mécanismes de survenue des anomalies chromosomiques de ... · levée du blocage par changement de...
TRANSCRIPT

Mécanismes de survenue des anomalies chromosomiques de structure
JM DUPONTLaboratoire de CytogénétiqueGroupe Hospitalier Cochin Saint Vincent de PaulUniversité Paris Descartes - Faculté de MédecineParis

Anomalies de structure Fréquence = 0,5% équilibrées 0,06% déséquilibrées
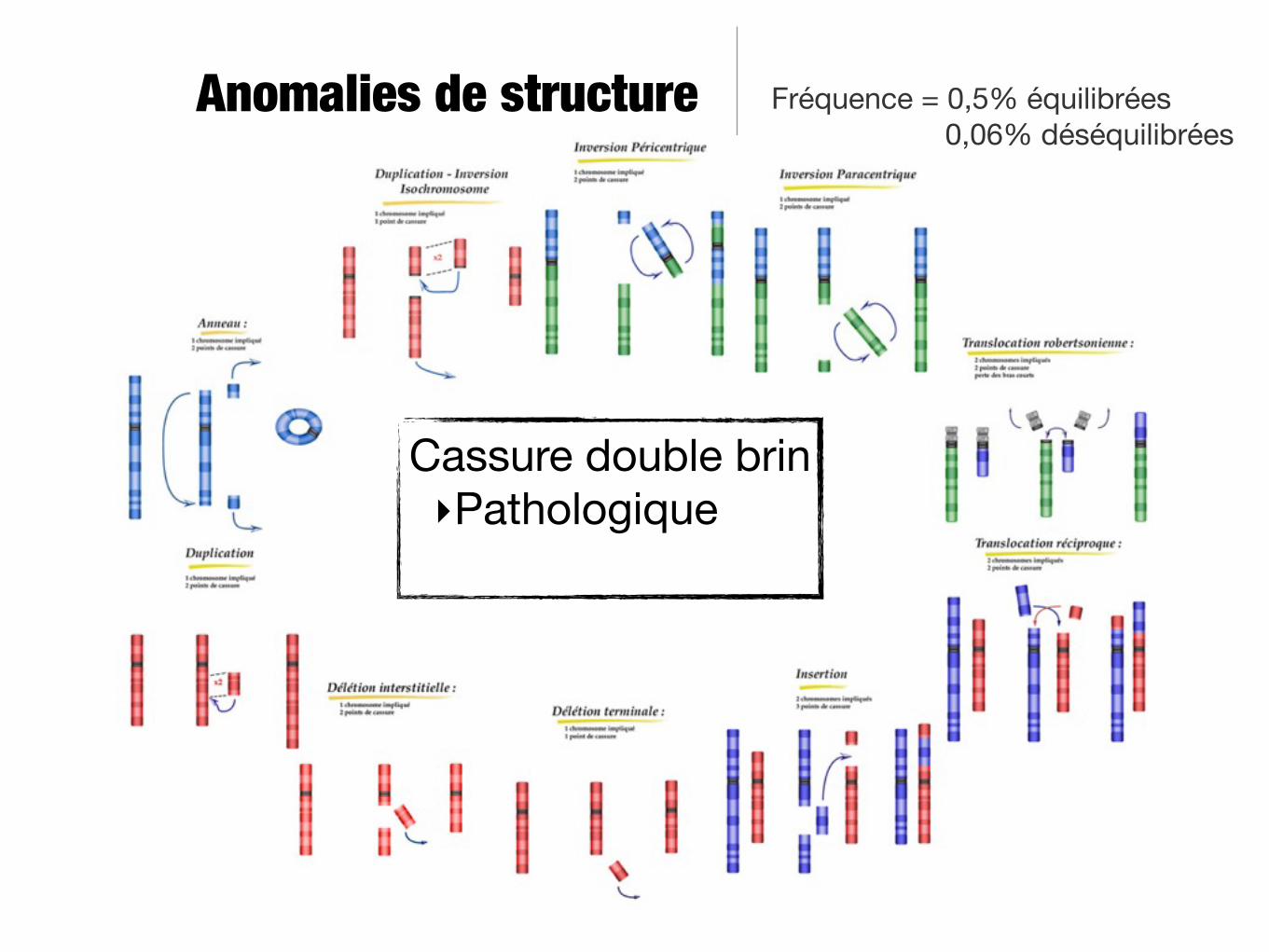
Anomalies de structure
Cassure double brin‣Pathologique
Fréquence = 0,5% équilibrées 0,06% déséquilibrées

Anomalies de structure
Cassure double brin‣Pathologique‣Physiologique
Fréquence = 0,5% équilibrées 0,06% déséquilibrées

CDB physiologiques

CDB physiologiques
• CDB méiotiques déclenchées par Spo11
• Première étape de la recombinaison méiotique

CDB physiologiques
• CDB méiotiques déclenchées par Spo11
• Première étape de la recombinaison méiotique
• CDB déclenché par le complexe RAG
• Recombinaison V(D)J

Cassures double-brin (CDB) pathologiques
• ≈ 10 CDB / j / cell
• Stress réplicatif
• O2- provenant du métabolisme oxydatif -> radicaux hydroxyl (≈ 100/h/cell)
• Radiations ionisantes
• rayons γ, rayons X (≈ 300 Millions/personne/heure)
• Erreur de processus enzymatiques normaux
• Défaut de la Topoisomérase II (pas de ligation), action du complexe RAG en dehors de la région V(D)J
• Stress mécanique

Redondance des mécanismes de réparation
R-S Mani & A M Chnnaiyan., Nature Rev Genetics 2010-11;819

Redondance des mécanismes de réparation
R-S Mani & A M Chnnaiyan., Nature Rev Genetics 2010-11;819
Modification des extrêmités et ligation
«peu» fiableTout au long du cycle
Quelque soit l’état de l’extrêmitéGénère de la diversité par les
réarrangements formés

Redondance des mécanismes de réparation
R-S Mani & A M Chnnaiyan., Nature Rev Genetics 2010-11;819
Remplacement de l’ADN cassé par une copie
Essentiellement S et G2Fidélité «parfaite»
Modification des extrêmités et ligation
«peu» fiableTout au long du cycle
Quelque soit l’état de l’extrêmitéGénère de la diversité par les
réarrangements formés

