirodoslovce XVIII. stoljeća znatno je utjecao i Leibnizov "Zakon kontinuiteta" ili neprekidnosti, po kojem priroda ne pravi skokove, već je u njoj sve povezano postupnim prijelazima, kako u području fizičkih, tako i psihičkih pojava.
9 Tales Milećanin (grč.: Θαλῆς ὁ Μιλήσιος; Milet, Mala Azija, 640. ili 624. pr. Kr. – oko 547. pr. Kr.), grčki filozof predsokratovac, tradicionalno se smatra prvim zapadnjačkim filozofom i ocem znanosti.

10 Anaksimen (585. pr. Kr. - 525. pr. Kr.) bio je starogrčki filozof predsokratovac, pripadnik Miletske škole koja je kao temeljni filozofski problem postavila pitanje o počelu (pratemelju, prauzroku) svijeta. Možemo ga smatrati praznanstvenikom, kozmologom i astronomom (točno je, recimo, ustvrdio da je Mjesečeva svjetlost reflektirana, a zanimao ga je i niz meteoroloških problema), no on je u biti, prema načinu postavljanja pitanja, svakako filozof. 11Heraklit Efežanin (grč. Ἡράκλειτος ὁ Ἐφέσιος; 535. pr. Kr. - 475. pr. Kr.) je grčki filozof predsokratovac. 12 Кreacionizam ili kreacijska teologija je vjerovanje da je vrhovno biće ili božanstvo kreiralo čovjeka, Zemlju i Svemir.

3.6.5. Lamarckova teorija evolucije
Jean-Baptiste Lamarck objavio je 1801. "Sustav životinja bez kralježnice" (Système des animaux sans vertèbres), u kojem je prvi put iznio ideju o zajedničkom podrijetlu organizama i njihovu postupnom razvoju, koju je 1809. u poznatom djelu "Filozofija zoologije" (Philosophie zoologique) razradio u prvu cjelovitu evolucijsku teoriju. Po Lamarcku u prirodi postoji polagan, neprekidan proces preobrazbe vrsta. Čimbenike koji su uvjetovali taj proces, Lamarck svodi na promjene okoliša po kojima se mijenjaju i potrebe životinje, pa time životinja stječe nove navike. U skladu s novim navikama i potrebama jedni se organi više upotrebljavaju i jače razvijaju; obratno, neupotrebljavanje organa dovodi do njihova slabljenja i iščezavanja u potomaka. Te su promjene po Lamarcku uvijek adekvatne uvjetima sredine, a roditelji ih prenose na potomstvo. Tako su npr. krtici reducirane oči, jer ih pod zemljom ne upotrebljava. Te prilagodbe nastaju zbog volje životinje, napregnute želje, težnje da zadovolji svoje potrebe i navike. Lamarck je prvi obuhvatio u cjelini evolucijski problem, ali se zbog manjkavih rezultata tadašnje znanosti zapleo u teleološke pogreške. 3.6.6. Darwinova teorija evolucije Sredinom XIX. stoljeća prirodne su znanosti već raspolagale mnogobrojnim činjenicama koje su govorile u prilog evoluciji. Postignuti su važni uspjesi na području poredbene anatomije, osobito kralježnjaka (Georges Cuvier i dr.), i poredbene embriologije (Baer utvrdio je sličnost zametaka svih kralježnjaka); Theodor Schwann otkrio je 1839. jedinstvo stanične građe svih živih bića (celularna teorija) i osnovao znanost o stanici – citologiju. Charles Lyell u djelu "Načela geologije" (Principles of Geology, 1831.) postupne promjene Zemlje tumačio je sporim i neprekidnim djelovanjem prirodnih čimbenika: vode, vjetra, sunca itd. U poljoprivredi se razvijala tehnika selekcije – engleski stočari i ratari uzgajali su nove pasmine domaćih životinja i nove sorte kulturnoga bilja. Za obradbu i dokumentaciju svoje evolucijske teorije Charles Darwin je, kako se vidi, imao znatno povoljniji znanstveni teren. Uza sve to on je dugo oklijevao s izdavanjem svojega glavnog djela "O podrijetlu vrsta posredstvom prirodne selekcije" (On the Origin of Species by Means of Natural Selection) i objavio ga je tek 1859., kada ga je na to potaknuo Alfred Russel Wallace, koji je te probleme rješavao gotovo na isti način kao i sam Charles Darwin. Darwinova teorija evolucije poziva se ponajprije na prirodni odabir ili selekciju koja uništava jedinke slabije prilagođene uvjetima života, a podupire one bolje prilagođene. Individualne varijacije koje ulaze u proces selekcije mogu, po Darwinovu mišljenju, biti determinirane vanjskim (okoliš) i unutarnjim (poslije nazvani genetičkim) čimbenicima. Darwin je nadalje dokazao da evolucija ima adaptivan značaj i da je svrhovitost relativno izražena u prilagođenosti organizma na određeni povijesno uvjetovani kompleks vanjskih uvjeta. Njegova je teorija u znanosti poznata kao darvinizam. Često se i sama znanost o evoluciji

poistovjećuje s pojmom darvinizma. Darwinovo naučavanje dalje su razradili, proširili i produbili Huxley, Ernst Haeckel, Kliment, Arkadijevič, Timirjazev i drugi. Haeckel osobito se istaknuo kao vatreni pristaša Darwinove teorije. On je znanstveno razradio problem jedinstva žive i nežive prirode, postanak života kemijskim putem iz nežive prirode, monofiletsko13 podrijetlo živih bića itd. Na kraju Haeckel je sastavio biogenetski zakon koji u suvremenoj evoluciji ima samo povijesno značenje. Prema Haeckelu filogenija je osnovna znanost o promjenama oblika kroz koje prolaze organizmi tijekom cijeloga svojeg razvoja. Haeckel je tako prvi postavio teoriju descendencije14 , koju je nazvao transformizmom, spojivši u njoj uz darvinizam i neke lamarkističke misli.

3.6.7. Evolucionistički smjerovi nakon Darwina

13 - porijeklo iz jedne razvojne linije (jednog predačkog organizma). monofilija - odnosi se na cvijet kojeg oprašuje samo jedna vrsta oprašivača 14 pogled koji vidi sav svijet u rodbinskoj povezanosti, sve postojeće prirodne skupine (vrste) nastale su iz zajedničkog pretka. Osnovna su pitanja po kojima se darvinizam i lamarkizam razilaze pitanje nasljeđivanja stečenih svojstava i problem biološke svrhovitosti. Za lamarkiste je osnovno načelo evolucijskoga procesa neki teleološki postulat, "unutrašnja svrha", pa u svezi s time oni umanjuju ili sasvim ignoriraju ulogu prirodnog odabira, a to je po darvinizmu odlučujući faktor biološke evolucije. Na temelju lamarkizma razvio se cijeli niz idealističkih mehanicističkih, tzv. neolamarkističkih hipoteza, koje se mogu svrstati u dvije skupine: psiholamarkizam i mehanolamarkizam. Psiholamarkizam je utemeljio 1871. Edward Drinker Cope i po njemu je glavni pokretač evolucije psihička energija, koja se kod nižih životinja očituje u "nastojanju pod utjecajem nužde", kod viših životinja u "izboru", a kod razumnih životinja u "svjesnom izboru". Mehanolamarkizam se razvio na osnovi nazora engleskog filozofa Herberta Spencera (1820.–1903.), koji je evoluciju organizama tumačio prijelazom tvari iz jednoličnog i indiferentnog stanja u raznolično i diferencirano stanje. U skupinu mehanolamarkističkih strujanja ulazi i teorija Theodora Eimera (1843.–1898.), poznata pod imenom Eimerova ortogeneza. Po toj je teoriji evolucijski proces koji ima sam po sebi točno određen smjer (tzv. usmjerena evolucija). Nasuprot različitim lamarkističkim smjerovima i hipotezama, koje podcjenjuju ili negiraju Darwinovo načelo prirodnog odabira, zoolog August Weismann ne priznaje nikakve druge faktore evolucije osim prirodnog odabira. Ta se teorija naziva neodarvinizam ili vajsmanizam. Weismann proširuje načelo prirodnog odabira i na procese koji se zbivaju u samom organizmu. On smatra da je nasljedna tvar ili zametna plazma sastavljena od niza nasljednih jedinica koje su okupljene u jezgri stanice. Najvažnije od njih bile bi posebne materijalne jedinice, "determinante", koje određuju svojstva pojedinih stanica. Determinante se mogu k tome mijenjati, i to pojedinačno, neovisno o drugima, tako da cijeli

organizam postaje mozaik pojedinih svojstava. Na taj način Weismann svu raznolikost evolucijskoga procesa svodi na kombinaciju nasljednih elemenata. Weismannove determinante prihvatio je botaničar Hugo de Vries i postavio svoju mutacijsku teoriju evolucije. Po toj teoriji organizmi se mijenjaju samo u iznenadnim, skokovitim promjenama (mutacijama), a one se javljaju neovisno o utjecaju okoliša. Te mutacije nenadano proizvode nove vrste, koje su odmah stalne. Na temelju novih znanstvenih činjenica Stephen Jay Gould i Niles Eldredge dali su nov model evolucijskih promjena koji su nazvali isprekidanom ravnotežom (engl. punctuated equilibria) ili punktualizmom, prema kojem se evolucijske promjene događaju naglo, nakon dugotrajna razdoblja stagnacije, za razliku od gradualizma – mišljenja da se evolucijski proces odvija postupno kroz dugo vrijeme. Osim navedenih, postoji još cijeli niz evolucionističkih hipoteza, smjerova i pravaca. Sva su ta strujanja dala poneki doprinos razvoju biologije, ali kao evolucijske teorije imaju uglavnom povijesno značenje. Sintetička teorija o evoluciji plod je istraživanja niza suvremenih znanstvenika (Ronald Aymler Fisher, Julian Huxley, Sewall Wright, Hermann Joseph Muller, Theodosius Dobzhansky, Ernst Mayr, George Gaylord Simpson, Richard Goldschmidt, Nikolaj Vladimirovič Timofejev-Resovski, Nikolaj Petrovič Dubinjin, Ivan Ivanovič Šmaljgauzen), a napustila je sve one hipoteze koje svojim spekulativnim primjesama priječe ili ograničavaju mogućnost egzaktnog istraživanja i eksperimentalnog provjeravanja. Baza je te moderne teorije darvinizam, u koji su se, uz neke izmjene, mogli izvanredno ugraditi rezultati moderne biologije. Evolucija živih bića saglediva je: a) kao povijesna činjenica koja je imala svoj tijek i b) kao proces kojemu su na svoj način i sada podvrgnuta živa bića i po čemu se može dobiti uvid u njezine faktore. Kao povijesna činjenica ona je dokumentirana mnogobrojnim paleontološkim nalazima i komparativnim proučavanjima. Na temelju toga materijala uspješno su rekonstruirani tijekovi razvoja nekih biljnih skupina, nekih odjeljaka životinja, osobito kralježnjaka, a među njima specijalno najviših sisavaca i čovjeka. Ima i pokušaja prikazivanja drveta (stabla) života sa svim prijelaznim oblicima od njegovih prvih početaka do čovjeka. Te rekonstrukcije pružaju često dobru opću orijentacijsku sliku, ali vrijede samo kao bolje ili slabije obrazložene spekulacije o razvoju, jer su rađene bez fosilnih dokaza. Budući da prvi oblici života nisu mogli ostaviti tragova, evolucijska se proučavanja stalno nalaze pred problemom kako su se i u kojim okolnostima pojavila najjednostavnija živa bića. Problem ostaje i razvoj jednostaničnih organizama , npr. ameba, u kojima već postoje svi osnovni procesi karakteristični za živa bića. G. G. Simpson smatra da je razvoj od prvih jednostavnih bića do amebe bio najteži i najkompleksniji, te da je vjerojatno trajao najmanje onoliko vremena koliko i razvoj od amebe do čovjeka. U pitanju razvoja višestaničnih organizama, neki znanstvenici tvrde da put od jednostaničnih do višestaničnih organizama ide preko kolonija jednostaničnih bičaša; zoolog Jovan Hadži zastupa teoriju celularizacije. Prava "povijest" živih

bića počela je u najstarijim razdobljima paleozoika, u kambriju i siluru, gdje ima fosila svih glavnih tipova životinja (jednostanične životinje, člankonošci, preteče kralježnjaka ili kordati, kralježnjaci). Krajem devona pojavili su se fosili djelomično kopnenih životinja (vodozemci), a u karbonu kopnene životinje, gmazovi. Mezozoik se smatra erom gmazova, koji su osobito brojni u njegovu posljednjem razdoblju, u kredi; u kenozoiku su izumrli mnogi njihovi redovi. Potkraj trijasa pojavili su se fosili gmazova sa svojstvima sisavaca, a u juri gmazova i ptica. Nalazi gmazova toliko su brojni i raznoliki da se upravo s pomoću njih mogla prikazati jedna od tipičnih pojava u evoluciji, tzv. proces adaptivne radijacije. Tu se vidi kako se unutar jedne skupine na temelju jednoga zajedničkog oblika razvijaju različiti oblici »u najrazličitijim smjerovima uvjetovanima vlastitim mogućnostima i sredinama u koje ulaze« (G. G. Simpson). Nakon znatnoga prorjeđenja gmazova, proširile su se ptice i sisavci. Oni su u tercijaru bili malobrojni, ali je postojalo mnogo njihovih vrsta. Sisavci su se razvili od gmazova na temelju potpuno nediferenciranoga, prijelaznog oblika. Njihovo dobro zaštićeno potomstvo (rađaju živu mladunčad koju hrani majka), stalna tjelesna toplina i uravnoteženi metabolizam vitalne su prednosti koje su im omogućile da se mnogostrano razvijaju i prošire. Među njima je osobito zanimljiv red primata, u kojih se predstavnik pojavio u paleocenu. Iz tog osnovnog tipa u procesu adaptivne radijacije odvojili su se Lemuroidea , Tarsioidea i Hominoidea, među kojima se pojavio i čovjek. Iz činjenice da postoji oko 2 milijuna različitih vrsta živućih organizama, a utvrđeno je i mnoštvo izumrlih, te odatle da svi organizmi imaju neka zajednička svojstva (reprodukcija, asimilacija, iskorištavanje energije) proistječe da su raznolikost i jedinstvo temeljne karakteristike života. Evolucija kao razvoj vrsta tijekom vremena, postupnim slijedom promjena od jednostavnoga do složenog stanja, uključuje u sebi navedene karakteristike. Moderna teorija evolucije razvija se na temeljima darvinističkoga shvaćanja unoseći daljnje evolucijske faktore otkrivene ponajprije u okvirima genetike. Prvotno shvaćanje o nasljeđivanju stečenih svojstava modificirano je stečevinama molekularne biologije, gdje se raspravlja o povratnoj vezi između citoplazmatskih bjelančevina (sposobnih da prime utjecaje iz okoline) i DNK informacija. Razradbom nekih temeljnih bioloških činjenica i pojava (zakoni nasljeđivanja, vrsta, rasa, mutacija, selekcija , okoliš), prevladavanjem filozofskih gledišta teorije preformacije i duboko uvriježenoga tipološkog mišljenja (da u osnovi pojavnim različitostima leže »ideje« kao nešto jedino realno i nepromjenjivo), evolucija je postala središnja biološka znanost koja je zahvatila u sve ogranke biologije i fiziologije do genetike, morfologije i ekologije. Njezino je područje osobito unaprijeđeno dostignućima populacijske genetike. Osnovni evolucijski procesi zbivaju se unutar populacije. Evolucija započinje narušavanjem genske ravnoteže. Ravnoteža se remeti onda kada se mijenjaju uvjeti u populaciji ili u okolišu. Analize uzročnih osnova evolucije

utvrđuju da su njezine osnovne sile mutacija, prirodni odabir ili selekcija i genetska tendencija; to su procesi koji remete nasljednu ravnotežu.

3.6.8 Mutabilnost Mutabilnost je sposobnost promjene nasljednoga materijala. Osim promjena u strukturi gena (genske mutacije), u stanicama se mogu pojaviti i promjene u broju kromosoma. Različiti oblici mutacije gena proširuju genetsku raznolikost populacije, odnosno njezinih genskih zaliha, udaljujući je time od "divlje" forme. Budući da je mutacijska učestalost, koja djeluje u smislu udaljavanja od "normalnog" alela divlje forme, mnogo veća od "povratne mutacije" prema divljoj formi, izražena je stalna tendencija k razbijanju jednoličnosti genskih zaliha ili tzv. mutacijski pritisak. Mutacijskim pritiskom, iz čisto nasljednih, nastaje populacija raznolikih genotipova. Budući da su mutacije moguće stalno i kod svakoga gena, one su izvor nasljedne varijacije. Mutirani geni imaju pojačane interakcije s drugim genima, i taj posredni učinak još je značajniji za raznolikost genotipova i fenotipova, a njihov je utjecaj na evoluciju veći od neposrednog učinka samoga mutiranog alela. Rekombinacijom kromosoma ili izmjena dijelova kromosoma s odgovarajućim dijelovima homolognoga kromosoma koja se zbiva tijekom mejoze poznata je kao crossing-over. Rekombinacije pojačavaju učinak mutacije stvarajući širok spektar genskih kombinacija.
3.6.9. Prirodni odabir ili selekcija
A.) Prirodni odabir ili selekcija je druga je osnovna snaga evolucije, ona je prirodni izbor između nositelja različito vrijednih nasljednih faktora. Kao otkriće, načelo selekcije (odabira) glavni je Darwinov doprinos teoriji evolucije, koju on objašnjava borbom za opstanak među pojedinim organizmima, od kojih preživljavaju najsposobniji. Selekciju, Darwin primjenjuje na jedinke a ne na populacije, a okoliš kao selektivna sila odabire varijante koje su joj se najbolje prilagodile. Darwin je time naglasio negativno gledište selekcije, ističući da je ona neumoljiva snaga koja uništava jedne a unaprjeđuje druge. To je stajalište Darwin temeljio na diferenciranom mortalitetu koji može dovesti do diferencirane reprodukcije. Danas je modificirano načelo selekcije temeljna orijentacija u rješavanju evolucijskih problema. Prema Hardy-Weinbergovu pravilu, kao polazištu u populacijskoj genetici, u standardnim uvjetima okoline svi geni populacijskih genskih zaliha dolaze do ravnoteže koja se stalno održava (idealna populacija). Zato je prirodni odabir, osim mutacija, jedina snaga koja uzrokuje promjene u genskoj ravnoteži populacije. Prirodna selekcija je utjecaj bilo kojega faktora iz okoliša na organizme. Kao selekcijski faktori mogu djelovati: ekstremne temperature, oborinski omjeri (sušna razdoblja, poplave), kemijski uvjeti, prirodni neprijatelji različitih štetočina i nametnika koji

unaprjeđuju evoluciju pojedinih vrsta, zatim paraziti i uzročnici bolesti koji tijekom širenja i djelovanja zaraze selekcijom i umanjenjem populacije snažno utječu na evoluciju domaćinova organizma, a kroz njegovu obranu također i na svoju vlastitu. B.) Natjecanje i borba oko hrane, životnoga prostora i ostalih važnih životnih uvjeta ubraja se u unutar vrstnu selekciju koja podređene skupine potiskuje u nove ekološke niše ili vodi njihovu izumiranju. Tako djeluje selekcijski pritisak. Djelujući tijekom milijuna godina, selekcija omogućuje razvoj novih adaptacija u najrazličitijim sredinama na Zemlji. C.) Genska tendencija (drift) treća je osnovna sila evolucije, a uvjetuje ustaljivanje neutralnih ili neadaptivnih svojstava. Genski drift djeluje samo u malim populacijama, i to po načelu vjerojatnosti. Rezultati su slučajni i uzrok su slučajnom ustaljivanju gena ili gubitku mutiranog gena. Gubitak ili fiksacija događa se bez obzira na selekcijski pritisak, prema čemu i naziv pojave: genetski drift, tj. genetski nanos. Tako osim selekcijskog istrjebljenja, postoji i mogućnost da se oni geni, inače samo usputni, koji se u velikoj genskoj zalihi nisu mogli istaknuti, odjednom istaknu u slučajem uvjetovanom genetskom driftu u malim populacijama i tako postanu valjani za neki genetski sastav, dok se oni geni koji su prije toga bili zastupljeni sada gube. Kao primjer za tu pojavu uzimaju se Darwinove zebe na otočju Galápagosu .
Slika 8. Genska tendencija (drift)
3.6.10. Podjela evolucijskih procesa S obzirom na opseg i određene karakteristike evolucijskih procesa, evoluciju je moguće razlučiti na: 1. mikroevoluciju, 2. makroevoluciju i 3. megaevoluciju kao stupnjeve ili razine u neprekidnom evolucijskom nizu.

a. Mikroevolucija - sadrži manje sukcesivne promjene u genskoj zalihi određene populacije od jedne do druge generacije. U njoj djeluju osnovne sile evolucije. Procesi mikroevolucije vode stvaranju novih vrsta, odnosno populacija. Specijacija je odjeljivanje i razvoj novih populacija, tj. nastanak novih vrsta. Još se naziva i populacijska divergencija. Cijeli tijek evolucije ovisi o podrijetlu novih populacija koje imaju adaptivnu sposobnost veću od svojih predaka. U nastanku novih vrsta uz osnovne sile djelatne su i dodatne sile, koje djeluju dulje vrijeme. Glavni faktor je izolacija. Možemo izdvojiti nekoliko oblika izolacije:
• geografska,
• ekološka,
• reproduktivna i
• genska.
U geografskoj izolaciji proces se odvija tako da se početna populacija širi od sredine svojega prebivališta u susjedna područja. U početku se izmjena gena nastavlja između svih dijelova populacije. Postupno se male populacije počinju fragmentirati i ustaljivati u najpogodnijim prebivalištima, pa jedne od drugih postaju odijeljene ekološki nepovoljnim područjima. Nakon određenog vremena izolacije započinje fragmentacija u još manje populacije ili demove. Dijeljenje početne populacije u homogene serije dema poznato je kao ponašanje svih živućih organizama. Svaki dem zadržava isti genski sastav kakav je imala populacija predaka, ali su ekološke barijere genskoj izmjeni između demova temelj genske divergencije između njih. Svaki izolirani dem biva od početka modificiran silama mikroevolucije. Nakon nekog vremena demovi se počnu međusobno razlikovati, iako još uvijek ostaju otvoreni genskoj izmjeni. Primarni je način postanka vrste alopatrijska specijacija; u njoj je prostorna izolacija, pojačana geografskim ili ekološkom barijerom, djeluje tako da dvije prvotno vrlo slične populacije postaju genski izolirane. Ekološka izolacija sastoji se u tome da populacije koje žive na istom geografskom području postaju izolirane faktorima okoliša tako da zauzmu odijeljene biotope ili nastanjuju specijalne ekološke niše. Taj proces vodi stjecanju specifično usklađenih gena i gubljenju prvotno zajedničkih oznaka, a to unaprjeđuje fiziološku izolaciju. Dogodi li se da se takve populacije, nakon dugog vremena, ponovno nađu zajedno, unatoč činjenici da nastanjuju isti areal, one se više međusobno ne križaju. Reproduktivna izolacija sastoji se u razlikama u ponašanju u doba parenja (npr. u ptica razlike u glasanju), u razlikama s obzirom na razdoblje i sezonu parenja, u morfološkoj neusklađenosti kopulacijskih organa što priječi nesmetano neograničeno križanje (panmiksiju). Čini se da geografsko odjeljivanje odlučno utječe na mehanizam te izolacije. Genetska izolacija nastupa kao posljedica promjena u kromosomskoj garnituri ili u poretku gena. Kada nastupe takve promjene, kromosomi dotičnih

vrsta ne mogu se spojiti, pa više-manje nepravilno djeluju ako i dođu u dodir. Zbog antagonističnosti genetskoga sastava roditeljskih vrsta, gamete ili ugibaju već u samom početku procesa oplodnje ili pak, ako se nastavi razvoj zametka, rađaju se životno nesposobni ili sterilni hibridi. Nastanak novih vrsta kroz samu genetsku izolaciju je rijedak, ali zajedno s geografskom izolacijom dobiva veliko značenje.
b. Makroevolucija ili adaptivna radijacija je evolucija iznad razine vrste, a karakterizira ju dijeljenje ili fragmentacija, koja dovodi do postanka i razvoja većeg broja adaptivnih tipova. Dok u mikroevoluciji i specijaciji nastaju posebne adaptacije, u makroevoluciji razvoj se odvija od opće adaptacije divergentno prema mnogobrojnim posebnim adaptacijama. Makroevolucija obuhvaća razvoj viših sistematskih skupina (porodica, red, razred). Nju karakterizira: (i) dioba skupina u mnogo novih podskupina; (ii) prodor u nove sredine; (iii) organiziranje različitih struktura i načina života.
Osnovna karakteristika evolucije iznad razine vrste je pomicanje skupine organizama u nove adaptivne zone. Kako bi organizam mogao proći kroz zonu ekološke nestabilnosti u novu adaptivnu zonu, on mora imati otvoren evolucijski i ekološki pristup u nju, tj. skupina mora već imati neka prilagodljiva svojstva, a zona ne smije u sebi imati jakoga konkurenta. Prvi prodor u novu zonu zahtijeva samo opću adaptaciju; poslije toga evolucija vodi specijaciji. Važno je svojstvo makroevolucije također usmjeravanje razvoja nekih svojstava prema samo jednoj razvojnoj liniji. Evolucija gmazova paleontološkim15 je nalazima najbolje dokumentiran primjer makroevolucije. Čini se da su određene karakteristike adaptivne radijacije, utvrđene u evoluciji gmazova, osnovne značajke makroevolucije u svim skupinama, a to su: 1. do makroevolucije uvijek dolazi nakon stjecanja nove opće adaptacije ili nakon ulaska u novu adaptivnu zonu (kod gmazova je nastala opća adaptacija za život na kopnu). 2. makroevolucija nije linearna već zrakasta. Radijacija se nastavlja na opću adaptaciju i na ulaz u novu adaptivnu zonu time što se razvija specijalna adaptacija u različitim divergentnim novim linijama.
3. adaptivna radijacija stvara evolucijske linije koje, s obzirom na specijalne adaptacije, konvergiraju prema drugim sistematski udaljenim skupinama, a koje se razlikuju po načinu opće adaptacije. (Ihtiosauri16 su primjer evolucijske konvergencije prema ribama

4. makroevolucija stvara skupine paralelnih specijalnih adaptacija između divergentnih ali srodnih linija, koje imaju zajedničku osnovu opće adaptacije. (Fitosauri i krokodiliiz trijasa danas su primjer takva paralelizma i ekološke zamjene.)

17 Premosnici (često , ponekad i ) () su jedini recentni rod reda (u nekoj literaturi [1] ) u razredu gmazova. Pored toga, jedini su recentni predstavnici porodice . Danas postoje još samo dvije vrste, koje žive na sjeveru Novog Zelanda, i često ih se naziva i "živim fosilima", jer se njihovo postojanje može slijediti fosilnim nalazima unatrag sve od razdoblja trijasa, u vrijeme prije oko 200 milijuna godina.
5. u makroevoluciji zbiva se definitivno propadanje skupina organizama. Kako je opća adaptacija zamijenjena specijalnom, skupine se kruto specijaliziraju na uske adaptivne podzone i gube sposobnost za pomak u nove veće zone. Budući da se sve adaptivne zone moraju konačno promijeniti, skupine zatvorene u usku zonu moraju propasti. (Primjer su za to Rhynchocephaliakrede, od kojega živi još samo jedan rod (premosnici) sa samo dvije vrste u Novom Zelandu .)
c. Megaevolucija je pojava novih bioloških sustava ili tipova organizacije. Kroz tri milijarde god. razvilo se gotovo 200 takvih tipova. Njihovo podrijetlo najvažniji je događaj u evoluciji, vrlo kompliciran i najmanje istražen.
Megaevoluciju karakteriziraju: 1. pokušaji i istraživanja raznosmjernih linija od polaznoga pretka da prijeđu ekološku granicu i uđu u novu zonu, dok jednoj konačno to i ne uspije; 2. prodor i promjena uvijek su brzi; 3. nova zona uvijek je ekološki pristupačna i nema suparnika, a zahtijeva novi opće adaptivni tip za invaziju (npr. razvoj gmazova od vodozemaca). Glavno je svojstvo organske evolucije divergencija koju usmjeruje prirodna selekcija kao glavna sila. Na svim razinama evolucije djelatne su osnovne sile, ali se javljaju i sve složenija djelovanja manje poznatih sila.
3.6.10.1. Molekularna evolucija
Molekularnom evolucijom se bavi moderna grana biologije. Predmet istraživanja nastanak je prvih živih bića iz predbioloških molekula i njihova daljnja evolucija, a koristi se strukturnom srodnošću makromolekula u danas živućim vrstama. Budući da je sveukupna informacija o ustrojstvu živih bića zapisana u genomu, biološka evolucija očito je evolucija genoma. Varijabilnost populacije zasniva se na slučajnoj varijabilnosti genoma (mutacija). Prirodna se pak selekcija zbiva samo na razini produkata gena. Filogenetsku srodnost pokazuju zato sveukupni

genomi, geni unutar tih genoma te produkti gena – bjelančevine. Upravo analiza redoslijeda nukleotida u DNK pruža potpunu informaciju o sličnostima i razlikama među genima. Premda se sve mutacije zbivaju jednakom učestalošću, geni i njihovi produkti ne evoluiraju jednako brzo, jer preživljavaju samo prihvatljive promjene. Jedinično evolucijsko vrijeme je potrebno da se promijeni 1% aminokiselina u nekoj bjelančevini; u eukariota to može biti od milijun do čak 600 milijuna godina. Tako se histoni graška H4 i krave (vrste koje su se odvojile prije više od milijardu godina) razlikuju samo u dvije aminokiseline. S druge strane, strukture citokroma c i brojnih drugih gena ili produkata gena omogućuju da se prvi put u povijesti konstruiraju filogenetska stabla na osnovi kvantitativnih podataka a ne slobodne procjene. Evolucija genoma svakako uključuje porast količine genetičke informacije. Genomi najjednostavnijih danas živućih samostalnih organizama sadrže oko milijun parova nukleotida, a genomi sisavaca oko tri milijarde. Današnji genomi jasno pokazuju put evolucije gena: prvo je pri repliciranju nastala identična kopija nekoga gena, a zatim su se nizom mutacija dvije kopije razilazile u svojim strukturama (divergentna evolucija). U nekim slučajevima iz strukturno različitih pragena nastali su geni za bjelančevine srodnih funkcija (konvergentna evolucija). Analiza genoma omogućuje klasifikaciju živoga svijeta u tri velike domene: Archaea, Bacteria i Eucarya. Ona se međusobno razlikuju u sustavu za prevođenje genetičke informacije s nukleinskih kiselina na bjelančevine, što

očigledno pokazuje na njihovo razdvajanje u najranijoj fazi evoluciji
Filogenetska slika živih bića

3.7. Izumiranje
Izumiranje je pojava koja postoji otkako postoji i život na Zemlji, izumrle su mnoge biljne i životinjske vrste. Ponekad se to događa zbog prirodnih promjena klime, grabežljivaca i biljnog pokrova, no u zadnje vrijeme je glavni krivac čovjek. Izumiranje u najužem smislu riječi znači potpuno nestajanje nečega, ono može biti postupno ili naglo, odnosno može biti masovno izumiranje. Slika 10. Dodo (Raphus cucullatus), izumrla vrsta Od pretpostavljenog broja životnih vrsta na Zemlji negdje oko 4 milijarde, danas ih postoji manje od 50 milijuna.
3.7.1. Ugrožene vrste
Danas posebno ugrožene vrste su velike mačkeosim lavova (lov zbog krzna), razni jeleni i ptice (meso), i životinje u šumama koje gube staništa (sječa). Ako se ovako nastavi, za desetak godina bi mogle nestati stotine vrsta. Posebna opasnost prijeti divljim životinjama kao što su tigrovi, koji bi mogli izumrijeti do 2015. 3.7.2.Borba protiv izumiranja
Organizacija IUCNcrveni popis biljnih i životinjskih vrsta prema stupnju ugroženosti. 18se bavi ovim problemom. Radi podizanja svijesti o ovome problemu i sprječavanja daljnjeg izumiranja biljnih i životinjskih vrsta, sastavila je 18 Međunarodni savez za očuvanje prirode (nternational nion for the onservation of ature and Natural Resources) osnovan je . godine i obuhvaća 83 države članice, 108 vladinih agencija, 766 nevladinih agencija i 81 međunarodnu organizaciju. Okuplja oko 10.000 stručnjaka i znanstvenika iz država širom svijeta. Cilj rada mu je korištenje prirodnih resursa na znanstvenim osnovama te zaštita rijetkih vrsta i ugroženih prirodnih staništa.
GEOBOTANIKA Geobotanika ili biljna geografija bavi se rasprostranjenjem biljaka na Zemljinoj površini. Ona ga pokušava utvrditi, analizirati njegova pomicanja i pravilnosti te razjasniti uzroke tog rasprostranjenja. Oni su dijelom ekološki, tj. nalaze se u izmjeničnim odnosima između životnih potreba biljaka i životnih uvjeta koji vladaju na Zemlji, a dijelom su sadržani u povijesti postanka i rasprostranjenja biljaka. Geobotanika se većinom dijeli na četiri dijela:
(1) nauka o arealu (ili flori), tj. floristička biljna geografija ili horologija, bavi se rasprostranjenjem pojedinih sistematskih jedinica,
(2) nauka o vegetaciji (uključivši i biljnu sociologiju - fitocenologiju) bavi se rasprostarnjenjem biljnih zajednica,
(3) nauka o staništu i ekološka biljna geografija istražuju ekološke osnove biljnog rasprostranjenja, a
(4) genetska (ili historijska) biljna geografija proučava povijest flore i vegetacije.
Mi ćemo se u okviru geobotanike detaljnije upoznati s dva geobotanička kompleksa, i to s florom i vegetacijom.

