prefrontal cortical projections to the hypothalamus in macaque monkeys
TRANSCRIPT

Prefrontal Cortical Projections to theHypothalamus in Macaque Monkeys
D. ONGUR, X. AN, AND J.L. PRICE*Department of Anatomy and Neurobiology, Washington University School of Medicine,
St. Louis, Missouri, 63110
ABSTRACTThe organization of projections from the macaque orbital and medial prefrontal cortex
(OMPFC) to the hypothalamus and related regions of the diencephalon and midbrain wasstudied with retrograde and anterograde tracing techniques. Almost all of the prefrontalcortical projections to the hypothalamus arise from areas within the ‘‘medial prefrontalnetwork,’’ as defined previously by Carmichael and Price ([1996] J. Comp. Neurol. 371:179–207). Outside of the OMPFC, only a few neurons in the temporal pole, anterior cingulate andinsular cortex project to the hypothalamus. Axons from the OMPFC also innervate the basalforebrain, zona incerta, and ventral midbrain.
Within the medial prefrontal network, different regions project to distinct parts of thehypothalamus. The medial wall areas 25 and 32 send the heaviest projections to thehypothalamus; axons from these areas are especially concentrated in the anterior hypotha-lamic area and the ventromedial hypothalamic nucleus. Orbital areas 13a, 12o, and Iai, whichare related to the medial prefrontal network, selectively innervate the lateral hypothalamicarea, especially its posterior part. The cellular regions of the paraventricular, supraoptic,suprachiasmatic, arcuate, and mammillary nuclei are conspicuously devoid of cortical axons,but many axons abut the borders of these nuclei and may contact dendrites that extend fromthem. Areas within the orbital prefrontal network on the posterior orbital surface andagranular insula send only weak projections to the posterior lateral hypothalamic area. Therostral orbital surface does not contribute to the cortico-hypothalamic projection. J. Comp.Neurol. 401:480–505, 1998. r 1998 Wiley-Liss, Inc.
Indexing terms: prefrontal cortex; autonomic nervous system; feeding; emotions; amygdala;
Macaca fascicularis
The medial and ventral parts of the frontal lobe havelong been recognized to have the ability to influence theautonomic nervous system in mammals. Kaada et al.(1949) found that stimulation of a large continuous strip ofcortex in the temporal pole, the agranular insula, theposterior orbital cortex, the ventral part of the medialsurface of the frontal lobe, and the anterior cingulatecortex causes changes in blood pressure, heart rate, andrespiration rate in monkeys. A large number of stimula-tion studies in rats, rabbits, cats, and monkeys support thecontention that these areas can modulate autonomic pa-rameters (Wall and Davis, 1951; Delgado, 1960; Neafsey,1990 and references therein; Yasui et al., 1991).
A further indication of the role of the ventromedialfrontal lobe in autonomic function is that individuals withlesions of this region are unable to generate autonomicresponses to emotional stimuli. Remarkably, they are alsoimpaired in making judgments about the consequences oftheir actions in social situations, despite possessing theknowledge necessary to make the correct decision (Dama-
sio et al., 1990; Bechara et al., 1994; Damasio, 1994).Damasio and his coworkers have suggested (Bechara etal., 1997) that these two deficits are related and that thesociopath-like behavior of these patients is due to theirinability to generate ‘‘somatic markers’’ (autonomic andother internal changes) that tag behavioral options asdesirable or not.
Although anatomical studies of prefrontal projections toautonomic areas in primates (e.g., Johnson et al., 1968;Nauta, 1969) were not very detailed, studies in rats showthat both medial and lateral prefrontal/insular corticalareas project to the hypothalamus and the periaqueductal
Grant sponsor: USNIH; Grant number: DC000093; Grant sponsor:Division of Biology and Biomedical Sciences of Washington University;Grant sponsor: McDonnell Center for Higher Brain Function.
*Correspondence to: Dr. J.L. Price, Department of Anatomy and Neurobi-ology, Washington University School of Medicine, 660 S. Euclid Ave., St.Louis, MO 63110. E-mail: [email protected]
Received 10 February 1998; Revised 3 June 1998; Accepted 4 June 1998
THE JOURNAL OF COMPARATIVE NEUROLOGY 401:480–505 (1998)
r 1998 WILEY-LISS, INC.

gray (PAG), both of which are well known to generatecoordinated autonomic and somatic responses to emo-tional stimuli (Beckstead, 1979; Kita and Oomura, 1981;Sesack et al., 1989; Neafsey, 1990; Hurley et al., 1991;Price et al., 1991; Allen et al., 1991; Bacon and Smith,1993). Further, projections from the prefrontal cortex(PFC) in rats have been observed to reach more caudalregions involved in autonomic control, including the para-brachial nucleus and the nucleus of the solitary tract in thebrainstem and even the intermediolateral cell column inthe spinal cord (Van Der Kooy et al., 1982; Terreberry andNeafsey, 1987; Hurley et al., 1991). Functional studies inthe rat have demonstrated a monosynaptic pathway fromthe PFC to the hypothalamus and shown that stimulationof the PFC causes autonomic and somatic changes medi-ated by the hypothalamus (Kita and Oomura, 1981; Hardyand Holmes, 1988; Cechetto and Chen, 1990; DeBruin,1990). Based on these results, Neafsey (1990) has proposedthat parts of the PFC constitute a ‘‘visceromotor’’ cortex.
Recent studies of the anatomical structure and connec-tions of the orbital and medial prefrontal cortex (OMPFC)in monkeys have defined a large number of cortical areaswith distinctive afferent and efferent connections. Basedon their differential connections with other prefrontalareas (Carmichael and Price, 1996), and with other brainregions such as the mediodorsal thalamus and the amyg-dala (Ray and Price, 1993; Carmichael and Price, 1995a),these areas can be divided into two distinct but overlap-ping networks. The orbital prefrontal network includesmost areas on the orbital surface and receives visual,somatosensory, gustatory, olfactory, and visceral inputs.The medial prefrontal network consists of areas on the medial
wall of the frontal lobe and a few selected areas on the orbitalsurface that receive few if any direct sensory inputs.
In this study, the origin and termination of prefrontalcortical projections to the hypothalamus have been demon-strated in monkeys with retrograde and anterograde axo-nal tracers, injected into several parts of the hypothala-mus, and into restricted areas of the prefrontal cortex,respectively. In particular, the origin of the projections wasdefined in relation to the previously described architec-tonic areas of the OMPFC, and to the medial and orbitalprefrontal networks. The results indicate that almost all ofthe prefrontal projections to the hypothalamus arise fromareas of the medial prefrontal network.
MATERIALS AND METHODS
Animals
Macaca fasicularis adults of either sex weighing be-tween 2 and 3.5 kg. were used in this study. All animalprotocols were reviewed and approved by the AnimalStudies Commitee of Washington University. The animalswere initially anesthetized with ketamine (10 mg/kg i.m.)and xylazine (0.67 mg/kg i.m.). They were then placed in astereotactic apparatus and anesthesia was maintainedwith a gaseous mixture of oxygen, nitrous oxide, and 1%halothane. After surgery, a long-lasting analgesic wasgiven (buprenorphine, 0.1 mg/kg i.m.) before the animalwas brought out of anesthesia.
Injections
The procedure for the microinjection of tracers has beendescribed previously (Carmichael et al., 1994). Briefly,
Abbreviations
AAA anterior amygdaloid areaAB accessory basal nucleus of amygdalaac anterior commissureAHA anterior hypothalamic areasAONl or
AONm anterior olfactory nucleus, lateral and medial subdivisionsArh arcuate hypothalamic nucleusB (i, mg, or
pc) basal nucleus of the amygdala (magnocellular, intermedi-ate, or parvicellular subdivision)
BDA biotinylated dextran amineBNST bed nucleus of the stria terminalisBvl basal amygdaloid nucleus, ventrolateral partCa caudate nucleusCe (m/l) central nucleus of the amygdala (medial/lateral)CN3 cranial nerve IIICoA cortical nucleus of amygdalaCP cerebral peduncleDH dorsal hypothalamusDMH dorsomedial hypothalamic nucleusDLPFC dorsolateral prefrontal cortexdlPAG dorsal column of PAGEn endopiriform nucleusFB Fast Blue (tracer)FR Fluoro Ruby (tracer)fx fornixG gustatory cortexGP (i/e) globus pallidus (pars interna/externa)H hippocampal formation3H-leu tritiated leucineIal, Iapl, Iai,
Iam, Iapm agranular insula, lateral, posterolateral, intermediate, me-dial and posteromedial subdivisions
ic internal capsuleIP interpeduncular nucleusL lateral nucleus of amygdala
LH lateral hypothalamusLHA lateral hypothalamic arealPAG lateral column of periaqueductal grayLPOA lateral preoptic areaLV lateral ventricleMe medial nucleus of the amygdalaMH medial hypothalamusMN mammillary nucleiMH medial hypothalamusMPOA medial preoptic areaNBM nucleus basalis of Meynertoc optic chiasmOMPFC orbital and medial prefrontal cortexot optic tractOT olfactory tuberclePAC periamygdaloid cortexPAG periaqueductal grayPC piriform cortexPFC prefrontal cortexPH posterior hypothalamusPrCO precentral operculumPVN paraventricular hypothalamic nucleusRN Red nucleusS or Sub subiculumSC superior colliculusSCN suprachiasmatic nucleusSON supraoptic nucleusSN, r or c substantia nigra, pars reticulata or compactaSTN subthalamic nucleusTP (g, d, or a) temporal polar cortex (granular, dysgranular, or agranular
parts)vlPAG ventrolateral column of periaqueductal grayVMH ventromedial hypothalamic nucleusVTA ventral tegmental areaZI zona incerta
PREFRONTAL PROJECTIONS TO THE HYPOTHALAMUS 481

