nuclear distribution of pcna during embryonic development in
TRANSCRIPT

Journal of Cell Science 102, 63-69 (1992)Printed in Great Britain © The Company of Biologists Limited 1992
63
Nuclear distribution of PCNA during embryonic development in Xenopus
laevis: a reinvestigation of early cell cycles
MICHEL LEIBOVICI1*, GUILLAUME MONOD1, JACQUELINE GERAUDIE1, RODRIGO BRAVO2
and MARCEL MECHALI1
'CNRS - Institut Jacques Monod, Laboratoire d'Embryologie Moliculaire, 2, place Jussieu - Tour 43, 75251 Paris Cedex 05, France2Squibb Institute for Medical Research, Department of Molecular Biology, PO Box 4000, Princeton, NJ 08543, USA
•Present address: Institut d'Embryologie Cellulaire et Moliculaire, 49 bis, avenue de la Belle Gabrielle, 94736 Nogent sur Marne Cedex,France
Summary
The immunocytological distribution of the proliferatingcell nuclear antigen (PCNA), a protein involved in DNAreplication, has been examined during the early develop-ment of Xenopus laevis. The protein is uniformlydetected in nuclei during early stages up to the neurulastage. PCNA is detected by its distinctive cyclical patternduring early development, remaining detectable onlyduring the period of S phase of each cell cycle.Immunological detection of PCNA is therefore a usefuland specific non-isotopic marker of S-phase cells in theembryo. PCNA associates with typical karyomeric
structures, suggesting that DNA replication starts beforethe nuclear compartment is entirely formed. At themidblastula transition, a new pattern of PCNA stainingbecomes apparent. First, a new type of PCNA staining isdetected at the nuclear periphery. Second, mitoticclusters with different PCNA distributions suggest thatthe onset of desynchronization of the cell cycle at thisstage is not random.
Key words: cell cycle, PCNA, Xenopus, nucleus,development.
Introduction
The gene coding for the proliferating cell nuclearantigen (PCNA) has been characterized in a wide rangeof species from higher plants to man (Nakane et al.,1989). The product identified in vertebrate cells is anuclear protein with an apparent relative molecularmass of 36X103. Because of the specific nuclearlocalization and the increase in the synthesis of PCNAduring the late Gi and S phases of the cell cycle, a rolein DNA replication was suggested for this protein(Bravo and Macdonald-Bravo, 1984). More recently, arequirement for PCNA in the replication of SV40 DNAin vitro in human cell-free extracts has been demon-strated and emphasizes the role of PCNA during DNAreplication (Bravo et al., 1987; Prelich et al., 1987a,b;Prelich and Stillman, 1988). The PCNA may cooperatewith other replication factors to increase the processi-vity of DNA polymerase 6 during elongation of theleading strand (for review, see Stillman, 1989).
Previous immunofluorescence studies performed onmammalian cultured cells and with cell-free extracts ofXenopus eggs showed that PCNA appears at the G]/Stransition and during S phase (Celis and Celis, 1985;Hutchison and Kill, 1989). However, in some specific
conditions, PCNA protein has been detected in thenucleoplasm of quiescent cells (Bravo and Macdonald-Bravo, 1987; Leibovici et al., 1990). Furthermore,biochemical studies of the expression of the XenopusPCNA gene during development showed that stage VIoocytes contained a high level of PCNA protein, whichremained constant until the swimming tadpole stage(Leibovici et al., 1990).
The early development of Xenopus laevis has manyadvantages for studying the cell cycle, including naturalsynchrony for up to 12 cycles and the possibility ofobserving three kinds of cell cycles. The first cell cyclefollowing fertilization encompasses the classical Ga, S,G2 and M phases. Then, during segmentation, 11synchronous and biphasic cell cycles including only Sand M phases occur every 30 min. During this period,the embryos are transcriptionally quiescent. Finally, atthe 12th cell cycle, new zygotic gene expression occursand Gj and G2 phases are again present (Newport andKirschner, 1982; Kimelman et al., 1987). We haveanalysed the localization of the PCNA protein as amarker for DNA replication during the dynamicchanges associated with this developmental period. Theanalysis employed the whole-mount technique adaptedfrom Dent et al. (1989), and permitted screening of a

64 M. Leibovici and others
large number of synchronized embryos. The obser-vations were verified by classical histological techniquesusing conventional or confocal microscopy. We showthat the detection of PCNA in embryonic cells isassociated with the period of the cell cycle dedicated toDNA synthesis. This association reveals distinguishablesubnuclear structures associated with this processduring early development, and validates the use ofPCNA as a general suitable non-isotopic marker forcells engaged in the process of DNA replication.
Materials and methods
EmbryosAll experiments were done with albino Xenopus laevis. Invitro fertilization was carried out according to Gurdon (1967).
For the kinetics experiments, five batches of about 50fertilized eggs were prepared. We observed that, with thisprotocol, synchrony during successive cleavages was betterthan if we fertilized 250 eggs in a large Petri dish. Allexperiments were conducted at 21°C, and the embryos werestaged according to Nieuwkoop and Faber (1956).
Whole-mount immunocytochemistryWe essentially followed the protocol described by Dent et al.(1989). All experiments were repeated at least twice with twodifferent fixative reagents. The first fixative was Dent'sfixative (20% dimethyl sulphoxide:80% methanol), in whichdividing embryos were immersed for 60 min, at 4°C. Thesecond one was MEMFA (0.1 M MOPS pH 7.4, 2 raMEGTA, 1 mM MgSO4,3.7% formaldehyde) used according toBrivanlou and Harland (1989). Final detection was performedwith diaminobenzidine (DAB) as described by Dent et al.(1989), and the signal was optimal after 10 to 15 min ofincubation at room temperature. At the end of the exper-iment, dehydrated embryos were cleared using Murray'sclearing solution (1: 2 (v/v), benzyl alcohol/benzyl benzoate).
