kauno medicinos universitetas farmacijos …taigi, kaip ir visų organizmo baltymų, fermentų,...
TRANSCRIPT

KAUNO MEDICINOS UNIVERSITETAS FARMACIJOS FAKULTETAS
MONTPELLIER 1 UNIVERSITETAS FARMACIJOS FAKULTETAS
Vacis Tatarūnas
POLIMERAZĖS GRANDININĖS REAKCIJOS METODO TAIKYMAS FARMAKOGENETINIUOSE TYRIMUOSE
MAGISTRO DARBAS
Darbo vadovai : Prof., dr. Catherine Gozé Prof., dr. Vitalis Briedis
KAUNAS MONTPELLIER
2007

TURINYS SANTRUMPOS (5) ĮVADAS (6) I. LITERATŪROS APŽVALGA (8)
A. FARMAKOGENETIKA (8) 1. Vaistų metabolizmas ir transportas (8) 2. Genetiniai polimorfizmai (9) 3. Genotipo ir fenotipo nustatymo metodai (11)
3.1. Fenotipavimas (11) 3.2. Genotipavimas (11)
B. PRIEŠVĖŽINIŲ VAISTŲ FARMAKOGENETIKA (12) 1. Tiopurin-S-metil transferazė (TPMT) (12)
1.1. Tiopurinai (13) 1.2. Geno mutacijos (14) 1.3. Mutacijos aleliuose (14) 1.4. Geno TPMT seka su mutacijomis (15) 1.5. Nepageidaujamas tiopurinų poveikis (16)
2. Uridin-difosfat-gliukuronoziltransferazė (UGT1A1) (16) 2.1. Geno mutacijos (16) 2.2. UGT1A1 TATA-BOX (16) 2.3. Topoizomerazės – 1 inhibitoriai (17) 2.4. Metabolizmas (18) 2.5. Nepageidaujamas vaistų poveikis (19)
3. Aromatazė (19) 3.1. Aromatizacijos reakcijos mechanizmas (20) 3.2. Geno mutacijos (21) 3.3. Vaistai (22) 3.4. Pašaliniai poveikiai (23)
C. DARBO POLIMERAZĖS GRANDININĖS REAKCIJOS METODU PRINCIPAS (24)
1. DNR ekstrahavimo principai (24) 2. DNR ekstrahavimo metodai (24)
2.1. Ekstrahavimas natrio chloridu (druskinis metodas) (24) 2.2. Ekstrahavimas panaudojant DEAE polimerus (25)
3. DNR kiekio nustatymas spektrofotometriškai (26) 4. DNR plazmoje bei serume (27)
4.1. DNR nustatymas plazmoje (27) 4.2. Padidintas plazmos DNR kiekis pas pacientus, sergančius
onkologinėmis ligomis (27) 5. DNR izoliavimas ir konservavimas (27) 6. Polimerazės grandininė reakcija (PGR) (28)
6.1. Principas (28) 6.2. Etapai (28)
6.2.1. Denatūravimas (29) 6.2.2. Hibridizacijos fazė (29) 6.2.3. Elongacijos fazė (29)
6.3. DNR kiekio didėjimas (29) 6.4. Pradmenys polimerazės grandininei reakcijai (30) 6.5. Lizdinės PGR metodas (30)
2

6.6. PGR produktų valymas (31) 6.7. Gel-elektrofarezė (31) 6.8. Nukleino rūgščių vizualizavimas (31) 6.9. Sekvenavimas (32)
D. RNR – RIBONUKLEINĖ RŪGŠTIS (33) 1. RNR ekstrahavimo metodas (33) 2. Atvirkštinės transkripcijos reakcija (34) 3. Geno ekspresijos matavimas (35)
3.1. Metodas (35) 3.2. Detekcijos ‘realiu laiku’ sistemos (36) 3.3. Strategijos (37)
II. EKSPERIMENTINĖ DALIS (38) A. KRAUJO, PLAZMOS BEI SERUMO GENOTIPAVIMAS (38)
1. Medžiagos ir instrumentai (38) 2. Sąlygos (39) 3. Gauti pavyzdžiai (39) 4. DNR ekstrahavimas (40)
4.1. Ekstrahavimo procedūros tobulinimas serumui ir plazmai, ekstrahuojant natrio chloridu (druskiniu metodu) (40) 4.1.1. Ekstrahavimo protokolas (40)
4.2. Ekstrahavimo procedūros, ekstrahuojant iš plazmos ir serumo, panaudojant QIAGEN (QIAAMP DNA BLOOD MIDI KIT) rinkinį, tobulinimas (41) 4.2.1. Ekstrahavimo protokolas (41)
5. Optinės skvarbos matavimas (41) 6. Genas TPMT (42)
6.1. “Ilgųjų” pradmenų patvirtinimo procedūra, panaudojant DNR, ekstrahuotą iš kraujo (42)
6.2. PGR reakcijos optimizavimas egzonui 5 (45) 7. Lizdinės PGR strategija : DNR, ekstrahuotos iš plazmos ir serumo,
genotipavimas (46) 7.1. Plazma paciento 42 (46) 7.2. Paciento 44 plazmos ekstraktas (48) 7.3. Sekvenavimas PGR produkto, gauto panaudojant DNR ekstraktą
iš 370µL paciento 44 plazmos (49) 7.4. Ilgalaikio užšaldymo įtaka : DNR buvo ekstrahuota iš serumo,
kuris buvo užšaldytas metus –200C temperatūroje (50) 8. UGT1A1 genas (51)
8.1. Pradmenys UGT1A1 genui (51) 8.2. Naujųjų pradmenų patvirtinimas, panaudojant kraujo DNR
ekstraktą (51) 8.3. Lizdinės PGR strategija : plazmos ir serumo genotipavimas (54)
8.3.1. Paciento 42 ir paciento 44 plazma (54) 8.3.2. Paciento 0007 serumas : pavyzdys buvo užšaldytas esant
– 200C ir laikomas metus (56) B. GENO EKSPRESIJOS MATAVIMAS (56)
1. Pasirinktas analizės metodas (56) 2. Medžiagos ir instrumentai (57) 3. Sąlygos (57) 4. RNR ekstrahavimas QIAGEN RNEASY MINI rinkiniu (58)
3

4.1. Ekstrahavimo protokolas (58) 5. Optinės skvarbos matavimo rezultatai (58) 6. kDNR gel-elektrofarezė (58) 7. Atvirkštinės transkripcijos reakcija (59) 8. Kiekybinis metodas (59)
8.1. Ląstelių, galinčių tapti pavyzdžiu, paieška (60) 8.1.1. MCF-7 ląstelės (60)
8.2. PGR sąlygų patvirtinimas (60) 8.3. Hela ląstelės (62) 8.4. Palyginamoji studija su MCF-7 (64) 8.5. Aromatazės geno ekspresija pirmos stadijos krūties vėžio ląstelėse
(65) IŠVADA (66) LITERATŪRA (67)
4

SANTRUMPOS
6-MP – 6-merkaptopurinas
6-TGN – 6-tioguaninas
B2 - genas, naudojamas kaip standartas
CYP19 – citochromo P450 šeimos fermentas, aromatazė
CYP2D6 – citochromo P450 šeimos fermentas
CPT-11 – irinotekanas
ddNTP – didezoksiribonukleotidai
DEAE – dietilamino etanas
DEPC – dietil pirokarbonatas
dNTP – dezoksiribonukleotidai
EDTA – etildiamino tetraacto rūgštis
GSTP1 – gliutation-S-transferazė
HPRT - genas, naudojamas kaip standartas
iRNR – informacinė RNR
kDNR – DNR kopija (cDNR)
Mg – magnis
oligo(dX) – oligonukleotidas sudarytas iš tam tikros bazės grandinės
PGR – polimerazės grandininė reakcija
poly(A) – RNR dalis, sudaryta iš adeninų
RT – atvirkštinė transkriptazė
SDS – natrio duodecil sulfatas
SN-38 – aktyvus irinotekano metabolitas
SNP – vieno nukleotido polimorfizmas
TATA-BOX – tai promotoriaus sritis, prie kurios jungiasi transkripcijos faktoriai ar
histonai
TBE – tris borat EDTA
TBP89 – genas, naudojamas kaip standartas
Topo-1 – topoizomerazė 1
TPMT – tiopurin-metiltransferazė
UGT1A1 – uridin-difosfat-gliukuronoziltransferazė 1
UV spinduliai – ultravioletiniai spinduliai
5

ĮVADAS Aš atlikau praktiką Prancūzijoje, Montpellier mieste, Arnaud de Villeneuve ligoninės
ląstelių biologijos ir hormonologijos laboratorijoje.
Laboratorijoje vykdoma veikla:
• Onkologinių ligų nustatymas, remiantis vėžio žymenimis
• Onkogenetika
• Žmogaus papilomos viruso nustatymas
• Farmakogenetika
Mano darbas – farmakogenetika. Rėmiausi 2 metodais:
1. lizdinė polimerazės grandininė reakcija, po kurios atliekamas
sekvenavimas
2. realaus laiko polimerazės grandininė reakcija
Žingsnis po žingsnio įvaldžiau abi technikas : pradėjau nuo DNR ir RNR
ekstrahavimo, bei klasikinės PGR, o užbaigiau realaus laiko PGR metodu.
1. Ligoninių laboratorijose, tiriant tam tikrą geną, naudojamas pacientų kraujas,
kuriame gausu DNR (10-15mg/mL) (Quinque D. ir kt., 2006). Daroma klasikinė
PGR, po kurios seka sekvenavimas. Tačiau dažna yra tokių situacijų, kai neturima
pakankamai kraujo, kaip pvz.:
• Siekiant atlikti retrospektyvius seroteko tyrimus
• Audiniams, fiksuotiems pat-anatomų, pvz.: parafinu
• Kriminologijoje, kai turimi tik maži kiekiai kraujo
Šių dienų technologijos neleidžia identifikuoti DNR sekų (atlikti sekvenavimą),
neturint pakankamai matricos.
2. Norint identifikuoti geno raiškos lygį, daroma kiekybinė PGR (realaus laiko). Tai
metodas, leidžiantis “suskaičiuoti” geno kopijas (mano darbo atveju – RNR),
remiantis cheminių medžiagų fluorescencija.
6

Eksperimento tikslai 1. Patikrinti galimybę gausinti serume ir plazmoje esančią DNR nauju
būdu ir atlikti genetinius tyrimus.
2. Patvirtinti realaus laiko polimerazės grandininės reakcijos (PGR)
metodiką aromatazės genui, panaudojus ląsteles, kurių šio geno raiška
yra pakankama.
Uždaviniai 1. Pritaikyti druskinį ir Qiagen metodus DNR ekstrahavimui iš plazmos
bei serumo.
2. Sumodeliuoti pradmenis lizdinei PGR ir nustatyti jų tinkamumą,
naudojant kraujo DNR ekstraktus.
3. Pritaikyti sąlygas bei patvirtinti lizdinę PGR darbui su plazmos bei
serumo ekstraktais, lyginant gautas eksperimento metu sekas su
patvirtintomis sekomis.
4. Palyginti druskinį ir Qiagen DNR ekstrahavimo metodus.
5. Sekvenuoti DNR, gautą iš mažo kiekio plazmos, ekstrahuotos Qiagen
metodu ir rasti tyliąją mutaciją 7 egzone.
6. Ištirti ilgalaikio biologinės medžiagos užšaldymo efektą : sekvenuoti
DNR iš serumo, laikyto metus esant –200C.
7. Surasti ląstelių tipą, kurių aromatazės geno raiška yra pakankama, kad
galėtų būti naudojama kaip standartas tolimesniems tyrimams.
Darbo objektas 1. Kraujo, plazmos bei serumo DNR:
• tiopurin-S-metiltransferazės, metabolizuojančios tiopurino grupės
vaistus, ir
• uridin-difosfat-gliukuronoziltransferazės, metabolizuojančios topo-1
inhibitorių grupės vaistus, genai.
2. MCF-7 ir Hela ląstelių RNR bei kDNR iš pirmos stadijos krūties naviko
ląstelių:
• Aromatazės genas
7

LITERATŪROS APŽVALGA A. Farmakogenetika Kiekvieno vaisto sukeltas tiek farmakologinis (veiksmingumo), tiek toksikologinis
poveikis kiekvienam pacientui yra skirtingas, todėl gana dažnai vaistų skyrimas ir
vartojimas tampa labai komplikuotas. Prancūzijoje 3.2% hospitalizacijos atvejų yra
sąlygoti vaistų. Tai sudaro 320 milijonų eurų sumą per metus. JAV 100000 mirčių per
metus sąlygojamos gydytojo išrašytų vaistų toksinio poveikio. Genetiniai faktoriai,
sąlygojantys vaistų farmakokinetiką ir farmakodinamiką, dalinai paaiškina skirtingą
vaistų poveikį žmogui (Allorge D. ir kt., 2004).
Farmakogenetikos era prasidėjo 1950 metais, kai mokslininkai nustatė, kad pašalinis
vaistų poveikis gali būti nulemtas individo genetiniame lygmenyje (Meyer A., 2000).
Tačiau tuo metu buvo galima tik fenotipo raiškos analizė, paremta fermentų įvairovės
nustatymu tarp individų grupių ir vaistų metabolitų tyrimu (Bouquier M., 2003).
Farmakogenetika nagrinėja genetinius veiksnius, kurie sąlygoja atsaką ir toleranciją į
pavartotą vaistą. Vaisto veiksmingumas ir toksiškumas yra šio mokslo prioritetai
(Bouquier M., 2003; IPSEN, 2000).
Farmakogenomika tai mokslas, kuris tiria genetinius veiksnius, lemiančius
farmakologinį poveikį. Šio mokslo mąstelis – visas genomas. T.y. identifikuojamas
visas genas tam tikros ligos atveju tam tikram vaistui (Bouquier M., 2003).
1. Vaistų metabolizmas ir transportas Vaistai – tai svarbi cheminių, organizme nesintetinamų (ksenobiotinių) medžiagų
grupė, su kuria žmogus susiduria kasmet vis dažniau. Vaistai dažniausiai yra
hidrofobinės medžiagos. Organizme jos dažniausiai verčiamos hidrofilinėmis
(metabolizuojamos) ir pašalinamos pagrinde su tulžimi ir šlapimu.
Pavartojus vaistų per os, vaistas turi praeiti tam tikrus barjerus (žarnyno sienelė,
skrandis, burnos gleivinė), kad patektų į kraują. Gana dažnai vaistus per barjerą
perneša specialūs nešikliai. Organizme metabolizmas pagrinde vyksta kepenyse.
Fermentai, dalyvaujantys biotransformacijoje, skiriami į dvi grupes :
1. pirmos fazės fermentai. Tai fermentai, tokie kaip oksigenazės,
oksidoreduktazės, hidrolazės, kurių poveikyje molekulės tampa poliškesnės
(hidroksilinimas, dealkilinimas). Dauguma pirmos fazės reakcijų
katalizuojamos citochromų superšeimos fermentų.
8

2. antros fazės fermentai. Tai transferazės, kurios katalizuoja konjugacijos
reakcijas, padarydamos molekulę dar poliškesnę. Konjugacijos būdu
prijungiamos karboksi, sulfo, gliukurono, gliutationo ir kt. grupės.
3. pagaliau, norint pašalinti iš lastelės metabolitus, pernašai per lastelės
membraną panaudojami trečios fazės fermentai : ABC baltymai (ATP binding
cassette) (Allorge D. ir kt., 2004)
Metabolizmo ląstelėje pavyzdys Pav.1.
Fermentai ir pernašos baltymai turi keletą savybių :
Pav.1. Vaisto metabolizmas ir transportas lastelėje (Allorge D. ir kt., 2004)
jie priklauso šeimoms (kaip cyp 450), turinčioms daug izoformų, kurios yra
labai homologiškos amino rūgščių sekos atžvilgiu.
Jie yra santykinai specifiški substratui
Jų ekspresija, palyginti su jų aktyvumu, kinta veikiant tam tikriems fiziologiniams,
patologiniams, aplinkos ir genetiniams faktoriams (Allorge D. ir kt., 2004).
2. Genetiniai polimorfizmai Genetinis polimorfizmas – tai buvimas tam tikroje genetinėje srityje mažiausiai 2
alelių populiacijos atžvilgiu (Kaplan J.C. ir kt., 1990). Genetiniu polimorfizmu
vadinama, kai mutacija ar genetinė variacija pasireiškia dažniau kaip pas 1% visų
populiacijos individų (Nebert D.W ir kt., 2001).
Yra trys DNR sekos variacijos, sukeltos polimorfizmų :
9

