rickettsia , orientia , ehrlichia , coxiella ,chlamydia tekst
DESCRIPTION
Porodica Rickettsiaceae---------------------------------------------------- 3Rod Rickettsia--------------------------------------------------------------- 4Rikecije skupine pjegavih tifusaRickettsia prowazekii------------------------------------------------------- 9Rickettsia typhi-------------------------------------------------------------- 10Rikecije skupine pjegavih groznica Rickettsia conorii------------------------------------------------------------ 11Rickettsia rickettsii---------------------------------------------------------- 12Rickettsia akari-------------------------------------------------------------- 13Rod Orientia----------------------------------------------------------------- 13Orientia tsutsugamushi----------------------------------------------------- 13Rod Ehrlichia---------------------------------------------------------------- 14Rod Coxiella----------------------------------------------------------------- 16Chlamydia-------------------------------------------------------------------- 17Chlamydia trachomatis----------------------------------------------------- 20Chlamydia psittaci----------------------------------------------------------- 24Chlamydia pneumoniae----------------------------------------------------- 25TRANSCRIPT

Bosna i HercegovinaUniverzitet u Zenici Zenica, april 2006 g.
Mikrobiologija i ParazitologijaVanredni studij
SEMINARSKI RAD :
Rickettsia , Orientia , Ehrlichia , Coxiella , Chlamydia
Studenti: Alić Nino Dizdarević Amar Drinić Venad Kahrić Alma Pašalić AdmiraMentor: Zec MersihaDoc.dr sc. Selma Marušić MirnesaUzunović –Kamberović
1

Sadržaj:
Uvod-------------------------------------------------------------------------- 3 Porodica Rickettsiaceae---------------------------------------------------- 3
Rod Rickettsia--------------------------------------------------------------- 4
Rikecije skupine pjegavih tifusa
Rickettsia prowazekii------------------------------------------------------- 9
Rickettsia typhi-------------------------------------------------------------- 10
Rikecije skupine pjegavih groznica
Rickettsia conorii------------------------------------------------------------ 11
Rickettsia rickettsii---------------------------------------------------------- 12
Rickettsia akari-------------------------------------------------------------- 13
Rod Orientia----------------------------------------------------------------- 13
Orientia tsutsugamushi----------------------------------------------------- 13
Rod Ehrlichia---------------------------------------------------------------- 14
Rod Coxiella----------------------------------------------------------------- 16
Chlamydia-------------------------------------------------------------------- 17
Chlamydia trachomatis----------------------------------------------------- 20
Chlamydia psittaci----------------------------------------------------------- 24
Chlamydia pneumoniae----------------------------------------------------- 25
2

Uvod
Pripadnost rikecija i klamidija dugo vremena nije mogla da se odredi. Te dvije grupe mikroorganizama bile su svrstavane između bakterija i virusa . I rikecije i klamidije su slične virusima po tome što se ne mogu razmnožavati van žive stanice. Rikecije , međutim imaju mnogo više osobina bakterija. One su vidljive u običnom optičkom mikroskopu , ne prolaze kroz bakterijske filtre ( izuzev roda Coxiella ) , imaju stanični zid koji sadrži muraminsku kiselinu , dijele se cijepanjem u dvije nove jedinke , mogu generisati energiju i proizvoditi ATP iz ADP i sadrže DNK i RNK – za razliku od virusa koji sadrže uvijek samo jednu od te dvije nukleinske kiseline . Zbog svih tih osobina rikecije su svrstane u bakterije . I klamidije imaju više osobina bakterija nego virusa . I one se mogu vidjeti običnim optičkim mikroskopom , primaju anilinske boje i po Gramu se boje negativno . Posjeduju stanični zid koji sadrži muraminsku kiselinu, dijele se cijepanjem u dvije nove jedinke i sadrže DNK i RNK. Zbog tih mnogih bakterijskih osobina i klamidije su svrstane u bakterije .
Rickettsiaceae
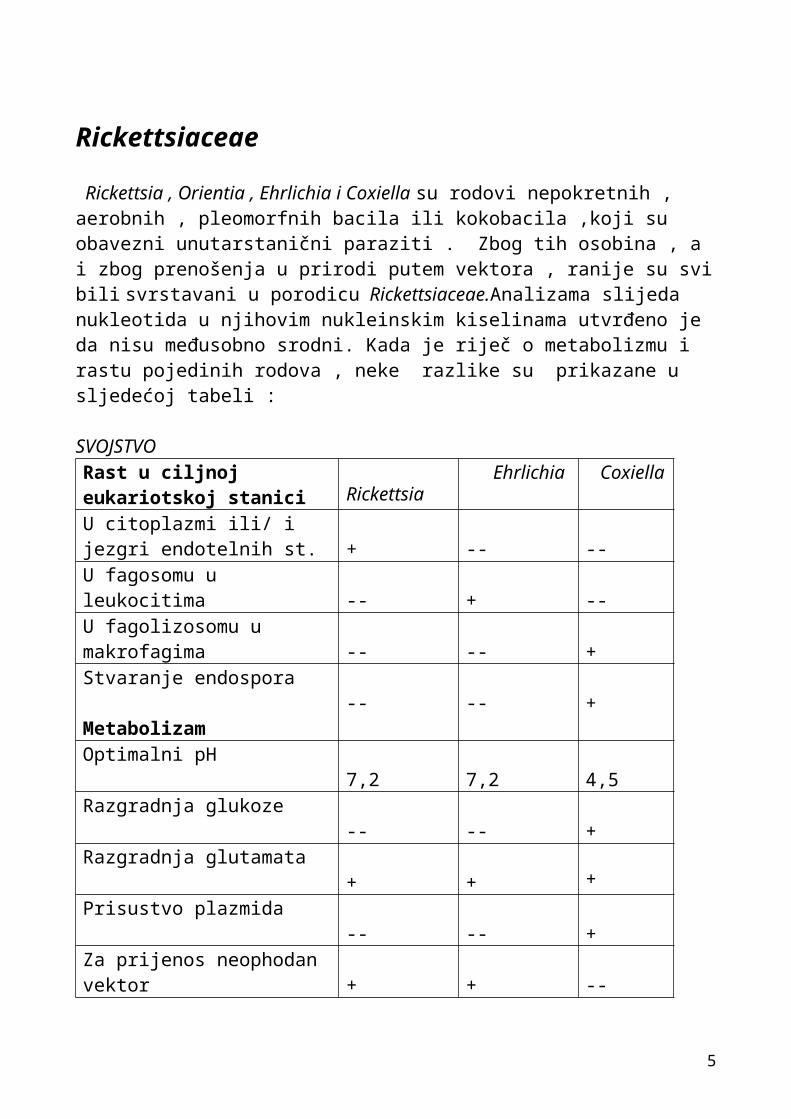
Rickettsia , Orientia , Ehrlichia i Coxiella su rodovi nepokretnih , aerobnih , pleomorfnih bacila ili kokobacila ,koji su obavezni unutarstanični paraziti . Zbog tih osobina , a i zbog prenošenja u prirodi putem vektora , ranije su svi bili svrstavani u porodicu Rickettsiaceae.Analizama slijeda nukleotida u njihovim nukleinskim kiselinama utvrđeno je da nisu međusobno srodni. Kada je riječ o metabolizmu i rastu pojedinih rodova , neke razlike su prikazane u sljedećoj tabeli :
SVOJSTVO Rast u ciljnoj eukariotskoj stanici
Rickettsia Ehrlichia Coxiella
U citoplazmi ili/ i jezgri endotelnih st.
+ -- --
U fagosomu u leukocitima -- + --U fagolizosomu u makrofagima -- -- +Stvaranje endospora
Metabolizam
-- -- +
Optimalni pH 7,2 7,2 4,5Razgradnja glukoze -- -- +Razgradnja glutamata + + +Prisustvo plazmida -- -- +Za prijenos neophodan vektor + + --
3