FLORA Svi biljni taksoni1 nekog određenog zemljopisnog područja tvore njegovu floru, pa pod florom podrazumijevamo sve biljke nekoga, prirodnim ili umjetnim granicama određenog prostora. Kako među biljkama postoje različite sistematske skupine kao što su alge (zelene, smeđe, crvene), lišajevi, mahovine, itd. možemo razlikovati npr. floru zelenih alga, floru mahovina itd. Međutim, kada u običnom govoru mislimo na floru bez posebne napomene o kojoj se flori radi, u pravilu se misli na floru stablašica s dobro razvijenim provodnim sistemom, koje se zbog toga označavaju imenom vaskularne biljke. Ako želimo točno označiti da se radi baš o takvim biljkama, tada se to i pobliže određuje kao «flora vaskularnih biljaka» ili kraće «vaskularna flora». Tako možemo govoriti npr. o flori odnosno vaskularnoj flori grada Splita, Mostara, Zagreba, Dalmacije, Hercegovine, odnosno o flori Hrvatske, flori Europe i flori čitave Zemlje. 1 Takson – sistematska jedinica biljaka višeg ili nižeg reda. Kada bismo gledali koliko tih vaskularnih biljaka ima, tada bi na području grada Splita mogli popisati 600 – 700 vrsta, u flori Dalmacije otprilike oko 3000 vrsta, u flori Hrvatske oko 4000 vrsta, u flori Europe nešto preko 11000 vrsta i u flori Zemlje oko 250 – 300 tisuća vrsta. Biljne vrste koje rastu zajedno na nekom ograničenom području našle su se tamo na temelju vrlo različitih čimbenika. Ti čimbenici mogu biti biološki, klimatski, geografski, orografski, a sve te čimbenike možemo zajedničkim imenom označiti kao recentni (lat. = skorašnji, današnji) čimbenici. Njima moramo pridodati i čimbenike iz bliže ili dalje geološke prošlosti, a to su najčešće paleografski i paleoklimatski čimbenici. U rasprostranjivanju i širenju biljnih vrsta od neolita do naših dana presudno značenje ima čovjek, što je poznato pod nazivom antropogeni čimbenici.

Sastav i analiza područne flore U sastavu dobro proučene područne ili regionalne flore mogli bismo zabilježiti vrlo različiti spektar različitih biljnih vrsta i njih bismo mogli analizirati na različite načine. Mogli bismo tako analizirati sistematsku pripadnost pojedinim porodicama, i to izraziti brojčano i u postocima. Kako različita područja imaju različiti broj biljnih vrsta brojčani pokazatelji pripadnosti pojedinim porodicama bili bi različiti, ali kada se to izrazi u postocima od ukupnog broja zabilježenih vrsta postoci bi bili vrlo slični. Tako naši otoci, kada bismo popisali njihovu floru najviše pripadnika imale bi porodice Poaceae, Asteraceae, i Fabaceae. Mogli bismo reći da je to opća značajka Sredozemlja. Drugi pristup analizi flore bio bi prema životnim oblicima, pa bismo mogli izraditi spektar životnih oblika. U našem Sredozemnom području dominiraju dva životna oblika i to: hemikriptofiti (H) i terofiti (T). Iz toga bismo mogli zaključiti da je zima razmjerno hladna, a ljeto vruće i suho. U još sušnijem i toplijem dijelu Sredozemlja prevladavaju terofiti, jer su tamo ljeta još toplija i suša, a zime gotovo i nisu hladne. Kao treća mogućnost analize područne flore može nam poslužiti današnja rasprostranjenost pojedinih biljnih vrsta i to je florni geoelemet. U nekim slučajevima uspijeva nam ustanoviti i podrijetlo pojedinih biljnih vrsta, pa govorimo o flornom genoelementu. Ako bismo vrste analizirali prema pripadnosti pojedinim biljnim zajednicama, tada govorimo o flornom cenoelementu. Na kraju mogli bismo unutar neke flore razlikovati autoktone biljne vrste, tj. biljne vrste koje su se rasprostranile nekim područjem na temelju prirodnih sila kao što su vjetar, voda, životinje i dr. i aloktone biljne vrste koje je u neko područje svojim aktivnostima unio čovjek. Takve biljke označavamo s dva termina i to antropokorne ili sinantropne biljke. Bolji je termin antropokorne biljke ili kraće antropokore, jer su terminološki analogne ornitokorama2, anemokorama3, hidrokorama4 itd. 2 ornitokorama-rasprostranjuju se pomoću ptica. 3 anemokorama –rasprostranjuju se pomoću vjetra 4hidrokorama – rasprostranjuju se pomoću vode. Smatra se da u Europi ima oko 20% antropokornih biljaka prenijetih iz Amerike. U Velikoj Britaniji od oko 1850 vrsta čak je oko 40% vrsta prenijetih na otok posredstvom čovjeka. (antropokorno). 59

Areali Pod pojmom areal 5 smatramo područje rasprostranjenosti neke biljne skupine (taksona). To može biti areal porodice, roda, vrste ili podvrste. Prema teoriji o monotropnom nastajanju biljnih vrsta smatra se da je u nastanku neke vrste sudjelovao jedan roditeljski par na jednom mjestu (monotropno) i samo jednom tijekom evolucije. Ukoliko se zbog nekih prednosti u odnosu na konkurente pojedini potomci ističu nekim korisnim svojstvima, oni počinju postupno prevladavati u populaciji, dok konačno ne istisnu sve one jedinke koje nisu prilagođene na neke novostvorene ekološke uvjete. Oni, pak, individui koji su najprilagođeniji takvim novonastalim uvjetima množe se sve više I više, te se koncentrično šire na sve strane tako dugo dok ne dođu do neke prepreke (barijere) koja ih zaustavlja u širenju. Barijera može biti geografska, kao npr. obala mora za kopnene biljke ili ekološka uvjetovana promjenom nekih ekoloških čimbenika. Tako širenje niza biljnih vrsta u Europi prema sjeveru zaustavlja hladnoća. 5 Areal, (definicija): područje, oblast, pojas prirodne rasprostranjenosti bilo koje skupine biljaka ili životinja. 6 Holarktik – holo – sav artik = područje sjeverne Zemljine polutke. 7 Cirkumarktičke – cirkum = naokolo 8 Borealis - sjeverni Tako nastali areali pojedinih vrsta vrlo su različiti i nastali su uglavnom davno tijekom geološke prošlosti, pa ih danas ne susrećemo u njihovom izvornom obliku za koji ne znamo kako je izgledao. Mi npr. danas znamo da je Ginko biloba u živom stanju poznata jedino iz Japana, a u fosilnom obliku iz vrlo različitih dijelova Euroazije. To znači da je Ginko biloba pred 200-tinjak milijuna godina bila rasprostranjena na ogromnom prostoru, a danas se je sačuvala samo u izuzetno malenom području. Takvih primjera ima mnogo. Kako je do danas proučeno veliko mnoštvo areala ogromnog broja vrsta, možemo vrste prema oblicima njihova areala, općenito klasificirati u 1) kozmopolite, 2) široko rasprostranjene, 3) endeme i 4) relikte. Zanimljivo je naglasiti, što je gotovo nevjerojatno, da kozmopolitskih biljaka koje su se čitavom Zemljom raširile prirodnim silama ima izuzetno malo, jer ima biljaka koje su danas kozmopolitski rasprostranjene ali zahvaljujući čovjeku. Jedna od takvih, rijetkih prirodno kozmopolitskih biljaka je obična trstika (Phragmites australis). Ne zna se točno objasniti kako se je uspjela rasprostraniti čitavom Zemljom. Široko rasprostranjene biljke – mogli bismo reći da među prirodno rasprostranjenim biljnim vrstama tj. skupni pripada najveći broj biljaka. Kada bismo proučavali današnju rasprostranjenost takvih vrsta na Zemlji, ona nam je značajna u fitogeografskom smislu za diferencijaciju pojedinih flornih carstava (regija). Ako bismo se ograničili na našem flornom carstvu sjeverne polutke – Holarktiku 6, možemo razlikovati cirkumarktičke biljke 7, borealne 8 biljke, europske i azijske biljke, i mediteranske biljke (sl. 39).

Prema tome, u okviru holarktika Europe možemo razlikovati arktički, borealni, europski, odnosno azijski, euroazijski i mediteranski florni element u smislu geoelemenata, tj. na temelju recentnog areala pojedinih vrsta. Međutim, u smislu genoelementa, a s obzirom na naše geografsko područje arktičke i borealne vrste su glacijalni relikti, a to znači da su one zaostatak biljnog pokrova ledenog doba, dok je niz vrsta u našem dijelu Sredozemlja ostatak toplog tercijarnog razdoblja, pa ih zovemo tercijarni relikti. Među glacijalnim reliktima arktičkog podrijetla na mjestima gdje na Velebitu tijekom proljeća leži snijeg, susreću se npr. Viola biflora i Dryas octopetala (Rosaceae – grm) kao tipični glacijalni relikti. U smrekovim šumama našega pretplaninskog pojasa mogu se susresti npr. Clematis alpina, Bellidiastrum michelii kao borealni glacijalni relikti. U granicama Hrvatske nemamo Alpe, ali već u Sloveniji postoji izvjestan broj biljnih vrsta koje su rasprostranjene oko sjevernog pola i na Alpama, a to su arktoalpske vrste. U našem Sredozemlju koji je topao može se susresti i nekoliko vrsta zaostalih iz tercijara (tercijarni relikti), među njima se ističe gospin vlasak (Adiantum capilus-veneris, por. Polypodiaceae). Da je ta vrsta nekoga, davnog toplog razdoblja svjedoči nam činjenica da se je u unutrašnjosti Hrvatske i Bosne sačuvala samo oko izvora termalne vode uz rijeku Vrbas u Novom Šeheru pokraj Banja Luke. Adiantum capilus-veneris ja biljka polušpilja, gdje polagano kaplje voda i jedino takvo nalazište može se vidjeti na poluotoku Pelješcu u selu Dubrava.
Relikti
Relikti su biljne vrste koje su se u nekom području sačuvale na nekim za njih povoljnim mjestima pribježištima ili refugijama 10 iz razdoblja kada su u dotičnom području vladale druge ekološke prilike. Za ovaj dio Europe gdje je smještena Hrvatska i BiH, dva su razdoblja u širem smislu imala određeno značenje u oblikovanju biljnog pokrova. Starije razdoblje je konac tercijara (miocen 11-pliocen 12) pred 6-2 miliona godina i niz glacijacija 13
9 Relikti su stvar, pojava ili organizam sačuvan kao ostatak iz davnih vremena, a nalazi se još samo u malom broju na nekim ograničenim mjestima. 10 Refugije = lat. Utočište, sklonište, zaklon. 11 miocen = četvrta epoha tercijara 12 pliocen = gornji sloj terijara 13 glacijacija = ledeno doba u razvoju Zemlje. od pred 1 milijun do 12 tisuća godina. Tercijarno razdoblje značajno je po toploj i umjereno vlažnoj klimi, po prilici onakvoj kakva i danas vlada u tropima. Ledeno razdoblje značajno je po hladnoj klimi približno onakvoj kakva vlada u sjevernim dijelovima Euroazije, iznad šumske granice (područje tundre). Biljne vrste sačuvane iz razdoblja tercijara označavaju se kao tercijarni relikti, a biljke sačuvane iz ledenog doba nazivaju se glacijalni relikti. Kako Europa tijekom uzastopnih glacijacija izgubila najveći broj termofilnih biljnih vrsta to je ona vrlo siromašna tercijarnim reliktima. Najveće njihovo središte u Europi nalazi se u južnom dijelu Balkanskog poluotoka između Ohridskog jezera na sjeveru i planine Olimpa na jugu. Tu rastu npr. Aesculus hipocastanum, Dioscorea balcanica, Ramonda serbica, Haberlea rodopensis, Jankea heldreichii, Pinus peuce. Ovamo spada i Picea omorica, ali je ona rasprostranjena nešto sjevernije. Promatrajući globalno najveći broj tercijarnih relikata na području izvan tropskog dijela Holarktika sačuvao se je u zapadnom dijelu Sjeverne Amerike i u istočnoj Aziji. Razlog je tome što su planinski lanci Sjeverne Amerike i Središnje Azije smješteni transverzalno. Nastupom glacijacija kod merdijalnih planinskih lanaca biljke su mogle migrirati samo prema 61

jugu, dok su u Europi i Aziji migraciju zaustavile visoke planine i sve termofilne biljke sjeverno od transverzalnih planinskih lanaca morale su izumrijeti. Naime, nisu ih mogle prijeći i spustiti se na jug. Glacijalni relikti - Artičkog porijekla (Velebit) – Viola biflora, Dryas octopetala - Borealnog porijekla (Smrekove šume – pretplaninski pojas) Clematis alpina, Bellidiastrum michelii Tercijarni relikti - Adiantum capilus-veneris
Endemi Kako su se oblikovali i kako su nastali endemi? U ovom ćemo se poglavlju upoznati s endemičnošću i endemizmom, kao i s pregledom najvažnijih endema hrvatske flore. Pri proučavanju biljnog pokrova često se nameću pitanja: kako nastaju endemi? zašto endema negdje ima više, a negdje manje?, te napokon, što sve može uvjetovati pojavu i oblikovanje endema? Ako se jedna vrsta pojavljuje samo unutar nekog prirodno obilježenog (ograničenog) dijela Zemljine površine (npr. na nekom planinskom masivu, na otoku, ali i na čitavom kontinentu) ili unutar određenog floristički jedinstvenog područja, tada je ona tamo endemična. Takvi endemi mogu očito nastati tako da se ta vrsta razvila u tom području te se nije mogla dalje rasprostraniti (progresivni endemi), ali i tako da se neka vrsta koja je prije bila mnogo više rasprostranjena, zadržala se još ovdje, gdje zbog izolacije nije imala konkurencije ili joj je posebno odgovaralo stanište (konzervativni endemi). Pojavu ograničenja vrsta na određena floristički jedinstvena, većinom i fiziografski dobro omeđena područja, označavamo kao endemizam. Jedna od osnovnih poteškoća u objašnjenju postanka endema, između ostalog , leži i u činjenici da se endeme ne može proizvesti pokusima u laboratoriju, niti pokusima možemo dokazati kako je neki endem mogao nastati, jer je nemoguće u pokusu ponoviti sve one promjene odnosa kopna i mora, kao i sve promjene obličja zemljine površine (reljefa), koje su se događale tijekom zemljine prošlosti. Isto tako, paralelno s time i klimatske prilike su se umnogome promijenile, tako da je teško zamisliti kakva je stvarno bila klima pred kraj paleozoika ili tijekom mezozoika, u usporedbi s današnjom klimom. Da bismo, prema tome, mogli uopće shvatiti kako su se endemični oblici mogli razviti, osim općeg biološkog znanja, potrebno nam je i izvjesno geološko, geografsko i klimatološko predznanje. Danas je razmjerno dobro proučeno i objašnjeno kako se je tijekom geološke prošlosti oblikovalo današnje lice i izgled Zemlje. Smatra se da je kopneni dio Zemlje u početku bilo jedno jedinstveno kopno, nazvano Pangea, koje se je kasnije postepeno raspadalo na kontinente, a oni su zatim «putovali» i međusobno se razdvajali, da bi zauzeli današnji položaj. Što više, kontinenti još uvijek «putuju», po prilici 1 cm godišnje, pa se tako Afrika polako pomiče prema sjeveru i vrši pritisak na Eurazijsko kopno, pod njega se podvlači, a kao posljedica toga povremeno se javljaju jaki potresi. «Putovanje» kontinenata današnje južne zemljine polutke (hemisfere) bilo je daleko veće od putovanja kontinenata sjeverne polutke. Najveći put prevalile su Australija i Indijski potkontinent. Smatra se da je «indijski potkontinent» prešao najduži put, u razdoblju od 200

milijuna godina - naime, prevalio je čak 9000 kilometara, te svojim naslanjanjem na Aziju uvjetovao izdizanje planinskog lanca Himalaja. Ni polovi, a povezano s time niti ekvator Zemlje nisu uvijek bili tamo, gdje se nalaze danas. I oni su se znatno mijenjali, a to je dovodilo do vrlo velikih promjena klimatskih uvjeta na pojedinim dijelovima Zemlje. Kada bi neki dio Zemlje počeo dolaziti prema ekvatoru klima je u njemu postajala toplija, kada se je približavao polu njegova je klima postajala hladnija. Kakav su, dakle utjecaj imala navedena zbivanja na pojavu i razvitak endema? Sav onaj mnogoliki živi svijet - biljke i životinje- koji je u određeno vrijeme živio na jedinstvenom kopnu i činio jednu cjelinu, postepeno se je počeo razdvajati na manje dijelove. Tako su se na pojedinim novonastalim kontinentima nalazili oblici koji su se u vrijeme razdvajanja našli na svakom pojedinom dijelu, pa je već time izvršeno razdjeljivanje (diferencijacija) ukupnog fonda već postojećih živih bića i to je jedan od uzroka endemičnosti. Značajno je da upravo kontinenti južne polutke, kao npr. Južna Amerika, Afrika i Australija imaju veliki broj vlastitih endemičnih porodica, a svaka od njih ima veliki broj endemičnih rodova i vrsta. U slučaju da je neka skupina (primjerice, vrsta, rod) na jedinstvenom kopnu zauzimala veliko zemljopisno prostranstvo (areal) razdvajanjem kontinenata, na pojedinom novonastalom kontinentu su se našli samo dijelovi takva areala, oni su bili razdvojeni i nastavili su se dalje razvijati samostalno i neovisno jedan od drugoga. Došavši svaki od njih u uvjete drugačije od onih na zajedničkom kopnu, počeli su procesi specijalizacije koji su konačno kroz dovoljno dugo vremensko razdoblje doveli do stvaranja endemičnih oblika. Ako je odvojeno (izolirano) područje bilo veće, endemi su zauzeli veće prostranstvo, a ako je bilo manje zauzeli su manje prostranstvo, kao što je to slučaj sa stenoendemima .14
14 Stenos grč. = uzak, tijesan 15 Gondvana . pretpostavljeni velikikontinent na južnoj zemljinoj polutki u paleozoiku i mezozoiku. 16 Areal: područje rasprostranjenosti neke biljne skupine (taksona); takson sistematska jedinica bilja višeg ili nižeg reda. Kao dobro proučeni primjer mogli bismo navesti rod Nothofagus, srodnika našeg roda bukve (Fagus). Rodovi Fagus i Nothofagus počeli su se od svog zajedničkog pretka samostalno razvijati odvajanjem sjevernog kopna Laurazije od južnog Gondvane .15 Rod Fagus se širio sjevernom polukuglom, a rod Nothofagus južnom polukuglom. Danas su vrste roda Nothofagus rasprostranjene samo na južnoj hemisferi, a ograničene su samo na Južnu Ameriku s jedne strane, te Australiju s okolnim potkontinentom Novim Zelandom, Novom Kaledonjom, Novom Gvinejom i Tasmanijom s druge, dok je rod Fagus vezan isključivo na sjevernu hemisferu – Europu, Aziju i Sjevernu Ameriku. Planinski lanci što okružuju Sredozemno more zanimljiv su primjer formiranja mnogobrojnih endemičnih oblika znatno manje rasprostranjenosti. Možda je jedan od najslikovitijih primjera rod jela (Abies), koji je skoro na svakoj našoj usamljenoj i dovoljno velikoj sredozemnoj planini razvio posebnu endemičnu vrstu. Na većim planinskim sustavima rasprostranjenost je takve endemične vrste veća, a na manjim proporcionalno manja. Tako na međusobno povezanim planinskim sustavima Alpama, Dinaridima i Apeninima susrećemo običnu jelu (Abies alba) koja od svih sredozemnih vrsta zauzima najveće prostranstvo (areal 16). Ako krenemo u smjeru istoka susresti ćemo već na bugarskim i sjevernogrčkim planinama endemičnu bugarsku jelu Abies borisi-regis, na južnim grčkim planinama i Peloponezu . grčku jelu (Abies cephalonica), a na Kavkazu - kavkasku jelu (Abies nordmanniana). U sjeverozapadnom dijelu Male Azije na planinama Uludagu i susjednim planinama uz obalu Crnog mora raste bornmilerova jela (Abies bornmuelleriana), a na planinama povrh Troje, trojanska jela (Abies equi-trojani). Na planinama Cilicije, na Bliskom istoku raste endemična cilicijanska jela (Abies cilicica), na dijelu Atlasa prema Numidijskoj pustinji u

sjevernoj Africi živi numidska jela (Abies numidica), a iznad Giblatara marokanska jela (Abies marocana); na pirinejskom poluotoku raste španjolska jela (Abies pinsapo) i konačno na Nebrodima na Siciliji sicilijanska jela (Abies nebrodensis). Približimo li se još više našim krajevima, u bazen Jadranskog mora, možemo uočiti potpuno identični model, dakako s kombinacijom drugih endemičnih vrsta. Jedan od najzanimljivijih primjera je svakako rasprostranjenost endemičnih vrsta iz roda zvončića (Campanula) i to srodstvene skupine jednakolisnih (izofilnih – Isophyllae) zvončića. U sjevernohrvatskom, kvarnerskom primorju raste istarski zvončić (Campanula istriaca), na primorskim padinama Velebita raste prozorski zvončić (Campanula fenestrellata), na Biokovu i susjednim otocima Braču i Hvaru raste portenšlagov zvončić (Campanula portenschlagiana) i u dubrovačkom primorju raste pošarskov zvončić (Campanula poscharskiana). Znatno južnije, u kanjonu Crnog Drima pokraj Ohridskog jezera u Makedoniji raste debarski zvončić (Campanula debarensis), a na grčkom otoku Cefalinija u Jonskom moru raste cefalenski zvončić (Campanula cephalenica). Na suprotnoj strani Jadranskog mora, na Monte Garganu raste endemični garganski zvončić (Campanula garganica), a u Tiranskom primorju krhki zvončić (Campanula fragilis). Sve su se navedene vrste razvile najvjerojatnije pred kraj tercijara, trećeg doba u razvitku života na Zemlji, u razdoblju pred nekih 4 milijuna godina, kada se je vjerojatno suvisli areal praroditeljske vrste jednakolisnih zvončića pod utjecajem promjena ekoloških uvjeta i procesa izolacije uvjetovane promjenom reljefa oblikovao niz paralelnih (vikarnih, = pomoćnih zamjenik) vrsta razmjerno malene rasprostranjenosti. Pojava ledenih doba na sjevernoj polukugli, pred nešto više od milijun godina uvjetovalo je velike promjene u biljnom pokrovu Europe, što je također dovelo do pojave i razvitka endemičnih oblika. Svi navedeni i mnogi drugi slični primjeri oblikovanja usko srodnih vrsta koje se obično nalaze u području u kojem su nastale («in situ»), a nastale su u relativno bliskoj geološkoj prošlosti označavaju se kao mladi endemi (neoendemi). Drugu skupinu endema tvore one vrste koje su vrlo stare, vuku svoje porijeklo iz mezozoika ili pojedinih razdoblja tercijara, kada su bile široko rasprostranjene, kasnije su najvećim dijelom izumrle, a do danas se sačuvale samo ma malenim i ograničenim područjima. Takav je primjer nadaleko poznata vrsta Ginko biloba. Ta je vrsta pred nešto više od 200 milijuna godina bila široko rasprostranjena na sjevernoj polukugli, a tijekom vremena je sve više izumirala, da bi se danas sačuvala samo u istočnoj Aziji (Kina, Japan). Slična je situacija i s poznatom pančićevom omorikom (Picea omorica) koja je također, ali tijekom tercijara bila široko rasprostranjena u Europi, opisana pod imenom Picea omoricoides, a do danas se je sačuvala samo na nekoliko mjesta u gudurama središnjeg Balkana. I veliki broj zeljastih biljaka što rastu u našim bukovim šumama filogenetski su vrlo stare vrste, koje su negdje kod nas preživjele sve nepogode ledenih doba. One više nemaju bližnjih srodnika, a daljni su im srodnici rasprostranjeni tek u udaljenim krajevima Azije, ili uopće nemaju srodnika. Takve su vrste npr. velika mrtva kopriva (Lamium orvala) koja je bez srodnika, biskupska kapica (Epimedium alpinum) s daljim srodnicima na Kavkazu i u Sjevernoj Americi, bijeli kranjski bun (Scopolia carniolica) sa srodnicima u sjeveroistočnoj Aziji, praseće zelje (Aposeris foetida) uopće bez srodnika i još neke druge vrste. Sve su to zaostale (reliktne) vrste iz nekih drugih razdoblja, pa ih se označava kao stare endeme (Paleoendeme). Kako su te vrste zaostatak (relikt) neke davno izumrle flore, a ujedno su i endemične označavamo ih još i kao endemorelikti. Najveći broj paleoendema su istovremeno i endemorelikti. Tijekom oblikovanja endemičnih oblika naročito kod cvjetnica jednu od najvažnijih uloga odigrala je pojava poliploidije. Ona se sastoji u povećanju broja kromosoma, točnije umnožavanju osnovnog broja kromosoma (n) nekoliko do više puta (2n = 2x; 3n = 3x; 4n = 64

4x; 5n = 5x; 6n = 6x; 8n = 8x itd.), pa se od osnovnog diploida (2n) mogu nastati triploidi (3x) i tetraploidi (4x), od njih dalje pentaploidi (2x + 3x = 5x), heksaploidi (3x + 3x ili 4x + 4x), oktoploidi i napokon poliploidi gdje je broj kromosoma višestruko povećan. U slučajevima kad se broj kromosoma povećava (umnožava) unutar jedne populacije ili vrste govorimo o autopoliploidiji, a ako se broj kromosoma povećava križanjem dviju vrsta , govorimo o alopoliploidiji. Danas samim povećanjem broja kromosoma nova vrsta ne nastaje automatski. Naime, umnožavanje osnovnog broja kromosoma u prirodi je dosta česta pojava i uvjetovana je različitim čimbenicima. Ako ta pojava ne pridonosi boljoj prilagođenosti na odgovarajuće ekološke uvjete, individui s povećanim brojem kromosoma biti će eliminirani iz populacije. Nasuprot tome, ukoliko povećani broj kromosoma povećava podatljivost za neke korisne prilagodbe, broj poliploidniih individua se u populaciji povećava iz generacije u generaciju, pa u jednoj populaciji prevladavaju diploidni, a u nekoj drugoj poliploidni individui. Tada govorimo da je dotična vrsta zastupljena s dva ili više citotipova. To je osnovni preduvjet za oblikovanje endema poliploidijom. Kada nastupi potpuno razdvajanje (separacija) diploidnih i poliploidniih (tri-, tetra-, penta-, heksa-, itd. ploidnih) citotipova u zavisnosti od geografskih čimbenika dolazi do pojave geografski razdvojenih (izoliranih) citotipova i to je drugi korak u razvitku endema. Navedeni citotipovi već mogu predstavljati geografske rase, odnosno endemične podvrste. U novije vrijeme iscrpno je proučeno više poliploidniih kompleksa koji mogu poslužiti kao model geneze citotipova, geografskih rasa i poliploidniih vrsta. Kao takav primjer nastanka pojedinih vrsta procesima poliploidije i hibridizacije poliploida može poslužiti primjer kompleksnog roda stolisnika (Achillea millefolium). Osnovni broj kromosoma navedenog kompleksa n = 9. Vrste A. asplenifolia koja raste na vlažnim livadama panonskog prostora i A. setacea koja raste na stepama istočne Europe i zapadne Azije su diploidi i imaju 2n = 18 kromosoma, a može ih se u razvojnom (filogenetskom) smislu smatrati najstarijima. One su križanjem, kako se smatra, dale vrstu A. roseo-alba, koja može biti diploidni i tetraploini citotip, te vrstu A. collina koja je tetraploid i rasprostranjena je po suhim travnjacima toplijeg dijela Europe, nastalim čovjekovim djelovanjem (krčenjem šuma radi dobivanja pašnjaka). Kao slijedeći član poliploidnog niza je A. millefolium (obični stolisnik) koji je heksaploid (2n = 6x), a naseljava, također čovjekovim djelovanjem nastale livade i travnjake, te različita druga antropogena staništa. Kao zadnji član navedene serije razvija se oktoploidna (2n = 8x) vrsta A. pannonica. Oblikovanje endema može biti uvjetovano i razlikom u kemijskoj građi stijena povrh kojih rastu. Vulkanske i njima slične stijene građene su u osnovi od aluminosilikata i u sebi ne sadrže ni kalcij (Ca) niti magnezij (Mg). To su najstarije stijene i kada su biljke osvojile kopno, najprije su se naselile na silikatne stijene. Kasnije su se iz morskih prostranstava uzdigle mnogo mlađe taložne karbonatne i dolomitne stijene. Karbonatne – vapnenačke stijene građene su više –manje od čistog vapnenca (CaCO3), a dolomitne od smjese kalcijskog i magnezijskog karbonata (CaCO3 + MgCO3). Ustanovljeno je da su genetička varijabilnost, a isto tako i poliploidija pomogle biljkama silikatnih stijena da osvoje karbonatne stijene. Neke su to skupine mogle, a neke nisu. Tako vrste roda Azalea žive samo na silikatima, a vrste roda Rhododendron adaptirale u se i na karbonata. U Alpama poznati pjenišnici, alpski endem smeđi pjenišnik (Rhododendron ferrugineum) raste isključivo na silikatima, a jugoistočnoalpski endem dlakavi pjenišnik (Rh. hirsutum) raste isključivo na vapnencima, a koji na planinama u Hrvatskoj i BiH postiže svoju jugoistočnu granicu. Jedan od čestih preduvjeta formiranja endema jest i utjecaj vlažnosti, odnosno suhosti tla. Kao poučan primjer upravo su naše endemične vrste roda zvonca (Edraianthus). Dalmatinsko zvonce (E. dalmaticus) raste u močvarnim livadama Hrvatskog primorja, a uskolisno zvonce (E. tenuifolius) na kamenjarskim pašnjacima istog područja, pa govorimo da su se navedene endemične vrste razvile sinpatrički, tj. imaju «zajedničku domovinu». 65

Potpuno jednak primjer «sinpatrične diferencijacije» je u roda zlatica (Ficaria). Diploidna vrsta (2n = 16), primorska zlatica (F. calthaefolia) raste u suhim submediteranskim hrastovim šumama i na kamenjarskim pašnjacima, njezin je srodnik, ali tetraploid (2n = 32), velecvjetna zlatica (F. grandiflora) u poplavnim šumama Sredozemlja, odakle se je kasnije naselila u plitke odvodne kanale i uopće na poplavna i izrazito vlažna mjesta. Zahvaljujući tetraploidnosti F. grandiflora je znatno veća biljka od diploidne vrste F. calthaefolia, što je upravo česta pojava da su poliploidi gorostasniji od diploida. Iako su obje vrste zlatica rasprostranjene širom Sredozemlja, one su sredozemni endemi, a obje su zastupljene u Hrvatskom primorju. Poseban oblik sinpatriške diferencijacije je oblikovanje endema na serpentinima (smeđi mineralni kamen), tzv. serpentinofita. Oni se iz općeg fonda vrsta u procesu adaptivne evolucije specijaliziraju na posebne uvjete koje pruža serpentinska podloga. Iako u Hrvatskoj nema serpentina, ima ih već u susjednoj Bosni, gdje su serpentini vrlo rasprostranjeni i na njima raste više serpentinskih endema. Već iz nekoliko netom navedenih primjera možemo uočiti da su mogućnosti oblikovanja endema upravo nepregledne, pa područja bogata endemičnim oblicima, kao što je npr. naš Velebit, ukazuje nam na burna i mnogolika zbivanja tijekom dalje i bliže geološke prošlosti u tom dijelu Europe.
Endemične biljne vrste Hrvatske i Bosne i Hercegovine Na području hrvatske duž lanca primorskih Dinarida dva su dobro izražena genetička središta diferencijacije – velebitski i biokovski endemični centar (velebitski I biokovski stenoendemi). Isto tako maleni jadranski otoci i otočići oko Korčule značajna su mjesta diferencijacije, ali tu uz posebne ekološke faktore djeluje I faktor izolacije (tzv. Insularna diferencijacija). Iako ovdje ne bismo mogli navesti sve endemične vrste zastupljene u hrvatskoj flori, navest ćemo bar najvažnije od onih koje su u svojoj rasprostranjenosti ograničene isključivo na područje Hrvatske ili u hrvatskoj nalazi središte njihove rasprostranjenosti. Takve smo endeme rasporedili u nekoliko značajni skupina, ovisno o njihovom arealu. 1. Istarski stenoendemi: Campanula justiana -justinov zvončić Campanula tommasiniana -tomazinijev zvončić 2. Kvarnersko-liburnijski endemi: Aristolochia croatica -hrvatska vučja stopa Campanula istriaca -istrski zvončić Centaurea dalmatica -dalmatinska zečina Gentianella liburnica -liburnijska sirištara Iris illyrica -ilirska perunika Leucanthemum platylepis -liburnijska ivančica 3. Ilirsko-jadranski endemi: Alyssanthus sinuatus -naduta gromotuljka 66

Alyssum litorale -primorska gromotulja Astragalus muelleri -krčki kozlinac Campanula pyramidalis -piramidalni zvončić Centaurea ragusina -dubrovačka zečina Centaurea rupestris -žuta krška zečina Corydalis acaulis -primorska kacigača Crataegus transalpina -primorski glog Dianthus dalmaticus -dalmatinski karanfil Edraianthus tenuifolius -uskolisno zvonce Festuca illyrica -ilirska vlasulja Frangula rupestris -primorska krkavina Genista dalmatica -dalamtinska žutilovka Helleborus multifidus -mnogoperni kukurijek Inula verbascifolia -jadranska slavulja Rhamnus intermedia -plošnasti pasjak Salvia officinalis -ljekovita kadulja Satureja montana -ljekovita kadulja Satureja subspicata -ružičasti vrisak Viola adriatica -jadranska ljubica 4. Velebitski stenoendemi: Arenaria orbiculata -paklenička pješčarka Campanula fenestrellata -prozorski zvončić Campanua velebitica -velebitski zvončić Cardaminopsis croatica -hrvatsak parežuha Degenia velebitica -velebitska degenija Dianthus bebius -bijeli velebitski karanfil Euphorbia triflora -trocvjetna mlječika Dianthus velebiticus -crveni velebitski karanfil Iberis velebitica -velebitska ognjica Knautia velebitica -velebitska prženica Laburnum alschingeri -velebitska zanovjet Leucanthemum croaticum -hrvatska ivančica Thalictrum velebiticum -velebitski vrednovac 5. Biokovski stenoendemi: Astragalus angustifolius subsp. biokovoensis -biokovski kozlinac Centaurea biokovoensis -biokovska zečina Centaurea cuspidata -osjava zečina Cerinthe tristis -tužna visika Edraianthus pumilio -biokovsko zvonce Minuartia clandestina -biokovska pjeskarnica 6. Dalmatinsko-otočni stenoendemi: Asperula staliana -biševska lazarkinja Asperula visianii -vizianijeva lazarkinja Aurinia scopulorum -kamnička gromotulja 67