1. SNP – vienetinis nukleotidų polimorfizmas (liečia vieną mutaciją – punktuali
mutacija)
2. bazių įterpimas ar delecija, kuri liečia vieną, keletą ar net tūkstančius
nukleotidų
3. pasikartojančių DNR fragmentų įterpimas arba delecija (mikrosatelitai, AluI
segmentai ir kt.) (Bouquier M., 2003).
Taigi, kaip ir visų organizmo baltymų, fermentų, dalyvaujančių biotransformacijoje,
kokybė ir kiekis priklauso nuo juos koduojančio geno. Skirtingos to geno versijos
išreiškiamos skirtingų alelių ir kiekvienas individas turi 2 to pačio geno alelių versijas
(viena iš mamos, kita - iš tėvo), kurios ir apibrėžia genotipą.
Mutacijos, kurios yra genuose, koduojančiuose baltymus, gali sąlygoti ekspresijos
įvairovę (pakeisdamos amino rūgštis baltymo sekoje), tuo pačiu gali sąlygoti baltymo
funkcijų praradimą. Pvz., fermentų aktyvumas gali būti sumažėjęs, padidėjęs arba jo
gali išvis nebūti (Allorge D. ir kt., 2004).
Priklausomai nuo metabolizmo greičio, populiacija skirstoma (pav.2):
lėtai metabolizuojantys (pasireiškia toksiškumas)
greitai metabolizuojantys (normalus metabolizmas)
labai greitai metabolizuojantys (nepakankamas terapinis efektas)
Pav.2. vaistų metabolizmas trijose individų grupėse ( Allorge D. ir kt., 2004).
10

Fermentų polimorfizmas svarbus, kai vaistas daugiau nei 50% metabolizuojamas
vieno fermento, kai vaistas turi siaurą terapinį langą, ar kai vaisto aktyvumas
priklauso nuo polimorfinio fermento suformuoto metabolito (Samer C.F. ir kt., 2004).
3. Genotipo ir fenotipo nustatymo metodai 3.1. Fenotipavimas
Fenotipavimo metodas paremtas tiesioginio fermento aktyvumo matavimu, arba, gana
dažnai, naudojant substrat-testą (dažniausiai vaistinį preparatą) kai tiriamas likusio
substrato ar suformuotų metabolitų kiekis, paveikus fermentu (Allorge D. ir kt.,
2004).
3.2. Genotipavimas
Metodas paremtas tiesioginiu genetinių anomalijų nustatymu, kurios lemia fermentų
ekspresijos ir jų aktyvumo įvairovę. Genotipavimo metodai sąlygoja tokių
molekulinės biologijos įrankių kaip polimerazės grandininė reakcija, panaudojimą
(Allorge D. et al., 2004).
Genotipavimas netiesiogiai susietas su fenotipavimu, nes pvz. analizuojant 1-5000000
SNP, gaunama 1-5000 klaidingai teigiamų rezultatų (Wilkins M.R. ir kt., 2000).
Taigi, fermento aktyvumas priklauso nuo jį koduojančio geno azotinių bazių sekoje
esančių polimorfizmų (mutacijų). Mutacijos
gali būti (pav.3):
A. promotoriuje (UGT1A1, CYP19)
B. introninėje dalyje (TPMT, CYP19)
C. egzone – koduojančioje srityje
(TPMT, CYP19, CYP2D6). Pav.3. supaprastinta geno struktūra
Kadangi amino rūgštys baltymuose koduojamos nukleotidų tripletais, tai SNP, esantis
koduojančioje srityje gali pakeisti amino rūgštį baltyme. Tai sąlygoja baltymo savybių
pablogėjimą arba netgi išnykimą. Pavyzdžiui, ląstelių atsparumas oksaliplatinui buvo
stebėtas pas žmones, kurie turi ERCC2 (kinta amino rūgštis 751 lys/gln pozicijoj) ir
taip pat, GSTP1 (kinta amino rūgštis 105 ile/val) geno polimorfizmą (Chaisemartin
L., 2005)
11
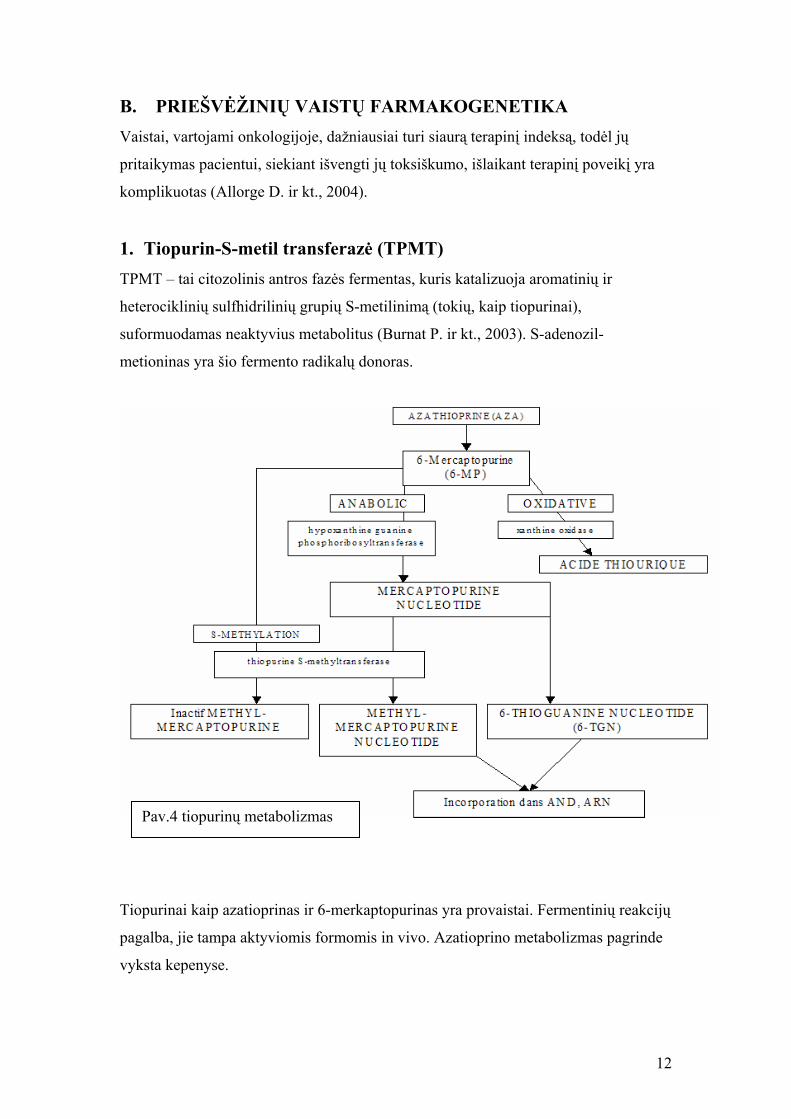
B. PRIEŠVĖŽINIŲ VAISTŲ FARMAKOGENETIKA Vaistai, vartojami onkologijoje, dažniausiai turi siaurą terapinį indeksą, todėl jų
pritaikymas pacientui, siekiant išvengti jų toksiškumo, išlaikant terapinį poveikį yra
komplikuotas (Allorge D. ir kt., 2004).
1. Tiopurin-S-metil transferazė (TPMT) TPMT – tai citozolinis antros fazės fermentas, kuris katalizuoja aromatinių ir
heterociklinių sulfhidrilinių grupių S-metilinimą (tokių, kaip tiopurinai),
suformuodamas neaktyvius metabolitus (Burnat P. ir kt., 2003). S-adenozil-
metioninas yra šio fermento radikalų donoras.
Pav.4 tiopurinų metabolizmas
Tiopurinai kaip azatioprinas ir 6-merkaptopurinas yra provaistai. Fermentinių reakcijų
pagalba, jie tampa aktyviomis formomis in vivo. Azatioprino metabolizmas pagrinde
vyksta kepenyse.
12

6-tioguanino nukleotidai (6-TGN) atsakingi už imunosupresinį ir citotoksinį tiopurinų
poveikį. Priešingai, metilinant per TPMT ar XO (ksantinoksidazę), tiopurinai
inaktyvuojami (pav.4).
1.1. Tiopurinai 6 – merkaptopurinas (6-MP) – naudojamas ūmios limfoblastinės vaikų leukemijos
atveju.
Mercaptopurine (APRD01096)
Tioguaninas – ūmiai mieloidinei ir ūmiai limfoblastinei vaikų leukemijai.
Thioguanine
(APRD00290)
Azatioprinas – naudojamas (IBD) uždegiminei žarnyno ligai, autoimuniniam
hepatitui, sisteminiam lupus erytematosus (SLE), reumatoidiniam artritui, įvairioms
odos būklėms ir organų transplantacijos metu.
Azathioprine (APRD00811)
13

1.2. Geno mutacijos Gene TPMT buvo rastos 4 mutacijos, kurios atsakingos už fermentinio aktyvumo
praradimą pas 80-95% kaukaziečių : tai aleliai TPMT*2, 3A, 3B ir 3C. Pacientai,
turintys šiuos alelius, lėtai metabolizuoja tiopurinus, todėl jiems pasireiškia šių vaistų
toksinis poveikis – mielosupresija. Tuo tarpu pacientai, kurių šio fermento aktyvumas
padidėjęs, nepakankamai gerai išgydomi. Pacientai su vidutiniškai greitu tiopurinų
metabolizmu yra heterozigotai mutavusiam aleliui, o pacientai su sumažėjusiu –
homozigotai mutavusiam aleliui (Zhou S., 2006).
1.3. Mutacijos aleliuose TPMT*2 : 238 G → C (EXON 5) TPMT*3A: 460 G → A (EXON 7) 719 A → G (EXON 10) TPMT*3B: 460 G → A (EXON 7) TPMT*3C: 719 A → G (EXON 10) (Zhou S., 2006).
TPMT genas sudarytas iš 10 egzonų ir 9 intronų (chromosoma 6p22.3)
TPMT geno aleliai (pav.5) (TAESPET, 2001):
Pav.5 TPMT aleliai, atsakingi už tiopurinų toksiškumą
14

DNR mutacijos keičia koduojamą amino rūgštį fermento sekoje, transliacijos
momentu. TPMT fermento atveju aukščiau esančiame paveikslėlyje grafiškai
pavaizduoti aleliai su juose esančiomis mutacijomis (pav.5). Tokie piešiniai
palengvina darbą modeliuojant pradmenis.
1.4. Geno TPMT seka su mutacijomis
EXON 5 11221 taagtcagtt tttcagaatt tttataaggt ttgaaaataa tatagatctg ctttcctgca 11281 tgttctttga aaccctatga acctgaattc atataaattc ctctaaatta aagaaaatat 11341 atgcttactc taatataacc ctctatttag tcatttgaaa acataattta agtgtaaatg 11401 tatgatttta tgcaggtttg cagaccgggg acacagtgta gttggtgtgg aaatcagtga 11461 acttgggata caagaatttt ttacagagca gaatctttct tactcagaag aaccaatcac 11521 cgaaattcct ggaaccaaag tatttaaggt ttgttttgat ttgggtaaat aattgtatcc 11581 atatccccac aaaagttttt ctcagtgtga gtattatgag gataccattc atgtgtccga 11641 tggttcctat ttagcacgca gattcactgt agatactata tagtataaga agcaagggct 11701 taaaaatata ggtgatagct acctaaatag gtatagacat atgtatataa aagctgaggt
EXON 7
15901 aggctgctgc cacaggctcc taaaaccatg aggggatgga cagctctcca cacccaggtc 15961 cacacattcc tctaggagga aacgcagacg tgagatccta ataccttgac gattgttgaa 16021 gtaccagcat gcaccatggg ggacgctgct catcttctta aagatttgat ttttctccca 16081 taaaatgttt tttctctttc tggtaggacaaatattggca aatttgacat gatttgggat 16141 agaggagcat tagttgccat caatccaggt gatcgcaaatggtaagtaat ttttcttttt 16201 ttgtttagct gtcttaattt tttagtatac tatacttttt ctgggttcta gaaaatcagc 16261 ttagacttct atgagtttga aataggttat tatgtttgga atttataaaa acctaaatcc 16321 aatactagct ttgtctaaaa agtataggat ttgactacag agacctgggt gtaaggctat
EXON 10 24181 tttcaccatc ttggccaggc tggtcatgaa ctcctgacct caagtgatcc acctgcctcg 24241 gcctcccaaa gtgttgggat tacaggtgtg agccaccgca cccagccaat tttgagtatt 24301 tttaaaagaa tccctgatgt cattcttcat agtattttaa catgttactc tttcttgttt 24361 caggtaaaatatgcaatata cgttgtcttg agaaggttga tgcttttgaa gaacgacata 24421 aaagttgggg aattgactgt ctttttgaaa agttatatct acttacagaaaagtaaatga
238 G → C
460 G → A
719 A → G
24481 gacatagata aaataaaatc acactgacat gtttttgagg aattgaaaat tatgctaaag 24541 cctgaaaatg taatggatga atttttaaaa ttgtttataa atcatatgat agatctttac 24601 taaaaatggc tttttagtaa agccatttac tttttctaaa aaagttttag aagaaaaaga 24661 tgtaactaaa cttttaaagt agctcctttg gagaggagat tatgatgtga aagattatgc (PUBMED - NC_000006)
Žalios spalvos sekos dalis – egzonas ; bespalvė sekos dalis – intronas; mutacija
pažymėta geltonai.
15

1.5. Nepageidaujamas tiopurinų poveikis
Pas pacientus, kurių TPMT aktyvumas sumažėjęs, 6-MP verčiamas į 6-TGN. Šie
toksiški metabolitai kaupiasi ląstelėse ir lemia ryškų hematotoksiškumą :
leukoneutropeniją, trombopeniją, kaulų čiulpų aplaziją. (Welti M. ir kt., 2001).
2. Uridin-difosfat-gliukuronoziltransferazė (UGT1A1)
UGT1A1 – tai antros fazės fermentas. Jis metabolizuoja estrogenus, prijungdamas
gliukurono rūgštį. Hormonas praranda sugebėjimą jungtis su savo receptoriumi
(Duguay Y. ir kt., 2004). Jis taip pat atsakingas ir už bilirubino konjugaciją (Gilbert`o
sindromas) (O'Dwyer ir kt., 2006). Taip pat už kai kurių vaistų metabolizmą, kaip
irinotekanas.
2.1. Geno mutacijos
Buvo nustatyta, kad UGT fermento kiekis yra reguliuojamas transkripcijos
reguliacijos metodu (promotoriumi) ir, kad tam tikrų grupių kiekis promotoriuje
tiesiogiai veikia transkripciją. UGT atveju promotoriuje buvo rasta TA grupė, kuri
įvairuoja nuo 5 iki 8 : kai TA yra mažiau, tai geno transkripcijos efektyvumas yra
didesnis. Pvz. : Gilberto sindromas – žmogus heterozigotas TA7 (alelis –
UGT1A1*28) (Monaghan G. ir kt., 1996).
2.2. UGT1A1 TATA-BOX Mutacijos detekcijos skirtumai genui UGT1A1
Gene UGT1A1 mutacija nustatoma “matuojant” promotoriaus DNR sekos ilgį, kuris
priklauso nuo TA grupių promotoriuje : pvz TA6/TA7 (viena DNR grandinės dalis
ilgesnė už kitą).
Principas : daroma polimerazės grandininė reakcija, naudojant pradmenis, žymėtus
fluoroforais 5` pozicijoje. Padaroma daug promotoriaus srities, kurioje yra TA grupės
16

(lemiančios transkripciją) kopijų. Denatūruojama kaitinant ir staigiai šaldant.
Denatūravimo metu perskiriama dvigrandė DNR. Pridedama specifinio masės
žymeklio (100-600 bazių porų mišinys). Denatūratas leidžiamas per chromatografines
kolonėles. Matuojama fluorescencija. Sekvenavimo aparatas brėžia pikus,
priklausomai nuo to, kokio ilgio fragmentai pereina per kolonėlę : 1 pikas –
homozigotas, 2 pikai – heterozigotas.
2.3. Topoizomerazės – 1 inhibitoriai
Natūralus CAMPTOTECINAS (campto)
Tai medžio camptotheca acuminate derivatų mišinys. Jie turi topo-1 inhibitoriaus
savybių. (topo-1 yra fermentas, kuris kontroliuoja ir modifikuoja DNR genetinės
medžiagos replikacijos ir transliacijos metu). (CPA, 2006).
Topotekanas
Topotecan D.B. (APRD00687)
Tai pusiausintetinis vaistas, campto analogas. Topotekanas
jungiasi prie TOPO1-DNR komplekso, stabdydamas DNR
molekulės religaciją (DrugBank, APRD00687). Topotekanas,
priklausomai nuo pH yra dvejose formose : laktono ir
karboksilato. Aktyvus yra laktonas. Topotekanas pašalinamas
per kepenis ir inkstus (D.Armstrong ir kt., 1998). Vaistas
naudojamas antrosios linijos kietųjų navikų terapijoje,
pagrinde metastatinėms kiaušidžių karcinomoms, plaučių
vėžiui, esant įvairioms kraujo vėžio formoms (D.Armstrong
ir kt., 1998).
Irinotekanas (CPT-11)
Tai pusiausintetinis vaistas. Campto analogas. Tai provaistas.
Topo-1 inhibitorius (SN-38 ir irinotekanas jungiasi prie topo-
1-DNR komplekso ir stabdo grandinės religaciją) (CPA,
2006).
SN-38, priklausomai nuo pH yra 2 formų : laktono ir karboksilato. Irinotecan D.B. (APRD00579)
Tik laktonas aktyvus. Irinotekanas pagrinde pašalinamas per
kepenis (O'Dwyer ir kt., 2006).
17

