organization of amygdaloid projections to the mediodorsal thalamus and prefrontal cortex: a...
TRANSCRIPT

THE JOURNfi OF COMPARATIVE NEUROLOGY 262:46-58 (1987)
Organization of Amygdaloid Projections to the Mediodorsal Thalamus and Prefrontal
Cortex: A Fluorescence Retrograde Transport Study in the Rat
ALEXANDER J. McDONALD Department of Anatomy, University of South Carolina,
Columbia, South Carolina 29208
ABSTRACT Previous studies have shown that the amygdala projects to both the
mediodorsal thalamic nucleus (MD) and its cortical projection area, the prefrontal cortex (PFC). In this investigation rats received injections of different fluorescent retrograde tracers (true blue and diamidino yellow) into MD and either the lateral, polar, or medial PFC in order to examine the relationship of amygdaloid neurons with cortical andor thalamic projec- tions. PFC injections labeled neurons in the basolateral (BL), basomedial (BM), ventral endopiriform (En,), and rostra1 lateral nuclei as well as the periamygdaloid cortex (PAC) and the medial part of the amygdalohippocarn- pal area (AHA). In BL, which contained the great majority of neurons projecting to PFC, most labeled cells were concentrated in particular parts of the nucleus and were topographically organized. The overwhelming ma- jority of labeled neurons in BL were large pyramidal or piriform cells that correspond to class I neurons described in Golgi studies. Occasional small neurons with thin dendrites were also observed; these cells may be class I1 neurons.
MD injections labeled numerous cells in the anterior division of the cortical nucleus, medial nucleus, and caudomedial part of the central nu- cleus. Moderate numbers of labeled cells were found in caudal portions of BM and PAC, whereas scattered cells were observed throughout the rest of the amygdala with the exception of the lateral nucleus. In BL and AHA many MD-projecting neurons were observed along nuclear boundaries and in the adjacent white matter. Neurons in BL, BM, and AHA usually had large elongated or irregular somata and two to four primary dendrites that branched sparingly. Other cells had smaller ovoid somata. The morphology and distribution of MD-projection cells in the basolateral amygdala indicate that they are primarily large class I1 neurons.
Double-labeled amygdaloid neurons, labeled by both cortical and thal- amic injections, were observed only in a small number of animals. Control experiments suggest that most of the double-labeled cells in these cases were artifacts caused by spread of the thalamic injectate into the third ventricle with subsequent uptake by fibers in the anterior commissure. Thus the findings of this study suggest that different neuronal populations in the amygdala project to the two poles of the MD-PFC system. In the basolateral amygdala class I neurons are the predominant cell type involved in PFC projections, whereas a subpopulation of class I1 neurons, hitherto thought to be primarily local-circuit neurons, project to MD.
Key words: amygdala, true blue, diamidino yellow, frontal cortex, limbic system
The prefrontal cortex (PFC) and its associated thalamic nucleus, the mediodorsal nucleus (MD), appear to play an Accepted February 11,1987.
0 1987 ALAN R. LISS. INC.

AMYGDALOID PROJECTIONS TO THALAMUS AND CORTEX 47
important role in learning and memory, and the elabora- tion of complex behavior (e.g., Markowitsch, '82; Kolb, '84). Since these activities are often modulated by motivational and affective factors, it is not surprising that both MD and PFC receive projections from the limbic system, particu- larly the amygdala. In primates, and also in rodents, the amygdala projects primarily to the medial portions of MD and to the part of the PFC that receives projections from the medial MD, namely, the ventral and medial PFC (Nauta, '61, '62; Jacobson and Trojanowski, '75; Krettek and Price, '77; Porrino et al., '81; Amaral and Price, '84).
The PFC, which by definition consists of the portion of the cortex that receives afferents from MD (Rose and Wool- sey, '481, receives amygdaloid projections mainly from the basolateral portion of the amygdala in both primates and rodents. In the rodent the basolateral amygdala projects to prefrontal cortical areas located on lateral, polar, and me- dial aspects of the frontal lobe (Krettek and Price, '77; Sarter and Markowitsch, '83, '84). These areas are, respec- tively, the dorsal agranular insular area, the rostralmost part of the medial precentral area, and the prelimbic-in- fralimbic cortical region. In rodents and primates neurons projecting to MD are found in small numbers in almost all amygdaloid nuclei (Siege1 et al., '77; Price and Slotnick, '83; Aggleton and Mishkin, '84).
Although both poles of the MD-PFC system receive pro- jections from the basolateral amygdala, it is not known whether individual neurons send axonal collaterals to both regions or whether different cells project to each area. In the present investigation this question has been studied in the rat by the multiple retrograde fluorescent tracer tech- nique (Kuypers and Huisman, '84).
MATERIALS AND METHODS In this study 35 Sprague-Dawley rats (225-275 g) re-
ceived injections of different fluorescent retrograde tracers into MD and one of the three major PFC regions that receive projections from the basolateral amygdala (lateral, polar, or medial PFC). The fluorescent dyes true blue (TB) and diamidino yellow hydrochloride (DY) were used in each animal. Cells retrogradely labeled by TB exhibit a blue fluorescent labeling of the perikarya and nucleolus; DY- labeled cells display a yellow fluorescent labeling of the nucleus and, in some instances, a light labeling of the perikarya and dendrites. TB and DY have similar retro- grade transport rates and have been used effectively in many different systems in the rat brain to identify individ- ual neurons (double-labeled cells) projecting axonal collat- erals to two different brain areas wokfelt et al., '83; Kuypers and Huisman, '84).
Injections of TB (5% in distilled water; 0.2-0.3 pl) and DY (2% in 0.2 M phosphate buffer, pH 7.2; 0.4 pl) were made stereotaxically through a 1.0-p1 Hamilton microsyringe equipped with a 26-gauge needle. Stereotaxic coordinates were obtained from the atlas by Paxinos and Watson ('82). Since MD and the lateral and medial parts of PFC extend for several millimeters in the rostrocaudal direction, most attempts to inject these regions involved two iqiections, one into the rostra1 half and the other into the caudal half of these regions. Survival times and injection volumes re- quired to label each amygdaloid projection effectively were determined in preliminary experiments. Attempts to inject MD sometimes resulted in spread of the tracer into the third ventricle dorsomedial to MD and along the needle track through the cingulate cortex dorsal to MD. In control
animals (n = 10) injections were made into the third ventri- cle or cingulate cortex to determine whether the involve- ment of these areas might contribute to retrograde labeling seen in the amygdala.