Brassica botteri -palagruški kupus Brassica mollis -maljavo zelje Centaurea friederici -palagruška zečina Centaurea jabukensis -jabučka zečina Centaurea crithmifolia -Zelena jabučka Limonium vestitum -jabučka mrižica 7. Dalmatinski endemi: Campanula portenschlagiana - portenšlagov zvončić Campanula poscharskiana - pošarskov zvončić Edraianthus dalmaticus -dalmatinsko zvonce Iris adriatica -jadranka perunika Iris pseudopalida -dalmatinska perunika Pevalekia triquetra -tribridna pevalekija Pinus dalmatica -dalmatinski crni bor Salvia brachyodon -kratkozuba kadulja 8. Ilirsko-dinarski (planinski) endemi: Arabis scopoliana -skopolijeva gušarka Carex kitaibeliana -kitajbelov šaš Euphrasia illyrica -ilirska očanica Leucanthemum chloroticum -blijeda ivančica Lonicera borbasiana -borbaševa kozokrvina Lonicera glutinosa -ljepiva kozokrvina Micromeria croatica -hrvatski čubar Pedicularis acaulis -prizemni ušljivac Primula kitaibeliana -lički jaglac Pulmonaria visianii -vizianijev plućnjak Rhamnus fallax -ilirski pasjak Ribes pallidigemum -blijedi ribiz Serratula cetinjesis -cetinjska pilica Sesleria juncifolia -sitolisna šašika Silene marginata -pušina točilarka Teucrium arduini -arduinov dubačac U kopnenom dijelu Hrvatske endemičnost flore je manja, a među najznačajnije endeme mogli bismo uvrstiti skupinu. 9. Jugoistočni endemi: Centaurea fritschii -fričeva zečina Dianthus croaticus -hrvatski karanfil Doronicium croaticum -hrvatski divokozjak Helleborus atrorubens -tamnocrveni kukurijek Helleborus niger -kukurijek božičnjak Iris croatica -hrvatska perunika Scopolia carniolica -kranjski bijeli bun Sesleria kalnikensis -kalnička šašika 68

Neki od važnijih BiH endema: Prenjska kamenika -(Saxifraga prenja) Frajnov karanfil -(Dianthus freynii) Bosanska perunika -(Iris bosanica) Bosanska zvončika -(Symphyandra hofmanni) Bosanski ljiljan -(Lilium bosanicum) Hercegovački zvončić -(Campanula hercegovina) Gregersenova mlječika -(Euphorbia gregersenii) Nježna kockavica -(Fritillaria gracilis) Dinarska kandilka -(Aquilegia dinarica) Vraničko zvonce -(Edraianthus niveus) Bekova ljubica -(Viola beckiana) 69

NAČINI ŠIRENJA BILJNIH VRSTA I OBLICI AREALA
Težnja je svakog živog bića da se proširi na što većem prostoru. Tamo gdje ima takvih uvjeta to je širenje lagano i brzo. Međutim u prirodi takvi uvjeti praktički ne postoje. U pravilu postoji borba velikog broja potomaka za isti prostor i iste izvore hrane s time da već unaprijed ne postoje uvjeti za preživljavanje svih. Kod životinja, naročito ptica osvajanje novih prostora mnogo je lakše, jer postoji manji broj potomaka i veća mogućnost savladavanja prostora. Kod kopnenih biljaka koje su u pravilu pričvršćene za podlogu (ukorijenjene) ne postoji mogućnost aktivnog kretanja i savladavanja prostora. Da bi se biljke ipak «kretale» u tu svrhu proizvode niz različitih dijaspora pomoću kojih također mogu savladati prostor. Dijaspore mogu biti sjemenke, plodovi, rasplodni pupovi, gomolji, lukovice i sl. Za širenje biljnih vrsta i formiranje areala svakako su najvažnije sjemenke i plodovi. Biljke imaju sposobnost razbacivanja svojih sjemenaka vlastitom aktivnošću, ali je ta pojava izuzetno rijetka i naziva se autkorija. Takva je npr. štrcalica (Ecbalium allaterium) ili nedirak (Impatiens noli-tangere). Mnogo je češći slučaj da biljke za širenje svojih dijaspora koriste neke prirodne sile koje mogu biti biotske i abiotske i to je alokorija. Biotske sile su životinje, a širenje pomoću životinja zove se zookorija. Ona može biti dvojaka; onda kada životinje pojedu plodove, sjemenke prolaze kroz njihov probavni trakt, tada govorimo o endozookoriji. Te biljke imaju u pravilu plodove koštunicu ili koštuničavu bobu. Endokarp takvih plodova stvara čvrstu ljusku (košticu) koja obavija sjemenku. Taj je omotač tako čvrst da probavni sokovi životinja razgrade djelomično samo vanjski dio, tako da nakon izlaska iz probavnog trakta sjemenka može nicati. Ako sjemenka nije prošla kroz probavni trakt životinja ne može nicati. Kao primjer navodimo trešnju (Prunus avium). Kada plodovi ili sjemenke nose na sebi kukice ili zupce različita oblika s kojima se mogu prihvatiti za krzno ili runo životinja govorimo o epizookoriji, kao npr. čičak (Actium tomentosum, A. lappa), bročika (Galium aparine) i dr. Mnoge vodene biljke mogu se prilijepiti za noge ptica močvarica pa se tako mogu raširiti na vrlo velike udaljenosti. Tako mesožderna biljka (Aldrovanda vesiculosa) sa svojim područjem rasprostranjenosti u središnjem dijelu južne Afrike ima u Europi rasprostranjenost vrlo sličnu rasprostranjenosti ptice rode. Naime, europske rode odlaze na zimovanje upravo u dijelove južne Afrike. Slično se šire i neki sočni plodovi sjemenaka koje ne smiju proći kroz probavni trakt životinja, kao npr. agrumi (Cistus), jabuka (Malus) i dr. Ti su plodovi tako veliki da ih životinje ne mogu niti izgristi niti progutati cijele, već ih na nekom skrivenom mjestu trgaju i komadaju, a pri tome komadanju sjemenke se obično razlete u okolicu. Kada sjemenke i plodove šire ptice to se naziva ornitokorija, a kada ih šire mravi zove se mirmekokorija. Takve sjemenke imaju na sebi naročito masno tjelašce – elajosom kojim se mravi hrane. U našim geografskim širinama to je najčešće slučaj kod rodova kacigača (Corydalis) i ljubica (Viola) Geokorija je kada biljka sama zasijava svoje sjemenke u zemlju, kao npr. kikiriki (Arachis hypogea), djetelina (Trifolium subterraneum) ili u pukotinu zida (Cymbalaria muralis). Barikorija – plodovi koji se šire pomoću vlastite težine, npr. jabuke se kotrljaju niz padinu, žir hrasta (Quercus) i itd. Hidrokorija plodovi koji su unutrašnjosti šuplji, pa plivaju na vodi, kao npr. tikva (Cucurbita), vodeni orah (Trapa natans), kokosov orah (Coccos nucifera) koji može izdržati više godina u moru, jer ima plutasti (spužvasti) omotač i tako može prevaliti više tisuća milja. Antropokorija – je širenje biljnih vrsta posredstvom čovjeka. To je vrlo uobičajeni način širenja mnogobrojnih biljnih vrsta iz jednog kraja svijeta u drugi, iz kontinenta na itd. 70

Sve takve biljne vrste koje su se izvan svoga prirodnog areala raširile zahvaljujući čovjekovoj aktivnosti nazivaju se antropokore. Kako se čovjekova aktivnost može sustavno pratiti od neolita pa tijekom povijesnog doba sve do naših dana možemo antropokore podijeliti na arheofita, paleofita, neofita i neotofita. Arheofiti – su se širili u pretpovijesno doba i kada bismo ih htjeli fitografski definirati, najčešće bi pripadali azijskom geoelementu, a kako su se udomaćili u Europi najčešće ih označavaju kao Euroazijsku skupinu vrsta. Paleofiti – su skupina biljnih vrsta koje je širio čovjek tijekom povijesnog doba sve do otkrića Amerike. Neofiti – otkriće Amerike je početak Novog vijeka pa se one biljne vrste različitog podrijetla koje su se širile od tada, pa do drugog svjetskog rata označavaju kao neofiti. One podrijetlom iz Amerike su amerikanoneofiti, iz Indije – indikoneofiti, iz Turske – turcikoneofiti, kao npr. iđirot (Acorus calamus). Neotofiti su biljke koje se Europom šire poslije II. Svjetskog rata. Za hrvatsku floru to su npr. Solanum elaeagnifolium, S. sodomeum, Cyperus rotundus. Efemerofiti – Često se događa da pojedini neofiti, naročito jednogodišnji, negdje niknu i budu zabilježeni kao pridošlice, pa se nakon nekoga vremena izgube, a onda od odbačenog sjemena opet niknu i pojave se. Takve biljke koje se u nekoj flori povremeno pojavljuju nazivamo efemerofiti, kao primjer spomenimo rajčica (Lycopersicum esculentum) i lubenicu (Colocyntis citrulus).
AREAL
Kada bi površina Zemlje bila ravna ploča bez razlika u ekološkim čimbenicima pojedinih njezinih dijelova, areali pojedinih biljnih vrsta bili bi pravilni krugovi, slično kao što su pravilni krugovi kolonije čistih kultura na hranidbenoj podlozi. Kako Zemlja ima određenu orografiju (reljef), a ni ekološki uvjeti nisu jednaki, čak i na manjim razdaljinama, areali se biljnih vrsta oblikuju prema mogućnostima koje dotičnoj vrsti pruža teritorij i ekološki uvjeti. Ako je npr. neka biljka značajna za slatkovodne bazene nazivamo ih hidrofiti. Ako je primjerice biljka u potpunosti obrasla bazen, onda areal takve biljke ima oblik dotičnog bazena. Ako na nekom kopnu postoji više odvojenih vodenih bazena, tada vodena biljka ima raspršeni ili disperzni areal. Ako se sveukupni areal neke vrste sastoji od nekoliko ili više odvojenih dijelova, govorimo o disjunktnom arealu. Svi takvi areali mogu se označiti i kao diskontinuirani areali. Nasuprot tome, areali onih šumskih vrsta (drveta) koje homogeno obrašćuju veća ili manja područja, ali se izvan njih više ne pojavljuju, onda govorimo o suvislom ili kontinuiranom arealu. Taj areal je još i homogen, jer se individui praktički nalaze jedan do drugog. Međutim, kontinuirani areal i ne mora biti homogen, jer individui zbog svojih bioloških značajki ne rastu jedan do drugog, ali kontinuirano zauzimaju jedno područje. Često se puta događa da je neka vrsta šire rasprostranjena na nekom području i stvara kontinuirani areal, ali osim toga u nekom drugom području stvara neki mali kontinuirani areal. Tu pojavu zovemo disjunkcija, a areali su disjunktni. Vrlo mnogo disjunktnih areala imaju jadransko-apeninske endemične vrste. Tako npr. oman ili slavulja (Inula verbascifolia) obuhvaća veći dio istočnojadranskog primorja sa središtem u Hrvatskom primorju, a isto tako na Monte Garganu u Italiji ima jednu disjunkciju. 71

Već je odavno postavljena teorija da je nekada postojalo jedinstveno kopno na Zemlji koje se kasnije postepeno raspadalo u kontinente. Ta se teorija morala dokazati činjenicama zasnovanim na znanstvenim istraživanjima. Rezultatima istraživanja na raznim područjima prirodnih znanosti sve se više podupiru ta teorija. U posljednje vrijeme su se pridružili i rezultati biljnogeografskih usporednih istraživanja na raznim kontinentima. Utvrđeno je, naime da postoji sličnost u biljnom sastavu pojedinih kontinenata, a ta se sličnost može utvrditi na osnovu bližeg ili daljeg srodstva pojedinih biljnih predstavnika na raznim kontinentima. O ovome će biti riječi nešto kasnije. Novija istraživanja posebno ona o paleomagnetizmu, te rezultati istraživanja i sondiranja dna Atlantskog oceana idu u prilog teoriji o horizontalnom pomicanju kontinenata. Međutim, danas se smatra da do pomicanja dolazi uslijed endogenih sila što vladaju u unutrašnjosti Zemlje, a ne zbog «plivanja» pojedinih kontinenata. Osim tog gledišta o jednom jedinstvenom kopnu postoji općenito prihvaćena spoznaja da su još početkom mezozoika, odnosno u trijasu postojala dva velika kontinenta, i to sjeverno kopno – Laurazija i južno kopno – Gondwana, koji su međusobno razdvojeni mediteranskom geosinklinalom18, ali su povremeno i mjestimično bili međusobno spojeni. Takve su prilike morale vladati bar do sredine mezozoika, srednjega doba u geološkom razvoju Zemlje. 18 Geosinklinala – labilan, udubljen morem pokriven prostor Zemljine kore između kontinentskih blokova; isp. geoantiklinala. Danas raspolažemo s nizom zanimljivih florističkih i faunističkih podataka o rasprostranjenosti mnogih biljnih i životinjskih skupina, čije se rasprostranjenje praktički ne bi moglo objasniti na neki drugi način, osim postojanja jednog više-manje jedinstvenog kopnenog kompleksa od kojeg su pucanjem i razmicanjem nastali pojedini kontinenti. Moderna gledišta filogenije najsavršenijih stablašica – cvjetnica ili kritosjemenjača – to još više potvrđuju. Na današnjem stupnju znanosti o evoluciji viših biljaka općenito je prihvaćeno gledište da su se najsavršenije stablašice – cvjetnice ili kritosjemenjače (Magnoliophyta (Anthophyta, Angiospermae) morale razviti svakako prije donje krede (mezozoik), jer već u donjoj kredi ili nešto kasnije, u gornjoj kredi, postoje takvi oblici u kojih direktne potomke nalazimo još i danas u biljnom pokrovu Zemlje. Dakle, tijekom krede postojale su visoko organizirane cvjetnice. Širenje najvećeg broja skupina kopnenih biljaka na površini Zemlje zavisi u prvom redu o reljefu i o klimatskim prilikama, a zatim o osobinama geološke podloge (npr. silikati i karbonati). Kada se tijekom geološkog razvoja Zemlje jedan dio kopna odvojio bilo na koji način i ostao izoliran, biljni svijet koji se tada na njemu našao, ostao je «zarobljen» i dalje se razvijao sam neovisno o ostalim kopnenim masama. Proces evolucije nije se zaustavio niti na jednom odvojenom dijelu, već teče dalje , ali neovisno jedan od drugoga, pa nakon nekog vremena dolazi do razlika između predstavnika pojedinih izoliranih kopnenih dijelova. Što se je izolacija prije dogodila to su razlike između prvotno identičnih ili srodnih biljnih skupina sve veće i veće. Na brzinu evolucijskog procesa utječe i stupanj filogenetskog razvoja i stupanj specijalizacije pojedinih biljnih skupina. Evolucija teče brže kod skupina višeg stupnja evolucije – dakle kod filogenetskih mlađih oblika, te kod oblika na nižem stupnju specijalizacije, tj. kod nesavršenih, nespecijaliziranih oblika. Velike kontinentalne disjunkcije, naročito veza Južna Amerika – Afrika bile su pobudom njemačkom meteorologu Wegeneru da postavi hipotezu o horizontalnom razmicanju kontinenata Danas se zna da je proces evolucije na sjevernoj hemisferi tekao od kraja tercijara, tijekom kvartala, dakle u razdoblju od posljednjih milijun godina, neobično brzo. Od tada je naime nastupom ledenih doba veza između Sjeverne Amerike i Eurazije, koja je postojala

preko današnjega sjevernog pola, bila prekinuta, pa se mnoge biljne skupine koje su se pred nastupom leda povlačile prema jugu, na područja središnjeg dijela Sjeverne Amerike i istočnu Aziju, a vrlo rijetko i u Europu, razvijaju potpuno samostalno. U nekih je skupina evolucijski proces išao brže pa je dostigao stupanj roda (npr. rod Thuja u Sjevernoj Americi i rod Biota u istočnoj Aziji (sl. 40), a negdje sporije pa je dostigao samo razinu vrste (npr. Platanus occidentalis u Sjevernoj Americi i P. orientalis u Europi i Aziji – sl. 41), ili samo razinu podvrste (npr. Anemone nemorosa subsp. quinquefolia u Sjevernoj Americi, A. nemorosa subsp. nemorosa u Europi i A. nemorosa subsp. amurensis u istočnoj Aziji – sl. 42.) Sličnih primjera ima mnogo, još neke od njih donosimo u slijedećoj tablici: Europa Sjeverna Amerika Ostrya carpinifolia Ostrya virginiana Ulmus minor (=U. campestris) Ulmus fulva Ulmus laevis Ulmus americana Sorbus aucuparia Sorbus americana Pinus sylvestris Pinus banksiana Pinus peuce Pinus strobus Viburnum lantana Viburnum lantanoides Viburnum opulus Viburnum trifidus Oxalis acetosella Oxalis montana Vallisneria spiralis Vallisneria americana Maianthemum bifolium Mainthemum canadense Polypodium vulgare Polypodium virginianum Skoro da i nema biljnog roda a kamo li porodice koja ne bi imala predstavnika istovremeno i u Sjevernoj Americi s jedne strane i u Euraziji s druge strane. To je uočeno već davno pa je najveći dio sjeverne hemisfere zbog velike srodnosti svoje flore ujedinjen u jedno jedinstveno florno carstvo – holarktis. Sl. 40. Karakteristično rasprostranjenje međusobno srodnih rodova Thuja (Sjeverne Amerike) i Biota (Jugoistočna Azija) na sjevernoj hemisferi.

Sl. 41. Rasprostranjenost nekih vrsta roda Platanus: P. occidentalis (Sjeverna Amerika) i P. orie
ntalis (Europa – Azija). To je izrazita tercijarna disjunkcija.

Sl. 42. Varijabilnost vrste Anemone nemorosa, uvjetovana izolacijom poslije tercijara (Pleistocen – holocen): A. nemorosa subsp. quinquefolia (Sjeverna Amerika); A. nemorosa subsp.nemorosa (Europa); A. nemorosa subsp. amurensis (Istočna Azija). Situacija u južnoj polukugli znatno je drugačija. Iste takve velike disjunkcije kao i u sjevernoj polukugli povezuju dijelove Južne Amerike, južne Afrike i Indomalajskog arhipelaga. Pokazalo se je da niz vrsta i rodova Južne Amerike raste na istočnoj izbočini obale, a u Africi upravo tamo, gdje se na njezinoj zapadnoj obali nalazi udubljenje obalne linije (sl. 43).


Sl. 43. Nekoliko značajnih biljnogeografskih primjera međusobne paleogeografske povezanosti Južne Amerike i Afrike: lijevo, rekonstrukcija; desno, stvarno stanje. A – Areal roda Sacoglottis 75

B – Areal roda Aptandura C – Areal roda Maprounea D – Areal roda Chrysobalanus Prvotno jedinstveno južno kopno - Gondwana raspada se na niz kontinenata (Južnu Ameriku, Afriku, Indomalajski dio Azije, Australiju s Novim Zelandom i Antarktik). Na temelju florističkih podataka taj je proces otpočeo vrlo davno već krajem mezozoika, pa se na svakom tako razdvojenom i izoliranom kontinentu razvija poseban i za njega svojstven biljni svijet. Stoga fitogeografi čitavu južnu hemisferu zajedno s ekvatorijalnim dijelom sjeverne hemisfere dijele u više samostalnih flornih carstava (sl. 46). Iako flore pojedinih kontinenata južne hemisfere razlikuju se tako jako da predstavljaju samostalna florna carstva, u njihovu sastavu nalazimo i razmjerno veliki broj takvih biljnih skupina (kritosjemenjača, a i golosmjemenjača) prvenstveno na razini porodica i rodova, vrlo rijetko vrsta, koje su na pojedinim kontinentima tako karakteristično raspoređene da nam tu pojavu može objasniti samo hipoteza o jednom jedinstvenom kontinentu, koji se postepeno lomio i raspadao pa su jedne biljne skupine zajedničke Južnoj Americi i Africi, druge Africi i Australiji, treće Južnoj Americi i Australiji itd. Na slici 43 prikazana je veza Južna Amerika – Afrika i povezanost Južne Amerike, Afrike i Indomalajskog dijela Azije (sl. 44). Izvjestan broj biljnih skupina ograničen je u svom rasprostranjenju na Zemlji samo na dijelove Južne Amerike i Afrike. Najzanimljivije je da se dijelovi takvih areala na spomenutim kontinentima međusobno skoro idealno podudaraju, što se najbolje može vidjeti na slici 43.


Sl. 44. Nekoliko primjera karakterističnih areala koji pokazuju biljngeografsku povezanost Južne Amerike, Afrike i Indomalajskog dijela Južne Azije, kao odraz starih paleogeografskih prilika tog dijela Zemlje. A – areal roda Xylopja; B – areal roda Buddleja; C – areal porodice Styracaceae; D – areal roda Chaetocarpus. 77

Kao daljnji primjer možemo spomenuti disjunkcije koje povezuju Južnu Ameriku – Australiju (Novi Zeland, Nova Gvineja). Svakako najneobičnije rasprostranjenje imaju malobrojni oblici koje danas nalazimo lokalizirane uglavnom na jugozapadnoj obali južne Amerike i na istočnoj obali Australije, odnosno na Novom Zelandu ili u Novoj Gvineji (sl. 45).


Sl. 45. Karakteristično rasprostranjenje nekih vrlo starih oblika koji su bili ograničeni na rubni pojas Gondwana -kontinenta. Areali su vrlo isprekidani osobito na potezu Australija (s Novom Zelandom, Tasmanijom i Novom Gvinejom), jer je Antarktik došao u nepovoljni položaj za opstanak najvećega broja životnih oblika, što je dovelo do tako velike disjunkcije. A- areal roda Embothrium; B – areal roda Nothofagus; C – areal roda Araucaria; D – areal roda Coriaria. Postoji veliki broj ekosustava koje možemo podijeliti u dvije velike cjeline:
Kopneni ekosustavi - to je skup ekosustava gdje je okolni zrak prirodni okoliš
Vodeni ekosustavi – to je skup ekosustava gdje je voda prirodni okoliš
Kopnene zajednice
I. Terestički BIOMI
BIOMI – NAJVEĆI TIPOVI VEGETACIJE NA ZEMLJI Biomi (ekozone ili ekoregije) - čine makro biocenoze ili velika geografska područja ekozona na Zemlji, gdje makroklimatski uvjeti određuju tipove ekoloških zajednica koje ih izgrađuju. Biome čine različite skupine kopnene biljnih i životinjskih zajednica prilagođenih na zajedničke makroklimatske uvjete. Biom čini specifičan spoj biljaka i životinja koje su prilagođene životu u određenim uvjetima u okolini, među kojima klima ima najznačajniji utjecaj. Npr. grafički prikaz kopnenih bioma sa srednjom godišnjom temperaturom zraka i srednjom godišnjom količinom oborina ima specifičan obrazac (slika x). Na slici x prikazane su jasne granice između bioma, međutim treba imati na umu da se biomi postupno izmjenjuju. Također, iako ćemo opisati svaki biom posebno treba znati da je svaki biom povezan sa svakom kopnenom i vodenom zajednicom u biosf eri.


Slika xx. Biomi najveći tipovi vegetacije na Zemlji Slika xx1. Odnos između nadmorske visine, geografske širine i bioma . Veća nadmorska širina, klima je hladnija. Čak i u tropskim područjima vrhovi visokih planina mogu zadržavati snijeg tijekom cijele godine. Tako je moguće susretati se s istim tipovima vegetacije idući u visoke planinske lance, ali i krećući se od ekvatora prema polovima (od manje prema većoj geografskoj širini)

Slika xx2. Utjecaj prosječne godišnje količine oborina i prosječne godišnje temperature na razvoj bioma na Zemlji. U područjima s niskim oborinama i niskim temperaturama razvijaju se tundre. S druge pak strane u područjima s visokom količinama oborina i visokim temperaturama razviaju se tropske kišne šume. Slično možemo navesti i za sve ostale biome.
Rasporostranjenost bioma na Zemlji vezani su uz osnovna obilježlja makroklime i njezinim razlikama, čime su određeni i uvjeti života na Zemlji, odnosno već opisanom količinom sunčevog zračenja ovisno o geografskoj širini i nadmorskoj visini kao i topografskim obilježjima (Slika xx1). Dakle, utjecaj temperaturnog gradijenta može se vidjeti ne samo s geografskom širinom već i s nadmorskom visinom. NA svakih 100m nadmorske visine temperatura opada 1oC. Idući od ekvatora prema Sjevernom polu prvo nailazimo na tropske kišne šume, slijede listopadne šume umjerenih područja, pa redom šume četinjača i tundre. Gotovo isti slijed možemo uočiti uspinjanjem na visoke planine. Šume četinjača u planinama nazivamo planinske četinjače, a tundre blizu planinskih vrhova nazivaju se alpske tundre. Međutim, s druge pak strane idući od prema Južnom polu, nećemo sresti područja koja odgovaraju šumama četinjača i tundri sjeverne polutke, jer se kopno na južnoj polutki nalazi smješteno više prema sjeveru. Rasprostranjenost bioma na Zemlji određena je makroklimatskim paramertima kao što je prosječna godišnja temperatura i prosječna godišnja količina oborina, a koje variraju prema geografskim širinama i nadorskim visinama. Biomi su klimatski i geografski određeni prema sličnim klimatskim uvjetima koji vladaju na Zemlji, a također ih obilježavaju karakteristične zajednice biljaka, životinja kao i obiježlja tla, zbog čega se biomi definiraju kao veliki ekosustavi ili ekozone odnodno ekoregije.

Neki dijelovi Zemlje imaju više ili manje iste ili slične biotičke ili abiotičke čimbenike koji su prošireni na velikom području stvarajući tipični ekosustav karakterističan za to područje. Takvi veliki ekosustavi nazivaju se biomi. Biomi se često uspoređuju s pojedinim obrascima ekološke sukcesije i klimaks vegetacije (prividnim ravnotežnim stanjem lokalnih ekosustava). Ekosustav uključuje veliki broj biotopa, a biom u cjelini predstavlja glavni stanišni tip. Biološka raznolikost karakterizira svaki biom, a posebno raznolikost flore i faune, ali uključujući i manje dominantne biljke i životinje. U kopnenim biomima, biološka raznolikosti biljnih vrsta često se odražava na veću neto primarnu proizvodnost, što je u uskoj vezi s dostupnosti vlage i većim temperaturama. U osnovi biome možemo podijeliti na: 1. Terestičke /kopnene biome, i 2. Akvatičke biome (uključujući slatkovodne i morske biome) Kao što je već istaknuto, makroklima je glavni čimbenik koji određuje rasprostranjenost terestičkih bioma. Među važnije klimatske čimbenike možemo ubrojiti: 1. Geografsku širinu: Arktik, sjeverni klimatski pojas, umjereni klimatski pojas, suptropski pojas i tropski pojas, 2. Vlažnost: vlažno, polu-vlažno, polu-sušno i sušno područje. 3. O sezonske varijacije: Oborine mogu biti ravnomjerno raspoređena tijekom cijele godine ili mogu postojati sezonska razdoblja bez kiše, to jest sušna razdoblja, kao npr u Sredozemlju gdje su izražena suha ljeta i vlažne zime: Većina regija na Zemlji dobivaju većinu svojih oborina tijekom ljetnih mjeseci, dok u Sredozemnom Bazenu većina oborina dolazi tijekom zimskih mjeseci. 4. Nadmorska visina, povećanje nadmorske visine uzrokuje raspored stanišnih tipova slično kao povećanje geografske širine. Biološka raznolikost se općenito povećava od polova prema ekvatoru i raste s porastom vlage. Biomi su široko rasprostranjeni ekosustavi, koji se odlikuje određenim dominantnim tipovima vegetacije kao na primjer: tundra, tajga, tropska kišna šuma, stepa, savana, bjelogorične i crnogorične

šume

Slika xxc razvoj bioma u odnosu prema klimatskim uvjetima na zemlji
1. Tundra
Tundra je biom koji se prostire u predjelima, gdje je rast drveća onemogućen nižim temperaturama i kratkom vegetacijskom sezonom. Ime potječe od kildin-samske (laponske) riječi "tūndâr", koja označava "ledinu bez drveća". Postoje tri tipa tundre: (i) arktička, (ii) antarktička i (iii) alpska. Flora palearktičkih1 tundri često se naziva arkto-alpskom uslijed postojanja zajedničke povijesti vegetacije i velikog broja zajedničkih vrsta biljaka u arktičkoj, antarktičkoj i alpskoj tundri. Biom artičke tundre okružuje Zemlju malo južnije od ledom prekrivenog polarnog mora na sjevernoj polutki, te prekriva 20% zemljine površine. Slična zajednica zvana alpska tundra nalazi se iznad planinskih područja. Artička tundra je hladna i mračna tijekom većeg dijela godine. S obzirom da godišnja količina oborina iznosi svega 200 mm, tundra bi se mogla smatrati hloadnom pustinjom, no topljenje snijega uzrokuje nastanak vodenih

bazena tijekom ljeta. U tundi se voda zadržava jer je isparavanje iznimno malo. Samo površinski sloj tla se odmrzava tijekom ljtnih mjeseci, a ispod tog sloja se nalazi permafrost koji je uvijek smrznut, pa je stoga procjeđivanje vode minimalno. 1 Palearktička regija je staro-sjeverno područje (palearktička regija). U razdoblju terercijar-kvartar kopnena veza između sjeverne amerike i euroazije egzistirala je u području beringovog prolaza i grendlanda. Šuma u tundrama nema jer je vegetacijska sezona prekratka, njihovo korjenje ne može prodrijeti u permafrost, i ne mogu se ukorjeniti u blatnom tlu tijekom ljeta. Ljeti tlo je prekriveno

niskom travom i trskom te područjima s lišajevima i mahovinom. Patuljasti drveni grmovi, kao što je patuljasta breza ( Betula papyrifera), cvatu i osjemenjuju se brzo dok ima dovoljno sunca za fotosintezu. Nekoliko vrsta životinja živi u tundrama preko cijele godine. Npr. mišu sličan leming ostaje ispod snijega; tetrijeb se zakopava u snijeg za vrijeme oluja; a mošusno govedo čuva toplinu zahvaljujući debelom krznu te kratkom i zdepastom tijelu. Ljeti tundre preplave brojni insekti i ptice, posebno obalne ptice, guske i patke koji kreću u unutrašnjost. Jeleni i srne također migriraju u i iz tundre, kao i vukovi koji tamo love, a polarni medvjevi su česti uz obalu. Slika.xxx. Biom tundra Slika xxc. Patuljasta breza ( Betula papyrifera)
(Ovibos moschatus) mošusno govedo

1.1. Arktička tundra
U skorijoj geološkoj prošlosti (ledena doba kvartara) velike ledene mase pokrivale su dio Sjeverne Amerike i Europe. U periodu poslije posljednje glacijacije led se povukao prema sjeveru ili se zadržao na planinama (ledenjaci). Područja koja su posljednja oslobođena leda, a susjedna su Slika. Xcx Arktička tundra, Aljaska ledenim pokrivačima pripadaju biomu tundri. Oko 75% živog svijeta tundre zajednički je arktičkim tundrama u Europi i Aziji. Biljke su malih dimenzija. Od drvenastih biljaka ima jako malo vrsta. To su obično breze i patuljaste vrbe. Od zeljastog bilja brojne su mahovine i lišajevi. Klima je hladna i oštra, ima jako malo padalina, tlo je veći dio godine zaleđeno. Od životinjskih vrsta ističu se mošusno govedo i irvasi, koje u Americi nazivaju karibu. Vukovi i polarne lisice predatori su koji su prilagođeni životu u tundri tako što imaju gusto krzno i sloj potkožne masti. Također su uši, rep i njuška jako skraćeni kako ne bi došlo do smrzavanja. U tundri je veoma mali broj ptica koje su stanarice, dok ptice selice dolaze ljeti u milijunima primjeraka. Neke od njih su plovke i guske. Veliki broj ptica se tada hrani kukcima i to najčešće

dvokrilcima (Diptera) koji inače zimuju u stadiju ličinki pod ledom koji se stvara na površini bara
Karibu Rangifes tarantus
1.2. Antarktička tundra Antarktička tundra pojavljuje se na Antarktiku i na nekoliko subantarktičkih otoka. Antarktika je uglavnom previše hladna i suha za uspijevanje vegetacije. Većina kontinenta pokrivena je ledom. Međutim, neki dijelovi kontinenta, osobito antarktički poluotok, imaju područja krša na kojem je moguć razvoj biljnog svijeta. To su najčešće mahovine, lišajevi i kopnene alge. Slika xxc. Antarktička tundra Za razliku od arktičke tundre, na Antarktici nema velikih kopnenih sisavaca, uglavnom zbog fizičke izolacije od drugih kontinenata. Ima morskih sisavaca i morskih ptica, uključujući pingvine. Nastanjuju područja u blizini obale, a neki mali sisavci, kao što su zečevi i mačke došli su posredstvom čovjeka. Flora i fauna Antarktike i antarktičkih otoka (južno od 60 ° južne zemljopisne širine) zaštićene su Ugovorom o Antarktici.