Šis vaistas naudojamas kietųjų navikų, ypač kolorektalinio ir plaučių vėžio gydymui
(D. Allorge ir kt., 2004), o taip pat įvairiems hematologiniams vėžiams (K.Seiter,
2005).
2.4. Metabolizmas (pav.6)
APC – AMINOPENTANO CARBOXY ACIDE NPC – 7-ETIL-10(4(1-PIPERIDINO)-1-AMINO)-CARBONYLOXYCAMPTOTECINE CPT-11 – IRINOTECAN SN-38 – 7-ETIL-10-HIDROCAMPTOTECINE SN-38-G - 7-ETIL-10-HIDROCAMPTOTECINE GLUCURONIDE UGT1A1 – UDP GLUCURONOSYLTRANSFERASE B-GLUC – BETA GLUCURONIDASE CYP3A4 – CYTOCHROME P-450 NF-KB – FACTEUR NUCLEAIR KAPPA CMOAT – FACTEUR CANNALICULIER DU TRANSFERT
Pav.6 irinotekano metabolizmas (Thorn C. F. ir kt., 2005).
18

2.5. Nepageidaujamas vaistų poveikis Kamptotecinai sukelia mielosupresiją (anemija, neutropenija, trombocitopenija,
leukopenija) (Armstrong D. et al., 1998). Topotekanas turi silpną ekstrameduliarinį
toksinį poveikį pacientams su sumažėjusia inkstų filtracija (Seiter K., 2005).
Irinotekanas gali sukelti diarėją, mielosupresiją, žarnyno epitelio apoptozę (Yang X. ir
kt., 2006).
Dėl irinotekano toksiškumo yra ginčijamasi. Yra dvi nuomonės :
1. Vieni kalba, kad UGT1A1 geno polimorfizmas yra atsakingas tik už
hematotoksiškumą (leukopenija, neutropenija) : jie randa neutropeniją pas
individus, homozigotus ar heterozigotus genui UGT1A1*28, bet neranda
jokio sąryšio su diarėja (McLeod H.L., 2006).
2. Kiti pabrėžia, kad pastarojo geno mutacijos lemia ir diarėją : ūmi diarėja
buvo stebėta pas pacientus, heterozigotus (4 laipsnio diarėja) ir
homozigotus (3 laipsnio diarėja/4 laipsnio neutropenija) (TA)7TAA sekai
(Iyer L. ir kt., 2002).
UGT1A1 genotipavimas, prieš pradedant chemoterapiją, leidžia aptikti individus,
turinčius polinkį į šio vaisto sukeltus ūmius toksinius efektus (Allorge D. ir kt., 2004).
3. Aromatazė Aromatazė (citochromas P450) – tai fermentas, kuris katalizuoja C18 steroidų
konvertavimą į estrogenus. Aromatazės raiška vyksta tam tikruose audiniuose kaip
kiaušidės, placenta, sėklidės, smegenys, oda, o taip pat ir patologijos apimtuose
audiniuose, kaip krūtys, vėžio apimtuose endometriumo audiniuose (Shozu M. ir kt.,
1998).
Aromatazė koduojama vieno geno, kuris yra 15q21.2 ir kurio ilgis 123 kbp
(tūkstančiai bazių porų). Genas sudarytas iš 9 (10) egzonų : 2-10. Egzonas 1
nekoduojantis (Haiman C.A. ir kt., 2003).
Aromatazė – tai sudėtinis fermentas. Aromatazės kompleksas sudarytas iš 2
polipeptidinių dalių : NADPH-citochromo P450 reduktazės ir ubikvitinflavoproteino.
C18 Androgenų konversija į C19 estrogenus katalizuojama 3 hidroksilinimo reakcijom
(Mahendroo M.S. ir kt., 1993) (pav.7, pav.8).
19

Pav.7. Konversijos reakcija (Kao Y.C. ir kt., 2001)
3.1. Aromatizacijos reakcijos mechanizmas
Aromatazė – aktyvusis centras ir hemas
Pav.8. estrogenų sintezė. A- hidroksilinimas B- aromatizacija (Kao Y.C.ir kt., 2001)
20

3.2. Geno mutacijos
Padidinta fermento ekspresija buvo pastebėta vėžio apimtuose krūtų audiniuose. Buvo
rastas I.4 promotoriaus (specifinis adipocitų, gliukokortikoidų stimuliuojamas)
pakeitimas proksimaliniu II promotoriumi, kuris kontroliuoja kiaušidžių aromatazės
ekspresiją ir promotoriumi I.3.
Labiausiai žinomas polimorfizmas – (TTTA)N tetranukleotido pasikartojimas I.4 –
ame. Šiuo metu žinoma, kad tetranukleotido pasikartojimas nėra dominantinis
faktorius daugumoje atvejų, kai padidėjęs aromatazės aktyvumas (Haiman C.A. ir kt.,
2003). Variacija C790T (C826T) egzone 7 (8) keičia amino rūgštį Arg264Cys (Hirose
K. ir kt., 2004). Variacija C602T egzone 5(6), koduojanti Thr201Met, yra labai svarbi
kaukaziečių krūties vėžio atveju (0.05) (Ma C.X. ir kt., 2005).
Egzone 2 esanti variacija, koduojanti Trp39Arg yra susijusi tik su japonų ir havajiečių
krūties vėžiu (Miyoshi Y. ir kt., 2000).
Polimorfizmų variacija (pav.9)
pav.9 Geno CYP19 mutacijos (Ma C.X. ir kt., 2005)
21

3.3. Vaistai Yra 2 struktūriniai aromatazės inhibitorių tipai : steroidiniai, substrato tokio kaip 4-
hiroksiandrostenedionas (formestanas) ir eksemestanas analogai (pirmas tipas) ir
nesteroidiniai, kurie jungiasi su prostetine citochromo P450 hemo grupe (antras tipas).
Azoto atomas antro tipo vaistų struktūroje leidžia jungtis su geležies atomu (Kao ir
kt., 1996). Antro tipo inhibitoriai yra svarbesni nei pirmo, nes jie inaktyvuoja
fermentus (Lonning P.E., 2000). Fermentas konvertuoja vaistą, vaistas negrįžtamai
jungiasi prie fermento kovalentinėmis jungtimis (Dowsett M. ir kt., 1987).
Vaistai vartojami pagrinde krūties vėžiui gydyti.
Aminogliutetimidas – grįžtamas konkurencinio tipo pirmos kartos aromatazės
inhibitorius. Nenaudojamas šiuo metu, nes sukelia daug šalutinių reakcijų. Jungiasi su
fermentais ir slopina hidroksilinimo reakcijas (Patrick G.L., 2005).
Aminogluthetimide (Patrick G.L., 2005)
Formestanas (4-hidroksiandrostendionas) – antros kartos vaistas. Vartojamas tik
intramuskuliariai, todėl nepopuliarus (Fentiman I.S., 2004).
Formestane (Patrick G.L., 2005)
22

Anastrozolis ir letrozolis – trečios kartos grįžtamo konkurencinio tipo inhibitoriai,
labiau selektyvūs CYP 19 (Patrick G.L., 2005)
Anastrozole D.B. (APRD00016)
Letrozole D.B.(APRD01066)
Eksemestanas – trečios kartos grįžtamas konkurencinio tipo inhibitorius. Jis yra
steroidinės struktūros. Jungiasi 2,6 karto stipriau nei natūralus substratas
androstendionas (Coombes R.C. ir kt., 1999).
Exemestane D.B.(APRD00144)
Trečios kartos vaistai yra peroraliniai, naudojami, kai navikas atsparus hormono-
terapijai, postmenopauzinis, metastatinis (Hamilton A. ir kt.,1999).
3.4. Pašaliniai poveikiai Endometriumo vėžys, kraujavimas iš makšties, tromboembolija, ūmi venų trombozė,
širdies-kraujagyslių išemija, skeleto raumenų skausmas, kaulų lūžis, artritas, artralgija
(Arora A. ir kt., 2004).
23

C. DARBO POLIMERAZĖS GRANDININĖS REAKCIJOS
METODU PRINCIPAS DNR (DEZOKSIRIBONUKLEINO RŪGŠTIS) DNR – tai daugumos gyvybinių formų genetinė informacija. Tai polimeras, sudarytas
iš struktūrinių vienetų, vadinamų nukleotidais (mononukleotidais) (Huret J.L., 2006).
1. DNR ekstrahavimo principai DNR ekstrahavimas iš biologinės medžiagos reikalauja ląstelės lizės, ląstelės
nukleazių inaktyvavimo ir DNR atskyrimo nuo liekanų. Ląstelių suardymo procedūra
turi būti atitinkamai suderinta, kad tinkamai ir efektyviai suardyti ląstelę, bei
neprarasti DNR.
Lizės procedūros:
mechaninis suardymas (hipotoninė lizė)
cheminis suardymas (lizė detergentais, chaotropiniais agentais, redukcija
tioliais)
suardymas fermentais (proteinazė)
Membranos suardymas ir nukleazių inhibavimas dažnai suderinami (Somma M. ir kt.
2004).
2. DNR ekstrahavimo metodai Praktikoje DNR ekstrahavimas dažniausiai daromas iš žmogaus kraujo. Naudojama
keletas ekstrahavimo technikų:
1. ekstrahavimas fenoliu-chloroformu
2. ekstrahavimas natrio chloridu arba acetatu
3. komercinių rinkinių, kaip Qiagen panaudojimas
Metodai pritaikomi prie kiekio, kokybės ir kitokių biologinio objekto faktorių
(Francina A., 2003).
2.1. Ekstrahavimas natrio chloridu (druskinis metodas) Raudonųjų kraujo kūnelių lizė
Ši operacija atliekama, kad pašalinti hemoglobiną, kuris stabdo PGR. Lizė atliekama
panaudojant specialų buferinį tirpalą, skirtą raudoniesiems kraujo kūneliams lizuoti
24

(0,155M NH4Cl, 0,01M KHCO3, 0,001M EDTA), kuris sudaro hipotoninę terpę
(Bienvenu T. ir kt., 1999). EDTA jungiasi su Mg ir mažėja laisvo Mg kiekis, nes
didelis Mg kiekis didina DNR praradimo nuostolius (Lahiri DK ir kt., 1993).
Baltųjų kraujo kūnelių lizė
Pridedama buferinio tirpalo, skirto baltiesiems kraujo kūneliams lizuoti (0,4M NaCl,
0,01M trishidroksimetilaminometano, 0,002M EDTA). Baltieji kraujo kūneliai suyra.
Branduolio DNR atpalaiduojama į terpę (Bienvenu T. ir kt., 1999).
Proteinų ardymas
Pridedama fermento proteinazės, kad lizuoti proteinus, kurie gali būti prisijungę prie
DNR. Pridedama SDS 10% tirpalo (natrio duodecil sulfato), kad destabilizuoti lipido-
proteinines membranas ir sensibilizuoti nelizuotas ląsteles lizuojantiems reagentams
(Kang T.J ir kt., 2004).
Ląstelės liekanų atskyrimas
Pridedama NaCl, kad precipituoti liekanas, kurios praranda savybę tirpti vandenyje,
prisotintame stipraus elektrolito.
DNR precipitavimas
Pridedama 2 tūriai 100% etanolio, laikyto esant -200C (Kang T.J ir kt., 2004).
Precipitacija greitinama šaldant. Po to daromas DNR plovimas 70% etanoliu. Darant
plovimą, prarandamas tam tikras kiekis DNR. (Kang T.J ir kt., 2004).
DNR tirpinama mažame buferinio tirpalo kiekyje (TRIS, EDTA – slopina DNR`azes,
esančias tirpale).
2.2. Ekstrahavimas panaudojant DEAE polimerus Metodas turi privalumų, nes galima pašalinti tokias atliekas kaip hemoglobiną,
plazmos proteinus arba jonus (geležies, mangano ir kt.) (Bienvenu T. ir kt., 1999).
Ląstelių lizė
Pirmiausiai kraujas maišomas su lizės buferiniu tirpalu (NaOH/SDS/guanidino
tiocianatas). Guanidino tiocianatas – tai chaotropinis agentas, kuris lengvai tirpina
proteinus (Vallejo L.F. ir kt., 2004). Pridedama proteinazės K, kuri yra pakankamai
efektyvi, nes po lizės, kuri vykdoma esant 700C, dauguma proteinų lizuojami
(Dahlgren L., 2005).
Po ląstelių lizės pridedama etanolio, kad precipituoti DNR ir pagerinti sąlygas DNR
fiksacijai ant DEAE polimerų.
25

DNR fiksavimas ant DEAE polimerų
Centrifūguojamas mišinys. DNR fiksuojasi ant membranos, sudarytos iš specifinių
polimerų (dietilaminoetanolio darinys). Polimerai turi 100µm dydžio daleles, dideles
poras ir hidrofilinį paviršių. DNR fosfatai turi neigiamą krūvį, o polimerai – teigiamą
(pav.10). Reikiamos molekulės fiksavimas ir eliuavimas priklauso nuo NaCl
moliaringumo ir buferinio tirpalo pH. Nešvarumai, tokie kaip proteinai, valomi
panaudojant silpnas elektrolitų koncentracijas (pav.11).
Pav.11 eliuavimas skirtingom druskų koncentracijom
Pav.10. fiksavimo molekulinis mechanizmas
Plovimas
Vykdomas panaudojant buferinį tirpalą, kuriame NaCl koncentracija yra didelė 1,25M
(papildomai yra (CH3COO)2KH, kuris neutralizuoja NaOH).
Po to druskos pašalinimas ir precipitacija vykdoma su izopropanoliu ir natrio azidu
(0.25 V/V NaN3), kuris skirtas apsaugoti nuo DNR gedimo (Nicole T. Vu ir kt.,
1999).
DNR išplovimas iš kolonėlės
Pagaliau DNR eliuojama panaudojant eliuavimo buferinį tirpalą (10mM TRIS-Cl,
0,5mM EDTA, pH 9). Ši technika leidžia išgauti 90-95% pradinės DNR kiekio ir
pašalinti fragmentus, kurie mažesni nei 100 bazių porų (Cere N., 1998).
3. DNR kiekio nustatymas spektrofotometriškai DNR, RNR, oligonukleotidų ir netgi mononukleotidų kiekis gali būti nustatytas
vandeniniuose tirpaluose, matuojant tirpalo absorbciją (optinį tankį) ultravioletinių
26

bangų ilgyje. Pagrindinę funkciją atlieka purino bazės, kurios ir absorbuoja
matavimams naudojamo bangos ilgio spindulius.
Kai pavyzdys neužterštas tokiomis medžiagomis kaip baltymai, fenolis, agarozė,
optinio tankio (absorbcijos) matavimas yra labai paprasta ir tiksli operacija.
Vietoj gryno vandens dažnai naudojami buferiniai tirpalai (TBE). Nukleino rūgščių
nustatymas vykdomas panaudojant pagrinde 260nm bangos ilgio spindulius.
Priemaišos nustatomos matuojant absorbciją tirpalo prie 260nm ir lyginant su
absorbcija, matuojant kito bangos ilgio spinduliais, leidžiančiais išmatuoti aktualias
priemaišas (pvz. baltymams tai 280nm). Imamas santykis A260/A280. Jei DNR gryna,
tai santykis turi būti apie 1,8, o RNR apie 2,0. Matuojant absorbciją esant 230nm,
nustatomas užteršimas tokiais teršalais kaip angliavandeniliai, fenoliai, peptidai,
aromatiniai dariniai (Somma M., 2004).
4. DNR PLAZMOJE BEI SERUME
4.1. DNR nustatymas plazmoje DNR plazmoje cirkuliuoja pas sveikus ir ligonius. Yra aprašyta keletas metodų, kai
DNR ekstrahuojama iš 20-250µl plazmos. Rašoma, kad šios mikroekstrakcijos leidžia
amplifikuoti fragmentus iki 3789 bazių porų, panaudojant PGR. Gauti produktai
tokios pačios kokybės kaip ir gauti panaudojant ekstraktus, padarytus iš kraujo
(Andolfato S. ir kt., 2003).
4.2. Padidintas plazmos DNR kiekis pas pacientus, sergančius
onkologinėmis ligomis Onkologinėmis ligomis sergančių pacientų plazmoje ir serume dažnai aptinkama
dvigrandės DNR fragmentų. Buvo nustatyta, kad pas sveikus individus serume yra
13ng/ml laisvos DNR, kai tuo tarpu, pas onkologinėmis ligomis sergančius pacientus
aptinkama vidutiniškai 180ng/ml. Buvo padaryta išvada, kad šis genetinės medžiagos
kiekis yra navikų kilmės (Jahr S. ir kt., 2001).
5. DNR izoliavimas ir konservavimas DNR gali būti išskirta iš įvairių objektų:
1. kraujo, kuriame yra antikoaguliantų
2. koaguliavusio kraujo
27