Rickettsia Prvu rikeciju je otkrio Ricketts pri istraživanjima američkog pjegavca. Morfološki identičan mikroorganizam izolovao je Prowazek 1913 g. za vrijeme velike epidemije pjegavca u Srbiji. Da Rocha – Lima je takve mikroorganizme izolovao iz osoba koje su bolovale od pjegavog tifusa kao i iz bijelih ušiju . On je dao i prvi detaljan opis tih organizama i nazvao ih Rickettsia prowazeki.
Morfologija
Rikecije su kratki, štapićasti ili kokobacilarni organizmi dugi od 800 – 2000 nm i široki od 300 – 500 nm. Po Gramu se slabo boje negativno , pa se u stanici jasnije prikazuju bojenjem po Giemsi ( grimizno), Gimenezu ( crveno na zelenoj pozadini), ili po Macchiavellu ( crveno na plavoj pozadini ) . Elektronskim mikroskopom vidi se tipična gram- negativna prokariotska stanica sa troslojnim staničnim zidom , sastavljenim od unutarnje i vanjske membrane sa slojem peptidoglikana između . Izvan vanjske membrane rikecije imaju proteinsku mikrokapsulu i bogati sluzavi sloj koji se vidi kao svijetlo područje oko nje.
Razmnožavanje i uzgoj
Rikecije se razmnožavaju u citoplazmi ili u jezgrama eukariotskih stanica , pa se za uzgoj u laboratoriju koriste pokusne životinje ili češće kulture stanica i žumanjčana vrečica oplođenih kokošijih jaja. Optimalna temperatura za rast rikecija je 33-35*C, a obilnije se razmnožavaju u stanicama usporenog metabolizma. Rikecije ulaze u eukariotsku stanicu pomoću fagocitoze, u kojoj i sama učestvuje. Rikecijska fosfolipaza A kida lipide citoplazmatske membrane i ulazi. Da bi rikecije izbjegle razgradnju i preživjele u stanici, moraju nakon ulaska razoriti membranu fagosoma i ući u citoplazmu , prije nego se fagosom spoji sa lizosomom. U citoplazmi se dijele na dvoje a generacijsko vrijeme im je 8-12 sati. Rikecije se razmnožavaju dok potpuno ne ispune stanicu, nakon čega citoplazmatska membrana puca, a rikecije se oslobađaju i inficiraju nove stanice.
4
Metabolizam
Rikecije imaju metaboličke sposobnosti kao i druge bakterije, ali su im za pravilan razvoj potrebni neki metaboliti koje dobivaju od stanice domaćina pa su razvile dvojnu strategiju za preuzimanje molekula bitnih za svoj metabolizam .
Posjeduju enzim ADP/ ATP translokaza , koji zamjenjuje molekulu ADP iz rikecije za molekulu ATP iz inficirane stanice domaćina ali kad nestane tih visokoenergetskih fosfata, sposobne su sintetizirati vlastiti ATP.
Sličnim mehanizmom prijenosa rikecije uvlače i uridin-5-difosfoglukozu koja im je potrebna za sintezu polisaharida. Antigena struktura Na osnovi razlike u antigenima i biološkim svojstvima, vrste u rodu Rickettsia svrstane su u dva različita biotipa : - biotip pjegavih tifusa – PT ( R. prowazekii i R. typhi ) - biotip pjegavih groznica- PG (R rickettsii, R sibirica, R conorii, R. akari, R australis, R africae, R japonica, R slovaca, R helvetica, R felis, te čitav niz drugih rikecija koje su izolovane iz krpelja a za koje nije dokazano da izazivaju bolest kod čovjeka ) .
Antigeni specifični za skupinu
Termostabilni topljivi lipopolisaharid (LPS) koji se nalazi u staničnom zidu rikecija nosi antigene determinante specifične za skupinu , odnosno biotip. LPS se oslobađa iz rikecija nakon obrade sa eterom. Ti su antigeni specifični za sve članove istog biotipa pa se po njima razlikuje biotip tifusa od biotipa pjegavih groznica a dokazujemo ih pomoću reakcije vezanja komplementa ( RVK).
Zagrijavanjem topljivog grupno specifičnog antigena u alkalnom mediju oslobađa se tvar koja senzibilizira eritrocite (ESS) . ESS se adsorbira na ovčije ili ljudske eritrocite O- skupine i pokazuje grupnospecifičnu reaktivnost za rikecije u jednom biotipu. ESS dokazujemo testom indirektne hemaglutinacije (IHA) ili latex-aglutinacijom .Taj antigen ne možemo dokazati u RVK .
LPS u vrstama oba biotipa u rodu Rickettsia ( osim R.akari ) ima zajedničke epitope s OX-19 ( srodniji više s biotipom tifusa ) i OX-2 ( srodniji s biotipom pjegave groznice) . Ova je unakrsna reakcija osnova nespecifičnog testa aglutinacije po Weil-Felixu koji pokazuje djelomičnu grupnu specifičnost .
5

Antigeni specifični za vrstu
U vanjskoj membrani rikecija se nalaze termolabilni proteinski antigeni koji sadrže epitope specifične za vrstu rikecija, a izvan vanjske membrane rikecije stvaraju mikrokapsulu , površinski proteinski sloj , koji je specifičan za vrstu . Ovi proteinski antigeni potiču stvaranje zaštitnih antitijela . Antitijela za ove antigene se dokazuju : - reakcijom vezanja komplementa- testom imunofluorescencije- mikroaglutinacije- ELISA-testom neutralizacije toksičnosti za miša
Osjetljivost na fizikalne i hemijske faktore okoline
Bakterije roda Rickettsia vrlo su neotporne prema faktorima iz okline pa brzo ugibaju . Izvan žive stanice rikecije gube svoju metaboličku aktivnost, gube kroz svoju membranu proteine, nukleinske kiseline i ostale važne molekule kao što su NAD, ATP, pa budući da im za prodor u stanicu treba energija, prestaju biti infektivne. Brzo ih inaktiviraju uobičajeni dezinficijensi uključujući Na-hipoklorit, 5% H2O2,
8% formaldehid, 1% fenol, 70% etanol i UV zrake. Pri temperaturi od 56*C rikecije ugibaju za 30 min.
Iako rikecije brzo propadaju pri sobnoj temperaturi u sasušenom fecesu zaraženih uši može R. prowazekii ostati infektivna više mjeseci ako su vlažnost i temperatura okoliša niski. Infektivne rikecije čuvaju se pri –70*C ili liofilizirane .
Osjetljivost na antimikrobne lijekove
Osjetljivost rikecija prema antibioticima se određuje na kulturi stanica postupkom inhibicije stvaranja plakova u prisustvu određenog antibiotika. Tetraciklini, kloramfenikol, fluorirani kinoloni , neki makrolidi i rifampicin djeluju inhibitorno na rikecije in vitro. Najbolji učinak imaju tetraciklini pa se zbog toga najviše upotrebljavaju u liječenju. Sulfonamidi pojačavaju rast rikecija , pa su kontraindicirani u liječenju rikecioza. Penicilini , cefalosporini i aminoglikozidi ne djeluju na rikecije.
6