1.3. Alpska tundra
Snježno bijela koza

Aplska tundra je biom u kojem ne raste drveće, zbog prevelikih nadmorskih visina. Razlikuje se od ostalih tundri po boljoj kvaliteti tla, koje nije zaleđeno. Zauzima područja visokih planina. Nazvana je po Alpama, ali se naziv isto tako odnosi ne samo na Alpe, već i na Kordiljere, Pireneje, Tibetansku visoravan itd.. Slika. Xxc.Alpska tundra Slika. Xxv. Planinska (alpska ) tundra, snježno-bijela koza. Biljni svijet predstavlja patuljasto grmlje. Hladna klima uzrokovana je niskim tlakom zraka, slična je polarnoj klimi.
2. Tajge -Šume četinjača
Šume četinjača nalaze se na tri lokacije: u tajgama koje se nalaze u sjevernom djelu Sjeverne Amerike i Euroazije; blizu planinskih vrhova (gdje se zovu planinske šume četinjača); te duž sjevernoameričke obale Pacifika sve do sjeverne Kalifornije. Obilježja tajgi su šumske vrste s češerima kao što su smreka (Picea) , jela (Abies) i bor (Pinus). Ta su stabla dobro prilagođena na hladnoću i niske temperature, jer im listovi i kora drveta imaju debelu prevlaku. Također igličasti listovi mogu podnjeti težinu snijega. Grmlje je rijetko, a tlo je prekriveno mahovinom i lešajevima ispod sloja iglica. Ptice jedu sjemenke četinjača, a medvjedi, jeleni, sobovi, losovi, dabrovi i mošusni štakori žive oko hladnih jezera i duž potoka i rijeka.Vukovi love veće sisavce. Gorska kuna (Gulo gulo) i planinski lav ili puma (Puma concolor) također nastanjuju ova područja.
Gorska kuna ( Gulo gulo)
Puma ( Puma gonkolor

TAJGA – najveća vegetacija, pojas šuma četinjača na sjevernoj hemisferi (Škotska, Skandinavija, Sibir). Karakteristike: hladna kontinentalna klima (duge, sniježne i suhe zime, hladna, kratka i vlažna ljeta). BILJKE: obična smreka, bijeli bor i breza u Europi, sibirska smreka, sibirski bor i ariš u Sibiru. ŽIVOTINJE: vjeverica, voluharica, tekunica, ptice djetlići, bjelokrili krstokljun, kreja, lješetarke; biljojedi los i sob, altajski i mošusni jelen, mrki medvjed, ris, sove ušara, sjeverna i velika soba, ćug batoglavac.
2.1. Rasprostranjenost tajgi u Svijetu
Tajge su borealne šume, tip vazdazelenih šuma četinjača sjevernih dijelova umjerene zone sjeverne polutke koje su cirkupolarno rasprostranjene u Europi, Aziji ("sibirska tajga") i Sjevernoj Americi ("kanadska tajga"). Naziv tajga potiče iz narodnog govora a prvobitno se odnosio na četinarske šume zapadnog Sibira. Danas se kao znanstveni naziv primjenjuje na sve šume četinjače sjevernih dijelova sjeverne hemisfere. Tajge su jedan od najvećih bioma na našem planetu. Obuhvaćaju područje od oko 12 milijuna kvadratnih kilometara, tj. oko 8% kopna na Zemlji i čine trećinu svjetskih šuma. Geografski, govorimo o području koje se proteže od Kanade i Aljaske do Skandinavije, Rusije, Kine, Mongolije te dijelova Japana i Korejskog poluotoka. Ovakve šume nisu ograničene samo na sjeverna područja, možemo ih pronaći u Sjevernoj Americi na višim nadmorskim visinama Apalači gorja, Stijenjaka i Sierra Nevade te u sličnim planinskim regijama diljem Euroazije. Primarno su sastavljene od četinjača (Coniferales), i to od svega nekoliko različitih vrsta, što znači da je raznolikost u ovim šumama niska. Naime, dominantni rodovi cirkumpolarne tajge su smreka (Picea), jela (Abies), bor (Pinus) i ariš (Larix). Također, niska je i produktivnost, kao i stupanj raspadanja (tome pridonose niske temperature), te su tla u ovakvim šumama uglavnom kisela. Zbog niskog stupnja raspadanja dolazi do akumulacije organskog materijala, što znači da su ova područja zapravo velika skladišta ugljika. Zato imaju veoma važnu ulogu u održavanju globalne ravnoteže ugljika na Zemlji, budući da godišnje apsorbiraju 700
zone područja tajgi

milijuna tona ugljika (što je otprilike jednako godišnjoj emisiji ugljika u Kini). Tajge se u sjevernim subarktičkim oblastima Europe, Azije i Sjeverne Amerike pružaju zonalno širokim pojasom od zapada ka istoku, preko ogromnih oblasti između Tihog i Atlanskog oceana (slika, xxc), pa se stoga može govoriti o zoni tajge, za koju su tamne četinarske šume, prije svega šume smreke i jele, najkarakterističnije. To su tamne četinarske šume u kojima potpuno nedostaju florni elementi širokolisnih listopadnih šuma. Dakle, te šume najčešće izgrađuju jela i smreka. Te tamne, zatvorene četinarske šume relativno su siromašne faunom. Stalni stanovnici tajge su mrki medvjed, vuk, lisica, divlja svinja, vjeverica, puh, hermelin, polarni zec, srna, dabar. Od vodozemaca tu su smuk, crni daždevnjak, živorodni gušter. Zimska klima tajgi je vrlo surova pa živi svijet opstaje zahvaljujući životnim formama koje ih karakteriziraju npr. gusto krzno, bogato perje, potkožno masno tkivo, te sposobnost nalaženja i pripremanja dobrog skrovišta i jazbina, fiziološke osobine tijekom razdoblja zimskog sna i dr.
Slika.xx. Mrki medvjed (Ursus arctos) Slika.xx Vuk (Canis lupus)
Mrki medvjed (Ursus arktor)
Vuk ( Kanis lupus)
2.2. Tipovi tajgi
Možemo istaknuti nekoliko tipova odnosno podtipova tajgi:
listopadne ariševe tajge (graniče sa tundrama)
svijetle tajge (dominiraju borovi)
tamne tajge, dominiraju vrste iz redova smreka (Picea), jela (Abies), bor (Pinus)
mješovite tajge u kojima dominiraju širokolisne listopadne vrste otpornije na niske temperature.
2.3. Ruske borealne šume (ruska tajga)
Ruska Federacija država je na istoku Europe i sjeveru Azije. Površinom je najveća država na svijetu (oko 11,5% zemaljskog kopna), ali je većina državnog teritorija slabo naseljena pa po broju stanovnika zauzima tek osmo mjesto (poslije Kine, Indije, SAD-a, Indonezije, Brazila, Pakistana i Bangladeša). Glavni grad Moskva nalazi se u europskom dijelu države. Ruske borealne šume (tajge) su najveće šumsko prostranstvo na svijetu, trenutno prolaze velike promjene u vegetacijskim tipovima, što je posljedica globalnih i regionalnih promjena klime. Zbog značajnog dokumentiranog zagrijavanja tijekom zadnjih nekoliko desetljeća vrste drveća koje su tolerantnije na toplije vrijeme napreduju prema sjeveru dok vrste koje su manje tolerantne na topliju klimu gube na brojnosti.
Ruska borealna tajga

Slika xxc. Ruska borealna (tajga)
Konkretno, zimzelene četinjače potiskuju stabla ariša koji je jedna od rijetkih listopadnih četinjača. Ariš (Larix decidua) u jesen odbacuje svoje iglice i na taj način dozvoljava da snijeg prekrije površinski sloj tla te reflektira sunčevo zračenje i toplinu natrag u svemir. No, vazdazelene četinjače, poput smreke, zadržavaju iglice tijekom cijele godine, apsorbiraju sunčevo zračenje, te uzrokuju zadržavanje topline. Taj proces stvara idealne uvjete za rast i razvoj vazdazelenih vrsta koje potiskuju ariš što uzrokuje dodatnu progresije vazdazelenih vrsta četinjača prema sjevernijim područjima te potiskivanje ariša. Spomenuti proces je tipičan primjer sukcesije vegetacije u ruskoj borealnoj šumi. Klimatski modeli predviđaju da bi se širenjem vazdazelenih borealnih šuma četinjače dovelo do dodatnog povećanja površinskog zagrijavanja tla. Model uzima u obzir još detaljnije informacije o prirastu drveća, te se rezultati poklapaju s dokumentiranim promjenama na terenu u produkciji i pomicanju vazdazelenih šumskih granica prema sjeveru. Takve promjene u tako velikim šumskim regijama imaju potencijal da utječu i na područja izvan te regije. Ruska tajga nalazi se na površinama koje čine ogromno skladište ugljikom bogato tlo koje je smrznuto u obliku permafrosta. Kako se u širem području ruske tajge mijenja distribucija vrsta od ariša do vazdazelenih četinjača, zagrijavanje površinskog prizemnog sloja uzrokovalo bi odmrzavanje i razgradnju tla što bi dovelo do otpuštanja golemih količina ugljikovog dioksida (CO2) u atmosferu. Smatra se da je to količina od čak 15% CO2 koji se trenutno nalazi u atmosferi. Takav scenarij bio bi iznimno nepovoljan, jer bi još dodatno došlo do povećanja zagrijavanja na globalnoj razini.
2.4. Crnogorične šume umjerenog pojasa
Crnogoricom odnosno crnogoričnim šumama nazivamo šume sa prevladavajućim vrstama drva koje ne odbacuje lišće, nego ga zadržavaju tijekom cijele godine. Od crnogoričnih vrsta možemo istaknuti jelu (Abies), smreku (Picea), planinski bor (Pinus sylvestris), crni bor (Pinus nigra), munjika (Pinus heldreichii) ,

molika (Pinus peuce), ariš (Larix), tisa (Taxus baccatta), tuja (Thuja), borovnica (Vaccinum myrtillus). Crnogorične šume četinjača u kojima prevladava smreka i jela najviše se javljaju na Karpatima, Alpama, Dinaridima, zatim u SAD-u u području Stijenjaka. Tlo je podzol. Na tlu se nalazi debeli sloj iglica i češera koji prekrivaju raspadnuti humus.

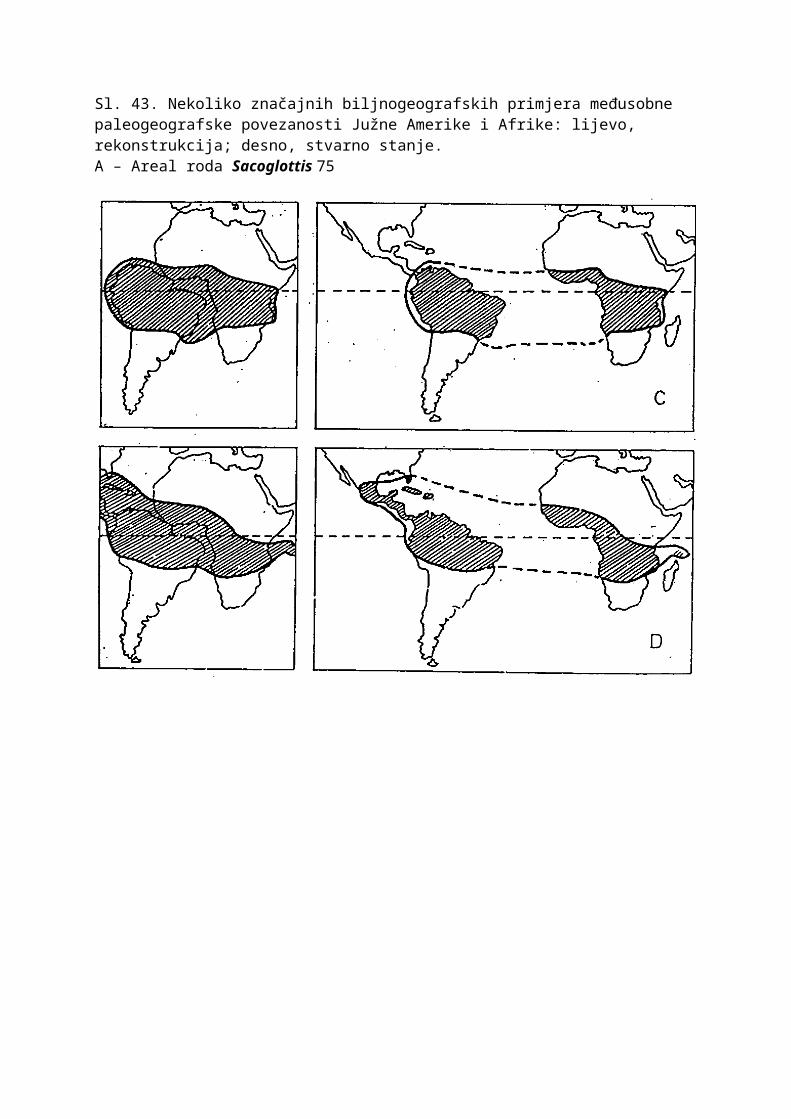
Slika xxy. Rasprostranjenost šuma smreke i jele u Europi (lijevo) i Svijetu (desno)
To je dio prostranih planinskih područja, planinski prevoji i bogatstvo prirodnih ljepota. Velike guste bjelogorične i crnogorične šume u planinama nekada su bile nepristupačne. Na području crnogoričnih šuma umjerenog pojasa može se izdvojiti područje do 400 m nadmorske visine koje je obraslo pretežno bjelogoricom. Tu prevladava bukva (Fagus silvatica), zatim grab (Carpinus) , brijest (Ulmus) , jasen (Fraxinus) , lipa (Tilia) i dr. Od niskog raslinja, značajne su razne režuhe (Cardamine), lazarkinja (Asperula) i mnogobrojne biljke pratilice: šumarica (Anemone), kopitnjak (Asarum europaeum) i dr. U višem gorskom području (400-1000 m nadmorske visine), značajne su samonikle, mješovite ili čiste šume, šume četinjača, uglavnom jele (Abies) i smreke (Picea). Od crnogorice najviše ima smreke, zatim jele i vrlo rijetko tise (Taxus). Pretplaninsko područje obuhvaća prostor iznad 1000 m nadmorske visine. Veoma snažan vjetar, snijeg i led, tu su odlučujući ekološki činioci. Tu je značajna pretplaninska šuma bukve (Fagetum croaticum subalpinum Horv.), a u prizemnom raslinju bijeli žabljak (Ranuculus), kamenika (Saxifraga) i planinska

paprat, zatim je uočljiva pretplaninska šuma smreke (Piceetum croaticum subalpinum Horv.) sastavljena od smreke vitkog uzrasta i niske krošnje, a na tlu različite krušćice (Pirola), crvotočine (Lycopodium), borovnica (Vaccinum myrtillus), brusnica (Vitis idaeae), mahovina i druge acidofilne biljke. Konačno, u najvišim pretplaninskim predjelima iznad 1400 m nadmorske visine (Bjelolasica), javlja se klekovina bukve (Fagetum suffrutcosum Horv.). Na rubovima i udubinama ponikava, gdje se dugo zadržava snijeg, održava se velelisna vrba (Salicetum grandifoliae Horv.). Krajoliku daju posebna obilježja livade, rudine, vrištine, utrine i druga zemljišta koja nisu obrasla šumama. Prašuma umjerenih krajeva U pojasu umjerene klime goleme se prašume nalaze u zapadnoj Rusiji i Kanadi. U Europi je jedina prašuma Perućica kod Vučeva u Bosni i Hercegovini. U prašumama umjerenih krajeva žive: mrki medvjed, ris, divlja mačka, vjeverica, kuna itd. Slika. Xxc. Prašuma Peručica u području Istočne Bosne

3. Listopadne šume Umjerenog pojasa Listopadne šume umjereno klimatskih područja rasprostranjene su u predjelima s relativno hladnom zimskom i toplom ljetnom sezonom. Tu se prosječna mjesečna temperatura kreće od 3oC koliko iznosi za siječanj do 15,6 oC za srpanj. Klima u području širokolisnih listopadnih šuma je relativno topla i kišovita, katkada tijekom ljeta se pojavi duže sušno razdoblje. Suha sezona tijekom zime se pojavljuje u istočnoj Aziji. Druga područja rasprostranjenja listopadnih šuma imaju česte padaline, tako da ukupna godišnja količina padalina u ovoj zoni se kreće od 600 mm do često i preko 1500 mm. Temperatura je obično umjerena, osim u nekim dijelovima Azije gdje ove šume opstaju unatoč surovim uvjetima i vrlo hladnim zimama.
Područje rasprostranjenja listopadnih šuma umjerenog pojasa

3.1. Rasprostranjenost listopadnih šuma umjerenog područja Listopadne šume nalazimo na svim kontinentima umjerenog pojasa, kao na primjer na istočnoj obali Sjeverne Amerike, na krajnjem jugu Južne Amerike i Afrike, najvećem dijelu Europe, istočnoj Aziji i istočnoj obali Australije. Rastu u umjereno kontinentskoj klimi. Karakteristično i dominantno širokolisno drveće listopadnih šuma umjerenog područja (bioma) su hrastovi (vrste roda Quercus),
Slika. Xxc. Područje rasprostranjenja listopadnih šuma umjerenog pojasa bukve (Fagus), javori (Acer) i breze (Betula). U ovom području često se javljaju i "mješovite šume" što se odnosi na šume u kojima ima elemenata listopadnog i zimzelenog drveća. Tipično zimzeleno drveće su: borovi (vrste roda Pinus), jele (Abies) i smreke (Picea). U nekim područjima u okviru ovog bioma zimzeleno drveće može biti važnije od listopadnog. U nešto hladnijim područjima listopadna šuma prelazi u zimzelenu šumu četinjača u kojima prevladava jela i smreka. U ovim šumama, tlo je većinom smeđe, bogato humusom, stoga je pogodno za razvoj poljoprvrede. Zbog krčenja i sječe šuma danas su ta šumska postranstva potisnuta (ostale su manje površine tih šuma), no u novije vrijeme sve se više vodi briga o pošumljavanju. Ovdje su živjele brojne životinjske vrste, neke od njih su i nestale (medvjed, ris, vuk, jazavac, divlje govedo...). Životinje koje žive u umjerenim listopadna šuma moraju biti u stanju prilagoditi se promijenama godišnjih doba. Neke se životinje sele ili hiberniraju tijekom zime.

Struktura širokolisnih listopadnih šuma se obično sastoji iz četiri dijela odnosno sloja. Najveći sloj je (i.) sloj krošnji odraslog listopadnog visokog drveća, koje može biti visoko između 30 i 65 m. Ispod sloja krošnji nalaze se ostala tri sloja, koja su otprilike visoka između 9 i 15 metara. Niži sloj, ispod sloja krošnje visokog drveća javljaju se biljke prilagođene na manju količinu svjetlosti. (ii) Taj sloj uključuje manje odraslo drveće ili mlado drveće koje se probija prema svjetlosti. To se drveće obično razvije u visoka stabla ako im se ukaže prilika u borbi za svjetlost i prostor, posebno ako se uruši ili osuši neko veće drvo. Ispod ovog sloja je (iii) sloj grmlja, koji se sastoji od niskih drvenastih biljaka. Najniži (i biološki najraznovrsniji) je (iv) prizemni sloj zeljastih biljaka. Biom - umjerena listopadna šuma uvijek se mijenja. Ona ima četiri različita godišnja doba: zima, proljeće, ljeto i jesen. Zime su hladne, a ljeta su topla. Listopadne šume dobivaju između 800 i 1500mm oborina godišnje koje su raspoređene tijekom cijele godine. Tlo u listopadnim šumama vrlo je plodno. Šume sadrže tvrdo drveće koje se koristi u građevinarstvu i drvoprerađivačkoj industriji. U području listopadnih šuma umjerenog pojasa razvili su se najveći gradovi Svijeta. Slika. Xxc. Struktura (slojevi) listopadne šume umjerenog klimatskog područja Listopadne šume rastu u umjerenom klimatskom pojasu, gdje su ljeta topla, zime umjereno hladne, s dosta oborina tijekom cijele godine. Prema nekim istraživanjima je utvrđeno da listopadne šume dva do četiri puta rastu brže nego što su rasle prije 200-njak godina. Tako je prema proučavanju (više od 20 godina) miješanih listopadnih šuma u državi Maryland utvrđeno da u prosjeku šume na istoku SAD-a godišnje narastu više za oko 2t po 0,4 ha, nego prije 200-njak godina objavio je časopis Proceedings of the National Academy of Sciences. Studija ukazuje da su čimbenici ubrzana rasta šuma povećanje količine ugljikova dioksida u atmosferi, porast temperature i produljenje vegetacijske sezone. Tijekom protekle 22 godine razina CO2 porasla je 12 %, prosječna temperatura za skoro tri desetine stupnja, a vegetacijska sezona produljila se za 7,8 dana. Cjelokupni učinak na ekosustave tek će se utvrditi.Postoji veliki broj ekosustava koje možemo podijeliti u dvije velike cjeline:
Kopneni ekosustavi - to je skup ekosustava gdje je okolni zrak prirodni okoliš
Vodeni ekosustavi – to je skup ekosustava gdje je voda prirodni okoliš
Kopnene zajednice
I. Terestički BIOMI
BIOMI – NAJVEĆI TIPOVI VEGETACIJE NA ZEMLJI Biomi (ekozone ili ekoregije) - čine makro biocenoze ili velika geografska područja ekozona na Zemlji, gdje makroklimatski uvjeti određuju tipove ekoloških zajednica koje ih izgrađuju. Biome čine različite skupine kopnene biljnih i životinjskih zajednica

prilagođenih na zajedničke makroklimatske uvjete. Biom čini specifičan spoj biljaka i životinja koje su prilagođene životu u određenim uvjetima u okolini, među kojima klima ima najznačajniji utjecaj. Npr. grafički prikaz kopnenih bioma sa srednjom godišnjom temperaturom zraka i srednjom godišnjom količinom oborina ima specifičan obrazac (slika x). Na slici x prikazane su jasne granice između bioma, međutim treba imati na umu da se biomi postupno izmjenjuju. Također, iako ćemo opisati svaki biom posebno treba znati da je svaki biom povezan sa svakom kopnenom i vodenom zajednicom u biosferi.

Slika xx. Biomi najveći tipovi vegetacije na Zemlji Slika xx1. Odnos između nadmorske visine, geografske širine i bioma . Veća nadmorska širina, klima je hladnija. Čak i u tropskim područjima vrhovi visokih planina mogu zadržavati snijeg tijekom cijele godine. Tako je moguće susretati se s istim tipovima vegetacije idući u visoke planinske lance, ali i krećući se od ekvatora prema polovima (od manje prema većoj geografskoj širini)

Slika xx2. Utjecaj prosječne godišnje količine oborina i prosječne godišnje temperature na razvoj bioma na Zemlji. U područjima s niskim oborinama i niskim temperaturama razvijaju se tundre. S druge pak strane u područjima s visokom količinama oborina i visokim temperaturama razviaju se tropske kišne šume. Slično možemo navesti i za sve ostale biome. Rasporostranjenost bioma na Zemlji vezani su uz osnovna obilježlja makroklime i njezinim razlikama, čime su određeni i uvjeti života na Zemlji, odnosno već opisanom količinom sunčevog zračenja ovisno o geografskoj širini i nadmorskoj visini kao i topografskim obilježjima (Slika xx1). Dakle, utjecaj temperaturnog gradijenta može se vidjeti ne samo s geografskom širinom već i s nadmorskom visinom. NA svakih 100m nadmorske visine temperatura opada 1oC. Idući od ekvatora prema Sjevernom polu prvo nailazimo na tropske kišne šume, slijede listopadne šume umjerenih područja, pa redom šume četinjača i tundre. Gotovo isti slijed možemo uočiti uspinjanjem na visoke planine. Šume četinjača u planinama nazivamo planinske četinjače, a tundre blizu planinskih vrhova nazivaju se alpske tundre. Međutim, s druge pak strane idući od prema Južnom polu, nećemo sresti područja koja odgovaraju šumama četinjača i tundri sjeverne polutke, jer se kopno na južnoj polutki nalazi smješteno više prema sjeveru. Rasprostranjenost bioma na Zemlji određena je makroklimatskim paramertima kao što je prosječna godišnja temperatura i prosječna godišnja količina oborina, a koje variraju prema geografskim širinama i nadorskim visinama. Biomi su klimatski i geografski određeni prema sličnim klimatskim uvjetima koji vladaju na Zemlji, a također ih obilježavaju karakteristične zajednice biljaka, životinja kao i obiježlja tla, zbog čega se biomi definiraju kao veliki ekosustavi ili ekozone odnodno ekoregije.

Neki dijelovi Zemlje imaju više ili manje iste ili slične biotičke ili abiotičke čimbenike koji su prošireni na velikom području stvarajući tipični ekosustav karakterističan za to područje. Takvi veliki ekosustavi nazivaju se biomi. Biomi se često uspoređuju s pojedinim obrascima ekološke sukcesije i klimaks vegetacije (prividnim ravnotežnim stanjem lokalnih ekosustava). Ekosustav uključuje veliki broj biotopa, a biom u cjelini predstavlja glavni stanišni tip. Biološka raznolikost karakterizira svaki biom, a posebno raznolikost flore i faune, ali uključujući i manje dominantne biljke i životinje. U kopnenim biomima, biološka raznolikosti biljnih vrsta često se odražava na veću neto primarnu proizvodnost, što je u uskoj vezi s dostupnosti vlage i većim temperaturama. U osnovi biome možemo podijeliti na: 1. Terestičke /kopnene biome, i 2. Akvatičke biome (uključujući slatkovodne i morske biome) Kao što je već istaknuto, makroklima je glavni čimbenik koji određuje rasprostranjenost terestičkih bioma. Među važnije klimatske čimbenike možemo ubrojiti: 1. Geografsku širinu: Arktik, sjeverni klimatski pojas, umjereni klimatski pojas, suptropski pojas i tropski pojas, 2. Vlažnost: vlažno, polu-vlažno, polu-sušno i sušno područje. 3. O sezonske varijacije: Oborine mogu biti ravnomjerno raspoređena tijekom cijele godine ili mogu postojati sezonska razdoblja bez kiše, to jest sušna razdoblja, kao npr u Sredozemlju gdje su izražena suha ljeta i vlažne zime: Većina regija na Zemlji dobivaju većinu svojih oborina tijekom ljetnih mjeseci, dok u Sredozemnom Bazenu većina oborina dolazi tijekom zimskih mjeseci. 4. Nadmorska visina, povećanje nadmorske visine uzrokuje raspored stanišnih tipova slično kao povećanje geografske širine. Biološka raznolikost se općenito povećava od polova prema ekvatoru i raste s porastom vlage. Biomi su široko rasprostranjeni ekosustavi, koji se odlikuje određenim dominantnim tipovima vegetacije kao na primjer: tundra, tajga, tropska kišna šuma, stepa, savana, bjelogorične i crnogorične šume

Slika xxc. Razvoj bioma u odnosu prema klimatskim uvjetima na Zemlji
1. Tundra
Tundra je biom koji se prostire u predjelima, gdje je rast drveća onemogućen nižim temperaturama i kratkom vegetacijskom sezonom. Ime potječe od kildin-samske (laponske) riječi "tūndâr", koja označava "ledinu bez drveća". Postoje tri tipa tundre: (i) arktička, (ii) antarktička i (iii) alpska. Flora palearktičkih1 tundri često se naziva arkto-alpskom uslijed postojanja zajedničke povijesti vegetacije i velikog broja zajedničkih vrsta biljaka u arktičkoj, antarktičkoj i alpskoj tundri. Biom artičke tundre okružuje Zemlju malo južnije od ledom prekrivenog polarnog mora na sjevernoj polutki, te prekriva 20% zemljine površine. Slična zajednica zvana alpska tundra nalazi se iznad planinskih područja. Artička tundra je hladna i mračna tijekom većeg dijela godine. S obzirom da godišnja količina oborina iznosi svega 200 mm, tundra bi se mogla smatrati hloadnom pustinjom, no topljenje snijega uzrokuje nastanak vodenih bazena tijekom ljeta. U tundi se voda zadržava jer je isparavanje iznimno malo. Samo površinski sloj tla se odmrzava tijekom ljtnih mjeseci, a ispod tog sloja se nalazi permafrost koji je uvijek smrznut, pa je stoga procjeđivanje vode minimalno. 1 Palearktička regija je staro-sjeverno područje (palearktička regija). U razdoblju terercijar-kvartar kopnena veza između sjeverne amerike i euroazije egzistirala je u području beringovog prolaza i grendlanda. Šuma u tundrama nema jer je vegetacijska sezona prekratka, njihovo korjenje ne može prodrijeti u permafrost, i ne mogu se ukorjeniti u blatnom tlu tijekom ljeta. Ljeti tlo je prekriveno

niskom travom i trskom te područjima s lišajevima i mahovinom. Patuljasti drveni grmovi, kao što je patuljasta breza ( Betula papyrifera), cvatu i osjemenjuju se brzo dok ima dovoljno sunca za fotosintezu. Nekoliko vrsta životinja živi u tundrama preko cijele godine. Npr. mišu sličan leming ostaje ispod snijega; tetrijeb se zakopava u snijeg za vrijeme oluja; a mošusno govedo čuva toplinu zahvaljujući debelom krznu te kratkom i zdepastom tijelu. Ljeti tundre preplave brojni insekti i ptice, posebno obalne ptice, guske i patke koji kreću u unutrašnjost. Jeleni i srne također migriraju u i iz tundre, kao i vukovi koji tamo love, a polarni medvjevi su česti uz obalu. Slika.xxx. Biom tundra Slika xxc. Patuljasta breza ( Betula papyrifera) Slika xxx. Tetrijeb Slika.xy Mošusno govedo (Ovibos moschatus)

1.1. Arktička tundra
U skorijoj geološkoj prošlosti (ledena doba kvartara) velike ledene mase pokrivale su dio Sjeverne Amerike i Europe. U periodu poslije posljednje glacijacije led se povukao prema sjeveru ili se zadržao na planinama (ledenjaci). Područja koja su posljednja oslobođena leda, a susjedna su Slika. Xcx Arktička tundra, Aljaska ledenim pokrivačima pripadaju biomu tundri. Oko 75% živog svijeta tundre zajednički je arktičkim tundrama u Europi i Aziji. Biljke su malih dimenzija. Od drvenastih biljaka ima jako malo vrsta. To su obično breze i patuljaste vrbe. Od zeljastog bilja brojne su mahovine i lišajevi. Klima je hladna i oštra, ima jako malo padalina, tlo je veći dio godine zaleđeno. Od životinjskih vrsta ističu se mošusno govedo i irvasi, koje u Americi nazivaju karibu. Vukovi i polarne lisice predatori su koji su prilagođeni životu u tundri tako što imaju gusto krzno i sloj potkožne masti. Također su uši, rep i njuška jako skraćeni kako ne bi došlo do smrzavanja. U tundri je veoma mali broj ptica koje su stanarice, dok ptice selice dolaze ljeti u milijunima primjeraka. Neke od njih su plovke i guske. Veliki broj ptica se tada hrani kukcima i to najčešće dvokrilcima (Diptera) koji inače zimuju u stadiju ličinki pod ledom koji se stvara na površini bara.

Slika. Xxx karibu (Rangifer tarandus) Slika. Xxx. Polarna lisica (Alopex lagopus),
1.2. Antarktička tundra Antarktička tundra pojavljuje se na Antarktiku i na nekoliko subantarktičkih otoka. Antarktika je uglavnom previše hladna i suha za uspijevanje vegetacije. Većina kontinenta pokrivena je ledom. Međutim, neki dijelovi kontinenta, osobito antarktički poluotok, imaju područja krša na kojem je moguć razvoj biljnog svijeta. To su najčešće mahovine, lišajevi i kopnene alge. Slika xxc. Antarktička tundra Za razliku od arktičke tundre, na Antarktici nema velikih kopnenih sisavaca, uglavnom zbog fizičke izolacije od drugih kontinenata. Ima morskih sisavaca i morskih ptica, uključujući pingvine. Nastanjuju područja u blizini obale, a neki mali sisavci, kao što su zečevi i mačke došli su posredstvom čovjeka. Flora i fauna Antarktike i antarktičkih otoka (južno od 60 ° južne zemljopisne širine) zaštićene su Ugovorom o Antarktici.
1.3. Alpska tundra

Aplska tundra je biom u kojem ne raste drveće, zbog prevelikih nadmorskih visina. Razlikuje se od ostalih tundri po boljoj kvaliteti tla, koje nije zaleđeno. Zauzima područja visokih planina. Nazvana je po Alpama, ali se naziv isto tako odnosi ne samo na Alpe, već i na Kordiljere, Pireneje, Tibetansku visoravan itd.. Slika. Xxc.Alpska tundra Slika. Xxv. Planinska (alpska ) tundra, snježno-bijela koza. Biljni svijet predstavlja patuljasto grmlje. Hladna klima uzrokovana je niskim tlakom zraka, slična je polarnoj klimi.
2. Tajge -Šume četinjača
Šume četinjača nalaze se na tri lokacije: u tajgama koje se nalaze u sjevernom djelu Sjeverne Amerike i Euroazije; blizu planinskih vrhova (gdje se zovu planinske šume četinjača); te duž sjevernoameričke obale Pacifika sve do sjeverne Kalifornije. Obilježja tajgi su šumske vrste s češerima kao što su smreka (Picea) , jela (Abies) i bor (Pinus). Ta su stabla dobro prilagođena na hladnoću i niske temperature, jer im listovi i kora drveta imaju debelu prevlaku. Također igličasti listovi mogu podnjeti težinu snijega. Grmlje je rijetko, a tlo je prekriveno mahovinom i lešajevima ispod sloja iglica. Ptice jedu sjemenke četinjača, a medvjedi, jeleni, sobovi, losovi, dabrovi i mošusni štakori žive oko hladnih jezera i duž potoka i rijeka.Vukovi love veće sisavce. Gorska kuna (Gulo gulo) i planinski lav ili puma (Puma concolor) također nastanjuju ova područja.