3. sušaldytų leukocitų
4. sušaldytų limfocitų
5. ląstelių limfoblastoidinių linijų
6. burnos epitelio ląstelių ir kt.
Pagrinde, esant pakankamai medžiagos, naudojamas kraujas su antikoaguliantu, nes
patogiausia vykdyti ekstrahavimą. Patariama DNR išekstrahuoti 2 savaičių
laikotarpyje (Francina A., 2003).
6. POLIMERAZĖS GRANDININĖ REAKCIJA (PGR)
PGR išrasta 1983 metais K. Mullis. Principas paremtas DNR polimerazės
panaudojimu, kai vykdoma replikacija in vitro tam tikro DNR sekos fragmento. Per
kelias valandas galima sintezuoti 106 kopijų tam tikro matricos DNR fragmento.
Vienintelė sąlyga yra ta, kad būtų sekos dalis, kurią norima replikuoti, matricinėje
DNR. Teoriškai, matricinės DNR kiekis nėra limituojantis faktorius (Siatka Ch.,
2006; Kubista M. ir kt., 2006).
6.1. Principas Imama pora oligonukleotidinių pradmenų, kurie komplementarūs tiriamai DNR sekai,
bet nekomplementarūs viens kitam (Mullis K. ir kt., 1986 ; Guatelli J.C. ir kt., 1989).
Pradmenų ilgis 16-30 nukleotidų. Veikiant DNR polimerazei, kiekvienas pradmuo,
kuris prisijungęs prie viengrandės DNR, ilgėja 3’-5’ kryptimi, komplementariai sekai.
Kopijų kiekis didėja eksponentiškai (Wahbi K., 1996).
Vartojamas termostabilus fermentas, kuris tampa aktyviu tik po tam tikro sužadinimo
padidinta temperatūra (Kubista M.ir kt., 2006). Fermentas yra aktyvus tam tikrą kiekį
PGR ciklų (dažniausiai tai 35 ciklai) (Guatelli J.C. ir kt., 1989).
6.2. ETAPAI (pav.12)
Pradinės sąlygos
Kambario temperatūroje dvigrandė DNR yra dvivijės struktūros.
Pradinis denatūravimas
Pirmas reakcijos etapas – DNR denatūravimas, kaitinant (Guatelli J.C. ir kt., 1989).
Šis etapas leidžia išvynioti dviviję DNR, išardyti antrines struktūras, homogenizuoti
28

reakcinį turinį, termiškai maišant, aktyvuoti “hot start’ polimerazes, denatūruoti kitus
fermentus, kurie galėtų būti terpėje (atvirkštinė transkriptazė) (Wikipedia).
6.2.1. Denatūravimas
Pav.12. PGR. (Guatelli J.C. ir kt., 1989)
Šis etapas leidžia galutinai suardyti DNR struktūrą,
“atplėšti” polimerazes, kurios buvo prisijungusios
prie matricos ir homogenizuoti reakcijos terpę
(Kubista M. ir kt., 2006).
6.2.2. Hibridizacijos fazė Pradmenys šio etapo metu jungiasi su viengrande
DNR matrica, kuri perskirta denatūravimo metu.
Pradmenys jungiasi tik tuo atveju, jei temperatūra
yra atitinkama:
jei ji per žema, tai pradmuo gali jungtis
nespecifiškai, gali būti “susirietęs” dėl galimo
komplementarumo (A-T, G-C) tarp pradmens
nukleotidų rinkinių ir kt.
Jei per aukšta, tai pradmuo gali nesijungti su
matrica dėl per silpnų jėgų.
Kuo didesnis kiekis G-C nukleotidų, tuo hibridizacijos temperatūra didesnė, kuo
daugiau A-T, tuo temperatūra mažesnė. Dalis pradmenų gali susijungti vieni su kitais,
bet ne su matrica, nes jų kiekis reakcijos terpėje ženkliai didesnis (Kubista M. ir kt.,
2006; Mullis K. ir kt., 1986).
6.2.3. Elongacijos fazė Šis etapas polimerazei leidžia sintetinti komplementarią matricinei DNR grandinę,
esant optimaliai temperatūrai. Ši grandinė sintetinama iš mononukleotidų, kurie
pridedami į terpę. Šio etapo trukmė priklauso nuo sintetinamo fragmento ilgio
(Kubista M. ir kt., 2006).
6.3. DNR kiekio didėjimas Kartojant denatūravimą, hibridizaciją ir elongaciją, DNR kiekis didėja eksponentiškai
(Mullis K. ir kt., 1986).
29

6.4. Pradmenys polimerazės grandininei reakcijai Kad empyriškai nustatyti pradmenų hibridizacjos temperatūrą, pasitelkiamos
formulės:
1. Tm=4(G+C)+2(A+T)*0C
Ši formulė tinka tokiu atveju, jei tai taisyklingi dupleksai, sudaryti iš 11-20 bazių
porų nukleotidų. Esant didesniam kiekiui nukleotidų duplekse, temperatūra
mažėja apie 5-100C kas nukleotidas.
Praktikoje naudojama 2-50C aukštesnė temperatūra, nei apskaičiuota panaudojant
formulę (Wahbi K., 1996).
2. Mc Conaughy formulė
Td=81,5-16,6(log(Na+))+0,41%(G-C)-(600/N)
Ši formulė naudojama, kai dupleksai didesni nei 20 bazių porų ir:
G ir C nukleotidai
N – bendras pradmens azotinių bazių skaičius (Wahbi K., 1996).
6.5. lizdinės PGR metodas Fragmentas, gautas pirmos PGR metu (su vienais pradmenimis) yra gausinamas dar
kartą su pradmenimis, kurie jungiasi fragmento vidinėje dalyje (pav.13). Išauga
metodo jautrumas ir specifiškumas, nes įvykdomos dvi viena paskui kitą sekančios
PGR. Sumažėja veiksnių, kurie inhibavo pirmąją PGR (Durand G. ir kt., 2006)
(Mullis K. ir kt., 1986).
30Pav.13. lizdinė PGR

6.6. PGR produktų valymas Atlikus PGR-ją, daromas PGR-jos produktų valymas, kadangi tirpale kartu su
galutiniu produktu yra nesureagavusių reagentų bei šalutinių produktų (polimerazė,
pradmenys, mažos molekulinės masės PGR šalutiniai produktai) (pav.14).
Valymas dažniausiai atliekamas panaudojant komercinius rinkinius. Valymo principas
remiasi tuo, kad amplifikuota DNR molekulė yra santykinai didesnė už kitas mišinyje
esančias molekules. Taigi, pridėjus papildomai vandens į mišinį, kad būtų galima
geriau išplauti ir patirti mažesnius DNR nuostolius, tirpalas leidžiamas per
membranas, kuriose yra tam tikro dydžio poros : santykinai mažesnės molekulės
praeina pro šias poras, o tokios kaip 100bp DNR – pasilieka ant membranos (pav.15).
DNR, esanti ant membranos yra tirpinama vandenyje ir gaunamas grynos DNR
tirpalas (Millipore, 2007).
Pav.15 PGR produkto
Pav. 14 PGR mišinys
6.7. Gel-elektrofarezė Elektrofarezė – tai krūvį turinčių molekulių judėjimas elektriniame lauke. DNR ir
RNR molekulė yra įkrauta teigiamai, todėl elektriniame lauke juda link neigiamo
elektrodo.
Gel-elektrofarezė – DNR bei RNR migruoja tarp gelio porų. Molekulių migravimo
greitis priklauso nuo molekulės dydžio ir gelio porų kiekio bei dydžio (Brown T.A.,
2006). Metodas leidžia atskirti DNR ir RNR molekules priklausomai nuo jų dydžio
(Sinniger S., 2001).
6.8. Nukleino rūgščių vizualizavimas Atliekamas veikiant etidiumo bromidu, kuris dedamas į gelį. Etidiumo bromidas
įsiterpia į nukleino rūgščių struktūrą ir, veikiant UV spinduliais (200-300nm), spindi
oranžine spalva, tuo pačiu leisdamas vizualizuoti ir nukleino rūgštis.
31

Norint žinoti gauto produkto molekulės dydį, į gelį, šalia tiriamo produkto dedamas ir
standartinis molekulinis markeris. Migruojama tam tikrą laiko tarpą. Ir pagal
molekulinio markerio fragmentų išsidėstymą gelyje nustatomas tiriamo produkto
molekulės dydis. Gali būti nustatomas ir produkto kiekis, dedant atitinkamą kiekį
molekulinio markerio ir tiriamo produkto. Pastaruoju atveju lyginamas
fluorescencijos intensyvumas (INRP, 2001).
6.9. Sekvenavimas DNR sekvenavimas – tai nukleotidų grandinės nustatymas (Delarue M. ir kt., 2005).
Principas
Sekvenavimui naudojami fermentai – DNR polimerazės. Reakcijos vykdomos esant
panašios komponentinės sudėties reakcijos mišiniui kaip ir PGR, tačiau pridedama
didezoksiribonukleotidų (ddNTP), kurie yra pažymėti fluoroforu. Jų dedamas labai
nedidelis kiekis. Taigi, DNR polimerazė, kaip ir PGR reakcijos metu sintetina DNR
grandinę, naudodama dNTP (nukleotidus), kol prie grandinės nepridedamas ddNTP.
ddNTP sustabdo reakciją (pav.16). ddNTP, pakeisdamas dNTP, leidžia ‘perskaityti’
seką.
Reakcijos mišinyje yra 4 tipų ddNTP, kurių bazės : adeninas, citozinas, guaninas,
timinas. Visi jie žymėti skirtingais fluoroforais ir, paveikus tam tikro bangos ilgio
spinduliais, fluorescuoja skirtingo bangos ilgio spindulius (Delarue M. ir kt., 2005).
Pav.16. DNR sekvenavimas
Automatiniais sekvenatoriais “perskaitoma” seka. Gauti fragmentai išskirstomi pagal
ilgį, panaudojant chromatografiją kolonėlėse. Sekvenatorius “perskaito” seką, tuo
32

pačiu nustatydamas ir fragmento ilgį. Rezultatas vaizduojamas kreivių pagalba
(pav.17), lyginant fluorescencijos pobūdį ir intensyvumą (Delarue M. ir kt., 2005).
Pav.17 nuskaityta seka
D. RNR – ribonukleinė rūgštis
Pav.18. RNR vaidmuo baltymų sintezėje
1. RNR ekstrahavimo metodas Etapai :
1) Ląstelių lizė guanidino tiocianatu ir beta merkaptoetanoliu. Guanidino
tiocianatas denatūruoja baltymus, padeda atskirti branduolyje esančias
nukleino rūgštis nuo proteinų. Beta merkaptoetanolis soliubilizuoja proteinus,
perskirdamas inter ir intramolekulines S-S jungtis, kurios gali susidaryti po
kontakto su deguonimi (Guillaume G. ir kt., 1978 ; Vallejo L.F. ir kt., 2004).
2) RNR precipituoja veikiant 70% etanoliu.
33

3) Vėl pridedama guanidino tiocianato, kad pašalinti nepašalintus baltymus.
Pridedama DNR`azės, siekiant suskaidyti DNR (nes reikia gauti tik RNR).
4) Vykdomas eliuavimas, panaudojant RNR`se free vandenį (vanduo iš kurio
pašalinta RNR`azė, nes ji skaido RNR). Šiame vandenyje yra nedidelė
koncentracija NaCl.
Visas vanduo, naudotinas darbui su RNR, turi 0,1% DEPC (dietilpirokarbonatas).
DEPC inhibuoja RNR`azę ją denatūruodamas.
2. Atvirkštinės transkripcijos reakcija Kad sintezuoti kDNR, naudojamas fermentas, rastas pas retrovirusus : atvirkštinė
transkriptazė. Jis sintezuoja komplementarią RNR matricai viengrandę DNR (Lodish
H. ir kt., 2004).
Mišinys, naudojamas atvirkštinės transkriptazės reakcijoje, turi turėti 2 pagrindinius
elementus :
• Oligo(dT), kuris jungiasi su poly(A) RNR dalimi : 3` padėtyje yra 1-2 ne
timinai. Šis oligonukleotidas duoda startą fermentui veikti.
• Pasirenkamieji pradmenys (6-9 bazių dariniai), kurie gali būti visokiausių
galimų sekų variantas, besijungiantys įvairiose RNR sekos vietose. Šie
pradmenys būtini, kad fermentas galėtų sintetinti ilgos grandinės fragmentus
(Kubista M. ir kt., 2006).
Mišinyje naudojamas DTT (ditiotreitolis), kad
pašalinti S-S jungtis ir inhibuoti RNR`azę
(Carlsson J. ir kt., 1978).
Buferinis tirpalas, kad pritaikyti terpę
fermentui veikti (250 mM Tris-HCl, pH 8,3 ;
375mM KCl ; 15mM MgCl2).
Įvykdžius reakciją, iRNR yra išplaunama šarmu, kuris ją hidrolizuoja. Gautos
viengrandės DNR molekulės konvertuojamos į dvigrandę DNR, vykdant elongaciją
nuo 3` veikiant terminalinei transferazei (specifinė polimerazė), į reakcijos mišinį
pridedama deoksinukleotidų. Sintetinis oligo dC jungiasi su 3`esančiu oligo dG. Oligo
polimerazė sintetina kDNR. Pastaroji polimerazė dC naudoja kaip startą. Taigi,
34

kiekviena kDNR turi viename gale oligo dC – oligo dG sritį ir oligo dT – oligo dA
sritį kitame (Lodish H. ir kt., 2004 ; Guatelli J.C. ir kt., 1989).
3. Geno ekspresijos matavimas Metodas paremtas informacinės RNR
ekspresijos matavimu (pav.19). Atvirkštinės
transkripcijos būdu gaunamas kDNR kiekis
atitinkantis iRNR kiekį (Kubista M. ir kt.,
2006).
Pav.19 ekspresijos matavimas
3.1. Metodas PGR reakcijos realiu laiku principas
• PGR reakcijos produktų eksponentinis didėjimas
PGR produktų kiekis didėja pagal šią formulę :
N = N0*2n
N – reakcijos pabaigoje gautas molekulių kiekis
N0 – Matricos kiekis reakcijos pradžioje
n – ciklų skaičius (Tse C. ir kt., 2003).
• Reakcijos naudingumas
Eksperimento metu gauto produkto kiekis priklauso nuo amplifikacijos naudingumo
faktoriaus (E). Reakcijos metu gali būti : 0<E<1 (0 – nesintezuojama DNR ; 1-
sintezuojamos 2 DNR po 1 ciklo) (Tse C. ir kt., 2003).
Reakcijos pabaigoje gaunama :
N=N0(1+E)n
logaritminės formos : Log N = Log N0 + nLog (1 + E) (Kubista M. ir kt., 2006).
• PGR reakcijos “PLATO” fazė
PGR reakcijos produktų kiekio eksponentinis didėjimas yra matomas tik po 20-35
ciklų. Reakcijai artėjant link pabaigos, sulėtėja reakcija ir produktų akumuliacija,
35

kadangi pasiekiamas limitas. Tai “PLATO” efektas. Paskutiniųjų ciklų metu DNR
polimerazė inaktyvuojama, mažėja dNTP ir pradmenų, todėl lieka tik reikalingi
galutiniai produktai, (Tse C. ir kt., 2003).
Ciklas
PGR realiu laiku principas tas, kad reakcijos metu, ciklas po ciklo, matuojama
fluorescencija. Fluorescencija (pav.20) kinta priklausomai nuo sintezuoto produkto
kiekio, nuo panaudotų pradmenų, įsiterpiančių į DNR medžiagų (SYBRGREENTM) ir
kt.
Taigi, lyginant fluorescenciją ciklas po ciklo, galima nustatyti gautų produktų kiekį.
Fluorescencijos intensyvumas vaizduojamas kreivėje (Tse C. ir kt., 2003).
Pav. 20. 3 fazės : iniciacijos, eksponentinė ir plato
3.2. Detekcijos “realiu laiku” sistemos Medžiagos, įsiterpiančios į dvigrandę nukleotidų seką
Šios medžiagos įsiterpia į dvigubą nukleotidų grandinę ir fluorescuoja specifinio
bangos ilgio spindulius. Tai pati paprasčiausia detekcijos “realiu laiku” sistema.
Yra keletas komercinių produktų, kaip SYBRTM Green I ar BOXTOTM , kurie įsiterpia
į dvigrandę DNR, tačiau gali įsiterpti į trumpus DNR fragmentus, į susijungusius
pradmenis (pradmenų dimerus) ir skleisti parazitinę (klaidinančią) fluorescenciją.
Todėl metodas nėra labai tikslus (Tse C. ir kt., 2003).
36