burr holes were introduced at the appropriate site in theskull guided by the atlas of Szabo and Cowan (1984).Electrophysiological recording of spontaneous, multiunitactivity allowed determination of structural boundariesand adaptation of the atlas to the individual animal.Anterograde and retrograde axonal tracers were pressure-injected through micropipettes. The size of the injectionwas determined from the known inner diameter of thepipette combined with monitoring the change in the levelof the meniscus. It ranged from 200 to 1,000 nl dependingon the structure injected and the properties of the tracerbeing used. The anterograde tracers were biotinylateddextran amine (BDA), Fluoro Ruby (FR), and tritiatedleucine (3H-leu); the retrograde tracers were Fast Blue(FB) and Diamidino Yellow (DY).
Perfusion and tissue processing
After a survival period of 14 or 28 days, anesthesia wasinduced with ketamine (10 mg/kg i.m.) and xylazine (0.2mg/kg i.m.), and deep anesthesia was maintained withsodium pentobarbital (25–30 mg/kg i.v. or i.p.). The ani-mals were then perfused transcardially with phosphate-buffered saline and a sequence of 4% paraformaldehydesolutions, first at pH 6.5 and then at pH 9.5. The brainswere transferred through 5% and 30% sucrose solutionsover 48 hours at 4°C, frozen in isopentane and dry ice andcut on a freezing microtome. For each brain, several seriesof 50-µm sections were cut. These series were processed forNissl, acetylcholinesterase, myelin, or one of several immu-nohistochemical stains for detection of cytoarchitectonicborders. Other series were processed for fluorescence,autoradiography, or to demonstrate BDA label for detec-tion of anatomical tracers. In the cases in which 3H-leuwas used as a tracer, autoradiography was used to demon-strate the radioactive label (Cowan et al., 1972), with anexposure period of 6 weeks. In cases where BDA was usedas a tracer, the label was demonstrated with the avidin-biotin method previously described (Carmichael et al.,1994).
Methods of analysis
Labeled cells, fibers, or axonal varicosities from coronalsections through the OMPFC or the hypothalamus wereplotted on a microscope digitizer system with encodersattached to the microscope stage and interfaced with apersonal computer (Minnesota Datametrics, St Paul, MN).Cortical boundaries and other landmarks were added tothese plots using camera lucida drawings of adjacentsections stained for Nissl, myelin, and acetylcholinester-ase (AChE). With retrograde tracers, each labeled cell bodywas mapped as a point. In anterograde cases, axonalvaricosities in the hypothalamus were marked in the samemanner. This was possible because the number of labeledvaricosities was not overwhelmingly large in a givenhigh-power field. Because this type of mapping was notpossible for axons labeled autoradiographically by triti-ated amino acid injections, only the location of labeledfibers were mapped in these cases.
In this study, we adhered to the divisions of the OMPFCdelineated recently by Carmichael and Price (1994). Thesedivisions were made on the basis of patterns of differentialcyto- and chemoarchitectonics. In all, the OMPFC wasshown to contain more than 20 areas with distinctivestructure and connections.
Unfolded maps of the prefrontal cortex were created foreach retrograde tracer experiment, based on the techniqueof VanEssen and Maunsell (1980). A line was tracedthrough the middle layer of the cortex in individualsections, with a cut introduced at the depth of the principalsulcus. Cortical areal boundaries as well as labeled cellcounts were carried over to the unfolded map to create asingle representation of the whole PFC. Labeled cells werecounted in each 1-mm-wide column of cortex and plottedon the unfolded map. If the column contained no cells, thecorresponding area on the map was left blank. If itcontained between 1 and 5 cells, it was marked by a dot onthe map; if it had between 5 and 10, it was representedwith a plus sign, and if it had more than 10 labeled cells,with a plus sign and a square around it.
The density of labeled cells, fibers, or varicosities incortical and hypothalamic areas was quantified with acustomized computer program that counted points withindesignated and outlined areas on the section maps. Thesecounts were not corrected for section thickness and pos-sible double counting, and they should therefore be consid-ered as relative instead of absolute values. Because of this,and because of inherent differences between cases infactors such as the size and efficacy of the injections, thenumbers were used only for analysis of the relative patternof labeling within a given case. The density of labeled cellsin each cortical area or the number of labeled axonalvaricosities within the parts of the hypothalamus wastherefore scaled to the maximum value within that brain.
For this analysis, the hypothalamus was divided intomedial, dorsal, and lateral zones. The medial zone in-cluded the medial preoptic area (MPOA) anteriorly, andthe ventromedial hypothalamic nucleus (VMH) and thepre- and supramammillary nuclei posteriorly. The dorsalzone included the dorsomedial hypothalamic nucleus(DMH) and the dorsal hypothalamic area lateral to theparaventricular nucleus (PVN), the medial part of theperifornical area and the dorsal part of the posteriorhypothalamus. The lateral zone included the lateral preop-tic and lateral hypothalamic areas (LPOA and LHA).Nuclei not having label in any injection (such as the PVN,arcuate nucleus [Arh] or mammillary nuclei [MN]) wereexcluded from the area analyzed. In some anterogradetracer experiments, camera lucida drawings at severalhypothalamic levels were made by hand to demonstratethe organization of labeled fibers.
RESULTS
Retrograde tracer experiments
Injections of retrograde axonal tracers were made intothe hypothalamus in five monkeys, distributed from theanterior hypothalamus and preoptic area, to the caudaledge of the hypothalamus and adjacent ventral midbrain.In all cases, there are labeled cells in the area contralat-eral to the injection and in parts of the hypothalamus,basal forebrain, and ventral tegmentum. All of theseinjections also produce considerable numbers of labeledcells in the OMPFC, but the pattern varies with theanteroposterior placement of the injection. The more ros-tral injections label a restricted number of cytoarchitec-tonic areas, including areas in the medial prefrontalcortex, and select areas on the orbital surface. In contrast,the injections in more caudal parts of the hypothalamuslabel cells in a more extensive region of cortex, including
482 D. ONGUR ET AL.

all of the areas in the posterior orbital and agranularinsular cortex as well as the medial prefrontal areas; thereis less substantial labeling in the central and rostralorbital areas. Outside of the OMPFC, the amygdala,entorhinal cortex, subiculum, and other medial temporallobe structures contain cells labeled from the hypothala-mus, and in some cases labeled cells are found in theanterior cingulate cortex, temporal polar cortex, and dys-granular insula.
In all cases, the retrogradely labeled cells are largeneurons, concentrated in layer V, in a pattern very similarto that of cells labeled from the PAG (An et al., 1998). Inthe region of the agranular insula, many cells are alsolabeled deep to the cortex, in the anterior part of theclaustrum, and some labeled cells are scattered throughlayer VI and the white matter.
OM24. In this monkey, an injection of the tracer fastblue was placed in the anterodorsal part of LHA, whichinvolves the perifornical region just rostral to the level ofthe ventromedial hypothalamic nucleus (Fig. 1). The areasthat show intense label include area 25, and the posteriorportion of area 32 on the medial wall. On the orbitalsurface, label is concentrated in areas Iai, 14c, followed byareas 13a and 12o. Although there are a few scattered cellsin other parts of the agranular insula and posterior orbitalcortex, the density of cells in these areas is much lower.The overall areal pattern of label observed in this case istherefore very selective, avoiding much of the agranularinsula, the central and rostral orbital cortex, the anteriorcingulate cortex, and the dorsolateral PFC.
There are no labeled cells in the cortex of the temporalpole and the caudal part of the anterior cingulate cortex.The dysgranular insular cortex in the depth of the inferiorlimiting sulcus contains a few scattered labeled cells(Fig. 6).
OM37. In this case, an injection of fast blue was placedin the medial part of the anterior hypothalamus, involvingprimarily the medial preoptic area and the anterior hypo-thalamic area. The pattern of label in the OMPFC issimilar to that seen in case OM24, although there are morelabeled cells in the medial PFC, and fewer cells in theorbital cortical areas (Fig. 2). Medial areas 25, 32, and 10mare particularly heavily labeled, with the largest totalnumber of labeled cells. These are followed in intensity oflabel by medial areas 24b, 14c, 14r, and 9. The only areason the orbital surface with substantial numbers of labeledcells are Iai and to a lesser extent 13a. Scattered cells arealso found in areas 13b, Iam, and Iapm.
There are no labeled cells in the agranular temporalpolar cortex in this case, and only a small number oflabeled neurons are found in the ventral part of the insula.In contrast, there are a large number of cells in thegranular temporal pole, the perirhinal cortex, and theinferior temporal gyrus. This label is not seen in any othercases with hypothalamic injections, and may be due to thefact that the injection site also involved the anteriorcommissure.
OM27. The injection of the retrograde tracer fast bluein this case involved the lateral part of the ventromedialhypothalamic nucleus (VMH), posterior to both of thepreceding injections (Fig. 3). It also extended to thecell-sparse shell lateral to the nucleus. It produced a morewidespread pattern of cell labeling in the OMPFC than thetwo more anterior injections. In particular, many labeledcells are found throughout the posterior orbital and ante-
rior insular regions as well as in the medial wall. Medialnetwork areas 32 and 25 again have the largest number oflabeled cells, followed by other medial areas 24b, 10m, 14c,and the related orbital areas Iai and 12o. Within theorbital network, areas Iam, Iapm, Ial, Iapl, 13b, 13m, and14r are also substantially labeled. There are smallernumbers of labeled cells in the caudal parts of areas 11l,11m, 10o, and 10m, but these decrease rostrally. There arerelatively few labeled cells in areas 12r, 12m, 12l, and 13l,and there is virtually no label in the dorsolateral prefron-tal cortex.
Moderate numbers of labeled cells are found in theanterior cingulate and in the agranular temporal polarcortex (not shown). There are a few labeled cells in theventral, dysgranular insula as well.
OM38. This animal received an injection of fast bluelateral to the fornix, near the caudal edge of the ventrome-dial nucleus. The large injection site involved the lateraland dorsal hypothalamic areas. As in case OM27, but incontrast to OM24 and OM37, there are substantial num-bers of labeled cells throughout much of the OMPFC (Fig.4). On the medial wall, areas 25, 32, 24b, and 14c areheavily labeled. There are also many labeled cells in thecaudal part of areas 14r and 10m, although the labelbecomes lighter more rostrally. The rostral and centralorbital areas contain fewer cells than the medial wall, butin the caudal orbital cortex, areas 13a and 12o, andagranular insular areas Iai, Iapm, Iam, and Ial are heavilylabeled. Areas 13b, 13m, and 13l contain many labeledcells as well. On the dorsomedial surface of the frontallobe, area 9 contains substantial numbers of cells. Thereare also scattered cells in most parts of the dorsolateralprefrontal cortex (DLPFC).
Outside the prefrontal cortex, a number of labeled cellsare found in the caudal agranular insular cortex and theanterior cingulate cortex (Fig. 6). This label extends cau-dally to area 23b on the posterior cingulate gyrus and todysgranular portions of the insula in the depth of theinferior limiting sulcus. There are also scattered cells inthe agranular temporal pole. Most of the rostral basalforebrain, including the shell of the nucleus accumbens, isalso strongly labeled in this case, although the cells aremostly devoid of label in other cases.
OM26. A fast blue injection was centered just caudal tothe lateral hypothalamus in this case (Fig. 5). This is atransitional area between the hypothalamus and theventral midbrain and is close to the dopaminergic cellbodies of the ventral tegmental area and the substantianigra. Part of the injection site also extended rostrally intothe caudal LHA. This injection produced a pattern of labelsimilar to that of OM27 and OM38. There are largenumbers of labeled cells in the posterior medial and orbitalareas, including the agranular insula. Areas 32, 24b, 12o,and Iai are particularly heavily labeled. The number oflabeled cells decreases markedly in the central and rostralorbital areas, in the rostral part of the medial areas and inthe dorsolateral prefrontal areas.
As in the previous case, there are many labeled cells inthe anterior cingulate cortex but in this case, label extendsinto the premotor areas on the dorsomedial and dorsolat-eral parts of the hemisphere (Fig. 5). Scattered labeledcells are also found in the agranular temporal pole and theventral insula. The wider variety of cortical areas containing labeled cells in this case indicates that there is more
PREFRONTAL PROJECTIONS TO THE HYPOTHALAMUS 483
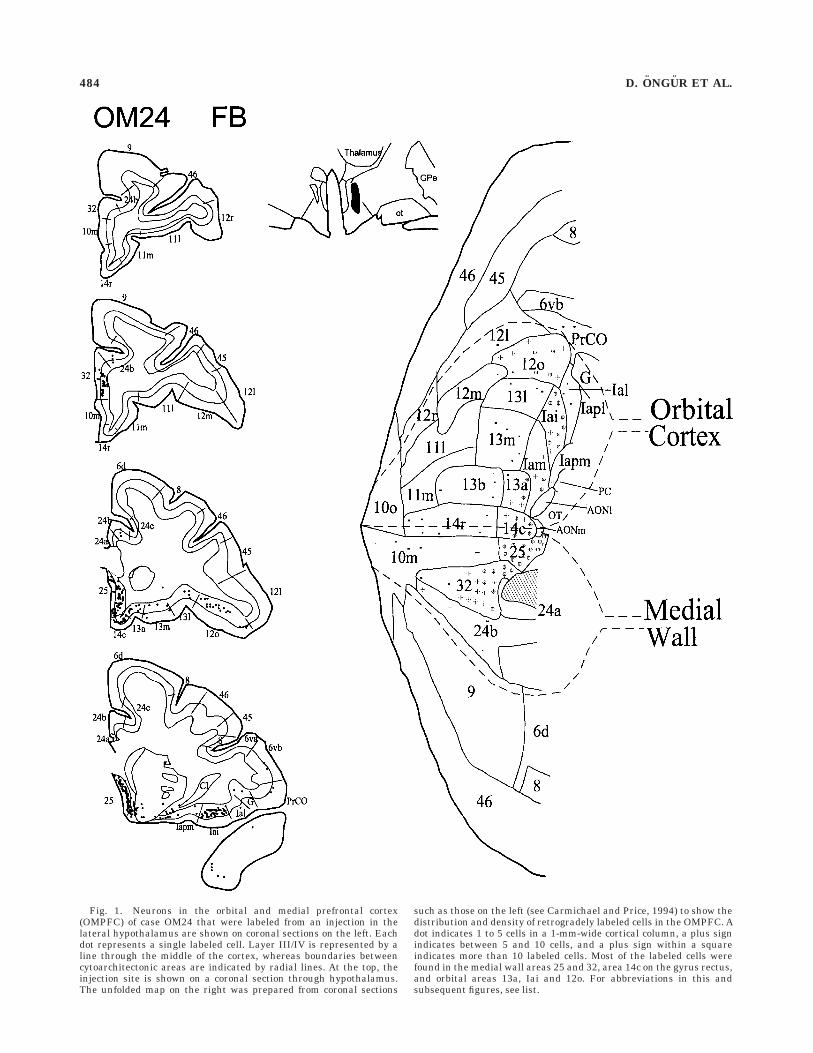
Fig. 1. Neurons in the orbital and medial prefrontal cortex(OMPFC) of case OM24 that were labeled from an injection in thelateral hypothalamus are shown on coronal sections on the left. Eachdot represents a single labeled cell. Layer III/IV is represented by aline through the middle of the cortex, whereas boundaries betweencytoarchitectonic areas are indicated by radial lines. At the top, theinjection site is shown on a coronal section through hypothalamus.The unfolded map on the right was prepared from coronal sections
such as those on the left (see Carmichael and Price, 1994) to show thedistribution and density of retrogradely labeled cells in the OMPFC. Adot indicates 1 to 5 cells in a 1-mm-wide cortical column, a plus signindicates between 5 and 10 cells, and a plus sign within a squareindicates more than 10 labeled cells. Most of the labeled cells werefound in the medial wall areas 25 and 32, area 14c on the gyrus rectus,and orbital areas 13a, Iai and 12o. For abbreviations in this andsubsequent figures, see list.
484 D. ONGUR ET AL.