Slika xxy. Gorska kuna (Gulo gulo) Slika xxc. puma (Puma concolor) TAJGA – najveća vegetacija, pojas šuma četinjača na sjevernoj hemisferi (Škotska, Skandinavija, Sibir). Karakteristike: hladna kontinentalna klima (duge, sniježne i suhe zime, hladna, kratka i vlažna ljeta). BILJKE: obična smreka, bijeli bor i breza u Europi, sibirska smreka, sibirski bor i ariš u Sibiru. ŽIVOTINJE: vjeverica, voluharica, tekunica, ptice djetlići, bjelokrili krstokljun, kreja, lješetarke; biljojedi los i sob, altajski i mošusni jelen, mrki medvjed, ris, sove ušara, sjeverna i velika soba, ćug batoglavac.
2.1. Rasprostranjenost tajgi u Svijetu
Tajge su borealne šume, tip vazdazelenih šuma četinjača sjevernih dijelova umjerene zone sjeverne polutke koje su cirkupolarno rasprostranjene u Europi, Aziji ("sibirska tajga") i Sjevernoj Americi ("kanadska tajga"). Naziv tajga potiče iz narodnog govora a prvobitno se odnosio na četinarske šume zapadnog Sibira. Danas se kao znanstveni naziv primjenjuje na sve šume četinjače sjevernih dijelova sjeverne hemisfere. Tajge su jedan od najvećih bioma na našem planetu. Obuhvaćaju područje od oko 12 milijuna kvadratnih kilometara, tj. oko 8% kopna na Zemlji i čine trećinu svjetskih šuma. Geografski, govorimo o području koje se proteže od Kanade i Aljaske do Skandinavije, Rusije, Kine, Mongolije te dijelova Japana i Korejskog poluotoka. Ovakve šume nisu ograničene samo na sjeverna područja, možemo ih pronaći u Sjevernoj Americi na višim nadmorskim visinama Apalači gorja, Stijenjaka i Sierra Nevade te u sličnim planinskim regijama diljem Euroazije. Primarno su sastavljene od četinjača (Coniferales), i to od svega nekoliko različitih vrsta, što znači da je raznolikost u ovim šumama niska. Naime, dominantni rodovi cirkumpolarne tajge su smreka (Picea), jela (Abies), bor (Pinus) i ariš (Larix). Također, niska je i produktivnost, kao i stupanj raspadanja (tome pridonose niske temperature), te su tla u ovakvim šumama uglavnom kisela. Zbog niskog stupnja raspadanja dolazi do akumulacije organskog materijala, što znači da su ova područja zapravo velika skladišta ugljika. Zato imaju veoma važnu ulogu u održavanju globalne ravnoteže ugljika na Zemlji, budući da godišnje apsorbiraju 700

milijuna tona ugljika (što je otprilike jednako godišnjoj emisiji ugljika u Kini). Tajge se u sjevernim subarktičkim oblastima Europe, Azije i Sjeverne Amerike pružaju zonalno širokim pojasom od zapada ka istoku, preko ogromnih oblasti između Tihog i Atlanskog oceana (slika, xxc), pa se stoga može govoriti o zoni tajge, za koju su tamne četinarske šume, prije svega šume smreke i jele, najkarakterističnije. To su tamne četinarske šume u kojima potpuno nedostaju florni elementi širokolisnih listopadnih šuma. Dakle, te šume najčešće izgrađuju jela i smreka. Slika. Xxc. Zone pružanja tajgi Te tamne, zatvorene četinarske šume relativno su siromašne faunom. Stalni stanovnici tajge su mrki medvjed, vuk, lisica, divlja svinja, vjeverica, puh, hermelin, polarni zec, srna, dabar. Od vodozemaca tu su smuk, crni daždevnjak, živorodni gušter. Zimska klima tajgi je vrlo surova pa živi svijet opstaje zahvaljujući životnim formama koje ih karakteriziraju npr. gusto krzno, bogato perje, potkožno masno tkivo, te sposobnost nalaženja i pripremanja dobrog skrovišta i jazbina, fiziološke osobine tijekom razdoblja zimskog sna i dr. Slika.xx. Mrki medvjed (Ursus arctos) Slika.xx Vuk (Canis lupus)

Slika xxx. Divlja svinja (Sus scrofa) Slika xxx. Polarni zec (Lepus arcticus)
2.2. Tipovi tajgi
Možemo istaknuti nekoliko tipova odnosno podtipova tajgi:
listopadne ariševe tajge (graniče sa tundrama)
svijetle tajge (dominiraju borovi)
tamne tajge, dominiraju vrste iz redova smreka (Picea), jela (Abies), bor (Pinus)
mješovite tajge u kojima dominiraju širokolisne listopadne vrste otpornije na niske temperature.
2.3. Ruske borealne šume (ruska tajga)
Ruska Federacija država je na istoku Europe i sjeveru Azije. Površinom je najveća država na svijetu (oko 11,5% zemaljskog kopna), ali je većina državnog teritorija slabo naseljena pa po broju stanovnika zauzima tek osmo mjesto (poslije Kine, Indije, SAD-a, Indonezije, Brazila, Pakistana i Bangladeša). Glavni grad Moskva nalazi se u europskom dijelu države. Ruske borealne šume (tajge) su najveće šumsko prostranstvo na svijetu, trenutno prolaze velike promjene u vegetacijskim tipovima, što je posljedica globalnih i regionalnih promjena klime. Zbog značajnog dokumentiranog zagrijavanja tijekom zadnjih nekoliko desetljeća vrste drveća koje su tolerantnije na toplije vrijeme napreduju prema sjeveru dok vrste koje su manje tolerantne na topliju klimu gube na brojnosti.

Slika xxc. Ruska borealna šuma (tajga) Konkretno, zimzelene četinjače potiskuju stabla ariša koji je jedna od rijetkih listopadnih četinjača. Ariš (Larix decidua) u jesen odbacuje svoje iglice i na taj način dozvoljava da snijeg prekrije površinski sloj tla te reflektira sunčevo zračenje i toplinu natrag u svemir. No, vazdazelene četinjače, poput smreke, zadržavaju iglice tijekom cijele godine, apsorbiraju sunčevo zračenje, te uzrokuju zadržavanje topline. Taj proces stvara idealne uvjete za rast i razvoj vazdazelenih vrsta koje potiskuju ariš što uzrokuje dodatnu progresije vazdazelenih vrsta četinjača prema sjevernijim područjima te potiskivanje ariša. Spomenuti proces je tipičan primjer sukcesije vegetacije u ruskoj borealnoj šumi. Klimatski modeli predviđaju da bi se širenjem vazdazelenih borealnih šuma četinjače dovelo do dodatnog povećanja površinskog zagrijavanja tla. Model uzima u obzir još detaljnije informacije o prirastu drveća, te se rezultati poklapaju s dokumentiranim promjenama na terenu u produkciji i pomicanju vazdazelenih šumskih granica prema sjeveru. Takve promjene u tako velikim šumskim regijama imaju potencijal da utječu i na područja izvan te regije. Ruska tajga nalazi se na površinama koje čine ogromno skladište ugljikom bogato tlo koje je smrznuto u obliku permafrosta. Kako se u širem području ruske tajge mijenja distribucija vrsta od ariša do vazdazelenih četinjača, zagrijavanje površinskog prizemnog sloja uzrokovalo bi odmrzavanje i razgradnju tla što bi dovelo do otpuštanja golemih količina ugljikovog dioksida (CO2) u atmosferu. Smatra se da je to količina od čak 15% CO2 koji se trenutno nalazi u atmosferi. Takav scenarij bio bi iznimno nepovoljan, jer bi još dodatno došlo do povećanja zagrijavanja na globalnoj razini.
2.4. Crnogorične šume umjerenog pojasa Crnogoricom odnosno crnogoričnim šumama nazivamo šume sa prevladavajućim vrstama drva koje ne odbacuje lišće, nego ga zadržavaju tijekom cijele godine. Od crnogoričnih vrsta možemo istaknuti jelu (Abies), smreku (Picea), planinski bor (Pinus sylvestris), crni bor (Pinus nigra), munjika (Pinus heldreichii) ,

molika (Pinus peuce), ariš (Larix), tisa (Taxus baccatta), tuja (Thuja), borovnica (Vaccinum myrtillus). Crnogorične šume četinjača u kojima prevladava smreka i jela najviše se javljaju na Karpatima, Alpama, Dinaridima, zatim u SAD-u u području Stijenjaka. Tlo je podzol. Na tlu se nalazi debeli sloj iglica i češera koji prekrivaju raspadnuti humus. Slika xxy. Rasprostranjenost šuma smreke i jele u Europi (lijevo) i Svijetu (desno) To je dio prostranih planinskih područja, planinski prevoji i bogatstvo prirodnih ljepota. Velike guste bjelogorične i crnogorične šume u planinama nekada su bile nepristupačne. Na području crnogoričnih šuma umjerenog pojasa može se izdvojiti područje do 400 m nadmorske visine koje je obraslo pretežno bjelogoricom. Tu prevladava bukva (Fagus silvatica), zatim grab (Carpinus) , brijest (Ulmus) , jasen (Fraxinus) , lipa (Tilia) i dr. Od niskog raslinja, značajne su razne režuhe (Cardamine), lazarkinja (Asperula) i mnogobrojne biljke pratilice: šumarica (Anemone), kopitnjak (Asarum europaeum) i dr. U višem gorskom području (400-1000 m nadmorske visine), značajne su samonikle, mješovite ili čiste šume, šume četinjača, uglavnom jele (Abies) i smreke (Picea). Od crnogorice najviše ima smreke, zatim jele i vrlo rijetko tise (Taxus). Pretplaninsko područje obuhvaća prostor iznad 1000 m nadmorske visine. Veoma snažan vjetar, snijeg i led, tu su odlučujući ekološki činioci. Tu je značajna pretplaninska šuma bukve (Fagetum croaticum subalpinum Horv.), a u prizemnom raslinju bijeli žabljak (Ranuculus), kamenika (Saxifraga) i planinska

paprat, zatim je uočljiva pretplaninska šuma smreke (Piceetum croaticum subalpinum Horv.) sastavljena od smreke vitkog uzrasta i niske krošnje, a na tlu različite krušćice (Pirola), crvotočine (Lycopodium), borovnica (Vaccinum myrtillus), brusnica (Vitis idaeae), mahovina i druge acidofilne biljke. Konačno, u najvišim pretplaninskim predjelima iznad 1400 m nadmorske visine (Bjelolasica), javlja se klekovina bukve (Fagetum suffrutcosum Horv.). Na rubovima i udubinama ponikava, gdje se dugo zadržava snijeg, održava se velelisna vrba (Salicetum grandifoliae Horv.). Krajoliku daju posebna obilježja livade, rudine, vrištine, utrine i druga zemljišta koja nisu obrasla šumama. Prašuma umjerenih krajeva U pojasu umjerene klime goleme se prašume nalaze u zapadnoj Rusiji i Kanadi. U Europi je jedina prašuma Perućica kod Vučeva u Bosni i Hercegovini. U prašumama umjerenih krajeva žive: mrki medvjed, ris, divlja mačka, vjeverica, kuna itd. Slika. Xxc. Prašuma Peručica u području Istočne Bosne
3. Listopadne šume Umjerenog pojasa Listopadne šume umjereno klimatskih područja rasprostranjene su u predjelima s relativno hladnom zimskom i toplom ljetnom sezonom. Tu se prosječna mjesečna temperatura kreće od 3oC koliko iznosi za siječanj do 15,6 oC za srpanj. Klima u području širokolisnih listopadnih šuma je relativno topla i kišovita, katkada tijekom ljeta se pojavi duže sušno razdoblje. Suha sezona tijekom zime se pojavljuje u istočnoj Aziji. Druga područja rasprostranjenja listopadnih šuma imaju česte padaline, tako da ukupna godišnja količina padalina u ovoj zoni se kreće od 600 mm do često i preko 1500 mm. Temperatura je obično umjerena, osim u nekim dijelovima Azije gdje ove šume opstaju unatoč surovim uvjetima i vrlo hladnim zimama.

3.1. Rasprostranjenost listopadnih šuma umjerenog područja Listopadne šume nalazimo na svim kontinentima umjerenog pojasa, kao na primjer na istočnoj obali Sjeverne Amerike, na krajnjem jugu Južne Amerike i Afrike, najvećem dijelu Europe, istočnoj Aziji i istočnoj obali Australije. Rastu u umjereno kontinentskoj klimi. Karakteristično i dominantno širokolisno drveće listopadnih šuma umjerenog područja (bioma) su hrastovi (vrste roda Quercus), Slika. Xxc. Područje rasprostranjenja listopadnih šuma umjerenog pojasa bukve (Fagus), javori (Acer) i breze (Betula). U ovom području često se javljaju i "mješovite šume" što se odnosi na šume u kojima ima elemenata listopadnog i zimzelenog drveća. Tipično zimzeleno drveće su: borovi (vrste roda Pinus), jele (Abies) i smreke (Picea). U nekim područjima u okviru ovog bioma zimzeleno drveće može biti važnije od listopadnog. U nešto hladnijim područjima listopadna šuma prelazi u zimzelenu šumu četinjača u kojima prevladava jela i smreka. U ovim šumama, tlo je većinom smeđe, bogato humusom, stoga je pogodno za razvoj poljoprvrede. Zbog krčenja i sječe šuma danas su ta šumska postranstva potisnuta (ostale su manje površine tih šuma), no u novije vrijeme sve se više vodi briga o pošumljavanju. Ovdje su živjele brojne životinjske vrste, neke od njih su i nestale (medvjed, ris, vuk, jazavac, divlje govedo...). Životinje koje žive u umjerenim listopadna šuma moraju biti u stanju prilagoditi se promijenama godišnjih doba. Neke se životinje sele ili hiberniraju tijekom zime.
Struktura širokolisnih listopadnih šuma se obično sastoji iz četiri dijela odnosno sloja. Najveći sloj je (i.) sloj krošnji odraslog listopadnog visokog drveća, koje može biti visoko između 30 i 65 m. Ispod sloja krošnji nalaze se ostala tri sloja, koja su otprilike visoka između 9 i 15 metara. Niži sloj, ispod sloja krošnje visokog drveća javljaju se biljke prilagođene na manju količinu svjetlosti. (ii) Taj sloj uključuje manje odraslo drveće ili mlado drveće koje se probija prema svjetlosti. To se drveće obično razvije u visoka stabla ako im se ukaže prilika u borbi za svjetlost i prostor, posebno ako se uruši ili osuši neko veće drvo. Ispod ovog sloja je (iii) sloj grmlja, koji se sastoji od niskih drvenastih biljaka. Najniži (i biološki najraznovrsniji) je (iv) prizemni sloj zeljastih biljaka. Biom - umjerena listopadna šuma uvijek se mijenja. Ona ima četiri različita godišnja doba: zima, proljeće, ljeto i jesen. Zime su hladne, a ljeta su topla. Listopadne šume dobivaju između 800 i 1500mm oborina godišnje koje su raspoređene tijekom cijele godine. Tlo u listopadnim šumama vrlo je plodno. Šume sadrže tvrdo drveće koje se koristi u građevinarstvu i drvoprerađivačkoj industriji. U području listopadnih šuma umjerenog pojasa razvili su se najveći gradovi Svijeta. Slika. Xxc. Struktura (slojevi) listopadne šume umjerenog klimatskog područja Listopadne šume rastu u umjerenom klimatskom pojasu, gdje su ljeta topla, zime umjereno hladne, s dosta oborina tijekom cijele godine. Prema nekim istraživanjima je utvrđeno da listopadne šume dva do četiri puta rastu brže nego što su rasle prije 200-njak godina. Tako je prema proučavanju (više od 20 godina) miješanih listopadnih šuma u državi Maryland utvrđeno da u prosjeku šume na istoku SAD-a godišnje narastu više za oko 2t po 0,4 ha, nego prije 200-njak godina objavio je časopis Proceedings of the National Academy of Sciences. Studija ukazuje da su čimbenici ubrzana rasta šuma povećanje količine ugljikova dioksida u atmosferi, porast temperature i produljenje vegetacijske sezone. Tijekom protekle 22 godine razina CO2 porasla je 12 %, prosječna temperatura za skoro tri desetine stupnja, a vegetacijska sezona produljila se za 7,8 dana. Cjelokupni učinak na ekosustave tek će se utvrditi.


1 Fynbos (South Afrikaans izgovor: [fəinbɒs]) je prirodna grmolika vegetacija koja se javlja u malom pojasu oko Western Cape (Rt Dobre Nade) u Južnoj Africi. Područje je s mediteranskom klimom, okruženo obalnim i planinskim područjima. Fibnos je biom koji uključuje mediteranske šume i šikare. Glavni tipovi vegetacije U glavne tipove vegetacije ovog bioma mediteranske vegetacije spadaju:
Mediteranske šume - uglavnom se sastoje od širokolisnog drveća poput hrastova i mješovitih sklerofilnih šuma većinom rasprostranjenih u Kaliforniji i Sredozemlju, zatim od šuma Eucalyptusa na jugozapadu Australije i od šuma Nothofagusa u centralnom dijelu Čileu. Šume su većinom rasprostranjene na dubljim tlima koja više akumuliraju vode. Vazdazelene šume najvećim dijelom čine vrste rodova bora (Pinus) i vazdazelenih hrastova (Quercus).
Slika. Xxc. Izgled sredozemne vazdazelene šuma (lijevo); jadranska šuma alepskog bora (Pinus halepensis) (desno).

1 Fynbos (South Afrikaans izgovor: [fəinbɒs]) je prirodna grmolika vegetacija koja se javlja u malom pojasu oko Western Cape (Rt Dobre Nade) u Južnoj Africi. Područje je s mediteranskom klimom, okruženo obalnim i planinskim područjima. Fibnos je biom koji uključuje mediteranske šume i šikare.
Šumarci vazdazelenih hrastova/vazdazelena makija koja uključuje visoke i guste vazdazelene šikare koje su karakteristične za Sredozemlje i Kaliforniju, zajedno s manjim šumama alepskog (Pinus halepense) i crnog bora (P. nigra) , a u Kaliforniji šumarcima oraha (Juglans).

Slika. Xxc. Guste, neprohodne šikare vazdazelene mediteranske makije i "finbos" (Južna Afrika)

Niske prorijeđene šikare vazdazelenih bušika ili gariga - to su niske i razmjerno prorijeđene heliofilne šikare nazvane bušici (ili garizi), a sastoje se od vazdazelenih sklerofilnih grmova i niskih drvenastih vrsta, koje se nazivaju „vazdazeleni bušici“ (Hrvatska), "chaparral" (Kalifornija), "matorral" (Čile i južna Španjolska), "maquis" (Francuska i drugdje u Sredozemlju), "macchia" (Italija, a odatle potiče i hrvatski naziv - makija), "finbos" (Južna Afrika) i "kwongan" (jugozapadna Australija)..U nekim područjima ove prorijeđene šikare su glavni oblik vegetacije. U nekim područjima su se razvili degradacijom prvobitnih šuma i sastojina makije, u prvom redu sječom,nekontroliranom ispašom i/ili požarom. Uz degradaciju biljnog pokrova, degradiralo je i tlo odnosno stanište. Slika xxc. Niske, prorijeđene šikare vazdazelenih bušika ili gariga (lijevo); šikara u kanjonu Aliso u Kaliforniji (desno). Niski grmoliki pašnjaci kontinentalnih klimatskih područja Niski grmoliki pašnjaci (engl. heath ili heathland) obuhvaća biljne zajednice patuljastih grmova, uglavnom rasprostranjeni na kiselijim tlima niske kvalitete. Te se biljne zajednice odlikuju otvorenim, niskim drvenastim raslinjem u kojima često dominiraju biljne vrste iz porodice Ericaceae. Niski grmoliki pašnjaci su široko rasprostranjena u cijelom svijetu. Oni čine velike i vrlo raznolike zajednice diljem Australije na vlažnim i poluvlažnim staništima. Slika xxc. Niski grmoliki pašnjaci (Engleska). Ljubičasti cvjetovi pripadaju biljci Calluna vulgaris a žuti cvjetovi vrsti Ulex gallii.
Mediteranske savane, suhi travnjaci i kamenjarski pašnjaci

Slika xxc. Mediteranska savana na Korzici (lijevo); suhi mediteranski travnjaci, Dalmatinska Zagora (desno); prostrani kamenjarski pašnjaci, jadranski otoci (dolje).
Vatra kao ekološki faktor – može nastati slučajnim prirodnim putem ili namjerno izazvana od strane ljudi. Čovjek često koristi

kontroliranu vatru kao oruđe u gospodarenju mediteranskim biljnim pokrovom. Vatra nastala na bilo koji način igra veliku ulogu u oblikovanju svih gore navedenih tipova vegetacije u mediteranskom vegetacijskom biomu. Zbog klimatskih uvjeta (suha i vruća ljeta) i

Mediteranske savane su terestički biomi koji uključuju kamenjarske pašnjake, suhi travnjake, savane (pašnjaci izmiješani s raštrkanim drvećem) i grmolike pašnjačke zajednice (niske prorijeđene šikare) čiju dominantnu vegetacija uglavnom čine trave, grmovi i drveće. Klima je umjerena s izraženim ljetnim sušnim razdobljem. Pašnjačka vegetacija mediteranske eko-regije (bioma) razvijena je dijelom u obliku savane, zatim suhih mediteranskih travnjaka2 i prostranih kamenjarskih pašnjaka3. Travnjaci u Centralnoj dolini u Kaliforniji su najveća travnjačka u području mediteranskog vegetacijskog bioma. Međutim, te su površine većinom preorane i iskorištene za poljoprivredu. 2 Ovaj se travnjak (košanica) u Mediteranu jednom do dva puta godišnje se kosi, a ostali dio godine služi za ispašu.
2
Vatra kao ekološki faktor – može nastati slučajnim prirodnim putem ili namjerno izazvana od strane ljudi. Čovjek često koristi kontroliranu vatru kao oruđe u gospodarenju mediteranskim biljnim pokrovom. Vatra nastala na bilo koji način igra veliku ulogu u oblikovanju svih gore navedenih tipova vegetacije u mediteranskom vegetacijskom biomu. Zbog klimatskih uvjeta (suha i vruća ljeta) ilako zapaljive vegetacije (visoki sadržaj eteričnih ulja i smola, suho nisko raslinje) ova područja iznimno su osjetljiva na požare, koji se ponekad mogu izazvati samim udarom munje. U mediteranskoj vegetaciji mnoge su biljke pirofiti, tj. otporni na požare ili čak ovise o vatri koja im služi za razmnožavanje, ubrzan protok (recikliranje) hranjivih tvari (mineralizacija organske tvari) i odbacivanje mrtvih i/ili starijih biljnih dijelova to jest obnavljanjem (pomlađivanjem) vegetacije, izbijanjem mladih biljnih izbojaka iz spajajućih pupova vrata korijena. Domorodci su i u australskoj i kalifornijskoj mediteranskoj eko-regiji ekstenzivno koristili vatru kako bi uništili drveće i grmlje, obnovili i stvorili pašnjačku vegetaciju za ispašu domaćih i divljih životinja. Biljne sastojine u ovim područjima prilagođene su na česte požare, bilo da su izazvani na prirodni način ili čovjekovom djelatnošću. Na taj način pirofitne biljke počinju dominirati u vegetaciji, a manje otporne na vatru ubrzo iščeznu. Nakon kolonizacije ovih regija od strane Europljana, požari su postali rjeđi, što je izazvalo neke neželjene posljedice u ovim ekosustavima. Odbačena organska tvar nakupljala se u ekosustavu, pa su posljedice od požari postajale znatno veće nego inače, a neke vrste čije je razmnožavanje i razvoj ovisio o vatri postale su u novim uvjetima ugrožene. Sredozemna vazdazelena vegetacija makija i bušika ili gariga i vegetacija suhih travnjaka i kamenjarskih pašnjaka također je oblikovana antropogenim požarima, koji su povijesno vezani uz uzgoj ovaca i koza.
5. Tropske i suptropske kišne širokolisne šume Tropske i suptropske kišne širokolisne šume, također se nazivaju tropske kišne šume, su tropski i suptropski šumski biomi. Pod tim se nazivom podrazumijeva tip vegetacije koji srećemo samo u tropskom klimatskom području. Za ove vazdazelene šume karakterističan je gusti biljni pokrov oblikovan od krošnji visokog drveća. Te šumska sastojine bogate su florom i faunom. U tom klimatskom području ne postoje izmjene godišnjih doba, pa tako ni razdoblje vegetacijskog mirovanja. Pored koraljnih grebena, u ovim područjima je najveća gustoća biljnih i životinjskih vrsta na svijetu.

Tropske kišne šume (vazdazelene prašume ili džungle) tvore ekološke sustave4 prilagođene tropskoj i suptropskoj klimi, čije je osnovna obilježja uključuje prosječnu godišnju temperaturu od 25 °C s kolebanjima od 0,5-0,6 °C između godina, i dnevnim temperaturnim amplitudama od 6-10 °C. Pored toga, godišnja količina padavina je iznad 2.500 mm, tako da je više od sedam mjeseci godišnje klima humidna ili perhumidna. Tropske kišne šume se rasprostiru u tropskim područjima Južne i Srednje Amerike, Afrike i Južne Azije kao iAustralije, s obje strane ekvatora. Iznimka su područje Anda u Južnoj Americi i pasatno-monsunsko područje Istočne Afrike.
Slika xxc. Rasprostranjenost tropskih kišnih šuma
Utvrđeno je da su 1950. tropske kišne šume pokrivale od 16-17 miliona km2, dakle oko 11% površine Zemlje. Do 1980. oko 50% tih šumskih površina je uništeno čovjekovim djelovanjem. Nažalost, proces uništavanja tih šumskih sastojina nastavljen je i traje još uvijek.
Šume tropskog pojasa Šume tropskog pojasa se prostiru na ekvatorijalnom području, vlažna i topla klima, obilje padalina, velika bioraznolikosti i vertikalna slojevitost (5 slojeva). (Amazona najveća!). Monsunska šuma – indomalajski poluotok; kiše dolaze s monsunom, biljke listaju 1 mjesec prije dolaska monsuna. Tropske kišne šume su rasprostranjene između ekvatora i 23° sjeverne i južne geografske širine gdje padaline redovne premašuju i 250o mm godišnje. U tim šumama živi najveći broj vrsta po jedinici površine uopće, a visina stabala dosiže i do 60 metara. Temperatura i dužina dana su u tim geografskim širinama relativno su stalne, tako da raznolikost vegetacije određena samo različitim količinama padalina. Tako razlikujemo tropske sušne šume koje imaju 42-postotni udio listopadnog drveća; tropske šume s 32% udjela listopadnog drveća i tropske kišne šume s 25% udjela listopadnog drveća. U tropskim krajevima i njihovim prašumama, kiše su vrlo obilne. Najveća prašuma

na svijetu je tropska kišna šuma (džungla) Amazonija, velika je kao Europa. Proizvede oko 40% kisika na Zemlji. Klimatski dnevni raspored u tropskoj kišnoj šumi - 6.00 sati sunce vrlo brzo izlazi - šuma je u magli (20°C) - do 10.00 sati puno se vode isparava (20°C-25°C) - do 13.30 sati nastaju veliki oblaci (28°C) - između 14-17.00 sati snažne oluje/pljuskovi u kišnom razdoblju (između 30°C i 31°C) - nakon 17.00 sati sunce ponovo sja (28°C) - 18.00 sati sunce brzo zalazi (26°C
Slika xxc. Izgled tropske kišne šume (Hawai, SAD) (lijevo); amazonska džungla u Peru (dolje).

U biom tropskih i suptropske kišnih širokolisnih šuma spada nekoliko tipova šumskih ekosustava:
Nizinske ekvatorijalne vazdazelene kišne šume, obično zvane samo tropske kišne šume, su šume koje dobijaju veliku količinu oborina, više od 2000 mm godišnje.Te se šume nalaze u pojasu oko ekvatora, a najveća područja pod tim šumama su amazonska prašuma u Južnoj Americi, zavala Konga u centralnoj Africi i Indonezija i Nova Gvineja.
Vlažne listopadne i mješane-vazdazelene šume dobijaju najveću količinu oborina tijekom ljeta koje je toplo i vlažno, a zime su hladne i suhe. Neke drvenaste listopadne vrste u ovim šumama gube lišće tijekom suhe zimske sezone. Ove šume se mogu naći u dijelovima Južne Amerike, u Srednjoj Americi, oko Kariba, u obalnoj zapadnoj Africi, u nekim dijelovima indijskog potkontinenta i u većem dijelu Indo-Kine.
Planinske kišne šume se mogu naći na planinskim područjima sa hladnijom klimom.
Poplavne šume
Rasprostranjenost tropskih i suptropskih širokolisnih kišnih šuma Tropske i subtropske širokoline kišne šume javljaju se na nekoliko kopnenih ekozona na više kontinenata, uključujući i afričke trope (ekvatorijalna Afrika), Indomalajski podkontinent (dijelovi indijskog potkontinenta i jugoistočne Slika
Slika xxc. Rasprostranjenost tropskih i suptropskih širokolisnih kišnih šuma

Akvatični (Vodeni) Biomi Vodeni ekosustavi su smješteni u depresijama Zemlje. To je zajednica organizama koji ovise jedni o drugima ali i o vodenom okolišu odnosno vodenom ekosustavu u kojem žive. Postoje dva glavna tipa vodenih ekosustava, (i) slatkovodni ekosustavi i morski ekosustavi. I. Slatkovodni ekosustavi Slatkovodni ekosustavi pokrivaju svega 0,80% Zemljine površine, a od ukupne vode mase čine nastanjuje 0,009%. Slatkovodni ekosustavi stvaraju gotovo 3% neto primarne proizvodnje (procesom fotosinteze) na Zemlji i sadrže 41% svjetskih poznatih vrsta riba. Postoje tri osnovna tipa slatkovodnih ekosustava: • Lentički ekosustav: sporo kretanje vode, uključujući jezera, bazene i ribnjake, • Lotički ekosustav: brzo kretanje vode, na primjer, izvori, potoci i rijeke. • Močvare: područja gdje je tlo prezasićeno ili poplavljeno vodom barem određeni dio vremena. Osim toga, kopnene vode možemo podijeliti u tri osnovne skupine:
Vode stajaćice: jezera, bare, lokve;
Vode tekućice: izvori, potoci, rijeke;
Podzemne vode
Slika. Xxc- Ovo planinsko jezero je primjer lentičkog ekosustava (lijevo), a planinska rijeka je primjer lotičkog ekosustava (desno). 1. Jezero Jezero je vodom ispunjena prirodna depresija na kopnu, koja nema neposredne veze s morem. Većina jezera su slatkovodna i smještena su na sjevernoj hemisferi u području viših geografskih širina. Umjetna jezera su akumulacijski bazeni hidrocentrala, ribnjaci i dr. Dijelovi mora odijeljeni od kopna pješčanim prudovima nisu jezera. Velika

jezera ponekad se nazivaju "unutrašnjim morima" (Kaspijsko i Crno more) a malena se mora ponekad nazivaju jezerima. Termin jezero također se koristi za opisivanje prirodnih obilježja poput jezera Eyre1 koje je većinu vremena suho ali se napuni vodom tijekom sezonskih obilnih kiša. Mnoga su jezera umjetne akumulacije vode koje se grade za proizvodnju električne energije, rekreaciju (plivanje, daskanje na vjetru,...), opskrbu vodom, itd. Finska je poznata kao Zemlja tisuću jezera, a Minnesota (država u SAD-u) kao Zemlja deset tisuća jezera. Velika jezera Sjeverne Amerike podrijetlom su iz ledenog doba. Preko 60% svjetskih jezera nalazi se u Kanadi pretežito zato jer zemljom dominira neulančani sustav otjecanja vode. 1 Jezero Eyre (engl. Lake Eyre) je zatvorena slane jezero u Južnoj Australiji. To je najveće australsko jezero, smješteno u depresiji u suhim klimatskim uvjetima. Ima površinu od 15 000 km ², a doseže maksimalnu dubinu od 20 m. Nalazi se 12 m ispod razine mora.

2 Ladoga je jezero u europskom dijelu Rusije, u blizini rusko-finske granice, po površini je najveće jezero u Europi, a petnaesto u svetu. Po postanku jezera se dijele na:
1. tektonska u tektonskim depresijama (Mrtvo more, istočnoafrička jezera, Bajkalsko, Ohridsko i dr.);
2. vulkanska u kraterima vulkana (Trasimeno u Italiji);
3. glacijalna (ledenjačka) smještena u depresijama (Ladoga2, Bledsko jezero i dr.);
4. krška u kršu (Plitvička jezera) i
5. reliktna jezera, koja su preostala od nekadašnjih većih jezera ili mora i
6. mrtvi riječni rukavi.
Jezera su važna za privredu i promet pojedinih zemalja (Jezera u Sjevernoj Americi, Kaspijsko jezero); dobivanje soli (Mrtvo more); neka su bogata ribom, služe za snabdijevanje vodom (Vransko jezero na Cresu) i reguliranje vodostaja u rijekama. 1.1. Podrijetlo prirodnih jezera Mnoga su jezera mlada zbog erozijskih posljedica koji teže odnošenju jedne od strana bazena u kojemu se jezero nalazi. Postoje brojni prirodni procesi koji oblikuju jezera. Nedavno tektonsko izdizanje planinskog lanca može stvoriti depresije zdjelastog oblika koje mogu akumulirati vodu te stvoriti jezero. Napredovanje i povlačenje ledenjaka može struganjem napraviti depresije u površini gdje će se onda akumulirati jezero. Takva jezera uobičajena su u Skandinaviji, Sibiru i Kanadi. Jezera također mogu nastati klizanjem zemljišta ili glacijalnim preprekama. Primjer nastanka takvog jezera pojavio se tijekom zadnjeg ledenog doba u državi Washington kada je nastalo ogromno jezero iz glacijalnog toka. Naime, kada se led povukao, posljedica je bila ogromna poplava koja je formirala jezero.

Slana jezera su nastala u područjima gdje nema prirodnog otjecanja vode i gdje voda površinski brzo isparava pa dolazi do značajnog povećanja koncentracije soli u vodi. Primjeri slanih jezera uključuju Veliko slano jezero u Utah (SAD-u, Kaspijsko jezero u Rusiji i Mrtvo more u Izraelu. Malena potkovasta jezera nazvana mrtva jezera mogu nastati u riječnim dolinama kao posljedica vijuganja rijeke. Rijeke koje sporo teku stvaraju zavojit oblik pa se vanjska strana zavoja otkida mnogo brže od unutarnje strane. Kao posljedica toga nastaje potkovast zavoj kojeg presijeca rijeka koja počinje teći novim koritom. Potkovasti zavoj biva začepljen muljem rijeke i tako nastaje jezero. Jezero Vostok je jezero na Antarktici koje se nalazi pod ledom i vjerojatno je najveće na svijetu. Pritisak koji stvara led na površinu jezera i unutrašnji kemijski sastav rezultiralo bi štrcanjem vode iz jezera na isti način kao protresena limenka soka ako bi se probušio led iznad jezera. Neka jezera poput Bajklaskog i jezera Tanganyike vulkanskog su podrijetla i leže na geološkim rasjednim linijama. Kratersko jezero u Oregonu nalazi se unutar kaldere ugaslog vulkana. Slika. Xxcc. Jezero Vostok na Antarktici.
1.2. Karakteristike jezera
Promjena razine jezera uočavaju se razlikom između pritjecanja i otjecanja vode u usporedbi s ukupnim obujmom jezera. Značajniji ulazni izvori vode su oborine nad jezerom; višak vode donesen tokovima i kanalima s jezerskog razvodnog područja; kanali podzemne vode i akviferi (vodonosnici), te ljudski izvori izvan razvodnog područja. Izlazni izvori su isparavanje vode iz jezera; površinski i podzemni vodeni tokovi, te bilo kakvo uzimanje jezerske vode za ljudske potrebe. Budući da se klimatski uvjeti i ljudske potrebe za vodom razlikuju, stvaraju se fluktuacije u jezerskoj razini.