Réparation par recombinaison homologue
• Remplacement de la région lésée par une copie homologue
PJ Hastings et al., Nat Rev Genetics, 2009-10;551
Préparation des extrémités
Copie
Réparation

Réparation par recombinaison homologue

Réparation par recombinaison homologue
• Homologie de séquence
• > 97% sur 50 - 300 pb
PJ Hastings et al., Nat Rev Genetics, 2009-10;551
EXO I + BLM (Bloom)

Réparation par recombinaison homologue
• Homologie de séquence
• > 97% sur 50 - 300 pb
• Stabilisation de l’ADN simple brin
• RPA, RAD52
PJ Hastings et al., Nat Rev Genetics, 2009-10;551
EXO I + BLM (Bloom)

T Sugiyama et N Kantake, J Mpl Biol 2009-390;45
Réparation par recombinaison homologue
• Homologie de séquence
• > 97% sur 50 - 300 pb
• Stabilisation de l’ADN simple brin
• RPA, RAD52
PJ Hastings et al., Nat Rev Genetics, 2009-10;551
EXO I + BLM (Bloom)

Réparation par recombinaison homologueT Sugiyama et N Kantake, J Mpl Biol 2009-390;45
RAD51 + BRCA2 + cohesines
• Homologie de séquence
• Stabilisation de l’ADN simple brin
• Formation d’un nucléofilament
• RAD51-ssDNA
• Formation d’une synapse -> Recherche d’homologie
ME Moynahan, M Jasin, Nat Rev Mol Cell Biol, 2010-11;196
ATP
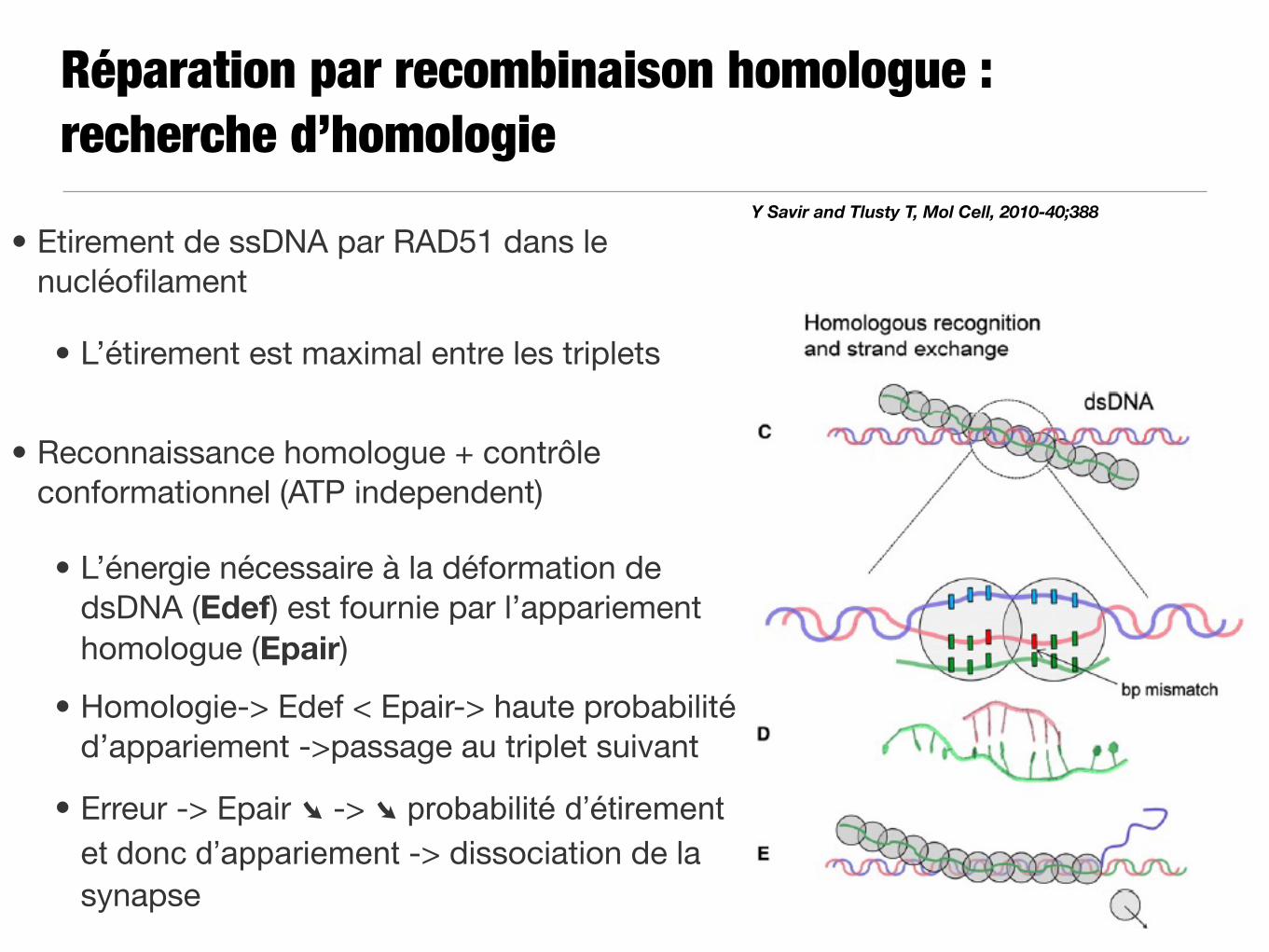
Réparation par recombinaison homologue : recherche d’homologie
Y Savir and Tlusty T, Mol Cell, 2010-40;388
• Etirement de ssDNA par RAD51 dans le nucléofilament
• L’étirement est maximal entre les triplets
• Reconnaissance homologue + contrôle conformationnel (ATP independent)
• L’énergie nécessaire à la déformation de dsDNA (Edef) est fournie par l’appariement homologue (Epair)
• Homologie-> Edef < Epair-> haute probabilité d’appariement ->passage au triplet suivant
• Erreur -> Epair ➘ -> ➘ probabilité d’étirement et donc d’appariement -> dissociation de la synapse

