localized alterations in pre- and postsynaptic serotonin binding sites in the ventrolateral...
TRANSCRIPT

BRAIN RESEARCH
E L S E V I E R Brain Research 688 (1995) 121-133
Research report
Localized alterations in pre- and postsynaptic serotonin binding sites in the ventrolateral prefrontal cortex of suicide victims
Victoria Arango a,* Mark D. Underwood a, Ananda V. Gubbi b, j. John Mann ~ Department of Neuroscience and the NIMH Clinical Research Center for the Study of Suicidal Behat,ior, New York State Psychiatric Institute, Columbia
UniL,ersity College of Physicians and Surgeons, New York, NY, USA Department of Psychiatry, Western Psychiatric Institute and Clinic, Unil,ersity of Pittsburgh, Pittsburgh, PA, USA
Acceptcd 18 April 1995
Abstract
Altered serotonin indices have been reported in the brain of suicide victims. We sought to localize the changes in presynaptic and postsynaptic serotonin receptors and identify an area of prefrontal cortex that may influence suicide risk. Quantitative autoradiography was performed in coronal sections of prefrontal cortex to determine whether serotonin 5-HT~A receptor (postsynaptic in cortex) and serotonin transporter (presynaptic) binding are different in suicide victims compared to matched controls. 5-HTIA receptor binding was higher in 85 of the 103 sampled areas in the suicide group (n = 18 pairs; P < 0.0001). The increase ranged from 17 to 30%. The increase was more pronounced in the ventrolateral prefrontal cortex. Serotonin transporter binding was found to be lower in the suicide group in all but one of the 43 sampled regions (n = 22 pairs; P < 0.0001). The reduction in binding was most pronounced in the ventrolateral prefrontal cortex, where the difference between suicides and controls ranged between 15 and 27%. Serotonin transporter and 5-HTIA binding were negatively correlated (r = - 0.35 to - 0.44, P = 0.04 to 0.007) within the same brain areas, suggesting common regulatory factors with opposite effects on binding to the two receptors. We conclude that suicide victims have an abnormality in the serotonin ,system involving predominantly the ventrolateral prefrontal cortex, and hypothesize that the serotonergic dysfunction in this brain region contributes to the risk for suicidal behavior.
Keywords: Quantitative receptor autoradiography; [3H]8-OH-DPAT; [3 H]Cyanoimipramine; 5-HT1A receptor; Serotonin transporter; HPLC: 5-HIAA
1. Introduct ion
Suicide claims more than 30,000 lives a year in the United States [20]. There is evidence suggesting that there are alterations in the serotonergic system in the brain of suicide victims and related altered serotonin (5-hydroxy- tryptamine, 5-HT) system indices in patients who have made serious suicide attempts [9,10,12,13,17,48,60,68]. Such findings have led to the hypothesis that reduced serotonergic activity is associated with increased suicide risk [50,53]. This reduction in serotonergic activity corre- lates with suicidal acts in several diagnostic groups includ-
" Presented in part at the Society for Neuroscience Annual Meetings [3,4,69].
* Corresponding author. Department of Neuroscience, New York State Psychiatric Institute, Columbia University College of Physicians and Surgeons, 722 West 168th Street, Box 28, New York, NY 10032. USA. Fax: (1) (212) 781-0503. E-mail: [email protected]
0006-8993/95/$09.50 © 1995 Elsevier Science B.V. All rights reserved SSDI 0 0 0 6 - 8 9 9 3 ( 9 5 ) 0 0 5 2 3 - 4
ing major depression, schizophrenia and personality disor- ders. These diagnostic groups make up the vast majority of suicides not associated with alcohol or drug abuse. Thus, this serotonin deficiency was not restricted to a specific diagnosis.
Studies of the serotonergic system fall into two major domains. The first involves studies of serotonin and metabolites in postmortem brain samples from suicide completers and in cerebrospinal fluid (CSF) of suicide attempters. Levels of serotonin and its metabolite 5-hy- droxyindoleacetic acid (5-HIAA), have been found in most studies to be lower in the brainstem of suicide victims compared to controls (see [2] for review), although we find no reductions in cerebral cortex [43]. Lower levels of CSF 5 -HIAA have been reported in suicide attempters com- pared to nonattempters [10,11,51,76], and a blunted release of prolactin in response to the indirect serotonin agonist fenfluramine has been observed in suicide attempters [22,51,52].

122 V. Arango et al. /Brain Research 688 (1995) 121-133
The second domain of study involves assessment of serotonin receptors. Serotonin receptors were originally classified as 5-HT 1 and 5-HT 2 [65]. Subtypes of 5-HT~ receptors, namely 5-HTIA and 5-HTI~, were soon pro- posed [32,64]. More recently, other 5-HT 1 receptor sub- types as well as 5-HT 3 [14,40], 5-HT 4 [28] and other [15,75] receptor subtypes have been reported. We previ- ously found that 5-HT 2 sites, as labeled by 125I-lysergic acid diethylamide ([125I]LSD) [1] or [3H]spiroperidol [54], are increased in the brain of suicide victims, compared to controls. We also found that 5-HT~ sites, as labeled by [3H]5-HT, are unaltered [54]. The 5-HT1A receptor, a postsynaptic serotonin receptor in the cerebral cortex, is of particular interest because of recent reports that 5-HT~A binding is increased in suicide victims [38,56], as well as its possible involvement in anxiety and affective disorders as suggested by the anxiolytic and antidepressant proper- ties of partial 5-HTIA agonists [74].
An index of the serotonin nerve terminal integrity may be obtained from binding studies of the serotonin trans- porter, which may be a better indicator of the abnormali- ties associated with suicide than cortical levels of serotonin or 5-HIAA [43]. Not all studies agree as to whether binding to the serotonin transporter on nerve terminals is reduced in cortical regions of suicide victims [2]. However, most studies of the serotonin transporter in suicide victims have employed [3H]imipramine as the ligand, which has been shown to bind to both high- and low-affinity sites [19,37], and to nonserotonergic receptors such as mus- carinic cholinergic sites [24]. Only the high-affinity site represents binding to the serotonin transporter [24]. In contrast, [3H]paroxetine [34] and [3H]cyanoimipramine ([3H]CN-IMI) bind to a single class of binding site [27,35,42], the high-affinity site. Studies of [3H]imipra- mine in homogenates from frontal cortex are divided in that about half found a decrease in the suicide group and the remainder found no change (see [2] for review). Three studies have used [3H]paroxetine as the ligand and two of those [45,46] reported no changes in the frontal cortex (Brodmann area 10) of depressed suicide victims com- pared to controls. The third study [36] was from our laboratory and found a 40% reduction in drug-free suicide victims.
The above studies suggest that there are serotonin re- ceptor alterations in the brain of suicide victims. Most studies examined homogenates from prefrontal cortex and provide little anatomical information as to the extent and localization of serotonin receptor changes in suicide. We therefore sought to localize where, within the prefrontal cortex of suicide victims, presynaptic (serotonin trans- porter) and postsynaptic (5-HT1A) serotonin receptor changes are present, and whether these changes in pre- and postsynaptic binding sites are colocalized.
We used the selective ligands [3H]8-hydroxy-2-(di-n- propyl-amino)-tetralin ([3H]8-OH-DPAT) [32] and [3H]CN-IMI [42] to perform quantitative receptor auto-
radiography of 5-HTIA and serotonin transporter binding, respectively, in adjacent sections of tissue taken from the prefrontal cortex of suicide victims and matched controls. This approach allowed mapping of binding sites across multiple regions of the prefrontal cortex and the compari- son of the precise localization of changes associated with suicide.
2. Materials and methods
Tissue was provided by local Coroner or Medical Ex- aminer offices in accordance with city or county regula- tions and the protocol was approved by the Institutional Review Board for Biomedical Research. An essential as- pect of the methods involved performing the assays for the two binding site populations (5-HT)A and serotonin trans- porter) on adjacent tissue sections cut from the same block of tissue from each case, in order to permit maximal comparability for assessing the degree of colocalization of changes in binding to the two sites.
We employed a matched-pairs design. The cause of death was determined by the Coroner or Medical Exam- iner. Each suicide case (n = 22) was matched with a control subject (n = 22) on the basis of postmortem inter- val (PMI, +5 h), age (___ 5 years), sex, season of death, side of brain and, whenever possible, race. The two groups were tested for differences in age, sex and race. The results are described below. The brain samples were coded and assayed together by personnel blind to the cause of death. Individuals with a history of cerebral trauma, central ner- vous system disease, chronic alcoholism, illicit or thera- peutic drug use or AIDS were excluded. Body fluids (blood, bile, aqueous humor and urine) underwent toxico- logical screening for cocaine, opiates, alcohol and other acidic and basic drugs.