Jezera se mogu kategorizirati prema bogatstvu hranjivih tvari, koje tipično utječu na rast biljaka. Jezera siromašna hranjivim tvarima nazivaju se oligotrofnim i općenito su prozirna te imaju nisku koncentraciju biljnog života. Mezotrofna jezera imaju dobru jasnoću i srednju razinu hranjivih tvari. Eutrofna jezera su obogaćena hranjivim tvarima, pa stoga imaju dobar rast biljaka kao i moguće cvjetanje algi. Hipertrofno jezero je vodena masa visoko obogaćena hranjivim tvarima. Ta jezera imaju slabu jasnoću i predmet su cvjetanja algi. Jezera tipično dostižu takvo stanje zahvaljujući ljudskim aktivnostima poput obilne upotrebe minealnih i organskih gnojiva u jezerskom razvodnom području. Takva jezera imaju gotovo nikakvu korisnost, a ekosustav je gotovo uništen.
1.3. Abiotička i biotička limnologija
Limnologija 3 ekosustav jezera dijeli na tri zone (slika xxc.): (i) litoralna zona, je rubno područje jezera, dakle nagnuto područje blizu zemlje;(ii) zona otvorene vode, gdje je sunčeva svjetlost obilna; i (iii) zona duboke vode, gdje dopire malo sunčeve svjetlosti. Dubina koju svjetlost može dostići u jezerima ovisi o gustoći i gibanju čestica. Čestice u vodi mogu biti sedimentarnog ili biološkog podrijetla i daju boju jezerskoj vodi. Raspadnuta biljna tvar daje žutu ili smeđu boju vodi, dok su alge daju zelenkastu boju vodi. U vrlo plitkim vodenim masama željezni oksidi daju vodi crvenkasto smeđu boju. U biološke čestice spadaju alge i detritus4. Sedimentna čestica je u suspenziji ako je njena težina manja od nasumičnih sila zamućivanja koje djeluju na česticu. Zamućivanje je presudni faktor u prozirnosti vode. Pridnevne detritivorne ribe odgovorne su za muljevite vode jer one u potrazi za hranom uzburkaju mulj. Piscivorne ribe jedu biljojede (planktonivorne) ribe, te stoga povećavaju broj algi (vidi akvatrofna kaskada). Dubina do koje dopire svjetlost ili prozirnost vode mjeri se upotrebom Secchijevog diska. To je disk promjera 20 cm s naizmjeničnim bijelim i crnim kvadrantima. Dubina na kojoj se disk više ne vidi je Secchijeva dubina, tj. mjera prozirnosti. Obično se koristi za testiranje eutrofikacije. 3 Limnologija (grčki λἰ jezero + -logija), grana hidrologije koja se bavi istraživanjem jezera, napose μνηfizikalnim i kemijskim svojstvima jezerske vode i biologijom jezera. François-Alphonse Forel (1841. - 1912.) je osnovao polje limnologije vlastitim proučavanjem Ženevskog jezera. Limnologija je tradicionalno blisko srodna hidrobiologiji koja se bavi primjenom principa i metoda fizike, kemije, geologije i geografije na ekološke probleme u jezerima. 4 Svježi do djelomično razloženi biljni i životinjski materijal
Jezero ublažava temperaturu i klimu okolnog područja zbog vode koja ima vrlo visok specifični toplinski kapacitet (4186). Osim toga jezero može tijekom dana ohladiti okolni prostor uz pomoć lokalnih vjetrovima, stvarajući povjetarac s vodene površine ili noću može zagrijati okolni prostor, stvarajući povjetarac s kopna.

Slika xxc. Zone jezera. Slika xxc. Litoralna zona jezera. Chara je alga, ostalo su biljke cvjetnjače.
1.4. Kako nastaju jezera?
Postoji važna razlika između nizinskih i gorskih jezera. Nizinska su jezera mirna, manje stjenovita/više sedimentna, imaju blago nagnuto dno te općenito sadrže više biljnog života. Velike vodene biljke (posebice trstika) značajno ubrzavaju taj proces zatvaranja jer zadržavaju sediment. Muljevita jezera i jezera s mnogo biljojednih riba teže polaganom nestajanju. "Nestajuće" jezero (jedva primjetno na čovjekovoj vremenskoj skali) tipično ima vodeni rub s ekstenzivnim biljnim pokrivačima. Oni postaju novo stanište za ostale biljke (poput maha tresetara kada su stvore za njega povoljni uvjeti) i životinje od kojih su mnoge vrlo rijetke. Postupno se jezero zatvara pa može nastati mladi treset koji stvara baru. U nizinskim riječnim dolinama (gdje rijeka može stvarati meandre), prisustvo treseta se objašnjava zatvaranjem bivših mrtvaja. U vrlo kasnim stadijima sukcesije raste više stabala konačno pretvarajući vlažno zemljište u šumu. Neka jezera mogu nestati u određenom dijelu godine. Takva jezera se nazivaju intermitentnim jezerima i tipična su za krški teren. Izvrstan primjer takvih jezera je jezero Cerknica u Sloveniji.

Slika. Xcc. Primarna sukcesija od jezera, močvare i bare do vlažne livade. U vodeni stupac bare vremenom se nataloži organska tvar od biljaka i životinja, koja postupno smanjuje vodeni stupac i bara se pretvara u vlažnu livadu gdje se useljavaju trave vlažnih livada i u konačnici se razvije šuma.
1.5. Značajnija jezera u Svijetu
Najveće jezero na svijetu je Kaspijsko jezero s površinom od 394,299 km² što je veće od područja koje zajedno pokrivaju sljedećih šest najvećih jezera.
Najveće slatkovodno jezero i drugo najveće jezero ukupno je jezero Superior s površinom od 82,414 km². Najveće je od pet Velikih jezera u Sj. Americi. Po volumenu slatkovodne vode treće u svijetu. Na sjevernoj obali jezera nalaze se kanadska pokrajina Ontario i američka savezna država Minnesota, a na južnoj američke savezne države Wisconsin i Michigan.
Najdublje jezero je Bajkalsko jezero u Sibiru dubine 1,741 m.
Najviše plovno jezero je jezero Titicaca nadmorske visine 3821 m. Ujedno je i drugo jezero po veličini u Južnoj Americi.
Najniže svjetsko jezero je Mrtvo more, 396 metara ispod morske razine. To je jezero s najvećom koncentracijom soli.
Najveći otok na slatkovodnom jezeru je otok Manitoulin na jezeru Huron površine 2,766 kvadratnih km. Huron je jezero u Sjevernoj Americi jedno od pet Velikih jezera, na granici SAD-a i Kanade, tj. američke savezne države Michigan i kanadske pokrajine Ontario. Jezero Huron je drugo po veličini od pet Velikih jezera površine 59.596 km2 i treće slatkovodno jezero po veličini u svijetu. Jezero Huron sadrži 3.540 km3 vode, duljina obale iznosi 6.157 km, jezero se nalazi nadmorskoj visini od 176 m prosječna dubina iznosi 59 m, a najveća je 229 m. Jezero je dugo 332 km, a najveća je širina 245 km. Na jezeru se nalazi otok Manitoulin najveći otok na svijetu koji je okružen slatkom vodom.
Najveće jezero smješteno na otoku je jezero Nettilling na Baffinovom otoku, Nunavut, Canada.
Jezero Toba na otoku Sumatri smješteno je u vjerojatno najvećoj probuđenoj kalderi na Zemlji.

Najveće slatkovodno jezero u Europi je Ladoga iza koje se nalazi Onega, obje smještene u Rusiji.
Jezero Victoria najveće je u Africi. Dio je Velikih jezera Afrike.
Jezero Superior je najveće jezero u Sjevernoj Americi. Dio je Velikih jezera.
Jezero Maracaibo je najveće jezero u Južnoj Americi. Jezero Titicaca je najveće slatkovodno jezero.
1.6. Eutrofikacija: problem u porastu
Eutrofikacija je proces obogaćivanja vode hranjivim tvarima, a jezera s visokim sadržajem hranjivih tvari su eutrofična. Voda u eutrofičnim jezerima mutna i nalikuje juhi od graška, u prvom redu zbog prisutnosti golemog broja algi i cijanobakterija koje se razvijaju na hranjivim tvarima. Prirodna eutrofikacija jezera se događa kada je voda obogaćena velikim brojem uginulih biljnih i životinjskih organizama koji s drugim tvarima stvaraju jezerski sediment tijekom dugog razdoblja. Postupno, vodene biljke počinju se intenzivno razvijati u hranjivim tvarima bogatim sedimentima, i tako počinju ispunjavati plitke vode, tvoreći močvare. Eutrofikaciju mogu znatno ubrzati ljudske aktivnosti, a to proizlazi iz obogaćivanja vode organskim i anorganskih hranjivim tvarima potrebnih za rast i razvoj biljaka i algi. To se događa ispiranjem mineralnih tvari iz

tla i pritjecajem otpadnih voda u jeze Slika xxc. Antropogeni i prirodni proces eutrofikacije.
2. Močvare
Močvare su plitke vode stajaćice, odnosno tlo natopljeno najčešće slatkom vodom, koje se odlikuje niskom pH vrijednošću (dakle kiselošću) i biološkom raznolikošću staništa za mnoge biljne i životinjske vrste, kao što su vodozemci, ribe, ptice, kukci, gmazovi i sisavci. Močvaru kao čest oblik voda stajaćica treba razlikovati od a) lokve, plitke udubine sa vodom koja presušuje tijekom sušnog razdoblja b) bare, plitka vodena masa kojoj sunce dopire do dna i odlikuje se bujnim biljnim raslinjem

c) jezera, koja na zemlji nastaju tek nakon ledenog doba i u pravilu se razlikuje eufotički5 i afotički sloj. 5 Plankton se javlja u zoni slobodne vode (limnion, pelagijal) koja nije jedinstvena već ju dijelimo na eufotički i afotički sloj. Eufotički je gornji, osvijetljeni sloj koji se, uglavnom, poklapa s dubinom prodora svijetla. U ovom sloju se odvija i organska produkcija, pa ga nazivamo i trofogeni sloj.

Afotički je donji, neosvjetljeni sloj koji se proteže od granice prodora svijetla do dna. Budući da ovdje nema svijetla, nema ni fotosinteze, pa nema ni fitoplanktona. Rijetki su i zooplanktonski oblici. Močvare se najčešće nalaze u riječnim nizinama ili uz jezera, odnosno u reljefnim depresijama s intenzivnom vegetacijom. U močvarama se, za razliku od tresetišta, ne stvara treset. Prema porijeklu i načinu dotoka vode močvare se dijele na:
niske, koje se "hrane" samo podzemnom vodom; u ovih močvara, vegetacijski sloj je deblji
visoke, koje se "hrane" samo površinskom vodom, praktično jedino oborinskim vodama; ovdje je sloj vegetacije tanak
mješovite, koje se prihranjuju iz oba izvora; to je najčešći tip močvara.
Isušivanje močvara obavlja se melioracijom pomoću drenažnih kanala i podizanjem nasipa. Na taj se način stvara novi životni okoliš u kojem ne rastu stabla, ponekad ima grmlja, a općenito se tu nalaze ekološke zajednice koje uspijevaju na mokrom mineralnom pa do organskom tlu, a mogu opstati samo uz neprekidni višak vode. Razgraničenje između močvara i drugih tipova biotopa nije jednostavno. One su među oblik između drugih biotopa kao što su tresetišta, vodene površine koje se isušuju ili mokre livade i grmišta. Tipične biljne vrste su iz rodova Eupatorium i Lysimachia, kao i neke druge biljne zajednice koje su vrlo važne kao "paša" za leptire i druge kukce. Mnoge su močvare ugrožene ili su u prošlom stoljeću i ranije potpuno isušene zbog potreba poljoprivrede za obradivim površinama ili iz zdravstvenih razloga (malarija). U novije vrijeme je došlo do promjene odnosa prema močvarama. Shvaćen je njihov značaj za ukupnost čovjekove životne sredine, i česti su uspješni napori za renaturiranjem tih područja.

Slika. Xxc. Izgled močvare u dijelu Hutovog Blata
3. Bare
Bare su prirodna udubljenja nepravilnog oblika, odnosno plitke vodene mase gdje žive razne biljke i životinje od zone niskih do zone visokih biljaka kao i od zone plitke i duboke vode. Bare se od močvara razlikuju zbog toga što svjetlost sunca u pravilu dopire do dna. Slika. Xxc. Posebni ornitološki rezervat Bara Dvorina, Slavonski Brod, Donja Bebrina,,
4. Ribnjak

Ribnjak je umjetno napravljen vodeni bazen s važećom dozvolom za rad, namijenjen za uzgoj ribe. Voda ribnjaka može da bude slatka ili slana. Ribnjak ima osiguranu infrastrukturu za punjenje i pretapanje vode te osigurava potpuno pražnjenje jezera za vrijeme ribolova. Tokom proizvodnje (uzgoja ribe), ribe se dodatno prehranjuju. Slika. Xxc. Ribnjaci Krupa na rijeci Vrbasu u Bosni i Hercegovini
5. Estuarij
Estuarij (lat. aestuarium; nisko riječno ušće) je naziv za riječno ušće oblikovano poput lijevka. Estuariji su rjeđi od ušća u obliku delte. Takav oblik imaju ušća imaju rijeke Temzea, Elbea, Oba, Jeniseja i niza drugih. Svakako je najupečatljiviji estuarij Amazone, čije je ušće široko preko 30 km. Na ušću postoji otok Marajo, kao i niz drugih, manjih otočića, koji nisu nastali kao riječni nanosi pijeska, nego je otočni pijesak nanos kojeg je donijela morska plima i oseka i pretvorila ga u otočiće. 5.1. Nastajanje i osnovna obilježja estuarija

Estuariji nastaju i oblikuju se pod utjecajem plime i oseke, pri čemu ušće dobijaju oblik lijevka. Na sjevernoj zemljinoj polutki pod utjecajem Coriolisove sile6 morska voda za vrijeme plime ulazi u rijeku snažnije uz lijevu obalu, dok se za vrijeme oseke u more vraća duž desne obale.

6 Coriolisova [Koriolisova] sila je komponenta inercijske sile uzrokovana rotacijom Zemlje. Djeluje okomito na zemaljsku os i na trenutačni smjer gibanja zračnih čestica i svakog tijela koje se giba na Zemlji. Posljedica djelovanja Coriolisove sile jest promjena smjera gibanja (posebice vjetrova i morskih struja) i to na sjevernoj polutki udesno, a na južnoj ulijevo. Estuariji se stvaraju naročito uz obale gdje su razlike između plime i oseke velike. Zbog toga su i rjeđi od delti rijeka. Mogu nastati samo kad je količina materijala koji donosi rijeka manji od količine koju morska voda u vrijeme oseke odnosi u more. Nasuprot tome ušća rijeka se oblikuju u obliku delte. Naime, u tom slučaju rijeka se dijeli na niz rukavaca koji nastaju zbog gomilanja sedimentnih nanosa različitog materijala koji sa sobom donosi rijeka. Daljnje obilježje estuarija je prijelaz od riječne, slatke vode prema boćatoj odnosno morskoj vodi. Nastajanjem boćatih voda stvaraju se uvjeti za razvoj posebnog biljnog i životinjskog svijeta koji se nalazi na prijelazu od područja rijeke prema moru. Slika xxc. Helgolandski zaljev s estuarijem rijeke Elbe na sjeveru Njemačke. 5.2. Neke poznatije rijeke s estuarijem
Amazona, sjeverni dio ušća, dok se na južnom dijelu tvori delta
Elba
Gironde kao zajednički estuarij Garonne i Dordogne
Jenisej
Loire
Ob
Temza

6. Laguna
Laguna je plitko područje slane ili bočatne vode7 uz morsku obalu koje je u manjoj ili većoj mjeri zatvoreno. Obično je odijeljena od otvorenog mora sa niskim pješčanim otocima ili koralnim prevlakama. Lagune se obično nalaze uz niske pješčane morske obale, posebno na ušćima rijeka u more. Lagunama se također nazivaju i kanali između kopna i koralnih grebena, i vode u atolima8 (jedna vrsta otoka). 7 Boćata voda u sebi sadrži više soli nego obična slatka voda, a manje soli nego što je ima morska voda; nastaje miješanjem morske i riječne vode. 8 Atol je koraljni otok. Okružuju ga korali koji rastu oko vulkanskih otoka u tropskim morima, a naročito u Tihom i Indijskom oceanu. Atoli imaju oblik prstena, ali i polukruga, u sredini je plitki zaljev — laguna. Veći atoli su naseljeni, tamo gdje nema izvora slatke pitke vode, život stanovnika ovisi o kišnici. Riječ atol dolazi iz maldivskog jezika. U engleskom jeziku prvi put je spomenuta 1625. god., a termin je usvojen od strane Charlesa Darwina 1824. god., koji je opisao atole kao posebnu vrstu otoka. Slika. Xxc. Područje lagune-di-Marano, sjeverna Italija Gotovo pola područja Kiritimata je pokriveno lagunama, slatkovodnim i slanim vodama. Poznate su lagune na obali Venecijanskog zaliva, Baltičkog mora, Crnog mora i Meksičkog zaliva. Riječ laguna se počela koristiti u engleskom jeziku 1769. godine, i odnosila se na Venecijansku lagunu (porijeklo riječi je iz Latinskog jezika, lacuna - prazan prostor). U španjolskom jeziku riječ laguna se koristi za pojam jezera poput Laguna Catemaco (Bos. Jezero Catemaco).
7. Riječna delta
Riječnom deltom se naziva karakteristični oblik ušća rijeke. Najpoznatija delta je delta Nila, koja je dobila naziv po sličnosti oblika grčkom slovu delta (Δ) i time postala sinonim za sve druge

delte širom svijeta. Nastajanje riječne delte je vezano je za odlaganjem sedimenta koji rijeka donosi sa sobom. Smanjenjem brzine toka vode kako se približava ušću, smanjuje se i snaga vode kojom nosi razne vrste sedimenta, koji se zbog toga taloži na dno i stvara prepreku. To opet dovodi do toga da voda mijenja smjer kako bi zaobišla prepreku, ili se razdvaja na rukavce. Veće delte se sastoje od niza brže i sporije tekućih ili čak stojećih rukavaca. Slika xxc. Delta rijeke Nila, snimljena iz satelita (lijevo). Delta rijeke Neretve (desno) To, međutim, nije moguće kad rijeke završavaju u morima sa velikim razlikama između plime i oseke, umjesto delte, kod tih se rijeka stvara ljevkasti estuarij. Kako rijeka teče u pravilu prema moru odnosno jezeru, kad stigne do vode koja ne teče, odlaže sediment, i time svoju deltu neprekidno "gura" dalje u more. To je vrlo vidljivo na snimci delte Nila (usp. sliku). Delta rijeke Neretve - na svome ušću, na jugu hrvatske obale Jadranskog mora deltu je formirala rijeka Neretva. Delta prekriva površinu od 12000 ha. Delta Neretve od Metkovića do ušća sa sjevera i sjeveroistoka omeđena je ograncima dinarskih planina, a s juga podgradinsko-slivanjskim brdima..Najvažniji gradovi su Ploče, Opuzen i Metković. Izvorno se sastojala od 12 rukavaca, ali je pod utjecajem čovjeka te nakon brojnih i opsežnih melioracija i kultivacije tog poljoprivredno bogatog prostora, broj rukavaca sveden na samo četiri. Vrste delti Pored ovdje navedene vrste delte, postoji još nekoliko drugačijih oblika delti: Unutrašnja delta – se naziva takva vrsta riječne delte gdje rijeka prolazi kroz depresiju ili nizinu, ili završava u bazenu iz kojeg ne izlazi niti jedan vodotok. Neki primjeri takvih unutrašnjih delti s poplavnim nizinama i vlastitim otjecanjem su:
Elba kod Hamburga (Njemačka, Europa)
Niger u nizini Massina (Mali, Afrika) (rijeka Niger na kraju svog toka tvori i riječnu deltu)
Ob u zapadnosibirskoj nizini u Sibiru (Rusija, Azija) (Ob na kraju svog toka tvori deltu ali i estuarij)

Delta obilježena plimom i osekom Delta obilježena valovima Delta kojom dominira rijeka Poznatije delte rijeka Dunav (Delta Dunava; 4.000 km², Rumunjska) - druga po veličini riječna delta u Europi Ganges-/Brahmaputra (Delta Gangesa Bangladeš; 80.000 km²) Ind (8.000 km²) Mekong (70.000 km²) Mississippi (Mississippi delta; 36.000 km²) Niger (Delta Nigera 24.000 km²) - u srednjem dijelu toka Niger tvori i unutrašnju deltu Nil (Delta Nila 24.000 km²) Irawadi (30.000 km²) Lena (45.000 km²) Orinoco (24.000 km²) Volga (Rusija) - najveća delta u Europi Neretva (Delta Neretve, 120 km²) - jedina hrvatska delta
II. Morski ekosustavi
Morski ekosustavi pokrivaju oko 71% Zemljine površine i sadrže oko 97% vode na planetu Zemlji. Oni generiraju oko 32% svjetske neto primarne proizvodnje. Od slatkovodni ekosustava se razlikuju po prisutnost otopljenih tvari u vodi, osobito soli,. Oko 85% otopljenog materijala u morskoj vodi su natrij i klor. Prosječna slanost morske vode je oko 35%. Stvarni salinitet morske vode varira između različitih morskih ekosustava. Morski ekosustavi mogu se podijeliti u slijedeće zone: 1. oceanska (otvoreni dio oceana, gdje žive morske životinje, kao što su kitovi, morski psi, i tune i druge); 2. donje ili duboke vode); 3. bentos (morsko dno). Brojni organizmi pronađeni u morskim ekosustavima uključuju smeđe alge, dinoflagellates, koralje, glavonožaca, fito i zooplankton. Mnogobrojne ribe koje potječu iz morskih ekosustava su najveći izvor mesa (hrane) dobivene od divljih populacija. Ekološki problemi koji se javljaju u morskim ekosustavima su

Slika xxc. Oceanska zone neodržive eksploatacije morskih resursa, kao na primjer pretjeran izlov pojedinih vrsta riba, onečišćenja mora otpadnim vodama i naftom uzrokovano naftnim havarijama, preizgađenost građevnih objekata u obalnoj zoni i slično.
1. Priobalje (litoralna zona)
1.1. Općenito o priobalju
Priobalje (litoral) je podijeljeno na više mikroregija koje, zavisno od toga o kojem se okružju radi (more, rijeka ili jezero), mogu biti različite. Tako npr. priobalje u jezerima obuhvaća priobalnu terasu do 30 metara dubine, a u plićim jezerima i manje. Isto tako, značenje izraza "priobalje" ili "litoral" u različitim dijelovima svijeta ili u različitim znanstvenim disciplinama može biti sasvim različito. Tako npr. u ratnoj mornarici kada se govori o "priobalju" ili "litoralnom području" onda se ono po svom značenju znatno drugačije interpretira od onog kako ga definiraju i shvaćaju pomorski biolozi. Priobalje općenito obilježava gibanje vode, erozija, jaka sedimentacija, kolebanje temperature, te bogatstvo biljnog i životinjskog svijeta. Sve to često stvara posebne mikroklimatske uvjete pogodne za razvoj jedinstvenih vrsta organizama. Priobalje je dio mora, jezera ili rijeke uz obalu. To je područje obale koje se proteže od crte visoke vode (plime ili visokog vodostaja) do crte niske vode (oseke ili niskog vodostaja). No, izraz

"priobalje" se koristi i za područja koja mogu biti znatno udaljena od područja između plime i oseke (na moru) ili između visokog i niskog vodostaja (na rijekama i jezerima) i ne postoji jedinstvena definicija značenja toga izraza. U hrvatskom jeziku, osim riječi priobalje, često se koristi i riječ litoral ili litoralno područje što dolazi od latinske imenice "litus, litoris" što znači "obala". U priobalnoj (litoralnoj) zoni nalazimo: (1.) supralitoral, (2.) mediolitoral, (3.) sublitoral, (3.a.) infralitoral i (3.b.) cirkalitoral.
1.2. Supralitoral
Supralitoral je prema definiciji prva po redu stepenica priobalja odnosno litorala, počevši od kopna prema moru. Supralitoral je stalno izvan vode, a more ga vlaži samo prskanjem i/ili razlijevanjem valova. Visina te stepenice vlaženja varira, ovisno o izloženosti obale prema moru, koja može biti od pola metra na zaštićenim mjestima pa do nekoliko metara u visinu na mjestima gdje je obala izložena vjetru koji nosi kapljice mora. U tome se području nastanjuju organizmi prilagođeni ekstremnim i vrlo promjenjivim životnim uvjetima. Slika xxc. Podjela litoralne zone počevši od kopna prema otvorenom moru.
1.3. Mediolitoral
Mediolitoral ili eulitoral je prema definiciji druga po redu stepenica priobalja odnosno litorala. To je područje izmjene plime i oseke koje se proteže od gornje granice visine plime do donje granice normale oseke. Za vrijeme plime područje mediolitorala je uronjeno u more, a za vrijeme oseke je izvan mora, pa stoga ekološki čimbenici kao što su temperatura, vlažnost, osvijetljenost i dr. u tom području jako variraju.

1.4. Sublitoral
Sublitoral je naziv za morsko priobalno (litoralno) područje koje se stalno nalazi pod morem. U oceanografskom smislu, sublitoral obuhvaća priobalje (litoral) na koje značajno utječu morske mijene9 i na kojima dolazi do gubitka energije valova, morskih struja i sl., a koje se ponekad proteže do dubine od 200 m. 9 Morske mijene su plima i oseka, te uz morske struje i valove jedini prirodni pokretači vodenih površina U pomorskoj biologiji sublitoral se shvaća kao područje mora do dubina na kojima sunčevo svjetlo dopire do morskog dna u kojemu živi najveći broj morskih organizama. Unutar sublitorala biolozi razlikuju dva priobalna morska područja, i to: a) Infralitoral i b) Cirkalitoral.
a.) Infralitoral
Infralitoral je treća po redu stepenica priobalja (litorala). Proteže se od donje granice oseke pa do dubine od oko 50 metara (može biti plići ili dublji ovisno o prodoru svjetla). To je područje fotofilnih alga koje se razvijaju na stjenovitoj podlozi i morskih cvjetnica na pomičnoj podlozi.
b.) Cirkalitoral
Cirkalitoral je četvrta po redu stepenica priobalja (litorala). Proteže se od donje granice rasprostranjenosti fotofilnih alga i morskih cvjetnica pa do dubine do koje se razvijaju scijafilne alge, obično od 120 do 200 metara dubine. Ispod 200 metara svjetlost koja prodire više nije dovoljna za fotosintezu pa ni razvoj alga više nije moguć. S dubinom se broj biljnih vrsta smanjuje, a životinjskih povećava.
2. Mangrove
Mangrove su poseban tip vegetacije koja se razvija na muljevitim terenima zaljeva, laguna i ušća rijeka u tropskim područjima. Glavna obilježja mangrova uglavnom čine vazdazelene drvenaste biljke. U području umjerenog klimatskog pojasa na ovim se terenima nalaze slane močvare.

Slika. xxc. Ekosustav mangrova (lijevo); vazdazeleno drveće mangrova s korijenjem (desno)
Ekološki uvjeti u mangrovama i njihova biologija U mnogim staništima mangrova može se pronaći velika raznolikost biljnih vrsta. Od utvrđenih 110 biljnih vrsta, samo 54 vrste koje spadaju u 20 rodova i 16 porodica čine prave mangrove, tj vrste koje se mogu pronađi samo u prirodnom staništu mangrova i retko drugde. Evolucija je mnoge od ovih vrsta gotovo podjednako prilagodila na uvjete staništa koja imaju vrlo različiti stupanj slaniteta vode, visinu plime i seke, anaerobne uvjete zemljišta i intenzivno sunčevo zračenje budući da se radi o tropskim područjima. Biološka raznolikost u mangrovama uglavnom je niska – rijetko se nalazi više od 20 biljnih vrsta. To se posebno odnosi na područja većih geografskih širina i u području obje Amerike. Najveća biološka raznolikost može se naći u šumama Nove Gvineje, Indonezije i Malezije. Šume mangrova su prilagođene malim količinama kisika. Tako npr. crvene mangrove, koje većinom žive u poplavljenim područjima izdignu se iznad razine mora putem dugačkog korijena i tako uzimlju kisik preko pora (lenticela) u kori drveta. S druge pak strane crne mangrove žive na višem zemljištu i obrazuju mnogo pneumatofora (specijaliziranih korjenovih struktura koje izbijaju iz zemlje poput slamčica za disanje) koje su prekrivene lenticelima. Ove „slamčice za disanje“ obično dostignu visinu od 30 centimetara, a u nekih vrsta i preko 3 metra. Postoji četiri tipa pneumatofora – štulaste, klinaste, kolenaste i trakaste. Kolenast i trakast tip se mogu kombinovati sa potpornim korenima u osnovi drveta. Korenja takođe imaju velike aerencime koje služe da ubrzaju kretanje kiseonika u biljci.

Slika xxc. Primjer crne mangrove (Avicennia germinans) na otoku Mustang. Obratite pažnju na pneumatofore oko mangrove (lijevo); primjer crvene mangrove (Rhizophora mangle) sa zračnim korijenjem (desno). Ograničavanje unosa soli Crvene mangrove ne propuštaju višak soli iz vode pomoću suberina koji se nalzi u kori korijena, a koji djeluje kao super-filtar za natrij-klorid. Analiza vode unutar biljaka mangrove pokazuje da se od 90% do 97% soli već odstranjuje u korijenu. Da bi se biljka oslobodila viška soli, ona se akumulira u biljnim izdancima, a zatim se koncentrira u starim listovima koji kasnije otpadaju. Isto tako višak soli se uskladišti u staničnim vakuolama. Bijele (ili sive) mangrove mogu lučiti sol izravno. Naime, bijele mangrove imaju dvije žlijezde pri bazi lista, gdje se kristalizira sol (biljka prekrivena kristalima soli), pa otuda se ta vrsta mangrova naziva bijele mangrove.

Slika. Xxc. Primjer bijele mangrove (Laguncularia racemosa) koja raste iznad plime i oseke. Ograničavanje gubitka vode Zbog ograničene dostupnosti sveže vode u slanom zemljištu, stanište mangrova je pod utjecajem plime i oseke, stoga su mangrove razvile način na koji ograničavaju gubljenje vode kroz lišće. Naime, biljke mangrova mogu smanjiti ili potpuno zatvoriti pore na površini listova (stome) kroz koje izmjenjuju kisik, CO2 i vodenu paru tijekom procesa fotosinteze. Biljke mangrove također mijenjaju položaj lišća kako bi se izbjegle jako popodnevno sunce i tako smanjile isparavanje iz lišća. Unošenje hranljivih tvari Najveći problem za biljke mangrova je unošenje hranljivih tvari iz okoliša. Pošto je stanište stalno poplavljeno u tlu se nalazi vrlo malo slobodnog kisika. Stoga anaerobne bakterije oslobađaju dušik, razgrađuju željezo, neorganske fosfate, sulfide i metan, čime tlo stvara još neplodnijim i manje hranjivim za biljke, a što još dodatno doprinosi oštrom i neugodom mirisu mangrova. Izdignuti sustav korijenja omogućava biljkama mangrova usvajanje plinova (O2, CO2 i H2O) izravno iz atmosfere, a ostale hranljive tari iz siromašnog i negostoljubivog tla. Atmosferski plinovi koje koristi biljka često su uskladišteni direktno unutar korijena i protječu kroz biljku čak i kada je korijenje potopljeno duboko pod vodom uslijed djelovanja plime. Biologija rasploda i širenje biljaka mangrova Biljke mangrova povećavaju uvjeta za preživljavanje potomaka. Naime, u vrlo oštrim uvjetima, biljke mangrova su razvile poseban mehanizam kojim pomažu/osiguravaju svojim potomcima da prežive i opstanu na ovim staništima. Tako npr. sve mangrove imaju plutajuće sjemenke koje su pogodne za širenje putem vode. Za razliku od većine drugih biljaka, čije sjemenke klijaju u zemlji, sjemenke mnogih biljaka mangrova (kao npr. crvenih mangrova) počinju već klijati dok se nalaze vezane za drvo biljke-roditelja. Kada sjemenke prokliju, klica raste unutar ploda (kao npr. u vrsta Aegialitis, Acanthus, Avicennia i Aegiceras) ili kroz plod (kao npr. u vrsta Rhizophora, Ceriops, Bruguiera i Nypa) i stvaraju propagulu (klicu) koja je sama sposobna stvarati hranu putem fotosinteze. Kada propagula sazri, ona pada u vodu, koja se može prenjeti na velike udaljenosti. Propagule mogu preživjeti isušivanje i ostati u dormantnom stanju nekoliko tjedana ili mjeseci, pa čak i godinu dana prije nego što ponovno dođe do pogodnih uvjeta za njihov rast i razvoj. Kada je propagula spremna da stvori korijen, ona mijenja gustoću određenih tkiva tako da izduženi dio biljke počinje

plutati vertikalno, a ne više horizontalno. U ovom će položaju plutati i pristati odnosno zabosti se u mulj gdje će se ukorijeniti. Ako se ne zabode u mulj, propagula će ponovno izmijeniti gustoću tkiva i ponovno će nastaviti plutati u potrazi za boljim uvjetima za ukorjenjenje. Korijenje drvenastih biljaka mangrova (kao što je slučaj kod Rhizophora koja ima potporno korijenje) zadržava sedimente koje nadalje nagomilavanjem stvara podlogu na kojoj se može nastaniti i rasti drugo drveće. Na taj se način širi šuma mangrova na račun mora. Slika. Xxc. Mangrove (Rhizophora sp.) u Queensland, Australija Ekološki uvjeti Kao i u svim drugim ekosustavima i u šumama mangrova svi se ekološki čimbenici odlikuju istovremenim uvjetima koji su ključni za razvoj vegetacije mangrova. Među posebno važne čimbenike koji utječu na životne forme svake biljne vrste koja nastanjuje šumu mangrova, ističe se prilagodba povećanoj koncentraciji soli, nedostatku kisika i promjeni razine vode, odnosno plimi i oseki. Ovi čimbenici također imaju utjecaj na prostornu i vremensku organiziranost, odnosno spratovnost šuma mangrova, a time i na sam izgled biocenoze mangrova. U pojedinim područjima Svijeta dominiraju različite biljne vrste, pa se tako mogu razlikovati bijele, crne, crvene, kao i mangrove palmi koje se susreću u jugoistočnoj Australiji. Rasprostranjenost

U području SAD-a šume mangrova su ograničene na obalni dio poluotoka Florida i izoliranih područja na južnoj obali zaljeva Luizijane i na jugu Teksasa. U Sjevernoj Americi močvare se na Floridi protežu južno od jezera Okičobi i zauzimaju više od 28.000 km površine. Južni dio ovih močvara pretvoren je u nacionalni park. U tim se područjima voda izljeva iz jezera u Meksički zaljev. Uz samu morsku obalu nalazi se pojas šuma mangrova koji je praktički neprohodan. Taj pojas čine crvene, crne i bijele mangrove. Dublji jarci su pod vodom cijelu godinu. U jugoistočnoj Aziji mangrove se javljaju u vidu pojaseva, od kojih se svaki odlikuje specifičnom skupinom vrsta drveća. Na te šume mangrova, koje se još nazivaju mangal obično se nadovezuju nizinske tropske šume u kojima se javljaju neke od istih vrsta koje se nalaze u šumama mangrova. U Australiji, mangrove se javljaju oko Nove Gvineje, Sulavesija i susjednim otocima. U jugoistočnoj Australiji se razlikuju pojedine biljne zone, od kojih šume mangrova čine zadnji pojas zapadnih i vlažnih padina okrenutih prema Tihom oceanu. U Australiji je oko 11.500 km² prekriveno šumama mangrova koje dopiru do krajnjeg juga do uvale u Viktoriji (37° 45') i Barkerovoj uvali u Adelejd. Novi Zeland ima šumovite mangrove na oko 38° (slično australijskom najjužnijem opsegu mangrova): krajnja geografska dužina na zapadnoj obali je luka Raglan (37 ° 48'), na istočnoj obali, luka Ohiva je krajnji jug gdje se javljaju mangrove (38° 00'). Mangrove se javljaju u Africi i Južnoj Americi, gdje „prerastaju“ u tropske kišne šume, ali se također javljaju i na brojnim otocima. Značajna područja pod šumama mangrova nalaze se u Keniji i na Madagaskaru, koje se miješaju sa obalskim madagaskarskim suhim listopadnim šumama. Nigerija ima najveća područja pokrivena mangrovama u Africi, koja zauzima oko 36.000 km². Veliki dio tih područja je uništen u posljednjih 50-tak godina uslijed izlijevanja nafte u more uglavnom zbog havarije tankera, što je imalo izravan negativan utjecaj na ribolov i gospodarstvo tih područja. Slika xxc. Područja Svijeta koje prekrivaju šume mangrova

Životni oblici biljaka mangrova Biljke mangrova su prilagođene na povišenu koncentraciju soli i mogu podnijeti periodično potapanje tokom plime. Veliki broj biljaka su sukulente jer u mangrovama vlada fiziološka suša. Naime, koncentracija soli u morskoj vodi i staničnom soku biljaka je jednaka, pa je tako onemogućena izmjena mineralnih tvari između ove dvije sredine. Da bi biljke preživjele u ovakvim uvjetima, one deponiraju rezervnu vodu u pojedinim organima. U različitih biljnih vrsta otpornost na povišenu koncentraciju soli u je različita. Isto tako, različite biljne vrste različito podnose i potapanje u morskoj vodi uslijed plime i oseke, pa se tako javlja nekoliko zona, koje su jasno uočljive u području jugoistočne Azije. Problem nedostatak kisika neke biljke su rješavaju putem korijenja za ventilaciju. Naime njihov korijen raste negativno geotropno i stvara se veliki broj takozvanih pneumatofora iz kojih izrasta bočno korijenje. Na periferiji takvog korijenja u kori se stvara tanki omotač koji se u gornjem dijelu ljušti i na taj način omogućava prolazak kisika do unutrašnjeg tkiva (arenhima). Takav je slučaj u vrsta roda Soneratia. U palme Areca catechu funkciju pneumatofora su preuzeli kratki bočni korjenovi koji odbacuju korjenovu kapu i u kojih se rastvara tkivo na vrhu kako bi prolazio kisik. U roda Avicenia postoje otvori u obliku lenticela za prolazak zraka. Uvrste Bruguiera korijenje se naglo savija prema tlu, poprimajući oblik koljena, na kojima također postoje lenticele. Zeljasta biljka Jussiera repens u korijenju ima tkivo za prozračivanje odnosno aerenhim koji je zapravo skladišni dio za zrak. Flora Vegetacija mangrova je podijeljena u posebne zone ovisno od ekoloških čimbenika koji utiču na same biljke. Biljne vrste koje su otpornije na povećanu koncentraciju soli čine vanjski pojas koji je uz more i na udaru plime svaki dan. Manje otporne biljke čine unutrašnji pojas do kojeg voda dopire tek nekoliko puta godišnje kada je plima izuzetno visoka. Te su biljke dominantne u određenim pojasevima i daju tipičan izgled biocenozi. Drveće mangrova Za razliku od drveća u mangrovama čije se korijenje posjeduje tkivo za prozračivanje, rod Rhizophora ima drvo čije korijenje izgleda poput mreže koje izvire iz vode, te uz to ima potpornu ulogu. Sjemenke ove vrste proklija prije nego što napusti roditeljsku biljku i na taj način može da ukorijeniti čim padne na prikladno tlo. Ova prilagođenost biljaka omogućava sjemenkama da ga plima ne odnese daleko na pučinu.