Réparation par recombinaison homologueT Sugiyama et N Kantake, J Mpl Biol 2009-390;45
RAD51 + BRCA2 + cohesines
DSBRDouble Strand Break Repair
SDSASynthesis-Dependant strand
annealing
• Homologie de séquence
• Stabilisation de l’ADN simple brin
• Formation d’un nucléofilament
• Recherche d’homologie
• Etape clé = invasion du brin homologue (D-loop) : RAD51

Réparation par recombinaison homologue : Crossing over / Conversion génique
• Homologie de séquence
• Etape clé = invasion du brin homologue (D-loop)
• Résolution
• DSBR (Holliday junction)
• Crossing over
• Conversion génique
PJ Hastings et al., Nat Rev Genetics, 2009-10;551
Endonucléase
Double Holliday Junction

Réparation par recombinaison homologue : Crossing over / Conversion génique
• Homologie de séquence
• Etape clé = invasion du brin homologue (D-loop)
• Résolution
• DSBR (Holliday junction)
• Crossing over
• Conversion génique
• Migration
PJ Hastings et al., Nat Rev Genetics, 2009-10;551
Double Holliday Junction
Hélicase + Topoisomérase
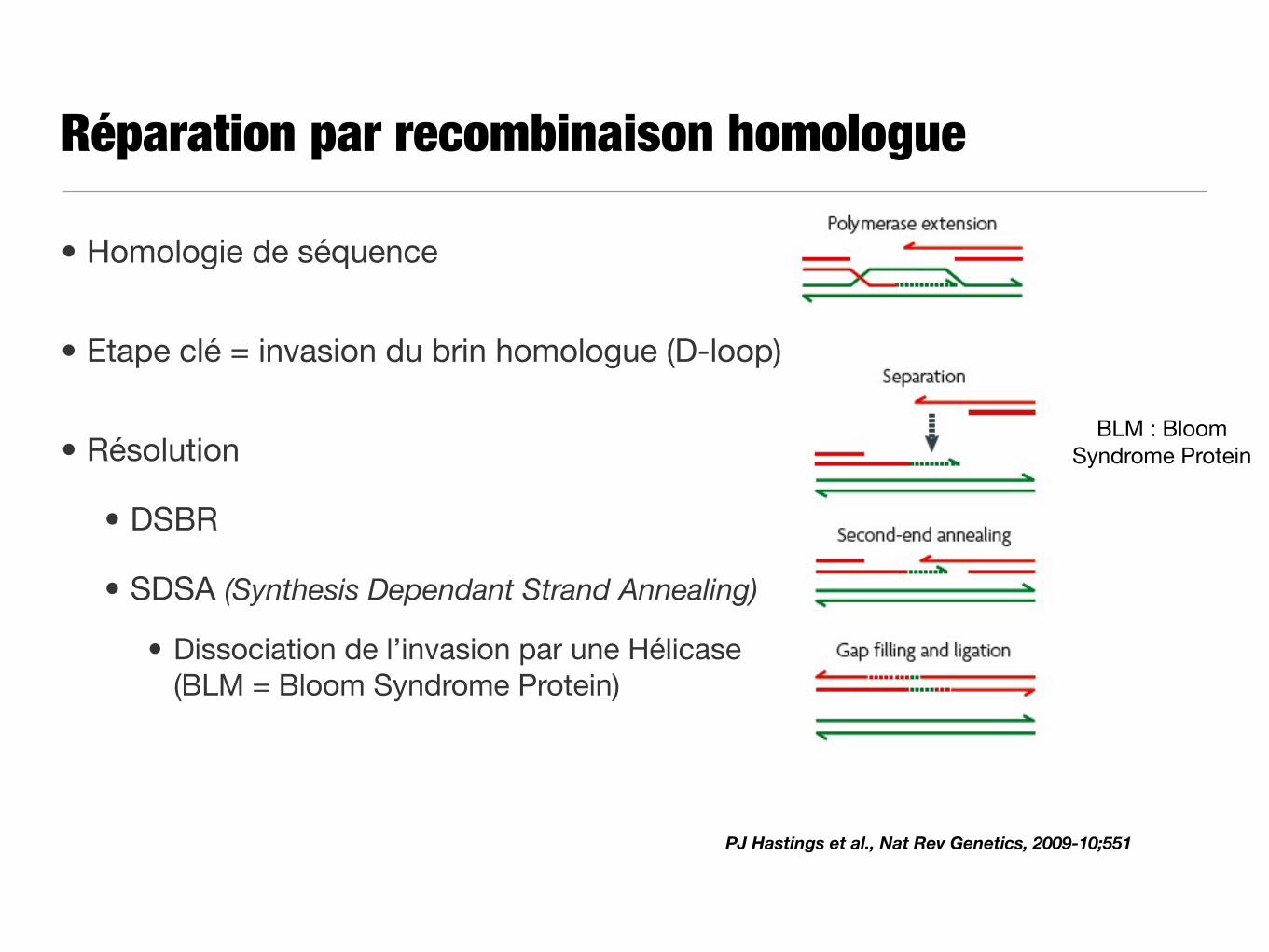
Réparation par recombinaison homologue
• Homologie de séquence
• Etape clé = invasion du brin homologue (D-loop)
• Résolution
• DSBR
• SDSA (Synthesis Dependant Strand Annealing)
• Dissociation de l’invasion par une Hélicase (BLM = Bloom Syndrome Protein)
PJ Hastings et al., Nat Rev Genetics, 2009-10;551
BLM : Bloom Syndrome Protein

Réparation par fusion des extrémités
± MRN(MRE11+RAD50+NBS1)
(Ku70+Ku80)
MR Lieber, Annu Rev Biochem, 2010-79;181

Réparation par fusion des extrémités• NHEJ : Non Homologous End Joining
• Nucléase, DNA polymérase, ligase
± MRN(MRE11+RAD50+NBS1)
(Ku70+Ku80)
MR Lieber, Annu Rev Biochem, 2010-79;181

Réparation par fusion des extrémités• NHEJ : Non Homologous End Joining
• Nucléase, DNA polymérase, ligase
• Fixation du complexe Ku (Ku70+Ku80)
• Facilite la fixation des autres enzymes
± MRN(MRE11+RAD50+NBS1)
(Ku70+Ku80)
MR Lieber, Annu Rev Biochem, 2010-79;181