2.1. Collection, dissection and storage o f brain samples'
Brains were collected and bisected at autopsy. The right hemispheres were cut coronally into 1.5-cm-thick sections using a Lipshaw macrotome (Lipshaw Corp., Detroit, MI). Blocks were placed on a glass slide, immersed in freon ( - 20°C) and stored at -80°C until sectioning. Samples from the left hemisphere were saved for neuropathological and toxicological analyses. After a control and suicide were matched, tissue was sectioned at 20 /~m with a large body Lipshaw cryotome (1800-N Research model, Lip- shaw Corp., Detroit, MI, - 1 4 to -18°C). Coronal sec- tions from the entire hemicerebrum were taken from a level just anterior to the genu of the corpus callosum. This level was chosen because it contains prefrontal cortical areas believed to be involved in higher cognitive functions, affect and emotion, as well as in the temporal organization of behavior [30]. Sections (n = 100) were collected serially and thaw-mounted onto 3"× 5" gelatin-subbed, acid-

v. Arango et al. / Brain Research 688 (1995) 121-133 123
cleaned glass slides. An intercalating set (every 200 /zm) was stained for Nissl substance. Six sections per brain were used for each receptor assay. This collection protocol allowed for the study of multiple receptor populations in essentially identical brain cortical areas. Tissue sections were desiccated and stored in sealed boxes (at -20°C for 24 h, and -80°C thereafter).
2.2. In Htro receptor autoradiography
Association / Dissociation / Competition Experiments To obtain optimal assay conditions, association, dissoci-
ation and competition curves were obtained (in quadrupli- cate) using slide-mounted rat brain sections prior to begin- ning the human experiments. Sections were wiped from the slide with filter discs (Whatman GF/B) and counted in a liquid scintillation spectrophotometer (Packard Instru- ment Co., Meriden, CT). Binding of [3H]8-OH-DPAT reached equilibrium in 60 min 01/,_ = 7 min), and the best specific/nonspecific ratio was achieved following a 10 min wash. Competition curves with 12 concentrations of serotonin (100 pM to 100 /xM) determined the IC5o to be 3.16 nM. Binding of [3H]CN-IMI reached equilibrium at 24 h (t~/2 = 4 h) and the IC5o for sertraline was 36 nM. The optimum wash time was determined to be 60 min.
General procedure Dry sections from one matched pair were surrounded
with dental wax (Miles Inc., South Bend, IN) and incu- bated with 8-10 ml of ligand in a humidity chamber. Washes were done in large plexiglass jars, thus ensuring the presence of excess buffer to remove nonspecific bind- ing. All buffers were isosmotic to preserve the integrity of the tissue. Following the washes, the slides were dipped in distilled H20 (4°C), the tissue was dried with cold, filtered air and vacuum desiccated overnight at 4°C. Dried sections from a matched pair were arranged in X-ray cassettes along with 3H-standards (ARC-138, American Radiola- beled Chemicals Inc., St. Louis, MO) and exposed to ~H-sensitive film (Ultrofilm, Amersham Co., Arlington Heights, [L) for 8 and 20 weeks for [3H]8-OH-DPAT and [3H]CN-IMI, respectively. At the end of the exposure period, the films were developed according to the manu- facturer's protocol (Eastman Kodak Co., Rochester, NY).
[ 3H]8-OH-DPA T binding Tissue sections were preincubated (30 min, 23°C) in
170 mM Tris-HC1 containing 4 mM CaCI 2 and 138 mM sucrose (pH 7.6). Incubations were carried out in the same buffer containing 2 nM [3H]OH-DPAT and 0.01% ascor- bic acid at 21°C for 60 min. Parallel incubations containing 1 /J.M 5-HT (oxalate salt) defined nonspecific binding, which was less than 10% of total binding. Sertraline (10 nM) was added to incubations in order to mask binding to serotonin uptake sites [29]. The addition of sertraline at this concentration (10 nM) did not affect binding to 5-HT]A
sites. Sections were washed for l0 rain (2 × 5 rain) in Tris-HCl buffer at 4°C.
[3H]CN-IM1 binding Sections were preincubated (30 rain, 23°C) in 50 mM
Tris-HCl buffer with 130 mM NaCI and 5 mM KCI (pH 7.4). Total binding was determined by incubation (4°C) in the same buffer containing 0.4 nM [3H]CN-IMI for 24 h. Nonspecific binding was defined by coincubation of adja- cent sections with 1 /xM sertraline (modification of [42]). Nonspecific binding was less than 20%. Sections were washed in cold buffer for 1 h (3 × 20 min).
Quantitation of autoradiograms Autoradiograms were analyzed using a PC-based image
analysis system (Imaging Research, Inc., St. Catherine, Ont., Canada). Films were digitized, shade corrected and calibrated against tritium standards (ARC. St. Louis, MO). Histological examination of Nissl-stained sections was used in the determination of the boundaries of cortical layers and their correspondence to isodensity bands in the auto- radiograms. Brodmann areas 8, 9, 46, 45, 47, 11, 12, 32 and 24 were identified using gyral and sulcal landmarks, cytoarchitecture and a standardized coronal atlas (Robert Perry and Edward Bird, personal communication). Images from sections incubated in the presence of the radioligand alone (total binding) were acquired first. Images produced from incubation in the presence of displacer (nonspecific binding) were then acquired and digitally subtracted from the total binding. We thus measured specific binding, in fmol /mg tissue, for each receptor. Measurements of bind- ing were taken from each of the individual isodensity bands, as well as across the full width of the cortical gray matter, from both the gyrus and sulcus. Measurements were taken from all areas available in the three resulting autoradiograms per brain. Multiple readings from each region were averaged. In instances where gray matter was discontinuous, measurements excluded missing tissue. Binding to white matter was taken as the average of multiple measurements throughout the sections. Standards were corrected daily for radioactive decay.
2.3. High pressure liquid chromatography (HPLC) analy- sis of serotonin, 5-HIAA, 5-hydroxytryptophan (5-HTP) and t.-tryptophan (L-try) le~'els
A modification of the method of Korpi and colleagues [41] was used for determination of the levels of serotonin, its precursors (5-HTP, L-try) and principal metabolite (5- HIAA). Tissue was sonicated in 0.1 M perchloric acid (4°C) containing N-methyl-5-hydroxytryptamine (NMET, 0.1 /xM) as the internal standard. An aliquot of the ho- mogenate was used for protein analysis [49], and the remaining homogenate was centrifuged at 10,000 X g (8 min, 4°C). Supernatants were diluted 2 fold with cold 0.1 M perchloric acid and injected into the HPLC column.

124 v. Arango et al. /Brain Research 688 (1995) 121-133
The HPLC consisted of a Waters M-501 high pressure pump, a WISP auto sampler, a Rainin Dynamax 5 p~M C18 reverse-phase analytical and guard columns and a Waters 460 electrochemical detector. The mobile phase consisted of 75 mM sodium phosphate buffer (pH 3.0) containing 7.5% acetonitrile (v /v ) , 0.9 mM 1-octane sul- fonic acid and 5 0 / z M disodium EDTA. The flow rate was 1.0 ml /min . Working electrode potential was kept at + 0.8V against an Ag / AgC1 reference electrode. Typical retention times (K") for 5-HTP, 5-HIAA, serotonin, L-try and NMET were 5.9 min, 11.4 min, 20.5 min, 24 min and 25.2 min, respectively. All data were expressed as pmol per mg protein.
Data were acquired and analyzed using a Waters Base- line 810 software package (Dynamic Solutions, Millipore, Milford, MA). Both external and internal standards were used for quantitation and data were calculated based on sample peak area relative to internal standard peak area. The standard curves were based on external standards
which encompassed the concentrations of the unknowns or samples.
2.4. S ta t i s t i ca l me thods
We employed a multivariate testing procedure, namely a forward, stepwise moving, logistic multivariate regres- sion model. This procedure was used not only to pick out the significant variables that explain the logistic model, but also to identify the distinctive profiles of the dichotomous dependent variable. This analysis was used to separately study differences in group, sex and dichotomized age, for binding to the 5-HTIA and the serotonin transporter recep- tors. Post-regression analyses were carried out, using paired t-tests for the significant variables. The descriptive statis- tics on these variables and correlations were included to support and complement the analysis. SPSSWlNDOWS (6.0) was used to analyze the data.
Binding to 5-HTjA sites in each brain was examined in
Fig. 1. Pseudocolor images of subtracted autoradiograms representing total specific 5-HTIA (A, left panel) and serotonin transporter (B, right panel) binding sites in adjacent coronal sections of the prefrontal cortex of the human. Note that: (1) the distribution of 5-HTIA receptors is laminar and most of the binding is localized in a band overlaying cortical layer II in all cortical areas; (2) Binding to the serotonin transporter is higher in the outer cortical layers and densest in the medial prefrontal cortex; (3) binding to the gray matter is much greater than to the white matter for both pre-and postsynaptic sites.

V. Arango et al. / Brain Research 688 (1995) 121-133 125
103 bands involving nine Brodmann areas (sulcus and gyrus) and 18 pairs were included in the analysis. The natural logarithms of these 103 measures were taken to reduce skewness, and constituted the variables to be tested for 5-HTIA differences. Binding to the serotonin trans- porter was studied in 43 bands, using data on 22 matched pairs of subjects. The binding levels in these 43 brain regions were not transformed as there was little skewness.