Four days after the dye injections animals were perfused through the heart with hypertonic saline (1.5% NaC1) fol- lowed by 20% formalin in 0.1 M citrate buffer (pH 7.18). Brains were removed, postfixed for 1-3 h, and sectioned at 40 pm in the coronal plane on a Vibratome. Alternate sections were mounted on gelatinized slides and stored in the dark at 4°C until examined with an Olympus BRA fluorescence microscope with a UG-1 excitation filter, DM400 dichroic mirror, and an L-420 barrier filter. Every other section was mounted and stained with a Nissl stain (pyronin-Y) to facilitate the localization of retrogradely la- beled neurons observed in adjacent sections. Nissl-stained sections at six levels of the amygdala (Bregma -1.8, -2.3, -2.8, -3.3, -3.8, -4.3 of the atlas by Paxinos and Watson, '82) were traced with the aid of a drawing tube using brightfield optics. Blood vessels were then drawn using darkfield optics. Since these same blood vessels could also be identified in adjacent unstained sections viewed by fluo- rescence microscopy, they provided landmarks that allowed the position of neurons retrogradely labeled with fluores- cent dyes to be accurately charted on Nissl tracings. Injec- tion sites were charted by using the same method used to map labeled neurons except that Nissl-stained sections were drawn with the aid of a Bausch and Lomb microprojector.
In general, the description of amygdaloid nuclei by Kret- tek and Price ('78) has been used to designate the location of labeled neurons and axons. However, Turner and Zim- mer's ('84) delineation of the anterior division of the baso- lateral nucleus (AB1 of Turner and Zimmer) and the two subdivisions of the lateral nucleus (AL1 and AL2 of Turner and Zimmer) was used to describe those nuclei. In the present account AL1 is termed the dorsolateral subdivision of the lateral nucleus (Ld) and AL,z is called the ventrome- dial subdivision (Lv). The central nucleus is subdivided according to the description by McDonald ('82a). This de- scription of the central nucleus recognizes four subdivisions arranged from medial to lateral: medial (CM), intermediate (GI), lateral (CL) and lateral capsular (CLC) subdivisions. Reference to several brain series stained for acetylcholin- esterase with alternate sections counterstained with cresyl violet (Butcher, '83) facilitated the identification of the ex- act boundaries between amygdaloid nuclei (Paxinos and Watson, '82; Turner and Zimmer, '84).
RESULTS In approximately two thirds of the animals used in this
study (n = 22), DY was injected into MD and TB was injected into the PFC. In the other 13 animals, the dye placements were reversed. The distribution of retrogradely labeled neurons seen with injections into each of the four areas under investigation (MD, lateral PFC, polar PFC, and medial PFC) was unique for each area and did not depend on the dye injected. Because of the larger size of most TB injections, these injections involved each cortical region more completely than injections of DY. The thalamic injections of DY in these same animals were large enough to involve all of the amygdalorecipient part of MD (medial MD). Although three representative cases in which this dye combination was used are described in detail, the results of animals with reversed dye placements were very similar.
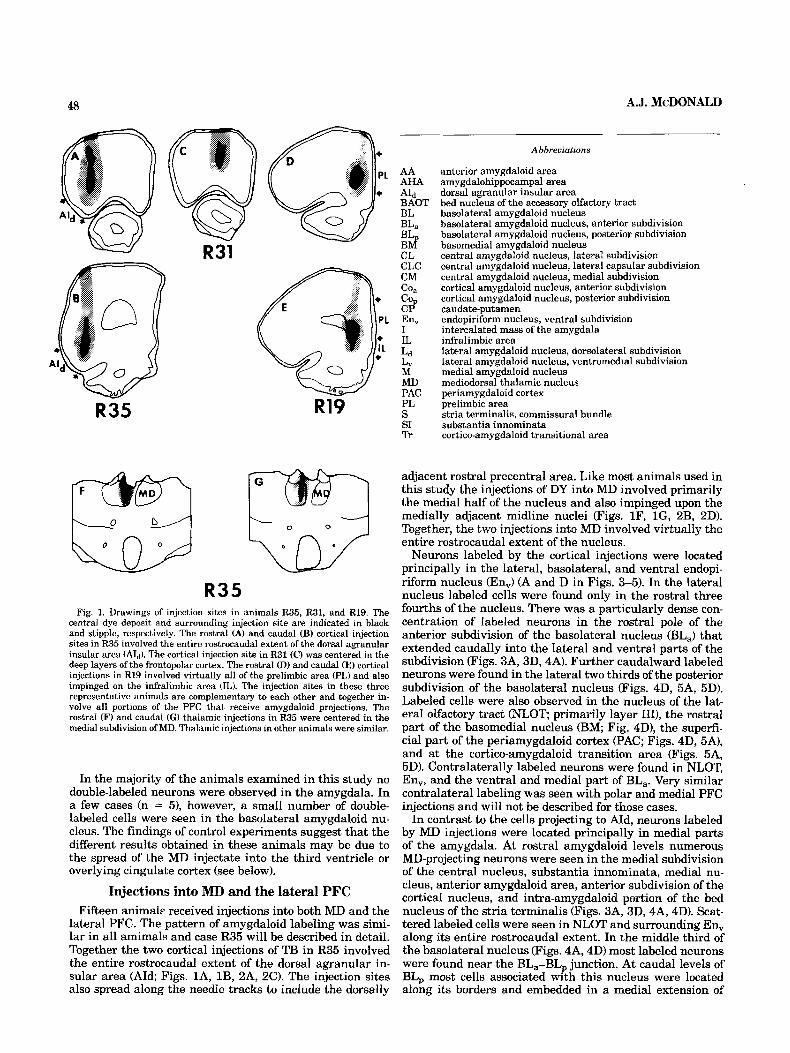
A.J. McDONALD 48
R35
R31
t
PL
+ AA AHA AId BAOT BL EL, BL
CL Bbf
A bhreuiatwns
anterior amygdaloid area amygdalohippocampal area dorsal agranular insular area bed nucleus of the accessory olfactory tract basolateral amygdaloid nucleus basolateral amygdaloid nucleus, anterior subdivision basolateral amygdaloid nucleus, posterior subdivision basomedial amygdaloid nucleus central arnygdaloid nucleus, lateral subdivision
CLC CM Go,
3 caudate-putamen
I
central amygdaloid nucleus, lateral capsular subdivision central amygdaloid nucleus, medial subdivision cortical amygdaloid nucleus, anterior subdivision cortical amygdaloid nucleus, posterior subdivision
pL En, endopiriform nucleus, ventral subdivision
+ IL infralimbic area '1 Ld lateral amygdaloid nucleus, dormlateral subdivision ' L, lateral amygdaloid nucleus, ventromedial subdivision
intercalated mass of the amygdala
M medial amygdaloid nucleus MD mediodorsal thalamic nucleus PAC periamygdaloid cortex PL prelimbic area R19 S stria terminalis, commissural bundle SI substantia innominata TY cortico-amygdaloid transitional area
R35 Fig. 1. Drawings of injection sites in animals R35, R31, and R19. The
central dye deposit and surrounding injection site are indicated in black and stipple, respectively. The rostral (A) and caudal (B) cortical injection sites in R35 involved the entire rostrocaudal extent of the dorsal agranular insular area (Aid). The cortical injection site in R31 (C) was centered in the deep layers of the frontopolar cortex. The rostra1 (D) and caudal (E) cortical injections in R19 involved virtually all of the prelimbic area (PL) and also impinged on the infralimbic area (IL). The injection sites in these three representative animals are complementary to each other and together in- volve all portions of the PFC that receive amygdaloid projections. The rostral (F) and caudal (G) thalamic injections in R35 were centered in the medial subdivision of MD. Thalamic injections in other animals were similar.