Fauna Isprepletano i gusto korijenje biljaka mangrova predstavlja praktično prirodnu branu koja sakuplja organski materijal koji donose rijeke ili plima. Mulj obogaćen hranjivim tvarima postaje stanište i hrana mnogobrojnim životinjskim vrstama. Rakovi (lat. Crustacea) Populacije od više stotina rakova (Uca) čija je jedna štipaljka dosta veća u odnosu na drugu, hitro se ukopavaju u mulj ukoliko se zateknu u opasnosti ili kada nadolazi plima. Njegova predimenzionirana štipaljka (teška koliko i cijelo njegovo tijelo) nije efikasna u borbi sa drugim mužjacima ili u odbrani od neprijatelja. Njena je uloga u privlačenju ženke, a njome i upozorava druge mužjake. Rak Seylla serrata nastanjuje mangrove na otocima. U slučaju opasnosti ukopava se u mulj. Dobro pliva, a hrani se svim na što naiđe: od strvina do živih bića. Carcinoscorpius rotundicauda je primitivni rak i zapravo „živi fosil“ koji također živi u mangrovi. Slika. Xxc. Rak (Uca annulipes) (lijevo), rak (Seylla serrata) (sredina) i rak Carcinoscorpius rotundicauda (desno) stalni su stanovnici mangrova. Ribe Muljski Skakači (Periophthalmus) su tipične ribe mangrova, ali ujedno i tipični predstavnici faune mangrova uopće. Ta se riba uz pomoć prsnih peraja može popeti na korijenje ili stabla drveća. Veliki dio disanja obavlja preko kože, a posjeduje vid da može vidjeti na kopnu i u vodi. Na suhom prostoru može dugo izdržati, jer ima uski škržni prorez koji sprečava brzo sušenje škrga. Neke vrste roda muljskih skakača nalaze kukce po drveću uz koje se penju, dok se druge vrste hrane algama. Zapravo, svaka vrsta ima poseban režim ishrane i zauzima posebnu ekološku nišu. Naziv su dobile po tome što se mužjaci udvaraju ženkama serijom skokova, a skaču tako da se odupiru o peraje i rep. Zanimljivo je da se tijekom udvaranja parovi ovih riba priljubljuju ustima. Riba strijelac (Toxotes jaculatrix) je dobila naziv po tome što lovi kukce izvan vode tako da ih nepogrešivo pogađa mlazom vode. Nakon toga kukci padaju na površinu vode, gdje postaju lak plijen. Riba strijelac izbacivanje mlaza vode postiže stezanjem škržnih poklopaca, a izbacuje nekoliko mlazova vode u vrlo kratkim razmacima

na daljinu od čak 120 cm. Iznimno se lako i brzo prilagođava slatkoj, bočatnoj ili slanoj vodi. Zbog toga, a i zbog svoje neobične osobine, omiljena je među akvaristima/biolozima širom sveta. Slika xxc. Muljski skakač (Periophthalmus sp.) (lijevo) i riba strijelac (Toxotes jaculatrix) Slika xxc. Riba Periophthalmus barbarus -tijekom udvaranja parovi ovih riba priljubljuju se ustima. Vodozemci Žaba Rana carnivora je vrlo rasprostranjena na staništima mangrova jugoistične Azije. Podnosi morsku vodu, čak ulazi u more gdje se hrani rakovima. Na kopnu se hrani škorpionima i kukcima. Gmazovi Tipični predstavnici gmazova su malajski varan (Varanus salvator), slanovodi krokodil (Crocodylus porosus), kao i više vrsta otrovnih morskih zmija, kao što je prugasta zmija (Laticauda colubrina). Svi se hrane rakovima, mekušcima i ribama, ali i dostupnim otpadnim organskim tvarima. Morski krokodil (još se naziva indomalajski ili estuarijski krokodil) najveći je od svih vrsta, može postići dužinu do 10 metara. Naziv je dobio po tome što često ispliva na pučinu. Mangrovska zmija (Boiga dendrophila) hrani se pticama koje u ova staništa dolijeću kako bi lovile ribe i mekušce. Ova zmija provodi život na drveću, kao i vaglerova jamičarka (Trimeresurus wagleri) koja je dosta „tromija“. Na

glavi između nosnih otvora i očiju ima par jamica koje su osjetljive na toplinu pomoću kojih pronalazi plijen. Slika. xxc. malajski varan (Varanus salvator) (lijevo) i slanovodi krokodil (Crocodylus porosus) s područjem rasprostranjenja. Slika. xxc. prugasta zmija (Laticauda colubrina) (lijevo) i vaglerova jamičarka (Trimeresurus wagleri) (desno). Ptice Neke od vrsta ptica koje se mogu naći u šumama mangrova su orao ribar (Haliaetus leucogaster), indijski marabu (Leptoptilus javanicus), crvenoglavi medojed i mrki pelikan (Pelecanus occidentalis). Mrki pelikan strmoglavim letom lovi ribu u moru. Čuva je u rastegljivoj kesi ispod kljuna, a javlja se i u tropskim područjima Amerike. Šarena roda (Ibis leucocephalus) i vodomar (Halcyon concreta) nastanjuju mangrove jugoistočne Azije. U šumama mangrova živi i vrsta darvinovih zeba (Cactospiza heliobates), to je jedina vrsta koja nastanjuje mangrove. Darvinove zebe su srodne vrste koje su nastale od jedne jedine vrste, a međusobno se razlikuju prema izgledu i snazi kljuna. Te razlike su nastale uslijed naseljavanja različitih ekosustava i prilagođavanja na različite vrste ishrane. Darvinove zebe nastanjuju skupinu Galapagos otoka. U šumama mangrova Nove Gvineje živi dvanaestopera rajska ptica.

Slika xxc. Ptice karakteristične za šume mangrova: mrki pelikan (Pelecanus occidentalis) (lijevo); orao ribar (Haliaetus leucogaster) (sredina) i Šarena roda (Ibis leucocephalus) (desno). Sisavci U šumama mangrova živi nekoliko vrsta životinja iz reda primata. Osim srebrnastog i tamnog langura (Presbytis) koji se hrani lišćem, na ovim staništima se može naći i bradavičavi makaki (Maccaca irus). Ovaj društveni majmun se hrani crvenim rakovima (Pseudosesarma moeshi) koje se kriju u mulju, pa zato dosta vremena provodi tragajući za njima po muljevitom tlu. Javanski makaki (Maccaca fascicularis) većinu vremena provodi tražeći rakove i školjke po plićacima. Znatno rjeđa vrsta majmuna je majmun nosonja (Nasalis larvatus). Kod mužjaka je nosni izraštaj dugačak i visi preko usta i brade. Miroljubiva je i društvena životinja. Za razliku od većine drugih vrsta majmuna on je vješt plivač i često će skočiti u vodu kako bi je preplivao, ali tada postaje lak plijen jedinog mu predatora — krokodila. Evolucija Fauna šuma mangrova jugoistočne Azije ukazuje na to da je evolucija živog svijeta išla od prelaska života iz mora na kopno. Naime, životinjske vrste koje naseljavaju ova područja su amfibijskog karaktera, dakle prilagođene su na kopneni način života, ali im se ciklus razvića odvija u vodi. Također, u obližnjim tropskim šumama se sreću vrste iz šuma mangrova, a čije osnovne značajke upućuju na njihovo porijeklo iz mora, a ne sa kopna. Poznate močvarne šume iz perioda Karbona od prije 350 milijuna godina, a čija je jedna od glavnih obilježja prilagođavanje amfibijskih životinja na kopneni način života. Zagađenje i zaštita

Kao i u slučaju mnogih drugih biljnih i životinjskih vrsta, antropogeni utjecaj na šume mangrova također je vrlo značajan i u većini slučajeva iznimno negativan. Veliki dio površina koje s obrastale šume mangrova u Africi, posebno Nigeriji, uništeno je u posljednjih 50 godina uslijed izlijevanja nafte u more, uglavnom zbog havarija tankera. Ovo je svakako imalo utjecaja i na kvalitetu vode u šumama mangrova.
3. Morski ekosustavi
Morski ekosustavi spadaju među najveće vodene ekosustave na Zemljia. Oni uključuju oceane, slane močvare, estuarije i lagune, mangrove, koraljne grebene, duboko more i morsko dno. Za razliku od slatkovodnih ekosustava imaju znatno veći sadržaj mineralnih soli. Morske vode prekrivaju dvije trećine površine Zemlje. Sva se ta područja smatraju ekosustavima, jer funkcioniraju kao zatvoren sustav koji uključuje proizvođače, (zelene biljke), potrošače (životinjski svijet) i razlagače saprofitski i parazitski organizmi). Usporedi hranidbeni lanac. Morski ekosustavi su vrlo važni za cjelokupno morsko i kopneno okruženje. Općenito se smatra da obalna staništa uključujući delte rijeka, estuarijima, slane močvarama, šume mangrova spadaju među najproduktivnija područja na planeti, te se računa da proizvode oko 1/3 morske primarne biološke produktivnosti. Uz to, drugi morski ekosustavi, kao što su koraljni grebeni, osiguravaju hranu i sklonište mnogim morskim organizmima i predstavljaju najviši stupanj biološke raznolikosti u svijetu. Morski ekosustavi obično imaju veliku biološku raznolikost i za njih se smatra da imaju dobru otpornost protiv invazivnih vrsta.
3.1. Morske struje
Morske struje koje susrećemo u svjetskim morima i oceanima su manje-više međusobno povezane. To su uglavnom horizontalno gibanje morske odnosno oceanske vode, te su uz valove i mijene jedno od triju osnovnih gibanja morske vode. Kreću se u određenom smjeru,a mogu biti: (i) stalne, (ii) periodične ili (iii) povremene pojave. Morske struje su tople ili hladne "rijeke" vode u oceanima. Povremene i periodične struje utječu na manja, uža litoralna područja, dok stalne morske struje imaju planetarni utjecaj. Morske struje uzrokuju sile i čimbenici koji djeluju na vodu, kao na primjer: rotacija Zemlje, prevladavajući vjetrovi koji pušu na površinu i usmjeravaju tok vode (stalni ili sezonski vjetrovi), razlike u temperaturi vode, razlika u salinitetu, privlačna sila Mjeseca i riječni tokovi. Na smjer i snagu morskih struja utječu i oblik morskog dna i obale, promjene u gustoći vode kao i druge struje.

Morske struje čiji su uzrok vjetrovi, rijetko kad zahvate vodu dublje od 200 metara, dok su struje koje su posljedica Coriolisovog učinka duboke i do 1000 metara. Javljaju se uz zapadne obale kontinenata i vrlo su brze i snažne. Najbrža među njima je Agulhaška struja u Indijskom oceanu i teče brzinom od 5 čvorova. Oceanske struje mogu "putovati" više tisuća kilometara. Imaju veliko djelovanje na klimu kontinenata, a naročito na područja uz obale oceana. Vjerojatno najizraženiji primjer je Golfska struja koja čini klimu sjeverozapadne Europe umjerenijom od drugih područja na jednakim zemljopisnim širinama. Drugi primjer su Havaji, gdje je klima suptropska a ne tropska zbog hladne tihooceanske odnosno Kalifornijske struje. Slika xxc. Karta Svijeta s oceanskim strujama Odjel za fizičku oceanografiju NASA-e je zaključio da nema usporavanja protoka velike oceanske struje koja donosi toplu vodu iz tropskih dijelova Pacifika na sjever Atlantika (slika xcc). Te struje funkcioniraju kao "pokretna traka" pokrenute zbog razlike u slanosti oceana. Tople oceanske struje stvaraju pogodne klimatske prilike za veliki dio Europe, što ne bi bio slučaj bez stalnog priljeva tople morske vode. Najnovijim proučavanjima morskih struja utvrđeno je da te morske struje ne usporavaju i vjerojatno nisu usporene sve od ranih 90-ih godina. To je dobra vijest za tople struje o kojima ovise regije poput Velike Britanije. Ova najnovija otkrića, koja se oslanjaju na podatke iz oceanskih senzora (postavljenih u oceanima) i sa satelita, dokazuju manju vjerojatnost da dolazi do zagrijavanja klime na Zemlji nego podupiru tezu da se Zemlja kreće prema ledenom dobu.

Slika. Xxc. Smjer kretanja toplih i hladnih oceanskih struja. Tablica 1.Oceanske struje s velikim značenjem, naziv oceana (more), temperatura Važnije oceanske (morske) struje
Ocean (more) Temperatura (t =topla/h= hladna)
Agulhaška struja Indijski ocean (Obala jugoistočne Afrike)
t
Antarktička cirkumpolarna struja u Atlantiku, u Pacifiku i u Indijskom oceanu)
h
Antilska struja Atlantik t Ekvatorska protustruja u pacifičkom i indijskom oceanu t Azorska struja Atlantik (Azorski prag) t Benguelska struja Atlantik (obala jugozapadne
Afrike) h
Brazilska struja Atlantik (Istočna obala Brazila) t Ekvatorska struja Pacifik t Kineska obalna struja Pacifik (Istočno kinesko more, Južno
kinesko more) t
Falklandska struja Atlantik (Falklandski otoci) h Floridska struja Atlantik (Meksički zaljev,
Floridski prolaz) t
Golfska struja Atlantik (Američka istočna obala, sjeverni Atlantik)
t
Gvinejska struja Atlantik (Gvinejski zaljev) t Humboldtova struja Pacifik (Zapadna obala Južne
Amerike) h
Irmingerova struja Atlantik (Europsko Sjeverno more)
t
Kalifornijska struja Pacifik (Američka zapadna obala) h Kanarska struja Atlantik (Obala zapadne Europe i
zapadne Afrike) h
Kap Hoornska struja Pacifik, Atlantik (Južni vrh Južne Amerike)
h
Karipska struja Atlantik (Karipsko more, Meksički zaljev)
t
Kurošio struja Pacifik (Zapadni Pacifik, Japansko more)
t
Labradorska struja Atlantik (Atlantik, Newfundlandski bazen)
h
Mozambička struja Indijski ocean (Mozambički prolaz)
t
Sjeverna ekvatorska struja Atlantik, Pacifik i Indijski ocean t Sjevernoatlantska struja Atlantik t Sjevernopacifička struja Pacifik t Norveška struja Atlantik (Sjeverno more) t Istočnoaustralska struja Pacifik (Tasmanovo more) t Istočna i Zapadna grenlandska
Atlantik (Atlantik, europsko Sjeverno more)
h

10. ZOOGEOGRAFSKE OBLASTI
Životinjske vrste i njihove populacije nisu ravnomerno raspoređene naZemlji. Na kojim se prostorima i u kojim uvjetimanalaze pojedine skupineživotinja danas, koliki su im i kakvi areali, zavisi od područja njihovognastanka i pravaca migracija, geotektonskih pomicana, utjecaja svih ekološkihfaktora u prošlosti i sadašnjosti, od evolutivnih procesa i od bioloških iekoloških odlika samih životinjskih skupina. Utjecaji svih povijesnih isuvremenih vanjskih i unutrašnjih faktora uvjeti i su da danas nekeživotinjske skupine žive u vodi, neke na kopnu, da pojedine naseljavaju staništa odjednog-dva hektara, a druge se javljaju na dva ili više kontinenata, da nekeopstaju isključivo u tropskim, a neke isključivo u polarnim oblastima.Na osnovu globalnog rasprostranjenost pojedinih životinjskih skupina,cjelokupnu kopnenu površinu Zemlje, Lopatin (1995) dijeli na četirizoogeografske oblasti u okviru kojih se izdvajaju pojedine specifičnezoogeografske oblasti:I Zoogeografsko oblast Arktogeja (Holarktik) obuhvaća Euro sibirsko iPalearktičko podoblast.II Zoogeografsko oblast Neogeja obuhvaća Neotropsku i Karipskuzoogeografsku podoblast.III Zoogeografska podoblast Paleogeja obuhvaća Etiopsku, Indo-Malajsku iMadagaskarsku zoogeografsku podoblast.IV Zoogeografska oblast Notogeja obuhvaća Novozelandsku, Australijskui Polinezijsku zoogeografsku podoblast.
I. ZOOGEOGRAFSKA OBLAST ARKTOGEJA (HOLARKTIK) Arktogeja ‒Palearktik (1. Sredozemna podoblast, 2. Centralnoazijska podoblast, 3. Euro sibirska podoblast) Nearktik (4. Kanadska podoblast, 5. Sonarska podoblast)
Zoogeografska oblast Arktogeja ili Holarktik, obuhvaća kopno sjevernehemisfere ‒ Evropu sa Grenlandom, Aziju bez njenog jugoistočnog dijela, SjevernuAfriku, gotovo cijelu Sjevernu Ameriku osim njenog krajnje južnog dijela. Ova velikateritorija dijeli se na dva zoogeografska podoblasti ‒ Palearktik i Nearktik.U granicama Palearktika nalaze se tri zoogeografske podoblasti ‒ Sredozemna,Centralnoazijska i Euro sibirska, dok se u granicama Nearktika nalaze dvijezoogeografske podoblasti ‒ Kanadska i Sonarska.Krajnji sjever ovog carstva nalazi se pod vječitim snijegom i ledom, doknjegovi južni dijelovi bez jasnih granica postupno prelaze u tropske oblasti.Naime, Palearktik se prema jugu graniči sa Paleogejom preko širokog pustinjsko-polu pustinjskog prelaznog pojasa Sjeverne Afrike i Arabije, preko Arabijskogmora, a dalje, preko visokog lanca Himalaja i jugoistočne Kine, do Istočno-kineskog mora. U Sjevernoj Americi, prelazna zona prema južnijem Neotropskomcarstvu obuhvata sjeverne dijelove Meksika, Kalifornije i Floride.Širina ovog područja koja podrazumijeva i smjenu određenih klimatskihpojaseva, uvjetovala je pojavu različitih tipove vegetacije, počevši od tundre nakrajnjem sjeveru, pa preko šumo tundre, tajge, listopadnih i vječno zelenih šuma,šumo stepa i stepa, do pustinja na jugu. Veoma složen i razdijeljen reljef, kao ivisinsko zoniranje vegetacije duž planinskih vijenaca, dodatno utiču na složen

biomski spektar, pa samim tim i na raspored životinjskih vrsta i razgraničenjepojedinih faunističkih oblasti. Specifično za Holarktik je da se izdvajaju samo dva reda slatkovodnih riba, prisutna isključivo na sjevernoj hemisferi Prvi je red grgeča (Percidae) rasprostranjen na čitavom Holarktiku. Drugi je red sunčanica (Centrarchidae) prirodno nearktičkog rasprostranjenosti, koji je naknadno prenijet i na sekundarna staništa širim Euroazije. Vodozemci i gmazovi su u odnosu na ostala carstva slabije zastupljeni, bez endemičnih redova koji bi bili specifični isključivo zaHolarktik. Među endemičnim porodicama sisara Holarktika izdvajaju sezečevi zviždači (Ochotonidae), dabrovi (Castoridae) i mišoliki skočimiši(Zapodidae). Također, samo u Nearktiku žive endemične porodice planinskog dabra(Aplodontidae) i viloroge antilope (Antilocapridae), dok samo u Palearktiku živeporodice miševa skakača (Dipodidae), panda (Ailuridae), hrčaka (Cricetinae), slijepihkučića (Spalacinae). Među pticama postoji samo jedan endemičan red, Gaviidae,koji obuhvata takozvane sjeverne gnjurce.
1. Mediteranska podoblast obuhvata mediteransko i submediteransko područjeEvrope, Sjeverne Afrike i prednje Azije, Kavkaz, Krim i južni dio Srednje Azije.Od vodozemaca, kao specifične vrste ove oblasti javljaju se čovječija ribica(Proteus anguinus), crni daždevnjak (Salamandra atra), šareni daždevnjak(Salamandra salamandra). Pored ovih, prisutni su i alpski mrmoljak (Triturusalpestris), vatrene žabe (Bombina sp.), žabe kreketuše (Hyla spp.), prave žabe(Rana spp.), krastave žabe (Bufo sp.), itd. Od gmazova, u ovoj oblasti se kaospecifičan javlja mediteranski kameneleon (Chamaeleo chamaeleon), a prisutnisu i brojni gušteri iz rodova Podarcis (zidni gušter), Lacerta (zelembaći,livadski gušteri), Ablepharus (kratkonogi gušteri), Anguis (slepić), Ophisaurus(blavor). Također, karakteristične su zmije iz porodica Elapidae (crna egipatskakobra, egipatska kobra), Boidae (različite pješčane boe), Colubridae (bjelouške,ribarice, smukovi, zmajur), Viperidae (šarka, poskok, planinski šargan), kao išumska i grčka kornjača (Testudo hoermanni, Testudo graeca). Međukarakteristične ptice ovog područja spadaju bjela kanja (Neophron percnopterus),bjeloglavi sup (Gyps fulvus), jarebica kamenjarka, (Alectoris graeca), pustinjskakreja (Podoces panderi), galeb kliktavac (Larus michahellis), vivak (Vanellusspinosus), a među karakteristične sisare iberijski ris (Linx pardina), pustinjskiris (Caracal caracal), divokoza (Rupicapra rupicapra), divlja mačka (Felis silvestris),srna (Capreolus capreolus), muflon (Ovis ammon), dvogrba kamila (Camelus ferus),jednogrba kamila (C. dromedarius), bezrepi makaki (Macaca sylvanus), itd.
2. Centralnoazijska podoblast obuhvaća centralnu i istočnu Aziju zajednosa Japanom. Specifične vrste ove oblasti su džinovski daždevnjaci (Andriasjaponicas i Andrias davidianus), kineski aligator (Alligator sinensis), mandžurijskifazan (Crossoptilum manchuricum), stepska lisica (Vulpes corsac), mali panda(Ailurus fulgens), veliki panda (Ailuropoda melanoleuca), tibetanska antilopa(Pantholops hodgsoni), tibetanska gazela (Procapra picticaudata), mongolijska gazela(Procapra gutturosa), crveni jelen (Cervus elaphus), azijski crveni pas (Cuonalpinus), azijski magarac (Equus hemionus), azijski divlji konj (Equus przewalskii),mošusno goveče (Moschus moschiferus), Davidov jelen (Elaphurus davidiensis),snježni japanski majmun (Macaca fuscata), tibetski majmun (Macaca thibetana),brojni specifični skočimiši, itd

3. Eurosibirska pdoblast obuhvata gotovo čitavu Evropu, Sibir i sjeverniKazahstan. Za ovu oblast karakteristične vrste su džinovski sibirski daždevnjak(Hynobius keyserlingi), gušter žaboglava agama (Phrynocephalus), sjeverni gnjurac(Gavia arctica), polarna jarebica (Lagopus lagopus), alpska koka (Lagopus mutus),šumska sova (Strix aluco), istočno europski jež (Erinaceus concolor), patuljastarovčica (Suncus etruscus), snježni zec (Lepus timidus), vuk (Canis lupus), polarnalisica (Alopex lagopus), obična lisica (Vulpes vulpes), stepska lisica (Vulpescorsac), kunopas (Nyctereutes procyonides), mrki medvijed (Ursus arctos), evroazijskijazavac (Meles meles), evroazijski ris (Lynx lynx), euroazijska divlja svinja (Susscrofa), jelen lopatar (Dama dama), sibirska snježna ovca (Ovis nivicola), itd.
4. Kanadska podoblast obuhvata zonu tajge Sjeverne Amerike i Grenland. Zaovu oblast karakteristične vrste su snježna sova (Bubo scandiacus), kanadskaguska (Branta canadensis), kanadski dabar (Castor canadensis), polarni medvjed(Ursus maritimus), crni medvjed (Ursus americanus), grizli (Ursus arctos horribilis)sjevernoameričko bodljikavo prase (Erethizon dorsatum), sjeverni ris (Lynxcanadensis), polarna lisica (Alopex lagopus), karibu (Rangifer tarandus), los(Alces alces), žderavac (Gulo gulo), mrki medvijed, itd.
5. Sonorska podoblast obuhvata središnji i južni dio Sjeverne Amerike. Zaovaj dio Nearktika karakteristične životinjske vrste su repati vodozemac Sirenlacertina koji može dostići dužinu i do 1 m, meksički daždevnjak aksolotl(Ambystoma tigrinum), američka čovječja ribica (Necturus maculosus), mala repatažaba (Ascaphus montanus), iguana poznata kao rogata žaba (Phrynosoma spp.)kalifornijski kondor (Gymnogyps californianus), prerijski tetrijeb (Tympanuchuscupido), američki kunići (Sylvilagus spp.), obični oposum (Didijphis virginiana),deveto pojasni oklopnik (Dasypus novemcinctus), planinski dabar (Aplodontia rufa),leteća vjeverica (Glaucomys volans), prerijsko kuče (Cynomys ludovicianus), kojot(Canis latrans), crni medvjed (Ursus americanus), puma (Puma concolor), američkibizon (Bison bison), bjelorepi jelen (Odocoileus virginianus) kengur pacovi(Dipodomys spp.), itd.
II ZOOGEOGRAFSKO oblast NEOGEJAOva oblast obuhvaća Neotropsku zoogeografsku oblast kojoj pripadajuJužna Amerika, dio Centralne Amerike do južnih oboda meksičke visoravni, kaoi Karibska otoci. U reljefu Južne Amerike izdvajaju se visoki vijenac Anda na zapadu, staregromadne planine sa visoravnima na istočnim i u centralnim dijelovimakontinenta, kao i prostrane ravnice. Andi predstavljaju visoki planinski vijenac koji se proteže u dužini od oko 7.000 km duž obale Tihog oceana od Panamskog kanala i Karipskog mora na sjeveru, do rta Horn na krajnjem jugu kontinenta. Širina Anda iznosi do 900 km,dok je najviši vrh Akonkagva koji doseže do 6.958 m nadmorske visine.Stare gromadne planina mogu se podijeliti u dvije skupine – gvajansku ibrazilsku. Svi imaju izgled visoravni sa kojih se uzdižu planinski vrhovido visine od 2.800 m. Između njih se nalaze ravnice Amazonije, visoko travnatesavane ljanosi na sjeveru u slivu rijeke Orinoko, travnate stepe pampase najugoistoku u slivu rijeke La Plata, ravnica Gran Čako i stepe Patagonije na jugu.

