protein profiling of milk from native nordic cattle breeds in …library.au.dk/fileadmin/ ·...
TRANSCRIPT

Master Thesis
Protein profiling of milk from native Nordic cattle breeds in
relation to technological properties
Protein profilering af mælk fra oprindelige nordiske kvæg racer i relation til
teknologiske egenskaber
Anette Kienke Rosengaard
Department of Food Science, Aarhus University
Student number 20094244

Preface
The present master thesis “Protein profiling of milk from native Nordic cattle breeds in relation
to technological properties” of 60 ECTS was part of the education “Molecular Nutrition and Food
Technology” at Aarhus University. The thesis was carried out at the Department of Food Science,
Foulum. The project was done with supervision from Lotte Bach Larsen and Nina Aagaard Poulsen.
Acknowledgement
I would like to thank my supervisors Lotte Bach Larsen and Nina Aagard Poulsen for good and
helpful supervision. Furthermore, I would like to thank laboratory technicians Hanne
Søndergaard Møller and Gitte Hald Kristiansen for experimental guidance. Thanks to Søren Drud
Nielsen for experimental guidance and good discussions of results and Guilherme de Moura
Maciel for help with statistical analysis and good discussions of results throughout the project.
Thanks to Helene Halkjær Jensen for critical revision of the thesis.
At last, thanks to my family and friends for moral support, with a special thanks to my husband
and daughter, Henrik and Clara, who always made me smile.
Aarhus University, Department of Food Science, March 21. 2016
Anette Kienke Rosengaard
20094244

Abstract
Bovine milk and milk products are important parts of human diet. The protein composition of
milk can vary depending on e.g. breed, diet and environment. Therefore, the dairy industry is
interested in identifying which factors affect the protein composition of the milk. Selecting for
special traits within breeds could promote some technological properties for special dairy
products. The milk and protein composition have therefore been an important research topic.
Both earlier and recent studies of current Scandinavian dairy milk have revealed strong relations
between milk protein genotype on one side and processing qualities on the other side. This has
been shown in milk from individual dairy cows and will, depending on genotype characteristic
and distribution of genotypes in milk at mixing, also manifest in tank milk. 69 milk samples from
two native Danish breeds, Red Danish and Jutland Cattle, and two native Swedish breeds,
Swedish Mountain and Swedish Red Polled, were characterized in milk composition and
processing qualities. By studying the protein profile and genotype variants in native Nordic
breeds, a possible difference in protein profile compared to current dairy breeds could be
observed. By LC-MS analysis, an identification of the most common casein (CN) proteins and
whey proteins, κ-CN, αs2-CN, αs1-CN, β-CN, β-lactoglobulin (β-LG) and α-lactalbumin (α-LA),
together with the protein isoform of κ-CN (glycosylated κ-CN) and αs1-CN (phosphorylated αs1-
CN), was performed. Furthermore, the total and ionic calcium content was measured, and a
subset of milk samples with different genetic variants and PTMs were collected and further
analyzed by 2-DGE. Genotype, relative protein content and calcium content were also compared
to two technological properties, rennet coagulation time and gel strength after 40 min, for
evaluation of native Nordic milk characteristics for further dairy processing.
The most frequently occurring genotype of the major milk proteins were identified as AB for κ-
CN, AA for αs2-CN, BB for αs1-CN, A1A2 for β-CN, BB for β-LG and BB α-LA. LC-MS analysis of milk
from native Nordic breeds revealed a higher frequency of β-CN F than previously reported in
Scandinavian breeds, and which have previously been identified in non-coagulating milk in Danish
Holstein-Friesian cattle. One native Nordic cattle breed, Swedish Mountain, had more suitable
properties for cheese production, compared to the other three breeds in this study. These
findings give evidence for further breeding of some native Nordic breeds to niche dairy products.

Resumé
Ko mælk og mælkeprodukter er en vigtig del af den menneskelige føde. Protein sammen-
sætningen af mælk kan variere afhængig af f.eks. race, kost og omgivelser. Mejeribranchen er
derfor interesseret i at identificere hvilke faktorer der påvirker proteinsammensætningen i mælk.
Udvælgelse af specielle træk i racerne kunne fremme nogle teknologiske egenskaber til
produktion af specielle mejeriprodukter. Mælke og protein sammensætningen har derfor været
et vigtigt forskningsemne. I både tidligere og nylig studier af mælk fra nuværende skandinaviske
køer, har vist en stærk relation mellem mælkeproteiners genotype på den ene side og
bearbejdnings kvaliteter på den anden side. Dette er set i mælk fra individuelle malkekøer og vil,
afhængig af genotype karakteristika og fordelingen af genotyper i mælken når den blandes, også
vise sig i tankmælk. 69 mælkeprøver fra to indfødte, danske racer, rød malkeko og jysk kvæg, og
to indfødte svenske racer, Fjällras og Rödkulla, blev karakteriseret for mælkesammensætning og
bearbejdnings kvaliteter. Ved at studere protein profilen og genotype varianterne i de indfødte
nordiske racer, er det muligt at kunne observere en forskel i protein profilen sammenlignet med
nuværende malkekøer. Via LC-MS analyse kan en identifikation af de mest almindelige kasein
(CN) proteiner og valle proteiner, κ-CN, αs2-CN, αs1-CN, β-CN, β-lactoglobulin (β-LG) og α-
lactalbumin (α-LA), sammen med isoform proteiner af κ-CN (glykosyleret κ-CN) and αs1-CN
(fosforyleret αs1-CN), blive udført. Udover dette blev total og ionisk Kalcium også målt, samtidig
med at et udsnit af mælkeprøver med forskellige genetiske varianter of PTMs blev samlet og
sendt til videre analyse via 2-DGE. Genotype, det relative protein indhold og calcium indhold blev
også sammenligne med to teknologiske egenskaber, rennet kogulerings tid og gel styrke efter 40
minutter, for evaluering af karakteristika i mælk fra indfødte nordiske køer, til videre
mejeribearbejdning.
Genotyperne af de større mælkeproteiner, som sås i med den højeste frekvens var AB for κ-CN,
AA for αs2-CN, BB for αs1-CN, A1A2 for β-CN, BB for β-LG og BB α-LA. LC-MS analyse af mælken fra
indfødte nordiske racer afslørede en højere frekvens af β-CN F I forhold til tidligere rapporteret I
de skandinaviske racer, og som tidligere er identificeret i ikke-koagulerende mælk fra danske
Holstein-Friesian køer. En indfødt nordiske ko-race, Fjällras, havde flere gavnlige egenskaber til

osteproduktion, sammenlignet med de tre andre racer i dette forsøg. Disse resultater giver
evidens for videre avl af nogle af de indfødte nordiske racer til niche mejeri produkter.

TABLE OF CONTENTS
1 Introduction .......................................................................................................................................... 1
Major milk proteins ....................................................................................................................... 1
1.1.1 The caseins ............................................................................................................................ 1
1.1.2 Whey ..................................................................................................................................... 6
Influence of protein profile on coagulation properties ................................................................ 8
Native nordic cattle ....................................................................................................................... 9
2 Project background ............................................................................................................................. 12
Objective ..................................................................................................................................... 14
3 Theoretic method background ........................................................................................................... 15
Liquid chromatography – mass spectrometry (LC-MS) .............................................................. 16
2D gel electrophoresis ................................................................................................................ 23
Total and Ionic calcium ............................................................................................................... 26
4 Materials and laboratory methods ..................................................................................................... 27
Milk collection ............................................................................................................................. 27
Milk protein profile by LC/ESI-MS ............................................................................................... 27
2-DGE .......................................................................................................................................... 29
total calcium ................................................................................................................................ 29
Ionic calcium ............................................................................................................................... 30
5 Statistical analysis ............................................................................................................................... 31
6 Results ................................................................................................................................................. 32
Protein profiling .......................................................................................................................... 32
Variation in milk protein profile between breeds ...................................................................... 40
Variation in milk protein profile between protein genetic variants ........................................... 42
Coagulation properties of milk from native Nordic cattle breeds .............................................. 46
7 Discussion ............................................................................................................................................ 52
Protein profiling .......................................................................................................................... 52
Variation in milk protein profile between breeds. ..................................................................... 55
Variation in milk protein profile between genetic variants ........................................................ 58
Coagulation properties of milk from native Nordic cattle breeds .............................................. 61
8 Conclusion ........................................................................................................................................... 65
9 Further perspectives ........................................................................................................................... 67

10 Reference ........................................................................................................................................ 67

1
1 INTRODUCTION
Milk is a fluid secreted from the mammary glands. In addition to being the main source of
nutrition for offspring, a large part of human diet is based on milk and milk products. Most of this
milk originates from cattle, but also mammals like goats, sheep and buffalos are milked to be
used for industrial products. However, as all milk discussed in the present thesis is bovine, it will
simply be referred to as milk. Since the earliest time, milk has been a part of the human diet, and
it has been of major importance for human nutrition and survival in some areas of the world.
Milk contains all nutrition necessary to sustain life of newborn offspring. It is composed of
approximately 87 % water, 3.2 % protein, 3.9 % fat, 4.6 % lactose, 2.6 % casein, 0.6 % whey and
0.7 % minerals and has a pH of 6.6-6.75 when it is fresh. However, the composition of the milk
can vary according to breed, genetic variation, feed composition, season, climate, lactation stage
and health (Belitz et al. 2009; Walstra et al. 2006).
MAJOR MILK PROTEINS
Milk can be divided into two major protein groups; caseins (CN) and whey proteins in the ratio of
approx. 80:20. CN and whey proteins are characterized by their stability, as CN proteins are
precipitated from raw milk at pH 4.6 at 20°C, and whey proteins exist as soluble proteins in the
serum phase. According to the homology of their primary structure, the CN proteins can be
divided into κ-, β-, αs1- and αs2-CN. The whey proteins consist mainly of β-lactoglobulin (β-LG) and
α-lactalbumin (α-LA). There are different genetic variants within each group, and every protein
variant is assign with a letter symbol based on their chemical difference from the reference
protein, and not based on differences in the DNA sequence (Farrell et al. 2004).
1.1.1 The caseins
Caseins (CN) are the predominant protein components in milk (80%). They have relatively low
levels of secondary and tertiary structure, and are characterized as relatively small proteins. CN
are to a high degree hydrophobic, but also contain some hydrophilic regions. These regions have

2
a quite high charge and contains many proline and few cysteine residues (Belitz et al. 2009). CN
can have four different forms which can be separated in the four groups: κ-, β-, αs1- and αs2-CN
in the approximate proportions of 4:10:11:3. Each form has a unique own amino acid
composition, which can also have different genetic variations and functional properties. Due to
the hydrophobic nature of CN in a hydrophilic phase, the molecule readily form hydrophobic
bonds by associating with other CN or self-association. CN mainly associate with other CN
molecules and form casein micelles in the milk together with colloidal calcium phosphate (CCP)
(Walstra et al. 2006).
The caseins are encoded by four genes mapped on chromosome 6 in a tightly linked 250 kb
cluster, also referred to as the CN locus. The four genes are placed in the physical order CSN1S1
(αs1-CN), CSN2 (β-CN), CSN1S2 (αs2-CN) and CSN3 (κ-CN) (Gallinat et al. 2013; Caroli et al. 2009).
In all four genes, genetic variance can occur. This means that e.g. in the coding genes of CSN2,
the genetic code can vary in one or more codons, resulting in more than one genetic variant for
β-CN. These codon changes may or may not be silent, leading to variance in the amino acid
composition of the β-CN protein, thus giving a range of different variants. However, not all
protein genetic variants varies in hydrophobicity or charge, and can thereby be difficult to
separate from each other (Walstra et al. 2006). Throughout this thesis, proteins with such
changes in amino acid composition are referred to as protein genetic variants. Several genetic
regions influence the transcription of CN genes and thus the amounts of protein products. First,
the transcription of the CN genes is regulated by a promoter region in front of the CN cluster,
from where the transcription process begins. Besides this region, the transcription of the genes
is influenced by other regions as well, among these non-coding regions, but also other CN loci
(Graml et al. 2003). This more complex as it is depended on the genetic variance in the gene, and
different regulatory sites also can be connected to differences in protein expression level. In
general, the CSN1S1 and CSN2 loci have reported to affect the synthesis of all caseins (Graml et
al. 2003).
The CN loci code for proteins mostly composed of 169-209 amino acid residues, depending on
the protein group and genetic variant. They have a molecular weight between 20-25 kDa. An
important aspect of CN biochemistry is posttranslational modifications (PTM). Phosphorylation

3
happens in all four CN groups, which is a characteristic not observed in the whey proteins. CN
proteins are thereby phosphoproteins, containing approx. 0.85% phosphorus, with the highest
degree of phosphorylations in αs2, ranging αs2 > αs1 > β > κ. The phosphate groups (organic
phosphate) are for the most part esterified to the hydroxyl group of serine. The number of
phosphate groups in the molecule are denoted with a P, e.g. αs2-12P-CN. Near the pH of milk
they are largely ionized, contributing to a negative charge of CN (Fox & Mcsweeney 1998).
Glycosylation happens only in κ-CN and can have from zero to six glycosylations, which are mainly
attached to threonine residues in in the hydrophilic C-terminal region. αs2-CN and κ-CN also
contain two cysteine residues per molecule, which form intermolecular disulphide bridges
(Holland et al. 2006; Fox & Mcsweeney 1998). Phosphorylation and glycosylation of CN will
further on be referred to as CN isoforms.
The protein expression level is influenced by many factors, where different breeds have reported
to have different protein contents (Gustavsson et al. 2014a; Jensen et al. 2012a; Hanne B Jensen
et al. 2015). The protein content is also affected by the genotypic profile of the proteins, with
correlation between the individual genotypes (Graml et al. 2003).
κ-casein (κ-CN)
κ-CN makes up about 9% of the proteins in milk (Belitz et al. 2009). To date, 12 different κ-CN
protein variants (A, B, C, D, E, F1, F2, G1, G2, H, I, and J) and one synonymous variant (AI) have been
identified. The most common variants are A and B. κ-CN A-1P is often used as reference protein
and has a molecular weight of 19,043 Da and a 169 amino acid residue long peptide chain (Caroli
et al. 2009).
The monomer contains two cysteine residues and most often one phosphoserine. In milk, it exist
as monomers or in association with other κ-CN as oligomers bound by disulphide bridges. κ-CN
is the only casein containing varying amounts and forms of carbohydrates. The C-terminal region
of κ-CN is hydrophilic and contains four threonine residues, which are subject for esterification
with a carbohydrate group. The levels of attached carbohydrates can vary from zero to six, and
about two thirds of the κ-CN molecules are glycosylated. The C-terminal region is hydrolyzed by
rennet during cheese making. This cleaved peptide is called the casein macropeptide (CMP). The

4
CMP contains the four threonine residues, and CMP is therefore often glycosylated (Walstra et
al. 2006; Fox & Mcsweeney 1998; Holland et al. 2006).
The content of κ-CN is dependent on the genetic variance in the gene, where the CSN3 locus
seems to account for very little of the variance in the κ-CN content. The κ-CN content seems to
be dominated by other regions in the DNA, with a clear influence from the CSN2 locus (Graml et
al. 2003). Two variants of β-CN (A1 and A2, see below) have also been correlated with altered κ-
CN content in milk. In addition, the genotype BB of κ-CN has been associated with higher κ-CN
and more glycosylated κ-CN content. The genotype of β-LG also has an effect on κ-CN content,
with a lower content in milk with β-LG genotype AA compared to genotype BB (Bobe et al. 1999;
Heck et al. 2009; H B Jensen et al. 2015).
αs1-Casein (αs1-CN)
There are to date identified nine different αs1-CN protein variants (A, B, C, D, E, F, G, H and I). The
most common variant is B (Caroli et al. 2009), and it is therefore often referred to as the reference
variant. αs1-CN B-8P is composed of a peptide chain consisting of 199 amino acid residues with
eight phosphoserines in the sequence. The molecular weight of the protein is 23,615 Da, and the
protein contributes to about 34% of the proteins in milk (Belitz et al. 2009; Caroli et al. 2009).
αs1-CN is one of the more heavily phosphorylated CN proteins, with the most common
phosphorylation degree on eight residues, but also a phosphorylation degree on seven and nine
is present in the milk (Heck et al. 2008).
A higher αs1-CN content has been associated with a heterozygote genotype. The major part of
variance in αs1-CN content has been related to CSN1S1 loci, which is less affected by other regions
in the DNA, but more affected by the genetic variance of the gene (Graml et al. 2003). The αs1-
CN genotype has, on the other hand, been associated with the protein expression β-LG (NG-Kwai-
Hang et al. 1987).
αs2-Casein (αs2-CN)
Four different αs2-CN protein variants (A, B, C and D) have been identified. The most common
variant is A. This variant is therefore often the reference protein. αs2-CN A-11P has a molecular

5
weight on 25,228 Da and a peptide chain of 207 amino acid residues. It is the most heavily
phosphorylated CN, containing from ten to thirteen phosphate groups. The most common
number of phosphate groups in αs2-CN is eleven. It contains two cysteines, resulting in
intermolecular disulfide bridges and exists as a monomer or dimer. αs2-CN is the most hydrophilic
CN and contributes to about 8% of the proteins in milk (Belitz et al. 2009; Caroli et al. 2009; Fox
& Mcsweeney 1998; Walstra et al. 2006; Farrell et al. 2004).
The content of αs2-CN in relation to genotype and regulation of transcription is not very deeply
studied. However, the content of αs2-CN seems to be affected by the genetic variance in the
CSN1S1 and CSN2 loci (Graml et al. 2003).
β-Casein (β-CN)
There has to date been identified 13 different β-CN protein variants (A1, A2, A3, B, B2, C, D, E, F,
G, H1, H2 and I). The most common variant is A2 and it is the common reference protein. β-CN A2-
5P has a molecular weight on 23,983 Da and a peptide chain consisting of 209 amino acid residues
(Caroli, Chessa, and Erhardt 2009). It contains five serine phosphorylation sites, which are mostly
phosphorylated. β-CN is the most hydrophobic CN and contributes to about 25% of the proteins
in milk (Belitz et al. 2009).
The CSN2 locus accounts for the major part of the β-CN protein variance. A heterozygote
genotype seems to result in a higher β-CN protein content, compared to a homozygote (Graml
et al. 2003). Variance in protein content is associated with β-CN variant A1, A2 and B, where A1 is
associated with a lower β-CN content compared to A2 and B (Heck et al. 2009; Heck et al. 2008).
Casein micelles and milk calcium
Most of the CN proteins in milk associate to form casein micelles, which are small droplet-like
structures. The exact structure of the CN micelle is still to debate, where several models is
proposed (Horne 2006), however the main features are agreed upon. The CN micelles are held
together with colloidal calcium phosphate (CCP). The CN micelle surface is mainly composed of
κ-CN, which acts as an emulsifier with hydrophilic C-terminal region of κ-CN in the milk serum
and the hydrophobic region inside the micelle. The size of the CN micelles ranging from <50 nm

6
to >500 nm, which have been reported to depend on factors such as κ-CN content and genotype.
Small micelle size is associated with high κ-CN content and genotype κ-CN BB. The phosphate
groups on CN have strong calcium binding properties and are some of the chemical forces that
cause CN proteins to associate in CN micelles. The phosphorylated serine residues in the CN
peptide chain bind to the calcium in CCP and form a phosphoserine cluster between all the CN
proteins, which are the building blocks of CN micelles. The CCP thereby plays a key role in the
formation and stability of the CN micelles (Gaucheron et al. 2005; Walstra et al. 2006; Fox &
Mcsweeney 1998).
The calcium in the CPP complex represents about 70% of all calcium in milk under normal pH and
room temperature. Calcium in the micelle as CCP is often referred to as micellar calcium, where
the remaining 30% calcium in milk is present in the serum phase, either as calcium complexes
with e.g. phosphate or citrate, or as free calcium, referred to as ionic calcium. Thus, these two
calcium groups, the micellar calcium and serum calcium, make up for the total calcium content
in milk. The total calcium concentration has been reported to vary according to protein
concentration. With a high protein concentration in milk, a higher level of phosphoserine
interactions with CCP occur. Moreover, the serum phosphate level and CCP level are positively
associated. (Gaucheron et al. 2005; Bijl et al. 2013).
Due to the important role of CCP in micelle stability, various studies about the role of CCP in milk
for further processing have been reported. Bovine milk contains approx. 30mM of total calcium,
where approx. 20mM exist as micellar calcium and 10mM exist as serum calcium. In raw milk
with normal pH, less than 10% of the total calcium is present as ionic calcium. Nevertheless, the
ionic calcium concentration has shown a major role in milk protein stability, rennetability, heat
stability and rheological properties (Lewis 2011).
1.1.2 Whey
Whey proteins are defined as the proteins which do not precipitate at pH 4.6. Most whey proteins
are globular proteins with compactly folded peptide chains. They are relatively hydrophobic with
a more compact hydrophobic core of the folded protein than caseins. They account for about