Correlations with age, PMI and freezer storage time were calculated using the Pearson's coefficient of correla- tion. Missing cells were replaced by their corresponding subgroup means for each of the receptor populations. Results are reported as mean + standard error of the mean. All P values are two-tailed. Alpha was preset at P < 0.05.
2.5. Subjects
Demographic variables from suicide and control sub- jects were similar. The mean age of the suicide group was 41.5 + 4.14 y (14-78) compared to 41.3 + 4.4 years (15 - 79) for the controls (t = 0.23, 21 df, P = 0.98). The PMI of the suicide group was 14.2 + 0.3 h (4 -22 h) compared to 13.9 + 0.2 h (4-22) for the controls (t = 0.83, 21 df, P = 0.78). The ratio of men to women was 14:8 in each group. The mean freezer storage time (time from tissue collection to assay) for samples assayed for 5-HTIA bind- ing in the suicide group was 518.8 + 55.5 days compared to 525.5 + 47.5 days for the control group (t = 0.14, 17 df, P = 0.82). Likewise, the storage time for samples assayed for the serotonin transporter was not different in the two groups (622.6 + 40.1 d vs. 587.5 + 31.2 days; t = 0.82, 21 df, P = 0.42). Causes of death in the suicide group were: hanging (10), fall from height (6), firearm (4), subway (1) and ingestion of lye (1). In the controls, causes of death were: fall (3), motor vehicle accident (8), firearm (3), cardiac failure (3), lacerations (2), explosion (1), aneurism (1) and respiratory failure (1). These criteria for matching pairs enabled us to limit possible variance due to PMI, sex, storage and age, that may affect receptor binding. Assay- ing the matched pairs together reduced interassay variance effects.
2.6. Drugs and reagents
[ 3 H]8-OH-DPAT ( 125-162 C i / m m o l ) and [ 3 H]CN-IMI (78.8-83.6 C i / m m o l ) were purchased from New England Nuclear (Boston, MA), stored in ethanol at - 8 0 ° C in the dark and diluted in buffer immediately prior to assay. Sertraline-HCl was generously supplied by Pfizer Pharma- ceuticals (Groton, CT). All other chemicals were pur- chased from Sigma Chemical Co (St. Louis, MO).
3. Results
Gyral and sulcal cortex exhibit different c y t o - a n d myeloarchitectonic characteristics [79], with further differ-
60
69 69
4O
Controls (n=l 8)
Suicides (n=18)
. p < 0.05
O3
e"
~5 < 12.
± o, ~, 20
I II III Ill-IV V-VI w/bite
Cortical Layers
Fig. 2. Laminar distribution of [~H]8-OH-DPAT binding in the sulcus of Brodmann area 46 in suicides vs. controls. Bar graph comparing thc specific binding of [3H]8-OH-DPAT to serotonin 5-HTIA receptors be- tween the suicide and control groups in layers of the lateral prefrontal cortex (sulcus of Brodmann area 46). Specific binding is expressed in fmol/mg tissue +_ S.E.M. for 18 subjects in cach group. Data for each subject is thc mean value obtained from multiple readings in 3 subtracted autoradiograms from 6 slide-mounted large coronal sections. Binding to this Brodmann area formed 5 isodensity bands and is representative of all Brodmann areas measured in both gyri and sulci. Paired t-tests were used to determine differences between groups.
ences between the sulcal wall and the fundus. Therefore, we examined binding to gyri and sulci separately in order to take into account potential anatomical differences.
Serotonin transporter and 5-HT~A binding data were analyzed separately for differences in each of the variables of group (suicide and control), sex and age.
3.1. [3H]8-OH-DPAT binding to serotonin 5-HTIA recep- tors
Five isodensity bands were observed in all cortical areas studied in suicides and controls (Fig. IA, Fig. 2), which corresponded to histologically identified cortical layers as follows: band I (cortical layer I), band II (cortical layer II), band III (upper layer III), band I I I - I V (inner layer III and layer IV) and band V - V I (layers V and VI).
Group differences The logistic procedure separated the two groups using
binding data from four regions. The data from these re-

126 V. Arango et al. / Brain Research 688 (1995) 121-133
Table 1
Model coefficients for 5-HTI~ receptor binding in prefrontal cortex: significant variables identified by the logistic regression analysis in a group of suicide victims and controls
Variable: Group
Variable Value of Sig. of Wald's (transformed) ~ coefficient stat. ( P )
LN 111ASI 16.5434 11.0275
LNI21AS56 - 12.9856 1/.0233
LN321AGGR 21.5996 11.0195
LN451ASGR 25.6795 11.0287
Constant - 53.2526 0.0265
Variable: Sex
Variable Value of Sig. of Wald ' s
(transformed) ~ coefficient stat. ( P )
LN321AG56 - 29.4346 0.0615
LN471AG34 29.8473 0.0614
LN471 AS34 20.9460 0.0824
Constant - 39.1049 0.0429
Variable: Age
Variable Value of Sig. of Wald's (transformed) ~ coefficient star. ( P )
LN 121 AS34 7.3877 11.0074
LN451AS56 - 9.3267 0.0055 Constant 3.1 (124 11.3988
~' Thc variable names include the following: LN = log; The first two digits indicate the Brodmann area; 1A identifies the 5-HT1A receptor, followed by S (sulcus) or G (gyms); the next one or two characters indicate the cortical laminae corresponding to a given isodensity band or to the entire gray matter (GR).
gions (natural log of f m o l / m g tissue) generated a mis- match of three out of thirty-six cases, two of which belonged to the suicide group. The four brain regions identified by the model are: band I of sulcal area 11; band V - V I of sulcal area 12; gyral gray matter of area 32 and sulcal gray matter from area 45. The accuracy of classifica- tion was 92%. When post hoc paired t-tests were per- formed, only Brodmann area 45 was significant (see be- low). (Table 1 presents pertinent model information on these four brain regions).
We studied in addition, the model information con- tained in the penultimate iteration of the multivariate logis- tic regression discussed above. Brodmann areas 45 and 46 contained many subregions where binding differences ap- proached statistical significance for both receptors. This led us to consider post-hoc paired t-tests for these two areas for both 5-HTtA and serotonin transporter. The mean values for 5-HT,~ binding in control and suicide groups differed significantly in seven bands of gyral and sulcal Brodmann area 45 ( P < 0.008 in six regions and P = 0.037 in the seventh). Serotonin 5-HTtA binding was higher for the suicide group in all the seven bands. Brodmann area 46 had three bands (one in gyrus and two in sulcus, Fig. 2) in which 5-HT1A binding for the two groups differed signifi- cantly, and it was higher for suicides. The t values ranged
from 2.37 to 3.67 ( P = 0.030 to 0.008). Table 2 presents these results.
Higher mean levels of binding were found in 85 of the 103 brain regions (bands) studied in the suicide group compared to the control group (Wilcoxon Matched-Pairs Signed Ranks Test, P < 0.0001). The differences were more pronounced in the 24 bands from the ventrolateral prefrontal cortex (Brodmann areas 45 and 46) with binding (untransformed) levels in the suicide group between 17 and 30% higher than the controls. In other brain areas suicides had between 15% fewer sites to 10% more sites than controls. When these variables were transformed us- ing the natural logarithm, in order to meet the requirements of statistical analysis, the percentage increases were be- tween 5 and 19%.
Sex d i f f e rences
The procedure identified the two sexes with only one mismatch out of the 36 cases, using receptor binding results (natural log of f m o l / m g tissue) from three regions: band V - V I of gyral area 32, (2.35 + 0.06 for males vs. 2.41 + 0.05 for females); band I l l - IV for both sulcal and gyral area 47 (1.83 + 0.06 for males vs. 2.24 _+ 0.05 for females and 1.89 + 0.07 for males vs. 2.04 +_ 0.07 for females, respectively). The accuracy of classification was 97%. See Table 1 for details on these variables. Binding was greater in females than males in 98 /103 brain regions
Table 2
Post-hoc paired t-tests for 5-HT1A receptor binding in Brodmann areas 45 and 46
Variable Mean _+ S.E.M. ~ t P (2-tailed)
Brodmann area 45
Band 11, Gyrus C: 3.59_+0.10
S: 3.80_+0.06 2.27 0.037
Band I l l - IV , Gyrus C: 1.48_+0.11
S: 1.76_+0.10 3 0.008 Band V-VI, Gyrus C: 2.11 _+ 0.09
S: 2.30_+0.09 2.7 11.015
Gyral gray C: 2.55 _+ 0.08
S: 2.77_+11.07 3.17 0.0(16 Band II, Sulcus C: 3.57 _+ 0.(17
S: 3.76-+(I.07 3.71 0.0(12 Band III-IV, Sulcus C: 1.56-+0.08
S: 1.81 _+(I.08 3.41 0.0(13 Sulcal gray C: 2.52_+ 0.06
S: 2.77_+0.06 4.46 0.000
Brodmann area 46
Band 11, Gyms C: 3.55 + 0.09
S: 3.64_+0.08 2.37 0.031/ Band 11, Sulcus C: 3.46_+0.10
S: 3.67_+0.08 3.67 0.002 Band Il l-IV, Sulcus C: 1.50+_11.11
S: 1.72+_0.12 3.02 0.008
Natural log of f m o l / m g tissue, obtained from 18 pairs of suicide and control. Degrees of freedom for all = 17. C = control group; S = suicide group.

