monoclonal antibodies to progesterone: characterization and selection for enzyme immunoassay in...
TRANSCRIPT

HYBRIDOMAVolume 18, Number 3, 1999Mary Ann Liebert, Inc.
Monoclonal Antibodies to Progesterone: Characterization andSelection for Enzyme Immunoassay in Bovine Milk
A. WALDMANN
ABSTRACT
Thirty-one stabile murine monoclonal antibody (MAb) producing cell lines to progesterone were generatedby using a short and a long immunization protocol. Long-term immunization with high doses of 11a-hy-droxyprogesterone-hemisuccinate-bovine serum albumin (11a-OH-P-HS-BSA) antigen led to very good anti-body response in Balb/c mice. The donor mouse produced antiserum with a high titre of 1/250,000. ElevenMAbs were selected for further characterization since they showed high sensitivities ( , 35 pg/well to inhibit50% of the tracer) in bridge homologous enzyme immunoassay (EIA). The results were compared to the donormouse polyclonal antiserum. The MAbs and the donor mouse antiserum were generally found to be highlyspecific, when tested with 30 different steroids. Employing MAb 9C11, with affinity constant, Ka , to 11a-OH-P-HS of 1.1 3 1010 M 2 1, a bridge heterologous microtitre plate EIA for milk progesterone was developed, us-ing the second-antibody coating technique and horseradish peroxidase (HRP) as an enzyme label. The assayis simple and convenient to use, as it permits direct addition of undiluted milk samples, at the same time main-taining high sensitivity, high precision, and a wide range of optical density (OD) values. The major advantageof the assay developed, compared to previously published direct addition milk progesterone immunoassays,is that progesterone concentrations, measured by the EIA, were not influenced by changing milk fat concen-trations, even when milk samples containing up to 10% of milk fat were used for analysis.
289
INTRODUCTION
PRO GESTER ON E IN BLO O D AN D M ILK in cyclic cows reflect theactivity of the corpus luteum and thus provide a precise in-
dicator of ovarian function. (1) Besides the importance of pro-gesterone analysis for scientific investigations, determination ofthe hormone is considered a useful parameter for veterinary andanimal production requirements. (2,3 ) Due to its noninvasive na-ture, simplicity of sample collection, and stability of proges-terone in milk, the determination of milk progesterone has be-come a sample of choice in lactating animals.
Several enzyme immunoassays (EIAs) for milk progesteronehave been described using sheep, rabbit, and goat polyclonalantisera. (4–11) To improve affinity and obtain substantial quan-tity of antibodies, many groups have prepared mouse mono-clonal antibodies (MAbs) to progesterone. (12–21) Some of theseMAbs have been utilized as reagents in different progesteroneimmunoassays developed for human plasma,(19,22–24) horse
plasma,(25) human saliva (18) and human urine. (26) Most an-tiprogesterone antibodies derived by hybridom a technologyhave affinities far below the corresponding conventional anti-sera. This might explain the relatively small number of publi-cations reporting the use of antiprogesterone MAb in quantita-tive progesterone immunoassays , compared to the use ofpolyclonal antibodies. However, the use of MAbs should leadto improvem ents in test standardization and avoid the depen-dency on animals producing high-quality antisera.
So far, only three MAb based nonisotopic quantitative milkprogesterone immunoassay s have been reported on.(27–29) Suf-ficient sensitivity of the milk progesterone assay, developed byStanley et al.,(28) was gained by the use of an enzyme amplifi-cation system of alkaline phosphatase. The amplification pro-cedure, in turn, has made the assay more complicated and ex-pensive than the conventional EIA. Groves et al.(27) producedan ovine MAb to progesterone for a bridge heterologous mi-crotitre-plate EIA. The described assay showed high sensitiv-
University of Tartu, Institute of General and Molecular Pathology, Veski 34, 51014 Tartu, Estonia.

ity, but utilized long incubation steps and elevated temperatureto obtain higher endpoint signal. This again has made the as-say unpractical. In a previous study Waldmann (29) reported abridge homologous MAb-based milk progesterone EIA usinghorseradish peroxidase as an enzyme label.
The present study aims to characterize 11 high-affinity an-tiprogesterone MAbs for use in EIA, in terms of sensitivity andspecificity, and compare the results with the polyclonal anti-serum obtained from the same mouse. We also report on a de-velopment of a simple direct-addition bridge heterologous mi-crotitre-plate EIA of progesterone in bovine milk using the“second antibody coating technique.” This technique, first re-ported by Meyer and Güven, (30) improves precision and over-comes time dependent drift, observed in microplate im-munoassays when hormone specific antibody is directly coatedto the wells.(31)
MATERIALS AND METHODS
Reagents
Unless mentioned otherwise, all chemicals, steroids, steroidderivates for immunization and synthesis, reagents, and mediaused in cell fusion and cell culture were purchased from SigmaChemical Co. (St. Louis, MO). Tween-20 was obtained fromFerak (Berlin, Germany), Ovalbumin (OVA) from Fluka(Switzerland). The labelled and unlabelled affinity purified sec-ond antibodies were prepared at the Institute of General andMolecular Pathology, University of Tartu.
Buffers
Phosphate-buffer ed saline (PBS) 0.15 M pH 7.2–7.4 con-tained per liter 8 g of NaCl, 0.1 g KCl, 1.15 g Na2HPO4, 0.2 gKH2PO4. PBS with casein, Tween, and Thimerosal (PBSCTT),and PBS with Tween (PBST) were prepared in the same man-ner, but PBSCTT contained, per liter, 0.8 g of casein, 0.5 mLof Tween-20, and 0.2 g of Thimerosal. PBST contained, perliter, 0.5 mL of Tween-20.
Preparation of hormone conjugates
11 a -OH-P-HS was conjugated with OVA as described byBacigalupo et al.(32) 11 a -OH-P-HS-HRP and progesterone 3-(O-carboxymethyl) oxime HRP (P-3-O-CMO-HRP) were syn-thesized by the mixed anhydride method according to Munroand Stabenfeldt. (31) The conjugates were chromatographe d ona Sephadex G-25 colum n, then tested for their titer and storedat 2 40°C.