Fig. 2. Representative coronal sections through the prefrontalcortex from case OM37 (on the left) illustrate cells labeled from aninjection in the anterior hypothalamus. The injection site also involvedthe medial preoptic area and the anterior commissure. The unfoldedmap on the right shows that labeled cells were seen in large numbers
in medial wall areas 25, 32, 24b, and 10m as well as areas 14c, 14r, andIai. This pattern is somewhat more restricted than in case OM24 andthe injection was in a more anterior and medial part of the hypothala-mus. The labeled cells in the temporal pole are probably due toinvolvement of the anterior commissure. Conventions as in Figure 1.
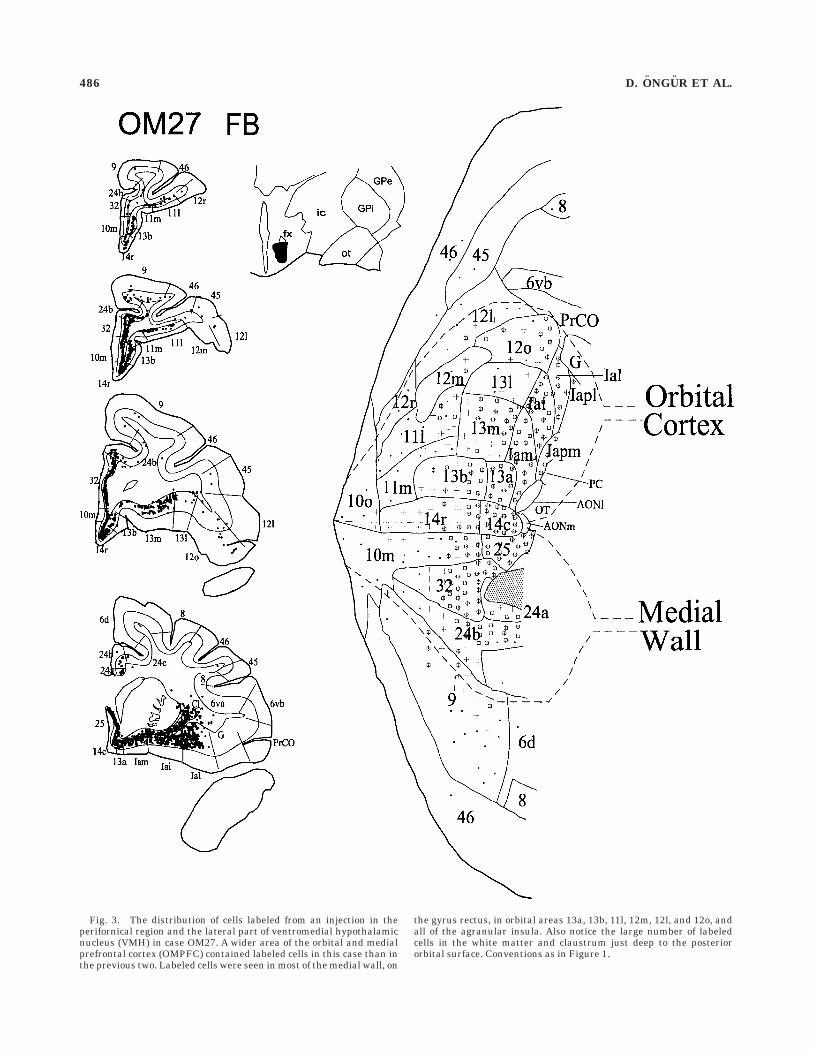
Fig. 3. The distribution of cells labeled from an injection in theperifornical region and the lateral part of ventromedial hypothalamicnucleus (VMH) in case OM27. A wider area of the orbital and medialprefrontal cortex (OMPFC) contained labeled cells in this case than inthe previous two. Labeled cells were seen in most of the medial wall, on
the gyrus rectus, in orbital areas 13a, 13b, 11l, 12m, 12l, and 12o, andall of the agranular insula. Also notice the large number of labeledcells in the white matter and claustrum just deep to the posteriororbital surface. Conventions as in Figure 1.
486 D. ONGUR ET AL.

Fig. 4. Prefrontal neurons labeled from the lateral hypothalamicarea in case OM38. The injection site is located at a more posteriorlevel than in the preceding cases. There are large numbers of labeled
cells in the medial areas 24b, 25, 32, and 14c and in orbital areas 13a,13l, Iam, Iai, Ial, 12o, and 12l, as well as in the rostral claustrum.Conventions as in Figure 1.
PREFRONTAL PROJECTIONS TO THE HYPOTHALAMUS 487

substantial non-OMPFC cortical input to the rostral ven-tral midbrain than to the hypothalamus.
Projections from the amygdala. As described in thepreceding paper, projections from the amygdala to the PAG
showed an interesting pattern in light of the corticalconnections of both structures. A similar observation isdescribed here about the amygdaloid projections to thehypothalamus.
Fig. 5. Prefrontal neurons labeled from an injection in the ventralmidbrain in case OM26. The injection site was close to the dopaminer-gic cells that project to the forebrain in the substantia nigra pars
compacta and the ventral tegmental area. There is heavy retrogradelabel in medial wall areas 32, 25, and 24b as well as in orbital areas14c, 13a, Iam, Iai, Ial, and 12o. Conventions as in Figure 1.
488 D. ONGUR ET AL.
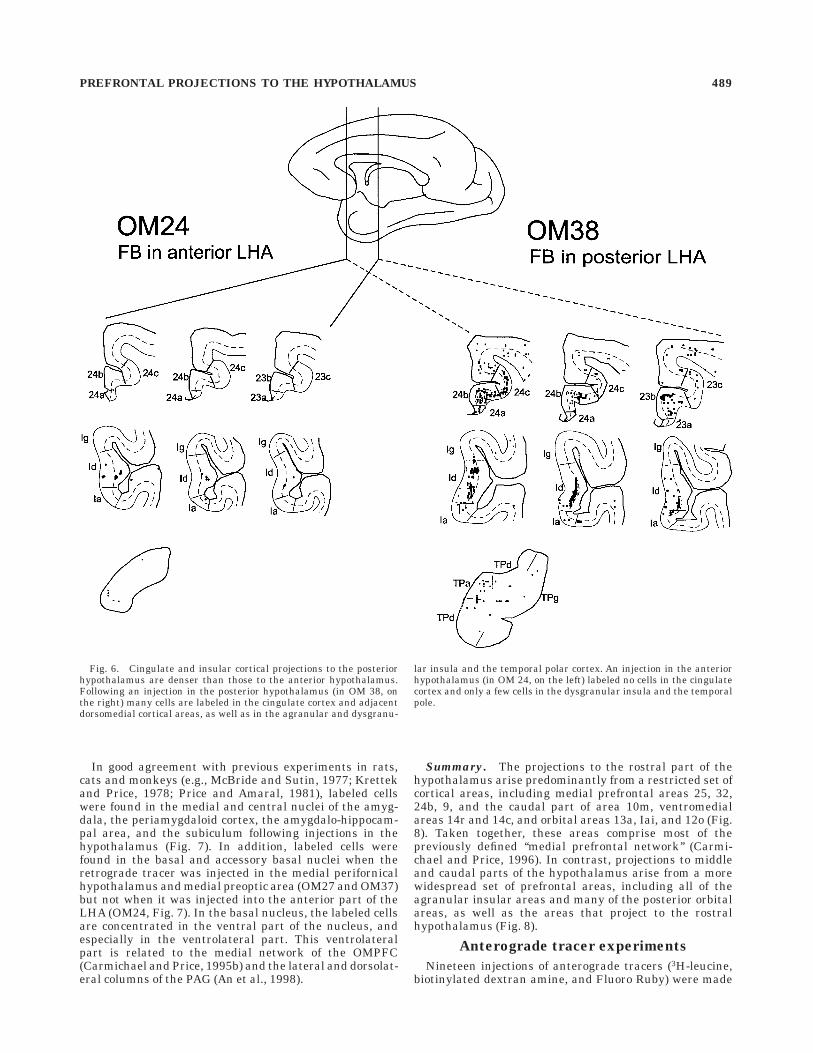
In good agreement with previous experiments in rats,cats and monkeys (e.g., McBride and Sutin, 1977; Krettekand Price, 1978; Price and Amaral, 1981), labeled cellswere found in the medial and central nuclei of the amyg-dala, the periamygdaloid cortex, the amygdalo-hippocam-pal area, and the subiculum following injections in thehypothalamus (Fig. 7). In addition, labeled cells werefound in the basal and accessory basal nuclei when theretrograde tracer was injected in the medial perifornicalhypothalamus and medial preoptic area (OM27 and OM37)but not when it was injected into the anterior part of theLHA (OM24, Fig. 7). In the basal nucleus, the labeled cellsare concentrated in the ventral part of the nucleus, andespecially in the ventrolateral part. This ventrolateralpart is related to the medial network of the OMPFC(Carmichael and Price, 1995b) and the lateral and dorsolat-eral columns of the PAG (An et al., 1998).
Summary. The projections to the rostral part of thehypothalamus arise predominantly from a restricted set ofcortical areas, including medial prefrontal areas 25, 32,24b, 9, and the caudal part of area 10m, ventromedialareas 14r and 14c, and orbital areas 13a, Iai, and 12o (Fig.8). Taken together, these areas comprise most of thepreviously defined ‘‘medial prefrontal network’’ (Carmi-chael and Price, 1996). In contrast, projections to middleand caudal parts of the hypothalamus arise from a morewidespread set of prefrontal areas, including all of theagranular insular areas and many of the posterior orbitalareas, as well as the areas that project to the rostralhypothalamus (Fig. 8).
Anterograde tracer experiments
Nineteen injections of anterograde tracers (3H-leucine,biotinylated dextran amine, and Fluoro Ruby) were made
Fig. 6. Cingulate and insular cortical projections to the posteriorhypothalamus are denser than those to the anterior hypothalamus.Following an injection in the posterior hypothalamus (in OM 38, onthe right) many cells are labeled in the cingulate cortex and adjacentdorsomedial cortical areas, as well as in the agranular and dysgranu-
lar insula and the temporal polar cortex. An injection in the anteriorhypothalamus (in OM 24, on the left) labeled no cells in the cingulatecortex and only a few cells in the dysgranular insula and the temporalpole.
PREFRONTAL PROJECTIONS TO THE HYPOTHALAMUS 489