Réparation par fusion des extrémités• NHEJ : Non Homologous End Joining
• Nucléase, DNA polymérase, ligase
• Fixation du complexe Ku (Ku70+Ku80)
• Facilite la fixation des autres enzymes
• Action itérative dans n’importe quel ordre et indépendamment sur les deux extrêmités -> modification pour permettre la ligation
• polymérases : Pol µ and λ
• nucléase: artemis:DNA-PKcs
• ligase
± MRN(MRE11+RAD50+NBS1)
(Ku70+Ku80)
MR Lieber, Annu Rev Biochem, 2010-79;181

Réparation par fusion des extrémités
MR Lieber, Annu Rev Biochem, 2010-79;181

Réparation par fusion des extrémités
• Ligation finale
• DNA ligase IV ±XRCC4±XLF
MR Lieber, Annu Rev Biochem, 2010-79;181

Réparation par fusion des extrémités
• Ligation finale
• DNA ligase IV ±XRCC4±XLF
• XRCC4 et XLF augmentent l’efficacité de DNA ligase IV sur des extrêmités non homologues
MR Lieber, Annu Rev Biochem, 2010-79;181

Réparation par fusion des extrémités
• Ligation finale
• DNA ligase IV ±XRCC4±XLF
• XRCC4 et XLF augmentent l’efficacité de DNA ligase IV sur des extrêmités non homologues
• «Cicatrice» au point de cassure
MR Lieber, Annu Rev Biochem, 2010-79;181

Réparation par fusion des extrémités
• Ligation finale
• DNA ligase IV ±XRCC4±XLF
• XRCC4 et XLF augmentent l’efficacité de DNA ligase IV sur des extrêmités non homologues
• «Cicatrice» au point de cassure
• Plusieurs résultats possibles pour un même point de cassure -> très flexible
MR Lieber, Annu Rev Biochem, 2010-79;181

Réparation par fusion des extrémités
• Ligation finale
• DNA ligase IV ±XRCC4±XLF
• XRCC4 et XLF augmentent l’efficacité de DNA ligase IV sur des extrêmités non homologues
• «Cicatrice» au point de cassure
• Plusieurs résultats possibles pour un même point de cassure -> très flexible
• Création de diversité ( réarrangement V(D)J)
MR Lieber, Annu Rev Biochem, 2010-79;181

Choix du mécanisme de réparation
M Shrivastav et al., Cell Res, 2008-18;134

Choix du mécanisme de réparation
M Shrivastav et al., Cell Res, 2008-18;134
• Compétition entre HR et NHEJ
• HR : S et G2
• NHEJ : Tout le cycle mais essentiellement G0 et G1

Choix du mécanisme de réparation
M Shrivastav et al., Cell Res, 2008-18;134
• Compétition entre HR et NHEJ
• HR : S et G2
• NHEJ : Tout le cycle mais essentiellement G0 et G1
• Eléments de régulation
• Partenaire disponible pour HR ++
• Régulation de la formation d’ADN simple brin par CDK (Cycline Dependant Kinases)
• Protéines participant au «choix» : DNA Protein kinase, BRCA1, ATM

Et si il n’y a pas de deuxième extrémité ?
Cassure télomérique
PJ Hastings et al., Nat Rev Genetics, 2009-10;551

Et si il n’y a pas de deuxième extrémité ?
D Branzei et M Foiani, Nat Rev Mol Cell Biol, 2010-11;208
J Atkinson et P McGlynn, Nucleic Acids Res, 2009-37;3475
Arrêt de la fourche de réplication
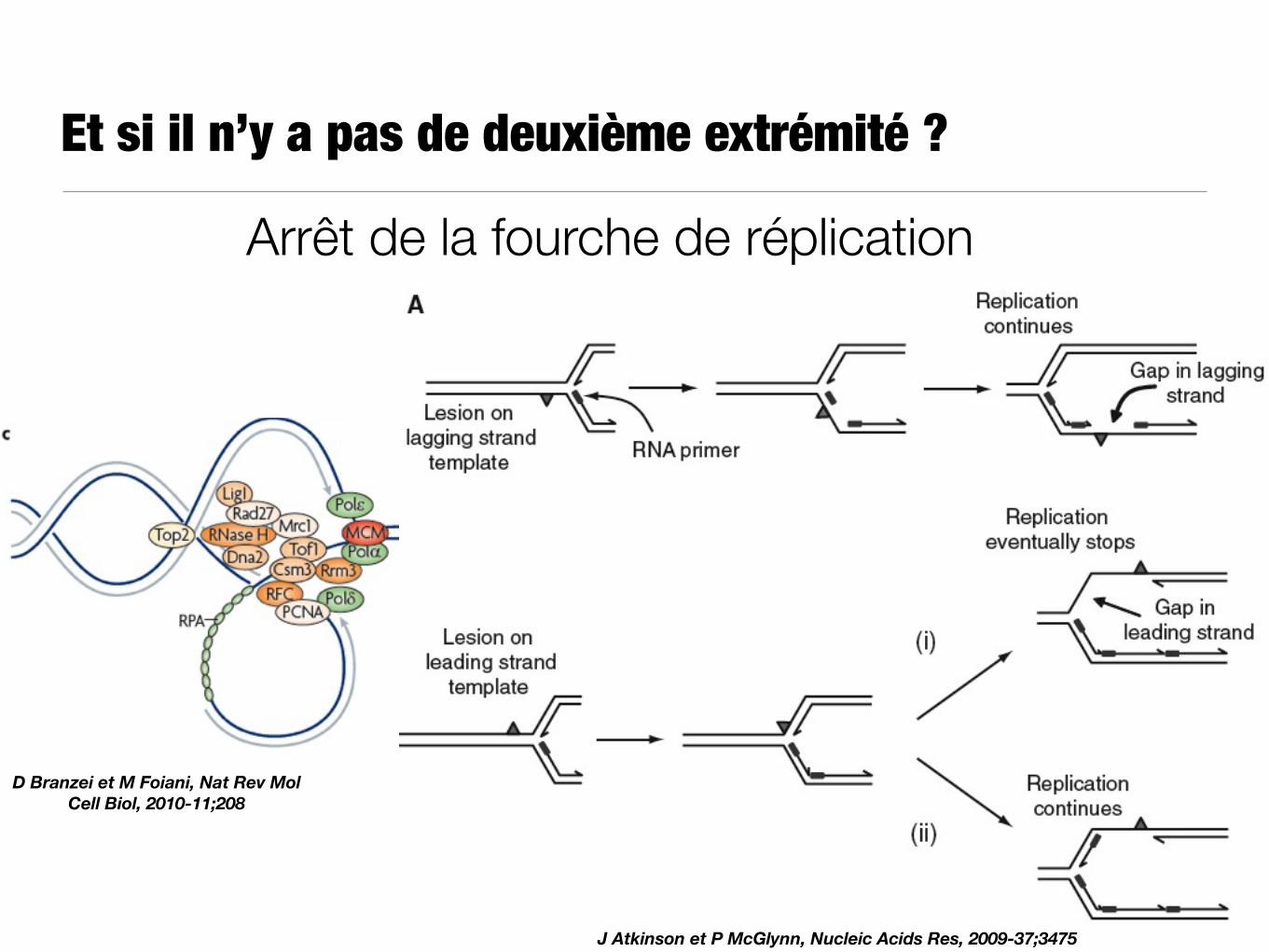
Et si il n’y a pas de deuxième extrémité ?
D Branzei et M Foiani, Nat Rev Mol Cell Biol, 2010-11;208
J Atkinson et P McGlynn, Nucleic Acids Res, 2009-37;3475
Arrêt de la fourche de réplication