To improve the topographical location of PCNA relative toDNA, sections (7 /jm) of embryos embedded in paraffin wereobtained according to routine histological procedures, andmounted on either poly-L-lysine-coated slides as described byHourdryetal. (1988) or 0.1% gelatin. DNA staining was donewith propidium iodide (PDI) at a final concentration of 0.5^g/ml or with 4,6-diamino-2-phenylindole (DAPI) at 10 f/g/mlapplied for 5 min at room temperature on the dewaxed slides.Slides were washed in phosphate-buffered saline (PBS) 5 to 6times, 5 min each. They were then mounted in Citifluor(Citifluor LTD, London).
AntibodiesWe used three different rabbit polyclonal antibodies raisedagainst (1) the human PCNA entire protein (a^PCNA), (2)the human PCNA C-terminal domain (oHLKY), or (3) thehuman PCNA N-terminal domain (o^GSIL). Their reactivi-ties against the Xenopus PCNA protein have been described(Leibovici et al., 1990).
For whole-mount immunocytochemistry, ar-PCNA and a-GSIL were diluted 1:1000, and O--HLKY was diluted 1:500.Non-immune rabbit serum was used as a negative control atthe appropriate dilution (e.g. 1:1000 or 1:500).
Results
Detection and localization of PCNA duringembryonic developmentOur previous results indicated that PCNA protein waspresent at a steady-state level from the egg to theswimming tadpole in Xenopus embryos (Leibovici etal., 1990). Indeed, western blot analysis did not showany decrease in the PCNA content after the midblastulatransition, at least up to the swimming tadpole stage.We analysed whether this constitutive level reflecteduniform or heterogeneous distribution of PCNA amongthe different tissues of the embryo.
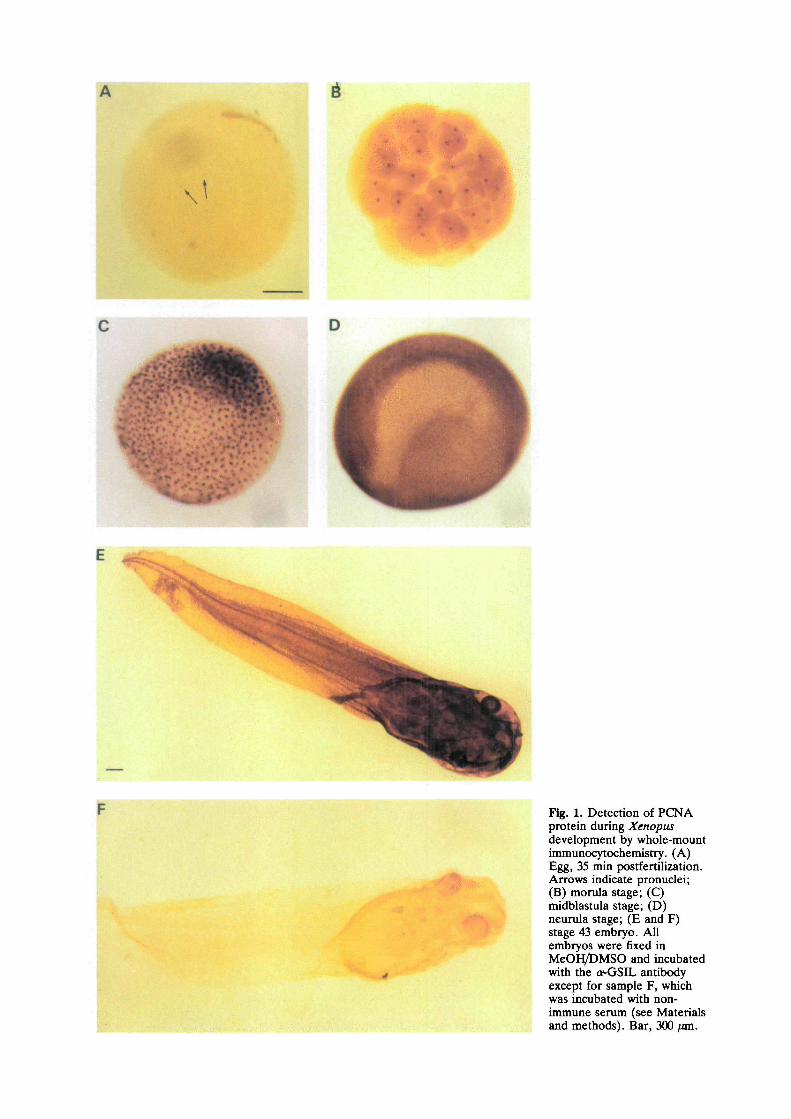
The localization of PCNA in the developing embryowas performed by in toto experiments on varioussamples of embryos fixed at different stages of develop-ment. Immunolocalizations were repeated with threeindependent sera raised against three distinct parts ofthe PCNA protein (see Materials and methods). Fig. 1shows that from the egg (Fig. 1A) to the swimmingtadpole (Fig. IE), PCNA staining was always detectedin the nuclei of embryonic cells. The maternal pool ofPCNA is entirely localized in the nucleus of the matureoocyte, when analysed by both biochemical proceduresand in histological sections of oocytes (Zuber et al.,1989; Leibovici et al., 1990), as well as by the whole-mount technique (not shown). The first appearance of aPCNA signal occurs in pronuclei of fertilized eggs at 35min after fertilization (Fig. 1A).
At morula and blastula stages (Fig. 1 B and C,respectively) the staining remained exclusively in thenuclei. At the blastula stage, we repeatedly observedthat numerous nuclei showed annular staining. Thispeculiar staining was further investigated using confocalmicroscopy (see below). Although variations of theexperimental conditions might explain differences inthe staining intensity, Fig. 1 clearly shows that thePCNA signal globally increases during development, inproportion to the number of nuclei (2, 32, 4000 andabout 20,000 in Fig. 1A, B, C, D respectively). As thereis no variation in the amount of PCNA estimated bywestern blot analysis (Leibovici et al., 1990), thisobservation probably reflects an unmasking of PCNAthat is correlated with the formation of embryonicnuclei. The reason for such a structural modification, orsubcellular association, accounting for this phenom-enon is unknown. However, a variation in PCNAdetection was also observed within each cell cycle, asshown below.