7
20% of the total protein content in milk. The main content of whey proteins is β-lactoglobulin
and α-lactalbumin, but also many proteins from the bloodstream in the udder diffuse into milk,
are categorized as whey proteins. These include e.g. bovine serum albumin, immunoglobulins
and several enzymes (Belitz et al. 2009; Walstra et al. 2006). The present study focuses only on
the major whey proteins, β-lactoglobulin and α-lactalbumin, which also are the two proteins
referred to when using the term whey.
β-lactoglobulin and α-lactalbumin are mapped on two different chromosomes, BLG on
chromosome 11 and LAA on chromosome 5, respectively. The gene coded for two small proteins
with 123-162 amino acid residues and a molecular weight of 14-18 kDa. Different genetic variants
exist in both groups, with one or more amino acid residue change per protein genetic variant
(Walstra et al. 2006; Caroli et al. 2009). The whey proteins are not phosphorylated and
glycosylated as the CN proteins, and β-lactoglobulin and α-lactalbumin are present in milk in the
ratio 10:4, respectively (O’Donnell et al. 2004)
β-Lactoglobulin (β-LG)
β-LG is the main whey protein. There has to date been identified 11 different genetic variants of
β-LG (A, B, C, D, E, F, G, H, I, J and W). The most common variants are A, B, and E (Caroli et al.
2009). The reference protein, β-LG B, has a molecular weight of 18,281 Da representing a peptide
chain of 162 amino acid residues. The protein contains five cysteine residues, which form two
intermolecular disulfide bonds. The last cysteine is free and buried within the structure. Thus, β-
LG is present as a dimer in milk at normal pH (Walstra et al. 2006).
The gene expression of β-LG is affected to a high degree of the BLG locus itself. Genetic variance
of the gene is reported to be the main source for changes in the β-LG protein content, with a
higher expression of β-LG A than β-LG B (Graml et al. 2003, Heck et al. 2009).
α-Lactalbumin (α-LA)
Three different protein variants (A, B and C) of α-LA have been identified. The most common is
α-LA B (Caroli et al. 2009). α-LA B has a molecular weight of 14,186 Da and a peptide chain of 123

8
amino acid residues. It contains eight cysteine residues and the protein has a biological function
as the B subunit of the enzyme lactose synthetase. (Belitz et al. 2009).
The protein expression level of α-LA is not deeply studied and do not seems to be influenced by
β-LG genotype or αs1-, κ-, β-CN haplotype (Hallén et al. 2008).
INFLUENCE OF PROTEIN PROFILE ON COAGULATION PROPERTIES
Variation in milk composition is well studied according to various factors such as breed
differences, milk protein genetic variant polymorphisms, and environmental factors. Milk
composition affects both the technological properties of the milk and the nutritional value.
Therefore, the detailed protein profile is important for the processability and functionality of
different dairy products, e.g. cheese, yoghurt, or milk powder (Walstra et al. 2006).
Coagulation properties such as curd firmness and rennet coagulation time are related to casein
amount and composition. As described above, the caseins are heterogeneous because of genetic
polymorphisms and posttranslational modifications. Thus, the protein composition in milk is to a
large extent determined by different genetic factors. Other factors such as season, lactation state,
parity, feeding and health status of the udder (somatic cell count, SCC) are other influencing
factors for milk protein composition (Walstra et al. 2006). Milk coagulation properties are
heritable and thus vary from breed to breed (Bittante et al. 2012). Extensive breeding programs,
which have resulted in large increases in milk yields from dairy cows over the years, have focused
more on fat content than milk composition. When working with a specific breed of cattle, it is
therefore important to characterize the protein composition.
Rennet coagulation is the critical step before the gelation initiates in cheese production. The milk
coagulation properties is important for both the quality and yield of the cheese (Belitz et al. 2009).
It is of great importance for the dairy industry to produce as good coagulating milk as possible.
Milk with a low rennet coagulation time (RCT), high curd firming rate (CFR) and high curd firmness
value (G’) are favorable during cheese production (Bittante et al. 2012). Many factors influence
the milk coagulation properties. These factors include the concentration of total CN, calcium, pH,

9
breed and genetic polymorphism of the milk proteins (Walstra et al. 2006). Cheese produced
from κ-CN BB milk has shown superior rennet coagulation properties compared to cheese from
κ-CN AA milk, resulting in a higher fat recoveries into the cheese due to the smaller average casein
micelle diameter in κ-CN BB milk (Walsh et al. 1998).
However, several studies evaluate the amount of poorly and non-coagulating milk, which results
in a weak or no gel formation (Wedholm et al. 2006; Frederiksen et al. 2011; Jensen et al. 2012a).
The properties and terms “poorly” or “non-coagulating” milk have no standard definition,
especially due to different methods for assessment of coagulation properties (Bittante et al.
2012). All studies, however, agree that some cows produce milk with poorer coagulations
properties, and therefore are less suited for further technological processing like cheese
production. Cheese making properties have been reported to be improved by a high casein
content of αs1-CN, β-CN and κ-CN (Wedholm et al. 2006). Jensen et al. (2012a) studied
coagulation properties in two Danish cattle breeds, according to protein genetic variants and
isoforms. The good coagulating milk was observed to have a different milk protein composition
compared to poorly coagulating milk. This was in correlation with the presence of different
genetic variants of the major milk proteins, with a prevalence of the B variant of κ- β-CN and β-
LG in good coagulating milk, and the poorly coagulating milk were associated with κ-CN A/E, β-
CN A2 and β-LG A/C. These results indicate a possibility to alter the milk composition by breeding
strategies for cattle containing the B variant of κ- β-CN and β-LG.
NATIVE NORDIC CATTLE
Denmark and Sweden have sixteen endangered native cattle breed, where five are classified as
extinct. Of these, six breeds are critically endangered, and suitable breeding and management
strategies are needed to prevent extinction of these breeds (FAO 2007). Saving genetic resources
of native livestock breeds is an important aim worldwide and is of great cultural importance. The
Food and Agricultural Organization of the United Nations (FAO) supports management and
utilization of natural resources, including genetic resources, for the benefit of the future
generations by maintaining population number (FAO). An investigation of Nordic native cattle

10
breeds can reveal if specific good production traits are available and thereby promote higher
awareness of the breed and prevent a population fall in these endangered cattle breeds. Most
living animals are mainly used locally as nursing cows for landscape protection and only a minor
fraction is being milked.
One of the ways to protect and keep native cattle breeds is to create a new use for them. Creation
of unique milk products from specific breeds may give an economic platform to put focus on
these breeds. In the present study, four breeds were studied to identify potential differences
between them. This could lead to development of niche products and thereby help conserving
them by increase the economic incitement for breeding of native Nordic cattle. Molecular
characterization suggest that the breeds are genetically different (Kantanen et al. 2000). This can
be reflected in a different milk composition between the four breeds, which also has been
showed in other studies of milk composition between breeds (Poulsen et al. 2013).
Red Danish Cattle (RDM)
In the 19th Century the Danish cattle “Ø-cattle” was crossed with several Red breeds. This
founded the “Red Danish Cattle”, which officially became a breed in 1878. Through pure breeding,
a uniformly red dairy breed with the highest milk yield from Danish cattle was developed. The
breed was exported to many countries until 1970, and in the 1950s 70% of all Danish cattle was
RDM. In 1970 the pure breeding principle was removed due to inbreeding problems. Since then,
RDM has been crossed with American brown cattle and other European red and brown breeds in
the modernization of the dairy industry (“Kvæg - NaturErhvervstyrelsen”; Sørensen and Nielsen
2012). Throughout this thesis, the term RDM is referring to RDM-1970, the pure breed cattle.
RDM is a dairy breed, but with good meat production qualities. Milk production per year per cow
is 6669 kg, with 4.78% fat and 3.70% protein. The population in 2007 was 21 breeding bulls and
186 heifers older than two years and the Danish conservation committee has preserved a stock
of RDM-1970 sperm which helps prevention of inbreeding (“Kvæg - NaturErhvervstyrelsen”;
Sørensen and Nielsen 2012).

11
Jutland Cattle (JUT)
In Jutland the original black- and gray-pied cattle was used throughout the 17th -19th Centuries,
with both dairy cattle qualities and meat cattle qualities. In 1881, the first herdbook was
published with 48 bulls and 53 heifers on the Jutland cattle SJM (K. Kortegaard 2004). The breed
was developed to dairy cattle through the first part of the 20th Century and crossbreeding with
Dutch black pied cattle became popular to obtain larger animals and higher milk production.
From 1949 SJM was renamed to SDM (black-pied Danish dairy breed), and insemination with
semen from SDM bulls replaced SJM bulls. The last registered purebred SJM bull lived in 1955
and the breed now only exists as part of SDM. However, a few farmers did not use Dutch cattle
bulls, but bulls from their own farms and a small group still looks like the original SJM. (Sørensen
and Nielsen 2012, K. Kortegaard 2004). The Danish Genetic Resources Committee has supported
the preservation of SJM since 1987. The meat has good meat production qualities with more
intramuscular fat than normal beef, which make it more tasty and tender. Milk production per
year is 6267 kg per cow, with 3.88% fat and 3.29% protein. The population in 2007 was 37
breeding bulls and 181 heifers older than two years (Sørensen & Nielsen 2012). Denmark contains
four main SJM herds, Kortegaard, Westergaard, Oregaard and Vesterboelle, together with some
smaller herds. Several studies have investigated the SJM breed through the Kortegaard herd
(Pertoldi et al. 2014; Brüniche-Olsen et al. 2012). The Kortegaard herd has been closed for several
generations, and evidence for inbreeding and very low levels of genetic variability have been
observed (Pertoldi et al. 2014). Several studies have investigated SJM and there is still big
inconsistency as to whether the remaining SJM herds in Denmark are purebred or not. The Danish
Genetic Resource committee recognize SJM as a closed genetic founder line, but these are
managed under Jutland cattle (JUT). The cows in the current project are not purebred SJM and
are thus referred to as JUT throughout this thesis.
Swedish Red Polled (SRP)
The SRP cattle is an old dairy cattle breed from Sweden, which has been low in population
number for a long time. In 1937, SRP breed was collected with Swedish Mountain cattle under a
new name, Svensk Kullig Boskap, but breeders were still separating in breeds (Bett et al. 2010).

12
This resulted in only one remaining SRP herd left in the 1970’s with 23 animals (Oklahoma Animal
Science Department). East-Norwegian Red Polled Cattle was used at foreign genetic breeding
material to save the breed. SRP varies in color from yellowish red to brown, and are used as
landscape protection in Sweden.
The females stand is 117 to 123 cm, with a weigh of 350-450 kg. The milk yield per year is up to
5500 kg (Oklahoma Animal Science Department).
Swedish Mountain (SM)
The SM breed was established by the end of the 19th century and is related to the Norwegian
breed “nordlandsfe”. Crossbreeding of the SM almost led to elimination of the oure breed in the
1980’s. By the use of frozen SM semen, an increase in the genetic size of the breed has increased
the population number since the 1990’s. The population in 1998 in Sweden was estimated to be
1000 breeding cows (Oklahoma Animal Science Department). The average milk yield and cow
weight have increased since the establishment of the breed. In 1893 the yield was 1200-1400 kg
milk per year, with a weight of the cow on 300-350 kg. Today, a SM cow can have a milk yield of
11000-12000 kg per year and the cows weigh about 450 kg. The female stand is 125 cm, with a
varying color of white with black or red spots (Oklahoma Animal Science Department).
2 PROJECT BACKGROUND
Throughout the last century, native Nordic cattle breeds have been diluted through
crossbreeding. This has been done to obtain high yielding dairy cattle, developed to fit the
modern industry with high milk yield and high milk fat content. Few living descendants of native
Nordic breeds exist today as endangered breeds, used as nursing cows and for landscape
protection, where only a minor fraction is being milked.
Molecular characterization of North European cattle breeds suggests that they are still
genetically different from modern dairy breeds. The present work of Kantanen et al. (2000)

13
reports an influence of breeding on the genetic composition of milk proteins, which have been
observed as a difference in allele frequencies between three commercial North European dairy
cattle breeds, compared to fifteen native North European cattle breeds. Moreover, Lien et al.
(1999) saw a high variance in milk protein allele frequencies between seventeen native Nordic
cattle breeds and five commercial North European dairy cattle breeds. However, the differences
in relative protein content between native Nordic cattle species were not studied. The four native
Nordic cattle breeds investigated in this thesis, Jutland cattle, Red Danish dairy cattle, Swedish
Mountain cattle and Swedish Red Polled cattle, have their origins in different evolutionary
lineages (Kantanen et al. 2000). Thus, these four breeds each represent different linages and have
genetically different background. This leads to a possible difference in milk protein composition
in the native Nordic breeds, compared to high yielding commercial dairy cattle breeds.
The protein content and composition is of major importance to the dairy industry. Here, the
casein profile of milk is important during cheese making, with a fundamental role in renneting
and ripening processes (Walstra et al. 2006). Research showed considerable differences in milk
composition and casein profile of the milk between cattle breeds. This can to some part be
ascribed to the frequencies of different genetic variants of the milk proteins between breeds,
where different genotypes have different transcriptional factors, as described above in section
2.1 (Gustavsson et al. 2014a; Poulsen et al. 2013; Wedholm et al. 2006). Recent studies have
provided new insight about coagulation characteristics correlation to protein isoforms of casein
in milk, which is the modification of casein by phosphorylation and glycosylation.
Therefore, a distinct milk and protein composition between native Nordic cattle breeds can alter
the technological properties of the milk. Favorable changes in technological properties can
contribute to development of niche-dairy products, e.g. yoghurt, cheese or other fermented
products. Development of niche-dairy products from native Nordic cattle can contribute to an
increase in population, which help preventing extinction of the native Nordic cattle breeds.

14
OBJECTIVE The hypothesis behind this thesis is that native Nordic cattle can have an altered milk protein
profile than modern dairy cattle, given the unique genetic background of the native Nordic
breeds (Kantanen et al. 2000). This can be manifested in both structure of the proteins due to
mutations and in the protein composition due to different expression levels of the various protein
genetic variants. Such variation might be beneficial for technological purposes. This can be used
in specific dairy products like drinking milk, butter, cheese or fermented dairy products.
Therefore, the objective of this master thesis was to study the protein profile of milk from native
nordic breeds, including protein genetic variants and post translational modifications.
To investigate this hypothesis, skim milk collected from four native Nordic cattle breeds were
studied by LC/ESI-MS (Figure 1). With a newly modified LC/ESI-MS method (Frederiksen et al.
2011), protein genetic variants could be more accurately identified and the impact of these on
technological properties were be studied. The proteins were also analyzed using complementary
proteomic techniques (2-DGE) when needed. LC/ESI-MS allowed identification of the milk protein
profile and relative protein content of κ-, αs- and β-CN, β-LG and α-LA. From these data, the
frequencies of the protein variants and variation in protein isoforms (glycosylation proportion of
κ-CN and phosphorylation proportion of αs1-CN) were identified. A comparison of the major
protein profile was performed with respect to breed and κ-, and β-CN genotypes.
This master thesis is part of a bigger investigation of milk originating from native Nordic breeds.
Collection of milk samples, analysis of rennet induced coagulation properties and overall milk
composition were already obtained on the samples (Figure 1, upper part). This was used to
compare results from this thesis and to discuss tecnological properties of the milk together with
calcium concentration measurements. Together, this allowed evaluation of the protein profile in
milk of the four investigated cattle breeds to use for futhere development of dairy products.

15
Figure 1 The design of the project “Technological properties and milk composition of native Nordic cattle”. The techniques within the box were performed during the preparation of in this thesis. JUT = Jutland cattle. RDM = Red Danish dairy cattle. SM = Swedish
3 THEORETIC METHOD BACKGROUND
This Master thesis was a part of an on-going project called “Technological properties and milk
composition of native Nordic cattle”. The project was initiated to preserve the native Nordic
cattle breeds, by creating a niche product from some of the breeds, thereby increasing the
population. Milk composition and milk coagulation properties were obtained by Milkoscan (milk
composition) and by ReoRox4 and Stresstech (milk coagulation properties). Milk protein profile

16
was obtained by LC-MS, and further investigation in the milk protein profile was performed with
2-DGE. Moreover, an expansion of the milk composition was made by measuring the total and
ionic calcium concentration (Figure 1, boxed area).
LC coupled to ESI-MS was used for genotype determination and quantification of the relative
protein content of the most common casein and whey proteins. This method has previously been
used to calculate the relative protein content in milk and genotyping of the major milk proteins
(V Bonfatti, Chiarot, & Carnier, 2014; Jensen et al., 2012a; Hanne B Jensen et al., 2015;
Frederiksen et al. 2011).
LIQUID CHROMATOGRAPHY – MASS SPECTROMETRY (LC-MS)
One of the methods of choice for analysis of complex protein samples is chromatography
followed by mass spectrometry. LC-MS can provide information on individual compounds in a
complex mixture as milk, including identification by average mass and quantification. By coupling
the LC-MS systems, minimum sample handling prevents sample loss and ensures maximal
efficiency of the analysis.. Every compartment of LC-MS are reviewed separately in the following
section.
Liquid chromatography (LC)
Liniquid chromatography (LC) is a widely used method for molecule separation. It is capable of
utilizing a number of separation principles, allowing separation of molecules based on several
different physico-chemical properties. LC is often coupled to high pressure (HPLC) and is generally
used with different separation techniques, e.g. size exclusion, affinity, hydrophobic interaction,
ion exchange and reversed-phase (Food Analysis by HPLC, 2012; O’Donnell, Holland, Deeth, &
Alewood, 2004). The LC column separation mechanism used in this study was reverse-phase (RP).
For this type of chromatography, a stationary phase of the column surface is hydrophobic and
complemented with a hydrophilic mobile phase. The column is made of silica and the surface is
covered with varying lengths of hydrophobic alkyl chains (C4-C18). This retains hydrophobic
compounds from the sample, while hydrophilic compounds will pass on through the column in

17
the mobile phase. By decreasing the polarity of the mobile phase, compounds elute in order of
their hydrophobicity, with elution of the most hydrophobic compounds last, if they elute at all
(Anon 2012). The columns used within this study were designed to analyze and separate intact
proteins with an Mw > 10,000 kDa. Moreover, they had high pH stability and were very good for
separation of highly hydrophobic proteins.
After elution of the column, the protein concentration of the fractions was estimated using UV
absorption. The area of each peak on the UV chromatogram refers to the intensity of peptide
bonds, corresponding to the amount of peptide bonds present. This can be used to estimate the
ratio between the different proteins. The higher the peak intensity, the higher the relative
protein content. No absolute concentration of each protein present in the milk can be calculated,
if no standard curve is made beforehand.
Milk proteins have a distinct pattern of elution that to a large degree reflects their hydrophobicity.
The solvent phase is most hydrophilic at the beginning of a sample run, and all the most
hydrophilic proteins, here glycosylated κ-CN, elute as the first proteins in milk, whereas
hydrophobic molecules such as the whey proteins, elute when at a more highly hydrophobic
solvent. The CN proteins have hydrophilic regions, but are to a high degree hydrophobic, with
several hydrophobic regions. The most hydrophilic CN is κ-CN, due to its C-terminal end with
hydrophilic regions and glycosylations, which contribute to the hydrophilicity. From the CN
proteins it elutes first, followed by the non-glycosylated κ-CN, as the solvent becomes more and
more hydrophobic (Fox & Mcsweeney 1998). Together, separation of intact milk proteins by RP-
HLPC reveals an elution pattern in a UV chromatogram in the following pattern:
Glycosylated κ-CN κ-CN αs2-CN αs1-CN β-CN β-LG (B) α-LA β-LG (A).
Due to a high resolution of the HPLC technique, differentiation between most of the various
intact protein genetic variants of both casein and whey proteins is possible. A few of the protein
genetic variants have the same hydrophobicity and cannot be separated by HPLC, but can be
separated by Mw in MS (denoted with * in Table 1). Glycosylation of κ-CN and phosphorylation
of αs1- and αs2-CN changes the hydrophobicity of the proteins and a separate peak with
glycosylated κ-CN appears in the chromatogram (A/B/E-G, Figure 2). Moreover, the

18
phosphorylation degree of αs1- and αs2-CN, where a higher phosphorylation degree makes the
protein more hydrophilic (denoted with B-8P and B-9P for αs1-CN and A-11P and A-12P for αs2-
CN, Figure 2).
The time it takes from application of the sample to the column until it elutes, is referred to as the
retention time. The retention time depends on the solvent gradient used during a selected time
range. However, the proteins still elute in the same order, which is termed the protein retention
pattern. The retention pattern is dependent on both differences between genetic variants and
degree of phosphorylation, and this affects the hydrophobicity. A retention pattern for protein
genetic variants of all proteins has been mapped in other studies. The retention times found by
Jensen et al (2012a) are noted in Table 1; however, as retention times depend on the timing of
the entire elution the retention pattern is of more interest for comparisons. The retention
pattern of protein genetic variants identified in Danish Holstein-Friesian was:
κ-CN A/B/E Glyco κ-CN A/E 1P κ-CN B 1P αs2-CN A 11P αs2-CN A 12P
αs1-CN B/C 8P αs1-CN B/C 9P β-CN B 5P β-CN A1 5P β-CN A2/I 5P β-CN F 5P β-LG
B α-LA B β-LG A
Selected protein genetic variants in Table 1 are in accordance with recorded genetic variant in a
current commercial dairy breed, Danish Holstein-Friesian cattle (Jensen et al., 2012a). Not all
known protein variants have been found within the Danish Holstein-Friesian breed. Identified
genetic variants and protein isoforms (κ-CN glycosylation and αs-CN phosphorylation) in Danish
Holstein-Friesian are illustrated in Figure 22, where the retention pattern is illustrated in an UV
chromatogram. β-CN F is reported in Holstein-Friesian, but is not in the chromatogram obtained
by Jensen et al. (2012a). The β-CN retention after β-CN A2/I is therefore added with an arrow.