Et si il n’y a pas de deuxième extrémité ?
D Branzei et M Foiani, Nat Rev Mol Cell Biol, 2010-11;208
J Atkinson et P McGlynn, Nucleic Acids Res, 2009-37;3475
Arrêt de la fourche de réplication

Levée du blocage par changement de matrice (FoSTeS : Fork Stalling and Template Switching)
Break Induced Replication
•Réplication en dehors d’un site initiateur de réplication
•Fourche relativement processive mais fréquent changement de matrice
•Taux de mutation élevé

Les Anomalies chromosomiques résultent d’une réparation anormale !
W Gu et al., PathoGenetics, 2008-I:4

Anomalies récurrentes : NAHR 2 fois plus de délétions que de duplications !
M Sasaki et al., Nat Rev Mol Cell Biol, 2010-11:182

Anomalies non récurrentes : NHEJ / BFB
• Organisation du génome dans le noyau Cassures
Machinerie de réparation
Intermédiaire de réparation
Translocation
K J Meaburn et al., Semin Cancer Biol 2007-17;80

Facteurs favorisants les anomalies de structure
• Organisation du génome dans le noyau
• Territoires chromosomiques

Facteurs favorisants les anomalies de structure
• Organisation du génome dans le noyau
• Territoires chromosomiques
• Juxtapositions de séquences éloignées
K J Meaburn et al., Semin Cancer Biol 2007-17;80

Breakage-first model
• Mobilité des fragments cassés
K J Meaburn et al., Semin Cancer Biol 2007-17;80
G1, RAD51- G2, RAD51+JA Aten et al., Science 2004-303;92

Contact-first modelK J Meaburn et al., Semin Cancer Biol
2007-17;80

• Association fonctionnelle de séquences éloignées
Contact-first modelK J Meaburn et al., Semin Cancer Biol
2007-17;80
T Cremer et al., Curr Opinion Cell Biol, 2006-18;307CS Osborne et al., Nature Genetics,
2004-36;1065
Kcnq1ot1 (chr7) hba (chr11)
hbb b1 (chr7) hbb b1 (chr7)

• Association fonctionnelle de séquences éloignées
• Corrélation entre volume de recouvrement et fréquence des translocations
Contact-first modelK J Meaburn et al., Semin Cancer Biol
2007-17;80
T Cremer et al., Curr Opinion Cell Biol, 2006-18;307
MR Branco and A Pombo, PLoS Biol, 2006-4;e138
CS Osborne et al., Nature Genetics, 2004-36;1065
Kcnq1ot1 (chr7) hba (chr11)
hbb b1 (chr7) hbb b1 (chr7)

Quid des anomalies complexes ?• Pelizaeus-Merzbacher
• Liée à l’X, gène PLP1Proteolipid Protein 1
• Non récurrente
P J Hastings et al., PLoS Genetics, 2009-5:e1000327
DuplicationDélétionTriplicationDiploïdie

Clonage des points de cassure
• Réarrangement complexe
• Zones tripliquées au sein de zones dupliquées
• Zones non dupliquées intercalées
• Microhomologies au points de cassure
J A Lee et al., Cell, 2007-131:1235

Erreurs de réplication : FoSTeS / MMBIR (Microhomology Mediated Break Induced Reparation)
P J Hastings et al., Nat Review Genetics, 2009-10:551
J A Lee et al., Cell, 2007-131:1235

Facteurs favorisant les remaniements
R-S Mani & A M Chnnaiyan., Nature Rev Genetics 2010-11;819

Facteurs favorisant les remaniements
• Proximité des molécules d’ADN
• Fréquence des cassures double brin
• Erreurs de réparation

Facteurs favorisant les remaniements
• Proximité des molécules d’ADN
• Territoires chromosomiques : taille, contenu en gènes, recouvrement
• Zones de transcription (Transcription factories), différenciation cellulaire
• Chronologie de réplication
• Translocations majoritairement entre séquences se répliquant en même temps
• Fréquence des cassures double brin
• Erreurs de réparation
E Gaffe, PLoS Genet, 2010-6;e1001011

Facteurs favorisant les remaniements
Triplex DNA Hairpins Quadruplex DNA
Cruciform DNA Left-handed Z-DNA
L Visser et al, Hum Mol Genet 2009-18;3579
• Proximité des molécules d’ADN
• Fréquence des cassures double brin
• Stress réplicatif
• Non-B DNA
• Erreurs de réparation

Facteurs favorisant les remaniements
• Proximité des molécules d’ADN
• Fréquence des cassures double brin
• Stress réplicatif
• Non-B DNA
• Séquences répétées
• Erreurs de réparation

Facteurs favorisant les remaniements
• Proximité des molécules d’ADN
• Fréquence des cassures double brin
• Stress réplicatif
• Non-B DNA
• Séquences répétées
• Erreurs de réparation
I Voineagu et al., PNAS 2008-105;9936