3.3. Strategijos Nukleino rūgšties kiekis gali būti matuojamas absoliutiniu metodu, po etaloninės
kreivės gavimo, panaudojant išorinį homologišką standartą arba reliatyviuoju metodu,
lyginant su genu, kuris yra validuotas, bei lyginant su kalibruojančiuoju pavyzdžiu.
Absoliutinis metodas simbolizuoja : kopijų kiekis/µL arba koncentracijos vienetai/µL
Reliatyvusis : koncentracija tiriamojo/koncentracija validuoto (Tse C. ir kt., 2003).
37

II. EKSPERIMENTINĖ DALIS
A. KRAUJO, PLAZMOS BEI SERUMO GENOTIPAVIMAS
1. Medžiagos ir instrumentai 1) DNR :
a. DNR iš 2 pacientų plazmos ir kraujo. DNR iš kraujo ekstrahuota
normaliomis sąlygomis ; DNR iš plazmos ekstrahuota modifikavus
operacijas ir sąlygas.
b. DNR iš serumo. Serumas laikytas metus, esant –20 C. ekstrahavimo
procedūros modifikuotos.
0
2) Ekstrakcija
a. QIAGEN rinkinys (QIAGEN, France). Rinkinys, DNR ekstrakcijoms
b. Centrifūgos : Jouan Cr412, Jouan Mr1812 (Saint-Herblain, France),
DNA 110 Speedvac (Savant, Waltham, MA, USA).
c. Spektrofotometras BIO RAD SMART SPEC 3000. Skirtas DNR
koncentracijai matuoti
3) PGR reakcija :
a. Pradmenys (Eurogentec S.A, Seraing, Belgium), (MWG-Biotech AG,
Martinsried, Germany).
b. Fermentas Taq polimerazė (Applied Biosystems, Foster City, CA,
USA).
c. Buferinis tirpalas 10x (Applied Biosystems, Foster City, CA, USA).
d. MgCl2 - (Applied Biosystems, Foster City, CA, USA).
e. Mišinys dNTP`s - (Applied Biosystems, Foster City, CA, USA).
f. Išgrynintas vanduo – Baxter (Baxter, Deerfield, IL, USA) ir aqua
B.Braun (B.Braun Melsungen AG, Deutscland).
g. Reakcijos buvo atliekamos THERMOCYCLERS`iuose (Applied
Biosystems, Foster City, CA, USA).
4) PGR produkto gryninimas
a. MILLIPORE MultiScreen® PCRµ96 (Millipore, Billerica, MA, USA).
5) Gel agarozė:
a. EUROBIO AGAROSE STANDARD (Laboratoires Eurobio,
Courtaboeuf, France).
b. Buferinis tirpalas TBE 10X gel-agarozei (Sigma-Aldrich, France).
38

c. Molekulinis markeris (AB Fermentas, Vilnius, Lietuva).
d. MP4+ INSTANT CAMERA – (Polaroid, S.A. Montigny le
Bretonneux, France) – kamera daryti gelio nuotraukas.
6) Sekvenavimo reakcija
a. Sephadextm g50 superfine (Ge Healthcare Biosciences AB, Uppsala,
Suede). Polimerai sekoms valyti.
b. KIT BIG DYE TERMINATOR V 1.1 (Applied Biosystems, Foster
City, CA, USA). Sekų reakcijos mišinys.
c. pradmenys M13 (forward).
d. Buferinis tirpalas 5x (Applied Biosystems, Foster City, CA, USA).
e. Išgrynintas vanduo – Baxter (Baxter, Deerfield, IL, USA) ir aqua
B.Braun (B.Braun Melsungen AG, Deutscland).
f. Sekų reakcijos vykdomos aparatu 3130 XL Genetic Analyser (Applied
Biosystems, Foster City, CA, USA).
7) Programos sekoms vizualizuoti :
a. AB sequencing analysis (Applied Biosystems, Foster City, CA, USA).
b. AB genemapper (Applied Biosystems, Foster City, CA, USA).
2. Sąlygos PGR – tai labai jautrus užteršimui metodas. Tam, kad reakcijos pavyktų, mes dirbome
visiškoje švaroje : stalai valomi NaOCl tirpalu, visuomet dėvimos pirštinės, specialūs
drabužiai “PGR mišinio” patalpoje. Patalpos, kuriose atliekamos skirtingos reakcijos,
yra atskirtos viena nuo kitos. Siekiant išvengti medžiagos praradimo, mes naudojome
pipečių antgalius ir mini mėgintuvėlius iš polipropileno, nes polipropilenas mažiau
adsorbuoja reagentus.
3. Gauti pavyzdžiai Gavom 3 pacientų pavyzdžius : paciento 42, paciento 44 ir pacento 0007
Tai plazma paciento 42 ir paciento 44 bei serumas paciento 0007. Pastarojo paciento
serumas leido ištirti ilgalaikio užšaldymo efektą biologiniame pavyzdyje ir ištirti
DNR.
39

4. DNR ekstrahavimas Mes atlikom 2 DNR ekstrahavimo metodus. Tai metodai, naudojami laboratorijoje,
todėl yra tam tinkamos sąlygos, bei medžiagos.
Metodai :
• ekstrakcija, panaudojant NaCl (druskinis metodas)
• ekstrakcija, panaudojant QIAGEN rinkinį
4.1. Ekstrahavimo procedūros tobulinimas serumui ir plazmai,
ekstrahuojant natrio chloridu (druskiniu metodu) Mes dirbome su tokiais mažais medžiagos kiekiais kaip 250µL plazmos ar serumo,
kadangi buvo naudojami serotekuose saugomi pavyzdžiai.
Eliuento tūrį, kuris panaudojamas eliuoti DNR, sumažinome iki 100µL, kadangi DNR
kiekis, gautas iš šaltinio, kurio kiekis sumažintas ir jame yra mažiau DNR (plazma,
serumas turi mažiau DNR, nei jos galima išgauti iš kraujo). Tačiau, eliuento tūris
pakankamai didelis, kad galima būtų gerai ištirpinti DNR, siekiant išvengti didelių
nuostolių, o taip pat, kad galima būtų įvykdyti atitinkamą kiekį reakcijų.
4.1.1. Ekstrahavimo protokolas 1. baltųjų kraujo kūnelių lizė
Mes įdėjome 750µL buferinio tirpalo WLB 1X (NaCl 0,4M;
trishidroksimetilaminometano 0,01M ; EDTA 0,002).
2. baltymų suskaidymas
Įdėjom 100µL proteinazės K (20mg/ml), 50µL SDS 10% (natrio duodecilsulfatas).
Maišėme lėtais judesiais, sukdami. Laikėme 370C temperatūroje visą naktį.
3. ląstelių liekanų atskyrimas
Įdėjome 300µL 6M NaCl tirpalo ir maišėme energingai 15s, kol susidarė baltos putos.
Centrifūgavome 15min, esant 4700aps/min. Ant dugno susidarė nuosėdos iš baltymų
ir druskų.
Perkėlėme tirpalą į kitą mėgintuvėlį ir į jį įpylėme 3 tūrius 100% etanolio, kuris
atšaldytas iki -200C.
Lėtai maišėme ir visą tūrį perkėlėme į 3 eppendorf`us po 1500µL. Centrifūgavom
30min, esant 16000aps/min, bei esant +50C temperatūrai. Išėmėme visą tirpalą ir
DNR, susidariusią ant mėgintuvėlio dugno, perplovėme 70% etanoliu : į kiekvieną
40

eppendorf`ą įdėjome po 500µL 70% etanolio ir centrifūgavome 30min, esant
16000aps/min ir +50C.
Džiovinimą atlikome SPEED VAC centrifūgomis, esant kambario temperatūrai, 10
min. DNR eliuavome 100-u mikrolitrų vandens.
4.2. Ekstrahavimo procedūros, ekstrahuojant iš plazmos ir
serumo, panaudojant QIAGEN (QIAAMP DNA BLOOD
MIDI KIT) rinkinį, tobulinimas Plazmos kiekį, naudotą ekstrakcijai, sumažinome iki 1000µL, o serumo – iki 370µL
(vietoj 2000µL kraujo, kurie naudojami standartinės ekstrakcijos iš kraujo, metu).
Eliuento tūrį sumažinome iki 100µL, kad gerai išeliuoti DNR iš QIAGEN kolonos ir,
kad gauto eliuato užtektų tam tikram reakcijų kiekiui.
4.2.1. Ekstrahavimo protokolas Įdėjome 100µL proteazės K (QIAGEN), 1,7mL buferinio tirpalo AL
(NaOH/SDS/guanidin tiocianatas). Gerai išmaišėme (2min minimum).
15 min patalpinom į vonelę, esant 700C.
Po to įdėjome 1mL 100% etanolio ir išmaišėme. 1,65 mL šio mišinio įleidome į
koloną ir centrifūgavome 3min esant 3000aps/min. Pakartojome operaciją su likusiu
kiekiu tirpalo.
Atlikome kolonos plovimą 1mL buferinio tirpalo AW1 (NaCl 1,25M) ir
centrifūgavom 1 min, esant 5000aps/min (operacija kartojama 2 kartus).
Atlikome kolonos plovimą 2mL buferinio tirpalo AW2 (izopropanolis ir natrio
azidas). Centrifūgavom 15 min, esant 5000aps/min (operacija kartojama 2 kartus).
Atlikom DNR eliuavimą 100µL buferinio tirpalo AE (10mM Tris-Cl ; 0,5Mm
EDTA ; pH 9). Užpylus AE buferiniu tirpalu, laikėm 5min ir po to 5 min
centrifūgavom 5min, esant 5000aps/min. (operacija kartojama 2 kartus, panaudojant
pirmą eliuatą).
5. Optinės skvarbos matavimas Siekiant išmatuoti optinę skvarbą, naudojamas 1/50 DNR tirpalas (tam, kad kuo
mažiau prarasti medžiagos). Buvo gauti rezultatai :
Pacientas 42 ekstraktas KIT Qiagen (1ml plazma) c→0
41

Pacientas 44 ekstraktas KIT Qiagen (1ml plazma) c→0
Pacientas 0007 ekstraktas KIT Qiagen (1ml serumas) c→0
Pacientas 0007 ekstraktas NaCl (250 µl serumas) c→0
Pacientas 44 ekstraktas KIT Qiagen (370 µl plazma) c→0
Pacientas 44 ekstraktas NaCl (250 µl plazma) c→0
Iš gautų rezultatų padarėme išvadą, kad jei ir yra DNR mūsų ekstraktuose, tai jos yra
toks nežymus kiekis, kad, atskiedus 50 kartų, naudotas spektrofotometras negali jos
aptikti.
6. Genas TPMT
6.1. “Ilgųjų” pradmenų patvirtinimo procedūra, panaudojant
DNR, ekstrahuotą iš kraujo Kadangi mes planavome dirbti su mažais kiekiais DNR, ekstrahuotos iš plazmos ir
serumo, reikėjo rasti metodą kaip sustiprinti signalą. Mes pasirinkom lizdinės PGR
metodą. Mes sumodeliavome 2 pradmenų poras : viena pora – “ilgieji”, kita pora –
“trumpieji”. Kad sumodeliuoti šiuos pradmenis, mes susiradome geno TPMT sekas
interneto tinklapyje “PUBMED”.
“Ilgieji” pradmenys 5F2 - 5`-CAT-TCC-AAC-TGT-TTC-ATA-C-3` 693 Bp 5R2 - 5`-GTC-TGG-CAC-TGA-TGG-GTT-C-3`
542 Bp 7F2 - 5`-TTG-CAC-ACT-GTC-CTT-TGT-TC-3` 7R2 - 5`-CAA-GCC-TTA-TAG-CCT-TAC-AC-3` 10F2 - 5`-TCC-CAA-GTA-GCT-GGG-ATT-AC-3` 631 Bp 10R2 - 5`-GCA-ATC-TGC-AAG-ACA-CAT-AG-3`
“Trumpieji” pradmenys ”Trumpieji” pradmenys jau naudojami laboratorijoje, DNR ekstrahuotai iš kraujo.
507 Bp 5F1 - 5`-CCT-GCA-TGT-TCT-TTG-AAA-CCC-TAT-GAA-3` 5R1 - 5`-GCT-TGA-GTA-CAG-AGA-GGC-TTT-GAC-CTC-3` 7F1 - 5`-CTC-CAC-ACC-CAG-GTC-CAC-ACA-TT-3` 280 Bp 7R1 - 5`-GTA-TAG-TAT-ACT-AAA-AAA-TTA-AGA-CAG-CTA-AAC-3`
401 Bp 10F1 - 5`-AAT-CCC-TGA-TGT-CAT-TCT-TCA-TAG-TAT-TT-3` 10R1 - 5`-CAC-ATC-ATA-ATC-TCC-TCT-CC-3`
42

Pradmenų paruošimas darbui
Pradmenys gauti liofilizuotos formos. Mes paruošėme 3 eiles skiedimų : 500µM,
100µM ir 10µM.
Pradmenų patvirtinimas
Mes atlikome naujų pradmenų patvirtinimą, panaudodami pacientų 41 ir 42 kraujo
DNR ekstraktus.
PGR reakcija. Mišinio sudėtis:
P. 41
P. 42
vanduo 38,75µl vanduo 38,25µl Buferis 10X 5µl Buferis 10X 5µl MgCl2 3µl MgCl2 3µl dNTP 0.5µl dNTP 0.5µl Pradmuo F2 1µl Pradmuo F2 1µl Pradmuo R2 1µl Pradmuo R2 1µl Taq polymerazė
0,25µl Taq polymerazė
0,25µl
DNR(200ng/µl) 0,5µl DNR(100ng/µl) 1 µl
DNR buvo gausinta, panaudojant programą TPMT BIS 620C :
Denatūravimas 950C – 5 min Denatūravimas - 950C – 1 min
35 ciklai Hibridizacija - 620C – 1 min Elongacija - 720C – 2 min Elongacija - 720C – 7 min
Po reakcijos gauti produktai buvo valomi plokštelėmis PCRµ96.
Buvo atlikta gel-elektrofarezė 2% agarozės gelyje. Elektrofarezė buvo atliekama
naudojant 1X TBE buferinį tirpalą, esant 21W galiai (200v/100mA).
Kad galima būtų vizualizuoti DNR, buvo naudojamas etidiumo bromidas. DNR
panirimas į šulinėlius gelyje buvo pagerintas panaudojant orange G.
Nufotografavus, matomi 3 ilgių fragmentai (pav.21) : egzonas 5 - 693Bp, – egzonas
10 – 631Bp ir egzonas 7 – 542Bp.
43

Pav.21. fragmentų išsidėstymas gelyje
542 Bp
693 Bp
631 Bp
Sekvenavimas
Atlikome sekvenavimą, kad išsiaiskintume ar gauname laukiamą geno seką.
Sekvenavimo reakcijos mišinys:
vanduo – 3,5µl buferinis tirpalas 5X - 2µl pradmuo – 2µl Big dye polimerazė – 1 µl DNR – 1,5µl
Programa:
Denatūravimas 960C – 2 min Denatūravimas - 960C – 30 min
25 ciklai Hybridizacija - 500C – 15 sec Elongacija - 600C – 4 min
Po to reakcijos mišinį išgryninome SEPHADEX`u (jo sudėtyje yra polimerų, kurie
adsorbuoja mažas molekules).
Iliustruoti rezultatui pasirinkome 10-ojo egzono pradžią (pav.22) ir pabaigą (pav.23):
Seka, kurios laukiama: AGGCATGTGCCACCATACCCAGCTCATTTTGTATTTTTAAGTTTCACCATCTTGGCPav.22
44

Seka kurios laukiama: GATTATGATGTGAAAGATTATGCCTATGTGTCTTGCAGATTGCPav.23
Gelio nuotraukoje matome 7 ir 10 egzonų fragmentus, kurie išsidėsto atitinkamai
pagal savo ilgį. Jie yra reikiamo ilgio. 5-ojo egzono fragmento sintezės sąlygas reikia
modifikuoti, nes jis nuotraukoje nėra tokio ryškumo kaip egz. 7 ir 10.
Analizuojant sekas, iliustravome naudodami 10-ojo egzono seką. Seka atitinka
laukiamą nukleotidų seką. Sekos pradžia ir pabaiga nėra gerai “nuskaitoma”, kadangi
sekvenavimo aparatas nevisada “nuskaito” sekos pradžią ir pabaigą, kurioje yra
pradmenų prisijungimo vieta.
6.2. PGR reakcijos optimizavimas egzonui 5 Modifikavome hibridizacijos temperatūrą egzonui 5, norėdami gauti ryškesnį jo
vaizdą gelyje, o tuo pačiu ir daugiau produkto sekvenavimui.Vietoj 620C
hibridizacijos temperatūros, atlikome reakciją, esant 600C. Naudojome paciento 42
kraujo DNR ekstraktą. Gelyje gavom tokį rezultatą (pav.24).
Sumažinus hibridizacijos temperatūrą, gavome
ryškų egzoną 5, mažiau ryškų egzoną 7, o egzono 10
pradmenys degradavę – tai nustatėme atlikdami
keletą reakcijų paeiliui ir panaudodami skirtingas
pradmenų hibridizacijos temperatūras.
Pav.24 fragmentų išsidėstymas gelyje
693 Bp
542 Bp
45