V. A ra ngo e t a 1. / Brain Research 688 (1995) 121 - 133 127
60
E 40
c--
:'5 ¢ -
121 -t- O +
"U
(,9
20
~ ] Males (n=26) Females (n=10)
, p < 0.05
4 * II III Ill-IV V-VI white
Cortical Layers
Fig. 3. Sex differences in [3H]8-OH-DPAT binding in cortical layers in the Sulcus of Brodmann area 46. Bar graph comparing the specific binding of [3H]8-OH-DPAT to 5-HTIA sites between males and femalcs once again utilizing the latcral prefrontal cortex (sulcus of Brodmann area 46) as a representative area. Specific binding is expressed in fmol/mg tissue + S.E.M. for 26 males (light stipple) and 10 females (dark stipple). Data for cach subject was obtained as described in Figs. 2 and 3.
( P < 0.0001). Females had higher b ind ing than males with
the difference ranging from 1 to 40%. Fig. 3 shows the
unt ransformed data for males and females in suical area
46.
A g e d i f f e rences
The age of each individual was transformed into a d ichotomous variable using the median age of 32.5 y.
Receptor b ind ing (natural log of f m o l / m g tissue) in two regions were identified by the model as significant. These
regions are: sulcal band I I I - I V of area 12 (2.15 _+ (I.09 4:br above-median group vs. 2.06 +_ 0.08 for below median
group); band V - V I of sulcal area 45 (1.92 +__ 0.08 for above median group vs. 2.17 + 0.07 for below median group). The accuracy of classification of this d ichotomous
variable was 77%, with ten misclassifications. 7 1 / 1 0 3 brain regions had lower b ind ing in the older group. Table 1 presents details on the logistic analysis.
3.2. [ 3 H / C N - I M I b i n d i n g to the s e ro ton in t r a n s p o r t e r
Binding to the serotonin transporter was mostly bi lami- nar (Fig. 1B). Therefore, we measured specific b ind ing
over cortical layers I, II and upper III (outer band), and over inner III, IV, V and VI ( inner band), as well as over the entire thickness of the gray matter. The outer cortical
layers had higher levels of binding. For consis tency with [3H]8-OH-DPAT binding, both sulcal and gyral regions
were measured separately, but did not differ ( P > 0.05). The b i laminar pattern was most str iking in the anterior cingulate gyrus, where the outer band had more than twice
25
20 E O
~ 15 c ~5 .c_ rn
.o_ 5
~ Control (n=22)
Suicide (n=22)
* p < 0.05
< 7
8 9 46 45 47 11 12 32
Brodmann Areas
Fig. 4. [3H]CN-IMI binding in sulci of Brodmann areas across prefrontal cortex: a comparison of suicides and controls. Bar graph comparing the specific binding of [ 3 HICN-IMI to serotonin transporter sites between the suicide and control groups across the entire thickness of the sulcal gray matter in multiplc Brodmann areas of the prefrontal cortex. Specific binding is expressed in fmol/mg tissue + S.E.M. for 22 subjects in each group. As with previous figures, data for each subject represents mean values obtained from 3 subtracted autoradiograms. Analyses between groups were done by paired t-tests.

128 V. Arango et al. / Brain Research 688 (1995) 121-133
the level of binding of the inner band. The bilaminar pattern of receptor distribution was preserved in suicide victims. Levels of binding differed greatly across cortical areas within the prefrontal cortex, and was more than four times higher in the anterior cingulate gyrus than in the association cortex of area 9. As with 5-HT~A binding, the pattern of differential binding to the serotonin transporter across Brodmann areas in prefrontal cortex was preserved in suicide victims.
The multivariate statistical procedure used for serotonin binding analyses is the same as above. The variables were not transformed because skewness in the receptor binding levels was negligible. As pointed out earlier, the logistic analysis was performed three times to study group, sex and age differences using binding levels ( f m o l / m g tissue) in all 43 regions (bands). The results are presented separately
for each of the three variables.
Group differences The control and suicide groups were identified by the
model using the binding data from only one region, namely, inner band of gyral area 46 (5.96 ___ 0.43 for the control group vs. 4.72 + 0.42 for the suicide group). There were thirteen mismatches out of a total 44 observations, yielding an accuracy of 71%. Table 3 presents the model parame- ters. Serotonin transporter binding was lower in the suicide group in 42 of the 43 brain regions (Wilcoxon Matched- Pairs Signed-Ranks Test, P < 0.0001, see Fig. 4). The exception was in the inner band of gyral area 32. The decrease in binding was more pronounced in Brodmann areas 45 and 46, the difference ranging between 16 and 27%. In the other cortical regions, the difference ranged between zero and 21%.
Table 3 Model coefficients for serotonin transporter receptor binding in prefrontal cortex: significant variables identified by the logistic regression analysis in a group of suicide victims and controls
Variable: Group
Variable Value of Sig. of Wald's coefficient stat.(P)
465UGIN - 0.3320 0.0472 Constant 1.7772 0.0730
Variable: Sex
Variable Value of Sig. of Wald's coefficient stat. (P)
325USIN - 0.4821 0.0081 Constant 2.7435 0.0261
a The variable names are constructed as follows: the first two digits indicate the Brodmann area; U identifies the serotonin transporter (up- take) site, followed by S (sulcus) or G (gyrus); the next two characters indicate the inner (IN) cortical laminae corresponding to an isodensity band.
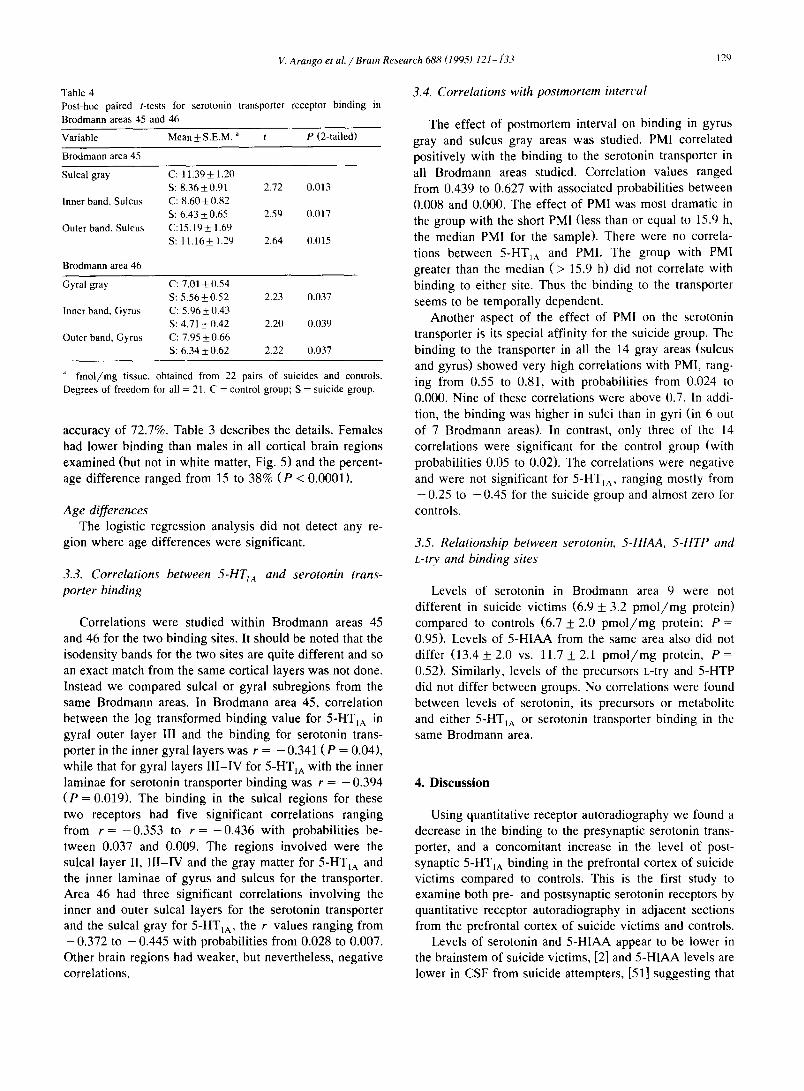
Post-hoe paired t-tests showed that probabilit ies associ- ated with the mean values for the serotonin transporter binding were significant in six bands in areas 45 and 46. The mean values for the suicide group was lower than the control group in all the six significant regions. Table 4 present details of these results.