Protection contre le stress réplicatif
J Atkinson et P McGlynn, Nucleic Acids Res, 2009-37;3475
• Régression de la fourche de réplication
• DNA supercoiling
• Hélicases et Translocases
• BLM, WRN (Werner syndrome) : suppriment les super tours
• FANCM : component of the Fanconi Anaemia core complex
• Protéines de recombinaison
• RecA / RAD51

Protection contre le stress réplicatif
J Atkinson et P McGlynn, Nucleic Acids Res, 2009-37;3475
Régression puis digestion de l’ADN double brin néo formé

Protection contre le stress réplicatif
J Atkinson et P McGlynn, Nucleic Acids Res, 2009-37;3475
Régression puis recombinaison pour passer la lésion
Régression puis digestion de l’ADN double brin néo formé

Cas de la t(11;22)(q23;q11)
• Palindromes en 11q 23 et 22q11
• ADN cruciforme
• Cassure double brin
• Réplication ? Autre ?
• Réparation par NHEJ
• Homologie qq nucléotides
• petite délétion au point de cassure

Facteurs favorisant les remaniements
• Proximité des molécules d’ADN
• Fréquence des cassures double brin
• Erreurs de réparation
• Structure du génome favorise les erreurs de recombinaison
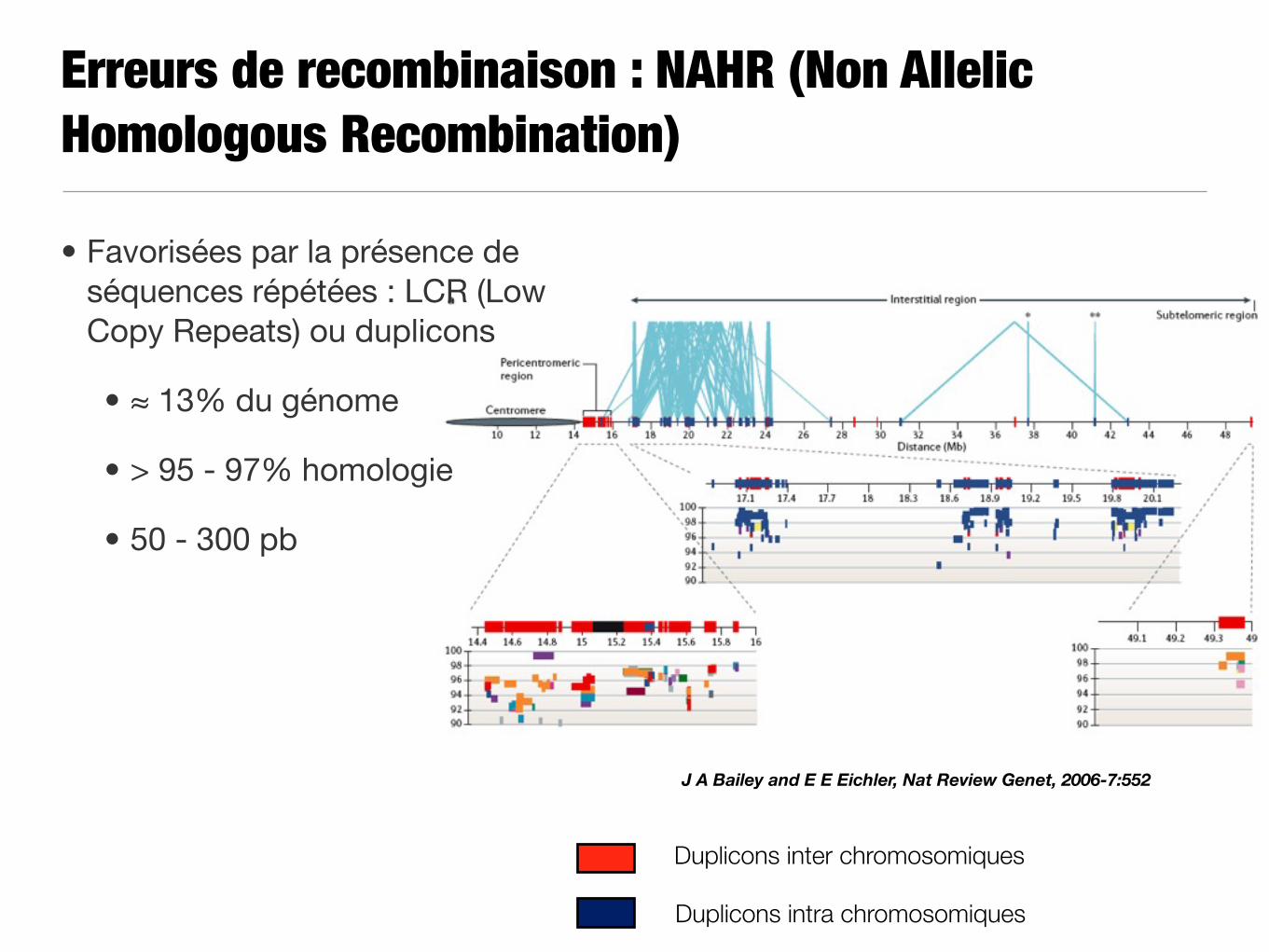
Erreurs de recombinaison : NAHR (Non Allelic Homologous Recombination)
J A Bailey and E E Eichler, Nat Review Genet, 2006-7:552
• Favorisées par la présence de séquences répétées : LCR (Low Copy Repeats) ou duplicons
• ≈ 13% du génome
• > 95 - 97% homologie
• 50 - 300 pb
Duplicons inter chromosomiques
Duplicons intra chromosomiques

Erreurs de recombinaison : NAHR (Non Allelic Homologous Recombination)
• Favorisées par la présence de séquences répétées : LCR (Low Copy Repeats) ou duplicons
• ≈ 13% du génome
• > 95 - 97% homologie
• 50 - 300 pb
• Hotspots recombinaison non allélique et recombinaison méiotique ont des positions semblables
M Sasaki et al., Nat Rev Mol Cell Biol, 2010-11:182

Déterminants de la fréquence des NAHR

Déterminants de la fréquence des NAHR
• Taille des duplicons
• Zone d’homologie divisée par deux (232 à 134 nucléotides) -> réduction par 20 de la fréquence de NAHR

Déterminants de la fréquence des NAHR
• Taille des duplicons
• Zone d’homologie divisée par deux (232 à 134 nucléotides) -> réduction par 20 de la fréquence de NAHR
• Structure locale du génome
• Palindrome, ADN non B, transposon…