19
A shift in retention time of milk proteins between cattle breeds has been shown by Jensen et al.
(2012a), between Holstein-Friesian and Jersey cows. Jersey milk proteins elute before Holstein-
Friesian milk proteins (e.g. Jersey αs1–CN retention time: 38-41 compared to 41-43.5 min from
Holstein-Friesian). This can be explained as differences in protein content can shift the retention
time a little. A small shift in retention time seems thereby possible between breeds (Jensen et al.
2012a).
Figure 2 Identification of protein genetic variants and isoforms of major milk proteins (horizontal text) in Holstein-Friesian. Identified milk proteins in vertical text. G= glycosylation, P= phosphorylation. Arrow with β-CN F retention time, found within the same study by Jensen. Modified from Jensen et al. (2012a).
β-C
N F
-5P

20
Protein1 Genetic variant and amino
acid substitution2
Mw3 modified /
unmodified
pI4 HPLC
retention
time5
Mutation of base pair6
κ-CN
169 aa
A: reference variant
B: Thr136 Ile; Asp148 Ala
E: Ser155 Gly
19,034 (1P)
19,002 (1P)
19,004 (1P)
5.93
6.34
5.93
10*
16
10*
-
ACC ATC, GAT CCT
AGC GGC
αs2-CN
207 aa
A: reference variant
25,228 (11P)
8.34
21-23.5
-
αs1-CN
199 aa
B: reference variant
C: Glu192 Gly
23,615 (8P)
23,542 (8P)
4.91
4.96
41-43.5*
41-43.5*
-
GAA GGA
β-CN
209 aa
A1: Pro67 His
A2: reference variant
B: Pro67 His; Ser122 Arg
F: Pro67 His, Pro152 Leu
I: Met93 Leu
24,023 (5P)
23,983 (5P)
24,092 (5P)
24,039 (5P)
23,965 (5P)
5.24
5.13
5.38
5.24
5.13
53
54.5*
50
57.5
54.5*
CCT CAT
-
CCT CAT, AGC AGG
CCT CAT, CCT CTT
AGT CTG
α-LA
123 aa
B: reference variant / 14,186 4.80 68 -
β-LG
162 aa
A: Gly64 Asp; Ala118 Val
B: reference variant
/ 18,367
/ 18,281
4.76
4.83
70
65
GGT GAT, GCC GTC
-
Table 1 Summarize of the identified genetic variants of the major milk proteins in Danish Holstein-Friesian cattle (Jensen et al.,
2012a), with amino acid substitution, calculated molecular weight, theoretical pI and identified HPLC retention.
*Proteins which co-elute within the same protein group. 1Protein name, number of amino acids in the mature reference protein. Mature protein = protein without signal peptide. 2Information about amino acid deletion or substitution in the genetic variant compared to the reference protein variant (Caroli
et al. 2009; Farrell et al. 2004). 3Average molecular weight (Mw; Da) of the mature protein with or without noted modifications. Calculated by ExPASy using
‘Compute pI/Mw’ (www.expasy.org) and by Farrell et al. 2004. 4Isoelectric point (pI) of the mature protein without modifications. Calculated by ExPASy using ‘Compute pI/Mw’
(www.expasy.org). 5Retention time (min) of peaks observed during HPLC separation (Abs 214 nm) of milk proteins from Holstein-Friesian cows, total
time for one sample run is 80 min (Jensen et al., 2012a). 6Information about the gene base pair mutation in positions according to the mature protein in each genetic variants in Bos Genus
according to reference variant (Caroli et al. 2009).
The HPLC retention pattern of milk protein variants has been identified in several studies as
summarized in Table 2. Also, glycosylation content of κ-CN and phosphorylation degree of αs1-

21
and αs2-CN can be detected by HPLC and have been investigated in several studies (Table 2). The
α-LA genotype has not been very intensively studied, where the protein content has been
observed, with no further instigation in genotype. β-CN F was identified by Visser et al. (1995),
and only Jensen et al. (2012a) have documented the presence of β-CN F with HPLC. The
information of β-CN F in North European cattle is therefore very low.
κ-CN* αs2-CN* αs1-CN* β-CN*
β-LG* α-LA
Genetic variant
Reference
Glyco A B E A Phos B C Phos B A1 A2 F I A B B
Visser et al.
(1995)**
√ √ √ √ √ √ √ √ √
Bobe et al.
(1998)**
√ √ √ √ √ √ √ √
Bonfatti et al.
(2008)***
√ √ √ √ √ √ √ √ √ √ (√) √ √ √ √
Frederiksen et
al. (2011)***
√ √ √ √ √ √ √ √ √ √
Jensen et al.
(2012a)***
√ √ √ √ √ √ √ √ √ √ √ √ √ √ √ √
Day et al.
(2015)****
√ √ √ √ √ √ √ √ √ √ √ √ √
Table 2 Overview of literature detecting genetic variant in milk by HPLC technique.
*All investigated protein variants is in accordance to genetic variants in Danish cattle found by Jensen et al. (2012a).
**Measured absorbance in HPLC at 220 nm
***Measured absorbance in HPLC at 214 nm
****Measured absorbance in HPLC at 280 nm
Glyco = glycosylated κ-CN. Phos = phosphorylated α-CN
After proteins were separated by LC they were introduced into the mass spectrometer. This can
be done with different techniques. In the present study ESI was used.

22
Electrospray ionization (ESI)
To be able to analyze protein content of a sample, it must be in gas phase. Electrospray ionization
(ESI) generates sufficiently high amounts of intact ionized proteins from liquid to gas phase,
without containing solvent, for detection and measurement the protein in MS. ESI is a soft
ionization technique, well suited for ionization of proteins and peptides, and the least invasive of
all ionization methods. ESI is based on pumping the solution containing the proteins from the
HPLC column over an electric field. This is done through a small metallic needle carrying a large
voltage. The voltage causes the proteins to become charged. Fine droplets containing solvent
and electrically charged proteins are then accelerated towards the opening of the needle, driven
by an electrical field between needle and plate. The force of the electrical field produces
gradually smaller droplets as the solvent evaporates from the droplets, concentrating the
multiply charged ions. Individual ions are separated and enters the MS (Berg et al. 2007; Georgiou
& Danezis 2015).
Mass spectrometry (MS):
Mass spectrometry (MS) is a powerful technique to identify and characterize the average mass
of an investigated component. If the compound can be transferred into gas phase (volatilizing)
and ionized, it can be measured by MS. MS measures the average mass of a compound by
separating and measuring mass-to-charge ratio (m/z). The result has form of a mass spectrum,
showing the signals which are proportional to the number of ions, as a function of m/z ratio (Berg
et al. 2007).
The mass analyzer used in this study was a single quadrupole. A quadrupole consists of four
parallel, cylindrical rods arranged in a square. The quadrupole filters the sample ions, they are
introduced in gas phase from the ESI, and directed down the center of the square. The rods of
the quadrupole create oscillating electric fields. This determines which m/z ratios of ions can pass
through the filter at a given time. Ions that do not have the correct m/z ratio will collide with rods.
Ions are thereby separated by their stability through the rods under a given voltage, and only
ions of a certain m/z will reach the detector. During scanning mode of the mass analyzer, a range

23
of m/z ratios are monitored, providing the qualitative information of the complete range of m/z
ratios detected in the sample. Moreover, from all the ions reaching the MS detector, the relative
abundance of each ion is calculated, resulting in a MS chromatogram, based on the ion intensity
and the time point at which the ion reaches the ESI from the LC separation. The time separation
by LC ensures separation of the molecules before reaching MS. This results in individual peaks in
MS chromatogram, containing the relative abundance of the ions in the peak, and by calculation,
the Mw of the detected molecules can be found (Georgiou & Danezis 2015; Kruve et al. 2015).
The Mw of the major milk protein genetic variants are listed in Table 1. Every peak in an MS
chromatogram contains one or more average masses of the protein(s) in the peaks. To detect the
intact protein genetic variant in the peak, a comparison with Mw and HPLC retention time of
known protein genetic variants can determine the origin of the protein(s) within the peak.
2D GEL ELECTROPHORESIS
Gel electrophoresis is a widely used method to separate biological macromolecules as protein
and nucleic acid, according to their net charge in an electrical field. The macromolecules are
placed in a gel, and by applying an electrical field they will move inside the gel. The velocity of
migration is depended of pore size of the gel, the electric field strength and the net charge of the
protein. Small proteins move readily through the gel, whereas larger proteins are more immobile
in the gel. The pores size are determined by the ratio of polyacrylamide:bis-acrylamide (Berg et
al. 2007).
For some purposes, the proteins can be separated in two dimensions to obtain a much higher
resolution, as the proteins can be separated both by their basis net charge and by their basis
molecular mass. This is called two dimensional gel electrophoresis (2-DGE). In other words, it is
the combination of two methods; isoelectric focusing (IEF) in the first dimension and sodium
dodecyl sulfate - polyacrylamide gel electrophoresis (SDS-PAGE) in the second
dimension(O’Donnell et al. 2004).
IEF separates proteins on the basis of their relative content of acidic and basic residues, summed
up as the proteins net charge. The pH where net charge is zero, is the proteins isoelectric point

24
(pI). IEF takes place on an immobilized pH gradient (IPG strips) where molecules move toward
the cathode or anode, until they reach their pI position within the pH gradient. Each protein will
move until it reaches the position where pH =pI of the protein. Proteins can readily be resolved
down to as small difference in pI as 0.01 (Berg et al. 2007). In SDS-PAGE, charged proteins are
separated on basis of their mass in an electrical field, under denaturing condition in a
polyacrylamide gel. By dissolving the proteins in a solution of the anionic sodium dodecyl sulfate
(SDS) and dithiothreitol (DTE), a disruption of nearly all noncovalent interactions in the native
protein and reduction of disulfide bonds will occur, respectively. The SDS-denatured protein
complexes have a homogeneous net charge, which added by SDS and proportional to the protein
size. Thereby, all proteins have a mobility in the gel towards the anode, with velocities dependent
on their Mw. Proteins with high Mw move slowly through the gel against the anode, while proteins
with low Mw moves faster towards the anode (Berg et al. 2007).

25
2-DGE is often used on complex mixtures, since the two used parameters are not
connected, so a uniform distribution of protein spots are often obtained. 2-DGE is
therefore often used on milk, due to only little changes in charge per protein genetic
variant, which can be detected by this method (Figure 3). The six major proteins in
milk is separated by 2-DGE, where clear spots is observed for all the six proteins.
The genetic variant is also separated as observed in figure 3, where e.g. β-LG and β-
CN is positioned in two spots. PTM degree on the caseins is also separated by 2-DGE, as seen with
κ-CN and αs2-CN. Milk protein genetic variants position in the first dimension of the gels depends
on pI, which is listed in Table 1. Two genetic variants with the same pI have same spot position
in the gel, where the change of molecular weight is too small for detection by 2-DGE. Distribution
of the major milk proteins is seen in figure 4, with spot detection by Holland, Deeth, and Alewood
(2004). The overview gel represents all the heavy caseins in the upper part of the gel and the
lighter whey caseins in the lower part of the gel.
Jensen, Holland and colleagues (2012) have identified the genetic variants position on the 2-D
gel from Danish Holstein-Friesian and Jersey cows. A separation of three dominant β-CN spots
Figure 3. 2-DGE of bovine milk proteins by Holland, Deeth, and Alewood (2004). Done by a pH 4-7 IPG strip (horizontal axes). Apparent MW on the vertical axes. Boxes indicate area with the major milk proteins. αS1-CSN = αs1-casein. αS2-CSN = αs2-casein. β-CSN = β-casein. κ-CSN = κ-casein. α-Lac = α-lactalbumin. β-Lg = β-lactoglobulin.
Figure 4 β-CN genetic variants in 2-DGE. pI on horizontal axes. Numbered spots is further identified in the article by Jensen, Holland et al. (2012)

26
with a pI range of 4.9 to 5.1 (β-CN variant A1, A2, B, I), seen in figure 4. Due to an identical pI, β-
CN variant A2 and variant I has the same spot position. Variant F was previous analyzed on 2-DGE
and was defined to have a spot position equal to A1 (Hanne B. Jensen, IMCG Workshop 2014,
unpublished data).
TOTAL AND IONIC CALCIUM
Ionic calcium
Ionic calcium is important for stabilisation, heat stability, rennetability and rheologocal
properties of milk proteins (Lewis 2011). Therefore, measurement of ionic calcium (Ca2+, free
calcium) in milk and milk products gives an understanding in the important role of calcium in milk
during further processing. The Ca2+ concentration in milk is very changeable, and small changes
in temperature, pH or addition of solutions to the milk can change the equilibrium between
insoluble calcium phosphate and Ca2+. There are a number of different known techniques to
measure ionic calcium, which have been roughly reviewed by Micheal J. Lewis (2011). Ion
selective electrodes have been used widely and compared within the different types (Lin et al.
2006). A calcium electrode consists of a thin membrane which allows permeation of only calcium
ions. Calcium ions are transported from a high concentration to a low concentration through the
selective binding within the membrane. This transport creates a potential difference, measured
by an electrode. This is compared to reference electrode of constant potential. Standard
solutions of CaCl2 with certain ionic strength are used as reference, so a concentration of an
unknown sample can be calculated through the electrode potential measured by the calcium
sensitive electrode.
Total calcium
Total calcium content in milk have reported to vary according to casein content of the milk. The
casein content have been increased during the last 75 years, together with the total calcium
content in the milk (Bijl et al. 2013). Total calcium content have been investigated in relation to

27
coagulation properties of the milk, where high total calcium content was associated with better
coagulation properties (Gustavsson et al. 2014b)
Potentiometric titration on Calcium use two electrodes, a Ca2+ ion-selective electrode and a silver
reference electrode. EGTA is used as a titrant, because it form a complex with calcium, which can
be detected by the Ca2+ ion-selective electrode (Methrom 2016).
The calcium content in % (wCa) is calculated as follow:
Where VEP1 is titrant consumption until the first equivalence point in ml, cEGTA is concentration of
EGTA in mol/L, f is titer of EGTA, MCa is Molecular weight of calcium, 49.078 g/mol, 0.1 is
conversing factor, which is all divided by ms, the sample size in gram (Methrom 2016).
4 MATERIALS AND LABORATORY METHODS
MILK COLLECTION
Milk samples from 69 cows were collected - 12 Jutland cattle, 28 Red Danish dairy cattle, 23
Swedish Mountain cattle and 8 Swedish Red Polled cattle by Gitte Hald Kristiansen. Milk was
collected from six different herds having native Nordic cows being milked in Denmark and
Sweden. One farm contained all the SRP, one all the SM, one with JUT, two with RDM and one
with RDM and JUT.
MILK PROTEIN PROFILE BY LC/ESI-MS
Analyses using LC/ESI-MS were performed with basis in Bonfatti et al. (2008) and modifications
from Frederiksen et al. (2011). The protein analysis was performed by a LC-MS system consisting
of the Agilent 1100 Series RP-HPLC equipment coupled with Agilent 100 LC/MSD equipment
(Agilent Technologies, Santa Clara, CA, USA). The RP-HPLC 1100 system consisted of a Jupiter C4
column (250 mm x 2 mm, 5µM particle size, 300 Å pores; Phenomenex, USA) and a diode array

28
detector (G1315B) with UV detection at 214 and 280 nm. ESI-single Q mass selective detector
coupled to RP-HPLC.
Prior to LC-MS analysis, skim milk samples were thawed in the refrigerator overnight before every
experiment. Milk samples were mixed in a 1:3 ratio with a reduction buffer of 6M Guanidine
hydrochloride (GdnHCl), 100mM Bis-Tris pH 6.8, 5.37mM Sodium-citrate dihydrate (Na-Citrate)
and to which a final concentration of DTE of approximately 15 mM was added. Samples were
incubated on a rocking platform at 37°C for 60 minutes and then centrifuged at 14000 x g for 10
minutes at 7°C. Before analysis, all samples were filtered through a 0.2-µm
polytetrafluoroethylene filter (Mini-UniPrep, Whatman, Florham Park, NJ, USA). Bulk milk from
Arla (Arla 24, “minimælk”), was used as a reference, sampled in duplicates every time a new set
of samples were prepared.
Protein separation was conducted using a binary solvent system with trifluoroacetic acid and
acetonitrile in a linear gradient. The HPLC equipment operated with a mixture of solvent A,
consisting of 0.05% trifluoroacetic acid (TFA) in miliQ water, and solvent B consisting of 0.05%
TFA in 99,95% acetonitrile (ACN). The column was equilibrated with 32% solvent B before sample
injection. For protein analysis, 10 µL sample was injected to the column and operated at 40°C.
Immendiately after sample injection, a linear gradient was initiated, from 32-44% solvent B in 80
min and from 44-32% solvent B in 1 min. A flow rate of 0.3 mL/min was used. The LC column was
cleaned between every sample with injection of 6M guanidine hydrochloride to remove
impurities, with a gradient from 32-90% B for 1 min, 90% B for 10 min, 90-32% B for 1 min and
finally, 32% B for 25 min. When eluted from the column, the sample protein content was
quantified by measuring UV absorbance at 214 nm, coupled with an ESI-MS. The samples were
analyzed in duplicate. The MS settings were as follows: Capillary voltage: 4,500 V; nitrogen drying
gas flow: 12 L/min; nitrogen drying gas temperature: 320°C; quadrupole temperature 99°C.
Data analysis from LC-MS was done using the deconvolution algorithm of the ChemStation
software (Rev. B.04.02 SP1 [208], Agilent Technologies). The average isotopic masses (Mw) of the
compounds were calculated from the total ion content. All relative protein quantification was
manually integrated for each protein genetic variant and protein isoform in ChemStation.