Enzyme substrate solutions
The substrate solution containing o-phenylened iamine(OPD/H2O2) was prepared by dissolving 1.5 mg of OPD in10 mL 0.1 M citrate/phosphate buffer, pH 5.0 and by adding10 m L of 10% H2O2 . The substrate solution containing3.3 9 .5.5 9 -tetramethylbenzidine (TMB/H2O2) was prepared byadding of 100 m L TMB stock solution (10 mg TMB dissolvedin 1 mL of dimethysul foxide) and 8 m L of 10% H2O2 to 10
mL of 0.1 M acetate/ citrate buffer pH 4.5 containing 1 mMethylenedia minetetraa cetic acid disodium salt (Reanal, Bu-dapest, Hungary).
Immunization
Short-term immunization . Male BALB/c mice received anintraperitoneal (i.p.) injection of 50 m g of 11 a -OH-P-HS-BSA(molar ratio for steroid: BSA, 22:1). The antigen was dissolvedin 50 m L of PBS and emulsified in an equal volum e of com-plete Freund’s adjuvant. After an interval of 1 week, 50 m g ofantigen was given as above, except for the use of incompleteFreund’s adjuvant. Five days later, final boosts with 400 m g ofantigen in 0.1 mL PBS were given i.p. at 4, 3, and 2 days be-fore fusion was performed.
Long-term immunization . Male BALB/c mice received ani.p. injection of 100 m g of 11 a -OH-P-HS-BSA (molar ratio forsteroid, 22:1) dissolved in 50 m L of PBS and emulsified in anequal volume of complete Freund’s adjuvant. After an intervalof 2 months and 1 month, booster injections were given asabove. One month after the third injection, final boosts with200 m g of antigen in 0.1 mL of PBS were given i.p. on foursuccessive days. Fusion was performed one day after the lastinjection with the spleen cells of the donor mouse showing thehighest serum antibody titer.
Generation of MAbs
In each experiment, cells from one spleen were fused withSP2/0 myeloma cells from one 280-mL tissue culture flask(80–90% confluence). The splenocytes and the myeloma cellswere washed twice with Hank’s solution. The final pellet wasmixed by tapping the tube and 1 mL 50% (v/v) polyethyleneglycol (mol. wt. 4000 E. Merck AG, Darmstadt, Germany) inRPMI-1640 was added over 1 min with gentle shaking. Themixture was centrifuged at 130 3 g for 4 min. The supernatantwas poured off and the mixture was diluted by slow (1 min)addition of 10 mL of RPMI-1640 followed by slow addition (1min) of 10 mL of RPMI-1640 supplem ented with 10% FCS.The mixture was centrifuged for 10 min at 200 3 g, the super-natant was discarded and the fused cell pellet was resuspendedin hypoxantine-ami nopterin-thymidi ne (HAT) medium and dis-tributed (150 m L per well) in the 96-well tissue culture plates.After 20% confluence was reached, aliquots of hybridom a su-pernatants (50 m L) were tested by EIA to detect antiproges-terone antibodies. Selected clones were subcloned by the lim-iting dilution. Ascitic fluids were produced in pristane-prim edBALB/c mice.
Primary screening of positive clones
Two-fold diluted cell culture supernatants in PBSCTT (100m L) were incubated for 1 h at room temperature in the wells ofan 11 a -OH-P-HS-OVA immobilized plate (Titertek, Flow Lab-oratories, Irvine, Scotland). After washing with PBSCTT, in-cubation was continued for 1 h with 100 m L of 1/1000 goatanti-mouse immunoglobu lins (GAM Ig) coupled to HRP. Fol-lowing a second washing 100 m L of OPD/H2O2 substrate wasadded. The color reaction was terminated after 30 min by theaddition of 50 m L of 2 M H2SO4 and ODs were read at 492nm with a Twin Reader Plus (Labsystem, S. Turku, Finland).
WALDMANN290
Determination of specificity
Microtiter plates (Titertek) were first coated by dispensing100 m L of 3.2 m g/mL affinity purified GAM Ig in PBS. Theplates were incubated overnight at 4°C. After threefold wash-ing with PBSCTT, plates were incubated with 100 m L of suit-able diluted (1/500–1/2500) positive cell culture supernatantsor the donor mouse antiserum (1/250,000) in PBSCTT for 2 hat room temperature (the supernatants and the donor mouse an-tiserum dilutions were selected by 50% inhibition of tracer bind-ing). After three washes with PBSCTT, 10 m L of progesteronestandards and the potentially cross-reacting steroids in the rangeof 0 to 2187 nmol/L in PBS were added to the wells, followedby 100 m L of 11 a -OHP-HS-HRP conjugate (1/30,000). Theplates were incubated for 2 h at room temperature under con-stant shaking, then washed three times with PBSCTT. The colorwas developed with OPD/H2O2 substrate. The cross-reactivi-ties were calculated according to Abraham.(33)
Typing and subtyping of immunoglobu lins
Immunoglobul in classes and subclasses were determined bydouble antibody microtiter-plate EIA. The antiserum raisedagainst mouse m -chain coupled to HRP was prepared at this in-stitute. Rabbit antibodies raised against mouse immunoglobu -lins IgG1, IgG2a, IgG2b, and IgG3 were obtained from MilesLaboratories (Elkhart, IN).