Patogeneza
Nakon uboda zaraženog člankonošca rikecije ulaze u endotelne stanice kapilara i u njima se razmnožavaju. Inficirane endotelne stanice nabreknu i nekrotiziraju; nastaje tromboza, pa to dovodi do začepljenja, nekroze i razdora kapilara. Oko takvih se mjesta uz kapilare nakupljaju mononuklearne upalne stanice ( Ly, Makrofagi ) i stvaraju patognomonične tifusne čvoriće. Iz raspuklih endotelnih stanica oslobađaju se rikecije i krvlju prenose po cijelom organizmu. Oštećenje malih krvnih žila najizraženije je u koži, miokardu i mozgu, ali i u drugim organima i čini se da je odgovorno za većinu kliničkih znakova , uključujući osip. Epidemični tifus, pjegava groznica Stjenovitog gorja i šikarski tifus spadaju među najteže poznate zarazne bolesti s visokom smrtnosti. Oštećenje stanica vjerovatno nastaje zbog : toksičnosti metaboličkih produkata rikecija , natjecanja rikecija sa stanicom za vitalne supstrate, iscrpljenja ATP-a i oštećenja stanične membrane zbog ulaska brojnih rikecija i kasnije zbog masovnog napuštanja stanice. Kao odgovor na infekciju rikecijama javlja se humoralni i stanični imunitet. Antitijela ne djeluju na rikecije koje se razmnožavaju u endotalnim stanicama, ali kad se rikecije razmnože i napuste stanicu fagocitiraju ih makrofagi. U makrofagima, ako nema antitijela rikecije ostaju u fagolizosomu i ugibaju. Da bi se ubile rikecije potrebna je suradnja humoralnih antitijela i makrofaga . Nasuprot tome ubijanje rikecija unutar endotelnih stanica ne ovisi o humoralnim antitijelima i vjerovatno je uzrokovano staničnim imunitetom, koji je važniji u kontroliranju unutarstaničnog rasta rikecija. Unatoč humoralnom i staničnom imunitetu, koji su izraženi i dugotrajni , rikecije mogu perzistirati u tkivima godinama nakon infekcije.
Mikrobiološka dijagnostika
Laboratorijska dijagnostika rikecioza može se postići izolacijom uzročnika, izravnim dokazom uzročnika u tkivu ili serološkim reakcijama.
Izolacija
Rikecije su vrlo infektivni mikroorganizmi pa se izolacija radi vrlo rijetko. Za izolaciju se uzima krv u epruvetu s heparinom na početku bolesti prije nego je započeto antimikrobno liječenje.Izolacija se radi na kulturi stanica . Nakon 2-3 dana inkubacije uzorka u kulturi stanica pravi se razmaz i dokazuje prisutnost rikecija u stanicama ( bojenje po Giemzi, Gimenezu ili s antitijelima obilježenim imunofluorescentnom bojom).
7

Direktno dokazivanje rikecija
Izravno se mogu dokazati u uzorku kože uzetom biopsijom s mjesta s osipom ili primarnom lezijom. Specifičnim antitijelima obilježenim fluoresceinom ili enzimom mogu se izravno dokazati rikecije u endotelnim stanicama kapilara kože. Istim postupkom mogu se rikecije dokazati i u postmortalno uzetim komadićima tkiva. PCR se može primijeniti za amplifikaciju DNA rikecija u punoj krvi, leukocitno- trombocitnom sloju stvorenom iznad koncentriranih eritrocita, plazmi ili u bioptatima kože s osipnom promjenom.
Serološke reakcije
Dijagnoza rikecijskih bolesti se najčešće potvrđuje serološkim reakcijama. Antitijele u serumu se stvaraju tek u drugoj sedmici bolesti pa je potrebno uzeti prvi serum na početku bolesti i drugi serum nakon 3 sedmice. Za potvrdu dijagnoze važno je dokazati barem četvorostruki porast titra specifičnih antitijela nastao kao odgovor na infekciju rikecijama. Razlikovanje antitijela IgM ili IgG omogućuje da se u jednom serumu odredi da li je infekcija akutna ili nije.
Weil-Felixova reakcija aglutinacije. Zasniva se na unakrsnoj reaktivnosti epitopa u LPS mnogih rikecija s onima u O antigenu sojeva OX-2 i OX-19 Proteus vulgaris. Antitijela koja aglutiniraju sojeve proteusa mogu se javiti 7-14 dana nakon pojave bolesti, pretežno su IgM klase a svoj maksimum dostižu u ranoj rekonvalescenciji. Reakcija vezanja komplementa. Antitijela koja vežu komplement obično se razvijaju kasnije u toku bolesti i godinama ostaju pozitivni. To su obično antitijela IgG klase čiji se klinički značaj dokazuje titrom većim od 1: 16. Test indirektne hemaglutinacije –IHA dokazuje antitijela za antigen ESS kojim se oblažu ljudski ili ovčiji eritrociti. Ovaj test otkriva antitijela i IgM i IgG klase. U testu latex- aglutinacije ESS je vezan na lateks čestice . Sa ovim testom moguće je dokazati antitijela 1 sedmicu nakon početka bolesti. Test indirektne imunofluorescencije- IFA . Mikrometoda kojom se može iz jedne kapi seruma istovremeno otkriti antitijela za više od 9 rikecijskih antigena. Otkrivanje antitijela IgM klase dokaz je svježe infekcije. ELISA tehnika služi za dokazivanje IgM i IgG antitijela ali je manje osjetljiva od IFA. Western blot je pouzdan potvrdni test serološke dijagnoze dobivene ranije spomenutim testovima.
RIKECIJE SKUPINE PJEGAVIH TIFUSA
8

U ovoj se skupini nalaze R. prowazekii i R. typhi.
Rickettsia prowazekii R. prowazekii uzrokuje epidemični pjegavi tifus ( typhus exanthematicus ). To je bolest siromaštva , nehigijenskih uvjeta i nesreća koji zbog kretanja neimunog stanovništva omogućavaju održavanje ciklusa prijenosa između čovjeka i prtene uši ( Pediculus humanus corporis ). Uši žive u odjeći , zaražavaju se uzimanjem obroka krvi na bolesniku sa rikecijemijom , i nakon što im se rikecije umnože u stanicama crijeva , izlučuju R. prowazekii fecesom. Osobe se zaražavaju utrljavanjem rikecija u kožu ozlijeđenu ubodom ili aerosolom preko sluznice oka i respiratornog sistema. R. prowazekii ubija svog vektora nakon 2-3 sedmice pa nema prijenosa na sljedeće generacije.
Klinička slika
Bolest počinje naglo nakon inkubacije od 9-12 dana s povišenom temperaturom, glavoboljom, bolovima u mišićima. Oko 5. dana bolesti javlja se osip na trupu koji se širi centrifugalno ali obično ne zahvata lice, dlanove i tabane. Mogu se javiti i neurološke promjene , delirij, stupor , ali i miokarditis, hipotenzija i gangrene na okrajinama. Nakon 2-3 sedmice od pojave simptoma, bolesnik će ili ozdraviti ili umrijeti. Smrtnost je niska u dječijoj dobi ali može biti i do 50% u osoba starijih od 50 godina. Imunitet je dugotrajan ali rikecije ostaju u retikuloendotelnim stanicama a da pri tom čovjek nema nikakvih simptoma bolesti. Brill-Zinsserova bolest je recidiv epidemičnog tifusa koji se javlja godinama nakon primarne infekcije najvjerovatnije uzrokovan slabljenjem imuniteta za rikecije koje su ostale skrivene u RES-u .
Prevencija i liječenje
Suzbijanje epidemičnog tifusa , kao i drugih rikecioza , može se postići prekidom lanca prijenosa i infekcije, uništavanjem vektora ili imunizacijom. Važno je i poboljšati higijenske navike ( redovno pranje odjeće i peglanje) . Vakcina pripremljena iz inficiranih žumanjčanih vrećica sadrži ubijene uzročnike i služi kao djelomična zaštita od bolesti . Atenuirana živa vakcina (Madrid-E-soj ) štiti od bolesti za vrijeme epidemijskog širenja bolesti. Tetraciklini i kloramfenikol su djelotvorni u liječenju i primarnog epidemičnog tifusa i Brill-Zinsserove bolesti.
Mikrobiološka dijagnostika
9