29
2-DGE The used 2-DGE method was based on the work by Jensen et al. (2012b).
50 µg skim milk samples were solubilized in a 1:10 ratio, corresponding to the milk protein
concentration in average milk which is 3.5 g protein/100 mL. For samples with altered protein
content, the dilution was adjusted to obtain the same final concentraion. The samples were
solubilized in lysis buffer, containing 7M Urea, 2M Thiourea, 1% DTE, 2% CHAPS, 2% Pharmalyte
(pH 3-10) and 40mM Tris Base pH 7.5 in miliQ water and allowed to lyse on a rocking platform at
RT for one hour. They were then centrifuged at 4°C, 14000 x g for 20 minutes. For isoelectric
focusing, the samples were added onto 11cm strips with immobilized pH gradient (IPG) (pH 3-
6)(Bio-Rad, Hercules, CA, USA) and passively rehydrated at room temperature, before focusing.
Isoelectric focusing was performed with a step gradient with constant current, 1mA, and power,
5W. The voltage gradient was 200 V for 5 hrs, from 200 – 500 V in 1 hr, 500 V for 2 hrs, from 500-
3500 V in 1 hr and 3500 V for 15 hrs.
The IPG strips were equilibrated for 15 min in a reducing equilibration buffer (6M Urea, 30%
Glycerol, 2% SDS, 0.05% Tris-HCl pH 8.8 and 65mM DTE). This was followed by a washing step in
an alkylating buffer (6M Urea, 30% Glycerol, 2% SDS, 0.05% Tris-HCl pH 8.8 and 270mM
Iodoacetamide), for 15 min. SDS-PAGE was performed on Criterion precast gels, 8-16% Tris-HCl
(Bio-Rad, Hercules, CA, USA) at 200 V for 35 min.
The gels were stained with Colloidal Coomassie Blue G-250 (CCB) containing 5% CCB, 10% ethanol,
2% phosphoric acid and 5% aluminium sulphate. They were destained in 1% acetic acid.
TOTAL CALCIUM
Frozen milk samples were thawed at room temperature, and aliquots of 20 mL were acidified to
pH 4.3 with 0.5 M HCl, followed by centrifugation at 3500 x g for 5 min. For each sample, about
2 grams of supernatant was transferred to titration tubes and weighed. Hereafter, 10 mL 0.1 M
borax buffer (Na2B4O7*10 H2O in miliQ water) and 80 mL miliQ water were added into the
tubes. Measurement of the total calcium concentration was performed using an automated

30
potentiometric titrator (Metrohm 862, Metrohm, Herisau, Switzerland), coupled with a double-
junction Ag/AgCl reference electrode (LL ISE Reference 6.0750.100, Metrohm, Switzerland) and
a Ca2+ ion-selective electrode (scION Ca Tip, 6.0509.000, Metrohm, Switzerland). Titration was
performed using a 0.01 M EGTA- 0.1 N KOH solution. Duplicates were made for each sample.
The titrator setup and method was provided by the manufacturer (Metrohm, 2016).
IONIC CALCIUM Measurements of ionic calcium concentration was based on Koutina et al. (2015) with a calcium
ion selective electrode using a calibrated standard curve (range from 0.1 – 40 mM). Ionic calcium
was measured in skim milk by a direct measurement with the ion-selective electrode Ca2+ Ion
Meter (HORIBA).
Samples were thawed from -18 °C for 24 hours at 4°C. Before measuring, the samples were
incubated in a water bath for 1 hour at 30°C to restore the calcium balance after the refrigeration,
and thereafter incubated at room temperature for 30 minutes before measuring.
HORIBA was calibrated with HORIBA standard solution, 150 ppm and 2000 ppm. Before sample
measurement, the calcium calibration curve was measured, using standard solutions of CaCl2 (0.1,
1.0, 10 and 32 mM) in 80 mM NaCl dissolved in miliQ water, as a background electrolyte. 300 µL
standard solution or sample were used to each measurement. Duplicates were made for each
sample. Between every measurement, HORIBA membrane was washed with miliQ water. All
measurements were done in electrode potential (pCa values for calcium, mV) and during
measurements of all samples the mV was noted when it reached an equilibrium after approx. 30
seconds. Determination of free calcium contents were done according to a standard curve, using
the linear relationship (Nernst equation) between the electrode potential (mV) measured in the
calibration solutions and the corresponding pCa value (electrode potential of Ca electrode, pCa
= -log [Ca2+]) measured in the milk (Koutina et al. 2015).
Ionic calcium measurements were also performed with a pH-adjusted standard curve, with the
same standard solutions of CaCl2 (0.1, 1.0, 10 and 32 mmol/L) with 80 mmol/L NaCl as
electrolyte background (Koutina et al. 2015), adjusted with 0.01 M NaOH to pH 6.75. Due to a
lower pH of the standard solutions (approx. 6.35), compared to milk (approx. pH 6.75), a pH-

31
adjusted and non pH-adjusted standard curve was made as described above, to test for pH
stability of HORIBA at both pH.
According to Lewis (2011), the output of a stable calcium ion selective electrode will change
about 29 mV for every 10-fold dilution of the standard solutions. The non pH-adjusted standard
curve had a change in mV closer to 29 mV for 0.1-1mM, and a little higher difference in 1-
10mM compared to pH-adjusted standard curve (data not shown). Due to the higher difference
from 0.1-1mM in pH-adjusted standard curve, all ionic calcium measurements was performed
with a non-pH adjusted standard curve.
5 STATISTICAL ANALYSIS
Relative protein content was quantified for all high abundance casein and whey proteins. In
addition, the specific protein isoforms covering glycosylated- and non-glycosylated-κ-CN (κ-Glyco
and κ-NonGlyco, respectively) and eight and nine attached phosphorylations αs1-CN (αs1-8P and
αs1-9P, respectively) were also quantified. Moreover, the glycosylation degree (GD) of κ-CN was
defined as κ-Glyco/κ-CN total. The phosphorylation degree (PD) of αs1-CN was defined as αs1-8P/
αs1-CN total. Calcium was quantified in total and ionic calcium fractions. Absolute protein
concentration, rennet coagulation time (RCT) and gel strength at 40 minutes of coagulation
(G’40) were measured in another study (ref) (coagulation factors). To explore variation between
breeds in the data set, Principal Component Analysis (PCA) of milk samples was made using
SIMCA software (SIMCA version 14, Umetrics AB, Umeå, Sweden). Relative protein content, GD
of κ-CN, PD of αs1-CN, κ-Glyco/κ-CN total, αs1-8P/ αs1-CN total, total protein, total and ionic
calcium, RCT and G’40 were used as X-variables.
One-way analysis of variance (ANOVA) was used to compare means between breeds, κ-CN and
β-CN to the average relative protein content or coagulation factors. The results of all analysis
were presented as the mean ± standard error. Pairwise comparisons for all tests were done by
Tukey’s test to detect significance levels of at least 95% (p<0.05) for each compared data set. All
tests were performed in R (version 3.2.1; http://www.r-project.org).

32
6 RESULTS
The protein content and distribution in milk is important for its technological properties and
handling in further dairy processing. Previous studies have shown that the genotype of casein
and whey proteins may influence the distribution of the protein composition. To further study
this, individual cows of different breeds, Jutland (JUT), Red Danish (RDM), Swedish Mountain
(SM) and Swedish Red Polled (SRP), were genotyped through determination of the protein
genetic variant in the milk and protein composition of the milk was measured and compared to
the genotyping.
The macro components and rennet induced coagulation properties of the milk samples used in
the present study were analyzed by Reorox/Stresstech and Milkoscan before freezing (Figure 1)
and used to compare with the results obtained in the present study.
PROTEIN PROFILING
To study how genotype is linked to protein composition in milk from native Nordic cattle, 69 cows
were analyzed to determine the genotype and the protein profile of their milk. As a way to study
the variation between animals, the milk of 69 cows was analyzed with respect to the six main
proteins: κ-, αs2-, αs1- and β-CN, β-LG and α-LA. These proteins are in high abundance in milk and
are known to exist as various protein genetic variants. The identification of protein genetic
variants of the high abundance milk proteins in native Nordic cattle were carried out by LC-MS.
UV in LC measured at 214 and 280 nm. A chromatogram obtained at 280 nm measures aromatic
amino acids. The results from 214 nm were used to monitor the amout peptide bonds, and thus
profile all present peptides. All peaks in MS chromatograms were listed and average mass
calculated and compared to all known intact casein, β-LG and α-LA protein masses. Protein
genetic variants identified in Danish Holstein-Friesian were the expected outcome of protein
genetic variants in native Nordic breeds (Table 1), and when in doubt, deeper investigation of
protein genetic variants was performed with inspiration from Caroli et al. (2009). LC-MS
identified the intact protein variants within each of the six main proteins in the 69 investigated

33
milk samples. This was based on retention time and Mw (Table 1). The retention pattern according
to this method was (Figure 5):
Glycosylated κ-CN κ-CN αs2-CN αs1-CN β-CN β-LG (B) α-LA β-LG (A)
As shown in Figure 5, none of the six major proteins eluted within the same time range. Thereby,
based on the retention time, each peak within a certain elution interval was preconceived to
belong to a certain protein group, as illustrated in Figure 5. For example, if the peak eluted in the
elution interval of κ-CN, then the average mass was compared to all known κ-CN genetic variants.
All protein genetic variants were identified manually. All samples were run in duplicate, and the
genotype was determined for both duplicate to make sure no sample was misread.
Figure 5 MS chromatogram of a milk sample with elution interval for the six major milk proteins. Elution interval is covered by a horizontal clamp { . The main protein in each elution interval is noted above the clamp. The vertical numbers give the retention times of the peaks. In the κ-CN elution interval, glycosylated κ-CN eluted first after approx. 10 minutes. Not all protein genetic variants were observed here. Genotype of sample: κ-CN BB, αs2-CN AA, αs1-CN BB, β-CN A1A2, β-LG BB and α-LA BB.
Validation of protein variants by 2-DGE
Apart from the general retention pattern, a number of samples had deviating chromatograms,
where a number of smaller peaks were identified besides the major ones. These peaks appeared
with low intensities in the chromatogram within the elution interval of κ- and β-CN, and generally
had low peak height and narrow peak width, compared to the major proteins peaks identified
within the sample (example in Figure 6). Generally, masses of these small peaks in the κ-CN
elution interval were identical with the molecular mass of a known protein variant of κ-CN and
small peaks within the β-CN elution interval were identical with the molecular mass of a known
κ-CN
αs2-CN
αs1-CN β-CN
α-LA β-LG B β-LG A

34
protein variant of β-CN. All samples were run in duplicates and in samples with smaller, low
intensity peaks, these peaks were observed in both of the sample runs. However, for correct
calculation of relative protein content these peaks should be correctly assigned and were
therefore further investigated. That is, samples with low intensity proteins in the κ- and β-CN
elution interval were examined to clarify the origin of the low intensity protein in the peak. Two
examples of low intensity proteins within β-CN elution interval are described below and
illustrated in Figure 6 by an LC chromatogram (sample no. 1228 and 7305).
Figure 6 Comparison of four different β-CN retention patterns in LC chromatogram. Elution interval 55-67 min. Investigated samples were sample no.1228 (β-CN A2A2) and 7305 (β-CN A2F) together with two normal samples as standards, STD.A2F (β-CN A2F) and STD.A1F (β-CN A1F). Standard sample peak retention time and peak width were in accordance with all sample runs in this trail. Peak height varied due to differences in relative protein content of the protein variants between the samples. Black arrow indicates and unknown peak in 1228, orange arrow indicates an unknown peak in 7305, and these were subject for further analysis.
Overall, it was observed that seventeen milk samples had smaller, low intensity peaks in the κ-
and β-CN elution interval, with a low peak height and narrow peak width compared to the protein
variant in the milk (data not shown). Two samples were selected to illustrate extra peaks in β-CN
elution interval (55-67 min. Figure 6). The two arrows (orange and black) indicate the peaks of
interest, which lead to further investigation of the milk samples. Two samples with genotype β-
CN A2F (STD.A2F is black) and A1F (STD.A1F is red) showed the expected retention time and peak
width.
0
50
100
150
200
250
55 57 59 61 63 65 67
UV
21
4 n
m (
mA
U)
Time (min)
β-CN elution pattern in LC
STD. A2F STD. A1F 1228 7305
β-C
N
A1 β-C
N
A2
β-C
N
F

35
Sample no. 1228 (blue) was identified as genotype β-CN A2A2. The small, low intensity peak of
investigation eluted after β-CN A2 (black arrow), which had a low peak height and narrow peak
width compared to the standard protein genetic variant peaks (black and red curve) and
compared to the β-CN protein variant in the sample. The retention time was within the range of
β-CN F (62.5-64 min.), and with a calculated mass equal to variant F (molecular mass in Table 1).
No other masses could be calculated in the additional low peaks/humps within the sample.
Sample no. 7305 (green) was identified as genotype β-CN A2F. The small unknown peak of
investigation eluted before β-CN A2 (orange arrow), which had a low peak height and narrow
peak width compared to the standard protein genetic variant peaks (black and red curve) and
compared to the β-CN protein variant in the sample. The retention time was within the range of
β-CN A1 (58-60 min.) and with a calculated mass equal to variant A1 (Table 1).
To further characterize these compounds, all seventeen samples were subjected to 2D gel
electrophoresis (2-DGE).
2-DGE is expected to identify the main protein variants in the milk as clear
spots, due to protein accumulation at the proteins isoelectric point (pH =
pI). For previously identified 2-DGE spot positions of β-CN, the reader is
referred to section 3.2, Figure 4. Due to the inability of 2-DGE to separate
and identify low abundance proteins, proteins in high amounts in the milk
will appear as clear spots in the gel, whereas proteins in low amounts in
the milk will appear as shadows on the gel. If proteins from the small, low
intensity peaks in sample 1228 and 7305 are a β-CN protein, and are
present in a relative high amount in the milk, a spot for the respective
protein will appear on the gels.
The zoom in areas of β-CN spot position in the gel, pI 4.8 - 5.1, of sample no. 1228 and 7305, are
shown in Figure 7. From the data presented in Figure 7 it was concluded that both samples
contained β-CN A2 with a clear spot around pI 4.9. No spots were further observed in the upper
panel, sample no. 1228. These results were in compliance with LC-MS results. One more clear
spot was observed in the lower panel, sample no. 7305, around pI 5.0. β-CN A1 and F have the
Figure 7. Spot position of β-CN, zoom area of pI 4.8-5.1 within 2-DGE. 1228 = Upper box with phenotype β-CN A2A2. 7305 = lower box with phenotype β-CN A2F.
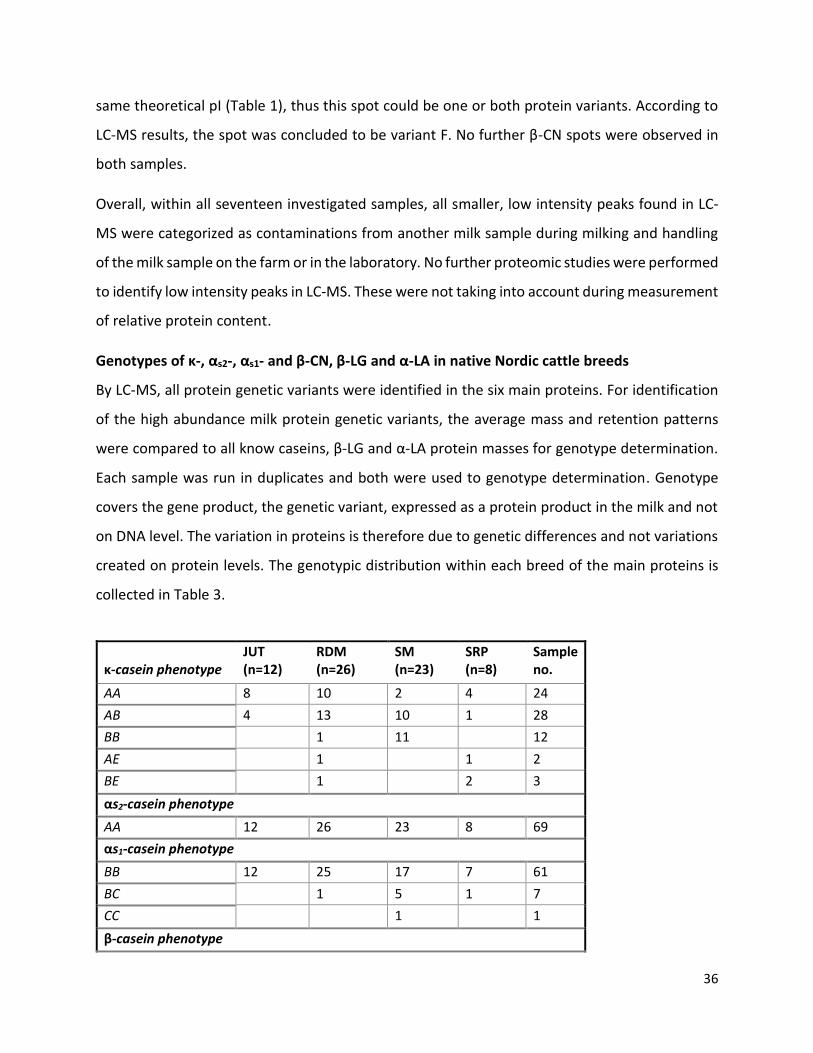
36
same theoretical pI (Table 1), thus this spot could be one or both protein variants. According to
LC-MS results, the spot was concluded to be variant F. No further β-CN spots were observed in
both samples.
Overall, within all seventeen investigated samples, all smaller, low intensity peaks found in LC-
MS were categorized as contaminations from another milk sample during milking and handling
of the milk sample on the farm or in the laboratory. No further proteomic studies were performed
to identify low intensity peaks in LC-MS. These were not taking into account during measurement
of relative protein content.
Genotypes of κ-, αs2-, αs1- and β-CN, β-LG and α-LA in native Nordic cattle breeds
By LC-MS, all protein genetic variants were identified in the six main proteins. For identification
of the high abundance milk protein genetic variants, the average mass and retention patterns
were compared to all know caseins, β-LG and α-LA protein masses for genotype determination.
Each sample was run in duplicates and both were used to genotype determination. Genotype
covers the gene product, the genetic variant, expressed as a protein product in the milk and not
on DNA level. The variation in proteins is therefore due to genetic differences and not variations
created on protein levels. The genotypic distribution within each breed of the main proteins is
collected in Table 3.
κ-casein phenotype
JUT (n=12)
RDM (n=26)
SM (n=23)
SRP (n=8)
Sample no.
AA 8 10 2 4 24
AB 4 13 10 1 28
BB 1 11 12
AE 1 1 2
BE 1 2 3
αs2-casein phenotype
AA 12 26 23 8 69
αs1-casein phenotype
BB 12 25 17 7 61
BC 1 5 1 7
CC 1 1
β-casein phenotype

37
.
Three different protein genetic κ-CN variants were found in this study, variant A, B and E,
combined in five different genotypes, AA, AB, BB, AE and BE. The κ-CN variants A and B were
present in all breeds, with AB as the most common genotype, present in 28 samples. All five
genotypes were present in RDM and only two genotypes were present in JUT, the breed with the
least diverse κ-CN protein genetic variant combination. Variant E was present in RDM and SRP
and was observed in low amount (two and three samples, respectively).
αs2-CN was present as one genetic variant, A, thus all samples had the genotype αs2-CN AA.
Two different αs1-CN genetic variants were present within the four breeds, variant B and C.
Variant B was the predominant and found in all breeds. Variant C was not found in JUT and found
in one animal in RDM and SRP. Six SM samples contained variant C, where one sample had variant
CC, which was not observed in the other breeds.
β-CN variants A1 and A2 were present in all breeds, whereas variant F was present in JUT and
RDM and variant B in SM. The β-CN variants were combined in seven different genotypes, A1A1,
A1A2, A2A2, A1F, A2F, A1B and A2B. The most common β-CN genotype was A1A2, present in 19
samples. RDM and SM were the breeds with the most diverse β-CN genotype, having five of the
seven observed β-CN genotypes. Moreover, they were also the breeds with highest sample
numbers (n=26 and n=23, respectively). SRP was the breed which had the most narrow β-CN
A1A1 3 4 2 1 10
A1A2 4 6 7 2 19
A2A2 3 8 5 16
A1F 3 2 5
A2F 2 11 13
A1B 4 4
A2B 2 2
β-lactoglobulin phenotype
AA 5 1 2 8
AB 6 3 9 3 21
BB 1 22 12 5 40
α-lactalbumin phenotype
BB 12 26 23 8 69
Table 3 Genotypic distribution of the six high abundance milk proteins in JUT, RDM, SM and SRP. All results were found by LC-MS. A total of every genotype found in the study is noted as “sample no.”

38
genotype pattern, having three of the seven observed β-CN phenotypes and also the breed with
the smallest sample number (n=8).
The most common β-LG phenotype was BB, as it was observed in 40 samples. Only two different
variants of β-LG, A and B, were observed, combined in genotype AA, AB and BB. Both protein
genetic variants were present in all breeds, where genotype AA was not observed in SRP.
Overall, the most common phenotypes in the four investigated breeds were κ-CN AB, αs2-CN AA,
αs1-CN BB, β-CN A1A2, β-LG BB and α-LA BB.
Together, this data showed that κ-CN and β-CN had a large genotype variation, with low variation
in αs1-CN and β-LG genotype. However, for αs2-CN and α-LA, all samples had the same genotype.
As the aim of this study was to study the protein profile, including variation in protein genetic
variants in native Nordic breeds, protein groups consisting of one protein genetic variant were
therefore not taken into account in the following experiments (αs2-CN and α-LA, Table 3). Sample
number of each breed varied a lot, from 8-26 samples per breed. A more clear view of the protein
genetic variant distribution can thereby be illustrated as protein genetic variant frequency
instead, where genotype is converted to protein genetic variant frequency by grouping all protein
genetic variants and dividing with sample number. Every genotype has two different alleles,
which can code for the same protein product or two different protein products. The protein
genetic variant frequency is therefore not found through DNA and not called alleles, but as the
protein product of different genetic codes.