At the neurula stage (Fig. ID), PCNA is still evenlydetected throughout the ectodermic cells of the em-bryo. However, histological analyses revealed differen-tial labeling of internal tissues of the embryo. Ectoder-mal cells were all PCNA positive, but staining wasscarcely observed in the neural tube, somites andendoderm, and not detected in the notochord (unpub-lished observations). At the tadpole stage, nuclearstaining was also observed (Fig. IE). In addition, strongdetection was reproducibly obtained in nervous tractsand in the cytoplasm of the ciliated cells (not shown).This unexpected localization was also obtained with the
\
Fig. 1. Detection of PCNAprotein during Xenopusdevelopment by whole-mountimmunocytochemistry. (A)Egg, 35 min postfertUization.Arrows indicate pronuclei;(B) momla stage; (C)midblastula stage; (D)neurula stage; (E and F)stage 43 embryo. Allembryos were fixed inMeOH/DMSO and incubatedwith the a^GSIL antibodyexcept for sample F, whichwas incubated with non-immune serum (see Materialsand methods). Bar, 300 ftm.

PCNA
Fig. 2. Changes in PCNA detection during the progression of the early development cell cycle. Embryos were fixed inMeOH/DMSO and incubated with a^GSIL to detect PCNA (right column). Sections were incubated with PDI to detectDNA; the left and middle columns show PCNA and DNA labeling, respectively, on histological cut sections (thebackground observed in the cytoplasm of PDI-stained embryos is essentially due to the autofluorescence of the vitellus.This is illustrated in M, a micrograph taken under a mixture of daylight and UV light). Rows 1, 2, 3, 4 and 5 are samplestaken at 0 min, 26 min, 30 min, 34 min and 40 min during the course of the experiment. In L, arrows indicate karyomericstructures. S, S phase; M, M phase. C, F, I, L and O are x27; A and B, x50; D and E, x200; and G, H, J, K, M and N,X127.
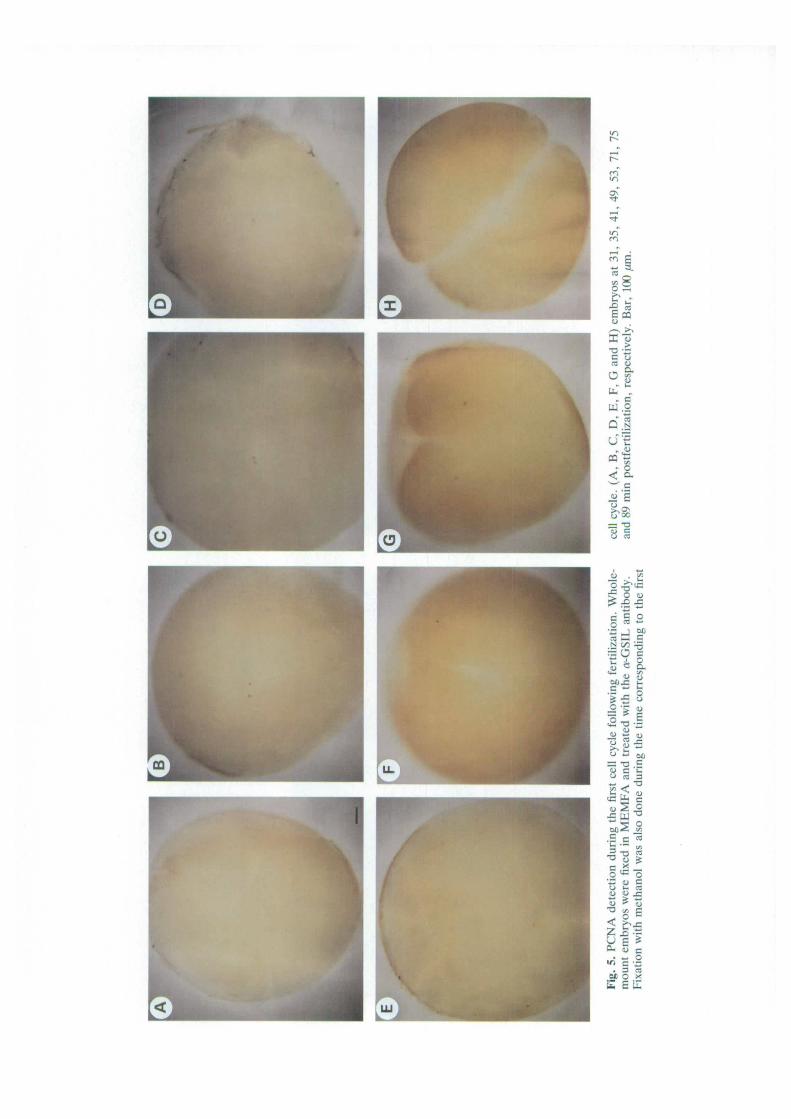

Fig. 7. Nuclearheterogeneous staining ofPCNA in a midblastula stageembryo. The whole-mountembryo was fixed inMeOI^/DMSO and incubatedwith the o^GSIL antibody(x60). Note that some areasdo not show any labelingwhile others exhibit annularor homogeneous nuclearstaining.
£ i< •
^ in M
Fig. 8. PCNA localization atthe midblastula transition.Embryos were fixed inMeOH/DMSO and incubatedwith a-GSIL. (A, B)Confocal micrographs of twodifferent fields of the samewhole-mount embryo (x450);(C, D) a histological sectiondouble-stained with DABand DAPI, respectively, todetect PCNA and DNA (asfor the PDI staining, weobserved background in thecytoplasm of D API-stainedembryos). The arrow in Dshows the condensedchromatin of a blastomerethat is not stained by anti-PCNA (C).