Determination of relative affinity constant
First, MAbs were purified from ascitic fluid by ammoniumsulfate precipitation and ion exchange chromatography on BlueDEAE-Toyopearl 650S, using the Pharmacia standard FPLCsystem. (34) The IgG concentration was estimated at 280 nm us-ing an extinction coefficient of 14. The affinity constants weredetermined by incubation of serial dilutions of each MAb (from2 m g/mL to 2 ng/mL) in the wells of 11 a -OH-P-HS-OVA im-
MONOCLONAL ANTIBODIES FOR MILK PROGESTERONE EIA 291
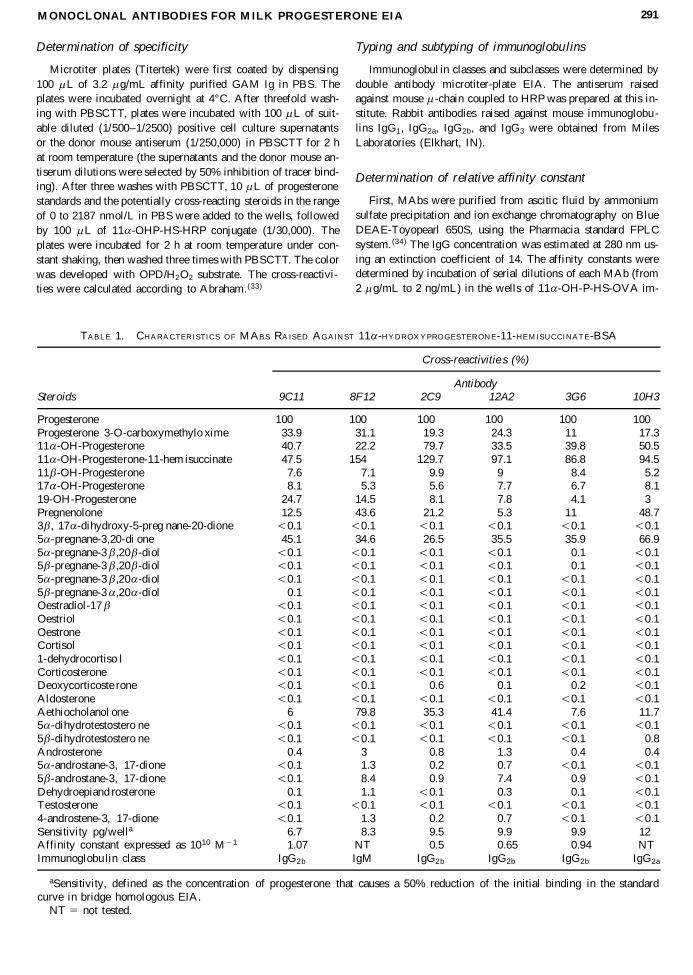
TABLE 1. CHARACTER ISTIC S O F MABS RA ISED AG AIN ST 11 a -H YDROX YPRO GES TER ON E-11-HE M ISUCCINAT E-BSA
Cross-reactivitie s (%)
AntibodySteroids 9C11 8F12 2C9 12A2 3G6 10H3
Progesterone 100 100 100 100 100 100Progesterone 3-O-carboxymethylo xime 33.9 31.1 19.3 24.3 11 17.311 a -OH-Progesterone 40.7 22.2 79.7 33.5 39.8 50.511 a -OH-Progesterone-11-hem isuccinate 47.5 154 129.7 97.1 86.8 94.511 b -OH-Progesterone 7.6 7.1 9.9 9 8.4 5.217 a -OH-Progesterone 8.1 5.3 5.6 7.7 6.7 8.119-OH-Progesterone 24.7 14.5 8.1 7.8 4.1 3Pregnenolone 12.5 43.6 21.2 5.3 11 48.73 b , 17 a -dihydroxy-5-preg nane-20-dione , 0.1 , 0.1 , 0.1 , 0.1 , 0.1 , 0.15 a -pregnane-3,20-di one 45.1 34.6 26.5 35.5 35.9 66.95 a -pregnane-3 b ,20 b -diol , 0.1 , 0.1 , 0.1 , 0.1 , 0.1 , 0.15 b -pregnane-3 b ,20 b -diol , 0.1 , 0.1 , 0.1 , 0.1 , 0.1 , 0.15 a -pregnane-3 b ,20 a -diol , 0.1 , 0.1 , 0.1 , 0.1 , 0.1 , 0.15 b -pregnane-3 a ,20 a -diol , 0.1 , 0.1 , 0.1 , 0.1 , 0.1 , 0.1Oestradiol-17 b , 0.1 , 0.1 , 0.1 , 0.1 , 0.1 , 0.1Oestriol , 0.1 , 0.1 , 0.1 , 0.1 , 0.1 , 0.1Oestrone , 0.1 , 0.1 , 0.1 , 0.1 , 0.1 , 0.1Cortisol , 0.1 , 0.1 , 0.1 , 0.1 , 0.1 , 0.11-dehydrocortiso l , 0.1 , 0.1 , 0.1 , 0.1 , 0.1 , 0.1Corticosterone , 0.1 , 0.1 , 0.1 , 0.1 , 0.1 , 0.1Deoxycorticoste rone , 0.1 , 0.1 , 0.6 , 0.1 , 0.2 , 0.1Aldosterone , 0.1 , 0.1 , 0.1 , 0.1 , 0.1 , 0.1Aethiocholanol one 6 79.8 35.3 41.4 7.6 11.75 a -dihydrotestostero ne , 0.1 , 0.1 , 0.1 , 0.1 , 0.1 , 0.15 b -dihydrotestostero ne , 0.1 , 0.1 , 0.1 , 0.1 , 0.1 , 0.8Androsterone 0.4 3 0.8 1.3 0.4 0.45 a -androstane-3, 17-dione , 0.1 1.3 0.2 0.7 , 0.1 , 0.15 b -androstane-3, 17-dione , 0.1 8.4 0.9 7.4 0.9 , 0.1Dehydroepiand rosterone 0.1 1.1 , 0.1 0.3 , 0.1 , 0.1Testosterone , 0.1 , 0.1 , 0.1 , 0.1 , 0.1 , 0.14-androstene-3, 17-dione , 0.1 1.3 0.2 0.7 , 0.1 , 0.1Sensitivity pg/wella 6.7 8.3 9.5 9.9 9.9 12Affinity constant expressed as 1010 M 2 1 1.07 NT 0.5 0.65 0.94 NTImmunoglobu lin class IgG2b IgM IgG2b IgG2b IgG2b IgG2a
aSensitivity, defined as the concentration of progesterone that causes a 50% reduction of the initial binding in the standardcurve in bridge homologous EIA.