39
Figure 8 Protein genetic variant frequency of heterogenic protein groups in Jutland (JUT), Danish Red (RDM), Swedish mountain (SM) and Swedish Red Polled (SRP) cattle (n=69). κ-A, κ-B and κ-E = κ-CN protein genetic variants. αs1-B and αs1-C = αs1-CN protein genetic variants. β–A1, β-A2, β-B and β-F = β-CN protein genetic variants. β–A and β-B = β-LG protein genetic variants. All frequencies were found within each breed with LC-MS analysis.
The frequencies of protein genetic variants are shown in Figure 8 Protein genetic variant frequency
of heterogenic protein groups in Jutland (JUT), Danish Red (RDM), Swedish mountain (SM) and Swedish
Red Polled (SRP) cattle (n=69). κ-A, κ-B and κ-E = κ-CN protein genetic variants. αs1-B and αs1-C = αs1-CN
protein genetic variants. β–A1, β-A2, β-B and β-F = β-CN protein genetic variants. β–A and β-B = β-LG
protein genetic variants. All frequencies were found within each breed with LC-MS analysisFigure 8, with
the different protein genetic variants for κ-CN, αs1-CN, β-CN and β-LG found in this study.
For κ-CN, the highest variant frequency observed was variant A in JUT, RDM, and SRP (0.83, 0.65
and 0.63, respectively), with the exception of SM having the highest κ-CN frequency of variant B
(0.70). κ-CN variant E was found in the red cattle, RDM and SRP (0.04 and 0.19, respectively). An
equally distributed κ-CN variant frequency between variant B and E was observed in SRP,
representing three samples with each variant (Table 3).
A low genetic diversity of αs1-CN was observed in all four breeds (Figure 8), with the highest
variant frequency of variant B in all breeds. Protein genetic variant frequencies of variant C in
RDM, SM and SRP were 0.02, 0.15 and 0.06, respectively. JUT did not contain variant C.
β-CN A2 was the variant with highest variant frequency within RDM, SM and SRP (0.44, 0.54 and
0.75, respectively), compared to variant A1, which was the variant with highest variant frequency
0
0.2
0.4
0.6
0.8
1
κ-A κ-B κ-E αs1-B αs1-C β-A1 β-A2 β-B β-F β-A β-B
Casein Whey
Freq
uen
cy (
%)
Protein genetic variants
Protein genetic variant frequency of κ-CN, αs1-CN, β-CN and β-LG
JUT RDM SM SRP

40
in JUT (0.54). Variant A1 was similarly distributed in the other three breeds, RDM, SM and SRP
(from 0.25-0.30). β-CN B was only found in SM in a frequency of 0.13. The two Danish breeds,
JUT and RDM, contained variant F in a frequency of 0.21 and 0.25, respectively.
In β-LG, two protein genetic variants were found, A and B. Variant B dominated the variant
frequency of RDM, SM and SRP (0.90, 0.72 and 0.81, respectively), while JUT had the highest
frequency of variant A (0.67).
Together, these data showed that JUT had the highest protein genetic variant frequency of κ-CN
A, β-CN A1, β-LG A, compared to the other breeds, and JUT contained β-CN F. The variants with
the highest variant frequency in RDM were κ-CN A, β-CN A2, β-LG B and it contained β-CN F. The
variants with the highest variant frequency in SM were κ-CN B, β-CN A1, β-LG B, and it contained
β-CN B. κ-CN B was found in the highest frequency in SM compared to all breeds. The variants
with the highest variant frequency in SRP were κ-CN A, β-CN A2, β-LG B. κ-CN E was in the highest
frequency in SRP compared to all breeds.
A relatively broad protein genetic variant distribution was observed between the breeds, which
could contribute to different characteristics of the milk, including differences in protein content
of each major protein group. However, these results did not reflect the differences in protein
content according to breeds.
VARIATION IN MILK PROTEIN PROFILE BETWEEN BREEDS
To investigate the protein content distribution within each breed, the relative protein content
was calculated. This was based on peak area in LC chromatograms as the relative protein content,
using the software ChemStation. As observed in Figure 6, peak heights were variable,
representing differences in relative protein content between samples. Various studies have
observed different amounts of milk protein isoforms (glycosylation degree of κ-CN and
phosphorylation degree of αs1-CN and αs2-CN) in different breeds. These were also investigated
with exception of αs2-CN, which in some samples was found in such low an amount, that the
isoforms were indistinguishable. The relative protein content was calculated by dividing the area
of the peak, from baseline to top with total area of all identified peaks (Glyco-κ-CN, κ-CN, αs2-CN,

41
αs1-CN, β-CN, β-LG and α-LA peak area). The relative protein content was thereby in percentage
according to all identified proteins. The relative protein content for each sample used throughout
this thesis was the same, but categorized according to the comparing parameters.
All statistical analysis was performed in R using ANOVA and Tukey’s test with significance level at
95% (P < 0.05). Means values significantly different from each other are specified from a-c, with
the highest mean value specified with “a“ and the lower mean value with “b” or “c”.
Protein
profile (%) JUT ± SE (n=12) RDM ± SE (n=26) SM ± SE (n=23) SRP ± SE (n=8) P-value
κ-CN 9.14 ± 0.55b 10.48 ± 0.77ab 11.16 ± 0.40a 10.94 ± 0.67ab 0.033
• κ-NonGlyco 6.27 ± 0.34ab 6.10 ± 0.23b 7.33 ± 0.24a 6.84 ± 0.41ab 0.003
• κ-Glyco 2.87 ± 0.41b 4.38 ± 0.28a 3.83 ± 0.30ab 4.10 ± 0.50ab 0.032
• κ-Glyco/κTotal 0.31 ± 0.02b 0.41 ± 0.02a 0.34 ± 0.02ab 0.37 ± 0.03ab 0.011
αs1-CN 32.10 ± 0.85 33.50 ± 0.57 32.20 ± 0.61 30.50 ± 1.04 NS
• αs1-8P 23.00 ± 0.71 23.70 ± 0.48 23.60 ± 0.51 23.30 ± 0.87 NS
• αs1-9P 9.02 ± 0.35ab 9.76 ± 0.24a 8.55 ± 0.25b 7.21 ± 0.47c < .0001
• αs1-8P/αs1Total 0.72 ± 0.01bc 0.71 ± 0.01c 0.73 ± 0.01ab 0.76 ± 0.01a 0.005
αs2-CN 6.64 ± 0.47ab 6.17 ± 0.32b 7.39 ± 0.34ab 8.07 ± 0.58a 0.014
β-CN 37.30 ± 0.82 36.80 ± 0.56 36.60 ± 0.59 36.60 ± 1.00 NS
β-LG 10.24 ± 0.54a 9.74 ± 0.37a 8.13 ± 0.39b 9.52 ± 0.66ab 0.006
α-LA 4.57 ± 0.25a 3.41 ± 0.17b 4.51 ± 0.18a 4.37 ± 0.31a < .0001
Table 4 Relative protein content within Jutland (JUT), Danish Red (RDM), Swedish mountain (SM) and Swedish Red Polled (SRP) cattle. Protein composition is calculated from LC chromatogram, where composition in % is peak area/total identified peak area. NS = non significant (P > 0.05). κ-NonGlyco= Nonglycosylated κ-CN. κ-Glyco= Glycosylated κ-CN. κ-Glyco/κ-Total= Glycosylated κ-CN/Total κ-CN. αs1-8P= αs1 with 8 phosphorylations. αs1-9P= αs1 with 9 phosphorylations. αs1-8P/αs1-Total= αs1 with 8 phosphorylations/Total αs1. Significant difference between breeds (P < 0.05) is indicated with a, b, c, the highest value a > b > c.
A comparison of the relative protein content between breeds is presented in Table 4. No
significant difference was found on total αs1-CN, αs1-8P and total β-CN between the breeds.
However, there were significant differences between the other major milk proteins when
comparing the four breeds. The biggest significant difference (P < 0.001) was observed in αs1-9P
and α-LA content. SRP has the lowest αs1-9P content, 7.21%, with the highest content in RDM of
9.76%. RDM had the lowest α-LA content, 3.41%, compared with the other breeds (no significant
difference, 4.37-4.57%). In total κ-CN amount, a significant difference (P = 0.033) between SM

42
and JUT was observed, with highest content in SM (11.16%) and lowest content in JUT (9.14%).
There was a significant difference in κ-NonGlyco content (P = 0.003) between RDM and SM,
where RDM had the lowest content on 6.10% and SM the highest content on 7.33%. κ-Glyco and
κ-Glyco/κ-Total were significant different, with the highest content in RDM (4.38% and 0.41,
respectively) and lowest in JUT (2.87% and 0.31, respectively). The lowest content of αs1-8P/αs1-
Total was observed in RDM (0.71) and highest in SRP (0.76) (P = 0.005). αs2-CN was significantly
different (P = 0.014) between the highest content in SRP (8.07%) and the lowest content in RDM
(6.17%). β-LG was in a high content in RDM and JUT and lower in SM (9.74%, 10.24% and 8.13%,
respectively) (P =0.006).
This is summarizes Table 5 (with P < 0.05).
Breed High content of Low content of
JUT β-LG κ-CN, κ-Glyco and κ-Glyco/κ-Total
RDM κ-Glyco, κ-Glyco/κ-Total, αs1-9P, β-LG κ-NonGlyco, αs1-8P/αs1-Total, αs2-CN, α-LA
SM κ-CN, κ-NonGlyco β-LG
SRP αs1-8P/αs1-Total, αs2-CN αs1-9P
Table 5 Summary of results obtained from Table 4. κ-NonGlyco= Nonglycosylated κ-CN. κ-Glyco= Glycosylated κ-CN. κ-Glyco/κ-Total= Glycosylated κ-CN/Total κ-CN. αs1-9P= αs1 with 9 phosphorylations. αs1-8P/αs1-Total= αs1 with 8 phosphorylations/Total αs1. Significant difference between breeds (P < 0.05) is indicated with a, b, c, the highest value a > b > c.
Many of the investigated protein groups seemed to be affected by breed differences. As observed
in Table 5, no breed showed superior differences within all protein groups, but RDM showed
differences in mean values in many of the protein groups, compared to the other breeds.
VARIATION IN MILK PROTEIN PROFILE BETWEEN PROTEIN GENETIC VARIANTS
The genotype profile and protein genetic variant frequencies in JUT, RDM, SM and SRP are unique
within each breed, with no breeds having the same genotype profile and variant frequencies
(Table 3, Figure 8). To investigate whether the protein profile of each breed, was due to genotype
differences between the breeds or could be ascribed to other breed characteristics as e.g. milk

43
composition, mineral content, feed and environment, the genotype influence on the protein
profile was investigated across the breeds.
Previous studies have shown correlations in variations of the protein profile (including
glycosylation degree of κ-CN and phosphorylation pattern of αs1-CN) and κ- and β-CN genotype.
To study if protein genetic variation of κ-CN were reflected in the protein production efficiency
in the milk, the relative protein content between κ-CN genotype was compared.
Protein
Profile (%) AA ± SE (n=24) AB ± SE (n=28) BB ± SE (n=12) AE ± SE (n=2) BE ± SE (n=3) P-value
κ-CN 9.13 ± 0.33b 11.02 ± 0.31a 12.38 ± 0.47a 9.30 ± 1.15a 10.47 ± 0.94a <.0001
• κ-NonGlyco 5.51 ± 0.20b 6.97 ± 0.19a 7.76 ± 0.28a 6.63 ± 0.71ab 7.02 ± 0.56ab <.0001
• κ-Glyco 3.52 ± 0.30 4.05 ± 0.28 4.62 ± 0.42 2.67 ± 1.04 3.45 ± 0.84 NS
• κ-Glyco/κTotal 0.37 ± 0.02 0.36 ± 0.02 0.37 ± 0.03 0.29 ± 0.07 0.33 ± 0.05 NS
αS1-CN 33.10 ± 0.61 32.10 ± 0.57 31.20 ± 0.87 33.10 ± 2.12 34.10 ± 1.73 NS
• αs1-8P 23.90 ± 0.49 23.30 ± 0.45 22.60 ± 0.69 23.70 ± 1.68 26.00 ± 1.37 NS
• αs1-9P 9.19 ± 0.30 8.87 ± 0.27 8.67 ± 0.42 9.47 ± 1.03 8.17 ± 0.83 NS
• αs1-8P/αs1Total 0.72 ± 0.01 0.73 ± 0.01 0.72 ± 0.01 0.72 ± 0.02 0.76 ± 0.02 NS
αs2-CN 6.84 ± 0.35 6.60 ± 0.32 7.95 ± 0.49 6.08 ± 1.21 6.05 ± 0.99 NS
β-CN 36.80 ± 0.57 37.20 ± 0.53 35.80 ± 0.81 36.90 ± 1.99 36.80 ± 1.63 NS
β-LG 10.25 ± 0.39a 8.96 ± 0.36ab 8.14 ± 0.55b 9.90 ± 1.34ab 8.37 ± 1.09ab 0.022
α-LA 3.89 ± 0.21 4.03 ± 0.19 4.50 ± 0.29 4.68 ± 0.71 4.20 ± 0.58 NS
Table 6 Relative protein content according to κ-casein phenotype AA, AB, BB, AE and BE. Protein composition is calculated from LC chromatogram, where composition in % is peak area/total identified peak area. NS= non significant (P > 0.05). NonGlyco= Nonglycosylated. Glyco= glycosylated. κ-Glyco/κ-Total= Glycosylated κ-CN/Total κ-CN. αs1-8P= αs1 with 8 phosphorylations. αs1-9P= αs1 with 9 phosphorylations. αs1-8P/αs1-Total= αs1 with 8 phosphorylations/Total αs1. Significant difference between breeds (P < 0.05) is indicated with a, b, c, the highest value a > b > c.
Table 6 shows the relative protein content within each κ-CN genotype group, AA, AB, BB, AE and
BE. A significant difference was observed in the relative protein content of total κ-CN and κ-
NonGlyco (P < 0.0001), together with the relative β-LG content (P = 0.022). There was not any
significant differences between the other protein groups and κ-CN genotype.
The BB genotype had the highest content of both κ-NonGlyco and κ-Glyco (7.76% and 4.62%,
respectively), which resulted in the highest total κ-CN content (12.38%). There was no significant
difference between the κ-Glyco content. The two relatively small groups, AE (n=2) and BE (n=3)

44
had a high standard error and statistical analysis of these small groups was therefore difficult to
validate. Genotype AA had the lowest total κ-CN content and κ-NonGlyco content (9.13% and
5.51%, respectively). A significant difference of κ-NonGlyco content was observed with grouping
of genotype BB and AB compared to AA. The β-LG content was highest in genotype AA (10.25%)
and lowest in genotype BB (8.14%). A pattern was observed between the total κ-CN content and
β-LG content, with the highest total κ-CN and lowest β-LG in BB, where the reverse is observed
in AA; lowest total κ-CN and highest β-LG.
It was observed that the κ-CN genotype had an influence on three of the investigated protein
groups, the content of total κ-CN (κ-CN), nonglycosylated κ-CN (κ-NonGlyco) and total β-LG (β-
LG) with the largest difference between genotype AA and BB. It is thereby possible that some
differences observed between breeds (Table 5) could be related to the influence of κ-CN
genotype in the breeds. Furthermore, β-CN had a high genotype diversity in the investigated
breeds and had been reported to influence the protein profile. To study if protein genetic
variation of β-CN was reflected in the protein profile, the relative protein content between β-CN
genotype was compared.
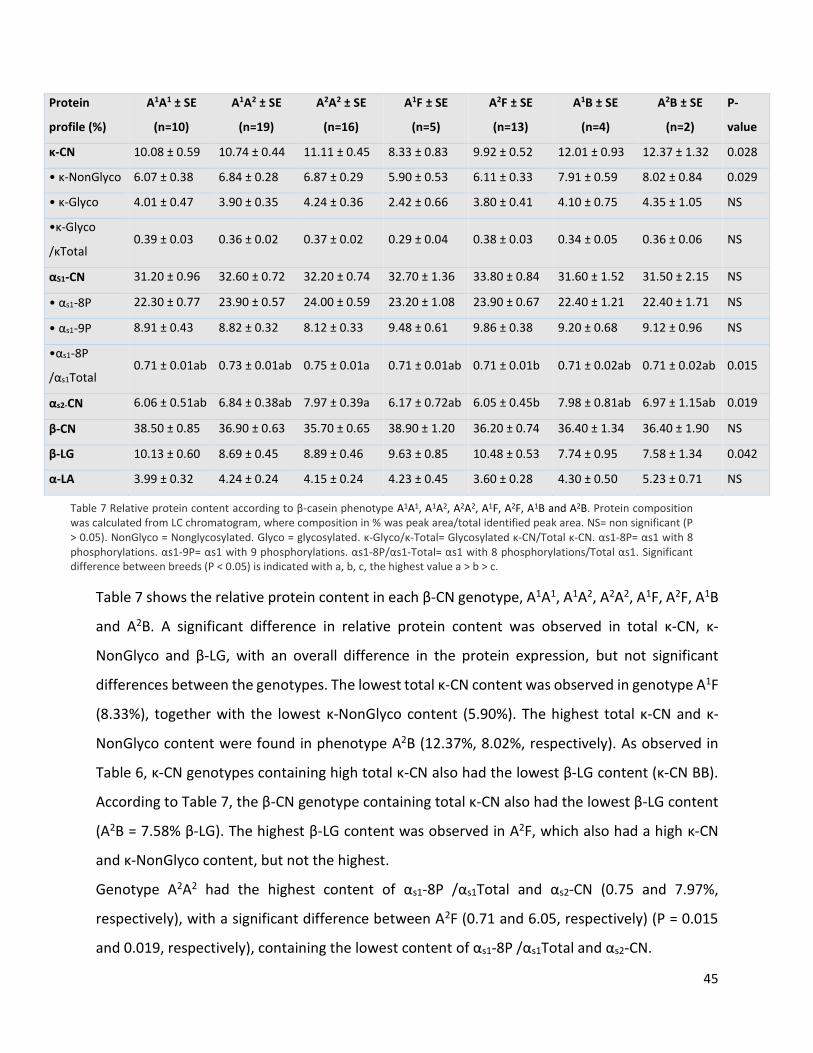
45
Table 7 shows the relative protein content in each β-CN genotype, A1A1, A1A2, A2A2, A1F, A2F, A1B
and A2B. A significant difference in relative protein content was observed in total κ-CN, κ-
NonGlyco and β-LG, with an overall difference in the protein expression, but not significant
differences between the genotypes. The lowest total κ-CN content was observed in genotype A1F
(8.33%), together with the lowest κ-NonGlyco content (5.90%). The highest total κ-CN and κ-
NonGlyco content were found in phenotype A2B (12.37%, 8.02%, respectively). As observed in
Table 6, κ-CN genotypes containing high total κ-CN also had the lowest β-LG content (κ-CN BB).
According to Table 7, the β-CN genotype containing total κ-CN also had the lowest β-LG content
(A2B = 7.58% β-LG). The highest β-LG content was observed in A2F, which also had a high κ-CN
and κ-NonGlyco content, but not the highest.
Genotype A2A2 had the highest content of αs1-8P /αs1Total and αs2-CN (0.75 and 7.97%,
respectively), with a significant difference between A2F (0.71 and 6.05, respectively) (P = 0.015
and 0.019, respectively), containing the lowest content of αs1-8P /αs1Total and αs2-CN.
Protein
profile (%)
A1A1 ± SE
(n=10)
A1A2 ± SE
(n=19)
A2A2 ± SE
(n=16)
A1F ± SE
(n=5)
A2F ± SE
(n=13)
A1B ± SE
(n=4)
A2B ± SE
(n=2)
P-
value
κ-CN 10.08 ± 0.59 10.74 ± 0.44 11.11 ± 0.45 8.33 ± 0.83 9.92 ± 0.52 12.01 ± 0.93 12.37 ± 1.32 0.028
• κ-NonGlyco 6.07 ± 0.38 6.84 ± 0.28 6.87 ± 0.29 5.90 ± 0.53 6.11 ± 0.33 7.91 ± 0.59 8.02 ± 0.84 0.029
• κ-Glyco 4.01 ± 0.47 3.90 ± 0.35 4.24 ± 0.36 2.42 ± 0.66 3.80 ± 0.41 4.10 ± 0.75 4.35 ± 1.05 NS
•κ-Glyco
/κTotal 0.39 ± 0.03 0.36 ± 0.02 0.37 ± 0.02 0.29 ± 0.04 0.38 ± 0.03 0.34 ± 0.05 0.36 ± 0.06 NS
αS1-CN 31.20 ± 0.96 32.60 ± 0.72 32.20 ± 0.74 32.70 ± 1.36 33.80 ± 0.84 31.60 ± 1.52 31.50 ± 2.15 NS
• αs1-8P 22.30 ± 0.77 23.90 ± 0.57 24.00 ± 0.59 23.20 ± 1.08 23.90 ± 0.67 22.40 ± 1.21 22.40 ± 1.71 NS
• αs1-9P 8.91 ± 0.43 8.82 ± 0.32 8.12 ± 0.33 9.48 ± 0.61 9.86 ± 0.38 9.20 ± 0.68 9.12 ± 0.96 NS
•αs1-8P
/αs1Total 0.71 ± 0.01ab 0.73 ± 0.01ab 0.75 ± 0.01a 0.71 ± 0.01ab 0.71 ± 0.01b 0.71 ± 0.02ab 0.71 ± 0.02ab 0.015
αs2-CN 6.06 ± 0.51ab 6.84 ± 0.38ab 7.97 ± 0.39a 6.17 ± 0.72ab 6.05 ± 0.45b 7.98 ± 0.81ab 6.97 ± 1.15ab 0.019
β-CN 38.50 ± 0.85 36.90 ± 0.63 35.70 ± 0.65 38.90 ± 1.20 36.20 ± 0.74 36.40 ± 1.34 36.40 ± 1.90 NS
β-LG 10.13 ± 0.60 8.69 ± 0.45 8.89 ± 0.46 9.63 ± 0.85 10.48 ± 0.53 7.74 ± 0.95 7.58 ± 1.34 0.042
α-LA 3.99 ± 0.32 4.24 ± 0.24 4.15 ± 0.24 4.23 ± 0.45 3.60 ± 0.28 4.30 ± 0.50 5.23 ± 0.71 NS
Table 7 Relative protein content according to β-casein phenotype A1A1, A1A2, A2A2, A1F, A2F, A1B and A2B. Protein composition was calculated from LC chromatogram, where composition in % was peak area/total identified peak area. NS= non significant (P > 0.05). NonGlyco = Nonglycosylated. Glyco = glycosylated. κ-Glyco/κ-Total= Glycosylated κ-CN/Total κ-CN. αs1-8P= αs1 with 8 phosphorylations. αs1-9P= αs1 with 9 phosphorylations. αs1-8P/αs1-Total= αs1 with 8 phosphorylations/Total αs1. Significant difference between breeds (P < 0.05) is indicated with a, b, c, the highest value a > b > c.