Sex differences The inner layers of sulcal area 32 (8.56 + 0.61 for
males vs. 5.52 + 0.51 for females) had significantly differ- ent binding. Male and female subjects were identified with a mismatch of twelve subjects, yielding a classification
25
~ 20
E v
t~ 15 ._=
~5
-~ 10
o
0
~ Males (n=28)
Females (n=16)
* p < 0.05
i * i i * .
l I11i i
8 9 46 45 47 11 12 32
Brodmann Areas
Fig. 5. Sex differences in [3H]CN-IMI binding in sulci of Brodmann areas across prefrontal cortex. Bar graph comparing the specific binding of [3H]CN-IMI to serotonin transporter sites between males (light stipple) and females (dark stipple) across multiple Brodmann areas in the prefrontal cortex. Specific binding is expressed in fmol/mg tissue + S.E.M. for 28 males and 16 females. Data for each subject was obtained as described in Figs. 2, 3 and 5. Differences between sexes were present in all areas measured.
V. Arango et al. / Brain Research 688 (1995) 121-1"33 129
Table 4 Post-hoe paired t-tests
Brodmann areas 45 and
for serotonin transporter receptor binding in
46
Variable Mean + S.E.M. " t P (2-tailed)
Brodmann area 45
Sulcal gray C: 11.39 -+ 1.20 S: 8.36+0.91 2.72 0.013
Inner band, Sulcus C: 8,60 + 0.82 S: 6.43+_0.65 2.59 0,017
Outer band, Sulcus C:15.19+ 1.69 S: 11.16+ 1.29 2.64 0.015
Brodmann area 46
Gyral gray C: 7.01 + 0.54 S: 5.56+_0.52 2.23 0.037
Inner band, Gyrus C: 5.96 _+ 0.43 S: 4.71 _+ 0.42 2.20 0.039
Outer band, Gyrus C: 7.95 + 0.66 S: 6.34+0.62 2.22 0,037
~' fmo l /mg tissue, obtained from 22 pairs of suicides and controls.
Degrees of freedom for all = 21. C = control group; S = suicide group.
accuracy of 72.7%. Table 3 describes the details. Females had lower binding than males in all cortical brain regions examined (but not in white matter, Fig. 5) and the percent- age difference ranged from 15 to 38% ( P < 0.0001).
Age differences The logistic regression analysis did not detect any re-
gion where age differences were significant.
3.3. Correlations between 5-1"ITIA and serotonin trans- porter binding
Correlations were studied within Brodmann areas 45 and 46 for the two binding sites. It should be noted that the isodensity bands for the two sites are quite different and so an exact match from the same cortical layers was not done. Instead we compared sulcal or gyral subregions from the same Brodmann areas. In Brodmann area 45, correlation between the log transformed binding value for 5-HTIA in gyral outer layer Ill and the binding for serotonin trans- porter in the inner gyral layers was r = - 0.341 ( P = 0.04), while that for gyral layers I l l - IV for 5-HT1A with the inner laminae for serotonin transporter binding was r = -0 .394 ( P = 0.019). The binding in the sulcal regions for these two receptors had five significant correlations ranging from r = -0 .353 to r = -0 .436 with probabilities be- tween 0.037 and 0.009. The regions involved were the sulcal layer II, I I I - IV and the gray matter for 5-HT1A and the inner laminae of gyrus and sulcus for the transporter. Area 46 had three significant correlations involving the inner and outer sulcal layers for the serotonin transporter and the sulcal gray for 5-HTtA, the r values ranging from -0 .372 to -0 .445 with probabilities from 0.028 to 0.007. Other brain regions had weaker, but nevertheless, negative correlations.
3.4. Correlations with postmortem interl~al
The effect of postmortem interval on binding in gyrus gray and sulcus gray areas was studied. PMI correlated positively with the binding to the serotonin transporter in all Brodmann areas studied. Correlation values ranged from 0.439 to 0.627 with associated probabilities between 0.008 and 0.000. The effect of PMI was most dramatic in the group with the short PMI (less than or equal to 15.9 h, the median PMI for the sample). There were no correla- tions between 5-HT1A and PMI. The group with PMI greater than the median ( > 15.9 h) did not correlate with binding to either site. Thus the binding to the transporter seems to be temporally dependent.
Another aspect of the effect of PMI on the serotonin transporter is its special affinity for the suicide group. The binding to the transporter in all the 14 gray areas (sulcus and gyrus) showed very high correlations with PMI, rang- ing from 0.55 to 0.81, with probabilities from 0.024 to 0.000. Nine of these correlations were above 0.7. In addi- tion, the binding was higher in sulci than in gyri (in 6 out of 7 Brodmann areas). In contrast, only three of the 14 correlations were significant for the control group (with probabilities 0.05 to 0.02). The correlations were negative and were not significant for 5-HT~A, ranging mostly from -0 .25 to -0 .45 for the suicide group and almost zero for controls.
3.5. Relationship between serotonin, 5-HIAA, 5-HTP and L-try and binding sites
Levels of serotonin in Brodmann area 9 were not different in suicide victims (6.9 _+ 3.2 pmo l /mg protein) compared to controls (6.7 _+ 2.0 pmo i /mg protein; P = 0.95). Levels of 5-HIAA from the same area also did not differ (13.4 _+ 2.0 vs. 11.7 _+ 2.1 p m o l / m g protein, P = 0.52). Similarly, levels of the precursors L-try and 5-HTP did not differ between groups. No correlations were found between levels of serotonin, its precursors or metabolite and either 5-HTlA or serotonin transporter binding in the same Brodmann area.
4. Discussion
Using quantitative receptor autoradiography we found a decrease in the binding to the presynaptic serotonin trans- porter, and a concomitant increase in the level of post- synaptic 5-HT1A binding in the prefrontal cortex of suicide victims compared to controls. This is the first study to examine both pre- and postsynaptic serotonin receptors by quantitative receptor autoradiography in adjacent sections from the prefrontal cortex of suicide victims and controls.
Levels of serotonin and 5-HIAA appear to be lower in the brainstem of suicide victims, [2] and 5-HIAA levels are lower in CSF from suicide attempters, [51] suggesting that

131} V. Arango et al. / Brain Research 688 (1995) 121-133
reduced serotonergic function is associated with suicide and attempted suicide. This reduction appears to be inde- pendent of diagnostic group and related to a history of a serious suicidal act. We therefore hypothesized that the postsynaptic 5-HTIA receptor may be upregulated and the nerve terminal serotonin transporter may be reduced in the suicide group compared to controls.
The differences in binding levels for the serotonin transporter were more uniformly widespread (42/43 re- gions) than for the 5-HT1A receptor (85/103 regions). Both binding site populations in suicide showed the great- est differences from controls in the orbital and lateral prefrontal cortex, suggesting the greatest abnormality in suicides are in these regions. Furthermore, the correlations between the 5-HTIA and serotonin transporter binding levels were found to be negative in more than 90% of the regions studied. Thus, we find both a negative correlation in binding to the 5-HT1A receptor and the serotonin trans- porter in both groups, as well as significant changes in these pre- and postsynaptic serotonin receptors in suicide victims compared to controls in ventrolateral prefrontal cortex. We interpret these results to suggest that there is an underlying common influence on 5-HT~A and serotonin transporter binding, and this influence results in changes in opposite directions on the two receptor subtypes.
Earlier binding studies of 5-HT t receptors in suicide victims [23,54,58,59] used [3H]5-HT in homogenates from frontal cortex and other regions. None of these studies reported any differences between groups, perhaps because such an approach fails to distinguish between serotonin receptor subtypes. [3H]5-HT binds with nanomolar affinity to 5-HT1A, 5-HTic, 5-HT1D and probably other receptor subtypes. More recent studies are in disagreement as to whether there is an increase in 5-HT1A binding in suicides (see [2,55] for review). One study found an increase in Bma x in the dorsal prefrontal cortex of nonviolent suicides (but not in violent suicides) compared to controls [56]. Another study found increased 5-HT1A binding in the entorhinal cortex and hippocampus of violent suicides [38]. The binding to 5-HTIA sites, as labeled by [3H]5-HT, was reported to be unaltered in frontal and temporal cortex of depressed suicide victims compared to controls [21]. More recent studies with negative results, using the selective 5-HT~A receptor agonist [3H]8-OH-DPAT as the ligand, have also had methodological limitations. For example, Dillon et al. [26] found no difference in 5-HTIA binding between suicide and control groups either in homogenates from pre/postcentral gyri or in several fronto-temporai cortical and subcortical regions by autoradiography, but their protocol did not include masking for serotonin trans- porter sites. Since serotonin transporter sites may be re- duced in number, not blocking [3H]8-OH-DPAT binding to the transporter may result in failure to detect increase in 5-HT1A binding.