NT 5 not tested.

mobilized plates. After 1 h incubation, the saturation curveswere evaluated by peroxidase labelled anti-m ouse IgG anti-bodies, as described above. The relative affinity constants werecalculated as the reciprocal of the MAb concentration giving50% saturation binding.
Evaluation of the performance of MAb 9C11 for EIAin bovine milk
Nunc-Immuno Plates MaxiSorp F-96 (Roskilde, Denmark)were coated by dispensing 200 m L of 3 m g/mL affinity puri-fied GAM Ig in PBS. The plates were incubated overnight at4°C. After being washed two times with PBST, plates weretreated for 10 min with PBS containing sucrose (50 g/L), BSA(5 g/L), and Thimerosal (0.2 g/L). The plates were air dried atroom temperature, sealed and stored at 4°C until use. Before
the assay, the plates were brought to room temperature. Twentymicroliters of milk samples or standards were added to thewells, followed by 100 m L of suitable diluted (1/12 500) P-3-O-CMO-HRP conjugate and 100 m L of MAb 9C11 (supernatantdiluted 1/2000) in PBSCT. The plates were coated with platesealing tape and incubated for 40 min at room temperature un-der constant shaking. The wells were washed four times withPBST. Two hundred microliters of TMB/H2O2 substrate wereadded and the color reaction stopped after 30 min by adding 50m L of 4 M H2SO4. The ODs were read at 450 nm with a Twin-Reader Plus (Labsystem, S. Turku, Finland).
Ovucheck method
Milk progesterone EIA kit based on polyclonal antiserumand alkaline phosphatase as enzym e label, Ovucheck™ (Cam-
WALDMANN292
TA BLE 2. CH ARACTER ISTIC S OF MABS AN D TH E DON OR MOU SE ANTIS ER UM
RA ISED AG AIN ST 11 a -H YDROX YPRO GES TER ON E-11-H EM ISUCCINA TE-BSA
Cross-reactivitie s (%)
DonorAntibody mouse
Steroid 6G12 9E7 7D1 1D7 10G11 antiserum
Progesterone 100 100 100 100 100 100Progesterone 3-O-carboxymethylo xime 25.5 36.4 25.6 22.1 8.9 10.911 a -OH-Progesterone 14.3 34.5 50 50 70.5 25.311 a -OH-Progesterone-11-hem isuccinate 55.6 106.1 109.5 66.3 176 237.811 b -OH-Progesterone 8.1 5.9 11.1 3.4 7 7.117 a -OH-Progesterone 3.2 6.1 55.2 28.3 3.2 1.419-OH-Progesterone 6 28 11.1 8.7 3.8 11Pregnenolone 23.6 19 4.5 2.7 15 4.13 b , 17 a -dihydroxy-5-preg nane-20-dione , 0.1 , 0.1 , 0.1 , 0.1 , 0.1 , 0.15 a -pregnane-3,20-di one 43.5 60 8.8 64.1 111 54.55 a -pregnane-3 b ,20 b -diol , 0.1 0.4 , 0.1 , 0.1 , 0.1 0.25 b -pregnane-3 b ,20 b -diol , 0.1 0.2 , 0.1 , 0.1 , 0.1 , 0.15 a -pregnane-3 b ,20 a -diol , 0.1 0.2 , 0.1 , 0.1 , 0.1 , 0.15 b -pregnane-3 a ,20 a -diol , 0.1 0.9 0.2 , 0.1 , 0.1 0.2Oestradiol-17 b , 0.1 , 0.1 , 0.1 0.2 , 0.1 , 0.1Oestriol , 0.1 0.2 , 0.1 , 0.1 , 0.1 , 0.1Oestrone , 0.1 , 0.1 , 0.1 , 0.1 , 0.1 , 0.1Cortisol , 0.1 , 0.1 , 0.1 , 0.1 , 0.1 , 0.11-dehydrocortisol 0.2 , 0.1 , 0.1 , 0.1 , 0.1 , 0.1Corticosterone , 0.1 , 0.1 , 0.1 , 0.1 , 0.1 , 0.1Deoxycorticoste one , 0.1 0.3 , 0.2 0.2 , 0.1 0.3Aldosterone , 0.1 , 0.1 , 0.1 , 0.1 , 0.1 , 0.1Aethiocholanol one 130.2 65.3 29.1 56 25.5 14.75 a -dihydrotestostero ne , 0.1 , 0.1 , 0.1 , 0.1 , 0.1 , 0.15 b -dihydrotestostero ne , 0.1 0.2 0.1 , 0.1 0.9 , 0.2Androsterone 8.5 1 0.3 1.1 0.6 , 0.15 a -androstane-3, 17-dione 0.4 0.3 0.9 0.1 , 0.1 , 0.15 b -androstane-3, 17-dione 3.1 0.5 , 0.1 10.4 , 0.1 0.5Dehydroepiandr osterone 1.7 0.9 0.5 , 0.1 0.9 0.3Testosterone , 0.1 , 0.1 , 0.1 , 0.1 , 0.1 , 0.14-androstene-3, 17-dione 0.4 0.3 0.9 0.1 , 0.1 , 0.1Sensitivity pg/wella 12.3 12.8 13.9 14.3 31.7 10.4Affinity constant expressed as 1010 M 2 1 NT NT NT NT NT NTImmunoglobu lin class IgG1 IgG1 IgG1 IgG1 IgG1 –
aSensitivity, defined as the concentration of progesterone that causes a 50% reduction of the initial binding in the standardcurve in bridge homologous EIA.
NT 5 not tested.