46
There were observed differences in the protein profiles between β-CN genotype with correlation
between A1F and A2B according to κ-CN protein expression. A correlation was also observed
between A2A2 and A2F according to αs1-8P /αs1Total and αs2-CN protein expression. The
expression of β-LG protein was inversely proportional to that of κ-CN protein expression. An
effect of β-CN genotype on milk protein profile from native Nordic cattle may therefore be
important to consider in future studies.
COAGULATION PROPERTIES OF MILK FROM NATIVE NORDIC CATTLE BREEDS
Genetic variance between breeds is a contributing factor to altering the milk composition. The
processing of milk is affected by milk composition, with the protein profile being an important
factor during further treatment of milk to different dairy products, especially cheese production.
Thus, some of the important technological properties during cheese production, rennet
coagulation time (RCT) and gel strength 40 minutes after rennet induced coagulation (G’40), were
compared to the protein profile from JUT, RDM, SM and SRP. The casein micelle size and stability
during processing are influencing the coagulation properties of the milk, and the calcium content
in milk has been related to casein micelle size and stability. To elucidate another milk
compositional trait according to breed differences, total and ionic calcium were included and
compared to coagulation properties between breeds, together with absolute protein
concentration.
Significant differences in relative protein content between breeds were observed (Table 4). The
largest differences in mean protein content were observed between the protein groups in
RDM, compared to the other breeds. To study if there were any patterns between breeds and
the relative protein content, calcium content, technological properties (RCT and G’40) and total
protein concentration, all measurements were subjected to Principal Component Analysis (PCA)
(scores plot Figure 9 and loading plot Figure 10). Relative protein content was used as X-
variables, together with relative protein content, total calcium (Total Calc) and ionic calcium
(Ionic Calc), which were also measured in this study.

47
Moreover, the absolute protein concentration (Protein), RCT and G’40, which were measured by
Milkescan, Reorox4 and Stresstech on Nordic native cattle, were included (Workflow, Figure 1).
G’40 was obtained in two different studies and thus measured with two different techniques.
Samples from RDM and JUT were measured with Reorox4, which measures G’max, maximum gel
strength, which is the highest value of the storage modulus G’, obtained within an hour after
rennet induced coagulation. Samples from SM and SRP were measured with Stresstech, which
measures G’40, gel strength 40 minutes after rennet induce coagulation. From Reorox4
coagulation data, storage modulus after 40 minutes was manually obtained, which corrected
G’max in RDM and JUT to G’40.
The PCA model showed some variance in specific breed genetics according to protein profile,
calcium content and coagulation factors, with a relative separation of the breeds by the first
component (Figure 9). No clear separation was observed between the breeds according to the
factors investigated here, but a little grouping of the Danish breeds (RDM and JUT) and the
Swedish breeds (SM and SRP) were detected, explaining 24.7% of the total variation in the data
set. The diverse protein profiles, calcium content and coagulation factors of the Danish breeds
and Swedish breeds were reflected in relatively higher content of β-LG, total calcium and αs1-
9P-CN together with higher RCT in the Danish breeds. In contrast, milk from SM and SRP were
characterized by relative higher content of κ-CN-NonGlyco, αs2-CN and αs1-8P/αs1Total and
higher G’40.
48
Figure 9 Principal component analysis, plot of scores of milk samples from Jutland (JUT), Danish Red (RDM), Swedish mountain (SM) and Swedish Red Polled (SRP) cattle. The relative protein content of each protein group, phosphorylation degree, glycosylation degree, protein concentration, total and ionic calcium and technological properties were used as X-variables.
Figure 10 Principal component analysis, plot of loadings of milk samples from Jutland, Danish Red, Swedish mountain and Swedish Red Polled cattle. The relative protein content of each protein group, phosphorylation degree, glycosylation degree, total protein concentration, total and ionic calcium together with technological properties were used as X-variables. RCT=Rennet coagulation time. G’40=Storage modulus at 40 minutes. Protein=Absolute protein concentration, g/100g. Calc=Calcium.

49
Based on PCA modelling, a grouping between breeds can to some extent be described by the
factors investigated in this thesis, with technological properties G’40 and RCT related to a breed
specific trait. To investigate the breed grouping in the PCA model, compositional (total and ionic
calcium, total protein concentration) and technological properties were compared for breed
differences (Table 8).
Comparing compositional and technological properties according to breeds showed no
significant difference in absolute protein concentration and G’40, even though RDM had a low
G’40. A high RCT was found in RDM (28.20 min) in contrast to SM and SRP (12.80 min and 11.10
min, respectively, p<0.0001), which was in line with the PCA model. JUT and SM contained the
lowest concentration of total calcium (25.60 mM and 25.80 mM, respectively) compared to RDM
with 36.70 mM (p<0.0001). Ionic calcium concentration was to some extent following total
calcium content and was significantly different (p=0.0004) between RDM and SM with 1 mM in
difference (2.99 mM and 1.92 mM, respectively).
Thereby, breed specific properties according to Table 8 were RCT, total calcium and ionic calcium.
RDM had the highest total calcium concentration, ionic calcium concentration and RCT, while JUT
and SM have the lowest total calcium concentration, reflected in the lowest ionic calcium
concentration and low RCT in SM. SRP have the lowest RCT, which is not reflected in calcium
content.
Compositional/
technological
properties JUT ± SE (n=12) RDM ± SE (n=26) SM ± SE (n=23) SRP ± SE (n=8) P-value
Protein (g/100g) 3.72 ± 0.18 3.95 ± 0.01 3.73 ± 0.10 3.49 ± 0.14 NS
RCT (min) 20.50 ± 2.51ab 28.20 ± 1.71a 12.80 ± 1.82b 11.10 ± 3.08b <.0001
G’40 (Pa) 145.50 ± 21.50 91.50 ± 15.90 142.70 ± 15.90 124.70 ± 26.30 NS
Total Calcium
(mM) 25.60 ± 1.88c 36.70 ± 0.92a 25.80 ± 0.96c 32.10 ± 1.33b <.0001
Ionic Calcium
(mM) 2.22 ± 0.25ab 2.99 ± 0.17a 1.92 ± 0.18b 2.16 ± 0.30ab 0.0004
Table 8 Compositional and technological properties within Jutland (JUT), Danish Red (RDM), Swedish mountain (SM) and Swedish Red Polled (SRP) cattle. NS= non-significant. G’40 = gel strength at 40 minutes. RCT = rennet coagulation time. Significant difference between breeds (P < 0.05) is indicated with a, b, c, the highest value a > b > c.

50
In correlation with the relative protein content in the κ-CN genotype (Table 6), κ-CN genotype
affected the protein expression of total κ-CN and κ-CN-NonGlyco, with lowest content in
genotype AA and highest content in genotype BB and AB. The κ-CN profile could thus be related
to RCT and G’40, based in κ-CN genotype AA and BB.
A haplotype effect of κ-β-CN on milk coagulation properties have been reported (Poulsen et al.
2013), but also a difference in coagulation properties between β-CN genetic variants is have been
studied (Heck et al. 2008). Therefore, compositional and technological properties according to β-
CN genotype were investigated
The technological properties G’40 and RCT varied significantly according to both κ-CN genotype
and β-CN genotype (p=0.002)(Table 10). In almost all β-CN genotypes were there a significant
difference in G’40 and RCT. G’40 and RCT is inversely correlated, with a low RCT and high G’40 in
β-CN A1A2 (15.04 min and 153.30 Pa, respectively) and β-CN A1B (7.54 min and 195.90 Pa,
respectively). On the other hand, β-CN A2F have a high RCT (28.79 min) and low G’40 (55.00 Pa).
β-CN A1F had a high RCT (31.38), with no significant effect on G’40. No significant differences
were observed in the compositional properties.
Compositional/
technological
properties AA ± SE (n=24) AB ± SE (n=28) BB ± SE (n=12) AE ± SE (n=2) BE ± SE (n=3) P-value
Protein (g/100g) 3.73 ± 0.11 3.80 ± 0.10 3.81 ± 0.15 3.59 ± 0.37 3.63 ± 0.30 NS
RCT (min) 25.40 ± 2.13a 17.50 ± 1.85ab 13.00 ± 3.01b 26.90 ± 7.36ab 17.70 ± 6.01ab 0.010
G’40 (Pa) 72.60 ± 15.40b 139.60 ± 14.2a 147.50 ± 21.70a 73.40 ± 53.2ab 109.00 ± 43.50ab 0.015
Total Calcium
(mM) 31.90 ± 1.42 31.90 ± 1.26 29.20 ± 1.93 30.70 ± 6.67 24.20 ± 3.85 NS
Ionic Calcium
(mM) 2.49 ± 0.20 2.50 ± 0.18 2.11 ± 0.28 1.97 ± 0.97 1.91 ± 0.56 NS
Table 9 Compositional and technological properties within Jutland (JUT), Danish Red (RDM), Swedish mountain (SM) and Swedish Red Polled (SRP) cattle. NS= non-significant. G’40 = gel strength at 40 minutes. RCT = rennet coagulation time. Significant difference between breeds (P < 0.05) is indicated with a, b, c, the highest value a > b > c.

51
No significant difference was observed in nether κ-CN and β-CN genotype comparison to protein
concentration, total calcium and ionic calcium. According to Table 8, differences in total and ionic
calcium concentration is more affected by breed specific genetic variation, then to κ-CN and β-
CN genotype.
Additional proteins observed by LC-MS
Within the genotyping of the major milk proteins there was discovered an extra mass within the
αs2-CN peak. The Mw of the protein was 24041 Da, and it was found within four Danish and eight
Swedish samples. The protein had an equally high ion intensity to αs2-CN A (25220 Da) present in
the sample, indicating that the two proteins were present in equal amount. Within five Swedish
samples, no other Mw than 24041 Da could be calculated, indicating a possible new variant
present, since no other Mw of αs2-CN have been observed. All samples containing a protein of
24041 Da within the αs2-CN peak was for the simplicity genotyped αs2-CN AA.
Table 10 Compositional and technological properties according to β-CN genotype (A1A1, A1A2, A2A2, A1F, A2F, A1B and A2B). NS= non-significant. G’40 = gel strength at 40 minutes. RCT = rennet coagulation time. Significant difference between breeds (P < 0.05) is indicated with a, b, c, the highest value a > b > c.
Composition
/technologic-
al properties
A1A1 ± SE
(n=10)
A1A2 ± SE
(n=19)
A2A2 ± SE
(n=16)
A1F ± SE
(n=5)
A2F ± SE
(n=13)
A1B ± SE
(n=4)
A2B ± SE
(n=2) P-value
Protein
(g/100g) 3.86 ± 0.16 3.72 ± 0.12 3.94 ± 0.12 3.43 ± 0.23 3.73 ± 0.14 3.71 ± 0.25 3.29 ± 0.36 NS
RCT (min)
17.82 ±
3.10ab
15.04 ±
2.31b
19.12 ±
2.32ab
31.38 ±
4.28a
28.79 ±
2.65a
7.54 ±
4.89b
14.49 ±
6.92ab 0.0002
G’40 (Pa) 135.20 ±
22.60ab
153.30 ±
16.80a
107.90 ±
17.30ab
56.30 ±
31.90ab
55.00 ±
19.80b
195.90 ±
35.70a
84.00 ±
50.50ab 0.002
Total Calcium
(mM) 30.40 ± 2.07 32.10 ± 1.55 27.90 ± 1.69 32.30 ± 2.93 34.80 ± 1.89 28.30 ±3.28 28.70 ± 4.64 NS
Ionic Calcium
(mM) 2.31 ± 0.30 2.52 ± 0.22 2.34 ± 0.23 1.97 ± 0.42 2.83 ± 0.27 1.88 ± 0.47 1.51 ± 0.67 NS

52
Moreover, an SM sample contained a peak with retention time between β-CN A2 and F, which
contained a compound with Mw 23991 Da. The peak height and ion intensity was equivalent to
β-CN A2 present in the sample. Calculation of relative protein content of the A2 peak showed that
the protein amount was half that of the protein amount normally seen in β-CN A2A2 milk samples.
Therefore the peak after β-CN A2 with an unknown protein of 23991 Da was enrolled under the
genotype β-CN A2A2. This was possibly a new β-CN genetic variant observed in SM cattle.
However, due to the time frame of this thesis, no further investigation within the origin of the
two proteins were done.
7 DISCUSSION
In this study, the protein profile from four native Nordic cattle breeds were identified. Major milk
proteins were analyzed, including κ-, αs2-, αs1- and β-CN, β-LG and α-LA. The protein genetic
variants and protein isoforms, consisting of the degree of glycosylated κ-CN and phosphorylated
αs1-CN were examined. Various comparisons of breed and genotype characteristics according to
protein profile, calcium concentration, RCT and G’40 were examined to detect any breed related
differences. Variations between breeds were compared to obtain and discuss possible
technological benefits or disadvantages.
PROTEIN PROFILING
LC-MS was used for protein profiling, detecting all high abundance proteins in the milk.
Throughout the analysis, various low intensity peaks, which could look like a protein from one of
the six major protein groups, appeared in the chromatogram. However, they could not be
detected on 2-DGE. Therefore, the proteins in low intensity peaks were concluded to be overrun
from other milk samples.
From these experiments, the protein profile of native Nordic cattle breeds representing thirteen
different protein genetic variants of the six main proteins in the milk, relative protein content
together with protein isoforms of κ-CN and αs1-CN were identified. Milk protein allele frequencies
in Nordic cattle breeds have previously been studied by Lien and colleagues (1999), to reveal

53
variants that are characteristic for these breeds. They compared five modern Nordic cattle breeds
to seventeen old cattle breeds by DNA sequencing of the CSN1S1, CSN2, CSN3 and LGB loci. The
results showed higher allele frequency of αs1-CN C, even though variant B was still predominant.
They also observed that SM contained a relatively high allele frequency of αs1-CN C (0.31), which
was consistent with results from this study (Figure 9), with SM having a variant frequency of 0.15.
αs1-CN C was also observed in very low frequency in SRP (0.06) and RDM (0.02) in this study, and
this also confirmed the observations in SRP (0.10) by Lien.
According to Lien, the dominating κ-CN genetic variant within the four investigated breeds were
variant A in JUT and RDM and variant B in SM and SRP. This corresponds to results in this study,
except from SRP, which had the highest amount of variant A. Interestingly, Lien reported κ-CN E
in JUT, RDM and SRP, with a high allele frequency in SRP (0.20), which was the highest frequency
reported in all 22 investigated breeds. κ-CN E was also observed in this study in SRP (0.19) and
RDM (0.04), in accordance to Lien. κ-CN B was observed in a lower frequency in current
conventional breeds, compared to milk from native Nordic breeds, with the highest frequency in
SM (0.71), which was also accordance with Lien.
Moreover, Lien and colleagues observed that β-CN A2 generally was the predominant allele, but
β-CN A1 was observed in higher frequencies in the native breeds from northern Europe. β-CN A1
and A2 were seen in approximately same frequency in the four breeds (Lien et al. 1999). Within
this study, JUT expressed a higher frequency of β-CN A1 (0.54) compared to A2 (0.25). However,
β-CN F was not studied by Lien. This may distort the comparison of β-CN variant frequencies, due
to the high frequency of variant F observed in this study. Variant B was reported within all four
breeds by Lien, but was only observed in SM in this study.
Since the sample numbers in the present study and that used by Lien and colleagues were
different this limits the comparison. With the low sample number in SRP, few individuals with e.g.
κ-CN E resulted in a high variant frequency of κ-CN E, compared to the sample number in RDM.
The Jutland breed studied by Lien was from year 1950. Whether the investigated animals was
indeed pure SJM or was a mix of SJM and JUT, as animals within this study, is not known. A
comparison of variant frequencies between the two studies should therefore be interpreted with
care. No big difference in allele frequencies reported by Lien and variant frequencies observed in

54
this study were found, except from SJM in Lien and JUT in this study. This could indicate a relative
homogeneous gene pool within each breed o RDM, SM and SRP, where JUT in this study can be
influenced by other gene populations, as mention in section 2.3.
Within all four breeds, 20 genotypes were identified (Table 3). None of the investigated breeds
showed clear relationship based on genotype and variant frequency, with diverse variant
frequencies in all breeds. According to the genetic diversity in North European cattle breeds
studied by Kantanen et al. (2000), the closest relationship among breeds reported is between
RDM and SRP, which are in two subgroups of Southern breed group. Both red cattle breeds are
more closely related to SM than JUT, and SM more closely related to RDM and SRP than JUT
(Kantanen et al. 2000). A correlation in breed specific genetic variation between RDM and SRP
could therefore be expected. According to genotypes found in this study, differences could be
observed related to the occurrence of κ-CN E, which was observed in RDM and SRP. κ-CN E has
previously been reported in Danish Holstein-Friesian and Swedish Red cattle (Jensen et al., 2012a;
Hanne B Jensen et al., 2015; Poulsen et al., 2013; Gustavsson, et al., 2014a), and in many old
Nordic cattle breeds (Lien et al. 1999). The genotypes of JUT had a high variant frequency of β-
CN F (0.21). Similarly, RDM also had a high frequency of β-CN F (0.25). β-CN F has not been
reported in many Danish and Swedish breeds, but Jensen et al. (2012a) reported β-CN F in Danish
Holstein-Friesian in non-coagulating milk in a frequency of 0.08. Therefore, the variant
frequencies observed in both Danish breeds are considered to be high. Again, whether Kantanen
et al. (2000) investigate Jutland breed containing both SJM and JUT is unknown, and a possible
relationship between investigated RDM and JUT cannot be excluded.
The most common commercial dairy cattle breed in Denmark is Holstein-Friesian. Jensen et al.
(2012a) compared good and poor coagulating milk from two Danish cattle breeds, Jersey and
Holstein-Friesian (DH) to the protein genetic variants and isoforms in each group. No αs1-CN C
variant was observed in DH and β-CN F was reported in low frequency (0.083) in non-coagulating
milk in DH. Both of these protein genetic variants were observed in native Nordic cattle in a higher
frequency, compared to DH. κ-CN BB is observed in a very high frequency in SM compared to DH.
Thereby, native Nordic cattle contains another variant frequency distribution, compared to DH.