In the majority of the animals examined in this study no double-labeled neurons were observed in the amygdala. In a few cases (n = 51, however, a small number of doubIe- labeled cells were seen in the basolateral amygdaloid nu- cleus. The findings of control experiments suggest that the different results obtained in these animals may be due to the spread of the MD injectate into the third ventricle or overlying cingulate cortex (see below).
Injections into RID and the lateral PFC Fifteen animal? received injections into both MD and the
lateral PFC. The pattern of amygdaloid labeling was simi- lar in all amimals and case R35 will be described in detail. Together the two cortical injections of TB in R35 involved the entire rostrocaudal extent of the dorsal agranular in- sular area (AId; Figs. lA, lB, 2A, 2C). The injection sites also spread along the needle tracks to include the dorsally
adjacent rostral precentral area. Like most animals used in this study the injections of DY into MD involved primarily the medial half of the nucleus and also impinged upon the medially adjacent midline nuclei (Figs. lF, IG, ZB, 2D). Together, the two injections into MD involved virtually the entire rostrocaudal extent of the nucleus.
Neurons labeled by the cortical injections were located principally in the lateral, basolateral, and ventral endopi- riform nucleus (En,) (A and D in Figs. 3-5). In the lateral nucleus labeled cells were found only in the rostral three fourths of the nucleus. There was a particularly dense con- centration of labeled neurons in the rostral pole of the anterior subdivision of the basolateral nucleus @La) that extended caudally into the lateral and ventral parts of the subdivision (Figs. 3A, 3D, 4A). Further caudalward labeled neurons were found in the lateral two thirds of the posterior subdivision of the basolateral nucleus (Figs. 4D, 5A, 5D). Labeled cells were also observed in the nucleus of the lat- eral olfactory tract (NLOT primarily layer 1111, the rostral part of the basomedial nucleus (BM; Fig. 4D), the superfi- cial part of the periamygdaloid cortex (PAC; Figs. 4D, 5A), and at the cortico-amygdaloid transition area (Figs. 5A, 5D). Contralaterally labeled neurons were found in NLOT, En,, and the ventral and medial part of BL,. Very similar contralateral labeling was seen with polar and medial PFC injections and will not be described for those cases.
In contrast to the cells projecting to AId, neurons labeled by MD injections were located principally in medial parts of the amygdala. At rostral amygdaloid levels numerous MD-projecting neurons were seen in the medial subdivision of the central nucleus, substantia innominata, medial nu- cleus, anterior amygdaloid area, anterior subdivision of the cortical nucleus, and intra-amygdaloid portion of the bed nucleus of the stria terminalis (Figs. 3A, 3D, 4A, 4D). Scat- tered labeled cells were seen in NLOT and surrounding En,, along its entire rostrocaudal extent. In the middle third of the basolateral nucleus (Figs. 4A, 4D) most labeled neurons were found near the BL,-BL, junction. At caudal levels of BL, most cells associated with this nucleus were located along its borders and embedded in a medial extension of

AMYGDALOID PROJECTIONS TO THALAMUS AND CORTEX 49
Fig. 2. A Brightfield photomicrograph of the caudal TB injection site in the dorsal agranular insular cortex in case R35 (section stained with py- ronin Y). Asterisks indicate the extent of the dorsal agranular insular area. Compare with Figure 1B. Rectangle indicates field shown in C. Scale bar = 1 mm. B: Brightfield photomicrograph of the caudal DY injection site in the mediodorsal nucleus in case R35. Section stained with pyronin Y. Dotted
the external capsule that is located dorsal to BL,, (Figs. 5A, 5D). Additional MD-projecting neurons were found in small numbers in BM, PAC, and the amygdalohippocampal area (AHA). There were also some labeled neurons located in the white matter just dorsal to the AHA. No double-labeled neurons were observed in the amygdala.
In R35 and all of the other animals used in this study the morphology of neurons in the basolateral amygdala project- ing to the PFC was different from that of neurons projecting to MD. This was true regardless of the dye used to label the cells. Neurons in the basolateral amygdala projecting to cortical areas were mostly large pyramidal or piriform cells
lines indicate the borders of the left and right MD. The injection is centered in the medial part of the left MD. Compare with Figure 1G. Rectangle indicates field shown in D. Scale bar = 250 pm. C, U: Fluorescence photom- icrographs of the rectangular fields indicated in A and R, respectively, photographed from adjacent unstained sections. Magnification is the same as in U.
(Fig. 6A). Occasionally smaller neurons with thin dendrites were observed. Neurons projecting to MD usually had large polygonal, irregular or elongated perikarya and two to foul; dendrites that branched sparingly (Fig. 6B-H). Other cells had smaller, more ovoid cell bodies. The MD-projecting neurons located deep to the piriform cortex resembled those in the basolateral amygdala.
Injections into RID and the polar PFC Four animals received injections into both MD and the
prefrontal cortex located at the frontal pole. The pattern of

50 A.J. McDONALD

AMYGDALOID PROJECTIONS TO THALAMUS AND CORTEX 51
amygdaloid labeling was similar in all cases and R31 will be described in detail. The cortical injection of TB in R31 involved the deep half of the frontopolar cortex (Fig. 1C). The injections of DY into MD were confined to the medial half of the nucleus and together involved the rostral three fourths of MD as well as medially adjacent midline nuclei. The distribution of MD-projecting cells was similar to that seen in R35, but somewhat fewer labeled neurons were seen in the vicinity of the rostral pole of the central nucleus (Fig. 3B), and more labeled cells were observed in the caudal part of the medial nucleus (Fig. 4E).
As with injections into the lateral PFC (R35), injections into the polar PFC (R31) resulted in many labeled cells in the basolateral amygdala and layer 111 of N M T (B and E in Figs. 3-5). However, the areas of the basolateral amyg- dala that contained concentrations of labeled neurons dif- fered. Thus, the number of labeled neurons in the lateral nucleus and BL, in R31 was considerably reduced com- pared to R35 (Figs. 4,5). In BL, the region containing dense concentrations of labeled cells was shifted caudally and medially compared to animals with lateral PFC injections. There were many labeled cells in medial and ventral por- tions of BL, with very few labeled neurons in the rostra1 pole and lateral part of the nucleus (Figs. 3, 4). No double- labeled cells were observed.