bridge Veterinary Sciences, England), was used for compari-son studies. Milk samples were analyzed according to the man-ufacturer’ s instructions.
RESULTS
Production and characterization of antibodies
Two out of three hybridizations were successful in terms ofproduction of antiprogesterone MAbs in the short-term immu-nization group, leading to 10 and 4 stabile hybridoma cell lines,respectively, producing antiprogesterone antibodies. However,all of the antibodies produced lacked sufficient sensitivity incompetitive EIA, and were, therefore, not characterized any fur-ther. On the day of sacrifice, one mouse from the long-term im-munization group produced antiserum with high titer of1/250,000, defined as the dilution of antiserum which resultedin 50% binding of tracer. The fusion experiment with the spleencells of this mouse resulted in 17 stabile antiprogesterone Igproducing cell lines. Eleven of the MAbs were selected for fur-ther characterization because the antibodies showed high sen-sitivity in competitive EIA, and were, therefore, considered po-tentially suitable for EIA in bovine milk. The results of thecharacterization of these MAbs and the donor mouse polyclonalantiserum are shown in Tables 1 and 2. The MAbs and the poly-clonal antiserum were generally found to be highly specificwhen tested with 30 different steroids. The MAbs and the donormouse antiserum showed remarkably high cross-reactivity withthe steroids differing primarily by a hydroxyl group in the 11th
position. None of the MAbs characterized was more specificthan the donor mouse antiserum. The standard curve midpointsensitivities of the MAbs in competitive EIA ranged from 6.7to 31.7 pg/well. One MAb from clone 9C11 demonstrated im-proved sensitivity (p , 0.05) requiring 6.7 pg of progesterone,to inhibit 50% of the standard curve, compared with 10.4 pgfor the donor mouse antiserum. Other MAbs produced equal orless sensitive standard curves in competitive EIA. The affini-ties of four antibodies, showing the most sensitive standardcurves, ranged from Ka 5 0.5 3 1010 to 1.1 3 1010 M 2 1. MAbfrom clone 9C11, which gave the most sensitive dose–responsecurve, also showed highest affinity to progesterone (Table 1).Of the MAbs, most were of IgG1 and IgG2b subclasses.
EIA of progesterone in bovine milk
MAb from clone 9C11 was chosen for EIA in milk since itshowed better characteristics than the other MAbs. The EIAwas optimized for measurement of progesterone in defattedmilk and whole milk. The standard curves ranged from 0.25 to10 ng/mL and from 1 to 60 ng/mL for the skim milk and wholemilk assay, respectively (Fig. 1), covering the whole range ofphysiological values of progesterone during the oestrus cycleand pregnancy. The corresponding range of OD values were2.3 to 0.25 and 2.5 to 0.2 for the skim-milk and whole-milk as-say, respectively. Sensitivities, as determ ined by the midpointof the calibration curves were 1.25 ng/mL (25 pg/well) for theskim-milk progesterone assay and 5.16 ng/mL (103.2 pg/well)for the whole-milk progesterone assay, respectively. The limitof sensitivity was about 0.1 and 0.25 ng/mL for the skim-milk
and whole-milk assay, respectively. The between-plate preci-sion was tested by duplicate measurement of 3 milk samples in14 consecutive assays. The results are given in Table 3. Withinassay variation (CV) was , 10% for both assays. Good corre-lation (r 5 0.92, p , 0.001) between progesterone values inbovine milk was obtained when 52 milk samples were assayedby EIA and Ovucheck™ EIA method. The regression line equa-tion was: y 5 0.92x 1 0.45, where y is the concentration ofprogesterone measured with EIA and x with the Ovucheck™method.
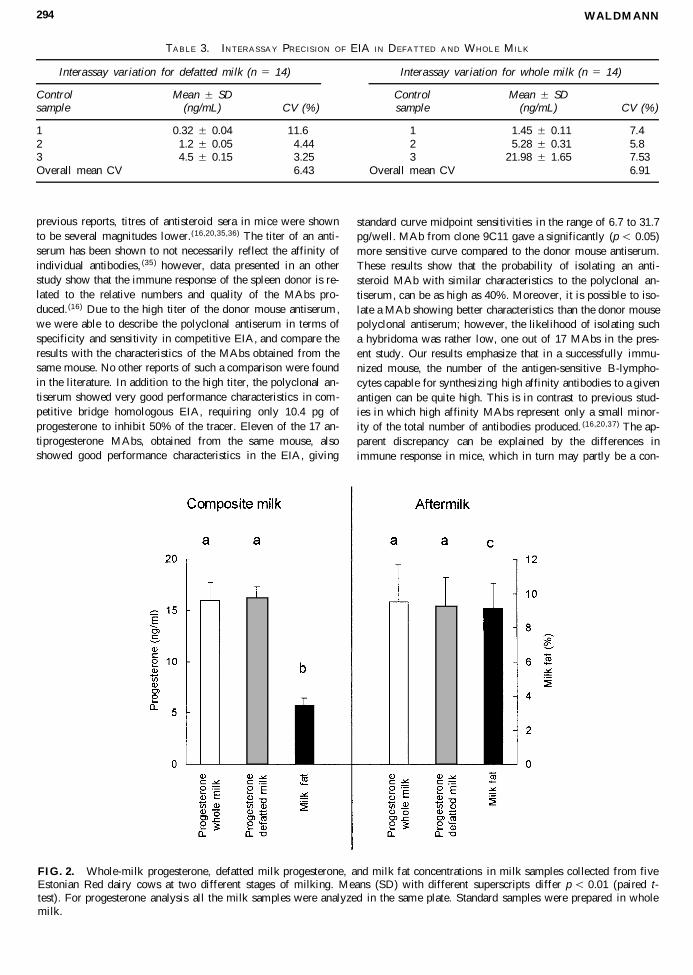
The influence of milk fat on the assay results was tested bymeasuring progesterone in whole milk and in defatted milk inmilk samples collected from five cows at two different stagesof milking. All the samples were analysed in the same plate byusing standard samples prepared in whole milk. The results arepresented in Fig. 2. No difference (p . 0.05) in mean whole-milk progesterone concentrations was found in milk samplesthat were taken from composite milk and aftermilk. At the sametime mean milk fat concentrations differed significantly, beinglower (p , 0.001) in composite milk than in aftermilk. Meanprogesterone concentrations in defatted milk, prepared from thesame milk samples, did not differ (p . 0.05) from those mea-sured in whole milk.