Déterminants de la fréquence des NAHR
• Taille des duplicons
• Zone d’homologie divisée par deux (232 à 134 nucléotides) -> réduction par 20 de la fréquence de NAHR
• Structure locale du génome
• Palindrome, ADN non B, transposon…
• Contexte de survenue
• Pour un même LCR, fréquence de NAHR peu varier entre mitose et méiose
• Fréquence variable de NAHR pour certains loci en fonction du sexe

Mécanismes de protection contre les NAHR

Mécanismes de protection contre les NAHR
• Inhibition des cassures au niveau des séquences répétées
• Homologie de séquence ≈ parfaite pour autoriser la recombinaison

Mécanismes de protection contre les NAHR
M Sasaki et al., Nat Review Mol Cell Biol 2010-11;182
• Inhibition des cassures au niveau des séquences répétées
• Homologie de séquence ≈ parfaite pour autoriser la recombinaison
• Utilisation préférentielle de l’homologue

Mécanismes de protection contre les NAHR
M Sasaki et al., Nat Review Mol Cell Biol 2010-11;182
• Inhibition des cassures au niveau des séquences répétées
• Homologie de séquence ≈ parfaite pour autoriser la recombinaison
• Utilisation préférentielle de l’homologue
• Contrôle de la qualité de l’homologie (RAD51 - ssDNA)

Facteurs favorisant les remaniements
• Proximité des molécules d’ADN
• Fréquence des cassures double brin
• Erreurs de réparation
• Structure du génome favorise les erreurs de recombinaison
• Rôle du stress cellulaire

Stress cellulaire et défaut de recombinaison
ME Moynahan, M Jasin, Nat Rev Mol Cell Biol, 2010-11;196
ATP

Stress cellulaire et défaut de recombinaison
• Mutation des protéines de réparation,
• Stress cellulaire
• Réduction RAD51ME Moynahan, M Jasin, Nat Rev Mol Cell Biol, 2010-11;196
ATP

Le stress cellulaire peut entraîner un déficit en RAD51

Le stress cellulaire peut entraîner un déficit en RAD51
• Mécanismes réplicatifs
• BIR : Homologie étendue (> 50 pb)
• MMBIR : Microhomologie de quelques nucléotides suffit

Le stress cellulaire peut entraîner un déficit en RAD51
• Mécanismes réplicatifs
• BIR : Homologie étendue (> 50 pb)
• MMBIR : Microhomologie de quelques nucléotides suffit
• BIR nécessite RAD51 pour l’invasion du brin homologue
• MMBIR : mécanisme alternatif impliquant RAD52
T Sugiyama et N Kantake, J Mpl Biol 2009-390;45
MMBIRBIR

Le stress cellulaire peut entraîner un déficit en RAD51
• En l’absence d’invasion d’un ADN double brin homologue :
• Réparation possible par Single-Strand Annealing (SSA) - RAD51 non nécessaire
• Résection 5‘ -> 3’ poursuivie
• Révélation de séquences homologues
• Appariement et résection des extrémités non appariées
• Ligation -> délétion entre les séquences répétées
• Mécanisme de délétion entre séquences AluPJ Hastings et al., Nat Rev Genetics, 2009-10;551

Le stress cellulaire peut entraîner un déficit en Cohésines
M Carratero et al. Curr Op Cell Biol, 2010
K Shintomi & T Hirano Chromosoma, 2010
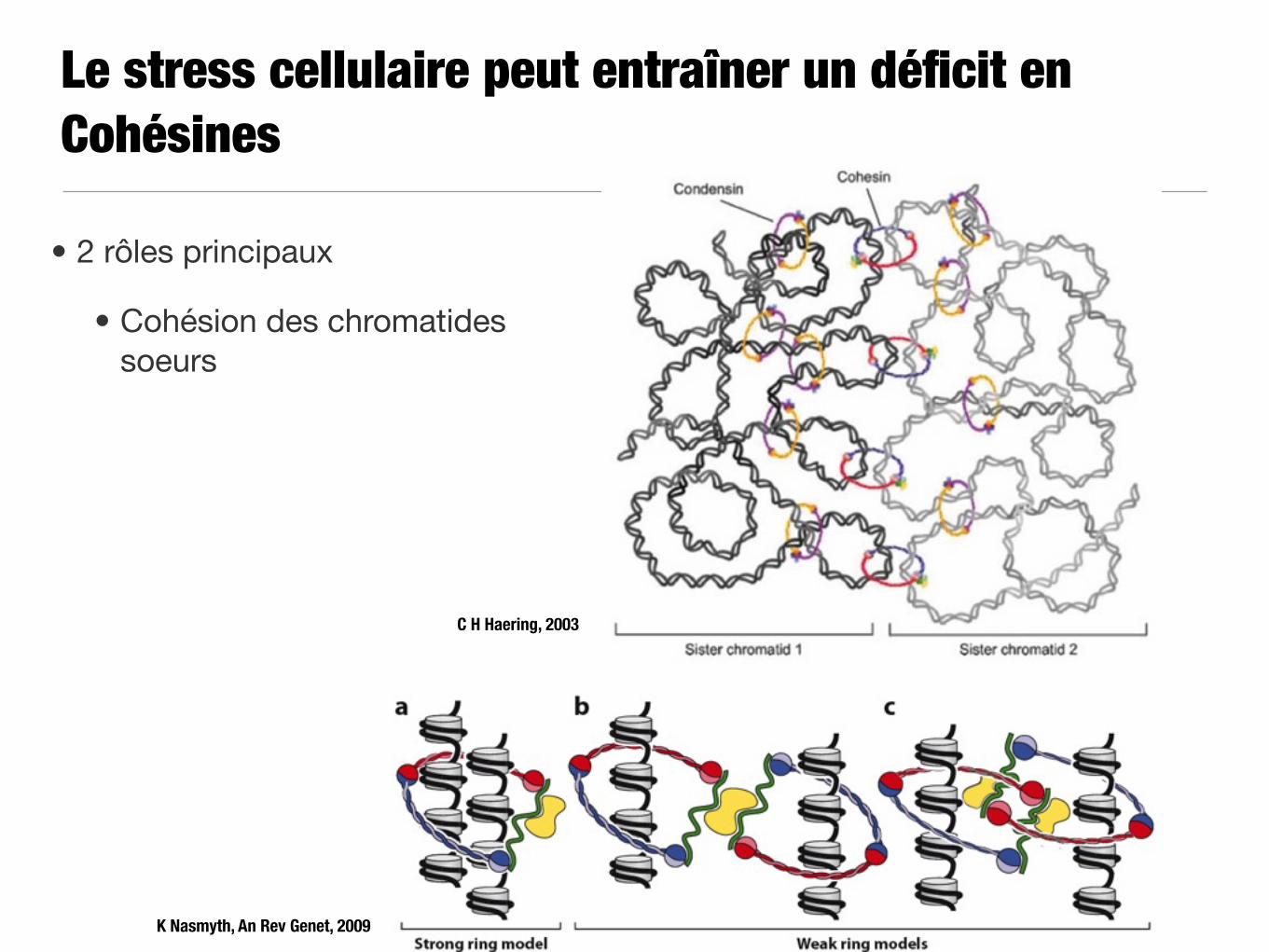
Le stress cellulaire peut entraîner un déficit en Cohésines
• 2 rôles principaux
• Cohésion des chromatides soeurs
C H Haering, 2003
K Nasmyth, An Rev Genet, 2009