Injections into MD and the medial PFC Sixteen animals received injections into both MD and the
medial PFC. As with the other combinations of injection sites in this study the pattern of amygdaloid labeling was very similar in all 16 animals. Case R19 will be described in detail and compared to animals that received cortical injections into the lateral and polar PFC (R35 and R31, respectively). The cortical injections of TB in R19 involved virtually the entire rostrocaudal extent of the prelimbic area and also impinged on the ventrally adjacent infralim- bic area (Figs. lD, 1E). Label along the needle tracks may have resulted in slight involvement of the anterior cingu- late cortex. Together the MD injections of DY involved the rostral three fourths of the nucleus and the medially adja- cent midline nuclei. The distribution of MD-projecting neu- rons in the amygdala was similar to that seen in R35 and R31 (Figs. 3-5). No double-labeled neurons were observed in the amygdala.
As with injections into other parts of the PFC, neurons labeled by the cortical injections were observed in the ba- solateral amygdala, ventral endopiriform nucleus, and layer 111 of NLOT (C and F, Figs. 3-5). Although very few neurons were labeled in Ld in R19, there were moderate numbers of labeled cells in the rostral part of L, (C and F in Figs. 3 and 4). There were far more labeled cells in the caudal parts of PAC than in R31 and R35 (Fig. 5C). The retrograde labeling in the lateral part of the amygdalohippocampal area Pigs. 5C, 5F) was not seen with injections into other regions of PFC .
Regions in BL, that contained concentrations of labeled ceIIs overlapped with amygdaloid areas that were densely labeled by polar and, to a lesser extent, lateral PFC injec-
Fig. 3. Tracings of sections through the amygdala in animals R35 (A,D), R31 (B,E), and R19 (C,F) at the Bregma -1.8 (A-C) and Bregma -2.3 (0- F) levels of the atlas by Paxinos and Watson ('82). In each animal solid symbols indicate neurons labeled by TB injections into the PFC and open symbols indicate neurons labeled by DY injections into MD.
tions but, in general, were shifted dorsomedially. Since a similar medial shift was noted when the amygdaloid label- ing patterns produced by lateral and polar PFC injections were compared, it appears that %La neurons projecting to different parts of PFC exhibit an overlapping topographical organization. This organization is seen most clearly in Fig- ure 4. The cortical projection of BL, is also topographically organized (Figs. 5A-5C). The lateral part of BL, projects to the lateral PFC, the central part to the polar PFC, and the medial part to the medial PFC.
Control experiments In five animals a small number of double-labeled cells
(two to four per section) were observed in the caudomedial part of BL,. These cases included animals from each of the three PFC groups. The double-labeled cells were always intensely labeled by the cortical injections and very lightly labeled by the MD injections. This labeling characteristic, and the fact that these cells were seen only in a small percentage of the animals in this study, suggest that the labeling by the thalamic injection may be due to the occa- sional spread of dye to structures surrounding MD.
The most likely cause of the double labeling was diffusion of the MD injectate into the third ventricle. The lateral edge of the third ventricle is located just dorsal to the amygdalorecipient portion of MD. As revealed by labeling of the ependyma, variable amounts of dye diffused into the third ventricle in several of the animals used in this study. This ependymal labeling was fairly intense in the five ani- mals with double-labeled cells. In these cases there were numerous cells in BL, and NLOT bilaterally that were retrogradely labeled by the MD injectate. Since both the medial BL, and NLOT project axons through the anterior commissure, which forms part of the rostral wall of the third ventricle, it seems likely that in the cases with dou- ble-labeled cells the dye injected into MD spread into the adjoining third ventricle and labeled axons fron NLOT and the medial BL,, which cross in the comrnissure. In support of this interpretation, control animals with injections into the third ventricle exhibited labeled cells bilaterally in NLOT, BL,, and other regions known to send fibers through the anterior commissure. When injections into the third ventricle were paired with PFC injections, numerous dou- ble-labeled cells were observed in BL,. Eisenman ('85) has reported similar false-positives with injections of fast blue into the cerebrospinal fluid of the subarachnoid space.
It is also possible that the double-labeled cells seen in these five animals could be caused by spread of the MD injectate up the needle track to involve the retrosplenial cingulate cortex. However, as in previous studies (Sripan- idkulchai et al., '84) even large injections of TB (0.2-0.3 pl) into the retrosplenial cingulate cortex overlying MD in control animals produce few labeled neurons in BL,. When these cingulate injections were paired with PFC injections very few double-labeled cells (0-5 cells per brain) were seen in the amygdala. Thus, the results of the control experi- ments suggest that most of the double-labeled cells seen in a small minority of the animals used in this study were due to spread of MD injectate into the third ventricle.
DISCUSSION The present investigation is the first study to examine
the organization of amygdaloid neurons projecting to the prefrontal cortex and mediodorsal thalamic nucleus by means of the multiple retrograde fluorescent tracer tech-


AMYGDALOID PROJECTIONS TO THALAMUS AND CORTEX 53
nique. The results suggest that separate neuronal popula- tions in the amygdala project to MD and the PFC and that the projections of the basolateral amygdala to the PFC are topographically organized.
Arnygdalocortical projections As in previous studies (Krettek and Price, '77; R e p and
Winans, '82; Sarter and Markowitsch, '84) the present in- vestigation found a strong projection from BLa to &. The projection of BL, to AId revealed in the present study was also observed in the hamster (Reep and Winans, '82), but perhaps because of the use of the sensitive retrograde flu- orescent tracer technique, more cells were seen in the pres- ent study in the rat. The failure of Krettek and Price ('77) to demonstrate this projection with autoradiographic tech- niques is probably due to the fact that their injections of BL,, which involved mainly central and medial portions of the nucleus, missed most of the cells of origin of this projec- tion. In addition, the number of cells involved in this projec- tion are fewer than those involved in the projection arising in BL,. The failure of Sarter and Markowitsch ('84) to observe with fluorescent retrograde tracers a projection from BL, to AId in Wistar rats is difficult to explain. It may be related to the exact placement of the injection sites or strain differences. The projection of the ventral endopiriform nu- cleus to Ah, and other parts of PFC, has not been reported in previous studies.
The projection of the lateral nucleus to AId seen in the present study was not noted in autoradiographic studies of amygdaloid efferents (Krettek and Price, '77) but has been observed in previous retrograde transport studies (Reep and Winans, '82; Sarter and Markowitsch, '84). Although Sarter and Markowitsch, ('84) appeared to label cells throughout the rostrocaudal extent of the lateral nucleus with lateral PFC injections, in the present study and in a study of AId afferents in the hamster (Reep and Winans, '82) neurons in the caudal portions of the lateral nucleus were not labeled. The present investigation is the first study to report a projection from the basomedial nucleus to the lateral PFC. Although some cells in the corticomedial amygdala were labeled in the present study with AId injections, previous studies have shown that this portion of the amygdala pro- jects primarily to the adjacent ventral agranular insular area (Krettek and Price, '77; Kevetter and Winans, '81; Reep and Winans, '82; Luskin and Price, '83).