DISCUSSION
MAbs to progesterone were obtained under a variety of im-munization conditions. High affinity MAbs to progesteronewere produced by using a short immunization protocol in oneprevious study, (19) as well as with longer periods of immu-nization. (12,13,17,20) In the present study, however, we failed toproduce suitable MAbs for milk progesterone EIA by usingshort-term immunization. Long-term immunization with highdoses of antigen led to a very good antibody response to pro-gesterone-BSA antigen in mice. The donor mouse produced an-tiserum with a titer as high as 1/250,000 at 50% binding. In
MONOCLONAL ANTIBODIES FOR MILK PROGESTERONE EIA 293
FIG. 1. Standard curves for skim milk ( j ) and whole ( m )milk progesterone EIA. Bars indicate standard deviations frommeans obtained from 14 consecutive standard curves analyzedin duplicate.
previous reports, titres of antisteroid sera in mice were shownto be several magnitudes lower.(16,20,35,36) The titer of an anti-serum has been shown to not necessarily reflect the affinity ofindividual antibodies, (35 ) however, data presented in an otherstudy show that the immune response of the spleen donor is re-lated to the relative numbers and quality of the MAbs pro-duced. (16) Due to the high titer of the donor mouse antiserum ,we were able to describe the polyclonal antiserum in terms ofspecificity and sensitivity in competitive EIA, and compare theresults with the characteristics of the MAbs obtained from thesame mouse. No other reports of such a comparison were foundin the literature. In addition to the high titer, the polyclonal an-tiserum showed very good performance characteristics in com-petitive bridge homologous EIA, requiring only 10.4 pg of progesterone to inhibit 50% of the tracer. Eleven of the 17 an-tiprogesterone MAbs, obtained from the same mouse, alsoshowed good performance characteristics in the EIA, giving
standard curve midpoint sensitivities in the range of 6.7 to 31.7pg/well. MAb from clone 9C11 gave a significantly (p , 0.05)more sensitive curve compared to the donor mouse antiserum.These results show that the probability of isolating an anti-steroid MAb with similar characteristics to the polyclonal an-tiserum , can be as high as 40%. Moreover, it is possible to iso-late a MAb showing better characteristics than the donor mousepolyclonal antiserum; however, the likelihood of isolating sucha hybridoma was rather low, one out of 17 MAbs in the pres-ent study. Our results emphasize that in a successfully immu-nized mouse, the number of the antigen-sensitive B-lympho-cytes capable for synthesizing high affinity antibodies to a givenantigen can be quite high. This is in contrast to previous stud-ies in which high affinity MAbs represent only a small minor-ity of the total number of antibodies produced. (16,20,37) The ap-parent discrepancy can be explained by the differences inimmune response in mice, which in turn may partly be a con-
WALDMANN294
TABLE 3. IN TER ASSA Y PREC ISION OF EIA IN DEF AT TED A ND WHOL E MILK
Interassay variation for defatted milk (n 5 14) Interassay variation for whole milk (n 5 14)
Control Mean 6 SD Control Mean 6 SDsample (ng/mL) CV (%) sample (ng/mL) CV (%)
1 0.32 6 0.04 11.61 1 1.45 6 0.11 7.412 1.2 6 0.05 4.44 2 5.28 6 0.31 5.813 4.5 6 0.15 3.25 3 21.98 6 1.65 7.53Overall mean CV 6.43 Overall mean CV 6.91
FIG. 2. Whole-milk progesterone, defatted milk progesterone, and milk fat concentrations in milk samples collected from fiveEstonian Red dairy cows at two different stages of milking. Means (SD) with different superscripts differ p , 0.01 (paired t-test). For progesterone analysis all the milk samples were analyzed in the same plate. Standard samples were prepared in wholemilk.

sequence of immunization protocol. The results of our studysupport the suggestion that to enhance the probability of iso-lating the desired hybridoma, animals whose sera show a goodimmune response should be used for fusion experim ents.
The specificities of the MAbs generated, show great simi-larity with MAbs produced by other groups against the sameantigen, (12,14,19 ,20) thus, confirming the finding that specifici-ties of the antisteroid MAbs depend on the nature of steroidlink to the protein carrier and the position on the steroid nu-cleus at which the linker is attached. (38)
De Boever et al.(39) studied the performance of antiproges-terone MAbs in a “solid” system, where progesterone-spec ificantibodies were coated to the microplate wells via second an-tibodies, and in a “liquid” system, where soluble antiproges-terone MAb was added to the wells, which had been coated inadvance with second antibody. The study showed that the het-erologous (P-3-CMO aminocaproic acid-HRP) and the homol-ogous (P-11-HS-HRP) conjugates produced highly sensitivecalibration curves in a “solid” system, but in a “liquid” systemthe binding of the heterologous conjugate to the solid phase wasalmost annihilated, reducing the sensitivity by several orders ofmagnitude. In contrast, MAb 9C11 produced highly sensitivecalibration curves in homologous “solid” system and heterolo-gous “liquid” system in the present study. These results showthat antiprogesterone MAbs, obtained against the same im-munogen, may show different performance characteristics, evenwhen tested in similar conditions. Our study is the first reportdescribing the use of antiprogesterone MAb in combinationwith the “second antibody coating technique” in milk proges-terone EIA.