into restricted areas of the OMPFC in 15 monkeys. Ofthese, twelve injections produced labeled axons in thehypothalamus. Confirming the retrograde tracing studies,these injections are all in a band of cortex, which extendsfrom the medial prefrontal cortex through the caudalorbital and agranular insular areas. Injections in therostral areas of the orbital or medial prefrontal cortex donot label axons in the hypothalamus.
Nine anterograde tracer experiments produced labelthroughout the anteroposterior extent of the hypothala-mus. These injections involve cortical areas within the‘‘medial prefrontal network’’ that are labeled after retro-grade tracer injections in both rostral and caudal hypo-
thalamus (i.e. areas 32, 25, 13a, Iai, 12o, and the caudalpart of area 12l). The other experiments have axonal labelonly in the posterior hypothalamus. There, injections weremade in areas of the caudal orbital cortex (areas Iam, 12m,and 13m) that only contained substantial numbers oflabeled neurons following retrograde tracer injections inthe caudal hypothalamus. In all cases, the label waspredominantly ipsilateral.
In cases with the anterograde tracers BDA and fluo-roruby, the labeled axons in the hypothalamus had numer-ous varicosities along their length. These varicositiesappear to represent synaptic boutons, and indicate thataxons make functional contacts with hypothalamic neu-
Fig. 7. The distribution of labeled cells in the amygdala followinginjections of the retrograde tracer fast blue (FB) in the hypothalamus.Note that all injections in the hypothalamus gave rise to label in thecentral and medial nuclei of the amygdala, but only those with
injections in the medial preoptic area and near the ventromedialhypothalamic nucleus (VMH) labeled cells in the basal nucleus of theamygdala. Labeled cells were also seen in the subiculum followinginjections in the hypothalamus.
490 D. ONGUR ET AL.

Fig. 8. Graphs of the relative density of retrogradely labeled cellsin prefrontal cortical areas after anterior and posterior hypothalamicinjections. The two cases with anterior injections and the three caseswith posterior injections were scaled to the maximum value withineach brain and then averaged to create single ‘‘anterior’’ and ‘‘poste-rior’’ values for each cortical area. The densities were again scaled to
the maximum value for each averaged data set to illustrate thepattern of label. The top panel shows that the anterior hypothalamusreceives input mainly from the medial prefrontal network areas,whereas the posterior hypothalamus receives inputs from the medialnetwork as well as additional posterior orbital areas.

rons. Therefore, the axonal varicosities, rather than thefibers themselves, are mapped in most cases.
Injections into areas 32, 25, 13a, Iai, 12, and 12l
OM35. BDA was injected into area 32 on the medialwall anterior to the corpus callosum. Intense axonal label
is seen in most parts of the hypothalamus throughout itsanteroposterior extent (Fig. 9), in agreement with thestrong cell labeling in this area following retrograde tracerinjections in all parts of the hypothalamus describedabove. Rostrally, there are many labeled fibers in both the
Fig. 9. The distribution of anterogradely labeled axons in OM35,following an injection of the anterograde tracer biotinylated dextranamine (BDA) in area 32. Camera lucida drawings of labeled axons insix sections through the hypothalamus are shown in c. The injectionsite is shown in a and the area of detail in b. There are labeled axons inthe anterior to posterior extent of both medial and lateral hypothala-
mus, with a particularly strong plexus of fibers in the anteriorhypothalamic area (AHA). The labeled axons avoid for the most partthe paraventricular, arcuate, suprachiasmatic, supraoptic, and mam-millary nuclei, although they may run immediately adjacent to thesenuclei. There is also label in many adjacent structures such as thenucleus basalis of Meynert and zona incerta.
492 D. ONGUR ET AL.

lateral and medial preoptic areas, and a particularly denseplexus of fibers in the anterior hypothalamic area. Furthercaudally, labeled fibers are found in the VMH and thecell-sparse shell around this nucleus, the dorsomedialhypothalamic nucleus, and the LHA. Caudally, there aremany labeled axons in the posterior hypothalamic regionsurrounding the third ventricle, dorsal to the premammil-lary and mammillary nuclei.
In this brain and in all the cases described below, thelabel avoids the supraoptic nucleus, the suprachiasmaticnucleus, the arcuate hypothalamic nucleus, the paraven-tricular nucleus and adjacent periventricular zone, and
the mammillary nuclei, although axons are observed abut-ting the borders of these nuclei.
The axons that innervate the hypothalamus coursethrough the basal forebrain anterior to the diencephalonand presumably innervate those areas as well (see Russ-chen et al., 1985). At caudal levels of the hypothalamus,the labeled axons also invade the adjoining zona incerta,the medial tip of the subthalamic nucleus, and the fields ofForel. Axonal label also continues caudally into the ventralmidbrain and the PAG (see below and An et al., 1998).
OM32. This animal had a large injection of BDA inarea 25, ventral to the genu of the corpus callosum. The
Fig. 10. Plots of labeled axonal varicosities in the hypothalamusfollowing two injections of the anterograde tracer biotinylated dextranamine (BDA) in areas 25 and 24b in the medial prefrontal cortex(PFC). a: With the injection in area 25, there are many labeled axonsin most parts of the hypothalamus with an especially dense concentra-
tion of varicosities in the ventromedial hypothalamic nucleus (VMH).b: With the injection in the anterior cingulate area 24b, there arefewer labeled varicosities, but with a relative concentration in thedorsal hypothalamus.
PREFRONTAL PROJECTIONS TO THE HYPOTHALAMUS 493
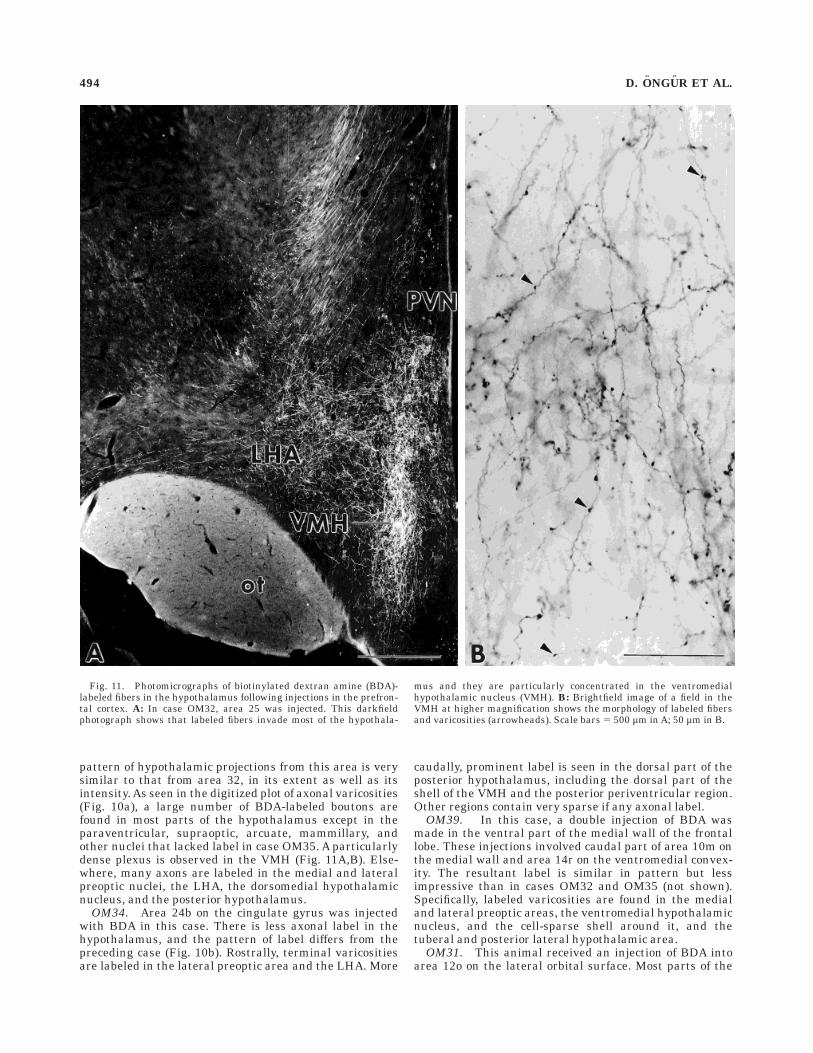
pattern of hypothalamic projections from this area is verysimilar to that from area 32, in its extent as well as itsintensity. As seen in the digitized plot of axonal varicosities(Fig. 10a), a large number of BDA-labeled boutons arefound in most parts of the hypothalamus except in theparaventricular, supraoptic, arcuate, mammillary, andother nuclei that lacked label in case OM35. A particularlydense plexus is observed in the VMH (Fig. 11A,B). Else-where, many axons are labeled in the medial and lateralpreoptic nuclei, the LHA, the dorsomedial hypothalamicnucleus, and the posterior hypothalamus.
OM34. Area 24b on the cingulate gyrus was injectedwith BDA in this case. There is less axonal label in thehypothalamus, and the pattern of label differs from thepreceding case (Fig. 10b). Rostrally, terminal varicositiesare labeled in the lateral preoptic area and the LHA. More
caudally, prominent label is seen in the dorsal part of theposterior hypothalamus, including the dorsal part of theshell of the VMH and the posterior periventricular region.Other regions contain very sparse if any axonal label.
OM39. In this case, a double injection of BDA wasmade in the ventral part of the medial wall of the frontallobe. These injections involved caudal part of area 10m onthe medial wall and area 14r on the ventromedial convex-ity. The resultant label is similar in pattern but lessimpressive than in cases OM32 and OM35 (not shown).Specifically, labeled varicosities are found in the medialand lateral preoptic areas, the ventromedial hypothalamicnucleus, and the cell-sparse shell around it, and thetuberal and posterior lateral hypothalamic area.
OM31. This animal received an injection of BDA intoarea 12o on the lateral orbital surface. Most parts of the
Fig. 11. Photomicrographs of biotinylated dextran amine (BDA)-labeled fibers in the hypothalamus following injections in the prefron-tal cortex. A: In case OM32, area 25 was injected. This darkfieldphotograph shows that labeled fibers invade most of the hypothala-
mus and they are particularly concentrated in the ventromedialhypothalamic nucleus (VMH). B: Brightfield image of a field in theVMH at higher magnification shows the morphology of labeled fibersand varicosities (arrowheads). Scale bars 5 500 µm in A; 50 µm in B.
494 D. ONGUR ET AL.