Most of the studies of the serotonin transporter in suicide victims have used homogenates and [3H]imipra-
mine as the ligand. These studies have not distinguished between [3H]imipramine binding to the high- vs. low-affin- ity sites [19,37], although only the high-affinity site corre- sponds to the serotonin transporter [24]. In contrast, [3H]CN-IMI only binds to a single, high-affinity site [27,35,42], probably the physiologically important one. [3H]paroxetine, a nontricyclic serotonin uptake inhibitor, has advantages over both [3H]CN-IMI and [3H]imipramine in terms of receptor specificity [34], and although it is not effective for human autoradiography (Arango and Erns- berger, unpublished work), it may be preferable to use this high-affinity selective ligand in studies of binding kinetics in membrane preparations.
Studies of [3H]imipramine in homogenates from frontal cortex have reported conflicting results. Most of these studies found a decrease in the number of binding sites in the frontal cortex of the suicide group [5,6,23,62,70,71]. Three studies found no change [7,8,58], and one study found an increase in these sites in the suicide group [57]. In looking at other brain regions, one study [62] found a 30% decrease in the hypothalamus of the suicide group, and another [46] found a decrease in the putamen of depressed suicide victims dying from an overdose. Four studies have used [3H]paroxetine as the ligand and two of those [45,46] reported no changes in the prefrontal cortex (Brodmann area 10) of depressed suicide victims com- pared to controls. The other studies [43,44] found a 35% and a 26% reduction in the prefrontal and temporal cortex, respectively, of drug-free suicide victims compared to controls [43], and a 43% reduction in prefrontal cortex [44]. As indicated above, [3H]imipramine is not a specific ligand for the transporter and therefore some of the differ- ences in results may be related to changes in other recep- tors, such as the non-transporter imipramine binding site (protease-resistant, not displaced by serotonin), o~j-adren- ergic or muscarinic cholinergic sites [24]. Moreover, if, as our present study indicates, the decrease in transporter binding is generalized but greater in specific subregions of the prefrontal cortex, then the results of homogenate assays are somewhat dependent on the accuracy of dissection of specific cortical regions.
Of the six studies that have reported no significant decreases in serotonin transporter binding in suicide vic- tims compared to controls, four had postmortem intervals greater than 35 h [2]. We found evidence of an increase in binding associated with long postmortem intervals in the suicide, but not the control group. Thus, differences be- tween these groups may be obscured in studies with longer postmortem intervals, because binding in the suicide group may increase and that of controls remain the same across the full range of PMI's. The other two studies reporting no reductions in binding had PMI's shorter than 27 h. On the other hand, all the studies reporting decreases in trans- porter binding in the suicide group had mean postmortem intervals shorter than 22 h [2,18,38]. The cause of the increase in binding with increased PMI is unknown.

V. Arango et al. / Brain Research 688 (1995) 121-133 131
One previous study utilized [3H]imipramine by auto- radiography [33] on coronal sections from one hemicere- brum and found reduced binding in the postcentral gyrus, the insular cortex and the claustrum, and increased binding in several areas of the hippocampal formation. Our study of the prefrontal cortex indicates a relatively localized decrease in the ventrolateral region. Thus the variability of results in studies of the serotonin transporter, may be due in part to the choice of iigand, the brain region examined, as well as the age and sex of the population studied, postmortem interval and the use of antemortem medica-
tion. Binding of [3H]8-OH-DPAT had a similar cortical lami-
nation in both suicide victims and controls, and therefore the greater binding was not due to a change in the distribu- tion or organization of 5-HT~A binding sites. In normal brain, binding to gyri and sulci had similar laminar organi- zation, although in the sulcus, cortical laminae were thin- ner and closer together. However, in both suicides and controls, binding varied similarly across cortical areas which emphasizes the need for precise comparison of the same brain regions between study groups. For example, binding was higher in the orbital cortex and medial cortex than in the dorsolateral cortex. Such regional comparisons in 5-HT~A binding within multiple regions at one coronal level are not addressed in previously published normative studies [63].
The suggestion that greater 5-HT~A binding represents upregulation of postsynaptic 5-HT~A serotonergic sites sec- ondary to decreased serotonergic innervation is, for the most part, not supported by animal models where no alterations have been reported after chemical lesions of the serotonergic system [39,73]. On the other hand, we have found increased 5-HT~A binding in frontal cortex of rats after electrolytic lesions of the dorsal raphe nucleus [77] and a 20-40% increase in 5-HT~A mRNA was reported in rat 11 days after 5,7-dihydroxytryptamine lesions [16], as well as an increase in 5-HT~A immunoreactivity in the hippocampus 14 days post-lesion [61]. Our results are consistent with an upregulation hypothesis for greater 5- HT~A binding in the ventrolateral prefrontal cortex of suicide victims. Studies of the effect of serotonin depletion on serotonin transporter binding are lacking.
The functions of the prefrontal cortex (PFC) in humans are unknown, but evidence suggests that it may be in- volved in behavioral inhibition and the expression of emo- tion (see [31,47] for review). Lesions of the PFC, either as a result of trauma or stroke [25,66,67,72], are commonly associated with the development of depression or aggres- sion. Disinhibition is also a consequence of a prefrontal lesion, suggesting that this area has an executive inhibitory action. Epilepsy involving the frontal cortex, particularly the right hemisphere, has been associated with pathological crying [67]. An early study [78] of patients with localized frontotemporal lobe contusion vs. brainstem contusion showed that 5-HIAA in the cerebrospinal fluid was re-
duced in the first group, compared to controls and to those patients with brainstem dysfunction, a condition which persisted for 6 months. Dysfunction of the prefrontal cor- tex may be the result of damaged subcortical afferent pathways including serotonergic fibers. Lesions of subcor- tical pathways may at least partly result in reduced sero- tonergic input. Alternatively, an abnormality may exist at the level of serotonin cell bodies in the raphe nuclei, which are yet to be studied directly. Regardless of the site of the lesion, the result of loss of serotonin input into the pre- frontal cortex may result in both disinhibition and affective changes that lead to suicidal acts.
Future postmortem studies should address the location of altered serotonin input in terms of cell bodies, fiber pathways, nerve terminals or target cortical neurons. These studies should also confirm that the association of altered serotonin function and suicide is independent of diagnosis by conducting careful psychological autopsies.
Acknowledgements
We gratefully acknowledge the assistance of the follow- ing people (listed alphabetically): Michelle L. Allen (auto- radiography), Victor V. Arkhipov (tissue collection), Mihran J. Bakalian (data management), David A. Brent, M.D., Denise M. Gillen (autoradiography), Theodore F. Lagattuta (HPLC assays), Patrick J. McDevitt (image anal- ysis), Tammy A. Mieczkowski (statistical analyses), William E. Miller (determination of binding kinetics and autoradiography assays), Brian W. Rigatti (tissue collec- tion) and Robert W. Smith (image analysis). Supported by PHS Grants MH40210, AA09004, MH47097 and MH46745.
References
[1] Arango, V., Ernsberger, P., Marzuk, P.M., Chen, J.-S., Tierncy, H., Stanley, M., Reis, D.J. and Mann, J.J. Autoradiographic demonstra- tion of increased serotonin 5-HT 2 and /3-adrenergic receptor binding sites in the brain of suicide victims, Arch. Gen. Psychiatry, 47 (1990) 1038-1047.
[2] Arango, V. and Mann, J.J., Relevance of serotonergic postmortem studies to suicidal behavior, Int. Rec. Psychiatry, 4 (1992) 131-140.
[3] Arango, V., Miller, W.E., Miller, M.L., Underwood, M.D., Smith, R.W. and Mann, J.J., Quantitative autoradiography of 5-HTIA bind- ing in suicide, Soc. Neurosci. Abstr. 17 (1991) 1472.
[4] Arango, V., Underwood, M.D., McDevitt, P.J., Gubbi, A.V. and Mann, J.J., Localized alterations in pre-and postsynaptic serotonin binding sites in prefrontal cortex of suicide victims, Soc. Neurosci. Abstr. 20 (1994) 26.
[5] Arat6, M., Tekes, K., Palkovits, M., Demeter, E. and Falus, A. Serotonergic split brain and suicide, Psychiatry Res. 21 (1987) 355-356.
[6] Arat6, M., Tekes, K., T6thfalusi, L., Magyar, K., Palkovits, M., Frecska, E., Falus, A. and MacCrimmon, D.J., Reversed hemispheric asymmetry of imipramine binding in suicide victims, Biol. Psychia- t~,, 29 (1991) 699-702.

132 V. Arango et at./Brain Research 688 (1995) 121-133
3 . . . . . [7] Arora, R.C. and Meltzer, H.Y., H-lmlpramlne binding m the frontal cortex of suicides, Psychiatry Res. 30 (1989) 125-135.
3 • , . [8] Arora, R.C. and Meltzer, H.Y., Laterality and H-lmlpramme bind- ing: Studies in the frontal cortex of normal controls and suicide victims, Biol. Psychiatry, 29 (1991) 1016-1022.
[9] Agren, H., Symptom patterns in unipolar and bipolar depression correlating with monoamine metabolites in the cerebrospinal fluid. II. Suicide, Psychiatry Res., 3 (1980) 225-236.