7. Lizdinės PGR strategija : DNR, ekstrahuotos iš plazmos ir
serumo, genotipavimas
7.1. Plazma paciento 42
Naudojome DNR, ekstrahuotą iš paciento 42 plazmos, pasitelkiant QIAGEN rinkinį.
Kad turėtume pakankamai matricos sekvenavimui, naudojome 2 poras pradmenų :
“ilguosius” ir “trumpuosius”. Darėme 2 PGR: iš pradžių su “ilgaisiais” pradmenimis
F2,R2, o po to – su “trumpaisiais” F1,R1.
Pakeitėm DNR ekstrakto kiekį, kuris dedamas į PGR mišinį. Dirbom su ekstraktais,
kuriuose yra labai maži DNR kiekiai, todėl į pirmos PGR reakcinį mišinį dėjom 10µL
ekstrakto, o į antros PGR reakcinį mišinį - 5µL pirmos PGR tirpalo. Nedarėm pirmo
PGR produkto valymo ant plokščių PCRµ96. Darėm valymą po antros PGR.
Naudojom 620C hibridizacijos temperatūrą..
Reakcijos mišinys:
P. 42 1-a PGR 2-a PGR
vanduo 29,25µl vanduo 33,25µl buferis 10X 5µl buferis 10X 5µl MgCl2 3µl MgCl2 3µl dNTP 0.5µl dNTP 0.5µl pradmuo F2 1µl pradmuo F1 1µl pradmuo R2 1µl pradmuo R1 1µl Taq polimerazė 0,25µl Taq polimerazė 0,25µl DNR (plazmos ekstraktas)
10 µl DNR (1-oji PGR) 5 µl
Gel-elektrofarezė (pav.25)
280 Bp
Pav.25 fragmentų išsidėstymas gelyje
401 Bp
46

Sekvenavom 10 egzoną
Panaudojom “trumpuosius” pradmenis. Iliustravom seką, imdami jos pradžią (pav.26)
ir pabaigą (pav.27).
Laukiama seka: TCATAGTATTTTAACATGTTACTCTTTCTTGTTTCAGGTAAAATATGCAATATACGTTGTCTTGAG
Pav.26
Laukiama seka: TAGAAGAAAAAGATGTAACTAAACTTTTAAAGTAGCTCCTTTGGAGAGGAGATTATGATGTG
Pav.27
Sekvenavom 5 egzoną
Abiejų PGR hibridizacijos temperatūrą pasirinkom 600C, kadangi prieš tai buvo gauti
geresni rezultatai esant 600C. Visos kitos sąlygos – kaip ir egz.10, bet į sekvenavimo
mišinį dėjom ne 1,5, bet 2,5µL antros PGR, tam, kad turėti daugiau matricos.
Iliustravimui pasirinkom egzono 5 pradžią (pav.28) ir pabaigą (pav.29)
Laukiama seka: CCTGCATGTTCTTTGAAACCCTATGAACCTGAATTCATATAAATTCCTCTAAATTAAAGAAAATATATGCTTACTCTAATATAACC
Pav.28
47

Laukiama seka: GGGCTTAAAAATATAGGTGATAGCTACCTAAATAGGTATAGACATATGTATATAAAAGCTGAGGTCAAAGCCTCTCTGTACTCAAGC
Pav.29
Gelyje gavom egzono 7 ir 10 laukiamo ilgio fragmentus. Egzono 10 sekos mums
rodo, kad yra fono triukšmas, lyginant su sekomis, darytomis panaudojant kraujo
ekstraktą, nes ekstraktai gauti iš plazmos, o ne iš kraujo ir darytos 2 PGR reakcijos. O
tai sąlygoja fono triukšmo atsiradimą.
Egzono 5 sekoje fono triukšmo daugiau nei egzono 10. Egzono 5 sekos pabaiga
prasčiau “nuskaitoma”.
7.2. Paciento 44 plazmos ekstraktas PGR mišiniui mes naudojome DNR ekstrahuotą iš paciento 44 plazmos, pasitelkiant
ekstrahavimą natrio chloridu (druskinis metodas).
Hibridizacijos temperatūrą pasirinkome 600C. DNR ekstrakto kiekį 1-ąjai PGR -
10µL, o PGR produkto kiekį 2-ąjai PGR - 5µL.
Mes sekvenavome 10 egzoną
Sekvenavimo reakcijos mišiniui naudojome 2,5µL 2-osios PGR (kad turėtume
daugiau matricos).
Rezultatas: sekos pradžia (pav.30) ir pabaiga (pav.31)
Laukiama seka: TCATAGTATTTTAACATGTTACTCTTTCTTGTTTCAGGTAAAATATGCAATATACGTTGTCTTGAGAAGGT
Pav.30
48

Laukiama seka: GAAAAAGATGTAACTAAACTTTTAAAGTAGCTCCTTTGGAGAGGAGATTATGATGTG
Pav.31
10 egzono sekos “perskaitomos”. Laukiama seka atitinka gautą eksperimento metu.
Yra daugiau fono triukšmo, lyginant su sekomis iš kraujo.
7.3. Sekvenavimas PGR produkto, gauto panaudojant DNR
ekstraktą iš 370µL p.44 plazmos Panaudodami QIAGEN rinkinį, ekstrahavom paciento 44 plazmos DNR.
Panaudojome 370µL plazmos, nes daugiau jos neturėjome.
PGR ir sekvenavimo sąlygos tokios pačios kaip ir prieš tai esančiame skirsnyje.
Vizualizuodami sekas, mes paaukštinome pikus, pasinaudodami programos
galimybėmis. Mes radom mutaciją, kuri buvo rasta pas šį pacientą, genotipuojant jo
kraujo DNR ekstraktą.
Palyginom savo gautą rezultatą (pav.32), su gautuoju panaudojant kraują (pav33):
Pav.32
Pav.33
49

Seka, gauta panaudojant 370µL plazmos (tai apie 6 kartus mažesnis tūris, nei kraujo,
kuris naudojamas rutininės operacijos metu), panaudojus 2 poras pradmenų, leidžia
mums nustatyti tyliąją mutaciją gene. Gautas pikas yra mažesnio aukščio nei kraujui.
7.4. Ilgalaikio užšaldymo įtaka : DNR buvo ekstrahuota iš serumo,
kuris buvo užšaldytas metus –200C temperatūroje Mes ekstrahavom paciento 0007 serumo DNR, panaudodami ekstrahavimą natrio
chloridu.
Naudojom lizdinę PGR. Hibridizacijos temperatūrą pasirinkom 600C. DNR ekstrakto
kiekis 1-ąjai PGR - 10µL, 2-ąjai - 20µL pirmosios PGR.
Į sekvenavimo mišinį dėjom 2,5µL antrosios PGR, kad turėtume kuo daugiau
matricos. Gavom tokias sekas 10 egzonui : pradžia (pav.34) ir pabaiga (pav.35)
Laukiama seka: TCATAGTATTTTAACATGTTACTCTTTCTTGTTTCAGGTAAAATATGCAATATACGTTGTCTTGAGAAGGT
Pav.34
Laukiama seka: TAGAAGAAAAAGATGTAACTAAACTTTTAAAGTAGCTCCTTTGGAGAGGAGATTATGATGTGGGTCATAG
Pav.35
50

Sekos mums leidžia spręsti, kad galima daryti genotipavimą net iš serumo, kuris buvo
laikomas užšaldytas –200C temperatūroje metus. Sekos yra “nuskaitomos”, nes
galima “perskaityti” tam tikras sekos dalis.
8. UGT1A1 genas Mes pasirinkom 2 poras pradmenų kaip ir genui TPMT, nes taip pat naudojom
lizdinės PGR metodą. Atlikom tas pačias procedūras kaip ir TPMT genui.
8.1. Pradmenys UGT1A1 genui
A. pradmenys, kai fragmentas gaunamas 253-255bp
SENS2 - 5`-M-AAG-TGA-ACT-CCC-TGC-TAC-CTT-3`
ANTISENS2 - 5`-CCA-CTG-GGA-TCA-ACA-GTA-TCT-3`
B. pradmenys, kai fragmentas gaunamas 98-100bp
SENS1 - 5`- M-GTC-ACG-TGA-CAC-AGT-CAA-AC-3`
ANTISENS1 - 5`-TTT-GCT-CCT-GCC-AGA-GGT-T-3`
8.2. Naujųjų pradmenų patvirtinimas, panaudojant kraujo DNR
ekstraktą Mes naudojome paciento 42 ir paciento 44 kraujo DNR ekstraktą.
PGR mišinys : P. 42
Vanduo 11,9µl buferis 10X 2µl MgCl2 4µl dNTP 0.5µl pradmuo sens bis 0,2µl pradmuo antisens 2 0,2µl Taq polimerazė 0,25µl DNR (100ng/µl) 1µl
51

Programa:
Denatūravimas 940C – 5 min Denatūravimas - 940C – 30sec Hybridisacija - 550C – 40sec 25 cycles Elongacija - 720C – 40sec Elongacija- 720C – 10 min
Mes nedarėme gel-elektrofarezės, nei PGR produkto valymo, nes metodas kitoks nei
prieš tai taikytas genui TPMT : PGR reakcijai atlikti naudojami pradmenys, žymėti
fluoroforu (pradmuo forward yra žymėtas fluoroforu 5`). Amplifikuojama dalis
promotoriaus, kuri turi TATA-BOX. Po to daromas denatūravimas formamido terpėje
ir esant aukštai temperatūrai. Gauti PGR metodu fragmentai yra 98-100bp ilgio ir
vadinami “trumpieji”, o 253-255bp – vadinami “ilgieji”.
Denatūruojantis mišinys:
Formamidas - 15µl
ROX – 0,5µl (molekulinio markerio fragmentų mišinys iki 600Bp).
DNR (PGR produktas) - 1µl paciento 42 kraujo DNR ekstraktui ir 0,5µl paciento 44
kraujo ekstraktui.
Mišinį denatūravome 3min esant 950C.
Kadangi, lėtai šaldant, DNR gali vėl susijungti į dvigrandę struktūrą, tai mišinys
tuojau pat dedamas į ledą 5-ioms minutėms (greitas atšaldymas neleidžia DNR grįžti į
pradinę struktūrą).
Rezultatas gaunamas mišinį perleidus per sekvenatoriaus chromatografines kolonėles.
Molekulinis markeris (ROX) leidžia nustatyti DNR fragmentų ilgį.
Analizuodami paciento 42 kraujo DNR ekstraktą, gavome 2 pikus 253 ir 255. Du
pikai rodo, kad pacientas yra heterozigotas UGT1A1 genui (pav.36).
Analizuodami paciento 44 kraujo DNR ekstraktą taip pat gavome 2 pikus (pav.37). Jie
išeina iš ribų, o tai rodo, kad PGR reakcijos mišinys yra persotintas DNR. 0,5 µL
paciento 44 DNR ekstrakto yra perdaug.
52

Pav.36
Pav.37
Naujieji pradmenys veikia. Gaunami laukiamo dydžio fragmentai. Reakcija yra
jautresnė, nei sekvenavimas, nes pacientui 44 0,5µL DNR kraujo ekstrakto yra
perdaug.
53
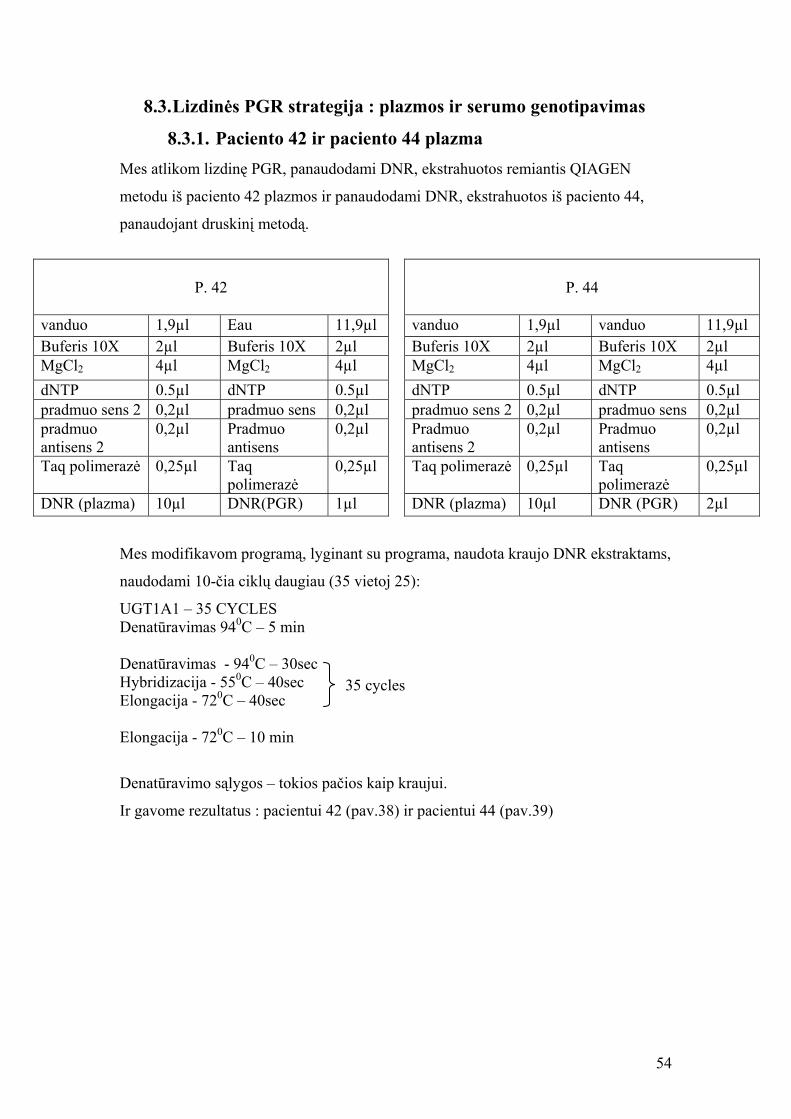
8.3. Lizdinės PGR strategija : plazmos ir serumo genotipavimas
8.3.1. Paciento 42 ir paciento 44 plazma Mes atlikom lizdinę PGR, panaudodami DNR, ekstrahuotos remiantis QIAGEN
metodu iš paciento 42 plazmos ir panaudodami DNR, ekstrahuotos iš paciento 44,
panaudojant druskinį metodą.
P. 44
vanduo 1,9µl vanduo 11,9µl Buferis 10X 2µl Buferis 10X 2µl MgCl2 4µl MgCl2 4µl dNTP 0.5µl dNTP 0.5µl pradmuo sens 2 0,2µl pradmuo sens 0,2µl Pradmuo antisens 2
0,2µl Pradmuo antisens
0,2µl
Taq polimerazė 0,25µl Taq polimerazė
0,25µl
DNR (plazma) 10µl DNR (PGR) 2µl
P. 42
vanduo 1,9µl Eau 11,9µl Buferis 10X 2µl Buferis 10X 2µl MgCl2 4µl MgCl2 4µl dNTP 0.5µl dNTP 0.5µl pradmuo sens 2 0,2µl pradmuo sens 0,2µl pradmuo antisens 2
0,2µl Pradmuo antisens
0,2µl
Taq polimerazė 0,25µl Taq polimerazė
0,25µl
DNR (plazma) 10µl DNR(PGR) 1µl
Mes modifikavom programą, lyginant su programa, naudota kraujo DNR ekstraktams,
naudodami 10-čia ciklų daugiau (35 vietoj 25):
UGT1A1 – 35 CYCLES Denatūravimas 940C – 5 min Denatūravimas - 940C – 30sec
35 cycles Hybridizacija - 550C – 40sec Elongacija - 720C – 40sec Elongacija - 720C – 10 min
Denatūravimo sąlygos – tokios pačios kaip kraujui.
Ir gavome rezultatus : pacientui 42 (pav.38) ir pacientui 44 (pav.39)
54

Pav.38
Pav.39
Rezultatai, dirbant su DNR, ekstrahuota panaudojant QIAGEN rinkinį, yra geresni,
nei ekstrahuojant natrio chloridu : yra daugiau parazitinių pikų ir jie labiau išreikšti,
nei ekstrahuojant QIAGEN`u. Tačiau, taip pat akivaizdu, kad ekstrahuojant natrio
chloridu gaunama daugiau matricos, nei ekstrahuojant QIAGEN`u : apie tai užsimena
ir gamintojas (QIAGEN).
55