The cortex of the frontal pole has been shown to be pre- frontal (MD projection) cortex in several nonprimate mam- malian species (Benjamin et al., '78; Golden et al., '78) and recently Sarter and Markowitsch ('83) reported a projection from MD to this area in the rat. These investigators also labeled neurons in the lateral and basolateral amygdaloid nuclei with injections into this area. The results of the present study suggest that most of the cells in the amygdala projecting to the polar PFC are found in BL,, although some neurons in BL,, En,, and the lateral nucleus also project to this area.
Although the results of initial autoradiographic studies suggested that the prelimbic and infralimbic areas of the
Fig. 4. Tracings of sections through the amygdala in animals B35 (A$), R31 (B,E), and R19 (C,F) at the Bregma -2.8 (A-c) and Bregma -3.3 (n- F) levels of the atlas bv Paxinos and Watson ('82). In each animal solid
medial frontal cortex receive only light projections from BLa (Krettek and Price, '77) subsequent retrograde trans- port studies (Sarter and Markowitsch, '84; present investi- gation) reveal that BL,, especially its medial portion, actually has strong projections to these cortical areas. The labeling in BM, B b , and the lateral nucleus described in the present study is similar to that reported by Sarter and Markowitsch ('84) except that we did not observe retrograde labeling in the caudal part of the lateral nucleus. The retrograde labeling in the ventral endopiriform nucleus and amygdalohippocampal area seen in the present study has not been described previously. The labeled cells seen in the corticomedial amygdala were probably due to the in- volvement of the infralimbic area in the injection site Kret- tek and Price, '77; Luskin and Price, '83).
The present study clearly indicates that the projections from the anterior and posterior subdivisions of the basola- teral nucleus to the PFC exhibit an overlapping topograph- ical organization. In general, displacement of injection sites from lateral to polar to medial PFC is paralleled by a shift of densely labeled areas of the basolateral nucleus in the caudomediodorsal direction. In addition, densely labeled areas of BLa tend to blend dorsally with labeled areas in the lateral nucleus. There appears, however, to be more overlap in the topographical organization of lateral nucleus efferents. These findings corroborate and extend the results of a recent fluorescent retrograde tracer study of amygda- loid efferents to the cortex in the rat (Sripanidkulchai et al., '84). These investigators studied amygdaloid projections to various cortical areas but focused on areas situated just caudal to the lateral, polar, and medial PFC (the gustatory insular cortex, motor cortex, and cingulate cortex, respec- tively). Comparisons of the maps of labeled neurons in the present study with those of Sripanidkulchai et al. ('84) suggest that displacement of injection sites from the medial PFC to the caudally adjacent cingulate area continues the lateral-to-medial shift of densely labeled areas of the baso- lateral nucleus noted in the present study. When injection sites are displaced from the polar PFC to the caudally adjacent motor cortex, the region of dense labeling in the basolateral nucleus remains relatively constant but the number of labeled cells diminishes as injections are moved more caudally within the motor cortex. Injection sites in the lateral PFC produce labeling in the basolateral nucleus that is very similar to that seen with injections into the caudally adjacent gustatory insular cortex.
The results of a retrograde transport study of amygdaloid projections to the posterior agranular insular cortex and the caudally adjacent perirhinal cortex (PR) indicate that neurons in the basolateral amygdala projecting to these cortical areas are also topographically organized WC- Donald and Jackson, '87). Neurons in the basolateral nu- cleus projecting to AI, overlap those projecting to the rostrally adjacent AId but are concentrated in the far lat- eral portion of the nucleus. Most neurons projecting to PR are found in the basomedial nucleus and the caudal part of the lateral nucleus, which was unlabeled after AI, and PFC injections. Thus taken together, the results of several re- cent studies of amygdaloid efferents to the cerebral cortex of the rat (Sripanidkulchai et al., '84; McDonald and Jack- son, '87; and the present study) suggest that the entire Droiection of the basolateral arnygdala to frontal, insular, a n i temporal cortex is topographically organized with ad- jacent portions of the amygdala projecting to adjoining por- tions of the amygdalorecipient cortex. The degree of overlap of domains in the basolateral amygdala Projecting to two
. .
symbols indicate new& labeled by TB injections into the PFC and open symbols indicate neurons labeled by DY injections into MD.

n
Figu
re 5

AMYGDALOID PROJECTIONS TO THALAMUS AND CORTEX 55
different cortical areas is related to the proximity of the cortical areas to each other, and some neurons may send collaterals to different cortical areas (Sarter and Markow- itsch, '84). Although it would seem likely that such a basic orderly pattern of connections between the basolateral amygdala and cortex would exist in other species, there have been no systematic attempts to examine this question in other animals.
The topographical organization of PFC Projections from the cat basolateral nucleus (Llamas et al., '85) resembles that seen in the rat in the present study. As in the rat, neurons in the anterior subdivision of the basolateral nu- cleus projecting to the lateral PFC tend to be located ven- trolateral to those projecting to the medial PFC (see Fig 3B of Llamas et al., '85). Studies of amygdaloid efferents to the primate PFC reveal that, as in rodents, most projections arise from the basal group of nuclei (Jacobson and Troja- nowski, '75; Porrino et al., '81; Amaral and Price, '84). Although there is evidence that different portions of the primate basal nuclear complex project preferentially to dif- ferent parts of the PFC, details regarding the exact topo- graphical ordering of these projections are not available. Since different portions of the PFC of both rodents and primates are involved in different functions (Rosenkilde, '79; Eichenbaum et al., '83; Kolb, '84), the elucidation of the topographical organization of amygdaloid efferents to these areas is a first step in developing an understanding of how the amygdala modulates the activity of this behav- iorally important brain region.
Amygdalothalamic projections In their classic autoradiographic study of amygdaloid ef-
ferents in the rat, Krettek and Price ('77) reported amyg- daloid projections to MD from the basolateral nucleus. Subsequent retrograde transport studies with HRP or wheat germ agglutinin-conjugated HRP as a tracer demonstrated that small numbers of neurons with MD projections were scattered throughout many other amygdaloid nuclei as well (Siegel et al., '77; Price and Slotnick, '83). The present study is the first investigation of amygdaloid projections to MD using the sensitive fluorescent retrograde transport tech- nique. The distribution of labeled neurons is basically sim- ilar to that seen in past studies of MD afferents in the rat (Siegel et al., '77; Price and Slotnick, ,831, but many more labeled neurons were observed in the present investigation. The labeling observed in the central nucleus in the present study may be due to involvement of the midline nuclei in the injection sites, since this nucleus did not contain labeled cells in previous HRP studies in which injection sites avoided the midline nuclei (Siegel et al., '77; Price and Slotnick, '83). In an HRP study in the opossum in which the injections involved both MD and the midline nuclei, there were numerous labeled cells in the central and medial nuclei and small numbers of labeled cells in other amygda- loid nuclei, as in the present study (Benjamin et al., '82). In the rat it appears that the media1 nucleus has a light
Fig. 5. Tracings of sections through the amygdala in animals R35 (A$), R31 (B,E), and R19 (C,F) at the Bregma -3.8 (A-C) and Bregma -4.3 (D- F) levels of the atlas by Paxinos and Watson ('82). In each animal solid symbols indicate neurons labeled by TI3 injections into the PFC and open symbols indicate neurons labeled by DY injections into MD.
projection to both the medial MD and the adjacent midline nuclei (see Fig. 8 of Luiten et al., '85).