The presence of milk has been shown to cause a significantdepression in antibody-antigen binding by lowering OD values,and by reducing the sensitivity of the assay. (9,27,40) Therefore,the prerequisite characteristic of the antibody, for use in milkprogesterone immunoassay, is to maintain the binding charac-teristics in the presence of milk. In contrast to previous re-ports,(5,40) the use of 20 m L of skim milk or whole milk pro-duced steep calibration curves (Fig. 1) with the wide range ofOD values, allowing to omit the extraction or dilution of milkprior to analysis.
The major advantage of the present assay, compared to pre-viously published direct-m ilk progesterone immunoassays , isthat progesterone concentrations measured by the EIA were notinfluenced by milk fat concentrations up to 10% of milk fat(Fig. 2). This phenomenon can be explained by the fact thatdue to the MAb used in the assay, the rate of binding of pro-gesterone to the antibody is fast compared with progesteronemoving from the fat phase. In previous studies, it has been con-sidered preferable to estimate progesterone concentration in fat-free milk or milk fat,(41) however, these techniques needed timeand labor consuming milk fat separation.
In conclusion, we have shown that very high titers(1/250,000) can be obtained in mice against steroid hormones.The likelihood of isolating a high affinity MAb from such ahigh titer donor mouse was substantially higher when comparedto data from the literature. However, more experiments have tobe undertaken to draw final conclusions. The carefully selectedantiprogesterone MAb 9C11 allowed us to develop a simpleEIA of progesterone in bovine milk, which offers considerableadvantages over previously described milk progesterone EIAs.
The assay is simple and convenient to use, as it permits directaddition of undiluted sample; at the same time this assay main-tains high sensitivity with a wide range of OD values, whichagain makes it possible to read the results with the naked eye.In addition, changing milk fat concentrations did not affectprogesterone concentrations in our assay, thus allowing the di-rect analysis of progesterone from milk samples with differentmilk-fat concentrations. These results suggest that the EIA testwith MAb 9C11 may be a simple and quick method to analyzeprogesterone for research and veterinary managem ent of dairycows.
ACKNOWLEDGMENTS
Dr. E. Juronen is gratefully acknowledged for MAb purifi-cation. Cambridge Veterinary Sciences Ltd. is acknowledgedfor providing Ovucheck™ milk progesterone kits.
REFERENCES
1. Dobson H, Midmer SE, and Fitzpatrick RJ: Relationship betweenprogesterone concentrations in milk and plasma during the bovineoestrous cycle. Vet Rec 1975;96:222– 223.
2. Foote RH, Oltenacu EAB, Kummerfeld HL, Smith RD, Riek PM,and Braun RK: Milk progesterone as a diagnostic aid. Br Vet J1979;135:550– 558.
3. Esslemont RJ: Economic appraisal of herd health schemes. In: TheVeterinary Annual 35, Raw M-E and Parkinson TJ (Eds.). Black-well Science Ltd., Oxford, UK 1995, pp. 243–280.
4. Arnstadt KI, and Schmidt-Adamopoulou B: Direct enzyme-im-munoassay for determination of progesterone in milk of cows. BrVet J 1982;138:436– 438.
5. Chang CF, and Estergreen VL: Development of a direct enzymeimmunoassay of milk progesterone and its application to pregnancydiagnosis in cows. Steroids 1983;41:173– 195.
6. Cleere WF, Gosling JP, Morris MC, Charleton MF, Moloney BT,Fottrell PF, and Sreenan JM: A high performance, high through-put enzymeimmunoassay for the analysis of progesterone in plasmaor milk. Ir Vet J 1985;39:6– 14.
7. Marcus GJ, and Hackett AJ: Use of enzyme-linked immunoassayfor measurement of bovine serum and milk progesterone withoutextraction. J Dairy Sci 1986;69:818– 824.
8. Prakash BS, Madan ML, Jailkhani S, and Singla SK: Developmentof a simple, direct, microtitre plate enzymeimmunoassay (EIA) forprogesterone determination in whole milk of buffaloes. Br Vet J1990;146:571– 576.
9. Sauer MJ, Foulkes JA, Worsfold A, and Morris BA: Use of pro-gesterone 11-glucuronide- alkaline phosphatase conjugate in a sen-sitive microtitre-plat e enzyme immunoassay of progesterone inmilk and its application to pregnancy testing in dairy cattle. J Re-prod Fertil 1986;76:375– 391.
10. Wimpy TH, Chang CF, Estergreen VL, and Hillers JK: Milk pro-gesterone enzyme immunoassay : Modifications and field trial forpregnancy detection in dairy cows. J Dairy Sci 1986;69:1115– 1121.
11. Van De Wiel DFM, and Koops W: Development and validation ofan enzyme immunoassay for progesterone in bovine milk or bloodplasma. Anim Reprod Sci 1986;10:201– 213.
12. Booman P, Tieman M, van de Wiel DFM, Schakenraad JM, andKoops W: Application of monoclonal antibodies in animal pro-duction: Pregnancy diagnosis in cattle. In: Innovations in Biotech-nology . Houwink EH and Van der Meer RR (Eds.). Elsevier Sci-ence Publishers, Amsterdam, 1984, pp. 259–266.
MONOCLONAL ANTIBODIES FOR MILK PROGESTERONE EIA 295

13. Brochu M, Veilleux R, Lorrain A, and Belanger A: Monoclonalantibodies for use with 125iodine-labeled radioligands in proges-terone radioimmunoas say. J Steroid Biochem 1984;21:405– 411.
14. Fantl VE, Wang DY, and Whitehead AS: Production and charac-terisation of a monoclonal antibody to progesterone. J SteroidBiochem 1981;14:405– 407.
15. Fantl VE, Wang DY, and Knyba RE: The production of high affin-ity monoclonal antibodies to progesterone. J Steroid Biochem1982;17:125– 130.
16. Fantl VE, and Wang DY: Simultaneous production of monoclonalantibodies to dehydroepiandros terone, oestradiol, progesterone andtestosterone. J Endocrinol 1984;100:367– 376.