Distribution of PCNA during development 65
two other PCNA antibodies; no signal was detectedwhen the embryos were incubated with a non-immuneserum (Fig. IF). Moreover, these results were observedwhen two different fixatives were used (methanol orformaldehyde). Although this pattern of staining wasnot examined in detail in this study, it is likely to be dueeither to the presence of PCNA in the cytoplasm of thepositive cells, or to an antigenic domain of a proteinimmunologically related to PCNA.
In summary, during segmentation, the detectabilityof the PCNA protein is restricted to nuclei that areuniformly stained with anti-PCNA up to the mid-blastula stage. The detection of PCNA in the nervoussystem and in ciliated cells of late-stage embryos has notbeen explained. During the course of this study, we alsoobserved that some samples within the same batch ofembryos treated in similar conditions did not show anyPCNA staining. We analysed this phenomenon in moredetail by close examination of the cell cycle during earlydevelopment, when cell cycles are synchronous.
Kinetic analysis of PCNA staining during the cellcycle at the segmentation stageThe behavior of PCNA during the cell cycle wasanalysed by fixation of several embryos at differenttimes, followed by treatment in identical conditions ofantibody dilution, washes and time of detection. Wechose to follow the transition from 8-cell stage to 32-cellstage embryos because the cleavage synchrony isgradually lost (Nieuwkoop and Faber, 1956; Satoh,1977). Results were reproducible using several batchesof embryos. Batches of 5 embryos were collected every2 min from the 8-cell stage through the 32-cell stage,and fixed in methanol/dimethylsulfoxide (MeOH/DMSO). Embryos were then incubated with anti-PCNA and PCNA was detected by the DAB-peroxi-dase procedure (Materials and methods). Fig. 2 showsrepresentative results from embryos taken over a 40min period, during progression from the 16- to the 32-cell stage. The detection of PCNA in whole-mountembryos is shown in Fig. 2, right column. Because wecould not detect DNA in whole-mount embryos,histological sections of the same embryos were stainedwith PDI as in Materials and methods (Fig. 2, middlecolumn). Thus, histological sections from the sameembryos analysed for PCNA in toto permitted thesimultaneous in situ detection of DNA and PCNA (Fig.2, middle and left columns).
Our observations indicate that staining of PCNA isrestricted to S phase. Row 1 of Fig. 2 shows an embryoat the 16-cell stage (time 0 of the kinetic study). All thenuclei are stained with anti-PCNA and histologicalsections confirmed the staining of round nuclei, closelysurrounded by cytoplasmic structures. The PDI stainingis faint, and spread out in the nuclear compartment asexpected for an S phase nucleus (compare Fig. 2B andN with E and H). Rows 2 and 3 show embryos at 26 minand at 30 min, respectively. During this period, noPCNA staining was observed either on all whole-mountembryos or on histological sections. PDI staining
B
IlkFig. 3. Detection of PCNA in karyomeric structures.(A) Whole-mount embryo 64- to 128-cell stage fixed inMeOI^/DMSO and incubated with o^GSIL antibody (x60);(B) a squash from the embryo presented in A (x560). Fivekaryomeres are clearly visible; (C) cut section of a 1024-cell embryo fixed in MEMFA and incubated with the a-GSIL antibody. Final detection with DAB was amplifiedwith a DAB-silver enhancement kit (Amersham). Eachblastomere encloses a cluster of karyomeres (x250).
indicated that this period corresponds to the M phase ofthe cell cycle. Indeed, Fig. 2E shows a typicalmetaphase plate where DNA is condensed in chromo-somes, and the nucleus presented in Fig. 2H exhibits acharacteristic punctate pattern, which may correspondto karyomeres present at the telophase step (Montag etal., 1988; and Fig. 3). Karyomeric structures aretransient individual chromosomal units surrounded by anuclear envelope, which progressively fuse finally toreconstitute a single nucleus. Row 4 shows a typicalembryo at 32-34 min; weak PCNA staining reappears inthe whole-mount embryo (Fig. 2L). Sections show thatthe nucleus is irregular when stained with DAB as wellas with PDI (Fig. 2J and K). The wide clear spacebetween the nucleus and the cytoplasmic structure (Fig.2J) indicates that mitosis has not been completed. Theirregular nuclear structures observed both in whole-mount embryos and on sections are most likely largefused karyomeres, at the end of telophase (Montag etal., 1988). PDI staining becomes faint again, indicatingthat the chromatin has started to decondense from itschromosomal state. This phase is usually followed byDNA replication and thus reappearance of PCNAstaining in karyomeric structures is correlated with the
66 M. Leibovici and othersry
os
. 0
ECJ
T31)
S tan
Oa?
100
80
60
40
20
0
/
//
1
1 • • ' ! — " — • -
R
\ f\\ j\ 1\ /
L 1
\ f\ /Av
10 20 30
Time (min)
40 50
Fig. 4. Periodic staining of PCNA during earlydevelopment. Synchronous dividing embryos were fixedevery 2 min in MeOH/DMSO (• • ) and then treatedwith Q--GSIL antibody as described in Material andmethods. Arrows indicate the appearance of the cleavagefurrow.
onset of the new S phase. This observation is inagreement with PCNA studies in in vitro cultured cells(Celis and Celis, 1985) as well as in Xenopus eggextracts (Hutchison and Kill, 1989). Forty minuteslater, the 32-cell stage is reached (Fig. 2, row 5), and theS phase of the next cell cycle is progressing, with allnuclei positive for PCNA staining.