Le stress cellulaire peut entraîner un déficit en Cohésines
• 2 rôles principaux
• Cohésion des chromatides soeurs
• Organisation du génome / Contrôle transcriptionnel en association avec CTCF
E Nunez et al., Curr Op Genet Dev, 2009-19;424

Le stress cellulaire peut entraîner un déficit en Cohésines
S Covo et al., PLoS Genetic 2010-e1001006
• Mutation des protéines de réparation,
• Stress cellulaire
• Réduction RAD51
• Déficit en cohésines
• Favorisent la recombinaison entre chromatides soeurs
• Suppriment la recombinaison entre homologues

Conclusion
Anomalies complexes
Cas sure

Conclusion
HR
NHEJBIR
Anomalies complexes
Cicatrice au point de cassure
Cas sure
Restauration fidèle

Conclusion
HR
NHEJBIR
CompétitionProtéines régulatrices : BRCA1
Cycle cellulaire
Anomalies complexes
Cicatrice au point de cassure
Cas sure
Restauration fidèle
Conclusion
HR
NHEJBIR
CompétitionProtéines régulatrices : BRCA1
Cycle cellulaire
Anomalies complexes
Cicatrice au point de cassure
Cas sure
Restauration fidèle
Position dans le noyau
Structure du génomeEnvironnement cellulaire

Conclusion
HR
NHEJBIR
NAHR
CompétitionProtéines régulatrices : BRCA1
Anomalies récurrentesmais pas seulement (ALU, LINE …)
Cycle cellulaire
Anomalies complexes
Cicatrice au point de cassure
LCR
Cas sure
Restauration fidèle
Position dans le noyau
Structure du génomeEnvironnement cellulaire

Conclusion
HR
NHEJBIR
NAHR
FoSTeS / MMBIR
CompétitionProtéines régulatrices : BRCA1
Anomalies récurrentesmais pas seulement (ALU, LINE …)
Cycle cellulaire
Stress cellulaire
Anomalies complexes
Anomalies complexes
Cicatrice au point de cassure
LCR
Cas sure
Restauration fidèle
Position dans le noyau
Structure du génomeEnvironnement cellulaire

Mais comment expliquer çà ??
Stephens, P. J. et al. Cell 144, 27-40 (2011).

Hypothèse d’un évènement catastrophique = chromothripsis
• Concernerait 2 à 3% des cancers
• jusqu’à 25% des cancers osseux
• Réparation par les mécanismes habituels (MMBIR / NHEJ)
• Très nombreux réarrangements créés en même temps

Accumulation d’anomalies ou chromothripsis ?
• Les cassures ne sont pas réparties aléatoirement
• Clustering sur un chromosome (ou une partie)
• Mais partenaires peuvent être distants
• Il n’y a pas de duplications, soit 1 copie, soit deux copies
Stephens, P. J. et al. Cell 144, 27-40 (2011).

Mécanisme ?
• Soit radiation ionisante / radicaux libres
• sur un chromosome en cours de condensation -> clustering des anomalies au niveau d’un bras, d’une bande…
• Soit un effet de l’attrition télomérique
• BFB pendant la cytokinèse
• Dans tous les cas remaniement en profondeur du génome
• En général mort cellulaire
• Sinon, très nombreux réarrangements entraînent
• déséquilibre d’expression génique (➘, fusion, ➚ (chromosomes minutes))
• avantage prolifératif et cancérisation

Le phénomène de chromthripsis peut-il expliquer des remaniements constitutionnels (très) complexes ?
• Réarrangements complexes
• localisés
• associent del / dup/ trp
• Pas de mosaicisme différent entre les différentes anomalies
Liu, P., et al. Cell 146, 889-903 (2011).

Mécanisme différent ?
• Eléments en faveur d’un mécanisme réplicatif type BIR
• Duplications /Triplications
• Microhomologies aux points de cassure
Liu, P., et al. Cell 146, 889-903 (2011).

Bibliographie
• A DNA Replication Mechanism for Generating Nonrecurrent Rearrangements Associated with Genomic Disorders. J A. Lee et al., Cell, 2007, 131:1235
• Regulation of DNA double-strand break repair pathway choice. M Shrivastav et al., Cell Resarch, 2008, 18:134
• Mechanisms of change in gene copy number. PJ Hastings et al, Nature review Genetics, 2009, 10:551
• The mechanism of double-strand DNA break repair by the Nonhomologous DNA end-joining pathway. MR Lieber, Annu Rev Biochem, 2010, 79:181
• Triggers for genomic rearrangements:insights into genomic, cellular and environmental influences.R-S Mani et al., Nature reviews Genetics, 2010, 11:819
• Mitotic homologous recombination maintains genomic stability and suppresses tumorigenesis. ME Moynahan et al., Nature reviews Molecular Cell Biology, 2010, 11:197
• Stephens, P. J., Greenman, C. D., Fu, B., Yang, F., et al. Massive genomic rearrangement acquired in a single catastrophic event during cancer development. Cell 144, 27-40 (2011).