The retrograde labeling in the amygdala following injec- tions of HRP into the monkey MD is remarkably similar t o that seen in the rat in the present study (Aggleton and Mishkin, '84). As in the rat, it appears that the central and medial nuclei in the monkey project primarily to midline nuclei rather than MD (Price and Amaral, '81; Aggleton and Mishkin, '84). Surprisingly, there is no significant amygdaloid projection to MD in the cat (Krettek and Price, '77; Irle et al., '84; Velayos and Reinoso-Suarez, '85).
Neuron populations projecting to the MI)-PFC system
The present study corroborates the finding of Siegel et al. ('77) that, in general, most neurons projecting to the dorso- medial thalamus in the rat are located in the corticomedial amygdala whereas most neurons projecting to the PFC are found in the basolateral amygdala. However, the present investigation demonstrates that neurons projecting to MD and the PFC are intermixed in the basolateral portion of the amygdala. Since most, if not all, of the double-labeled cells noted in some of the cases in this study can probably be dismissed as artifacts (see Results), it appears that few, if any, neurons in the basolateral portion of the amygdala send axonal collaterals to both MD and the PFC.
One of the most interesting and surprising results of the present study was the finding that neurons in the basola- teral amygdala projecting to the PFC differ morphologi- cally from cells projecting to MD. The perikaryal morphology of most PFC-projecting neurons in the basola- teral amygdala, as well as their bountiful numbers, clearly indicates that these cells correspond to the class I neurons described in Golgi studies (McDonald, '82b, '84). In Golgi preparations these cells are characterized by their pyrami- dal or semipyramidal shape and the dense covering of spines on their dendrites. The small neurons labeled by cortical injections, which were rarely observed, may correspond to the class I1 neurons described in Golgi studies (McDonald, '82b, '84). Some small labeled neurons were also seen by Sripanidkulchai et al. ('84) with various cortical injections.
Most neurons in the basolateral amygdala projecting to MD appear to correspond to large and, to a lesser extent, medium-size class I1 neurons described in Golgi studies (McDonald, '82b, '84). In Golgi preparations class I1 neu- rons have spine-sparse dendrites and sometimes exhibit dense local axonal arborizations. Like class I1 cells, neurons labeled by MD injections typically have irregular or elon- gated perikarya and two to four dendrites that branch sparingly. The morphological heterogeneity and sparse dis- tribution of MD-projecting cells is also characteristic of class I1 neurons. It has been shown that the white matter dorsal to BL, and AHA is populated almost entirely by class I1 neurons (McDonald, '84). This area contained many MD-projecting cells in the present study. Likewise, Golgi studies suggest that many class I1 neurons are located along the borders of the basolateral arnygdaloid nuclei, including the BL,-BL, border (McDonald, '84). Neurons labeled by MD injections demonstrated similar tendencies. Golgi studies have demonstrated that class I neurons in BL, are vertically oriented pyramidal cells with ventrally directed apical dendrites, although most class I1 neurons in this nucleus are oriented horizontally or obliquely (Mc- Donald, '84). The orientations of BL, neurons labeled by MD injections closely match those of class I1 cells.
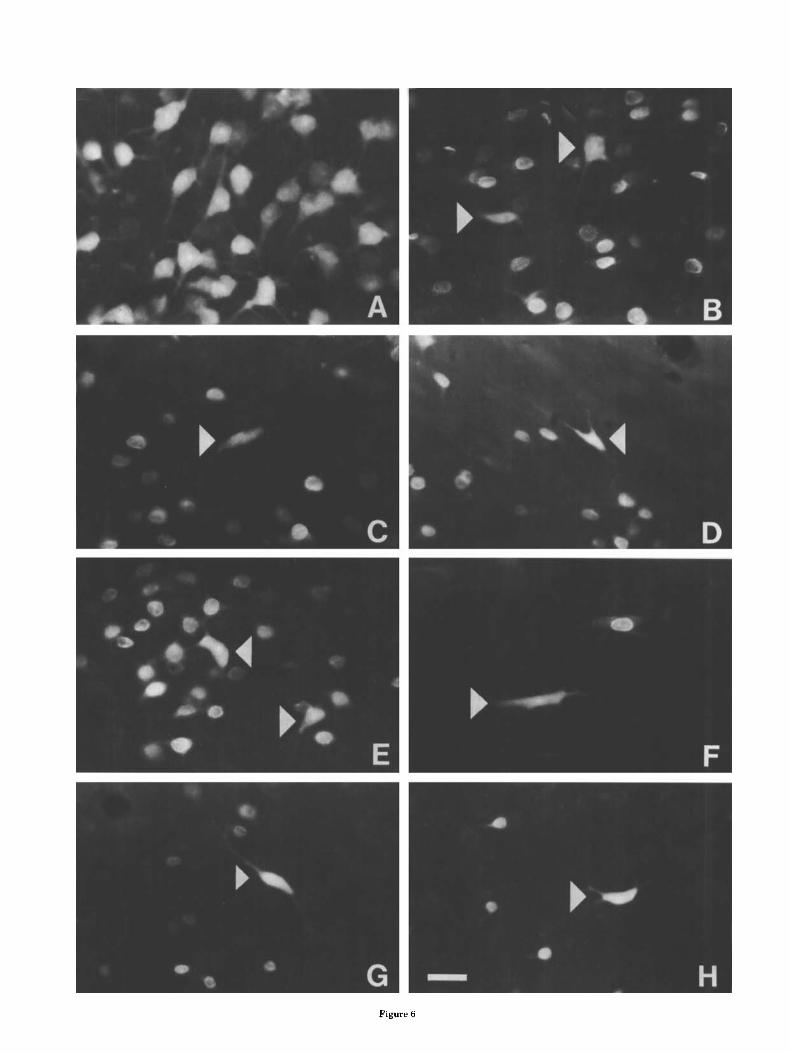
Figure 6

AMYGDALOID PROJECTIONS TO THALAMUS AND CORTEX 57
In Golgi preparations (McDonald, '82b) axons of class I1 neurons in the rat, when impregnated, often exhibit dense local axonal arborizations typical of local-circuit neurons (interneurons). The same is true of class I1 neurons in the opossum (McDonald and Culberson, '81) and similar cells in carnivores (Mukhina and Leontovich, '70; Tombol and Szafranska-Kosmal, '72; Kamal and Tombol, '75). The pres- ent investigation is the first experimental study to provide evidence that one subpopulation of class I1 neurons, mainly large class I1 cells, is the predominant cell type involved in a projection to an extrinsic brain area. It is of interest that in a Golgi study of the dog amygdala Mukhina and Leon- tovich ('70) predicted that their "reticular neurons," which appear to correspond to the large class I1 neurons of the rat basolateral amygdala, were "long-axon" projection neu- rons. Since axons of the large, pyramidal class I neurons are frequently observed coursing out of the amygdala in Golgi preparations (e.g., McDonald, '82b; Millhouse and DeOlmos, '831, it was not surprising to find cells of similar morphology labeled by cortical injections in the present study. The finding that the two poles of the MD-PFC sys- tem appear to receive projections mainly from two different cell types in the basolateral amygdala suggests that the type of information transmitted to MD and the PFC may differ.