The punctate PCNA pattern observed at the end ofmitosis was analysed in more detail, as shown in Fig. 3.When embryos taken at a stage between 64 and 128 cellsare stained by the anti-PCNA antibody, most of thenuclei have an irregular shape in a phase that lasts a fewminutes during the cell cycle (Fig. 3A). A squash of thisembryo permitted observation at higher magnification(Fig. 3B). We observed typical karyomeric structures,as previously shown by Montag et al (1988). Fivekaryomeres were stained by anti-PCNA. Fig. 3C showsanother observation of karyomeres done on a histologi-cal section of a 1024-cell stage embryo fixed inMEMFA. These peculiar chromatin structures appearsynchronously in the embryo and are strongly PCNApositive.
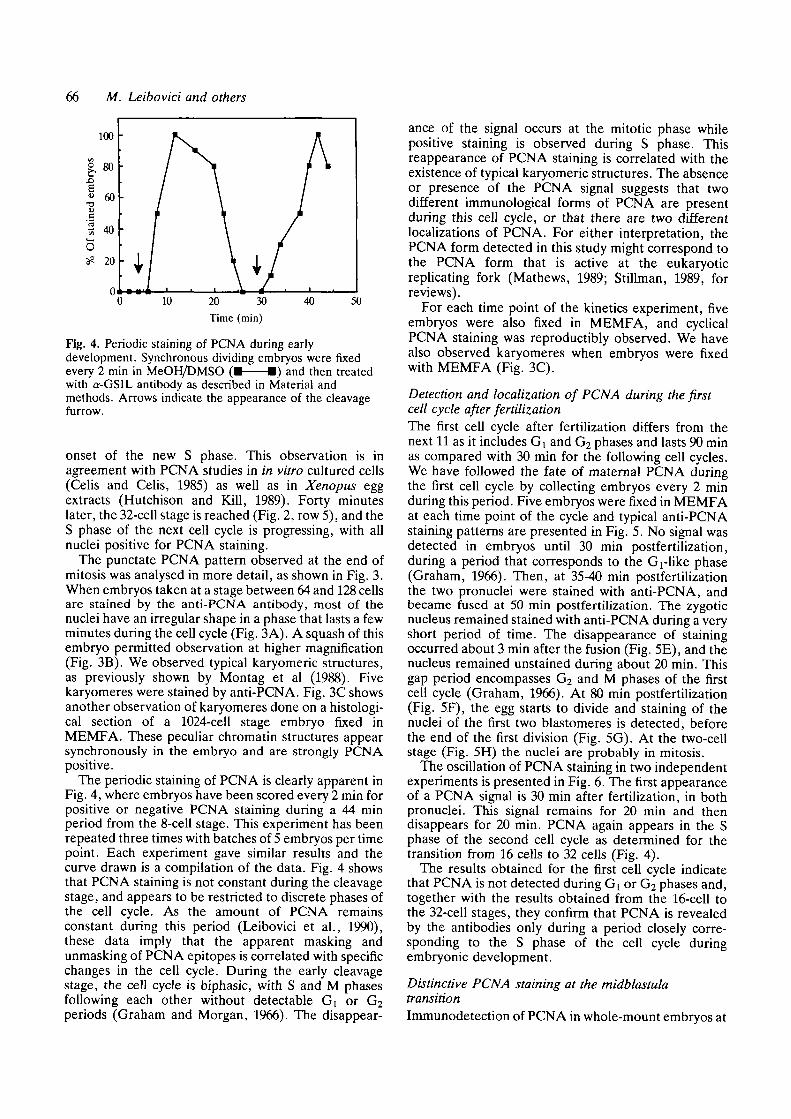
The periodic staining of PCNA is clearly apparent inFig. 4, where embryos have been scored every 2 min forpositive or negative PCNA staining during a 44 minperiod from the 8-cell stage. This experiment has beenrepeated three times with batches of 5 embryos per timepoint. Each experiment gave similar results and thecurve drawn is a compilation of the data. Fig. 4 showsthat PCNA staining is not constant during the cleavagestage, and appears to be restricted to discrete phases ofthe cell cycle. As the amount of PCNA remainsconstant during this period (Leibovici et al., 1990),these data imply that the apparent masking andunmasking of PCNA epitopes is correlated with specificchanges in the cell cycle. During the early cleavagestage, the cell cycle is biphasic, with S and M phasesfollowing each other without detectable Gx or G2periods (Graham and Morgan, 1966). The disappear-
ance of the signal occurs at the mitotic phase whilepositive staining is observed during S phase. Thisreappearance of PCNA staining is correlated with theexistence of typical karyomeric structures. The absenceor presence of the PCNA signal suggests that twodifferent immunological forms of PCNA are presentduring this cell cycle, or that there are two differentlocalizations of PCNA. For either interpretation, thePCNA form detected in this study might correspond tothe PCNA form that is active at the eukaryoticreplicating fork (Mathews, 1989; Stillman, 1989, forreviews).
For each time point of the kinetics experiment, fiveembryos were also fixed in MEMFA, and cyclicalPCNA staining was reproductibly observed. We havealso observed karyomeres when embryos were fixedwith MEMFA (Fig. 3C).
Detection and localization of PCNA during the firstcell cycle after fertilizationThe first cell cycle after fertilization differs from thenext 11 as it includes Gi and G2 phases and lasts 90 minas compared with 30 min for the following cell cycles.We have followed the fate of maternal PCNA duringthe first cell cycle by collecting embryos every 2 minduring this period. Five embryos were fixed in MEMFAat each time point of the cycle and typical anti-PCNAstaining patterns are presented in Fig. 5. No signal wasdetected in embryos until 30 min postfertilization,during a period that corresponds to the Grlike phase(Graham, 1966). Then, at 35-40 min postfertilizationthe two pronuclei were stained with anti-PCNA, andbecame fused at 50 min postfertilization. The zygoticnucleus remained stained with anti-PCNA during a veryshort period of time. The disappearance of stainingoccurred about 3 min after the fusion (Fig. 5E), and thenucleus remained unstained during about 20 min. Thisgap period encompasses G2 and M phases of the firstcell cycle (Graham, 1966). At 80 min postfertilization(Fig. 5F), the egg starts to divide and staining of thenuclei of the first two blastomeres is detected, beforethe end of the first division (Fig. 5G). At the two-cellstage (Fig. 5H) the nuclei are probably in mitosis.