8.3.2. Paciento 0007 serumas : pavyzdys buvo užšaldytas esant
– 200C ir laikomas metus Mes neturėjome atlikto kraujo genotipavimo šiam pacientui, todėl neturėjome kaip
patikrinti ar mūsų PGR rezultatai teisingi. Mes padarėme lizdinę PGR paciento 0007
serumo DNR ekstraktui, kuris buvo gautas panaudojant druskinį ekstrahavimą.
Mišinys : 10µL serumo DNR ekstrakto pirmai PGR, 3µL pirmojo PGR į antrosios
PGR reakcinį mišinį. Ir gavome piką (pav.40):
Pav.40
Gavom reikiamo dydžio piką. Fragmentas tinkamo ilgio. Pacientas turėtų būti
homozigotas, tačiau nėra kaip patvirtinti to, nes neturime kraujo. Yra fono triukšmo,
bet jis ne toks didelis kaip paciento 44 plazmos DNR ekstraktui.
B. GENO EKSPRESIJOS MATAVIMAS Tai iRNR, koduojančios aromatazę, ir esančios duotoje terpėje, kiekybinis
nustatymas. Mes pradėjome nuo šios technikos patvirtinimo.
1. Pasirinktas analizės metodas Geno ekspresijos matavimas, panaudojant ROCHE LIGHTCYCLERTM ir į DNR
įsiterpiančią medžiagą SYBRGREENTM.
56

2. Medžiagos ir instrumentai 1. Genetinės medžiagos šaltiniai :
a) MCF-7 ir Hela ląstelių RNR
b) pirmos krūties vėžio stadijos ląstelių cDNR (T1 ir T3)
2. Ekstrahavimas
a) QIAGEN RNEASY MINI KIT (QIAGEN, France) – rinkinys RNR
ekstrahavimui
b) Jouan 14A (Saint-Herblain, France) – centrifūga
c) BIO RAD SMART SPEC 3000. Spektrofotometras, skirtas RNR
koncentracijai matuoti
3. Atvirkštinės transkripcijos reakcija (reagentai)
a) Superscript 3 reverse transcriptase (Invitrogen, France) – rinkinys
atvirkštinės transkriptazės reakcijai :
i. dNTP (10mM)
ii. 5X first strand buffer
iii. 0.1 M DTT
iv. RNAse OUTTM recombinant RNAse inhibitor (40U/µl)
v. Superscript 3 RT – reverse transcriptase (200U/µl) (enzyme)
b) Hexamers (200ng/µl) (Ge Healthcare Biosciences AB, Uppsala, Suede).
c) RNA`se free water (Qiagen, France)
4. geno ekspresijos matavimo reakcija:
a) LIGHTCYCLER FASTSTART DNA MASTERPLUS SYBRGREEN 1
(Roche applied science, Mannheim, Germany)
b) ROCHE LIGHTCYCLER 1 (Roche applied science, Mannheim,
Germany)
c) Išgrynintas vanduo – Baxter (Baxter, Deerfield, IL, USA) ir aqua B.Braun
(B.Braun Melsungen AG, Deutscland).
5. Lightcycler software version 3 (Roche applied science, Mannheim, Germany)
– programa rezultatams analizuoti
3. Sąlygos Mes dirbom su RNR, kuri nėra labai stabili molekulė. Viena šio nestabilumo
priežąsčių – RNR`azė, kurios yra aplinkoje. Štai kodėl kiekvieną kartą prieš
57

imdamiesi darbo su RNR, mes kruopščiai nuvalydavome stalą su NaOCl,
apiplaudavome pipetes su vandeniu, turinčiu DEPC. Naudojom atskiras pipetes,
skirtas dirbti tik su RNR.
4. RNR ekstrahavimas QIAGEN RNEASY MINI rinkiniu iRNR sudaro 2-3% visų ląstelės RNR.
Mes atlikom RNR ekstrahavimą iš ląstelių MCF-7 ir Hela. Naudojom 7milijonus
ląstelių MCF-7 ir 4 milijonus Hela.
4.1. Ekstrahavimo protokolas Ant ląstelių užpylėme 600µL buferio RLT-BME (guanidino tiocianatas, 2-merkapto
etanolis). Homogenizavom mišinį. Lizuotą ląstelių mišinį perkėlėme į naują
mėgintuvėlį ir įdėjome 350µL 70% etanolio. Lizatą perkėlėme į QIAGEN koloną ir
centrifūgavom 15s esant 10000aps/min.
Įdėjom 350µL buferio RW1 (guanidino tiocianatas) ir vėl atlikom centrifūgavimą,
esant 10000aps/min 15s. Po to įdėjom 10µL DNA`zės, praskiestos 70µL-ų RDD
buferio ir palikom pusvalandžiui kambario temperatūroje.
Įdėjome 350µL buferio RW1 ir atlikom 15min centrifūgavimą esant 10000aps/min.
Vykdėm plovimą 500µL buferiu RPE (NaCl). Centrifūgavom 2 kartus po 2min esant
10000aps/min.
Eliuavom RNA`se free vandeniu (su DEPC) : ląstelėms MCF-7 naudojom 2 kartus po
30 µL, o ląstelėms Hela – 2 kartus po 50µL.
Eliuatą užšaldėm –800 C temperatūroje.
5. Optinės skvarbos matavimo rezultatai MCF-7 : 700µg/mL
Hela : 226µg/mL
6. kDNR gel-elektrofarezė Buvo naudojamas 2% agarozės gelis. RNR migravo
daugiau nei 2h (11W/100V/110mA).
Rezultatas:
RNR nedegradavusios, nes matosi 2 dydžių fragmentai,
kurie atitinka 18s ir 28s RNR (pav.41).
RNR 28S
RNR 18S
Pav.41
58

7. Atvirkštinės transkripcijos reakcija Mes atlikom RNR transformavimą į kDNR (cDNR), kad galėtume dirbti su stabilia
medžiaga (kDNR).
MCF-7 ląstelėms naudojome:
5µL RNR ekstrakto (3,5µg grynos RNR)
10µL RNR ekstrakto (t.y. 7µg grynos RNR)
Hela ląstelėms:
7µL RNR ekstrakto (1,6µg grynos RNR).
Padarėme mišinį:
1µL hegzamerų
1µL dNTP (10mM)
RNA`se free vandens iki 13µL
RNR ekstrakto kiekį
Inkubavom 5min esant 650C ir patalpinom 1min į ledą.
Įdėjom kitų komponentų:
4µL first strand buffer
1µL 0,1M DTT (ditiotreitolio)
1µL RNA`se OUT rekombinantinio RNA`zės inhibitoriaus
1µL SUPERSCRIPT 3 RT (200U/µL)
Naudojom programą : 250 C – 5min, 500C – 60 min, 700C – 15 min.
gautą kDNR užšaldėm esant -200C.
8. Kiekybinis metodas
Pasirinkti pradmenys Seka iš “PUBMED”
294 aagtcc 301 atgggaattg gacccctcat ctcccacggc agattcctgt ggatggggat cggcagtgcc 361 tgcaactact acaaccgggt atatggagaa ttcatgcgag tctggatctc tggagaggaa 421 acactcatta tcagcaagtc ctcaagtatg ttccacataa tgaagcacaa tcattacagc 481 tctcgattcg gcagcaaact tgggctgcag tgcatcggta tgcatgagaa aggcatcata 541 tttaacaaca atccagagct ctggaaaaca actcgaccct tctttatgaa agctctgtca 601 ggccccggcc ttgttcgtat ggtcacagtc tgtgctgaat ccctcaaaac acatctggac 661 aggttggagg aggtgaccaa tgaatcgggc tatgtggacg tgttgaccct tctgcgtcgt 721 tacaaaaagt atgagaag
59

294-448 – exon 3 449-624 – exon 4 625-738 – exon 5
Pasirinktas pradmuo forward (321-338) geltonos spalvos. Pradmuo reverse (676-696)
raudonos spalvos. Kadangi tai kDNR, tai gausinimas vyksta tik 3 egzonų. Gaunamas
376 bp ilgio fragmentas. Tikslas darbo su kDNR – išvengiama kontaminacijos
genetine medžiaga.
8.1. Ląstelių, galinčių tapti pavyzdžiu, paieška Reikėjo rasti ląsteles-RNR šaltinį, kurių aromatazės geno ekspresija yra pakankama,
kad patvirtinti techniką.
8.1.1. MCF-7 ląstelės Tai vėžinės krūtų epitelio ląstelės, kurios turi citoplazminius receptorius estrogenams
(ATCC:HTB-22).
8.2. PGR sąlygų patvirtinimas Reakcijos mišinio paruošimas.
9 µL vandens PCR GRADE po 1 µL pradmenų F ir R
4 µL buferio (sudarytas iš specifinės polimerazės, į DNR įsiterpiančios
medžiagos SYBRGREEN 1, MgCl2).
Šis mišinys dedamas į kapiliarus ir po to dedama kDNR.
Mes dėjom 5µL 1/2cDNR tirpalo (2,5µL vandens ir 2,5µL cDNR) į kiekvieną
kapiliarą.
Mes atlikom realaus laiko PGR, esant 55, 60 ir 650 C hibridizacijos temperatūroms.
Programa :
taq aktyvavimas : 950C – 6 min 45 Cycles : 950C – 15 sec pasirinkta hibridizacijos t0 (550C, 600C, 650C) – 7 sec 720C – 14 sec
lydymosi kreivė : 950C – 0 sec 650C – 30 sec 980C – 0 sec
šaldymas : 400C – 60 sec
60

Mes naudojom 2 pradmenų tipus : pradmenis aromatazei ir kalibracinio geno
pradmenis (HPRT). Atlikom reakciją su kalibraciniais pradmenimis, norėdami
sužinoti ar mūsų RT reakcija pavyko. Kalibracinis genas – tai genas, kurio yra visose
ląstelėse ir jis naudojamas ekspresijai monitoruoti.
Esant 55 ir 600C temperatūrai, gavom parazitinius pikus. Padarę gel-elektrofarezę,
naudodami 5µL kapiliaro turinio, gavom (pav.42):
Aromatase 600C Aromatase 650C
HPRT 550C
Aromatase 600C
376bp
Taigi, gelyje matom laukiamo dydžio fragmentą (376bp). Genui HPRT taip pat
matome tam tikrą fragmentą. Tai parodo, kad yra kDNR.
Iliustravimui pasirinkome lydymosi kreivę, kai hibridizacijos temperatūra yra 650C
(pav.43).
Pav.42
Skiedimas : M 1/40 ¼ 0 1/40 ¼ 0 ½ 0 ½ M
1/2
vanduo
Pav.43
61

Darbui su aromataze tinkamiausia temperatūra pradmenims yra 650C, nes gaunamas
specifinis gausinimas, o gelyje matomas vienas reikiamo dydžio fragmentas.
Pikas su vandeniu mums parodo pradmenų dimerų lydymosi temperatūrą.
Mes atlikom reakcijas su kalibraciniais genais TBP89 (pav.44) ir HPRT (pav.45).
Naudojom 2 kapiliarus 1 genui (vanduo “blanck” ir ½ kDNR tirpalas) :
TBP89 – 650C hibridizacija
HPRT - 550C hibridizacija
Pav.44
blanck
1/2
Pav.45
blanck1/2
Duotų genų gausinimas mums rodo, kad RT reakcija pavyko, nes esant ½ RT
skiedimui, gaunamas specifinis gausinimas.
8.3. Hela ląstelės Tai Henrietta Lacks gimdos kaklelio vėžinės ląstelės. Ląstelės auga daug greičiau nei
kitos vėžinės ląstelės. Ląstelės turi estrogeno receptorius ir galimai aromatazės geno
ekspresija jose padidėjus.
Tyrimas
Mes panaudojom tas pačias sąlygas kaip ir ląstelėms MCF-7. Naudojom aromatazės,
TBP89, HPRT ir B2 makroglobulino pradmenis. Naudojom 3 skiedimus: ½, ¼, 1/40 ir
vandenį.
62

Rezultatai : aromatazė (pav.46)
Rezultatai : TBP89 (pav.47)
Rezultatai : HPRT (pav.48)
Rezultatai : B2 (pav.49)
blanck 1/2
1/41/40
blanck
1/2
1/41/40
blanck
1/21/4
1/40
1/41/2Pav.46
1/40blanck
Pav.47
Pav.48
Pav.49
63

Aromatazės genui, esant praskiedimui ½ ir ¼ - specifinis gausinimas. 1/40 –
parazitinis, tikriausiai todėl, kad yra per mažai matricos. Kalibraciniai genai rodo, kad
RT pavykus. Lyginant kalibracinių genų ir aromatazės duomenis, galima padaryti
išvadą, kad Hela ląstelėse aromatazės ekspresija nėra didelė, kadangi, panaudojant
kalibracinius genus, net 1/40 skiedimas duoda teigiamą, panašų į didesnių
koncentracijų rezultatą.
8.4. Palyginamoji studija su MCF-7 Mes atlikom realaus laiko PGR su aromatazės, TBP, HPRT ir B2 pradmenimis. Tai
atlikom, kad išmatuoti aromatazės ekspresiją dviejuose ląstelių tipuose ir, kad
nustatyti transformuotos kDNR kiekį.
MCF-7 Hela
genai genai
aromatazė TBP HPRT aromatazė TBP HPRT B2 Blanck 9,296*10-12 5,938*10-8 8,549*10-7 9,296*10-12 - 1,775*10-6 2,424*10-5
½ 1,047*10-5 2,796*104 1,637*10-1 2,35*10-6 5,722*104 3,333*10-1 2,047 ¼ 7,718*10-7 1,317*104 1,167*10-1 1,188 1/10 1,523*10-6 4,609*102 2,441*10-2 1/40 8,786*10-7 1,358*102 1,798*10-2 1,197*10-6 32,03 1,154*10-2 9,77*10-2
Išvada: aromatazės geno raiška Hela ląstelėse didesnė, nei MCF-7, kadangi MCF-7
ląstelėms buvo panaudota didesnis kiekis grynos RNR. Tačiau tolimesniam darbui su
aromatazės genu šios dvi ląstelės nėra tinkamos.
64

8.5. Aromatazės geno raiška pirmos stadijos krūties vėžio ląstelėse Mes panaudojom kDNR iš krūties vėžio ląstelių, kuriose, mūsų spėjimu, aromatazės
geno raiška yra didesnė nei MCF-7 ar Hela.
Naudojom tas pačias realaus laiko PGR sąlygas.
Pasirinkom tokius T1 (pav.50) ir T3 (pav.51) ląstelių kDNR praskiedimus : ½, 1/10,
1/20
T3 pavyzdyje aromatazės geno raiška yra ryški. Tai mums rodo 1/10 ir 1/20
praskiedimai, kurių piko aukštis skiriasi. T3 pavyzdys duoda specifinį piką kai
koncentracija ½.
T1 ląstelių aromatazės geno raiška nėra ryški, nes praskiedimai nesąlygoja piko
aukščio pakitimų.
Taigi, T3 ląstelės labiau tinkamos aromatazės ekspresijai matuoti nei MCF-7 ar Hela.
blanck
1/2Pav.50 1/10
1/20
blanck
1/2Pav.51 1/10
1/20
65

Išvada: 1. Druskinis ir Qiagen metodai pritaikyti DNR ekstrahavimui iš plazmos bei
serumo, analizuojant rezultatus gautus darant gel-agarozės nuotraukas ir
sekvenavimą. Nustatyta, kad matuojant optinę skvarbą, negalima nustatyti
DNR, gautos ekstrahuojant iš plazmos bei serumo.
2. Sumodeliuoti pradmenys lizdinei PGR ir tyrimų rezultatais patvirtintas jų
tinkamumas kai naudojami kraujo DNR ekstraktai.
3. Lyginant eksperimento metu gautas sekas su patvirtintomis sekomis,
pritaikytos tyrimo sąlygos bei patvirtintas lizdinės PGR tinkamumas tirti DNR
plazmos bei serumo ekstraktuose.
4. Palyginus druskinį ir Qiagen DNR ekstrahavimo metodus nustatyta, kad
Qiagen metodu ekstrahuojant DNR, ji išgaunama geresnės kokybės, nei
druskiniu metodu, tačiau jos kiekis mažesnis.
5. Didelis Qiagen metodo efektyvumas bei bendras lizdinės PGR metodikos
jautrumas patvirtintas sekvenuojant DNR, gautą iš mažo kiekio plazmos, ją
ekstrahavus Qiagen metodu. Nustatyta tylioji mutacija 7 egzone.
6. Ištirtas ilgalaikio biologinės medžiagos užšaldymo poveikis: sekvenuota DNR
iš serumo, laikyto metus esant –200C. Tai parodo, kad galima atlikti
retrospektyvius tyrimus, naudojant ilgai laikytą genetinės medžiagos šaltinį,
pavyzdžiui, tiriant vaisto metabolizmo sutrikimus genetiniame lygmenyje.
7. Nustatytas ląstelių tipas, kuriose aromatazės geno raiška yra pakankama ir
todėl tokio tipo ląstelės gali būti naudojamos kaip standartas toliau tiriant
aromatazės geną.
66