lateral hypothalamus show strong axonal label (Figs. 12,13A,B). The LPOA, the anterior and posterior portions ofLHA and the posterior hypothalamus are all denselylabeled. More medial areas such as the MPOA, VMH, orPVN are devoid of label. This is consistent with theretrograde experiments, in which area 12o is most stronglylabeled after an injection in the LHA. The striking organi-zation of label in OM31 is similar to that in other caseswith injections in orbital areas connected with the medialprefrontal network (e.g., areas Iai, 13a, and the caudalpart of area 12l).
OM37. This animal received an injection of the fluores-cent tracer fluororuby in the caudal part of area 12l on theventrolateral convexity. Axonal varicosities are seen in theLPOA, anterior and particularly the posterior LHA, andthe posterior hypothalamus (Fig. 14a). There is no label inthe VMH or any of the other medial hypothalamic areas ornuclei.
In the same case, BDA was injected into the agranularinsular area Iai. The pattern of label is similar to that fromthe fluororuby injection but somewhat stronger. Axons andaxonal varicosities are labeled in the lateral preoptic area,the LHA anteriorly and posteriorly, the dorsomedial hypo-thalamic nucleus, and the posterior hypothalamus (Fig 14b).
OM19. An injection of the anterograde tracer 3H-leuwas made into area 13a in the medial orbital cortex. Thelabeled fibers innervate the lateral preoptic area and thefull anteroposterior extent of the lateral hypothalamus(Fig. 14c). There is label in the dorsal and posteriorhypothalamus as in OM37, but in this case, the label alsoextends medially towards the medial preoptic area, theanterior hypothalamic area, the VMH, and the premammil-lary nuclei.
OM28. This animal received an injection of BDA intothe orbital area 13l. This injection was large, however, andan appreciable part of it extended into the agranular
Fig. 12. Camera lucida drawings of fibers labeled from an injectionof biotinylated dextran amine (BDA) in area 12o on the lateral orbitalsurface . There are many fewer labeled fibers in the hypothalamus inthis case than in cases OM35 and OM32 with injections in areas 25
and 32. The fibers are concentrated in the lateral hypothalamic area(LHA) while largely avoiding the medial hypothalamus. There is alsostrong label in the nucleus basalis of Meynert and the zona incerta.
PREFRONTAL PROJECTIONS TO THE HYPOTHALAMUS 495
insular area Iai. There is light, scattered label in thepreoptic areas, the LHA, the dorsomedial hypothalamicnucleus, and the posterior hypothalamus (not shown). Thispattern is hard to attribute to a single architectonic areabut it is largely similar to that in the preceding three cases.
Injections into other orbital areas
OM30. BDA was injected into area 12m on the lateralorbital surface in this case. Strikingly, most hypothalamicnuclei and areas are devoid of label. The only exception isthe posterior LHA just lateral and dorsal to the mammil-lary nuclei (Fig. 15a). In this region, the number of labeledaxons observed is comparable to that seen in most other cases.
OM8. In this case, 3H-leu was injected into areasIam/Iapm in the agranular insula. Only light label is ob-served in the hypothalamus, which is restricted to the LHA,particularly the posterior portion of this area (Fig. 15b).
OM34. The fluorescent tracer FR was injected intoarea 13m in the central orbital surface in this case. Therewere small numbers of labeled axons in the hypothalamus(Fig. 15c). The only concentration of labeled fibers is in theposterior LHA, at the level of the mammillary bodies.(Monkey OM34 received an injection of BDA into area 24b;see above.)
OM16, OM27, OM29, OM30, OM36, OM38, OM39.Injections of 3H-leu, FR, and BDA into several other areasof the OMPFC produced no label in the hypothalamus inthese cases, regardless of the size of the injection. Theinjected areas include rostral area 12l, area 11l, area 11m,area 12r, rostral part of area 10m, area 10o, and area 9. Allof these injections are in the rostral part of the prefrontalcortex, five on the orbital surface, one on the medial wall,and one on the dorsomedial surface.
Ventral midbrain
The same injections that produced label in the hypothala-mus also label the substantia nigra pars compacta (SNpc)and nearby structures such as the interpeduncular nucleus(IP; Fig. 16).
Overall, the number of labeled axons in the mesencepha-lon is limited, but the relative intensity of label in theventral midbrain parallels the intensity of label in thehypothalamus in most cases. For example, injections ofanterograde axonal tracers in areas 25, 32, 12o, and Iairesult in numerous varicosities throughout the structure.In contrast, injections in areas 12m and Iam give only afew labeled varicosities and the injections that produce no
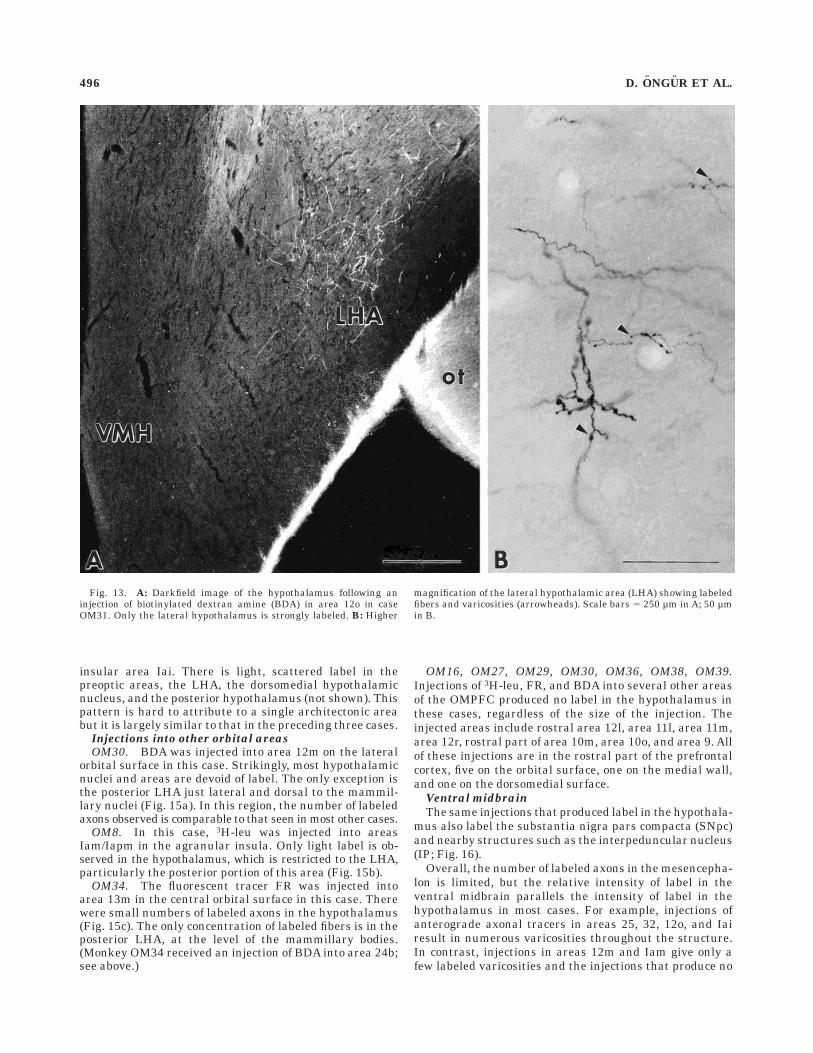
Fig. 13. A: Darkfield image of the hypothalamus following aninjection of biotinylated dextran amine (BDA) in area 12o in caseOM31. Only the lateral hypothalamus is strongly labeled. B: Higher
magnification of the lateral hypothalamic area (LHA) showing labeledfibers and varicosities (arrowheads). Scale bars 5 250 µm in A; 50 µmin B.
496 D. ONGUR ET AL.

Fig. 14. Plots of axonal varicosities or fiber label in the hypothala-mus in three cases with injections into orbital surface areas related tothe medial network. a: Case OM37 had an injection of fluororuby (FR)in the caudal part of the lateral orbital area 12l. A large number ofvaricosities can be seen in the lateral hypothalamus. There is alsolabel in the periventricular posterior hypothalamus, but the medialpreoptic area (MPOA) and ventromedial hypothalamic nucleus (VMH)are largely free of label. b: In case OM37, biotinylated dextran amine(BDA) was injected in the agranular insular area Iai . The labeledaxonal varicosities were mostly located in the lateral preoptic area and
the lateral hypothalamic area (LHA). c: In case OM19, an injection oftritiated leucine (3H-leu) was made in the medial orbital area 13a, andthe labeled fibers demonstrated by autoradiography. The distributionof autoradiographic silver grains above background in the hypothala-mus are shown. This is of necessity a more subjective depiction of datathan the varicosity plots shown in the first two parts of this figure.Labeled axons were concentrated in the lateral and posterior hypothala-mus, with a few in the dorsal hypothalamus (DMH), anterior hypotha-lamic area (AHA), and the lateral part of the VMH.
PREFRONTAL PROJECTIONS TO THE HYPOTHALAMUS 497

label in the hypothalamus also do not label the SNpc or theventral tegmental area (VTA). To reach the SNpc or VTA,labeled fibers commonly travel in the posterior hypothala-mus and PAG and take a ventrolateral turn to invade the
ventral midbrain. They then sweep dorsolaterally to makenumerous synapses along the dorsal edge of the SNpc. Thispattern is similar to that seen with amygdaloid afferentsentering the ventral midbrain (Price andAmaral, 1981).
Fig. 15. The distribution of axons and varicosities following injec-tions in areas associated with the orbital prefrontal network. a: In caseOM30 with a biotinylated dextran amine (BDA) injection in area 12mon the lateral orbital surface, there was no label in the hypothalamusexcept for in the posterior lateral hypothalamic area (LHA). b: In caseOM8 with a tritiated leucine (3H-leu) injection in agranular insularareas Iam and Iapm, no fibers were labeled in the anterior hypothala-
mus and only moderate label was found in the posterior LHA,although there was good label in areas adjacent to the hypothalamus.c: Case OM 34 had an injection of the fluorescent tracer fluororuby(FR) in area 13m on the central orbital surface. In the hypothalamus,there are only a few labeled fibers in the medial preoptic area (MPOA)and perifornical LHA, and in the posterior LHA.
498 D. ONGUR ET AL.