Dijagnoza epidemičnog tifusa najčešće se postavlja serološkim reakcijama. To je u prvom redu Weil-Felixova reakcija, a pomoću IFA i ELISA- testa možemo dokazati antitijela IgM klase što ukazuje na primarni imuni odgovor. U osoba sa Brillovom bolešću možemo dokazati IgG antitijela već krajem prve sedmice.
Rickettsia typhi
R. typhi morfološki i metabolički je slična R. prowazekii. Ova rikecija uzročnik je endemičnog ili murinog ( štakorskog ) tifusa , bolesti koja je klinički i patološki slična epidemičnom pjegavom tifusu, ali je obično blaža. Murini tifus je zoonoza , raširena po cijelom svijetu naročito u umjerenim i suptropskim priobalnim krajevima. Priridni rezervoar su štakori a glavni vektor koji prenosi rikeciju je štakorska buha ( Xenopsilla cheopsis ) . Buhe mogu prenositi rikeciju i transovarijalno pa su one i rezervoar i vektor za tu rikeciju. Čovjek se zarazi slučajno, kada se njegovom krvlju hrani inficirana štakorska buha , koja pri tom defecira i izlučuje rikecije.
Klinička slika
Nakon inkubacije 1-2 sedmice bolest počinje naglo s povišenom temperaturom, bolovima u mišićima. Makulopapulozni osip javlja se na trupu i širi se centrifugalno . Bolest je obično blaga, posebno u djece a smrtnost je niska , uglavnom starije osobe.
Prevencija i liječenje
U endemskim područjima primjenjuju se insekticidi i rodenticidi . Ne postoji djelotvorna vakcina. Lijek izbora u liječenju endemičnog tifusa je tetraciklin . Djelotvorni su i kloramfenikol i fluorirani kinoloni.
Mikrobiološka dijagnostika
Budući da R. prowazekii i R. typhi uzrokuju klinički sličnu bolest mogu se razlikovati biološkim postupkom. Ako se krv bolesnika od murinog tifusa inokulira zamorcu nastat će otok skrotuma i lezije na testisima, za razliku od blage bolesti bez promjena na skrotumu, koja nastaje kad se inokulira krv bolesnika od epidemičnog tifusa.
10

Serološka reakcija. Danas se u dijagnosticiranju murinog tifusa najčešće koristi IFA sa antigenom specifičnim za R. typhi.
RIKECIJE SKUPINE PJEGAVIH GROZNICA
Rikecije iz ove skupine se mogu razmnožavati i u jezgri i u citoplazmi inficiranih stanica , uzrokuju zoonoze , a ljudi su obično slučajni domaćini. U ljudi ove rikecije izazivaju pjegave groznice, koje su nazvane prema području u kojem se javljaju. U ovoj skupini se nalaze rikecije koje prenose krpelji ( Ixodidae ) ubodom prilikom uzimanja krvi na kičmenjaku. Rikecija se u krv prenosi tek nakon 6-10 sati jer to je vrijeme potrebno da se neaktivne rikecije aktiviraju , dođu do slinovnica i prodru u krv čovjeka. Krpelji mogu biti i vektori i rezervoari jer rikecije pjegavih groznica ne oštećuju krpelje. Do sada je dokazano da bolest u čovjeka može uzrokovati 12 rikecija ove skupine . To su : R. rickettsii, R. conorii, R. australis, R. sibirica, R .japonica , R. honei , R. africae, R. felis, R. mongolotimonae, R. slovaca, R. helvetica i R. akari.
Rickettsia conorii
R. conorii je najubikvitarnija od svih rikecija pjegavih groznica. Uzrokuje mediteransku pjegavu groznicu MPG koja se javlja u svim mediteranskim i suptropskim zemljama . Pas je glavni domaćin . Smeđi pseći krpelj Rhipicephalus sanguineus glavni je vektor ali i rezervoar R. conori.
Klinička slika
Nakon inkubacije od obično 7 dana bolest počinje naglo s povišenom temperaturom, glavoboljom i bolovima u mišićima i zglobovima. Karakteristična je pojava primarne lezije , malog ulkusa sa crnim nekrotičnim središtem, koji se javlja na mjestu uboda krpelja, gdje nastaje perivaskularni edem i nekroza kože zbog umnožavanja R. conorii u endotelu. Nakon nekoliko dana javlja se papularni osip prvo na podlakticama , a zatim se širi po cijelom tijelu. Mediteranska pjegava groznica je obično blaga bolest iako se mogu javiti i teži oblici s petehijalnim osipom , neurološkim i kardiovaskularnim simptomima.
11

Prevencija i liječenje
U endemskim krajevima za R. conorii ali i za ostale uzročnike pjegavih groznica najvažnije je izbjegavati dodir s krpeljima i ako je to moguće, potrebno ih je što češće odstranjivati sa tijela jer krpelj se mora hraniti najmanje 4 sata prije nego zarazna rikecija može biti prenijeta. Takođe je važno kontrolisati infestaciju pasa s krpeljima. Tetraciklini i kloramfenikol su djelotvorni u liječenju mediteranske pjegave groznice.
Mikrobiološka dijagnostika
Rana dijagnoza MPG može se postići već u prvoj sedmici bolesti ako se u komadiću kože koji je dobiven biopsijom , s mjesta primarne lezije ili osipa , pomoću specifičnih antitijela obilježenih fluorescentnom bojom dokaže R. conorii. – direktna imunofluorescencija. Za serološku dijagnostiku infekcija s R. conorii danas se najviše koristi IFA
Rickettsia rickettsii
Ova rikecija uzrokuje u SAD pjegavu groznicu Stjenjaka, vrlo tešku i po život opasnu bolest. Glavni vektori ali i rezervoari za R. rickettsii jesu krpelji iz roda Dermacentor . Životinjski rezervoari su divlji glodari , veće divlje i domaće životinje, posebno psi. U pasa se javlja izražena rikecijemija od koje ugibaju.
Klinička slika
Bolest počinje naglo s povišenom temperaturom, glavoboljom , slabošću i bolovima u mišićima . Osip makulopapulozan, ali u teškim slučajevima može biti i hemoragičan. Često se javljaju komplikacije , kao npr. Trombocitopenija , DIK, zatajivanje bubrega i srca. U neliječenih smrtnost je oko 20%.
Prevencija i liječenje je kao u MPG.
Mikrobiološka dijagnostika
12

I R. rickettsii se može dokazati pomoću DIF u komadiću kože dobivene biopsijom već krajem prve sedmice od pojave prvih simptoma bolesti. Za serološku dijagnostiku potrebno je skupiti 3 uzorka . Na početku infekcije, u drugoj i u četvrtoj sedmici od pojave bolesti . Kroz saerološke testove prati se titar, koji mora biti četvorostruko uvećan, ali problem je u tome što oni postaju relativno kasno pozitivni.
Rickettsia akari
Rickettsia akari razlikuje se od ostalih rikecija u skupini pjegavih groznica jer ne stvara toksični učinak na mišu i ne posjeduje zajedničke antigene s Proteus vulgaris. Ova rikecija uzrokuje nekrozu epitelnih stanica, nastaju intraepidermalne vezikule i javlja se vezikulozni osip, pa se zbog toga bolest naziva rikecijskim boginjama. Rezervoar R. akari je sivi kućni miš a vektor mišija grinja ( Allodermanyssus sanguineus ) . Čovjek je slučajni domaćin i obolijeva ako ga ubode inficirana grinja. Na mjestu uboda nastaje primarna lezija u obliku papule koja se pretvara u vezikulu s crnom krastom. Nekoliko dana nakon pojave primarne lezije javlja se povišena temperatura, glavobolja, mijalgija i generalizirani vezikulozni osip nalik variceli. Liječenje tetraciklinima i kloramfenikolom skraćuje tok bolesti na 1-2 dana. Kod serološke dijagnostike ove rikecije, najčešće se primjenjuje IFA.
ORIENTIA
Ova se bakterija razlikuje od rikecija po tome što u staničnom zidu ne sadrži LPS, peptidoglikan ni sluzavi sloj, a ima deblju vanjsku membranu s glavnim proteinom koji nosi glavne antigene determinante specifične za vrstu i tip.
Orientia tsutsugamushi
O. tsutsugamushi uzročnik je šikarske groznice. O. tsutsugamushi ima topljivi antigen, koji u RVK reagira pretežno specifično za vrstu i soj. Nije dokazano da ova vrsta ima ESS. Svega 3 soja ove vrste mogu uzrokovati toksičnu smrt miša. Samo neki sojevi imaju zajedničke antigene s Proteus mirabilis sojem OX-K ( ali ne i OX-19 i OX-2 ).
Prirodni rezervoar ove rikecije različiti su divlji glodari , među kojima ju prenosi larva grinje Leptotrombidium deliense. Grinja je i vektor i rezervoar. Grinja se hrani samo u stadiju larve i ako se u to vrijeme u blizini nađe čovjek, može se zaraziti. Na mjestu uboda larve javlja se papula, koja zatim ulcerira i nastaje crna krasta ( karakteristična nekrotična primarna lezija ) . Bolest počinje naglo s povišenom
13