17. Gupta SK, Rao DN, Das C, Johansson EDB, and Talwar GP: Highaffinity specific monoclonal antibodies against progesterone. In-dian J Med Res 1987;85:542– 549.
18. O’Rorke A, Kane MM, Gosling JP, Tallon DF, and Fottrell PF:Development and validation of a monoclonal antibody enzyme im-munoassay for measuring progesterone in saliva. Clin Chem1994;40:454– 458.
19. Schramm W, Yang T, and Midgley AR: Monoclonal antibodiesused in solid-phase and liquid-phase assays, as exemplified byprogesterone assay. Clin Chem 1987;33:1331– 1337.
20. Siklódi B, Barna-Vetró I, and Solti L: Latent immunization to pro-duce high-affinity monoclonal antibodies to progesterone. Hy-bridoma 1995;14:79– 84.
21. White A, Anderson DC, and Daly R: Production of a highly spe-cific monoclonal antibody to progesterone. J Clin Endocr Metab1982;54:205– 207.
22. Brochu M, Carrie ML, Terouanne B, Belander A, and Nicolas JC:Bioluminescent enzyme immunoassay for progesterone using mon-oclonal antibodies and glucose-6-phosp hate dehydrogenase labels.J Biolumin Chemilumin 1987;4:215– 222.
23. Miller SA, Morton MS, and Turkes A: Chemiluminesc ence im-munoassay for progesterone in plasma incorporating acridinum es-ter labelled antigen. Ann Clin Biochem 1988;25:27– 34.
24. Yoon DY, Song EY, Kwon DH, Choi MJ, Byun SM, Choe IS, andChung TW: Use of progesterone- 3(O-carboxymethyl oxime)-horseradish peroxidase in a sensitive microtitre-pla te EIA and itsapplication to a visual membrane EIA of progesterone. J Im-munoassay 1995;16:137– 153.
25. Nagy P, Solti L, Kulcsár M, Reiczigel J, Huszenicza Gy, AbaváryK, and Wölfling A: Progesterone determination in equine plasmausing different immunoassays . Acta Vet Hung 1998;46:501– 513.
26. Choi MJ, Choi J, Yoon DY, Park J, and Eremin SA: Fluorescencepolarization immunoassay of progesterone. Biol Pharm Bull1997;20:309– 314.
27. Groves DJ, Sauer MJ, Rayment P, Foulkes JA, and Morris BA:The preparation of an ovine monoclonal antibody to progesterone.J Endocr 1990;126:217– 222.
28. Stanley CJ, Paris F, Webb AE, Heap RB, Ellis ST, Hamon M,Worsfold A, and Booth JM: Use of a new and rapid milk proges-terone assay to monitor reproductive activity in the cow. Vet Rec1986;118:664– 667.
29. Waldmann A: Enzyme immunoassay (EIA) for milk progesteroneusing a monoclonal antibody. Anim Reprod Sci 1993;34:19– 30.
30. Meyer HHD, and Güven B: Improvement of microtitration plateenzymeimmunoassays for steroid determination by a second anti-body technique. J Steroid Biochem 1986;25:139.
31. Munro C, and Stabenfeldt G: Development of a microtitre plateenzyme immunoassay for the determination of progesterone. J En-docr 1984;101:41– 49.
32. Bacigalupo MA, Ferrara L, Meroni G, and Ius A: Time-resolvedfluoroimmuno assay of progesterone by Eu-labelled protein-A. Fre-senius J Anal Chem 1987;328:263– 264.
33. Abraham GE: Solid phase radioimmunoa ssay for estradiol17 b . JClin Endocr Metab 1969;29:886– 870.
34. Juronen E, Parik J, and Toomik P: FPLC purification of mousemonoclonal antibodies from ascitic fluid using blue DEAE andthiophilic sorbents. J Immunol Methods 1991;136:103– 109.
35. de Lauzon S, Le Trang N, Moreau MF, Gentin M, Christeff N,Desfosses B, and Cittanova N: Murine monoclonal antibody againstaldosterone: Production, characterization and use for enzymoim-munoassay. J Steroid Biochem 1987;28:459– 463.
36. Parvaz P, Mathian B, Patricot MC, Garcia I, Revol A, Mappus E,Grenot C, and Cuilleron CY: Production of monoclonal antibodiesto dehydroepiandr osterone-sulpha te after immunization of mousewith dehydroepiandro sterone-bovine serum albumin conjugate. JSteroid Biochem 1989;32:553– 558.
37. Eshhar Z, Kim JB, Barnard G, Collins WP, Gilad S, Lindner HR,and Kohen F: Use of monoclonal antibodies to pregnanediol-3 a -glucuronide for the development of a solid phase chemilumines-cence immunoassay. Steroids 1981;38:89– 109.
38. Gani M, Coley J, Piron J, Humphreys AS, Arevalo J, Wilson IA,and Taussig MJ: Monoclonal antibodies against progesterone: ef-fect of steroid-carrier coupling position on antibody specificity. JSteroid Biochem Mol Biol 1994;48:277– 282.
39. De Boever JG, Kohen F, and Bosmans E: Binding of homologousand heterologous isoluminol- and enzyme-labelle d progesteroneconjugates to monoclonal antibodies. Analytica Chimica Acta1994;290:239– 245.
40. Prakash BS, Meyer HHD, and van de Wiel DFM: Sensitive en-zyme immunoassay of progesterone in skim milk using second-an-tibody technique. Anim Reprod Sci 1988;16:225– 235.
41. Pope GS, and Swinburne JK: Reviews of the progress of dairy sci-ence: Hormones in milk: Their physiological significance and valueas diagnostic aids. J Dairy Res 1980;47:427– 449.
Address reprint requests to:Andres Waldmann
Institute of General and Molecular PathologyUniversity of Tartu
34 Veski str., 51014 TartuEstonia
Received for publication February 23, 1999. Accepted for pub-lication April 10, 1999.
WALDMANN296