Summary
The most substantial projections to all of the hypothala-mus originate from areas on the posterior medial wall.Related orbital areas send their axons mainly to thelateral hypothalamus. Finally, posterior orbital and agranu-lar insular areas send a relatively weak projection to theposterior part of the hypothalamus.
In order to quantify these relationships, the axonalvaricosities were counted from digitized maps in themedial, dorsal, or lateral parts of the hypothalamus (Fig.17). Cases OM8 and OM19, with injections in areas Iamand 13a, could not be included in this analysis because thetritiated amino acid tracer used in those cases did notallow varicosities to be visualized. The pattern of innerva-tion in these cases matched qualitatively the patterns oflabeled varicosities seen after injections in related areas.The strongest projections originate from areas 25 and 32and innervate all three parts of the hypothalamus. Areas14r/10m show a similar yet weaker input. Area 24bprojects most strongly to the dorsal hypothalamus. Areas12o, Iai, and caudal 12l all innervate prominently thelateral zone of the hypothalamus. Their input to themedial zone is meager. Finally, orbital areas 12m and 13msend a few axons to the lateral hypothalamus. All of thesepoints are summarized in Figure 18.
DISCUSSION
The cortical projections to the hypothalamus arise pre-dominantly from the OMPFC; within this region, themajor projections originate in areas of the previouslydefined ‘‘medial prefrontal network’’ (see Carmichael andPrice, 1966). Within this network, areas 25 and 32 on the
medial wall send the heaviest projection to all parts of thehypothalamus, with especially strong input to the anteriorhypothalamic area and the VMH. The ventromedial corti-cal areas 10m and 14r anterior to areas 25 and 32 show asimilar pattern of projections, only weaker. Orbital areasthat are part of the medial prefrontal network (areas Iai,13a, and 12o) and the caudal part of area 12l projectstrongly and selectively to the lateral hypothalamus.These areas project most strongly to the posterior LHA,and at the level of the mammillary nuclei also send fibersto the periventricular posterior hypothalamic region. Otherareas on the posterior orbital surface that are part of thepreviously defined ‘‘orbital network’’ (e.g., areas Iam, Iapm,Ial, 13m, 13l ,and 12m) send a much lighter projection,which is largely restricted to the posterior lateral hypotha-lamic area at the level of the mammillary bodies. The morerostral areas in the OMPFC (such as areas 10o, 11m, 11l,12r, or the rostral part of area 12l) do not send substantialprojections to the hypothalamus.
Taken together with the results of the preceding paperon the projections of the OMPFC to the PAG (An et al.,1998), these results indicate that areas of the medialprefrontal network project strongly to brain regions in-volved in autonomic and homeostatic regulation. Further-more, projections to subdivisions of both the hypothalamusand the PAG arise in distinct regions within the medialnetwork. The subdivisions within the hypothalamus andPAG also find support in the selective nature of amygda-loid projections to these structures. This organizationpoints to parallel but segregated output channels thatrelay forebrain activity to brainstem centers (Fig. 19).
The projection from the caudal part of area 12l to thehypothalamus corroborates the evidence from the compan-
Fig. 16. The distribution of labeled axons in the ventral midbrainin several cases with injections of anterograde tracers in the orbitaland medial prefrontal cortex (OMPFC). Those injections that pro-duced robust label in the hypothalamus (e.g., cases OM32, OM35,
OM31, and OM37) also produced substantial label in and around thesubstantia nigra pars compacta, whereas the injections that producedlittle label in the hypothalamus (e.g., cases OM8 ad OM30) resulted inlittle label in the midbrain.
PREFRONTAL PROJECTIONS TO THE HYPOTHALAMUS 499

ion paper that area 12l may be heterogeneous and itscaudal part may be related to the medial network. Ourresults also show that the posterior part of area 10mprojects to the hypothalamus, whereas its anterior partdoes not, indicating that this large area may also beheterogeneous. In this case, there may be differencesbetween the hypothalamus and the PAG, sbecause aninjection in the rostral part of 10m gave rise to label in thePAG, but not in the hypothalamus.
A similar situation exists with area 9 on the dorsomedialconvexity. This area is also substantially interconnectedwith medial prefrontal network areas and projects to thePAG. However, an anterograde tracer injection in this areafailed to label the hypothalamus. These observationsunderline an important characteristic of prefrontal net-works: they are groupings of interconnected cortical areas,but membership in a network does not dictate each point ofconnectivity for an area.
Previous studies
Anatomical studies in rats have consistently shown thatboth infralimbic and prelimbic areas in the medial wall ofthe prefrontal cortex and agranular insular areas in therhinal sulcus project strongly to the LHA (Beckstead,
1979; Sesack et al., 1989; Hurley et al., 1991; Price et al.,1991). Thus, the corticohypothalamic projection in ratsarises from restricted cortical fields as it does in monkeys.Several rat studies have suggested that the PFC neuronswhich project to the LHA cluster in the medial wall andlateral orbital areas, skipping the intervening medialorbital cortex (e.g., Beckstead, 1979; Price et al., 1991;Hurley et al., 1991). The networks of the OMPFC have notbeen studied in rats, but the mentioned data bear parallelsto our present findings.
The PFC projects mainly to the lateral hypothalamus inrats. Although we have found OMPFC inputs to the LHA,we also demonstrated that the medial wall areas projectstrongly to the medial parts of the hypothalamus. This isthe primary difference between our results and the ratstudies and may represent a prominent difference betweenprimates and rodents. Because the VMH and anteriorhypothalamic area are implicated in different functionsthan the LHA, the OMPFC may influence a wider varietyof processes in primates than in rats.
Previous reports of projections from the prefrontal cor-tex to the hypothalamus in primates are sketchy. Johnsonet al. (1968) used the Nauta axonal degeneration method
Fig. 17. Analysis of the distribution of axonal varicosities inmedial, lateral, and dorsal zones of the hypothalamus in several caseswith anterograde tracer injections in the orbital and medial prefrontalcortex (OMPFC). The density of the varicosities was measured in eachzone across four sections through the anteroposterior extent of thehypothalamus. Note that the absolute values depend on the size andeffectiveness of the tracer injection in each case as well as size of the
projections. Medial wall areas 25, 32, and 14r/10m send strongprojections to most of the hypothalamus and especially to the medialzone. Area 24b is similar but prefers the dorsal zone. The medialnetwork areas on the orbital surface (12o, 13a, Iai, and caudal 12l)project primarily to the lateral hypothalamus. The much weakerprojections labeled by injections in the orbital network areas 12m and13m are mainly distributed to the lateral hypothalamus.
500 D. ONGUR ET AL.
to show that degenerating fibers are seen in the septumand lateral hypothalamus only after lesions of the medialand lateral orbital areas (they did not study lesions of the
medial wall). This report is consistent with the presentfindings that areas on the orbital surface (13a, 12o, Iai,12m, 13l, Iam) project to the hypothalamus.
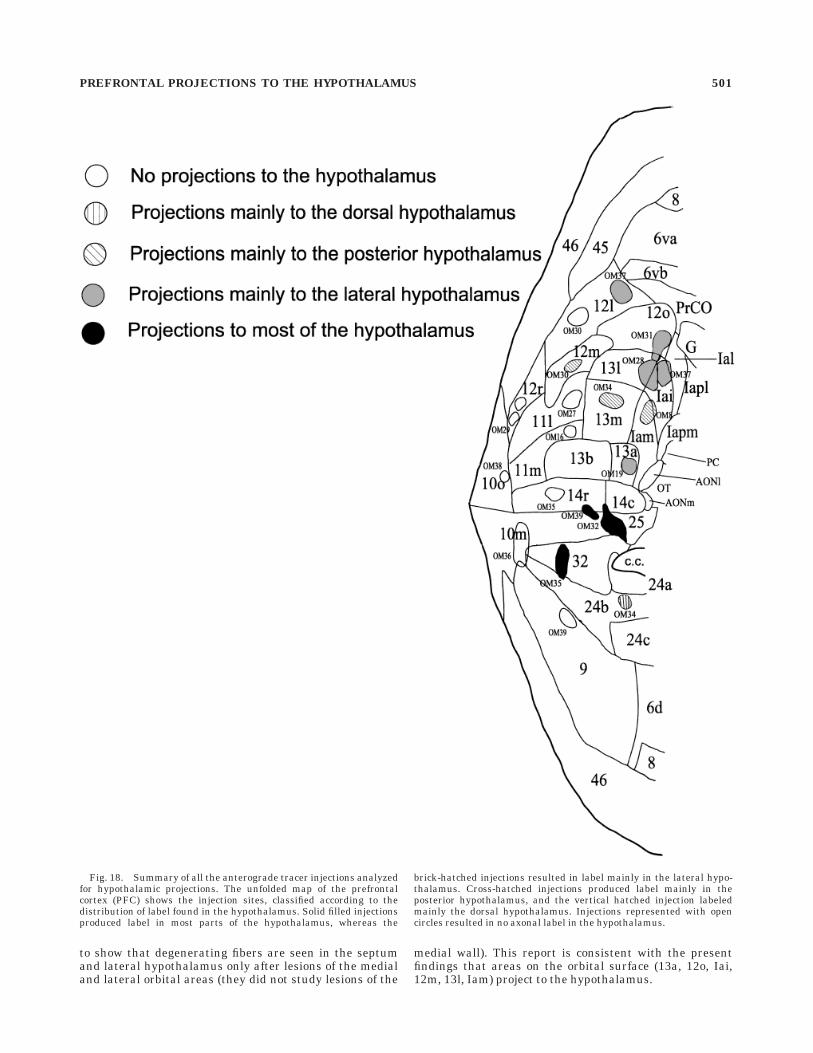
Fig. 18. Summary of all the anterograde tracer injections analyzedfor hypothalamic projections. The unfolded map of the prefrontalcortex (PFC) shows the injection sites, classified according to thedistribution of label found in the hypothalamus. Solid filled injectionsproduced label in most parts of the hypothalamus, whereas the
brick-hatched injections resulted in label mainly in the lateral hypo-thalamus. Cross-hatched injections produced label mainly in theposterior hypothalamus, and the vertical hatched injection labeledmainly the dorsal hypothalamus. Injections represented with opencircles resulted in no axonal label in the hypothalamus.
PREFRONTAL PROJECTIONS TO THE HYPOTHALAMUS 501

Fig. 19. The connections of the orbital and medial prefrontal cortex(OMPFC) are summarized with respect to prefrontal networks. Asimplified scheme of the connectivity of amygdala, hypothalamus, andperiaqueductal gray (PAG) is also shown. The orbital network areas ingreen in the top panel receive most of the sensory informationreaching the OMFPC, process it and relay it to the medial networkareas. The medial network has three components (yellow, red, andblue) identified by their preferred outputs to distinct parts of thehypothalamus and PAG shown in the same colors (dorsal hypothala-
mus and lateral PAG yellow; medial hypothalamus and dorsolateralPAG red; lateral hypothalamus and ventrolateral PAG blue). All ofthese cortical areas project to the basal nucleus of the amygdala whichis connected mainly to the medial hypothalamus and dorsolateralcolumn of PAG (dlPAG; shown in red). The lateral hypothalamus andthe ventrolateral column of PAG (vlPAG) receive inputs from thecentral nucleus of the amygdala (all shown in blue). Note that lateralorbital area 12l may be heterogeneous, with different portions relatedto the orbital and medial networks.