temperaturom, glavoboljom i bolovima u mišićima. Makulopapulozni osip prvo se javlja na trupu, a zatim se širi centrifugalno. Javlja se generalizirana limfadenopatija, splenomegalija, komplikacije CNS-a , a može doći i do zatajenja srca. Smrtnost nakon infekcije s nekim virulentnijim serotipovima može biti i do 60%. Tetraciklini i kloramfenikol djelotvorni su u liječenju akutne bolesti. O. tsutsugamushi može uspostaviti latentnu infekciju, pa se često javljaju recidivi bolesti. Zbog značajne raznolikosti sojeva , nije se uspjela napraviti djelotvorna vakcina.
EHRLICHIA
Erlihije su gram negativne , isključivo unutarstanične bakterije , koje obitavaju u fagocitnoj vakuoli limfocita, makrofaga ili polimorfnonuklearnih leukocita kičmenjaka . Primarno su bile poznate kao uzročnici bolesti u pasa, a kasnije i drugih životinja ( preživači, konji ) , pa su prema tome i dobivale imena : E. canis, E.equi, E. muris i druge. Prvi put je erlihioza čovjeka prepoznata u Japanu 1953 g.
Morfologija
Erlihije su gram negativne kokoidne vrlo pleomorfne bakterije koje se javljaju u dva oblika : kao malene stanice s gustom jezgrom i kao veće retikularne stanice. Oba se oblika Erlihija dijele diobom i u citoplazmi inficiranih leukocita stvaraju nakupine – morule, velike do 4000 nm.
Metabolizam
Erlihije imaju metabolizam sličan onome u rikecija . Razgrađuju glutamat da bi sintetizirali ATP . Ne razgrađuju glukozu.
Otpornost prema faktorima okoline
14

Erlihije su osjetljive na povišenu temperaturu, pa uzorke treba pri kratkotrajnom transportu čuvati na 4*C a pri dužem transportu na –20*C.Nakon uzgoja , stanice inficirane erlihijama mogu se čuvati mjesecima na –80*C.
Otpornost prema antimikrobnim lijekovima
Erlihije su rezistentne na peniciline, cefalosporine, aminoglikozide, makrolide i kloramfenikol. Osjetljive su na tetracikline, te na rifampicin.
Patogenost
Nakon uboda krpelja erlihije se šire limfnim i krvnim žilama i ulaze u ciljne stanice hematopoetskog i limforetikularnog sustava. Ulaskom u ciljne stanice sprječavaju spajanje fagosoma s lizosomom , pa na taj način izbjegnu razaranje. Infekcija mijenja funkciju inficiranih stanica i razara ih.
Bolest
U ljudi su opisana dva oblika erlihijoza: - HME – humana monocitna erlihijoza kod koje uzročnik E. chaffensis parazitira u makrofagima , -HGE – humana granulocitna erlihijoza kod koje uzročnik E. phagocytophila parazitira u granulocitima. Kod oba oblika bolesti javljaju se slični simptomi : povišena temperatura, glavobolja , mijalgija , osip , i kao teži oblik HME meningoencefalitis pa čak i smrt. Dokazuju se serološkim ispitivanjem porasta titra najmanje 4 puta.
COXIELLA
U rodu Coxiella postoji samo jedna vrsta . To je Coxiella burneti. Ona je uzročnik oboljenja koje se naziva Q – groznica. Tu je bolest prvi opisao Derrick 1937 g. među radnicima u jednoj mljekari u Queenslendu. Pošto nije znao kakva je to bolest , nazvao ju je Q ( od eng. riječi query- znak pitanja. )
Morfologija Coxiella burneti je kratki štapić dužine do 1 mikron. Ona se morfološki ne razlikuje od rikecija.
15

Otpornost
C. burneti je veoma otporna prema suhoći. Otporna je na povišene temperature, na 70*C živi nekoliko minuta, pa je zbog toga proširena u prirodi. Otporna je i prema korištenim koncentracijama dezinficijensa. Osjetljiva je na peniciline i tetracikline .
Patogenost Q- groznica je zoonoza . U prirodnim uslovima od te bolesti obolijevaju u prvom redu divlje životinje, zatim koze, ovce i goveda. Infekcija se među životinjama prenosi preko raznih vrsta krpelja iz porodice Ixodidae i Argasidae. Zaražene životinje obično ne pokazuju nikakve simptome bolesti . Infekcija u njih ostaje godinama , a kod tako zaraženih ženki gravidnost završava pobačajem.
Kod ljudi Q – groznica je akutno febrilno oboljenje, koje poslije inkubacije od 2-3 sedmice počinje jezom, glavoboljom, slabošću i pojačanim znojenjem. Ubrzo zatim javljaju se bolovi u grudima, lagani kašalj s oskudnom ekspektoracijom. Simptomi odgovaraju simptomima primarne pneumonije. Opisani su i slučajevi endokarditisa koji u neliječenih može završiti letalno.
Prevencija i liječenje
Rezervoar zaraze su divlje i domaće životinje kao i perad. Rezervoar su i zaraženi krpelji. Izvor zaraze su mokraća, izmet, meso i mlijeko zaraženih životinja, kao i izmet zaraženih krpelja. Čovjek se inficira prilikom udisanja kontaminirane prašine ili konzumiranjem nepasterizovanog mlijeka i nedovoljno termički obrađenog mesa zaraženih životinja. Prenos s čovjeka na čovjeka je veoma rijedak. Tetraciklini daju dobre rezultate u liječenju ovog oboljenja , a može se provoditi i specifična profilaksa vakcinom ubijenih uzročnika.
Mikrobiološka dijagnoza Dijagnoza se može postaviti bakteriološki dokazivanjem uzročnika u krvi i eventualno u sputumu bolesnika pomoću direktne imunofluorescencije. Serološki se prati porast titra najmanje 4 puta.
16