Južna Amerika se većim dijelom nalazi na južnoj hemisferi, između Tihog iAtlantskog oceana. Pošto je presijeca ekvator, njen najveći dio nalazi se podutjecajem ekvatorijalne, tropske i suptropske klime. Suptropska klima upojedinim kontinentalnim dijelovima poprima elemente aridne klime tako dašumsku vegetaciju smjenjuju travnate oblasti. Na zapadu kontinenta izdvaja sepojas sredozemne klime prema sjeveru, kao i pojas umjerene oceanske klime premajugu. Na velikim visinama vlada oštra planinska klima.Fauna Neotropske oblasti je veoma bogata, sa značajnim učešćemreliktnih i endemičnih vrsta. U pojedinim elementima pokazuje sličnost safaunom istočne Gondvane, prije svega Afrike, od koje se odvojila prije 130 milionagodina. Također, posjeduje niz zajedničkih faunističkih elemenata koji je povezujusa Sjevernom Amerikom (naročito Centralna Amerika), zahvaljujući Panamskommoreuzu koji je pre 15.000 miliona godina uspostavljen između Sjeverne i JužneAmerike. U raznovrsnoj fauni vodozemaca, kao specifične skupine treba spomenutibezplućne daždevnjake iz familije Plethodontidae, surinamsku pipu (Pipaamericana), otrovne žabe iz familije Dendrobatidae među kojima je najpoznatijaDarvinova žaba (Rhinoderma darwinii), kao i crvenooke žabe koje žive na drveću(Agalychnis callidryas) u šumama Centralne Amerike. Od gmizavaca, kao znakraspoznavanja ove oblasti, izdvajaju se galapagoska kornjača (Testudoelephantopus), zelena iguana (Iguana iguana), morska iguana (Amblyrhynchuscristatus), kubanska iguana (Cyclura cornuta), gušter bazilisk (Basiliscus basiliscus),zmijski car (Boa constrictor), anakonda (Eunectes murinus), mliječna meksička zmija(Lampropeltis triangulum), koralne zmije (Micrurus sp.), zvečarke (Crotalus sp.),orinoški krokodil (Crocodylus intermedius), meksički krokodil (Crocodylusmoreletii), kolumbijski kajman (Caiman sclerops), crni kajman (Melanosuchus niger) ibrojne druge vrste specifične za ove prostore.U bogatoj fauni ptica, kao vrste jedinstvene za ovo područje, izdvajaju senandu (Rhea americana), Darvinov nandu (Pterocnemia pennata), kraljevskilešinar (Sarcoramphus papa), andski kondor (Vultur gryphus), veliki karuso (Craxrubra), papagaj crveni makao (Ara macao), papagaj plavi makao (Ara ararauna),zeleni papagaj (Amazona brasiliensis), motmot (Momotus momota), todi (Todussubulatus), brojni kolibri koji pripadaju porodici Trochilidae, također brojnitukani koji pripadaju porodici Ramphastidae, itd.Osnovno obilježje faune sisavaca je prisustvo krezubica koje pripadaju reduPilosa. Za predstavnike ovog reda je svojstveno nedostatak zuba kao što je to slučajkod mravojeda, ili pojava samo kutnjaka kao što je to slučaj kod ljenjivaca ioklopnika. Mravojedi (Myrmecophagidae) su predstavljeni sa tri roda: mali dvoprstimravojedi (Mymecophaga mexicana), četvoroprsti mravojedi (Mymecophagatetradactyla) i veliki troprsti mravojedi (Mymecophaga tridactyla). Za mravojedeje svojstveno da se hrane mravima i termitima. Mali dvoprsti i srednjičetvoroprsti mravojedi hrane mravima koji žive na drveću, tako da pretežnonaseljavaju šumske oblasti. Veliki troprsti mravojedi koji mogu premašitidužinu od 2 m, uglavnom se hrane mravima na tlu, tako da najčešće naseljavajuotvorene prostore. Ishrana mravima i termitima kod ove skupine životinja dovelaje do niza prilagođenosti kao što su dugačka njuška, odsustvo zuba, dugačak iljepljiv jezik, duge i oštre kandže za kopanje zemlje.Lenjivci (Folivora) predstavljaju jednu od najstarijih skupina sisavaca. Krećuse veoma sporo, a nastanjuju isključivo krošnje drveća tropskih šuma SrednjeAmerike i Amazonije. Obično vise na granama za koje se pričvršćuju dugim,nazubljenim kandžama. Na tlu ne mogu stajati uspravno, nego se povlače uz pomoć

kandži. Hrane se uglavnom lišćem, rjeđe plodovima i kukcima.Oklopnici (Cingulata) su mali placentalni sisavci predstavljeni sa oko 20vrsta veličine 15 do 75 cm. Za ove životinje su svojstvene hrskavičave ilikoštane pločice spojene u vanjski oklop. Na površini oklopa nalaze serožnati zubići koji predstavljaju dodatnu zaštitu tijela. Žive u stepskim ipustinjskim oblastima, a najčešće se hrane korijenjem i drugim dijelovima biljaka,kukcima i lešinama. Pored južne Amerike, naseljavaju i dijelove sjeverneAmerike, naročito područja Teksasa, Nebraske, Južne Karoline i Floride.U neotropskoj oblast prisutan je i veliki broj torbara (Marsupilia).Najpoznaniji među njima su oposumi (Didijphis) koji predstavljaju vrstu torbarskihpacova. Naseljavaju sjeverne dijelove Južne Amerike, Centralnu Ameriku i južnedijelove Sjeverne Amerike. Oposumi imaju tijelo slično mačjem i dostižu težinu odpreko 6 kg. Aktivni su u potrazi za hranom, mada mogu i satima da visi okačen zagrane drveća zahvaljujući dugom i jakom repu. Među oposumima ima i vrstavezanih za vodu. Posebno mjesto među sisavcima zauzimaju brojni širokonosni majmuni(Platyrrhini): marmozet (Callithrix jacchus), zlatoglavi marmozet (Leontopithecusrosalia), Geldijev marmozet (Callimico goeldii), carski tamarin (Saguinus imperator),pinča tamarin (Saguinus oedipus), belogrli kapucin (Cebus capucinus), durukuli(Aotes trivirgatus), titi ogrličar (Callicebus torquatus), crveni uakari (Cacajaorubicundus), saki kaluđer (Pithecia monachus), obični saimiri (Saimiri sciureusi),crveni drekavac (Alouatta seniculus), crni drekavac (Alouatta caraya), runastimajmun (Lagothrix lagotricha), crni pauk majmun (Ateles paniscus), crnoruki paukmajmun (Ateles geoffroyi).Od ostalih sisavaca, za ovu podoblast naročito su karakteristični tapiri(Sylvilagus brasiliensis) i paka (Agouti paca) iz porodice glodara, kolumbijskidivlji pas (Speothos venaticus), grivasti vuk (Chrysocyon brachyurus), andskimedvjed (Tremarctos ornatus), jaguar (Panthera onca), puma (Puma concolor), koati(Nasua nasua), kinkadžu (Potos flavus), kuna tajra (Tayra barbara senilis), džinovskavidra (Pteroneura brasiliensis), obična lama (Lama glama), lama alpaka (Lamapacos), lama gvanko (Lama guanicoe), panamski jelen (Ozotoceros bezoarticus),belorepi jelen (Odocoileus virginianus), mali pudu jelen (Pudu pudu).
III ZOOGEOGRAFSKA OBLAST PALEOGEJAOva zoogeografska oblast obuhvaća etiopsku, madagaskarsku i indo-malajsku podoblast.
1. Etiopska zoogeografska oblastEtiopska zoogeografska oblast obuhvata Afriku južno od Sahare, južnidio Arabije i otok Sokotru u Arabijskom moru, kao i Zelenortska, Kanarska,Azorska otoci i Madeira u Atlantskom oceanu.Istočna Afrika pretežno je planinska vulkanska oblast sa Kilimandžaromnajvišom masivom Afrike (Uhur, 5.895 m n.v.). U rasjednim dolinama oko objeplaninske oblasti nalaze se velika jezera ‒ Tanganjika, Viktorijino jezero,Albertovo jezero, itd. Južna Afrika uglavnom je visoravan okružena primorskimplaninama od kojih je najveća Kvatlamba (3660 m n.v.). U njenom centralnom dijeluleži polu pustinjsko-pustinjska oblast Kalahari, a uz atlantsku obalu pustinjaNamib. Centralna Afrika uglavnom obuhvaća blago zatalasan sliv rijeke Kongo saProstorima prekrivenim tropskim kišnim šumama. Čitavu etiopskuzoogeografsku podoblast odlikuje bogatstvo ekosustava od kojih većina pripada

tropskim šumama, savanskim šumama i savanama.U fauni vodozemaca ove oblasti najbrojnije su žabe među kojima seizdvaja žaba Golijat (Conraua goliath) iz Kameruna i Ekvatorijalne Gvineje kojamože dostići težinu i do 3 kg, afrička žaba bik (Pyxicephalus adspersus) izjužnih dijelova Afrike koja dostiže do 2 kg, kao i brojne sićušne žabe koje živena drveću. Fauna gmazovas je također bogata, a kao tipične vrste mogu se navestiiguana Agama agama, kameleoni (Chamaeleonidae), brojni gušteri (Varanusniloticus, Lacerta echinata), kao i zmije među kojima su najpoznatije zmije jajožderi(Dasypeltis sp.), kobre (Naja sp.), mambe (Dendroaspis sp.), nilski krokodili(Crocodylus niloticus), patuljasti krokodili (Osteolaemus tetraspis).Fauna sisavaca etiopske podoblasti pokazuje određene sličnost sa faunomsisavaca madagaskarske i indomalajske oblasti. Sa faunom sisavaca Madagaskarapovezuje je porodica čekinjastih ježeva (Tenrecidae), dok je sa faunomindomalajske oblasti povezuju ljuskari, slonovi, nosorozi, damani i uskonosnimajmuni.Karakteristični predstavnici ove oblasti su sitne zlatne krtice izporodice Chrysochloridae kod kojih je uslijed podzemnog načina života došlo doredukcije očiju, kao i rovčice slične vidrama iz porodice Soricidae koje vodepolu vodeni način života hraneći se ribama. Prisutna je i endemična porodicaslonastih rovčica iz porodice Macroscelidae koje pokazuju veliku konvergentnusličnost sa skočimišima i imaju surlasto izduženu njušku.LJuskari (Pholidota) su još jedna interesantna autoktona skupina etiopskeoblasti predstavljena samo jednim rodom (Manis) sa sedam vrsta. LJuskari sukarakteristični po tome što im je tijelo prekriveno crijepasto raspoređenimkožnim ljuskama. Veličine su 30 do 90 cm. Nemaju zube, ali zato imaju dug,crvoliki, ljepljiv jezik dug do 25 cm. On im služi za hvatanje insekata, naročitomrava, što predstavlja konvergentnu sličnost sa mravojedima Južne Amerike.Među glodarima, po svojoj specifičnosti izdvajaju se afričke tekunice(Xerus sp.), afričke leteće vjeverice (Anomalurus sp.) koje imaju kožne opneizmeđu prednjih i zadnjih ekskremiteta, kao i kapski dugonog ili zec skakač(Pedetes capensis) koji liči na skočimiša zbog razvijenih zadnjih ekstremitetakoji mu omogućavaju kretanje u dugim skokovima.Zvijeri predstavljaju brojnu skupinu ove podoblasti. Kao znak raspoznavanjaAfrike izdvajaju se afrički divlji pas (Lycaon pictus) koji je vezan za prostraneoblasti savana, dugouha lisica (Otocyon megalotis), merkat (Suricatta suricatta) kojiživi u pustinji Kalahari, hijene (Hyaena), afrička cibetka (Civettictis civetta),leopard (Panthera pardus), gepard (Acinonyx jubatus), lav (Panthera leo), pustinjskiris (Caracal caracal), servak (Leptailurus serval), afrička zlatna mačka (Profelisaurata), itd.Endemična vrsta etiopske oblasti je i afrički slon (Loxodonta africana)koji predstavlja najkrupniju kopnenu vrstu sisavaca. Krupniji je od indijskog slona,od koga ima znatno veće kljove koje predstavljaju metamorfozirane sjekutiće, iuši koje mu između ostalog, služe za hlađenje. NJima su srodni damani(Procaviidae) koji se javljaju i u jugozapadnoj Aziji. Predstavljaju veoma staruskupinu čija je većina predstavnika izumrla tijekom miocena. Suvremeni damani suveličine zeca. Hrane se biljkama, nemaju očnjake, ali zato imaju male kljove.Od papkara (Artiodactyla), za ovu oblast je specifična porodica nilskihkonja (Hippopotamidae) koja pripada papkarima nepreživačima. Ova porodicapredstavljena je danas samo sa dvije vrste. To su patuljasti nilski konj (Choeropsisliberiensis) i nilski konj (Hippopotamus amphibius), krupna biljojedna životinja dugado 4,5 m i teške do 3.200 kg. Tijekom dana žive oko vode i u vodi, a tijekom noćiodlaze na okolne livade da pasu travu. Također, u nepreživače spadaju i

predstavnici porodice svinja (Suidae): riječna svinja (Potamochoerus porcus),šumska svinja (Hylochoerus meinertzhageni) i bradavičasta svinja (Phacochoerusaethiopicus). Specifične vrste papkara preživača ove oblasti su patuljasti jeleni izporodice Traguloidae, šupljorogi i žirafe. Patuljasti jeleni poznati i kaovodeni jelenčić ili vodeni ševroteni (Hyemoschus aquaticus), žive u šumama,močvarnim područjima i oko vodenih bazena. Veličine su zečeva tako da sunajmanji od svih papkara. Nemaju rogove, a mužjaci imaju razvijene gornje očnjakekoji štrče izvan usana.Šupljorogi objedinjuju bivole i brojne vrste antilopa koje pripadajuporodici Bovidae. Na afričkom tlu antilope su predstavljene sa 30 rodova iblizu 90 vrsta različitog ponašanja, izgleda i veličine koja varira odveličine omanjeg zeca do veličine krupnijeg konja. Tipični predstavnici oveskupine papkara su sabljaste antilope (Hippotragus niger), patuljaste antilope(Neotragus), oriks gazele (Oryx gazella), planinski skakači (Oreotragus sp.), gazelesavanski skakači (Antidorcas sp.), impale (Aepyceros melampus), adaks antilopa(Addax nasomaculatus), Tomsonove gazele ((Eudorcas thomsonii), gnu (Connochaetestaurinus), kane (Taurotragus derbianus), kudu (Tragelaphus strepsiceros), bivoli(Synceros caffer), itd. Porodica žirafa (Giraffidae) obuhvaća prave žirafe (Giraffacamelopardalis) koje predstavljaju karakteristične životinje savana.Ovoj porodici pripadaju i okapi (Okapia johnstoni) koji naseljavaju tropske šume.I za jedne i za druge predstavnike ove porodice, karakteristični su kratkikoštani rogovi pokriveni dlakavom kožom i dva papka na nogama, bez bočnihprstiju. Žirafu dodatno odlikuje veoma dug vrat. Prostrane oblasti savana uvjetovale su široko rasprostranjenost kopitara (Perissodactyla) među kojima se izdvajaju nosorozi i zebre. Nosorozi su krupni, debelokožni biljojedi koji obično naseljavaju savanske oblasti. Odlikuje ihjedan ili dva roga na njušci. Od ukupno pet vrsta nosoroga, u ovoj oblasti senalaze dvije vrste, crni nosorog (Diceros bicornis) i bijeli nosorog (Ceratotheriumsimum). Predstavnici obje ove vrste su dugi i preko 4 m, imaju po dva roga od kojihjedan može da dostigne dužinu do 1,3 m. Zebre pripadaju porodici konja(Equidae). Pored obične zebre (Equus burchelli), u ovoj podoblasti prisutne su još dvijevrste zebre: Grevijeva (Equus grevyi) i planinska (Equus zebra).Za veći dio etiopske oblasti karakteristične su i cijevozupke, čiji jetipičan predstavnik takozvani afrički mravojed (Orycteropus afer). Odlikuje secjevasto građenim zadnjim zubima. Dostižu dužinu do 1,3 m, ima dug rep i dugunjušku, dug i ljepljiv jezik kojim hvata insekte. Hrani se mravima i termitimaraskopavajući mravinjake oštrim kandžama.Polumajmuni (Prosimii) etiopske podoblasti pripadaju porodici lori majmuna.među kojima su najpoznatiji poto (Perodicticus potto), galago (Galago galago) ikalabarski poto (Arctocebus calabarensis). To su noćne životinje veličine do 40cm. Žive na krošnjama drveća i hrane se lišćem, plodovima, kukcima imanjim pticama. Majmini (Anthropoidea) Afrike pripadaju takozvanim uskonosnimmajmunima (Catarrhini). Tipični predstavnici su zeleni zamorac (Cercopithecusaethiops), diana zamorac (Cercopithecus diana), obični pavijan (Papiocynocephalus), grivasti babun (Papio hamadryas), mandril (Mandrillus sphinx),gvereza (Colobus abyssinicus), itd. U ovu skupinu spadaju i ljudomajmuni (Hominoidea)

gorila (Gorilla gorilla), čimpanza (Pan troglodytes) i patuljasti šimpanza (Pan paniscus).Od ptica ovog područja izdvajaju se afrički nojevi, batoglava čaplja(Scopus umbretta), čaplja cipelarka (Balaeniceps rex), svijeti ibis (Threskiornisaetiopica), afrička kašikara (Platalea alba), sekretar (Sagittarius serpentarius),afrički krunasti orao(Stephanoaetus coronatus), krunasti ždral (Balearicapavonia) pauni (Afropavo kongensis) žako (Psittacus erithacus), ljubavne ptice(Agapornis sp.), ljubičasti turako (Musophaga violacea), ptica nosorog (Bucorvusspp), kao i brojne vrste lasta, ševa, pliski, drozdova, sunčanica, strnadica, zebai drugih skupina ptica.
2. Madagaskarska zoogeografska oblastMadagaskarska zoogeografska oblast obuhvaća otok Madagaskar i nizmalih otoci (Sejšelska, Komorska, Mauricijus, Rodrigez...) u Indijskom oceanu, ublizini istočne obale Afrike. Istočna i zapadna obala Madagaskarapredstavljene su ravnicama koje vode prema centralnim planinama. Na sjeveruotoka dominira masiv Tsaratanana (2.876 m n.v.) i vulkanske planine, a najugozapadu se nalazi rajon visoravni i pustinja. Istočna strana otoka obraslaje tropskim kišnim šumama, dok su zapadna i južna strana otoka prekrivenetropskim suhim šumama, bodljikavim šumama i žbunastom vegetacijom.Madagaskar se od Afrike odvojio prije oko 165 miliona godina, a od Indijeprije oko 100 miliona godina, tako da fauna ove oblasti predstavlja staruizoliranu otočnu faunu sa visokim procentom endemizma, odnosno vrstama kojese nigdje više ne mogu sresti na planeti. Na Madagaskaru živi preko 400 vrsta gmazova i vodozemaca među kojimasu najpoznatiji slonovska kornjača (Testudo gigantea), zelena morska kornjača,blizu sto vrsta kameleona iz endemičnih rodova Furcifer i Calumma (Calumma parsonii), madagaskarski gušter (Chalarodon madagascariensis), madagaskarskaiguana (Oplurus cuvieri), madagaskarska boa (Acrantophis madagascariensis),madagaskarska šumska boa (Sanzinia madagascariensis), velika lažna boa (Bolyeriamultocarinata), nilski krokodil (Crocodylus niloticus).Od oko 300 vrsta ptica, trećina je endemična - veliki flamingo(Phoenicopterus roseus), južni pingvin (Eudyptes chrysocome), žutokljuni albatros(Thalassarche chlororhynchos), Barauova burnica (Pterodroma baraui),madagaskarski ibis (Lophotibis cristata), ružičast pelikan (Pelecanus rufescens),velika fregata (Fregata minor), madagaskarski orao ribar (Haliaeetus vociferoides),crni papagaj (Coracopsis nigra), madagaskarski crveni štiglić (Foudiamadagascariensis), vanga sa šljemom (Euryceros prevostii), itd. Ovde je živjela islonovska ptica (Aepyornis maximus) koja je pripadala ne letačima, a čija jedužina iznosila tri metra. Na žalost, istrijebljena je u XVII stoljeću. Ista sudbinazadesila je i pticu dodo (Raphus cucullatus) sa Mauricijusa, a u skorije vrijeme iDijakurovog malog gnjurca (Tachybaptus rufolavatus).Čekinjasti ježevi ili tenreci (porodica Tenrecidae) su kukcojedizastupljeni sa 30 vrsta najprimitivnijih placentalnih sisavaca u suvremenomživotinjskom svijetu. To su male životinjice veličine 4 do 40 cm. Odlikuju sedlakavim kožnim prekrivačem sastavljenim od čekinjastih ili tvrdih dlaka.Neke vrste tenreka žive na zemlji, neke riju hodnike ispod zemlje, a neke vodePolu vodeni način života. Zbog različitog načina života među sobom se imorfološki razlikuju. Jedan broj vrsta podsjeća na ježeve, dok neke druge vrste

imaju dugačke repove i liče na miševe. Od zvijeri Madagaskara, specifična je fosa (Cryptoprocta ferox). To jetipičan mesožder koji se hrani sitnijim životinjicama i veoma je aktivan u potrazi za hranom. Naseljava uglavnom planinske dijelove otoka. Dostiže dužinuod oko 70 cm, a po svom izgledu najsličniji je kombinaciji hijena i mačaka.Porodica Eupleridae kojoj fosa pripada, filogenetski je najbliža porodicimungosa čiji predstavnik, prstenorepi mungos (Galidia elegans), također naseljavaovo otočje. Od ostalih zvijeri, na Madakaskaru živi i fanolka (Fossa fossa).Madagaskar se odlikuje i bogatom faunom glodara (Rodentia). Među 30vrsta glodara, svojim specifičnim odlikama naročito se ističe malagaškiveliki pacov. Za razliku od glodara, papkari su zastupljeni samo jednom vrstom,riječnom svinjom (Potamochoerus porcus).Na otoku je registrirano 48 vrsta slijepih miševa od kojih većina predstavlja endemite ovog prostora. Polumajmuni lemuri koji pripadaju skupini polumajmuna sa dugim repom (Prosimiae), sa 99 pronađenih vrsta, predstavljaju jednu od najbrojnijih skupina madagaskarskih sisavaca. Za razliku od pravih majmuna, imaju slabije razvijenmozak i neke zaostale odlike zvijeri. Najpoznatiji među madagaskarskimpolumajmunima su lemuri ‒ lasičasti lemur (Lepilemur mustelinus), prstenorepilemur, (Lemur catta), crni lemur (Lemur macaco), veliki lemur bambusar(Prolemur simus), mali lemur bambusar (Hapalemur griseus), mišoliki lemur(Microcebus murinus), patuljasti lemur (Cheirogaleus medius), indri lemur (Indriindri), aj-aj (Daubentonia madagascariensis) i mnogi drugi. Na žalost, izravni iindirektni utjecaji čovjeka doveli su do iščezavanja brojnih sisavaca Madagaskara,između ostalog, 17 vrsta lemura, velike fose (Cryptoprocta spelea) imadagaskarskog nilskog konja (Hippopotamus lemerlei).
3. Indomalajska zoogeografska podoblastIndomalajska zoogeografska oblast obuhvaća Indiju, Indokinu i Malajskiarhipelag zaključno sa Filipinima. Odlikuje se bogatom faunom koja je zanijansu siromašnija od Etiopske oblasti. Tipični predstavnici ove podoblasti suleteća žaba (Rhacophorus reinwardtii), velikoglava kornjača (Platysternonmegacephalum), malajski vodeni gušter (Hydrosaurus amboiensis), indijska agama(Calotes versicolor), varan (Varanus komodoensis), leteća zmija (Chrysopelea spp),naočarka (Naja naja), kraljevska kobra (Ophiophagus hannah), estaurski krokodil(Crocodylus porosus), močvarni krokodil (Crocodylu. palustris), paun (Pavocristatus), ptica nosorog (Buceros bicornis), džinovska leteća vjeverica (Petauristaspp), sunčev medvjed (Helarctos malayanus), gepard (Acinonyx jubatus), zlatnaazijska mačka (Profelis temmincki), malajski tapir (Tapirus indicus), indijski nosorog(Rhinoceros unicornis), četvororožna antilopa (Tetracerus quadricornis), crnaantilopa (Antilope cervicapra), indijski bivol (Bubalus bubalis), Od primata,prisutni su veliki lori (Loris tardigradus), aveti (Tarsius spp.), rezus (Macacamulatta), beloruki gibon (Hylobates lar), orangutan (Pongo pygmaeus), itd.
IV ZOOGEOGRAFSKA OBLAST NOTOGEJA
Zoogeografska oblast Notogeja sastoji se iz tri podoblasti: novozelandske,australijske i polinezijske. Ova oblast se odlikuje dominacijom jedinstvenih

Austrakijska,Ppapuanska, Polinezijska, Havajska i Novozelendska podoblast

Podrijetlo tobolčara, kreda eocen prije 90 mil. G.

životinjskih skupina koje predstavljaju sponu sa drevnom faunom mezozoika.Osnovni razlog visokog endemizma i reliktnosti faune Notogeje leži u ranomodvajanju Australije i okolnih otoka od Gondvane, odnosno, u njenoj dugojizoliranosti koja je započela prije evolutivnog razmaha sisavaca s posteljicomOčuvanju mezozojske faune doprinijele su i relativno stabilne klimatskeprilike. Doduše, prije oko 15 milijardi godina klima je postala sušnija uslijedpromjene pravca morskih struja, ali ova promjena nije zahvatila sve dijeloveAustralije. Zbog toga, vlažniji dijelovi Australije i njeni otoci i dalje suostali utočište preživjele mezozojske faune, dok su sušna prostranstvaosvojile vrste koje su se postupno prilagođavale uvjetima kserotermije. Na tajnačin, fauna Notogeje dodatno je obogaćena.
1. Novozelandska zoogeografska podoblast
Novozelandska zoogeografska podoblast obuhvaća Novi Zeland, kao iobližnja manja otoci (Srjuart, Katam, Okland, Veliki Greben, Durvil, Vajheke).Ovi otoci nalaze se u jugozapadnom dijelu Tihog oceana, preko 1.500 kmjugoistočno od Australije. Sam Novi Zeland podijeljen je na Sjevernio i Južnootočje između kojih se nalazi 22 km širok Kukov morski prolaz. Gotovo čitavomdužinom Južnog otoka uzdižu se Južne Alpe čiji najviši vrh Maunt Kuk dostiževisinu od 3.754 m. U podnožju Južnih Alpa nalazi se oblast Fjordlend sadubokim fjordovima. Sjeverno otočje ima manje planinskih vijenaca, ali zato imaveliki broj vulkana. Vrh vulkana Taupo doseže do 2,797 m i predstavljanajvišu točku Sjevernog otočja. Klima na području Novozelandije je blagamaritimna, s tim što u kontinentalnim dijelovima nekih otoka prelazi usemiaridnu, a na samom sjeveru u suptropsku varijantu.Novozelandska otoci imaju faunu koja je u velikoj mjeri sačuvala svoj primarni mezozojski karakter i predstavlja najstariju faunu koja danas živi naZemlji. Odlikuju je džinovski kukci, prastari vodozemci i gmazovi, brojne

endemične vrste ptica i potpuno odsustvo autoktonih sisavaca.Od vodozemaca na ovim otocima žive samo četiri vrste primitivnihžaba roda Leiopelma. Veoma su slične fosilnim žabama koje su živjele prije 150miliona godina u doba Jure. Ove žabe ne prolaze kroz stadij punoglavaca,nemaju plovne kožice, uglavnom žive u šumama i nisu bezuvjetno vezane za vodu.Naročito su ugrožene Arčijeva žaba (Leiopelma archeyi) i Hohšteterova žaba(Leiopelma hochstetteri) koje se danas nalaze na veoma ograničenom prostoruSjevernog otočja. Gmazovi Novog Zelanda predstavljeni su sa samo osam autoktonih vrsta.Među gmazovima nema zmija, kopnenih kornjača i krokodila. Najinteresantnijigmazovi su dvije vrste koje pripadaju rodu haterija ili sfenodona (Sphenodon).Haterije Novog Zelanda predstavljaju izravnu potomačku vrstu iz trijasaprimitivnih guštera i predstavljaju takozvane „žive fosile“ čija se starostprocjenjuje na oko 200 miliona godina. Od gmizavaca koji su kasnije stigli na oveotoke, ističu se skinkovi (Scincus) i gekoni (Gekotta).Za razliku od faune gmazova koja je relativno siromašna vrstama,fauna ptica je veoma bogata. Mnoge od njih su izgubile sposobnost letenja jertijekom dugog razdoblja njihovog evolutivnog razvoja na Novom Zelandu nije bilosisavaca, pa samim tim ni glavnih predatora koji bi ih ugrožavali. Naročito seističu ptice trkačice. Jadan od rodova koji je obuhvaćao najveće ptice trkačicebio je rod moa (Dinornis) čiji su pojedini predstavnici živjeli do XVIII st. Odživih predstavnika ptica naročito su interesantni kiviji (Apterygidae)zastupljeni sa šest vrsta koje predstavljaju simbol Novog Zelanda. Također,izdvajaju se sove papagaji (Strigops habroptila) koje dostižu veličinu domaćekoke. Imaju razvijena krila, ali pošto im je perje mekano, ne mogu da lete.Specifične ptice ovih otoka su i papagaji nestori (Nestor), endemične vrstesokolova, barske koke, ronac iz roda Mergus, itd. Na Novom Zelandu se nalazinajveći broj vrsta pingvina, uključujući i endemične vrste među kojima seizdvajaju žutooki pingvin (Megadyptes antipodes), mali pingvin (Eudyptulaalbosignata albosignata), snerski pingvin (Eudyptes robustus) i fjorlandski pingvin(Eudyptes pachyrhynchus).Autoktonih vrsta sisavaca na novozelandskim otocima gotovo da nema. Dokčovjek nije doveo oko 600 vrsta sisavaca pretežno iz Evrope, na ovim su otocimaod sisavaca živjele samo dvije vrste slijepih miševa, morski lav i novozelandskafoka krznašica. Ove starosjedilačke vrste su na Novi Zeland stigle prelijetanjemi preplivavanjem. Autoktona novozelandska fauna se održavala sve dok otoci nisunaselili Maori u XII vijeku. Maori su naišli na izobilje hrane, pijre svega naveliku mou koja je imala velike količine mesa i krupna jaja. Zbog nekontroliranoglova, dvije vrste džinovskih moa (Dinornis maximus), kao i desetak vrsta manjih moa,istrijebljeno je u razdoblje od 1500 do 1800 godine. Ubrzo su nestale i neke vrstenovozelandskih gusaka, kao i Haastov orao (Harpagornis moorei), najveća pticagrabljivica na svijetu. Razlog njegovog iščeznuća bio je lov, ali i nestanak moakoje su mu bile osnovna hrana. Dolaskom Evropljana u XVIII vijeku, uništavanjefaune je nastavljeno. Nestale su i novozelandska prepelica, smijajuća sova, huijai druge jedinstvene vrste ptica.
2. Australijska zoogeografska podoblastOva zoogeografska oblast obuhvata kontinent Australija i okolne otokeu Južnom, Indijskom i Tihom oceanu (Novu Gvineju, Tasmaniju, Bizmarkovarhipelag, Solomonska otoci, Timor).

U reljefu Australije se izdvajaju tri velike prirodne regije: Istočnopobrđe, Središnja ravnica i Zapadni plato. U Istočnom pobrđu nalazi se najvišeplanine Australije koje dosežu do 2.228 m nadmorske visine. Središnja ravnica jenajniže područje Australije koje u zapadnom dviju prelazi u pješčane pustinje.Zapadni plato obuhvata pustinje, polupustinje i travnate ravnice iznad kojih seuzdižu niže planine. U klimatskom pogledu, najveći dio Australije nalazi se pod utjecajempustinjske aridne klime. Umjerena klima vlada samo na jugoistoku i jugozapaduzemlje, dok je tropska klima svojstvena sjevernim dijelovima kontinenta.Australija se odlikuje specifičnim i raznovrsnim ekološkim uvjetima zbog otočne izoliranosti, različitih klimatskih utjecaja, složenog reljefa i raznovrsne vegetacije, od prašumske do pustinjske. Australijska zoogeografska podoblast obuhvata faunu koja se po starostinalazi na drugom mjestu, odmah iza novozelandske oblasti. Čitava podoblast dijeli sena dvije provincijei – Australijsku koja pored Australije obuhvata i otokTasmaniju, i Papuansku koja obuhvata Novu Gvineju.Fauna gmazova i vodozemaca ove podoblasti veoma je osobna. Odautoktonih vrsta prisutne su brojne, uglavnom otrovne zmije, kratkonogi gekoniprekriveni krupnim krljuštima (Uromastyx sp.), agama gušteri među kojima seizdvaja Chlamydosaurus kingii koji trči veoma brzo oslanjajući se na zadnje noge,kusorepi cinkus (Tiliqua rugosa), vodena kornjača (Carettochelys insculpta),estaurski krokodil (Crocodylus porosus), australijski krokodil (Crocodylusjohnsoni), itd. Fauna ptica je veoma bogata endemičnim vrstama u koje između ostalih, spadaju emu nojevi (Dromaius sp.), brojni papagaji (nimfa – Nymphicus hollandicus, tigrica – Melopsittacus undulatus, crvena rozela – Platycercus elegans, ružičasti kakadu – Cacatua leadbeateri, crni kakadu – Calyptorhynchus funereus, brojni loripapagaji – Loriini), ptice nosorozi (Bucerotidae), rajske ptice (Paradisea sp.), kokehumkašice (Megapodius, Leipoa, Alectura), ptica-lira (Menura novaehollandiae),itd. Od drevnih aplacentalnih sisavaca najpoznatiji su kljunaši i torbari. Kljunaša su primitivni sisavaca iz reda Monotremata. Razlikuju se od svihostalih sisavaca po tome što polažu jaja, imaju elastičan kljun, zube prisutne samokod mladunaca i samo jedan izlazni tjelesni otvor (kloaku) koji služi zaoplodnju, izbacivanje mokraće i izmeta. Po ostalim osobinama slični susisavcima jer su prekriveni krznom, toplokrvni su i doje mladunce. Doduše,nemaju bradavice za dojenje, nego difuzno raspoređene mliječne žlijezde koje nagrudima majke obrazuju mliječna polja, tako da mladunci ne sišu, nego ližu mlijeko.U Australiji i Novoj Gvineji danas je prisutno pet vrsta kljunaša.Torbari (Marsupialia) su sisari koji zbog nedostatka posteljice rađajunepotpuno razvijene mladunce koji kod koale na primjer, teže svega 0,3 grama. Dabi preživio, mladunci se čvrsto drže za majčine bradavice dok ne dostignuodređeni stupanj samostalnosti. Neki torbari, na primjer kenguri (Macropus sp.),koala (Phascolarctos cinereus) i vombate (Vombatus sp.), imaju velike kožne torbeoko bradavica tako da mladunce osiguravaju od ispadanja. Pored navedenih, uAustraliji živi još oko 160 vrsta torbara koji su imena dobili na osnovusličnosti sa nekim placentalnim sisarima – torbarski mravojed (Myrmecobiusfasciatus), torbarska krtica (Notoryctes typhlops), torbarski jazavac (Peramelessp.), torbarski skočimiš (Antechinomys sp.), torbarska lisica (Trichosurusvulpecula), torbarska mačka (Dasyurus sp.), leteći torbarski miš (Acrobatespygmaeus), leteće vjeverice (Petaurus sp.), itd. Na žalost, torbarski vuk poznat i

kao tasmanijski đavo (Thylacinus cynocephalus), sa lica Zemlje nestao je u XX vijekuuslijed nekontroliranog lova.Za razliku od kljunaša i torbara, autoktona fauna sisavaca s posteljicomAustralijske podoblasti, veoma je siromašna. Predstavljaju je samo slijepi miševi,neki sitni glodari, pas dingo (Canis dingo) i rusa jelen (Cervus timorensis) koji suu davnoj prošlosti u ove predijele stigli sa Indomalajskog arhipelaga.Australijska oblast se dijeli na dvije provincije – Australijsku i Papuanskukoja obuhvata Novu Gvineju. Specifičnu faunu Nove Gvineje koja nije prisutna uAustralijskoj oblasti, predstavljaju prije svega, tobolčari iz roda Dendrolagusprilagođeni životu na drveću, torbolčar kuskus (Phalanger orientalis), kazuari kojipripadaju familiji nojeva (Casuariidae), veliki gekon (Rhacodactylus leachianus),novogvinejski krokodil (Crocodylus novaeguineae), zmija smrtnica (Acanthophisantarcticus), pacifička boa (Enygrus carinatus), džinovski golub (Ducula goliath),zeleni golub (Drepanoptila holosericea), papagaji kardinali (Erytrhrura psittacea),uvea papagaji (Eunymphicus cornutus uveaensis).
3. Polinezijska zoogeografska podoblastOva zoogeografska podoblast obuhvaća Novu Kaledoniju koju čini otokGrand Ter i mnogobrojna mala melanezijska otoka i arhipelazi u njegovojblizini. Sva otoci ovog dijela svijeta su vulkanskog ili koraljnog podrijetla osimGrand Tera koja predstavlja ostatak Gondvane. Najviša točka ovog otoci je MonPanije nadmorske visine od 1.628 m. Na otocima vlada tropska klima saprosječnom količinom padavina 1.200 mm godišnjeFauna na ovim otocima je veoma siromašna. Vodozemaca na većem brojuotoka nema. Izuzetak su samo Sendvička otoci i Havaji na kojima se mogu vidjetikrastave žabe iz familije Bufonidae i Fidži na kome žive obične žabe izfamilije Ranidae. Među gmazovima nedostaju krokodili, zmije, morske i kopnenekornjače. Karakteristični gmazovi su gušteri iz rodova skinkova (Scincus),gekona (Gekotta) i iguana (Iguania).Za razliku od faune vodozemaca i gmazova, fauna ptica je veoma bogata. Na otocima se sreću kagu ždralovi (Rhynochetus jubatus), pećinske čiope(Collocalia fuciphaga), havajske nektarice, brojni papagaji među kojima se izdvajajulori (Trichoglossus sp.), zvišci, autohtoni golubovi, vodomari, i još neke drugerijetke vrste. Starosjedilački sisavci su predstavljeni samo sitnim glodarima i slijepimmiševima.

Distribucija nojevki:
Podrijetlo pjevica:
Prije 60 mil. god. prije 20 mil. god.