Saper (1990) cites reports from human material thataxonal degeneration can be demonstrated with the Gleesmethod in the mammillary bodies and the ventromedialnucleus of postmortem brains from frontal leukotomypatients. However, both he and Nauta (1969) suggest thatthese findings are probably artifacts of the technique.
OMPFC and the hypothalamus
The hypothalamus is a complex region with widespreadintrinsic and extrinsic connections. Most of its majornuclei and areas (for example, the ventromedial hypotha-lamic nucleus and the lateral hypothalamic area) arereciprocally linked and this web-like organization seems tounderlie much of its function as well. Indeed, it is difficultto provide a simple and exhaustive account of hypotha-lamic control of any given function (Swanson, 1987; Saper,1995). Instead, we will correlate cortico-hypothalamic connec-tions with distinct but not separate functional zones.
The posterior lateral hypothalamic area lateral anddorsal to the mammillary nuclei and ventral to the subtha-lamic nucleus has emerged as a prominent zone of cortico-hypothalamic innervation. For some cortical areas, theposterior LHA was the only hypothalamic area to receivecortical input. This area has been noted to possess specialconnections in several studies in the rat. It has widespreadreciprocal connections with brainstem centers controllingthe autonomic nervous system, particularly the parabra-chial nucleus (Saper, 1995; Jansen et al., 1995). It alsoprojects to the dorsal vagal nucleus and the nucleusambiguus and thus can influence parasympathetic outflow(Loewy, 1991). The posterior LHA may thus be a focal pointin the hypothalamus controlling autonomic reactions un-der influence from the medial prefrontal cortex. Saper(1985) showed that the posterior LHA is also the majorsource of hypothalamocortical projections in the rat andthat these projections are topographically organized, imply-ing that it integrates autonomic changes with arousal andbehavioral states.
Studies with small injections of excitatory amino acidsshow that activation of the LHA causes a decrease in bloodpressure, heart rate, and cardiac output (Loewy, 1991).This effect parallels that seen with activation of theventrolateral column of the PAG (vlPAG). As we haveshown, both the LHA and the vlPAG receive selectiveinputs from the same cortical areas, namely 13a, 12o, Iai,and 12l. Both the LHA and vlPAG also receive inputs fromthe central nucleus of the amygdala but not from the basalnucleus of the amygdala. The basal nucleus projects tonearby areas in both structures while avoiding the LHAand vlPAG. We have also found that projections from theLHA to the PAG are concentrated in the vlPAG, wereasthose from the VMH are concentrated in the dlPAG(unpublished observations).
A number of physiological studies have shown that theOMPFC can influence a wide range of motivated behaviorsas well as autonomic function through the hypothalamus.In rats, stimulation of the frontal cortex produces monosyn-aptic as well as longer-latency responses in the hypothala-mus (Kita and Oomura, 1981). In monkeys, stimulation ofthe orbital cortex can also cause long-latency changes inthe firing patterns of most neurons in the LHA, especiallythose that are responsive to glucose (Karadi et al., 1990).This fits well with the long association of the LHA withhunger and thirst, as well as with aggression and reproduc-tion (see Swanson, 1987 for a review). In relation to
aggression, it is notable that stimulation of both the lateraland medial aspects of the PFC in rats (De Bruin, 1990) andin cats (Siegel et al., 1974, 1975) attenuate attack behaviorinduced by lateral hypothalamic stimulation. Further-more, autonomic changes that may be produced by corticalstimulation, such as an increase in sympathetic nervedischarge or hypotension, can be blocked by local injec-tions of cobalt or lidocaine in the LHA (Hardy and Holmes,1988; Cechetto and Chen, 1990).
The present finding that the posterior medial PFCprojects strongly to the medial hypothalamus in monkeysand especially to the VMH does not appear to have beenpreviously reported in rats or other animals. Cytoarchitec-tonically, the VMH is a heterogeneous nucleus and isdivided into dorsomedial, central, and ventrolateral por-tions. Cells in the ventrolateral part of the VMH concen-trate sex steroids and have been implicated in reproduc-tive behavior. The dorsomedial part of the nucleus isconnected with paraventricular and dorsomedial hypotha-lamic nuclei and seems to regulate appetitive behaviors(Simerly, 1995). In our study, both the dorsal and ventralparts of the VMH received heavy input from the medialcortical areas.
Classical studies indicated that the VMH contained a‘‘satiety center’’ in reciprocal relation to the ‘‘feedingcenter’’ in the LHA. Although this view is clearly inad-equate (Swanson, 1987), there is ample evidence to impli-cate the VMH in some sort of regulation of feedingbehavior (Hamilton et al., 1976). Indeed, recent work hasindicated that the VMH is activated following intravenousadministration of leptin, a hormone that is crucial inregulation of food intake and body weight (Elmquist et al.,1997). The VMH also couples energy intake to activity inthe hypothalamus-pituitary-adrenal axis through influenc-ing the release of adrenocorticotropic hormone (ACTH;Choi et al., 1996).
After the injection of BDA in area 32, the anteriorhypothalamic area (AHA) also contained a plexus of la-beled fibers comparable in strength to those of the VMH.The AHA has been implicated in several homeostaticfunctions, including the control of body temperature andelectrolyte balance, and there is also evidence that it isinvolved in the regulation of sexual behavior, especially infemales (Conrad and Pfaff, 1976). Taken together with thecortical input to the LHA, the finding of an importantprojection from the OMPFC to the anterior and medialhypothalamus indicates that the prefrontal cortex caninfluence a wide variety of behaviors essential to themaintenance of the individual and species.
OMPFC and the autonomic nervous system
Early investigators amassed a wealth of data about theability of prefrontal stimulation to modulate autonomicparameters (Kaada et al., 1949; Wall and Davis, 1951;Delgado, 1960). The suggestion that the functions of thePFC relate systematically to those of the autonomic ner-vous system was made prominently by Nauta (1971). Hedrew on animal data and human studies showing absentor reduced galvanic skin responses following frontal le-sions (Luria and Homskaya, 1964) and he proposed thatthe PFC is required for the visceral, ‘‘gut feelings’’ that weexperience in response to emotionally significant events.He postulated that an interoceptive agnosia may arisewith frontal lobe damage which would lead to loss ofaffective responsiveness and foresight. This concept hasbeen extended by Damasio (1994) to explain the ‘‘acquired
PREFRONTAL PROJECTIONS TO THE HYPOTHALAMUS 503

sociopathy’’ of patients with bilateral orbitomedial dam-age. His theory of somatic markers asserts that theOMPFC, driven by affective and motivational influences,modulates visceral states to evoke gut reactions whichlead our decision making to maximize benefit in compli-cated situations (Bechara et al., 1997). Thus, the controlover the autonomic nervous system that is exerted by theOMPFC may be of critical importance for its wider controlof behavior.
Recent anatomical work particularly in rats has delin-eated the relationship of the PFC to the autonomic nervoussystem. Neafsey has suggested (1990) that the somaticsensory and motor areas on the dorsal surface of thefrontal lobe are flanked ventrally by viscerosensory andvisceromotor areas in the agranular insula and infralimbicand prelimbic cortex, respectively. The viscerosensoryagranular insula receives input from visceral relatedthalamic areas. The putative visceral motor cortex of therat has direct connections with autonomic control regionssuch as the lateral hypothalamic area, periaqueductalgrey, nucleus of the solitary tract, the dorsal vagal nucleus,and even the intermediolateral column of the spinal cord.
The OMPFC has long been recognized as a higher-orderassociation cortex that is the target of converging pro-cessed sensory inputs. In primates, as in rodents, visceralsensory inputs reach specific areas within the agranularinsula (Carmichael and Price, 1995b). There are also othersensory inputs to the OMPFC from visual, somatosensory,olfactory, and gustatory modalities, which terminate pri-marily in areas on the posterior orbital surface (Rolls andBayliss, 1994; Carmichael et al., 1994; Carmichael andPrice, 1995b). Thus, as proposed by Neafsey (1990), theagranular insula and posterior orbital surface of themonkey do process visceral and other related sensoryinformation.
Our studies outline the visceromotor areas of the pri-mate prefrontal cortex, which constitute the second compo-nent of Neafsey’s (1990) model. Areas 25 and 32, theprimate counterparts of the rat infralimbic and prelimbicareas, provide the strongest input to both the hypothala-mus and PAG in our studies. The situation in the primate,however, is more complicated than a simple medial/lateralseparation as suggested in the rat because some orbitaland insular areas also project to autonomic related subcor-tical sites. These observations can be unified by the recentdescription of orbital and medial prefrontal cortical net-works because all visceromotor areas in our study belongto the medial network. The analogy with the rat should betempered, however, by the observation that visceral infor-mation is not the only modality reaching or leaving theOMPFC.
The primate prefrontal cortex diverges from that of therodent further in the specificity of its projections to dis-crete parts of the hypothalamus and the PAG. As discussedin the preceding article, projections from the medial wallareas and the orbital areas belonging to the medialnetwork terminate in different columns of the PAG. Asimilar pattern is found in the OMPFC projections to thehypothalamus. The visceromotor network can seeminglybe divided into regional subdivisions, perhaps based on thekinds of autonomic reactions that are elicited from each ofthose subdivisions.
The recognition of the two prefrontal networks as viscero-motor and viscerosensory systems raises the issue ofinteractions between the networks. The two networkswere initially defined based on the extensive cortico-
cortical connections inside each network and on the rela-tive paucity of internetwork connections. In spite of this,there are significant interconnections that may transferinformation between the two networks. For example,areas 12o and 13a have extensive input and outputconnections with both networks, and have been included inboth networks. Area 11m belongs to the medial prefrontalnetwork and most of its connections are with areas on themedial wall, but it also receives inputs from the adjacentarea 11l, which is part of the orbital prefrontal networkand receives multimodal sensory information from theposterior and lateral orbital areas. Areas 14r and 14c,which are situated just at the border between the medialand orbital surfaces, also have connections to adjacentareas of both networks. Together, these provide multiplepathways for crossover of information between the sensory-related orbital network and the visceromotor medial pre-frontal network, at several stages of processing.
All of these observations may be summarized in a circuitwhich includes the OMPFC as its central node (Fig. 19).Sensory information from several modalities related tomotivational or emotional aspects of ongoing behavior(particularly feeding) is sent to the orbital network whereit is processed in a hierarchical fashion. The salientfeatures of this information are then transferred to themedial network, which elicits an appropriate visceralreaction by modulating activity in the hypothalamus,PAG, and related structures such as the amygdala (Fig.19). This reaction can be autonomic (changes in bloodpressure, galvanic skin resistance) or somatic (vocaliza-tion, aggression). It is also stimulus- and context-specificdue to the specificity of hypothalamic and PAG innervation(e.g., approach vs. avoidance, hypo- vs. hypertension).Feeding and homeostasis functions may well be the pri-mary functions of this system in rats, but it is likely thatthe system has become abstracted to more general aspectsof reward and motivation in primates.
LITERATURE CITED
Allen, G.V., C.B. Saper, K.M. Hurley, and D.F. Cechetto (1991) Organizationof visceral and limbic connections in the insular cortex of the rat. J.Comp. Neurol. 311:1–16.
An, X., R. Bandler, and J.L. Price (1998) Prefrontal cortical connections tolongitudinal columns in the midbrain periaqueductal gray in macaquemonkeys. J. Comp. Neurol. 401:455–479.
Bacon, S.J. and A.D. Smith (1993) A monosynaptic pathway from anidentified vasomotor centre in the medial prefrontal cortex to anautonomic area in the thoracic spinal cord. Neuroscience 54:719–728.
Bandler, R. and M.T. Shipley (1994) Columnar organization in the midbrainperiaqueductal gray: Modules for emotional expression? TINS 17:379–389.
Bechara, A., A.R. Damasio, H. Damasio, and S.W. Anderson (1994) Insensi-tivity to future consequences following damage to human prefrontalcortex. Cognition 50:7–15.
Bechara, A., H. Damasio, D. Tranel, and A.R. Damasio (1997) Decidingadvantageously before knowing the advantageous strategy. Science275:1293–1295.
Beckstead, R.M. (1979) An autoradiographic examination of corticocorticaland subcortical projections of the mediodorsal-projection (prefrontal)cortex in the rat. J. Comp. Neurol. 184:43–62.
Carmichael, S.T. and J.L.Price (1994) Architectonic subdivision of theorbital and medial prefrontal cortex in the macaque monkey. J. Comp.Neurol. 346:366–402.
Carmichael, S.T. and J.L. Price (1995a) Limbic connections of the orbitaland medial prefrontal cortex in macaque monkeys. J. Comp. Neurol.363:615–641.
Carmichael, S.T. and J.L. Price (1995b) Sensory and premotor connectionsof the orbital and medial prefrontal cortex. J. Comp. Neurol. 363:642–664.
504 D. ONGUR ET AL.