Chlamydia
Klamidije su sićušne , kokoidne , nepokretne gram- negativne bakterije koje parazitiraju u citoplazmi stanice domaćina u kojoj stvaraju karakteristične uklopine (inkluzije) , koje se mogu vidjeti svjetlosnim mikroskopom. Kao i druge bakterije klamidije imaju unutarnju i vanjsku ovojnicu sličnu onoj u gram- negativnih bakterija, sadrže RNA i DNA , prokariotske ribosome, stvaraju vlastite nukleinske kiseline, proteine i lipide, a i osjetljive su prema antimikrobnim lijekovima. Razlikuju se od ostalih mikroorganizama po svome jedinstvenom načinu razvoja i razmnožavanja. U rodu klamidija nalaze se 4 vrste : C. trachomatis, C. psittaci, C. pneumoniae i C. pecorum.
Morfologija
Klamidije se javljaju u morfološki i funkcionalno različitim oblicima, što je usko vezano s razmnožavanjem tih bakterija. Osnovno ili elementarno tjelašce (ET) okrugla je ili kruškolika čestica i ona je zapravo zarazni oblik klamidija. Ima čvrstu vanjsku stijenku što mu omogućava da može kratko preživjeti izvan stanice domaćina . Inicijalno ili retikularno tjelašce (RT) veća je , metabolički aktivna čestica , koja se aktivno dijeli ali ne može preživjeti izvan stanice domaćina pa nije zarazna. C. pneumoniae (a vjerovatno i ostale vrste) mogu se javiti u stanici kao metabolički neaktivna čestica nazvana perzistentno tjelašce (PT), koje može u neaktivnom stanju ostati dugotrajno i uspostaviti perzistentnu inf. Antibiotici i imuni sistem ne mogu djelovati na PT.
Razmnožavanje klamidija
Da bi klamidije mogle inficirati stanicu i razmnožiti se , mora se ET pričvrstiti na specifične receptore na prijemljivim stanicama domaćina . ET zatim inducira specifičnu fagocitozu ( endocitozu) u stanicama koje inače nisu fagociti, pa ET ulazi u stanicu unutar vakuole (endosoma) dobivene od stanične opne domaćina. ET specifično sprječavaju spajanje endosoma s lizosomima i do kraja svog razvoja ostaju okružene ovojnicom endosoma. Nakon nekoliko sati od ulaska u stanicu nastaju promjene u staničnoj stijenci ET . DNA se rasprši pa nestaje centralni nukleoid, započinje sinteza DNA, RNA i proteina, povećava se citoplazma s ribosomima, nastaje veća RT. RT se dijele na
17

dvoje, citoplazma vakuola se povećava i nastaju inkluzijska tjelašca karakteristična za klamidije. Nakon toga RT se prestaju dijeliti, smanjuju se , zgušnjava im se citoplazma, javlja se središnji nukleotid i nastaju tipična zrela ET koja se oslobađaju iz stanice domaćina i započinju novi ciklus infekcije.
Metabolizam
RT sintetiziraju vlastite DNA, RNA, proteine i lipide i za tu sintezu trebaju energiju. RT ne mogu stvoriti ATP pa su klamidije potpuno ovisne o energiji stanice u kojoj parazitiraju, zato su nazvane energetskim parazitima. ATP stvoren u eukariotskoj stanici domaćina ulazi u RT pomoću enzima ATP/ADP translokaze. U klamidijama nisu dokazani citohromi ni flavoproteini pa su one vjerovatno anaerobni organizmi.
Antigena struktura
Svi članovi roda Chlamydia imaju zajednički termostabilni antigen. To je topljivi lipopolisaharid (LPS) , koji se nalazi staničnom zidu i ET i RT klamidija , a možemo ga dokazati RVK, elektroforezom u gelu , enzimskim imunotestom (EIA) i interdermalnim pokusom. Ustanovljeno je da klamidije imaju u glavnom proteinu vanjske membrane i za rod karakteristični proteinski antigen koji ima važnu ulogu u nastanku imunopatolških procesa u toku klamidijske infekcije.
Termolabilni površinski polipeptidi glavnog proteina vanjske membrane ET nose epitope specifične samo za određene izolate iste vrste (serovare). Ti su antigeni važni ne samo za serološku tipizaciju već oni vjerovatno djeluju i kao faktori virulencije i imaju ključnu ulogu u stvaranju zaštitnih antitijela. Dokazuju se postupkom neutralizacije toksičnosti na mišu i mikro imunofluorescencije.
Osjetljivost na fizikalne i hemijske faktore iz okoline
Klamidije su osjetljivije od drugih bakterija pa kratko preživljavaju izvan žive stanice, i to samo kao ET. Pri sobnoj temperaturi nešto sporije gube infektivnost ako se nalaze u mediju s bjelančevinama ili ostacima stanica. Pri 56*C gube infektivnost već za 5-10 min. Dobro podnose temperaturu od –70*C pa se na toj temp. i čuvaju. Eter , etilni alkohol, jod i Na-hipoklorit u dezinficijensima uspješno uništavaju klamidije. Ustanovljeno je da C. trachomatis može ostati infektivna u kloriranoj vodi bazena i nekloriranoj vodovodnoj vodi 2-3 dana, kao i na vlažnom ručniku 45-90 min. C. psittaci može preživjeti mjesecima u perju, prašini, fecesu.
18

C. pneumoniae gubi 50% infektivnosti pri +4*C u toku 24 sata.
Otpornost prema antimikrobnim lijekovima
Tetraciklini pokazuju najbolji inhibitorni učinak pa se smatraju lijekom izbora u liječenju infekcija klamidijama. Eritromicin i azitromicin nešto slabije djeluju. Eritromicin se koristi u trudnica kada je tetraciklin kontraindiciran. Sulfonamidi djeluju inhibitorno na većinu sojeva vrste C. trachomatis, dok su sojevi vrsta C. psittaci i C. pneumoniae otporni prema tim lijekovima. Aminoglikozidi, cefalosporini, linkomicin i vankomicin ne djeluju na klamidije. Penicilin nije koristan u liječenju.
Patogeneza
Inficirani domaćin redovno stvara antitijela protiv klamidija, ali ta antitijela imaju slabu zaštitnu moć protiv klamidija smještenih u stanici, pa je stoga značajniji imunološki odgovor posredovan stanicama.Može čak imati i štetan učinak , pa su nakon ponovnog dodira s uzročnikom opažene mnogo teže kliničke slike bolesti: nastaje kasna preosjetljivost i granulomatozne upale. Tako u novije vrijeme sve više seroloških i anatomskih podataka ukazuje na moguću ulogu hroničnih infekcija s C. pneumoniae u nastanku ateroskleroze i bolesti koronarnih arterija.
Mikrobiološka dijagnostika
Izolacija klamidija. Za izolaciju klamidija primjenjujemo oplođeno kokošije jaje, laboratorijske životinje i kulturu stanica. U stanicama žumanjčane vrećice pilećeg embrija mogu se naći za rod specifični antigen LPS i karakteristične inkluzije nakon određenog vremena inkubacije. Inkluzije i ET mogu se prikazati bojenjem po Macchiavellu ili po Gimenezu. Svi sojevi C. psittaci i LGV sojevi C. trachomatis ucijepljeni mišu intracerebralno uzrokovaće infekciju. Sojevi od A do K vrste C. trachomatis kao i C. pneumoniae ne mogu inficirati miševe. Kulture stanica (McCoy, HeLa, HL i Hep-2) pokazale su se najpogodnijim za uzgoj i izolaciju klamidija iz bioloških uzoraka. Prisutnost klamidija u kulturi stanica dokazujemo bojenjem po Giemsi, Lugolovim rastvorom ili specifičnim monoklonskim antitijelima obilježenim fluorescentnom bojom.
Direktno pretraživanje uzorka. Razmasci obrisaka sluznica , koje su zaražene klamidijama , mogu se obojiti po Giemsi pa će se svjetlosnim mikroskopom vidjeti grimizno obojene inkluzije u citoplazmi stanice domaćina. Direktnom
19