The oscillation of PCNA staining in two independentexperiments is presented in Fig. 6. The first appearanceof a PCNA signal is 30 min after fertilization, in bothpronuclei. This signal remains for 20 min and thendisappears for 20 min. PCNA again appears in the Sphase of the second cell cycle as determined for thetransition from 16 cells to 32 cells (Fig. 4).
The results obtained for the first cell cycle indicatethat PCNA is not detected during Gt or G2 phases and,together with the results obtained from the 16-cell tothe 32-cell stages, they confirm that PCNA is revealedby the antibodies only during a period closely corre-sponding to the S phase of the cell cycle duringembryonic development.
Distinctive PCNA staining at the midblastulatransitionImmunodetection of PCNA in whole-mount embryos at

Distribution of PCNA during development 67
20 30 40 50 60 70
Time after fertilization (min)
80 90 100
Fig. 6. PCNA labeling oscillations following fertilization.Whole-mount embryos fixed every 2 min in MEMFA(D • ) were incubated with the a^GSIL antibody.Fixation with MeOH/DMSO ( • • ) was also doneduring the time corresponding to the first cell cycle. Thecurve represents data from 2 experiments. The arrowindicates the time of pronuclei fusion.
the MBT revealed a heterogeneous pattern of stainingwithin discrete groups of nuclei of the same embryo, forboth fixatives used (Fig. 7). When different fields of thesame whole-mount embryo were observed with aconfocal microscope, three typical patterns of stainingwith anti-PCNA were detected: groups of cells withhomogeneous nuclear staining (Fig. 8A), or staining ofthe nuclear periphery (Fig. 8B) or absence of staining(not illustrated). Thus, at the midblastula stage,analysis of PCNA reveals two additional distinctivefeatures. First, patches of cells with different PCNAdistributions are observed within the same embryo.These data indicate that the desynchronization processthat occurs at the midblastula stage might not be astochastic event. Second, a new type of nuclear stainingis detected for this protein. During the early cleavagestage, homogeneous PCNA staining was detectedduring S phase whereas the absence of stainingcorresponded to Gi, G2 or M phases. Nuclear ring-shaped staining was never observed before the mid-blastula transition and might represent specific intranu-clear modifications, which occur at this stage.
Fig. 8C shows a cut section that confirms the annularnuclear staining of PCNA in groups of cells that can bedistinguished from those with homogeneous nuclearstaining. We have performed serial histological sectionsof some of these midblastula samples, and found thatPCNA staining was over the entire nucleus. Inside thering formed by DAB staining, the DNA appearshomogeneous by DAPI staining (Fig. 8C and D),although in other examinations the pattern of DAPIstaining was superimposed on the ring formed byPCNA staining. Thus the pattern of PCNA stainingdoes not correlate exactly with DNA staining at themidblastula stage. These observations reveal a subnuc-lear localization of PCNA, which might reflect dynamicnuclear changes at this stage.
Discussion
The biochemical studies of the expression of theXenopus PCNA gene during development showed thatstage VI oocytes contain a high level of PCNAtranscripts and protein (Leibovici et al., 1990). Thelarge amount of PCNA protein, 65 ng per single matureoocyte (Zuber et al., 1989), remained constant duringboth early and late development until the swimmingtadpole stage (Leibovici et al., 1990), as well as duringthe cleavages induced by activation (Zuber et al., 1989).However, in this study, we have shown that thedistribution of PCNA protein in the nuclei and itsappearance as a function of the cell cycle reveal majorchanges in its behavior during the early phases ofembryonic development.
The cyclical pattern of PCNA staining during earlydevelopmentIn contrast to the heterogeneous distribution of PCNAobserved in late stages of development, all early stageembryos examined showed homogeneous staining withanti-PCNA. This staining, exclusively nuclear, firstappears in eggs 35 min after fertilization. The kineticanalysis of the distribution of PCNA during the simplebiphasic cell cycles revealed a strong correlationbetween PCNA staining and the progression of the cellcycle. PCNA is exclusively detected during a periodcorresponding to the S phase of the cell cycle asdescribed in mammalian somatic cells (Celis and Celis,1985; Bravo and Macdonald-Bravo, 1985). As PCNAprotein content does not change during early develop-ment, we conclude that PCNA can assume at least twodifferent forms: one of them detectable during S phase,and the other remaining undetectable by three differentantibodies during M phase. An alternative explanationfor the absence of signal during M phase is that PCNAis diluted in the cytoplasm at a concentration below thelevel of detection.
PCNA labeling is absent during Gi and G2 phasesduring the first cell cycle. These in vivo results alsoagree with data obtained with in vitro Xenopus eggextracts that show that nuclei blocked in a G2-like stateby cycloheximide lose PCNA staining (Hutchison andKill, 1989). Immunoblots performed on isolated G2-likenuclei also showed that the antigen was not lost(Hutchison and Kill, 1989). Thus, the absence ofstaining observed in vivo during Gi and G2 phases couldbe due to a modification of the nuclear protein ratherthan its exclusion from the nuclear compartment.Analysis of the first cell cycle with anti-PCNA alsoshowed that the S phase of the second cell cycle startsbefore the first cleavage of the egg is accomplished, aspreviously reported by Graham (1966). During therapid cleavage stage, we also found PCNA in the largefused karyomeres that result from the fusion ofindividual chromosomal units during telophase (Mon-tag et al., 1988). The presence of PCNA in suchstructures might indicate that DNA replication starts atthis step of the cell cycle. This in vivo observationsuggests that karyomeres exhibit functions of the future

68 M. Leibovici and others
nucleus, particularly the ability to replicate DNA.However, the presence of PCNA in karyomericstructures does not necessarily imply that they arereplicating DNA at that time. A more accuratecorrelation between replication and PCNA stainingmay be provided by performing short pulse-labelingexperiments in vivo, although injection by biotinylateddNTPs into single cells at the segmentation stage mightnot be easy to achieve. It has been shown that theformation of a complete nuclear envelope is essential inorder to trigger the initiation of DNA replication(Newport et al., 1990). The formation of karyomeresmight permit DNA replication to start before theformation of the entire nucleus and suggests thatkaryomeres are adapted to rapid embryonic nucleardivision cycles, as first suggested by Montag et al.(1988).