55
VARIATION IN MILK PROTEIN PROFILE BETWEEN BREEDS.
The genotype distribution between the four breeds, JUT, RDM, SM and SRP was very different
(Table 3). Similarly, many studies have reported a diverse protein content distribution between
breeds and genotype (Jensen et al. 2012a; Gustavsson et al. 2014a). Investigation in protein
content both between breeds and according to κ- and β-CN genotype were done to compare the
variation in protein profile to the effect of breed specific genetics, or to κ- and β-CN genetic
variant specific variation. The relative content of the major proteins have been detected by RP-
HPLC or capillary zone electrophoresis (CZE) in different other studies; and variations between
breeds were observed (Heck et al. 2008; Heck et al. 2009; Bonfatti et al. 2011; Gustavsson et al.
2014a,b; Demeter et al. 2010)(Table 11). A variation in relative protein content would therefore
also be expected in native Nordic breed. However, it was observed that the relative protein
content was within the range of the reported except α-LA, which was observed in higher content
in this study compared to previously (Table 11).
Reported relative protein
content (%)1
Relative protein content in
native Nordic breeds (%)
κ-CN 4.01 - 10.46 9.14 – 11.16
αs2-CN 6.72 – 12.11 6.17 – 8.07
αs1-CN 29.48 – 35.61 30.50 – 33.50
β-CN 27.17 – 37.14 36.60 – 37.30
β-LG 8.30 – 9.13 8.13 – 10.24
α-LA 2.16 – 2.45 3.41 – 4.57
Table 11 Previously reported relative protein content by RP-HPLC and CZE in different breeds compared to relative protein
content in native Nordic breeds. 1(Heck et al. 2009; Heck et al. 2008; Bonfatti et al. 2011; F. Gustavsson et al. 2014a,b; Demeter
et al. 2010), by RP-HPLC or CZE.
Bovine milk protein largely (±90%) consist of the six major milk proteins investigated here. The
remaining 10% are low abundance proteins, not detected by LC or registered manually as a peak,
but maybe as a small hump up from baseline-level. The relative protein content was calculated
from the obtained LC chromatograms as the peak areas divided with total areas of all identified
peaks. Some studies calculate relative protein content as peak area divided with total protein

56
(Jensen et al. 2012a; Gustavsson et al. 2014a). This includes all proteins and compounds, e.g.
proteolytic CN degradation products and bovine serum albumin. By calculating the relative
protein content out from this interval only, the content of the genetic protein variants of CN and
whey are overestimated, as approximately 10% of the proteins in milk are not considered and CN
and whey are assumed to make up 100% alone. Conversely, by estimating the relative protein
content from the entire spectrum, background noise from the LC will be included in total protein
content. This causes an underestimation. Due to small errors in both calculations, the calculation
with all identified peaks performed in the present study. Moreover, CZE measured only the
nonglycosylated forms of κ-CN, which were then identified as the total κ-CN of the sample.
However, glycosylated κ-CN co-elutes with β-CN, which thereby is overestimated. Direct
comparison to reported relative protein content in other studies is thus not possible.
However, other few studies directly compare the relative protein content between breeds.
Gustavsson et al. (2014a) compared Swedish Red, Danish Holstein, and Danish Jersey cattle, to
investigate the relationship between casein genetic variants and detailed protein composition.
The protein profile was different between breeds, but also different in κ-, β-, and αs1-CN relative
protein contents compared to the genotype composition of κ-, β-, and αs1-CN. Moreover, it has
previously been reported in both Danish Holstein-Friesian and Danish Jersey, that the κ-CN
protein profile changed between breeds, when comparing κ-CN genotype and coagulation
properties of the milk (Jensen et al. 2012a; H B Jensen et al. 2015). Changes in the relative κ-CN
protein content could occur between the four investigated breeds, and one contributing factor
could be a different κ-CN genotype distribution between the breeds. Thereby, SM had
significantly higher total κ-CN relative content compared with the lowest in JUT. The high total κ-
CN content in SM could not be reflected in an increase in both glycosylated (κ-Glyco) and
nonglycosylated (κ-NonGlyco) κ-CN, since SM had the highest nonglycosylated content, but not
a high glycosylated content. The same was observed in JUT, which had the lowest glycosylated
content, but not nonglycosylated content. This could indicate that other regions in the DNA is
coupled with κ-CN protein profile, since no clear pattern is observed (Graml et al. 2003).
Glycosylation degree has been reported to vary from 22-76%, indicating that κ-Glyco variation
depends on many factors, which could be both variation in total κ-CN and in the efficiency of κ-

57
CN glycosylation (Bonfatti et al. 2014). In this study, RDM had the highest content of κ-Glyco and
lowest content of κ-NonGlyco, resulting in the highest κ-Glyco/κTotal ratio. From these results,
the high κ-Glyco content in RDM could be related to some high-efficient glycosylation traits in
RDM, while JUT with a low total κ-CN and a low κ-Glyco/κTotal ratio, maybe could be related to
some low-efficient glycosylation traits in JUT.
According to breed related to relative protein content in Table 4, there was no significant
difference in total αs1-CN and αs1-8P-CN (P=phosphorylations) between breeds, but a significant
difference in αs1-9P-CN between breeds. It was observed that RDM had the highest content
compared to SRP with the lowest content. The major part of variance in αs1-CN content has been
related to CSN1S1 loci and less affected by the genetic variance of the gene (Graml et al. 2003).
In respect to this, the αs1-CN genotype was the same in both breeds (All samples αs1-CN BB expect
one BC in each breed). Other factors then the αs1-CN genotype may affect the variation in αs1-CN
content in native Nordic breeds.
The most common phosphorylation states of are eight or nine phosphorylated serine residues.
Factors effecting the phosphorylation degree of αs1-CN have been investigated by Bijl and
colleagues (2014). They reported a low genetic and genotypic correlation between αs1-8P-CN
(0.18) and αs1-9P-CN (0.19). αs1-8P-CN was instead reported to be associated with β-LG A/B
polymorphism, and the β-LG AA genotype was associated with lower αs1-8P-CN concentration
compared to β-LG BB. Polymorphism in diacylglycerol acyltransferase 1 affected the
concentration of αs1-9P-CN. This indicates that the phosphorylation degree of αs1-CN is, to a large
extent, a genetically different trait. Also, it had a high heritability for αs1-9P-CN (0.76) compared
to αs1-8P-CN (0.48)(Bijl et al. 2014). The highly significant difference between αs1-9P-CN content
in RDM and SRP supported these results, since no difference in αs1-8P-CN content was observed.
Another regulatory mechanism could be an explanation. As an intermediate between RDM and
SRP, SM does also stand out. With a significant difference between three out of four breeds, the
αs1-9P-CN content could be associated with a breed-specific genetic trait. RDM contained a
higher αs1-9P-CN compared to αs1-8P-CN, which resulted in a low αs1-8P-CN/αs1Total and the low
αs1-9P-CN content in SRP resulted in a high αs1-8P-CN/αs1Total. JUT contained five samples with

58
genotype β-LG AA with a variant frequency on 0.67, but no differences in αs1-8P-CN content could
be observed.
The CSN2 locus accounts for the major part of the β-CN protein variance. A heterozygote
genotype seems to result in a higher β-CN protein content, compared to a homozygote (Graml
et al. 2003). Variance in protein content is associated with β-CN variant A1, A2 and B, where A1 is
associated with a lower β-CN content compared to A2 and B (Heck et al. 2009; Heck et al. 2008).
The variance in β-CN expression have reported to be affected mainly by the CSN2 locus (Graml
et al. 2003) No variance in β-CN content is observed between the breeds, even though they have
different β-CN protein variant frequencies. This could indicate a stable transcription of β-CN
genes in all breeds, with a very little effect of the β-CN genotype.
Genetic variance of the BLG gene (β-LG) is reported to be the main source for changes in the β-
LG protein content (Graml et al. 2003). Milk containing genotype β-LG AA have a significantly
higher β-LG content compared to milk with β-CN BB genotype (Ng-Kwai-Hang & Kim 1996). JUT
has the highest β-LG content, together with the highest variant frequency of β-LG A. The breed
difference in β-LG content seems related to the variant frequency of β-LG in the breeds.
VARIATION IN MILK PROTEIN PROFILE BETWEEN GENETIC VARIANTS
Studies indicate that casein polymorphism and relative protein content are connected (Heck et
al. 2008; NG-Kwai-Hang et al. 1987). The four casein genes are closely linked in a casein locus on
chromosome 6. This structure might lead to disequilibrium linkages between two casein variants,
as described in section 2.1.1. This can make it difficult to determine if an effect is specific for the
casein in question, or is an effect of another closely linked casein gene. Various studies have
investigated effects of genetic variants on milk protein composition and heritabilities between
the six major milk proteins (Gustavsson et al. 2014a; Heck et al. 2009; Bobe et al. 1999). Within
this study, the relative protein content from JUT, RDM, SM and SRP, varied significantly in many
of the protein groups. Genetic polymorphism of milk proteins is associated with milk composition,
technological properties and production traits of the milk. To investigate how much different
genotypes contribute to differences in the relative protein content, κ-CN and β-CN genotypes

59
were compared in terms of the relative protein content. During the comparisons, breeding
differences were neglected. By combining four breeds in one category, differences between
them were not considered. This was performed to ease the analysis, but may, however, have
introduced artefacts or hidden differences and trends in the data. Both the breed-specific genetic
variation and genotype-specific variation were compared to relative protein content. In that way,
investigation into each contributing factor for differences in protein profile was done, giving a
collected overview.
In other studies, especially κ-CN and β-CN have been investigated. Here, the casein and whey
genotype effects on relative protein content have been reported to vary. For example, milk with
κ-CN BB has been associated with lower content of αs1-CN and a higher content of κ-CN and
glycosylated κ-CN (Bobe et al. 1999; Heck et al. 2009; H B Jensen et al. 2015), compared to κ-CN
AA. Milk with β-CN A1 has been associated with higher contents of αs1-CN and κ-CN, together
with a lower content of αs2-CN and β-CN compared to β-CN A2 (Heck et al. 2008). Poulsen and
colleagues (2013) reported a haplotype effect of milk with β-CN A2A2 + κ-CN AA, which had lower
total casein, lower total protein and lower κ-CN content compared to other haplotypes. On the
other hand, Day et al. (2015) did not find any association between the total protein content and
different β-CN phenotypes. In this study, the different β-CN genotypes showed an overall
significant effect on κ-CN, κ- NonGlyco, αs1-8P/αs1Total, αs2-CN and β-LG, with only a significant
difference between the genotypes in αs1-8P/αs1Total and αs2-CN (Table 7). Graml et al. (2003)
reported an effect of the CSN2 gene variant on the content of κ-CN and β-LG. This was partly
observed in this study, with an overall β-CN genotype effect, but not between the genotypes.
The β-CN content was not affected by the β-CN genotype either, indicating a stable expression
mechanism of β-LG. A high content of αs2-CN was observed in β-CN A2A2, in accordance to Heck
et al. (2008), with the lowest content in β-CN A2F, which had not been reported before. A positive
correlation between β-CN genotypes containing variant B and a high content of κ-CN and κ-
NonGlyco were observed in this study, where β-CN A1F was correlated with low κ-CN content.
The β-CN genotype thus can be related to differences in protein content. However, no β-CN
genotype had a general effect on the protein content.

60
Different κ-CN genotypes have previously been associated with changes in the six major milk
proteins relative concentrations (P < 0.001)(Heck et al. 2009). Therefore, a correlation between
κ-CN genotype and relative protein concentration was investigated in this study (Table 6). It
showed that genetic variation of κ-CN affected the relative protein content of total κ-CN, κ-
NonGlyco and β-LG. The highest genetic variation was observed between genotype κ-CN AA and
BB, where κ-CN BB was associated with high total κ-CN and κ-NonGlyco content and low β-LG
content. Jensen et al. (2015) and Heck et al. (2009) have reported a significantly higher relative
κ-CN content in cows with κ-CN BB compared to κ-CN AA/AE, with the lowest relative κ-CN
content in κ-CN AE. This is partly in agreement with results in this study, where a relation of κ-CN
BB and higher κ-CN content were observed, and it was significantly different compared to κ-CN
AA. The genotype κ-CN AE was found in two animals and had a high standard error (Table 6). The
low sample number was not useful for statistical analysis and therefore the Tukey’s test criteria
to perform the test was not meet. Bonfatti and colleagues (2014) reported a relative higher
concentrations κ-CN in κ-CN BB milk compared to variant AA. This included both the glycosylated-,
nonglycosylated- and glycosylation degree (κ-Glyco/κ-CN). This is in accordance to previously
results (Coolbear et al. 1996). Based on this, a higher glycosylation degree of κ-CN BB was thus
to be expected. However, Jensen et al. (2015) did not find any significant difference for κ-Glyco/κ-
CN between genotypes containing κ-CN A/B/E. The relative content of glycosylated κ-CN and κ-
Glyco/κ-CN in native Nordic cattle did not differ significantly between κ-CN genotypes, whereas
the κ-NonGlyco content did. The higher total κ-CN content in κ-BB was a result of the increase in
κ-NonGlyco, and not a significant increase in κ-Glyco, where the low total κ-CN content in κ-AA
was a result in decreased κ-NonGlyco content.
Both the genotypic κ-CN effect, β-CN effect and breed effect have been compared to relative
protein content. A comparisons of these results showed that the content of κ-Glyco seemed to
be more regulated by breed genetic than by κ-CN or β-CN genotype. A significant different was
observed throughout the κ-CN profile between breeds. If the variation in κ-Glyco content
depends on total κ-CN content and the glycosylation efficiency (Bonfatti et al. 2014), RDM may

61
be predicted to have a strong glycosylation efficiency, with the highest κ-Glyco content and
lowest κ-NonGlyco content, but not a significantly high total κ-CN content.
In accordance with previously reported studies, β-LG content in this study was associated with κ-
CN genotype, where κ-CN BB was associated with lower content of β-LG compared to κ-CN AA
(Table 6)(NG-Kwai-Hang et al. 1987; Bobe et al. 1999).
COAGULATION PROPERTIES OF MILK FROM NATIVE NORDIC CATTLE BREEDS
There were six milk samples from RDM, which were non-coagulating, as no gel was formed and
G’40 was 0. For SM there was one non-coagulating milk sample. In samples where no RCT was
recorded, samples was noted with RCT = 45 min. This was done to obtain equally distributed data
in the statistical model. If RCT = 0 in the samples, the model calculate with a coagulation start
after 0 minutes. However, the samples were non-coagulating, meaning a really slow coagulation
start according to the model, thereby RCT = 45 minutes. Curd firming rate (CFR) is a coagulation
factor, used in many studies to measure coagulation properties of the sample. CFR data were
only available on samples from RDM and JUT. Therefore, the data were not enrolled in the
comparison models.
From Reorox4 coagulation data, storage modulus after 40 minutes was manually obtained, which
corrected G’max in RDM and JUT to G’40. Possible differences in G’40 between Danish and
Swedish breeds may be caused by differences in the two methods. One farm in this study
(Thorshøjgaard), showed a relatively diverse milk composition compared the other farms and
breeds (Nina A. Poulsen, unpublished data). Therefore, the twelve RDM from Thorshøjgaard were
omitted from the PCA model, resulting in a more comparable model. Future analysis can show
how the PCA model is affected by including these data.
During cooling of milk, calcium from the casein micelles dissociate out of the micelles. In that way
the soluble and ionic calcium concentration increase. By increasing the temperature, calcium will
be restored in the casein micelle. The concentration of ionic calcium was measured on thawed
samples, which stayed for 24 hours at 4°C, in a water bath at 30°C for 60 minutes and room

62
temperature for 30 minutes. Whether the calcium level in the casein micelles had become
identical to the original calcium micelle level before freezing is still a subject for investigation
(Walstra et al. 2006; Koutina et al. 2014). The mean values of ionic calcium levels in native Nordic
cattle was 2.39mM at room temperature, which was higher than previously reported (1.61mM,
20°C)(Koutina et al. 2014). A freezing effect on the stability of the CN micelle is a possible
explanation for the high ionic calcium (Gaucheron 2005).
By comparing compositional and technological properties for individual breeds, general breed
effects were observed for the parameters RCT, total calcium and ionic calcium, and there were
no significant effects in G’40 and total protein concentration. RDM had a high RCT, high total and
ionic calcium concentration. Milk from SM and SRP showed a low RCT, together with low total
calcium and low ionic calcium concentration in SM. Coagulation time has been reported to
decrease as ionic calcium increases, and addition of soluble calcium shortened the coagulation
time and increased the gel firmness (Sandra et al. 2012; Lewis 2011). Furthermore, higher
concentrations of total and ionic calcium were associated with both shorter gelation time and
higher G’40. The effect of ionic calcium in rennet induced gelation seems to be higher, compared
to total calcium (Gustavsson et al. 2014b). For RDM, the general high calcium level should thus
be correlated with a decrease in RCT, which was not the case. However, other factors can
influence on the onset of gelation, e.g. temperature, pH and casein profile; especially the κ-CN
profile (Bittante et al. 2012). Differences in calcium concentrations in milk have reported to be
more influenced by breed, compared to different genotypes (Gustavsson et al. 2014b), which is
also in accordance to results in the present study.
The comparison of the κ-CN genotype in terms of the compositional and technological properties
showed a clear significant difference between κ-CN AA and κ-CN BB according to technological
properties. A high RCT and low G’40 in κ-CN AA, and a low RCT and high G’40 in κ-CN BB were
observed. This is in accordance with the literature: Other studies have reported κ-CN BB to be
associated with improved rennet coagulation properties, compared to κ-CN AA. These
characteristics have been partly ascribed to a generally higher overall casein level in milk
containing κ-CN BB (Ikonen et al. 1999; Walsh et al. 1998; Gustavsson et al. 2014b). This is

63
especially true for higher glycosylated κ-CN BB content, with both higher levels of κ-CN-Glyco and
κ-CN-NonGlyco (Bonfatti et al. 2014). The κ-CN content in this study was also affected by the κ-
CN genotype, where κ-CN BB / AB contained significantly higher amounts of total κ-CN and κ-
NonGlyco, compared to κ-CN AA. Milk containing κ-CN BB had an overall favorable coagulation
properties. However, Jensen, Holland, et al. (2012) reported a consistent κ-CN isoform pattern
with only minor differences in relation to breed, κ-CN genetic variant and coagulation properties
of the milk. This contradictory results was also partly observed in this study, as the κ-CN isoform
pattern seemed only to be affect in some cases, according to κ-CN-NonGlyco, but not according
to κ-CN-Glyco content, when comparing to protein variant polymorphism.
SM was the breed with the highest κ-CN BB genotype frequency, nearly 50%, and a variant
frequency of κ-CN variant B on 0.70. SM also showed low RCT and high κ-CN content, which both
could be correlated to the high κ-CN variant B content in SM. κ-CN variant A was more equally
distributed in the three other breeds, with the highest variant frequency in JUT (0.83), where it
existed as κ-CN AA genotype in 67% of the samples. This could be related to the low total amounts
of κ-CN and κ-CN-Glyco in JUT, which did not seem to be reflected in a lower RCT, compared to
the other breeds. κ-CN variant E has been associated with poorly coagulating milk (Poulsen et al.
2013; Jensen et al. 2012a). However, this effect of RCT on κ-CN E was not described by Bittante
et al. (2012) who reported that there was no effect o RCT of the κ-CN E allele. In this study, κ-CN
E had the highest variant frequency in SRP (0.19), and it was present in 37% of the samples. SRP
was here correlated with low RCT. Thus, no negative effect of κ-CN E was observed on RCT,
indicating that other properties than the presence of κ-CN E may contribute to the low RCT
observed in this breed.
The β-CN genotype has been widely discussed according to coagulation properties, in relation to
both variant A1, A2, B and F. β-CN polymorphisms seems generally not to be the main contributors
to changes in coagulation properties of the milk on the same level of κ-CN polymorphism, but
correlations have been reported for β-CN A2, B and F. β-CN A2 and F have been coupled to poor
and non-coagulating milk samples, β-CN A2 often in combination with κ-CN A/E. A favorable effect

64
of β-CN B in terms of milk coagulation properties has been reported, often in combination with
κ-CN B, β-CN B and β-LG B (Bittante et al. 2012; Jensen et al. 2012a; Poulsen et al. 2013; H B
Jensen, Holland, et al. 2012; Bonfatti et al. 2010).
By combining the data from this study and the available literature, some major features can be
pointed out regarding the four discussed native Nordic breeds and the coagulation properties of
their milk:
SRP had the highest frequency of β-CN A2 together with κ-CN E, a combination which is associated
with poor or non-coagulating milk (Poulsen et al. 2013). Instead, SRP had a low RCT, with no
effect on G’40. High amounts of αs1-8P CN has been correlated to good coagulating milk
(Frederiksen et al. 2011), and a significantly higher proportion of αs1-8P CN compared to αs1-9P
CN was observed in SRP. This could be one of the contributing factors for a low RCT.
SM had a high β-CN B and κ-CN B variant frequency, which was not found in the other breeds in
this study; and they have been reported in a high content in good coagulating milk (Jensen et al.
2012a). In this study, milk containing a β-CN B protein genetic variant was related to a high κ-CN
content, together with a low RCT and high G’40 in β-CN A1B. Milk containing the κ-CN BB
genotype was also related to low RCT, low β-LG, high G’40, and high κ-CN in this study. Bobe et
al. (1999) have in accordance with these results reported a higher content of κ-CN in expense for
a low β-LG in the κ-CN BB genetic variant. Therefore, low RCT and high κ-CN content in SM was
to some degree related to the CN genotypic combination of the milk. SM had a low total and ionic
calcium content, which in this study seemed related to breed specific characteristics, as it did not
correspond to low RCT.
JUT had the highest frequency of κ-CN A of the four breeds. However, this was not combined
with a high variant frequency of β-CN A2, which is associated with poor or non-coagulating milk
(Poulsen et al. 2013), but contained a high β-CN A1 variant frequency. A high variant frequency of
F was also present in JUT. In this study, the β-CN A1F variant was related to a low κ-CN content
and high RCT. However, JUT was not related to either high or low RCT, and other factors than the

65
present of β-CN A1F seems to affect RCT in JUT. The κ-CN AA variant was related to high RCT, low
G’40, low κ-CN content and high β-LG content. Therefore, low κ-CN and high β-LG in JUT can be
related to the CN genotypic combination of the milk. Moreover, JUT had a low total calcium
concentration and κ-Glyco content, which both seemed to be related to breed specific
characteristics.
RDM had a high κ-CN A variant frequency, contained κ-CN E, had the highest β-CN F and the
highest β-LG variant frequency. In this study, genotypes containing β-CN F were related to a high
RCT, low G’40 and low κ-CN content. The genotype κ-CN AA was related to high RCT, low G’40,
low κ-CN content and high β-LG content. Therefore, the high RCT and high β-LG content could be
related to the CN genotypic combination of the milk. Moreover, RDM had a high total and ionic
calcium concentration and a high κ-Glyco content, which both seemed related to breed specific
characteristics. These properties have been associated with good coagulation properties (Jensen,
et al. 2012a; Gustavsson et al. 2014b), which were however not observed in this study.
It is of great importance for the dairy industry to produce as good coagulating milk as possible
for a better cheese production. Milk with a low rennet coagulation time (RCT) and high curd
firmness value (G’) are favorable during cheese production (Bittante et al. 2012). Mineral content
is also related to cheese production, where good coagulating milk contains a higher mineral
content, including higher concentrations of total and ionic calcium (Jensen, et al. 2012a;
Gustavsson et al. 2014b). Therefore, milk from SM could be of special interest in cheese
production, due to the low RCT and the high variant frequency of variant β-CN B and κ-CN B,
which are both related to high G’40, low RCT and high κ-CN content.
8 CONCLUSION
The present Masters thesis is a step towards better understanding of the genetic combination of
milk from native Nordic cattle. The objective of this master thesis was to study the protein profile
of milk from native nordic breeds, including protein genetic variants and post translational
modifications. This was investigated mainly by the use of LC-MS.