imunofluorescencijom i enzimskim testom moguće je dokazati prisustvo klamidijskih antigena u uzorku dobijenom od bolesnika.
Serološka dijagnostika. Zaražene osobe obično stvaraju antitijela za antigen specifičan za rod kao i ona specifična za vrstu i serovare klamidija. Stoga se danas najviše koristi test mikroimunofluorescencije jer dokazuje za vrstu tj. za serovar specifična antitijela.
Chlamydia trachomatis
Svi sojevi ove vrste su svrstani u tri biovara : -biovar trahoma,-biovar limfogranuloma venereum(LGV)-mišiji biovar U biovaru trahoma nalaze se serovari označeni A;B,Ba i C, koji uzrokuju trahom , i serovari od D do K , koji se prenose spolnim putem i uzrokuju bolesti oka i genitalnog sistema ali i druge brojne manifestacije u djece i odraslih. U LGV biovaru nalaze se serovari L1- L3 , koji se prenose spolnim putem i uzrokuju istoimenu spolnu bolest.
TRAHOM
Trahom je hronični keratokonjuktivitis , javlja se isključivo u ljudi i može uzrokovati sljepoću. Prenosi se direktnim kontaktom sa zaraženim okom ili kontaminiranim ručnicima. C. trachomatis se razmnožava u cilindričnim stanicama konjuktive gornje vjeđe, pa bolest počinje kao akutni purulentni konjuktivitis , koji se zbog nakupljanja limfocita i stvaranja folikula ispod konjuktive razvija u hronični folikularni konjuktivitis a dolazi i do prodora krvnih žila u rožnjaču(panus). Folikule nekrotiziraju pa nastali ožiljci uvrću vjeđu i trepavice ,koje stružu i oštećuju rožnjaću. Ta mehanička oštećenja uz sekundarne bakterijske infekcije pogoduju nastanku lezija koje će dovesti do sljepoće. Gubitak vida obično se javlja 20-30 godina nakon primarne infekcije.
Prevencija i liječenje
Najbolji način prevencije trahoma je poboljšanje ličnih higijenskih navika i uvjeta , a tek nakon toga upotreba tetraciklinske masti za oči kroz duže vrijeme. Do sada napravljene vakcine protiv trahoma proizvele su samo kratkotrajni imuni odgovor.
20

Mikrobiološka dijagnostika
Za mikrobiološku dijagnostiku uzimaju se strugotine ili obrisci epitelnih stanica konjuktive gornje vjeđe. Razmasci dobivenih stanica mogu se direktno obojiti po Giemsi ili antitijelima obilježenim fluoresceinom (DIF) ili enzimom (EIA) da bi se dokazala ET ili karakteristične inkluzije. Ti se uzorci mogu ucijepiti u kulturu McCoy –stanica. Testom mikro-IF mogu se dokazati za serovar specifična antitijela u serumima i to IgM tokom prvog mjeseca a zatim IgG. Titar antitijela u suzama korelira sa intenzitetom bolesti.
INFEKCIJE SPOLNOG SISTEMA I OKA
Serovari od D do K C. trachomatis inficiraju stanice cilindričnog i prijelaznog epitela sluznica uretre, endocerviksa, endometrija, jajovoda, anorektuma, disajnih puteva i konjuktive. Obično se prenose spolnim putem. U muškaraca C. trachomatis uzrokuje negonokokni uretritis sa karakterističnim sluzavo-gnojnim iscjetkom. Ovi serovari su najčešči uzročnici akutnog epididimitisa u spolno aktivnih muškaraca. Među homoseksualcima često se javlja klamidijski proktitis. U mlađih muškaraca u toku ili nakon negonokoknog uretritisa javlja se Reiterov sindrom (uretritis, konjuktivitis, poliartritis). Čini se da su infekcije s C. trachomatis mnogo češće u žena . Klamidije se razmnožavaju u stanicama cilindričnog epitela endocerviksa, vrlo često bez klinički jasnih znakova , ali se infekcija može očitovati i kao teški folikularni cervicitis. Iz cerviksa infekcija se može širiti na endometrij i jajovode , pa može nastati akutni salpingitis. Dokazano je da neliječene klamidijske infekcije u žena mogu uzrokovati izvanmaterična začeća i neplodnost. Spolno aktivne osobe mogu sekretima genitalnog trakta prenijeti serovare D-K na konjuktive pri čemu nastaje inkluzijski konjuktivitis. Vjeruje se da je glavni način prijenosa autoinokulacija prljavim prstima , ali se može javiti i nakon kontaminacije vode sa sekretima genitalnog trakta u bazenima.
INFEKCIJE U NOVOROĐENČADI
Prolazom kroz inficirani porodni kanal novorođenčad se mogu inficirati genitalnim serovarima D-K C. trachomatis. Mnoga od te novorođenčadi prerano su rođena , klamidijska infekcija majki povezana je sa brojnim komplikacijama i za majku i za dijete. 5-12 dana nakon poroda javlja se obično na jednom oku otok donje vjeđe, hiperemija i obilan gnojni iscjedak. Neliječene infekcije mogu trajati i do 12 mjeseci a mogu se javiti i ožiljci i vaskularizacija rožnice. Uz infekciju oka može se javiti i
21

infekcija srednjeg uha, rinitis i vulvovaginitis i u 10-20% slučajeva intersticijska pneumonija koja ima za posljedicu hroničnu bolest disajnog sistema.
Prevencija i liječenje
Infekcije genitalnog sistema i inkluzijski konjuktivitis uzrokovan klamidijama mogu se spriječiti sigurnijim spolnim običajima, pravovremenim liječenjem osoba sa simptomima i njihovih spolnih partnera, kao i traganjem za asimptomnim nosiocima. Lijek izbora su tetraciklini . Očne i genitalne infekcije odraslih mogu seLiječiti s jednom dozom azitromicina per os. U novorođenčadi eritromicinske ili tetraciklinske kapi nakon poroda mogu spriječiti nastanak inkluzijskog konjuktivitisa ali ne i pneumonije , pa bi zato trebalo redovito kontrolisati trudnice i one s infekcijom liječiti eritromicinom. Za liječenje pneumonije u novorođenčadi primjenjuje se eritromicin 14-21 dan.
Mikrobiološka dijagnostika
Uzorci za mikrobiološku dijagnostiku jesu strugotine ili obrisci epitelnih stanica s konjuktive, uretre, endocervikalnog epitela, aspirati iz jajovoda, Daglasovog prostora i epididimidisa. Dijagnoza se može postaviti inokulacijom uzročnika u kulturu McCoy stanica, a inkluzije u stanicama dokazujemo bojenjem Lugolovim rastvorom. Serološke reakcije nisu značajne u dijagnostici inkluzijskog konjuktivitisa i akutnih infekcija sluznica genitalnog trakta. Serološka je dijagnostika (IFA) značajna kod pneumonije novorođenčadi jer se obično tokom bolesti nalazi visok titar IgM antitijela.
LYMPHOGRANULOMA VENEREUM (LGV)
LGV biovar C. trachomatis razlikuje se od biovara trahoma po nekim biološkim odlikama. Serovari L1,L2,L3 invazivniji su od ostalih serovara C. trachomatis. Infekcije sa serovarima su sistemske i zahvaćaju i endotelne stanice i mononuklearne makrofage, što im omogućuje širenje limfnim putevima.
Patogeneza
LGV klamidije ulaze u organizam kroz sitne ozljede na koži ili sluzokoži i šire se u područne limfne čvorove u kojima nastaju apscesi . Limfni čvor može apscedirati pa nastaju rupture, sinusi i fistule, a kasnije se javlja fibroza i granulomatozni alergijski odgovor.
22