Carmichael, S.T. and J.L. Price (1996) Connectional networks within theorbital and medial prefrontal cortex of macaque monkeys. J. Comp.Neurol. 371:179–207.
Carmichael, S.T., M.-C. Clugnet, and J.L. Price (1994) Central olfactoryconnections in the macaque monkey. J. Comp. Neurol. 346:403–434.
Cechetto, D.F. and S.J. Chen (1990) Subcortical sites mediate sympatheticresponses from insular cortex. Am. J. Physiol. 258:R245–255.
Choi, S., C. Horsley, S. Aguila, and M.F. Dallman (1996) The hypothalamicventromedial nuclei couple activity in the hypothalamic-pituitary-adrenal axis to the morning or fed state. J. Neurosci. 16:8170–8180.
Conrad, L.C. and D.W. Pfaff (1976) Efferents from medial basal forebrainand hypothalamus in the rat. II. An autoradiographic study of theanterior hypothalamus. J. Comp. Neurol. 169:221–261.
Cowan, W.M., D.I. Gottlieb, A.E. Hendersen, J.L. Price, and T.A. Woolsey(1972) The autoradiographic demonstration of axonal connections inthe central nervous system. Brain Res. 37:21–55.
Damasio, A.R. (1994) Descartes’ Error. New York:Avon Science Publishing.Damasio, A.R., D. Tranel, and H. Damasio (1990) Individuals with socio-
pathic behavior caused by frontal damage fail to respond autonomicallyto social stimuli. Behav. Brain Res. 41:81–94.
DeBruin, J.P.C. (1990) Social behavior and the prefrontal cortex. Prog.Brain Res. 85:485–496.
Delgado, J.M.R. (1960) Circulatory effects of cortical stimulation. Physiol.Rev. 40:146–171.
Elmquist, J.K., R.S. Ahima, E. Maratos-Flier, J.S. Flier, and C.B. Saper(1997) Leptin activates neurons in the ventrobasal hypothalamus andbrainstem. Endocrinology 138:839–842.
Hamilton, C.L., P.J. Ciaccia, and D.O. Lewis (1976) Feeding behavior inmonkeys with and without lesions of the hypothalamus. Am. J. Physiol.230:818–830.
Hardy, S.G.P. and D.E. Holmes (1988) Prefrontal stimulus-produced hypo-tension in the rat. Exp. Brain Res. 73:249–255.
Hurley, K.M., H. Herbert, M.M. Moga, and C.B. Saper (1991) Efferentprojections of the infralimbic cortex of the rat. J. Comp. Neurol.308:249–276.
Jansen, A.S., X.V. Nguyen, V. Karpitskiy, T.C. Mettenleiter, and A.D. Loewy(1995) Central command neurons of the sympathetic nervous system:Basis of the fight-or-flight response. Science. 270:644–646.
Johnson, T.N., H.E. Rosvold, and M. Mishkin (1968) Projections frombehaviorally-defined sectors of the prefrontal cortex to the basalganglia, septum and diencephalon of the monkey. Exp. Neurol. 21:20–34.
Kaada, B.R., K.H. Pribram, and J.A. Epstein (1949) Respiratory andvascular responses in monkeys from temporal pole, insula, orbitalsurface and cingulate gyrus. J. Neurophysiol. 12:347–356.
Karadi, Z., Y. Oomura, H. Nishino, T.R. Scott, L. Leonard, and S. Aou (1990)Complex attributes of lateral hypothalamic neurons in the regulation offeeding of alert rhesus monkeys. Brain Res. Bull. 25:933–939.
Kita, H. and Y. Oomura (1981) Reciprocal connections between the lateralhypothalamus and the frontal cortex in the rat: electrophysiological andanatomical observations. Brain Res. 213:1–16.
Krettek, J.E. and J.L. Price (1978) Amygdaloid projections to subcorticalstructures within the basal forebrain and brainstem in the rat and cat.J. Comp. Neurol. 178:225–253.
Loewy, A.D. (1991) Forebrain nuclei involved in autonomic control. Prog.Brain Res. 87:253–268.
Luria, A.R. and E.D. Homskaya (1964) Disturbance in regulative role ofspeech with frontal lobe lesions. In J.M. Warren and K. Akert (eds): TheFrontal Granular Cortex and Behavior. New York: McGraw-Hill, p.352.
McBride, R.L. and J. Sutin (1977) Amygdaloid and pontine projections tothe ventromedial nucleus of the hypothalamus. J. Comp. Neurol.174:377–396.
Nauta, W.J.H. (1971) The problem of the frontal lobe: A reinterpretation. J.Psychiat. Res. 8:167–187.
Nauta, W.J.H. and W. Haymaker (1969) Hypothalamic nuclei and fiberconnections. In W. Haymaker, E. Anderson, and W.J.H. Nauta (eds):The Hypothalamus. Springfield, Illinois: Thomas, pp. 136–209.
Neafsey, E.J. (1990) Prefrontal cortical control of the autonomic nervoussystem: Anatomical and physiological observations. Prog. In Brain Res.85:147–166.
Price, J.L. and D.G.Amaral (1981) An autoradiographic study of theprojections of the central nucleus of the monkey amygdala. J. Neurosci.1:1242–1259.
Price, J.L., B.M. Slotnick, and M.-F. Revial (1991) Olfactory projections tothe hypothalamus. J. Comp. Neurol. 306:447–461.
Ray, J.P. and J.L.Price (1993) The organization of projections from themediodorsal nucleus of the thalamus to orbital and medial prefrontalcortex in macaque monkeys. J. Comp. Neurol. 337:1–31.
Rolls, E.T. and L.L. Bayliss (1994) Gustatory, olfactory and visual conver-gence within the primate orbitofrontal cortex. J. Neurosci. 14:5437–5452.
Russchen, F.T., D.G. Amaral, and J.L. Price (1985) The afferent connectionsof the substantia innominata in the monkey, Macaca fascicularis. J.Comp. Neurol. 242:1–27.
Saper, C.B. (1985) Organization of cerebral cortical afferent systems in therat. II. Hypothalamic projections. J. Comp. Neurol. 237:21–46.
Saper, C.B. (1990) Hypothalamus. In G. Paxinos (ed): The Human NervousSystem. San Diego: Academic Press Inc. pp. 389–413.
Saper, C.B. (1995) Central Autonomic System. In G. Paxinos (ed): The RatNervous System. San Diego: Academic Press Inc. pp. 107–135.
Sesack, S.R., A.Y. Deutch, R.H. Roth, and B.S. Bunney (1989) Topographi-cal organization of the efferent projections of the medial prefrontalcortex in the rat: An anterograde tract-tracing study with Phaseolusvulgaris leucoagglutinin. J. Comp. Neurol. 290:213–242.
Siegel, A., H. Edinger and H. Lowenthal (1974) Effects of electricalstimulation of the medial aspect of the prefrontal cortex upon attackbehavior in cats. Brain Res. 66:467–479.
Siegel, A., H. Edinger, and M. Dotto (1975) Effects of electrical stimulationof the lateral aspect of the prefrontal cortex upon attack behavior incats. Brain Res. 93:473–484.
Simerly, R.B. (1995) Anatomical substrates of hypothalamic integration. InG. Paxinos (ed): The Rat Nervous System. San Diego: Academic PressInc. pp. 353–376.
Swanson, L.W. (1987) The Hypothalamus. In A.Bjorklund, T.Hokfelt, andL.W. Swanson (eds): Handbook of Chemical Neuroanatomy, Vol. 5.Amsterdam: Elsevier, pp. 1–124.
Szabo, J. and W.M. Cowan (1984) A stereotaxic atlas of the brain of thecynomolgus monkey (Macaca fascicularis). J. Comp. Neurol. 222:265–300.
Terreberry, R.R. and E.J. Neafsey (1987) The rat medial frontal cortexprojects directly to autonomic regions of the brainstem. Brain Res. Bull.19:639–649.
Van Der Kooy, D., J.F. McGinty, L.Y. Koda, C.R. Gerfen, and F.E. Bloom(1982) Visceral cortex: A direct connection from prefrontal cortex to thesolitary nucleus in rat. Neurosci. Lett. 22:123–127.
VanEssen, D.C. and J.H.R. Maunsell (1980) Two-dimensional maps of thecerebral cortex. J. Comp. Neurol. 19:255–281.
Wall, P.D. and G.D. Davis (1951) Three cerebral cortical systems affectingautonomic function. J. Neurophysiol. 14:507–517.
Yasui, Y., C.D. Breder, C.B. Saper, and D.F. Cechetto (1991) Autonomicresponses and efferent pathways from the insular cortex in the rat. J.Comp. Neurol. 303:355–374.
PREFRONTAL PROJECTIONS TO THE HYPOTHALAMUS 505