[10] Asberg, M., Tr~iskman, L. and Thor~n, P., 5-HIAA in the cere- brospinal fluid. A biochemical suicide predictor? Arch. Gen. Psychi- atry, 33 (1976) 1193-1197.
[11] Banki, C.M., Arat6, M., Papp, Z. and Kurcz, M., Biochemical markers in suicidal patients. Investigations with cerebrospinal fluid amine metabolites and neuroendocrine tests, J. Affect. Disord., 6 (1984) 341-350.
[12] Beskow, J., Gottfries, C.G., Roos, B.E. and Winblad, B., Determina- tion of monoamine and monoamine metabolites in the human brain: post mortem studies in a group of suicides and in a control group, Acta Psychiat. Scand., 53 (1976) 7-20.
[13] Bourne, H.R., Bunney, W.E.J., Colburn, R.W., Davis, J.M., Shaw, D.M. and Coppen, AJ., Noradrenaline, 5-hydroxytryptamine, and 5-hydroxyindoleacetic acid in hindbrains of suicidal patients, Lancet, ii (1968) 805-808.
[14] Bradley, P.B., Engel, G., Feniuk, W., Fozard, J.R., Humphrey, P.P.A., Middlemiss, D.N., Mylecharane, E.J., Richardson, B.P. and Saxena, P.R., Proposals for the classification and nomenclature of functional receptors for 5-hydroxytryptamine, Neuropharmacology, 25 (1986) 563-:576.
[15] Branchek, T., Receptor diversity: more serotonin receptors, Curr. Biol., 3 (1993) 315-317.
[16] Brousseau, D., Wieland, S., Lucki, I. and McGonigle, P., 5-HT depletion alters the levels of 5-HTla receptor mRNA, Soc. Neurosci., 17 (1991) 719.
[17] Brown, G.L., Goodwin, F.K. and Bunney, W.E.J., Human aggres- sion and suicide: their relationship to neuropsychiatric diagnoses and serotonin metabolism, AdL,. Biochem. Psychopharmacol., 34 (1982) 287-307.
[18] Carpenter, W.T., Jr., Buchanan, R.W., Kirkpatrick, B., Tamminga, C. and Wood, F., Strong inference, theory testing, and the neu- roanatomy of schizophrenia, Arch. Gen. Psychiatry, 50 (1993) 825-831.
[19] Cash, R., Raisman, R., Ploska, A. and Agid, Y., High and low affinity [3H]imipramine binding sites in control and Parkinsonian brains, Eur. J. Pharmacol., 117 (1985) 71-80.
[20] Center for Disease Control and Prevention, Advance report of final mortality statistics, 1990, Mthly Vital Stat. Report, 41 (1993) 1-52.
[21] Cheetham, S.C., Crompton, M.R., Katona, C.L.E. and Horton, R.W. Brain 5-HT 1 binding sites in depressed suicides, Psychopharmacol- ogy (Berlin), 102 (1990) 544-548.
[22] Coccaro, E.F., Siever, L.J., Klar, H.M., Maurer, G., Cochrane, K., Cooper, T.B., Mohs, R.C. and Davis, K.L., Serotonergic studies in patients with affective and personality disorders, Arch. Gen. Psychi- atry, 46 (1989) 587-599.
[23] Crow, T.J., Cross, A.J., Cooper, S.J., Deakin, J.F.W., Ferrier, I.N., Johnson, J.A., Joseph, M.H., Owen, F., Poulter, M., Lofthouse, R., Corsellis, J.A.N., Chambers, D.R., Blessed, G., Perry, E.K., Perry, R.H. and Tomlinson, B.E., Neurotransmitter receptors and monoamine metabolites in the brains of patients with Alzheimer-type dementia and depression, and suicides. Neuropharmacology, 23 (1984) 1561-1569.
[24] D'Amato, R.J., Largent, B.L., Snowman, A.M. and Snyder, S.H., Selective labeling of serotonin uptake sites in rat brain by [3H]cita- lopram contrasted to labeling of multiple sites by [3 H]imipramine, J. PharmacoL Exp. Ther., 242 (1987) 364-371.
[25] Damasio, H., Grabowski, T., Frank, R., Galaburda, A.M. and Dama- sio, A.R., The return of Phineas Gage: clues about the brain from the skull of a famous patient, Science, 264 (1994) 1102-1105.
[26] Dillon, K.A., Gross-lsseroff, R., Israeli, M. and Biegon, A., Auto- radiographic analysis of serotonin 5-HTIA receptor binding in the human brain postmortem: Effects of age and alcohol, Brain Res., 554 (1991) 56-64.
[27] Dumbrille-Ross, A. and Tang, S.W., Binding of [3H]Ro 11-2465. Possible identification of a subclass of [3 H]imipramine binding sites, Mol. Pharmacol., 23 (1983) 607-613.
[28] Dumuis, A., Sebben, M. and Bockaert, J., The gastrointestinal prokinetic benzamide derivatives are agonists at the non-classical 5-HT receptor (5-HT 4) positively coupled to adenylate cyclase in neurons, Naunyn Schmiedebergs Arch. Pharmacol., 340 (1989) 403 -410.
[29] Frazer, A. and Hensler, J.G., 5-HTjA receptors and 5-HTiA-media- ted responses: effect of treatments that modify serotonergic neuro- transmission, Ann. NYAcad. Sci., 600 (1990) 460-475.
[30] Fuster, J.M., The prefrontal cortex and temporal integration. In A. Peters and E.G. Jones (Eds.), Cerebral Cortex: Association and Auditory Cortices, Plenum Press, New York, 1985, pp. 151-177.
[31] Fuster, J.M., The Prefrontal Cortex - - Anatomy, Physiology and Neuropsychology of the Frontal Lobe, Raven Press, New York, NY, 1989, pp. 1-255.
[32] Gozlan, H., E1 Mestikawy, S., Pichat, L., Glowinsky, J. and Hamon, M., Identification of presynaptic serotonin autoreceptors using a new ligand: 3H-PAT, Nature, 305 (1983) 140-142.
[33] Gross-lsseroff, R., Israeli, M. and Biegon, A., Autoradiographic analysis of tritiated imipramine binding in the human brain post mortem: effects of suicide, Arch. Gen. Psychiatry, 46 (1989) 237- 241.
[34] Habert, E., Graham, D., Tahraoui, L., Claustre, Y. and Langer, S.Z., Characterization of 3 H-paroxetine binding to rat cortical membranes, Eur. J. Pharmacol., 118 (1985) 107-114.
[35] Hensler, J.G., Ferry, R.C., Labow, D.M., Kovachich, G.B. and Frazer, A., Quantitative autoradiography of the serotonin transporter to assess the distribution of serotonergic projections from the dorsal raphe nucleus, Synapse, 17 (1994) 1-15.
[36] Henteleff, R.A., Arango, V. and Mann, J.J., Binding to serotonin uptake sites in prefrontal and temporal cortex of suicide victims, Soc. Neurosci. Abstr., 17 (1991) 1472.
[37] Hrdina, P.D., Differentiation of two components of specific ['a]im- ipramine binding in rat brain, Eur. J. Pharmacol., 102 (1984) 481-488.
[38] Joyce, J.N., Shane, A., Lexow, N., Winokur, A., Casanova, M.F. and Kleinman, J.E., Serotonin uptake sites and serotonin receptors are altered in the limbic system of schizophrenics, Neuropsy- chopharmacology, 8 (1993) 315-336.
[39] Juorio, A.V., Li, X.M. and Boulton, A.A., The effects of chronic trimipramine treatment on biogenic amine metabolism and on dopamine D2, 5-HT 2 and tryptamine binding sites in rat brain, Gen. Pharmacol., 21 (1990) 759-762.
[40] Kilpatrick, G.J., Jones, B.J. and Tyers, M.B., The distribution of specific binding of the 5-HT3 receptor ligand [3H]GR65630 in rat brain using quantitative autoradiography, Neurosci. Lett., 94 (1988) 156-160.
[41] Korpi, E.R., Kleinman, J., Goodman, S.I., Phillips, l., DeLisi, L.E., Linnoila, M. and Wyatt, R.J., Serotonin and 5-hydroxyindoleacetic acid in brains of suicide victims. Comparison in chronic schizophrenic patients with suicide as cause of death, Arch. Gen. Psychiatry, 43 (1986)594-600.
[42] Kovachich, G.B., Aronson, C.E., Brunswick, D.J. and Frazer, A., Quantitative autoradiography of serotonin uptake sites in rat brain using [3H]cyanoimipramine, Brain Res., 454 (1988) 78-88.
[43] Lagattuta, T.F., Henteleff, R.A., Arango, V. and Mann, J.J., Reduc- tion in cortical serotonin transporter site number in suicide victims in the absence of altered levels of serotonin, its precursors or metabolite, Soc. Neurosci. Abstr., 18 (1992) 1598.