Klinička slika
Oko 1-4 sedmice nakon spolnog dodira javlja se na mjestu ulaska primarna ozljeda – mali bezbolni ulkus ili vezikula ali brzo zacjeljuje. Nakon 1-3 sedmice (drugi stadij bolesti) područni limfni čvorovi otiču i postaju bolni. Tako uvečani gnoje se i gnoj izlazi kroz brojne kanaliće van. U tom stadiju izraženi su i opšti simptomi bolesti pa se uz povišenu temp, glavobolju, osip, hepatitis može javiti i meningitis. Limfni čvorovi će konačno zacijeliti ožiljcima ali infekcija može perzistirati i uzrokovati kasna razorna oštećenja (anogenitalni sindrom). Zbog preosjetljivosti kasnog tipa s granulomatoznim odgovorom nastaju ožiljci s posljedičnim začepljenjem limfnih puteva pa može nastati elefantijaza spolnih organa. LGV serovari mogu uzrokovati i okuloglandularni konjuktivitis.
Prevencija i liječenje
Mjere zaštite od LGV iste su kao i kod drugih bolesti koje se prenose spolnim putem. Djelotvorna vakcina do sada nije pripremljena. Lijek izbora u liječenju LGV su tetraciklini. Eritromicin i rifampicin su djelotvorni.
Mikrobiološka dijagnostika
LGV- klamidije mogu se izolirati iz primarnog ulkusa, aspirata ognojenog limfnog čvora, obriska rektuma i materijala dobivenih biopsijom iz zahvaćenih organa u kulturi neobrađenih McCoy i HeLa stanica, žumanjčanoj vrećici pilećeg embrija i u mozgu i peritoneumu miša. U serološkoj dijagnostici najviše se primjenjivala RVK. Kod te je bolesti teško dokazati značajan porast titra antitijela, jer zbog prirode bolesti bolesnik obično dolazi liječniku nakon akutnog stadija. Zato se smatra da titar viši od 1:64 uz odgovarajuću kliničku sliku ukazuje na vjerovatnu LGV infekciju. Mikro-IF je specifičniji i osjetljiviji test ali i ovim testom u LGV infekcijama je teško dokazati porast titra. Radi se i kožni test kasne preosjetljivosti – Freiev pokus, ali nije dovoljno specifičan.
Chlamydia psittaci
Sojevi C. psittaci razlikuju se od drugi dviju vrsta u rodu po antigenu specifičnom za vrstu. U inficiranoj stanici stvaraju više inkluzija, različitih oblika i veličina ali bez matriksa glikogena pa se ne mogu obojiti jodom. Prirodni domaćini C. psittaci su
23

različite ptice i sisari, kod kojih se najčešće uspostavlja asimptomatska, latentna infekcija, ali se može usljed stresa aktivirati i u ptica uzrokovati bolest psitakoza odnosno ornitoza, a u domaćih sisara pneumonije, pobačaje, artritise i enteritis. Inficirane životinje izlučuju velike količine klamidija u fecesu i slini , koje sasušene u prašini mjesecima mogu ostati žive. Ptičiji sojevi klamidija , naročito oni od papiga i purana su visoko infektivni za čovjeka. Čovjek najčešće udahne klamidije iz sasušenih ptičijih ekskreta .
Patogeneza
C. psittaci je invazivna i ima afinitet za stanice RES-a pa se lako širi i oštećuje mnoge organe uključujući pluća, bubrege, jetru, slezenu, srce i CNS. C. psittaci preko respiratornog sistema ulazi u krv i putuje u retikuloendotelne stanice jetre i slezene. U tim stanicama se umnožava i razara ih, pa nastaju nekrotična žarišta. U plućima nastaje upalni odgovor u kojem u alveolama i u intersticiju dominiraju limfociti. Nastaje zadebljanje stijenke alveola, infiltracija mononuklearnim stanicama , a javlja se i nekroza.
Klinička slika
Psitakoza se u čovjeka može očitovati kao latentna infekcija , blagi oblik sličan prehladi , kao pneumonija ilikao teška sistemska bolest , koja u neliječenih osoba može biti smrtonosna. Većina slučajeva očituje se kao atipična pneumonija . Nakon inkubacije od 1-3 sedmice bolest obično poćinje visokom temperaturom, klonulošću, fotofobijom i teškom glavoboljom. Zatim se javlja neproduktivni kašalj i simptomi pneumonije. Može se javiti i teška sistemska bolest (tifoidni oblik) sa simptomima encefalitisa, miokarditisa i hepatitisa. Smrt obično nastaje u starijih osoba zbog istovremenog zatajenja pluća, toksičnog šoka i cirkulatornog kolapsa. Nakon ozdravljenja od psitakoze osoba može ponovo oboljeti . C. psittaci može perzistirati u respiratornom traktu čovjeka godinama nakon preboljele bolesti , te se povremeno izlučivati u sputumu.
Prevencija i liječenje
Psitakoza je zoonoza koja se u ljudi najčešće javlja kao profesionalna bolest. Rezervoari zaraze za ljude su ptice s rijetkom pojavom prijenosa s čovjeka na čovjeka. Mjere za kontrolu širenja bolesti uključuju kontrolu uvoza ptica kao i iskorjenjivanje zaraze među domaćim peradima. Tetraciklini tokom 10-21 dan su lijek izbora u liječenju psitakoze, a može se upotrijebiti i eritromicin. Iako liječenje antibioticima utječe na ishod bolesti, ne mora
24

dovesti do eradikacije klamidije iz organizma, pa se može uspostaviti trajno nosilaštvo klamidije.
Mikrobiološka dijagnostika
C. psittaci možemo izolirati iz krvi uzete u ranom stadiju bolesti , iz iskašljaja (5-7 dana od početka bolesti) ili iz tkiva pluća postmortalno. Uzorci se ucjepljuju u kulturu stanica, u žumanjčanu vrećicu pilećeg embrija ili u miša. Inkluzije se mogu bojiti po Giemsi ili Macchiavellu, ali ne i Lugolovim rastvorom. Izolacija se radi samo u specijaliziranim referentnim laboratorijima zbog visoke infektivnosti. Zbog toga se psitakoza obično dijagnosticira serološkim testovima i to pomoću RVK i IFA. Titar antitijela u RVK od 1:64 ili viši u jednom serumu smatra se dijagnostički značajnim, naročito ako postoje epidemiološki podaci značajni za psitakozu. U IFA se koristi antigen specifičan za vrstu, pa dokazivanje IgM antitijela u titru 1:32 smatra se dijagnostički značajnim za psitakozu.
Chlamydia pneumoniae
Čovjek je jedini poznati domaćin C. pneumoniae. ET ove vrste je često kruškoliko, i ima širok periplazmatski prostor. Do sada je poznat samo jedan serovar ove klamidije (TWAR). Ne sadrži plazmide. Inkluzije su joj male, guste i okrugle.
Patogeneza
U respiratornom sistemu C. pneumoniae inficira i razara stanice trepetljikavog epitela respiratornog sistema. Brojna su seroepidemiolška ispitivanja ukazala na ulogu C. pneumoniae u patogenezi ateroskleroze. Budući da C. pneumoniae može inficirati stanice glatkih mišića krvnih žila, endotelne stanice koronarnih arterija i makrofage, pretpostavlja se da bolest nastaje zbog upalnog odgovora na hroničnu infekciju ovom klamidijom.
Klinička slika
Bolest se najčešće očituje kao faringitis s umjerenom temperaturom, ali se može javiti i sinusitis, bronhitis i pneumonija. Smatra se da je 5-20% izvanbolničkih pneumonija u mlađih osoba uzrokovano s C. pneumoniae. U starijih i onih s hroničnom respiratornom bolesti javlja se teška i ponekad smrtonosna atipična pneumonija.
25

Prevencija i liječenje
Tetraciklini i eritromicin djeluju na C. pneumoniae pa se mogu koristiti u liječenju infekcija. U novije vrijeme lijek izbora je azitromicin, jer dostiže višu koncentraciju u stanici, a i liječenje je kraće.
Mikrobiološka dijagnostikaU rutinskoj dijagnostici se najviše koriste serološke metode, i to RVK i IFA. U IFA se kao antigen koriste ET C. pneumoniae, pa je test specifičan za vrstu a za dokazivanje primarne infekcije prati se četverostruki porast titra antitijela bilo IgM ili IgG klase.Smatra se da bi povišeni titar IgA antitijela mogao biti biljeg za perzistentnu infekciju s C. pneumoniae.
26

27