66
For this project, milk from four different native Nordic breeds was analyzed: Red Danish, Jutland
Breed, Swedish Mountain and Swedish Red Polled cattle. It was observed that the Native nordic
cattle had a diverse protein genetic combination of the protein forms of κ-, αs2-, αs1- and β-CN,
β-LG and α-LA. Based on their genotype and variant frequency combination, none of the four
breeds, JUT, RDM, SM, or SRP, showed signs of close relationships, as there was a different
variant frequency combination between the breeds. Interestingly, the presence of β-CN F has not
previously been reported in such high a frequency in modern Scandinavian dairy cattle breeds,
as observed in native Nordic cattle breeds.
The protein profiles were significantly different between the breeds. The relative contents of all
the major proteins together with the protein isoforms content were different between breeds,
except the β-CN content and κ- and β-CN genotype, which were stable between breeds. The
protein profile and the technological properties were different for various κ-CN genotype. The κ-
CN AA genotype was correlated with a low κ-CN content, high β-LG content, high RCT and low
G’40. κ-CN BB had the reverse effect. The effect of the β-CN genotype was much less pronounced,
but with the highest correlation in the genotype β-CN A1B to low RCT, high G’40 and high κ-CN
content, and in β-CN A1F to high RCT and low κ-CN content.
The concentration of calcium in native Nordic breeds was more dependent on breed specific
regulation, than dependent on the genotypic distribution of κ- and β-CN. There was no clear
relation between total or ionic calcium and RCT in native Nordic cattle breeds. SM and SRP had a
low RCT and RDM had a high RCT. In all three breeds this was related to the both breed specific
genetics and κ- and β-CN genotype distribution.
Different favorable and unfavorable traits have been observed in native Nordic cattle, and further
investigation is needed to determine possible favorable products from milk from native Nordic
cattle. Swedish Mountain cattle show many favorable traits for cheese making, which could
promote a population increase of this breed.

67
9 FURTHER PERSPECTIVES
Native Nordic cattle have a diverse genotypic composition of the major proteins in milk,
compared between the breeds.
Further studies within the occurrence of a high variant frequency β-CN F in Jutland cattle and Red
Danish cattle is needed. A subset of study with the influence of β-CN F on coagulation properties,
to clear the effect of β-CN F, also in correlation with the presence of β-CN F in non-coagulating
milk in Danish Holstein-Friesian cattle (Jensen et al. 2012a).
Milk containing only variant A2 of the β-CN commonly sold in some countries, due to a proteolytic
product originating from β-CN A1, which have been related to an increased risk of coronary heart
disease and type 1 diabetes (Bell et al. 2006). SRP have a high variant frequency of β-CN A2, which
makes the breed a good candidate within this niche production by selective genetic breeding.
10 REFERENCE
Anon, 2012. Food Analysis by HPLC, Third Edition, CRC Press. Available at: https://books.google.com/books?id=yBEeEqzGCnsC&pgis=1 [Accessed March 5, 2016].
Anon, Kvæg - NaturErhvervstyrelsen. Available at: http://naturerhverv.dk/landbrug/genetiske-ressourcer/husdyrgenetiske-ressourcer/husdyrarter/kvaeg/#c9101 [Accessed December 1, 2015].
Belitz, H.D., Grosch, W. & Schieberle, P., 2009. Food chemistry,
Bell, S.J., Grochoski, G.T. & Clarke, A.J., 2006. Health implications of milk containing beta-casein with the A2 genetic variant. Critical reviews in food science and nutrition, 46(1), pp.93–100. Available at: http://www-tandfonline-com.ez.statsbiblioteket.dk:2048/doi/abs/10.1080/10408390591001144 [Accessed March 21, 2016].
Berg, J.M., Tymoczko, J.L. & Stryer, L., 2007. Stryer Biochemie, Available at: http://books.google.com/books?id=jQKGAAAACAAJ.
Bett, R.C. et al., 2010. Trajectories of Evolution and Extinction in the Swedish Cattle Breeds. Available at: https://www.researchgate.net/publication/265634310_Trajectories_of_Evolution_and_Extinction_in_the_Swedish_Cattle_Breeds [Accessed March 18, 2016].
Bijl, E. et al., 2014. Phosphorylation of αS1-casein is regulated by different genes. Journal of

68
dairy science, 97(11), pp.7240–7246. Available at: http://www.ncbi.nlm.nih.gov/pubmed/25200775.
Bijl, E. et al., 2013. Protein, casein, and micellar salts in milk: current content and historical perspectives. Journal of dairy science, 96(9), pp.5455–64. Available at: http://www.ncbi.nlm.nih.gov/pubmed/23849643.
Bittante, G., Penasa, M. & Cecchinato, a, 2012. Invited review: Genetics and modeling of milk coagulation properties. Journal of dairy science, 95(12), pp.6843–70. Available at: http://www.ncbi.nlm.nih.gov/pubmed/23021752.
Bobe, G. et al., 1999. Effect of milk protein genotypes on milk protein composition and its genetic parameter estimates. Journal of dairy science, 82(12), pp.2797–804. Available at: http://www.sciencedirect.com/science/article/pii/S0022030299755372 [Accessed November 30, 2015].
Bobe, G. et al., 1998. Separation and Quantification of Bovine Milk Proteins by Reversed-Phase High-Performance Liquid Chromatography. Journal of agricultural and food chemistry, 46(2), pp.458–463. Available at: http://www.ncbi.nlm.nih.gov/pubmed/10554263.
Bonfatti, V. et al., 2010. Effects of beta-kappa-casein (CSN2-CSN3) haplotypes, beta-lactoglobulin (BLG) genotypes, and detailed protein composition on coagulation properties of individual milk of Simmental cows. Journal of dairy science, 93(8), pp.3809–3817. Available at: http://dx.doi.org/10.3168/jds.2009-2779.
Bonfatti, V. et al., 2011. Genetic analysis of detailed milk protein composition and coagulation properties in Simmental cattle. Journal of dairy science, 94(10), pp.5183–93. Available at: http://www.ncbi.nlm.nih.gov/pubmed/21943768.
Bonfatti, V. et al., 2008. Validation of a new reversed-phase high-performance liquid chromatography method for separation and quantification of bovine milk protein genetic variants. Journal of Chromatography A, 1195(1-2), pp.101–106.
Bonfatti, V., Chiarot, G. & Carnier, P., 2014. Glycosylation of κ-casein: Genetic and nongenetic variation and effects on rennet coagulation properties of milk. Journal of dairy science, pp.1–9. Available at: http://www.ncbi.nlm.nih.gov/pubmed/24508440.
Brüniche-Olsen, a., Gravlund, P. & Lorenzen, E.D., 2012. Impacts of genetic drift and restricted gene flow in indigenous cattle breeds: evidence from the Jutland breed. Animal Genetic Resources/, 50(July 2011), pp.75–85. Available at: http://www.journals.cambridge.org/abstract_S2078633611000373.
Caroli, A.M., Chessa, S. & Erhardt, G.J., 2009. Invited review: Milk protein polymorphisms in cattle: Effect on animal breeding and human nutrition. Journal of Dairy Science, 92(11), pp.5335–5352. Available at: http://linkinghub.elsevier.com/retrieve/pii/S0022030209708653.
Coolbear, K.P., Elgar, D.F. & Ayers, J.S., 1996. Profiling of genetic variants of bovine κ-casein macropeptide by electrophoretic and chromatographic techniques. International Dairy

69
Journal, 6(11-12), pp.1055–1068. Available at: http://www.sciencedirect.com/science/article/pii/S0958694696000349 [Accessed March 14, 2016].
Day, L. et al., 2015. Casein polymorphism heterogeneity influences casein micelle size in milk of individual cows. Journal of dairy science, 98(6), pp.3633–44. Available at: http://www.sciencedirect.com/science/article/pii/S0022030215002167.
Demeter, R.M. et al., 2010. Relationships between milk protein composition, milk protein variants, and cow fertility traits in Dutch Holstein-Friesian cattle. Journal of dairy science, 93(11), pp.5495–502. Available at: http://www.sciencedirect.com/science/article/pii/S0022030210005916.
FAO, F. and A.O. of the U.N., 2007. THE STATE OF THE WORLD’s ANIMAL GENETIC RESOURCES FOR FOOD AND AGRICULTURE. Available at: http://www.fao.org/docrep/010/a1250e/a1250e00.htm [Accessed November 26, 2015].
FAO: http:/www.fao.org/docrep/meeting/027/mg015e/pdf
Farrell, H.M. et al., 2004. Nomenclature of the Proteins of Cows’ Milk—Sixth Revision. Journal of Dairy Science, 87(6), pp.1641–1674. Available at: http://linkinghub.elsevier.com/retrieve/pii/S0022030204733196.
Fox, P.F. & Mcsweeney, P.L.H., 1998. Dairy Chemistry and Biochemistry. Dairy Chemistry and Biochemistry, p.478.
Frederiksen, P.D. et al., 2011. Composition and effect of blending of noncoagulating, poorly coagulating, and well-coagulating bovine milk from individual Danish Holstein cows. Journal of dairy science, 94(10), pp.4787–99. Available at: http://www.ncbi.nlm.nih.gov/pubmed/21943730.
Gallinat, J.L. et al., 2013. DNA-based identification of novel bovine casein gene variants. Journal of dairy science, 96(1), pp.699–709. Available at: http://www.ncbi.nlm.nih.gov/pubmed/23102962.
Gaucheron, F., 2005. The minerals of milk. Reproduction, nutrition, development, 45(4), pp.473–83. Available at: http://dx.doi.org/10.1051/rnd:2005030.
Georgiou, C.A. & Danezis, G.P., 2015. Advanced Mass Spectrometry for Food Safety and Quality, Available at: http://www.sciencedirect.com/science/article/pii/B9780444633408000030.
Graml, R., Pirchner, F. 2003. Effects of milk protein loci on content of their proteins. Arch. Tierz., Dummerstorf 46 (2003) 4, 331-340
Gustavsson, F. et al., 2014a. Effects of breed and casein genetic variants on protein profile in milk from Swedish Red, Danish Holstein, and Danish Jersey cows. Journal of dairy science, 97(6), pp.3866–77. Available at: http://www.sciencedirect.com/science/article/pii/S0022030214002367 [Accessed November 27, 2015].

70
Gustavsson, F. et al., 2014b. Factors influencing chymosin-induced gelation of milk from individual dairy cows: Major effects of casein micelle size and calcium. International Dairy Journal, 39(1), pp.201–208. Available at: http://linkinghub.elsevier.com/retrieve/pii/S0958694614001290\nhttp://www.sciencedirect.com/science/article/pii/S0958694614001290.
Hallén, E. et al., 2008. Effect of beta-casein, kappa-casein and beta-lactoglobulin genotypes on concentration of milk protein variants. Journal of animal breeding and genetics = Zeitschrift fur Tierzuchtung und Zuchtungsbiologie, 125(2), pp.119–29. Available at: http://www.ncbi.nlm.nih.gov/pubmed/18363977 [Accessed March 17, 2016].
Heck, J.M.L. et al., 2009. Effects of milk protein variants on the protein composition of bovine milk. Journal of dairy science, 92(3), pp.1192–1202. Available at: http://dx.doi.org/10.3168/jds.2008-1208.
Heck, J.M.L. et al., 2008. Estimation of variation in concentration, phosphorylation and genetic polymorphism of milk proteins using capillary zone electrophoresis. International Dairy Journal, 18(5), pp.548–555. Available at: <Go to ISI>://000254982200014\nhttp://ac.els-cdn.com/S0958694607002063/1-s2.0-S0958694607002063-main.pdf?_tid=82f66dfb6b851fd510564d7b1821e4b8&acdnat=1335392828_2ad0c89b35ef5a153324b0020804ece9.
Holland, J.W., Deeth, H.C. & Alewood, P.F., 2006. Resolution and characterisation of multiple isoforms of bovine kappa-casein by 2-DE following a reversible cysteine-tagging enrichment strategy. Proteomics, 6(10), pp.3087–95. Available at: http://www.ncbi.nlm.nih.gov/pubmed/16619295 [Accessed March 17, 2016].
Horne, D.S., 2006. Casein micelle structure: Models and muddles. Current Opinion in Colloid & Interface Science, 11(2-3), pp.148–153. Available at: http://www.sciencedirect.com/science/article/pii/S1359029405001196 [Accessed January 27, 2016].
Ikonen, T. et al., 1999. Genetic Parameters for the Milk Coagulation Properties and Prevalence of Noncoagulating Milk in Finnish Dairy Cows. Journal of Dairy Science, 82(1), pp.205–214. Available at: http://www.sciencedirect.com/science/article/pii/S0022030299752252\nhttp://www.journalofdairyscience.org/article/S0022-0302(99)75225-2/pdf.
Jensen, H.B. et al., 2012b. Comparative proteomic analysis of casein and whey as prepared by chymosin-induced separation, isoelectric precipitation or ultracentrifugation. Journal of Dairy Research, 79(04), pp.451–458.
Jensen, H.B., Poulsen, N. a, et al., 2012a. Distinct composition of bovine milk from Jersey and Holstein-Friesian cows with good, poor, or noncoagulation properties as reflected in protein genetic variants and isoforms. Journal of dairy science, 95(12), pp.6905–17. Available at: http://www.ncbi.nlm.nih.gov/pubmed/23040012.
Jensen, H.B. et al., 2015. Genetic variation and posttranslational modification of bovine kappa -

71
casein: effects on caseino-macropeptide release during renneting. Journal of Dairy Science, 98(2), pp.747–758. Available at: http://www.sciencedirect.com/science/article/pii/S0022030214008509.
Jensen, H.B., Holland, J.W., et al., 2012. Milk protein genetic variants and isoforms identified in bovine milk representing extremes in coagulation properties. Journal of dairy science, 95(6), pp.2891–903. Available at: http://www.sciencedirect.com/science/article/pii/S0022030212002652 [Accessed November 27, 2015].
Kantanen, J. et al., 2000. Genetic diversity and population structure of 20 North European cattle breeds. The Journal of heredity, 91(6), pp.446–57. Available at: http://www.ncbi.nlm.nih.gov/pubmed/11218082.
Kortegaard, K. 2004. A study on the composition of milk from the native Danish Jutland cattle SJM and comparison with contemporary SDM - Danish Holstein. Master thesis, The Royal Danish Veterinary and Agricultural University.
Koutina, G. et al., 2015. Influence of colloidal calcium phosphate level on the microstructure and rheological properties of rennet-induced skim milk gels. LWT - Food Science and Technology, 63(1), pp.654–659. Available at: http://www.sciencedirect.com/science/article/pii/S0023643815001887 [Accessed January 27, 2016].
Koutina, G. et al., 2014. Temperature effect on calcium and phosphorus equilibria in relation to gel formation during acidification of skim milk. International Dairy Journal, 36(1), pp.65–73.
Kruve, A. et al., 2015. Tutorial review on validation of liquid chromatography-mass spectrometry methods: part I. Analytica chimica acta, 870, pp.29–44. Available at: http://www.sciencedirect.com/science/article/pii/S0003267015001956 [Accessed March 2, 2016].
Lewis, M.J., 2011. The measurement and significance of ionic calcium in milk - A review. International Journal of Dairy Technology, 64(1), pp.1–13.
Lien, S. et al., 1999. Comparison of milk protein allele frequencies in nordic cattle breeds. Animal Genetics, 30(2), pp.85–91.
Lin, M.J., Lewis, M.J. & Grandison, A.S., 2006. Measurement of ionic calcium in milk. International Journal of Dairy Technology, 59(3), pp.192–199.
Metrohm (2016) Application Bulletin 235/3 e. Available online:
http://partners.metrohm.com/GetDocument?action=get_dms_document&docid=1938468.
Accessed in 18-03-2016
NG-Kwai-Hang, K.F. et al., 1987. Variation in milk protein concentrations associated with genetic polymorphism and environmental factors. Journal of dairy science, 70(3), pp.563–70. Available at: http://www.ncbi.nlm.nih.gov/pubmed/3584600.

72
Ng-Kwai-Hang, K.F. & Kim, S., 1996. Different amounts of ??-lactoglobulin A and B in milk from heterozygous AB cows. International Dairy Journal, 6(7), pp.689–695.
O’Donnell, R. et al., 2004. Milk proteomics. International Dairy Journal, 14(12), pp.1013–1023. Available at: http://www.sciencedirect.com/science/article/pii/S0958694604000871 [Accessed March 5, 2016].
Oklahoma Animal Science Department,
http://www.ansi.okstate.edu/breeds/cattle/swedishredpolled
Pertoldi, C. et al., 2014. Genetic characterization of a herd of the endangered Danish Jutland cattle. Journal of animal science, 92(6), pp.2372–6. Available at: https://www-animalsciencepublications-org.ez.statsbiblioteket.dk:12048/publications/jas/abstracts/92/6/2372 [Accessed November 26, 2015].
Poulsen, N. a et al., 2013. The occurrence of noncoagulating milk and the association of bovine milk coagulation properties with genetic variants of the caseins in 3 Scandinavian dairy breeds. Journal of dairy science, 96(8), pp.4830–42. Available at: http://www.sciencedirect.com/science/article/pii/S0022030213004062.
Sandra, S. et al., 2012. Effect of soluble calcium on the renneting properties of casein micelles as measured by rheology and diffusing wave spectroscopy. Journal of dairy science, 95, pp.75–82. Available at: http://www.ncbi.nlm.nih.gov/pubmed/22192185.
Sørensen, L.H. & Nielsen, V.H., 2012. Danske Husdyrgenetiske Ressourcer. Available at: http://www.researchgate.net/publication/268387096_Danske_Husdyrgenetiske_Ressourcer [Accessed November 25, 2015].
Visser, S. et al., 1995. Identification of a new genetic variant of bovine beta-casein using reversed-phase high-performance liquid chromatography and mass spectrometric analysis. Journal of chromatography. A, 711(1), pp.141–50. Available at: http://www.ncbi.nlm.nih.gov/pubmed/7496485.
Walsh, C.D. et al., 1998. Influence of κ-casein genetic variant on rennet gel microstructure, Cheddar cheesemaking properties and casein micelle size. International Dairy Journal, 8(8), pp.707–714.
Walstra, P., Wouters, J.T.M. & Geurts, T.J., 2006. Dairy Science and Technology, Available at: http://www.worldcat.org/isbn/9780824727635.
Wedholm, a et al., 2006. Effect of protein composition on the cheese-making properties of milk from individual dairy cows. Journal of dairy science, 89(9), pp.3296–3305.