Recent studies have pointed out that neurons in the ba- solateral amygdala and neurons in the cerebral cortex in- cluding the hippocampus are very similar in both their morphology and immunohistochemistry (McDonald, '82b, '84, '85a, '85b; McDonald et al., '87; Millhouse and De- Olmos, '83; Carlsen and Heimer, '86). Both basolateral amygdala and cortex have spiny pyramidal neurons that appear to contain excitatory neurotransmitters, and spine- sparse nonpyramidal neurons that contain gamma-amino- butyric acid, acetylcholine, and certain neuropeptides. Tra- ditionally, it has been assumed that the pyramidal and nonpyramidal cells are, respectively, projection neurons and local-circuit neurons in both amygdala and cortex. The re- sults of the present study, however, demonstrate that spine- sparse nonpyramidal (class 11) neurons are the principal cell type of the basolateral amygdala projecting to the thal- amus (mediodorsal nucleus). It is of particular interest, therefore, that recent studies in both rat (Sikes et al., '77) and monkey (Aggleton et al., '86) indicate that spine-sparse nonpyramidal (polymorphic) neurons are also the principal cell type of the subicular portion of the hippocampal cortex projecting to the thalamus (anterior nucleus). Thus, the two major regions of the telencephalic limbic system, the amyg- dala and hippocampal formation, project to their respective thalamic targets via a similar cell type. It seems likely that these spine-sparse nonpyramidal neurons may process in- formation in a similar manner in both the amygdala and hippocampus and transmit a similar type of message to the
Fig. 6. Fluorescence photomicrographs illustrating the morphology of neurons in the basolateral amygdala retrogradely labeled by injections into MD and PFC. A Neurons in BL. labeled by an injection of TB into the lateral PFC. Thick apical dendrites can be observed emanating from many of these large pyramidal or piriform cells. No DY neurons labeled by the MD injection are seen in this field. B-H: Neurons labeled by TB injections into MD (arrowheads) are surrounded by neurons labeled by DY injections into the PFC, MD-projecting neurons were located in BL, (B,C), along the dorsal border of BL, (D), at the border separating BL, from the lateral nucleus (E) in the ventral endopiriform nucleus (F), nnd BM (G,H). Scale bar, 25 p m for all photomicrographs.
thalamus. Although the exact function of the morphologi- cally similar hippocampal and amygdaloid neurons is not known, it i s of considerable interest that they appear to participate in functionally equivalent, parallel, limbicoth- alamic connections that constitute an integral link in a cortico-limbico-thalamo-cortical circuit involved in recogni- tion memory (Mishkin, '82).
ACKNOWLEDGMENTS This work was supported by National Institutes of Health
grant NS19733. The author is grateful for the excellent technical assistance of Peggy Sullivan and secretarial assis- tance of Judy Lawrence,
LITERATURE CITED Aggleton, J.P., and M. Mishkin (1984) Projections of the amygdala to the
thalamus in the cynomolgus monkey. J. Comp. Neurol. 222:56-68. Amaral, D.G., and J.L. Price (1984) Amygdalocortical projections in the
monkey (Macaca f'ciculuris). J. Comp. Neurol. 230:465-496. Benjamin, R.M., J.C. Jackson, and G.T. Golden (1978) Cortical projections
of the thalamic medibdorsal nucleus in the rabbit. Brain Res. 141r251- 266.
Benjamin, R.M., J.C. Jackson, G.T. Golden, and C.H.K. West (1982) Sources of olfactory inputs to opossum mediodorsal nucleus identified by horse- radish peroxidase and autoradiographic methods. J. Comp. Neurol. 207:358-368.
Butcher, L.L. (1983) Acetylcholinesterase histochemistry. In A. Bjorklund and T. Hokfelt (eds): Handbook of Chemical Neuroanatomy, Vol. 1: Methods in Chemical Neuroanatomy. Amsterdam: Elsevier, pp. 1-49.
Carlsen, J., and L. Heimer (1986) A correlated light and electron microscopic immunocytochemical study of cholinergic terminals and neurons in the rat amygdaloid body with special emphasis on the basolateral amygda- loid nucleus. J. Comp. Neurol. 244.121-136.
Eichenbaum, H., R.A. Clegg, and A. Feeley (1983) Reexamination of func- tional subdivisions of the rodent prefrontal cortex. Exp. Neurol. 79:434- 451.
Eisenman, L.M. (1985) Uptake of the retrograde fluorescent tracer fast blue from the cerebrospinal fluid of the rat. Neurosci. Lett, 60:241-246.
Golden, E.T., J.C. Jackson, and R.M. Benjamin (1978) Cortical projection of the thaIamic mediodorsal nucleus in the opossum. Soc. Neurosci. Abstr. 4:221.
Hokfelt, T.G., G. Skagerberg, L. Skirboll, and A. Bjorklund (1983) Combi- nation of retrograde tracing and neurotransmitter histochemistry. In A. Bjorklund and T. Hokfelt (eds.): Handbook of Chemical Neuroanatomy, Vol. 1: Methods in Chemical Neuroanatomy. Amsterdam: Elsevier, pp. 228-285.
Irle, E., H.J. Markowitsch, and M. Streicher (1984) Cortical and subcortical, including sensory-related, afferents to the thalamic mediodorsal nucleus of the cat. J. Hirnforsch. 25.29-51.
Jacobson, S., and J.G. Trojanowski (1975) Amygdaloid projections to prefron- tal granular cortex in rhesus monkey demonstrated with horseradish peroxidase. Brain Res. 100:132-139.
Kamal, A.M., and T. Tombol (1975) Golgi studies of the amygdaloid nuclei of the cat. J. Hirnforsch. 17:175-201.
Keizer, K., H.G.J.M. Kuypers, A.M. Huisman, and 0. Dann (1983) Diami- din0 yellow dihydrochloride (DY SHCL); a new fluorescent retrograde neuronal tracer, which migrates only very slowly out of the cell. Exp. Brain Res. 51:179-191.