This study also shows that the PCNA protein is auseful marker of cells engaged in S phase and could bean alternative to thymidine labeling. PCNA is a subunitof DNA polymerase 6, which is implicated in chromo-somal replication (Bravo et al., 1987; Prelich et al.,1987a,b; reviewed by Thommes and Hiibscher, 1990).Its detection during S phase suggests that it is thefunctional form of PCNA that was identified. Duringthe other phases of the cell cycle the protein was notdetected, although present. Two explanations can begiven for such observations: either the active form ofPCNA has a structural form different from the inactiveprotein, or PCNA is sequestered in an unaccessibleform when the cell is not engaged in DNA replication.We favor the second explanation because three differ-ent antibodies failed to detect PCNA during M phase.In any case, these data emphasize that PCNA is asuitable S-phase marker but should not be taken to be acell division marker.
Distribution of PCNA at the MBTThe midblastula transition (MBT) is marked by a seriesof structural and functional events, which signal theswitch from the early embryonic cell cycle to a somatictype of cell cycle (Signoret and Lefresne, 1971; Newportand Kirschner, 1982). The cell cycle includes G\ and G2phases, nuclear organization is modified, lamins II and Iare integrated into the lamina structure (Benavente etal., 1985), the nuclear volume decreases and theirregular shape of the nucleus, typical of embryonicnuclei, disappears (Montag et al., 1988). At the sametime, the rate of DNA synthesis decreases, the cell cycleof individual blastomeres become desynchronized andzygotic transcription is turned on. At the MBT, a newPCNA-staining pattern is detected, with nuclei stainedperipherally. This pattern coexists in the same embryoswith cells with nuclei exhibiting homogeneous PCNAstaining and unstained cells. These three different celltypes are found in groups of up to ten cells, each groupsharing a given type of staining. These clusters appearto be randomly distributed among stage 12 embryos andprobably reflect the asynchrony of the cell cycles, whichis first observed at that stage (Gerhart, 1980). Such
patterns might be in a sequence of events linked to thecell cycle seen at the MBT, and the observation ofmitotic clusters suggests that the asynchrony is notrandomly set in the Xenopus embryo. However, we donot know if they directly reflect commitment of celldomains to specific developmental fates. In Drosophila,at the time of cellular asynchrony (cycle 14), mitoticdomains of cell clusters united by locally synchronousmitosis partition the early embryo (Foe, 1989; Edgarand O'Farrell, 1989). This pattern was interpreted toreveal an early commitment of cells to specific develop-mental fates. In Xenopus the midblastula transition isalso characterized by the onset of new zygotic ex-pression. Whether or not the onset of transcription is acause or consequence of the lengthening of the cellcycle is not yet understood. However, the asynchronyby groups of cells, as now revealed by PCNA staining,could indicate some features in common with thoseobserved in Drosophila.
The midblastula transition is also characterized by adistinct PCNA staining pattern in some groups of cells.In histological sections, DNA was either homo-geneously distributed inside the nucleus (see Fig. 8B) orsuperimposed on the ring formed by the PCNAstaining. A recent study of Hutchison and Kill (1989)indicated that nuclei incubated in Xenopus cell-freeextracts in the presence of aphidicolin (an inhibitor ofDNA polymerase a and 6) showed similar PCNA andDNA polymerase a staining patterns. These authorsconcluded that such a staining pattern might reflect thepresence of PCNA in inactive replication complexes.We propose the hypothesis that our in vivo resultsillustrate the physiological appearance of these com-plexes, which might be anchored at peripheral sites innuclei. The presence of these structures might reflectthe transition from active proliferation to a condition inwhich replication activity is reduced. PCNA annularstaining has also been observed in cultured cells whencellular proliferation decreases (S. Vriz, unpublisheddata). The coexistence of three types of staining at themidblastula stage could mirror the asynchrony in thecell cycle that starts at this step of development. Thestructural nuclear reorganization that occurs at theMBT might involve a transit of nuclear antigens fromthe nucleoplasm toward the nuclear envelope.
PCNA detection patterns during early developmentstrongly suggest dynamic changes associated with thisprotein, probably in relation to nuclear metabolism. Itwill be worthwhile determining if the structures re-vealed by PCNA staining represent discrete subnuclearstructures involved in the organization of the genomefor its replication.
We thank S. Vriz for helpful and stimulating discussionsand B. Maro for his help with confocal microscopy. We alsothank M. Gusse for his unpublished results (Fig. 3C) in thisarticle. Thanks are also due to P. Brooks for critical reading ofthis manuscript and M. Malet for typing it. This work hasbeen supported by the Association pour la Recherche sur leCancer, INSERM, Ligue Francaise contre le Cancer and theFondation de la Recherche M6dicale.

Distribution of PCNA during development 69
References
Benavente, R., Krohne, G. and Franke, W. W. (1985). Cell type-specific expression of nuclear lamina proteins during developmentof Xenopus laevis. Cell 41, 177-190.
Bravo, R., Frank, R., Blundell, P. H. and Macdonald-Bravo, H.(1987). Cyclin/PCNA is the auxiliary protein of DNA polymerase-6. Nature 326, 515-517.
Bravo, R. and Macdonald-Bravo, H. (1984). Induction of the nuclearprotein 'cyclin' in quiescent mouse 3T3 cells stimulated by serumand growth factors. Correlation with DNA synthesis. EMBO J. 3,3177-3181.