[44] Laruelle, M., Abi-Dargham, A., Casanova, M.F., Toti, R., Wein- berger, D.R. and Kleinman, J.E., Selective abnormalities of pre-

V. Arango et al. / Brain Research 688 (1995) 121-133 133
frontal serotonergic receptors in schizophrenia: a postmortem study, Arch. Gen. Psychiatry, 50 (1993) 810-818.
[45] Lawrence, K.M., De Paermentier, F., Cheetham, S.C., Crompton, M.R., Katona, C.L.E. and Horton, R.W., Symmetrical hemispheric distribution of ~H-paroxetine binding sites in postmortem human brain from controls and suicides, Biol. P~sychiatry, 28 (1990) 544- 546.
[46] Lawrence, K.M., Dc Paermentier, F., Cheetham, S.C., Crompton, M.R., Katona, C.L.E. and Horton, R.W., Brain 5-HT uptake sites, labelled with [3H]paroxetine, in antidepressant-free depressed sui- cides, Brain Res., 526 (1990) 17-22.
[47] Levin, H.S., Eisenberg, H.M. and Benton, A.L., (Eds.), Frontal Lobe Function and Dysfunction, Oxford University Press, New York, NY, 1991, pp. 1-427.
[48] Lloyd, K.G., Farley, l.J., Deck, J.H.N. and Hornykiewicz, O., Serotonin and 5-hydroxyindoleacetic acid in discrete areas of the brainstem of suicide victims and control patients, AdL~. Biochem. Psychopharmacol., 11 (1974)387-397.
[49] Lowry, O.H., Rosenbrough, N.J., Farr, A.L. and Randall, R.J., Protein measurement with the folin phenol reagent, J. Biol. Chem., 193 (1951) 265-275.
[50] Mann, J.J., Arango, V., Marzuk, P.M., Theccanat, S. and Reis, D.J., Evidence for the 5-HT hypothesis of suicide: a review of post-mortem studies, Br. J. Psychiatry, 155 (Suppl. 8) (19891 7-14.
[51] Mann, J.J., McBride, P.A., Brown, R.P., Linnoila, M., Leon, A.C., DeMeo, M.D., Mieczkowski, T.A., Myers, J.E. and Stanley, M., Relationship between central and peripheral serotonin indexes in depressed and suicidal psychiatric inpatients, Arch. Gen. Psychiatry, 49 (1992) 442-446.
[52] Mann, J.J., McBride, P.A., Malone, K.M., DeMeo, M.D. and Keilp, J., Blunted serotonergic responsivity in depressed patients, Neu- ropsychopharmacology, in press.
[53] Mann, J.J. and Stanley, M. (Eds.), Psychobiology of Suicidal Behau- ior, The New York Academy of Sciences, New York, NY, 1986, pp. 1-357.
[54] Mann, J.J., Stanley, M., McBride, P.A. and McEwen, B.S., In- creased serotonin~ and /3-adrenergic receptor binding in the frontal cortices of suicide victims, Arch. Gen. Psychiatry, 43 (1986) 954- 959.
[55] Mann, J.J., Underwood, M.D. and Arango, V., Postmortem studies of suicide victims. In S.J. Watson (Ed.) Biology of Schizophrenia and Affectit,e Disorders, Raven Press, New York, in press.
[56] Matsubara, S., Arora, R.C. and Meltzer, H.Y., Serotonergic mea- sures in suicide brain: 5-HTIA binding sites in frontal cortex of suicide victims, J. Neural Transm., 85 (1991) 181-194.
[57] Meyerson, L.R., Wennogle, L.P., Abel, M.S., Coupet, J., Lippa, A.S., Rauh, C.E. and Beer, B., Human brain receptor alterations in suicide victims, Pharmacol. Biochem. Beha~,., 17 (1982) 159-163.
[58] Owen, F., Chambers. D.R., Cooper, S.J., Crow, T.J., Johnson, J.A., Lofthouse, R. and Poulter, M., Serotonergic mechanisms in brains of suicide victims, Brain Res., 362 (1986) 185-188.
[50] Owen, F., Cross, A.J., Crow, T.J., Deakin, J.F.W., Ferrier, 1.N., Lofthouse, R. and Poulter, M., Brain 5-HT 2 receptors and suicide, Lancet, ii (1983) 1256.
[61)] Pare, C.M.B., Yeung, D.P.H., Price, K. and Stacey, R.S., 5- Hydroxytryptamine, noradrenaline, and dopamine in brainstem, hypothalamus, and caudate nucleus of controls and of patients committing suicide by coal-gas poisoning. Lancet, ii (19691 133- 135.
[61] Patel, T.D., Azmitia, E.C. and Zhou, F.C., Increased 5-HTIA recep-
tor immunoreactivity in the rat hippocampus following 5,7-DHT lesions in the cingulum bundle and fimbria-fornix, Behal. Brain Res., in press.
[62] Paul, S.M., Rehavi, M., Skolnick, P. and Goodwin, F.K., High affinity binding of antidepressants to a biogcnic amine transport site in human brain and platelet; studies in depression. In R.M. Post and C.J. Bellinger (Eds.) Neurobiology of Mood Disorders., Williams and Wilkins, Baltimore, 1984, pp. 846-853.
[63] Pazos, A., Probst, A. and Palacios, J.M., Serotonin receptors in thc human brain--Il l . Autoradiographic mapping of serotonin-I recep- tors, Neuroscience, 21 (1987)97-122.
[64] Pedigo, N.W., Yamamura, H.I. and Nelson, D.L., Discrimination of multiple [3H]5-hydroxytryptamine binding sites by thc neuroleptic spiperone in rat brain, J. Neurochem., 36 (1981) 22(/-226.
[65] Peroutka, S.J. and Snyder, S.H. Multiple serotonin receptors: differ- ential binding to H-5-hydroxytryptammc, 3H-lysergic acid dieth- ylamide and " H-splropendol, Mol. Pharmacol., 16 (19791 687-699.
[66] Robinson, R.G., Kubos, K.L., Starr, L.B., Rao, K. and Price, T.R., Mood disorders in stroke patients. Importance of location of lesion, Brain, 107 (1984) 81-93.
[67] Sackeim, H.A., Greenberg, M.S., Weiman, A.L., Gur, R.C., Hunger- buhler, J.P. and Geschwind, N., Hemispheric asymmetry in the expression of positive and negative emotions, Arch. Neurol., 39 (19821 210-218.
[68] Shaw, D.M., Camps, F.E. and Eccleston, E.G., 5-Hydroxytryp- tamine in the hind-brain of depressive suicides. Br. J. Psychiatry, 113 (19671 1407-1411.
[69] Smith, R.W., Arango, V., Miller, M.L., Underwood, M.D. and Mann, J.J., 3H-cyanoimipramine binding is reduced in suicide vic- tims, Soc. Neurosci. Abstr. 18 (19921 1599.
[70] Stanley, M., Mann, J.J. and Gershon, S., Alterations in pre- and postsynaptic serotonergic neurons in suicidc victims, Psvchophar- macol. Bull., 19 (1983) 684-687.
[71] Stanley, M., Virgilio, J. and Gershon, S., Tritiated imipraminc binding sites are decreased in the frontal cortex of suicides, Science, 216 (19821 1337-1339.
[72] Starkstein, S.E., Robinson, R.G., Berthier, M.L. and Price, T.R., Depressive disorders following posterior circulation as compared with middle cerebral artery infarcts, Brain, 111 (19881 375-387.
[73] Stockmeier, C.A. and Kellar, K.J., In vivo regulation of serotonin-2 receptor in rat brain. Life Sci., 38 (19861 117-127.
[74] Taylor, D.P., Serotonin agents in anxiety, Ann. NYAcad. Sci., 600 (1990) 545--557.
[75] Tecott, L.H. and Julius, D., A new wavc of serotonin receptors, Curt. Opin. Neurobiol., 3 (19931 310-315.
[76] Tr~iskman, L., ~,sberg, M., Bertilsson, L. and Sj6strand, L., Monoamine metabolites in CSF and suicidal bchavior, Arch. Gen. Psychiatry, 38 (1981) 631-636.
[77] Underwood, M.D., Bakalian, M.J., Miller, M.L., Katz, C.K., Lagat- tuta, T.F., Mann, J.J. and Arango, V., Effect of lesions of the dorsal raphe nucleus on serotonin transporter, 5-HTlA and 5-HT~ binding sites in rat, Soc. Neurosci. Abstr., 18 (1992) 928.
[78] van Woerkom, T.C.A.M., Teelken, A.W. and Minderhoud, J.M., Difference in neurotransmitter metabolism in frontotemporal-lobe contusion and diffuse cerebral contusion, Lancet, i (19771 812-813.
[79] Welker, W., Why does cerebral cortex fissure and fold? A review of determinants of gyri and sulci. In E.G. Jones and A. Peters (Eds.) Cerebral Cortex: Comparatit,e Structure and Et.olution of Cerebral Cortex, Part H, Plenum Press, New York, 1900, pp. 3-136.