Kevetter, G.A., and S.S. Winans (1981) Connections of the corticomedial amygdala in the golden hamster. 11. Efferents of the "olfactory amyg- dala." J. Comp. Neurol. 197.99-111.
Kolb, B. (1984) Functions of the frontal cortex of the rat: A comparative review. Brain Res. Rev. 8:65-98.
Krettek, J.E., and J.L. Price (1977) Projections from the amygdaloid complex to the cerebral cortex and thalamus in the rat and cat. J. Comp. Neurol. 172:681-W.
Krettek, J.E., and J.L. Price (1978) A description of the amygdaloid complex in the rat and cat with observations on intra-amygdaloid axonal connec- tions. J. Comp. Neurol. 178~255-280.
Kuypers, H.G.J.M., and H.M. Huisman (1984) Fluorescent neuronal tracers. Adv. Cell. Neurobiol. 51307-340.
Llamas, A., C. Avendano, and F. Reinoso-Suarez (1985) Amygdaloid projec- tions to the motor, premotor and prefrontal areas of the cats cerebral

58 A.J. McDONALD
cortex: A topographical study using retrograde transport of horseradish peroxidase. Neuroscience 15:651-657.
Luiten, P.G.M., J.M. Koolhaas, S. deBoer, and S.J. Koopmans (1985) The cortico-medial amygdala in the central nervous system organization of agonistic behavior. Brain Res. 3321283-297.
Luskin, M.B., and J.L. Price (1983) The topographic organization of associ- ational fibers of the olfactory system in the rat, including centrifugal fibers in the olfactory bulb. J. Camp. Neurol. 216:264-291.
Markowitsch, H.J. (1982) Thalamic mediodorsal nucleus and memory: A critical evaluation of studies in animals and man. Neurosci. Biobehav. Rev. 6:351-380.
McDonald, A.J. (1982a) Cytoarchitecture of the central amygdaloid nucleus of the rat. J. Comp. Neurol. 208t401-418.
McDonald, A.J. (1982b) Neurons of the lateral and basolateral amygdaloid nuclei: A Golgi study in the rat. J. Comp. Neurol. 212:293-312.
McDonald, A.J. (1984) Neuronal organization of the lateral and basolateral amygdaloid nuclei in the rat. J. Comp. Neurol222t589-606.
McDonald, A.J. (1985a) Immunohistochemical identification of gamma-ami- nobutyric acid containing neurons in the rat basolateral amygdala. Neurosci. Lett. 53:203-207.
McDonald, A.J. (1985b) Morphology of peptide-containing neurons in the rat basolateral amygdaloid nucleus. Brain Res. 3381186-191.
McDonald, A.J., and J.L. Culberson (1981) Neurons of the basolateral amyg- dala: A Golgi study in the opossum (Didelphis uirginiana). Am. J. Anat. 162:317-342.
McDonald, A.J., and T.R. Jackson (1987) Amygdaloid connections with pos- y terior insular and temporal cortical areas in the rat. J. Comp. Neurol.
262:59-77. McDonald, A.J., A.J. Beitz, J.E. Madl, and A.A. Larson (1987) Localization
of putative excitatory neurons in the hippocampus and amygdala: An immunohistochemical study using a monoclonal antibody to gamma glutamylglutamate. Submitted for publication.
Millhouse, D.E., and J. DeOlmos (1983) Neuronal configurations in lateral and basolateral amygdala. Neuroscience 10t1269-1300.
Mishkin, M. (1982) A memory system in the monkey. Phil. Trans. R. Soc. Lond. B. 298t85-92.
Mukhina, Y.K., and T.A. Leontovich (1970) Neuronal structure of some amygdaloid nuclei in dogs. Arkh. Anat. Gistol. Embriol. 59:62-70.
Nauta, W.J.H. (1961) Fibre degeneration following lesions of the amygdaloid complex in the monkey. J. Anat. (Lond.) 95515-531.
Nauta, W.J.H. (1962) Neural associations of amygdaloid complex in the
monkey. Brain 85505-520. Paxinos, G., and C. Watson (1982) The Rat Brain in Stereotaxic Coordinates.
New York: Academic. Porrino, L.J., A.M. Crane, and P.S. Goldman-Rakic (1981) Direct and indi-
rect pathways from the amygdala to the frontal lobe in rhesus monkeys. J. Comp. Neurol198t121-136.
Price, J.L., and D.G. Amaral(1981) An autoradiographic study of the projec- tions of the central nucleus of the monkey amygdala. J. Neurosci. lt1242-1259.
Price, J.L., and B.M. Slotnick (1983) Dual olfactory representation in the rat thalamus: An anatomical and electrophysiological study. J. Comp. Neurol. 215:63-77.
Reep, R.L., and S.S. Winans (1982) Afferent connections of dorsal and ventral agranular insular cortex in the hamster, Mesocricetus auratus. Neuroscience 7:1265-1288.
Rose, J.E., and C.N. Woolsey (1948) The orbitofrontal cortex and its connec- tions with the mediodorsal nucleus in rabbit, sheep, and cat. Assoc. Res. Nerv. Ment. Dis. 27:210-232.
Rosenkilde, C.E. (1979) Functional heterogeneity of the prefrontal cortex in the monkey: A review. Behav. Neural Biol. 25:301-345.
Sarter, M., and H.J. Markowitsch (1983) Convergence of basolateral amyg- daloid and mediodorsal thalamic projections in different areas of the frontal cortex in the rat. Brain Res. Bull. 10:607-622.
Sarter, M., and H.J. Markowitsch (1984) Collateral innervation of the me- dial and lateral prefrontal cortex by amygdaloid, thalamic, and brain- stem neurons. J. Comp. Neurol. 224t445-460.
Siegel, A,, T. Fukushima, R. Meibach, L. Burke, H. Edinger, and S. Weiner (1977) The origin of the afferent supply to the mediodorsal thalamic nucleus: Enhancement of HRP transport by selective lesions. Brain Res. 13511-23.
Sripanidkulchai, K., B. Sripanidkulchai, and J.M. Wyss (1984) The cortical projection of the basolateral amygdaloid nucleus in the r a t A retrograde fluorescent dye study. J. Comp. Neurol. 229:419-431.
Tombol, T. and A. Szafranska-Kosmal(1972) A Golgi study of the amygda- loid complex in the cat. Acta Neurobiol. Exp. (Warsz) 32t835-848.
Turner, B.H., and J. Zimmer (1984) The architecture and some of the interconnections of the rats amygdala and lateral periallocortex. J. Comp. Neurol. 227t540-557.
Velayos, J.L., and F. Reinoso-Suarez (1985) Proscencephalic afferent5 to the mediodorsal thalamic nucleus. J. Comp. Neurol. 242:161-181.