Bravo, R. and Macdonald-Bravo, H. (1985). Changes in the nucleardistribution of cyclin (PCNA) but not its synthesis depend on DNAreplication. EMBO J. 4, 655-661.
Bravo, R. and Macdonald-Bravo, H. (1987). Existence of twopopulations of Cyclin/Proliferating Cell Nuclear Antigen duringthe cell cycle: association with DNA replication sites. J. Cell Biol.105, 1549-1554.
Brivanlou, A. H. and Harland, R. M. (1989). Expression of anengra/7-related protein is induced in the anterior neural ectoderm ofearly Xenopus embryos. Development 106, 611-617.
Cells, J. E. and Cells, A. (1985). Cell cycle-dependent variations inthe distribution of the nuclear protein cyclin proliferating cellnuclear antigen in cultured cells: Subdivision of S phase. Proc. Nat.Acad. Sci. U.S.A. 82, 3262-3266.
Dent, J. A., Poison, A. G. and Klymkowsky, M. W. (1989). A whole-mount immunocytochemical analysis of the expression of theintermediate filament protein vimentin in Xenopus. Development105, 61-74.
Edgar, B. A. and O'FarreU, P. H. (1989). Genetic control of celldivision patterns in the Drosophila embryo. Cell 57, 177-187.
Foe, V. E. (1989). Mitotic domains reveal early commitment of cellsin Drosophila embryos. Development 107, 1-22.
Gerhart, J. (1980). Mechanisms regulating pattern formation in theamphibian egg and early embryo. In Biological Regulation andDevelopment, vol. 2 (ed. R.F. Goldberger), pp. 133-315. NewYork: Academic Press.
Graham, C. F. (1966). The regulation of DNA synthesis and mitosisin multinucleate frog eggs. J. Cell Sci. 1, 363-374.
Graham, C. F. and Morgan, R. W. (1966). Changes in the cell cycleduring early amphibian development. Develop. Biol. 14, 439-460.
Gurdon, J. B. (1967). In Methods in Developmental Biology (ed. F.H.Wilt and N.K.Wessels), pp.75-84. New York: Crowell.
Hourdry, J., Brulfert, A., Gusse, M., Schoevaert, D., Taylor, M. V.and Mechali, M. (1988). Localization of c-myc expression duringoogenesis and embryonic development in Xenopus laevis.Development 104, 631-641.
Hutchison, C. J. and Kill, I. (1989). Changes in the nucleardistribution of DNA polymerase alpha and PCNA/cyclin duringthe progress of the cell cycle, in a cell-free extract of Xenopus eggs.J. Cell Sci. 93, 605-613.
Kimelman, D., Klrschner, M. and Scherson, T. (1987). The events of
the midblastula transition in Xenopus are regulated by changes inthe cell cycle. Cell 48, 399-407.
Leibovlci, M., Gusse, M., Bravo, R. and Mechali, M. (1990).Characterization and developmental expression of XenopusProliferating Cell Nuclear Antigen (PCNA) Develop. Biol. 141,183-192.
Mathews, M. B. (1989). The proliferating cell nuclear antigen,PCNA, a cell growth-regulated DNA replication factor. In GrowthControl During Cell Aging (ed. E. Wang and M.R. Warner), pp.89-120. Boca Raton: CRC Press.
Montag, M., Spring, H. and Trendeknburg, M. F. (1988). Structuralanalysis of the mitotic cycle in pre-gastrula Xenopus embryos.Chromosoma 96, 187-196.
Nakane, P. K., Moriuchi, T. K., Tanlguchl, Y., Izumi, S. and Hul, L.(1989). Proliferating Cell Nuclear Antigen (PCNA/Cyclin): reviewand some new findings. Acta Histochem. Cytochem. 22, 105-116.
Newport, J. and Kirschner, M. (1982). A major developmentaltransition in early Xenopus embryos. I. Characterization andtiming of cellular changes at the midblastula stage. Ce//30, 675-686.
Newport, W. J., Wilson, K. L. and Dunphy, W. G. (1990). A lamin-independent pathway for nuclear envelope assembly. J. Cell Biol.I l l , 2247-2259.
Nieuwkoop, P. D. and Faber, J. (1956). Normal Table of Xenopuslaevis Daudin. Amsterdam: North-Holland.
Prelkh, G., Kostura, M., Marshak, D. R., Mathews, M. B. andStillman, B. (1987b). The cell-cycle regulated proliferating cellnuclear antigen is required for SV40 DNA replication in vitro.Nature 326, 471-475.
Prellch, G. and Stillnian, B. (1988). Coordinated leading and laggingstrand synthesis during SV40 DNA replication in vitro requiresPCNA. Cell 53, 117-126.
Prellch, G., Tan, C-K., Kostura, M., Mathews, M. B., So, A. G.,Downey, K. M. and Stillman, B. (1987a). Functional identity ofproliferating cell nuclear antigen and polymerase-<5 auxiliaryprotein. Nature 326, 517-520.
Sat oh, N. (1977). "Metachronous" cleavage and initiation ofgastrulation in amphibian embryos. Develop., Growth Differ. 19,111-117.
SIgnoret, J. and Lefresne, J. (1971). Contribution a l'dtude de lasegmentation de l'oeuf d'axolotl. I. Definition de la transitionblastuteenne. Ann. Embryol. Morph. 4, 113-223.
Stillman, B. (1989). Initiation of eukaryotic DNA replication in vitro.Annu. Rev. Cell. Biol. 5, 197-245.
Thommes, P. and HObscher, U. (1990). Eukaryotic DNA replication.Eur. J. Biochem. 194, 699-712.
Znber, M., Yasul, W., Tan, E. M. and Ryoji, M. (1989). Quantitationand subcellular localization of Proliferating Cell Nuclear Antigen(PCNA/Cyclin) in oocytes and eggs of Xenopus laevis. Expl CellRes. 182, 384-393.
(Received 6 January 1992 - Accepted 10 February 1992)