LITERATŪRA Allorge D., Loriot M. A. La pharmacogénétique ou la promesse d’une médecine personnalisée : variations du métabolisme et du transport des médicaments. Annales de Biologie Clinique 2004. Volume 62, Number 5 : 499-511 Andolfatto S., Namour F., Garnier A.L., Chabot F., Gueant J.L. and Aimone-Gastin I.. Genomic DNA extraction from small amounts of serum to be used for 1-antitrypsin genotype analysis. Eur Respir J 2003. 21:215-219 Anker, P., H. Mulcahy, X.Q. Chen & M. Stroun. Detection of circulating tumour DNA in the blood (plasma/serum) of cancer patients. Cancer Metastasis Rev. 1999. 18: 65–73 Armstrong Deborah, O’reilly Seamus. Clinical guidelines for managing topotecan-related hematologic toxicity. The Oncologist 1998. 3: 4-10 Arora Amit, and Jane Potter F. Aromatase Inhibitors: Current Indications and Future Prospects for Treatment of Postmenopausal Breast Cancer. The American Geriatrics Society. 2004. 52:611–616 Bienvenu T., Meunier C., Bousquet S., Chiron S., Richard L., Gautheret-Dejean A., Rouselle J.F., Feldmann D.. Les techniques d’extraction de l’ADN à partir d’un échantillon sanguin. Annales de Biologie Clinique. 1999. Vol. 57, N.1 : 77-84 Borde Isabelle. Polymerase chain reaction, ou réaction de polymérisation en chaîne. UFR. 2006 Bouquier M. Place de la pharmacogenomique dans les strategies de recherche et de developpement des medicaments : enjeux scientifiques, industriels et ethiques. These pour l'obtention du Diplôme d'Etat de docteur en pharmacie Universite Paris V- Rene Descartes faculte des sciences pharmaceutiques et biologiques 2003 Brown T.A.. Genomes 3. 2006 : 1-750 BURNAT Pascal, CEPPA Franck, MULLOT Héléne, BOUHSAIN Sanae, DELACOUR Hervé, OLIVER Manuela. Pharmacogenetique et pharmacogenomique des medicaments, deux nouveaux aspects de la prise en charge therapeutique. Pharm Hosp 2003. 38. 154 : 161-167 Canadian Pharmacists Association (CPA). Irinotecan. CCO Formulary. 2006/2007 Ceré Nicolas. Étude de la primo-infection à Pneumocystis carinii chez le lapin (Oryctolagus cuniculus): développement parasitaire et transmission. THESE présentée devant L'UNIVERSITE FRANCOIS RABELAIS DE TOURS U.F.R. Sciences et Techniques Pour l’obtention du Diplôme de Doctorat Sciences de la Vie Option : Parasitologie. 24 Février 1998 Chaisemartin L., Loriot M.A.. Pharmacogenetique des medicaments anticancereux. Pathologie biologie. 2005. 53: 116-124
67

Coombes R C, Harper-Wynne C and Dowsett M. Aromatase inhibitors and their use in the sequential setting. Endocrine-Related Cancer 1999. 6 : 259-263 Dahlgren L. Studies on the presence and influence of human papillomavirus (HPV) in head and neck tumors. Department of oncology-pathology karolynska institutet, Stockholm, Sweden. Reproprint AB. 2005 : 1-68 Delarue Michel et Furelaud Gilles. Le sequençage d'un ADN. UFR. 2005 Dowsett M, Goss PE, Powles TJ, Hutchinson G, Brodie AM, Jeffcoate SL, Coombes RC. Use of the aromatase inhibitor 4-hydroxyandrostenedione in postmenopausal breast cancer: optimization of therapeutic dose and route. Cancer research 1987. 47 : 1957–1961 Duguay Yannick, McGrath Monica, Le´pine Johanie, Gagne Jean-Francois, Hankinson Susan E., Colditz Graham A., Hunter David J., Plante Marie, Tetu Bernard, Belanger Alain, Guillemette Chantal, and De Vivo Immaculata. The Functional UGT1A1 Promoter Polymorphism Decreases Endometrial Cancer Risk. Cancer Research 2004. 64 : 1202–1207 Durand Geraldine, Frezet Sebastien. Biologie moleculaire en routine au laboratoire de bacteriologie. Des bacteriologie semestre hiver 2002/2003. Collège de Bactériologie-Virologie @ Centre Hospitalo-Universitaire de Lyon et Université LYON 1 FENTIMAN I.S. Aromatase inhibitors and breast cancer: time for a change? Int J Clin Pract. 2004. 58. 12:1152–1158 Francina A. Les Techniques De Base De La Biologie Moléculaire. Purification Des Acides Nucléiques. Fédération de Biochimie. 2002 Guatelli J. C., Gingeras T. R., and Richman D. D. Nucleic Acid Amplification In Vitro: Detection of Sequences with Low Copy Numbers and Application to Diagnosis of Human Immunodeficiency Virus Type 1 Infection. Clinical microbiology reviews. 1989. 2; 2 : 217-226 Guillaume G., Szymanowicz G., Randoux A., Borel J.P. A fractionation of insoluble rat skin collagen. Biomedicine. 1978. 29(1):5-6. Gustincich S., Manfioletti G., Del Sal G., Schneider C., Carninci P. A fast method for high-quality genomic DNA extraction from whole human blood. BioTechniques, 1991. vol. 11, no3 : 298-302 Haiman Christopher A., Stram Daniel O., Pike Malcolm C., Kolonel Laurence N, Burtt Noel P., Altshuler David, Hirschhorn Joel and Henderson Brian E. A comprehensive haplotype analysis of CYP19 and breast cancer risk: the Multiethnic Cohort. Human Molecular Genetics, 2003. Vol. 12, No. 20 : 2679–2692
68

Hamilton A. & Piccart M. The third-generation non-steroidal aromatase inhibitors: A review of their clinical benefits in the second-line hormonal treatment of advanced breast cancer. Annals of Oncology1999. 10: 377-384 Henegariu O., Heerema N. A., Dlouhy S. R., Vance G. H., Vogt P. H. Multiplex PCR : critical parameters and step-by-step protocol. Biotechniques 1997. vol. 23, no3 : 504-511 Hirose Kaoru, Matsuo Keitaro, Toyama Tatsuya, Iwata Hiroji, Hamajima Nobuyuki, and Tajima Kazuo. The CYP19 Gene Codon 39 Trp/Arg Polymorphism Increases Breast Cancer Risk in Subsets of Premenopausal Japanese. Cancer Epidemiology, Biomarkers & Prevention, 2004. vol. 13, no8 : 1407-1411 Huret JL . ADN: Structure Moléculaire. Atlas Genet Cytogenet Oncol Haematol. 2006 Hurley Carolyn Katovich, Vina Marcelo Fernandez, Gao Xiaojiang, Middleton Derek, Ng Jennifer, Noreen Harriet, Ee Chee Ren, Schmeckpeper Barbara, Smith Anajane, Ting Tang, Tokunaga Katsushi. HLA Class I and II DNA-Based Typing Sequence Specific Oligonucleotide Probe Typing Technical Manual / Reference Protocols. Version 1.1. Thirteenth International Histocompatibility Workshop. November 4, 1998 Institut National de Recherche Pedagogique (INRP). Techniques de biologie moleculaire les electrophoreses. 2001 IPSEN (Institut de produits de synthèse et d'extraction naturelle). Progrès génétique & HTA : lettre d'information médicale. [dir. publ. Norbert Claveille]. 2000. N° 9 Iyer L, Das S, Janisch L, Wen M, Ramírez J, Karrison T, Fleming G F, Vokes E E, Schilsky R L and Ratain M J. UGT1A1*28 polymorphism as a determinant of irinotecan disposition and toxicity. The pharmacogenomics journal. 2002. Volume 2, Number 1 : 43-47 Jahr Sabine, Hentze Hannes, Englisch Sabine Hardt, Dieter, Fackelmayer Frank O., Hesch Rolf-Dieter, and Knippers Rolf. DNA Fragments in the Blood Plasma of Cancer Patients: Quantitations and Evidence for Their Origin from Apoptotic and Necrotic Cells, Cancer Research 2001. 61 : 1659–1665 Kang T. J., Yang M. S. Rapid and reliable extraction of genomic DNA from various wild-type and transgenic plants. BioMed Central Biotechnology 2004. 4 : 20 Kao YC, Cam LL, Laughton CA, Zhou D, Chen S. Binding characteristics of seven inhibitors of human aromatase: a site-directed mutagenesis study. Cancer research 1996. 56 : 3451–3460 Kao Yeh-Chih, Korzekwa Kenneth R., Laughton Charles A. and Chen Shiuan. Evaluation of the mechanism of aromatase cytochrome P450. Eur. J. Biochem. 2001. 268, issue 2, 243-251
69

Kaplan J.C., M.Delpech. Biologie Moleculaire et Medecine. 1990. 597 Kubista M., Andrade J. M., Bengtsson M., Forootan A., Jonak J., Lind K., Sindelka R., Sjoback R., Sjogreen B., Strombom L., Stahlberg A., Zoric N. The real-time polymerase chain reaction. Molecular Aspects of Medicine. 2006. 27 : 95–125 Lahiri DK., Schnabel B. DNA isolation by a rapid method from human blood samples: effects of MgCl2, EDTA, storage time, and temperature on DNA yield and quality. Biochem Genet. 1993. 31(7-8) : 321-8 Lodish H., Berk A., Matsudaira P., Kaiser C. A., Krieger M., Scott M. P., Zipursky L., Darnell J.. Molecular Cell Biology. Fifth Edition. 2004 Lonning PE. Clinico-pharmacological aspects of different hormone treatments. Eur J Cancer 2000. 36 : S81–S82 Ma C.X., Adjei Araba A., Salavaggione O.E., Coronel J., Pelleymounter L., Wang L., Eckloff B.W., Schaid D., Wieben E.D., Adjei Alex A., Weinshilboum R.M. Human aromatase:gene resequencing and functional genomics. Cancer research 2005. 65 : 11071-11082 Mahendroo Mala S., Mendelson Carole R., and Simpson Evan R. Tissue-specific and Hormonally Controlled Alternative Promoters Regulate Aromatase Cytochrome P450 Gene Expression in Human Adipose Tissue. The journal of biological chemistry 1993. Vol. 268, No. 26 : 19463-19470 MANEN J. F., BOUBY L., DALNOKI O., MARINVAL P., TURGAY M., SCHLUMBAUM A. Microsatellites from archaeological Vitis vinifera seeds allow a tentative assignment of the geographical origin of ancient cultivars. Journal of archaeological science. 2003. vol 30, no6 : 721-729 McLeod H. L., Parodi L., Sargent D. J., Marsh S., Green E., Abreu P., Cisar L. A. and Goldberg R. M.. UGT1A1*28, toxicity and outcome in advanced colorectal cancer: Results from Trial N9741. Journal of Clinical Oncology 2006. Vol 24, No 18S (June 20 Supplement) 3520 Meyer A. Pharmacogenetics and adverse drug reactions. The Lancet. 2000. 356(9242):1667-71 Miyoshi Y, Iwao K, Ikeda N, Egawa C, Noguchi S. Breast cancer risk associated with polymorphism in CYP19 in Japanese women. Int. J. Cancer 2000. 89, 4 : 325-328 MONAGHAM G., RYAN M., SEDDON R., HUME R., BURCHELL B. Genetic variation in bilirubin UDP-glucuronosyltransferase gene promoter and Gilbert's syndrome. The Lancet. 1996. 347 : 578-581
Mullis K. B, Faloona F. A, Scharf S, Saiki R. K, Horn G, Erlich H. A., Specific enzymatic amplification of DNA in vitro : the polymerase chain reaction. Cold Spring Harbor Symposium in Quantitative Biology, 1986. 51 : 263-273.
70

Nebert D.W., Bingham E. Pharmacogenomics : out of the lab and into the community. Trends in Biotechnology 2001. Vol.19, N°12 : 519-523 O’Dwyer Peter J., Catalano Robert B. Uridine Diphosphate Glucuronosyltransferase (UGT) 1A1 and Irinotecan: Practical Pharmacogenomics Arrives in Cancer Therapy. Journal Of Clinical Oncology 2006. Vol 24, N28 : 4534-4538 Patrick G.L. AN introduction to Medicinal chemistry, third edition, 2005 : 489-522 Quinque D., Kittler R., Kayser M., Stoneking M., NasidzeI. Evaluation of saliva as a source of human DNA for population and association studies. Analytical Biochemistry 2006 Samer Caroline F., Piguet Valérie, Dayer Pierre, Desmeules Jules A. Polymorphisme génétique et interactions médicamenteuses : leur importance dans le traitement de la douleur. Canadian journal of anaesthesia. 2005. vol. 52, no8, pp. 806-821 Seiter Karen. Toxicity of the topoisomerase I inhibitors. Expert Opinion on Drug Safety. 2005. Vol. 4, No. 1 : 45-53 Shozu Makio, Zhao Ying, Bulun Serdar E., and Simpson Evan R. Multiple splicing events involved in regulation of Human aromatase expression by a novel promoter, i.6. Endocrinology 1998. vol. 139, no. 4 : 1610-1617 Siatka Christian La réaction de polymérisation en chaîne (PCR) principe et applications. Congres pluraliste des sciences. 2006 SINNIGER S. Electrophorèse d'ADN sur gel d'agarose. Stage de prise de fonction des nouveaux aides techniques de laboratoire. 2001 Somma M. Analyse d'echantillons alimentaires pour la presence d'organismes genetiquement modifies. Module 4. Extraction et purification de l'ADN. World Health Organisation, 2004 Thorn C. F., Carrillo M.W., Ramirez J., Marsh S.,. Schuetz E.G, Dolan M.E., Innocenti F., Relling M.V., Mcleod H.L.And Ratain M.J. Irinotecan Pathway. The Pharmacogenetics And Pharmacogenomics Knowledge Base, 2005 Tian Q, Zhang J, Chan SY, Tan TM, Duan W, Huang M, Zhu YZ, Chan E, Yu Q, Nie YQ, Ho PC, Li Q, Ng KY, Yang HY, Wei H, Bian JS, Zhou SF. Topotecan is a substrate for multidrug resistance associated protein 4. Curr Drug Metab, 2006. 7. 1 : 105-118 Tse C., Capeau J., Quantification des acides nucléiques par PCR quantitative en temps réel, Ann Biol Clin 2003. 61 n° 3 : 279–293 Vallejo L. F. and Rinas U.. Strategies for the recovery of active proteins through refolding of bacterial inclusion body proteins. Microbial Cell Factories 2004. 3:11
71

Vu N. T., Chaturvedi A. K., Canfield D. V.. Genotyping for DQA1 and PM loci in urine using PCR-based amplification: Effects of sample volume, storage temperature, preservatives, and aging on DNA extraction and typing. Forensic Science International. 1999. 102 : 23–34 Wahbi Kamal. Polymerase chain reaction (PCR) ou amplification elective in vitro. La gazette du laboratoire. Mars 1996. n°6 Weinshilboum Richard. Thiopurine Pharmacogenetics: Clinical And Molecular Studies Of Thiopurine Methyltransferase. The American Society for Pharmacology and Experimental Therapeutics. Drug Metabolism And Disposition 2001. Vol. 29, No. 4, Part 2 : 601–605 Welti Marie-Anne, Loriot Marie-Anne, Beaune Philippe. Prediction des effets indesirables des medicaments. ADSP. 2001, n° 34 Wilkins Martin R, Roses Allen D, Clifford C Piers. Pharmacogenetics and the treatment of cardiovascular disease. Heart 2000. 84 : 353–354 Yang XX, Hu ZP, Xu AL, Duan W, Zhu YZ, Huang M, Sheu FS, Zhang Q, Bian JS, Chan E, Li X, Wang JC, Zhou SF. A mechanistic study on reduced toxicity of irinotecan by coadministered thalidomide, a tumor necrosis factor-alpha inhibitor. J Pharmacol Exp Ther. 2006. 319. 1 : 82-104 Zhou Shufeng. Clinical Pharmacogenomics of Thiopurine S-methyltransferase. Current Clinical Pharmacology. 2006, 1 : 119-128 AKRIM M. Amplification et detection des sequences specifiques.http://www.sante.gov.ma http://en.wikipedia.org http://fr.wikipedia.org PUBMED - NC_000006. http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?DB=pubmed PUBMED - NC_000015 http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?DB=pubmed
72