effects of trace metals on mouse b16 melanoma cells in culture
TRANSCRIPT

@Copyright 1993 by The Humana Press, Inc. All rights of any nature, whatsoever, reserved. 0163-4984/93/36014)191 $02.20
Effects of Trace Metals on Mouse B16 Melanoma Cells
in Culture
ELIANE BAHBOUTH, ~ BRIGITTE SIWEK, j MARIE-CLAIRE D E
PAUW-GILLET, 2 E N R I C O SABBIONI, ~'* AND R O G E R B A S S L E E R 2
'CEC, JRC lspra Site, Environment Institute, 1-21020 lspra (Varese), Italy; and 2Department of Histology and Cytology, University of LiOge,
Rue de Pitteurs 20, B-4020 LiOge, Belgium
Received March 16, 1991; Accepted April 14, 1992
ABSTRACT
The effects of fourteen metal ions (As 3 +, As 5 § Cd 2 +, C o 2 + , Cr 3 +, Cr 6 +, Hg 2 +, Li +, Mg 2 +, Mn 2 +, Ni 2 +, Se 4 +, V 5 +, VO 2 +) on the proli- feration and differentiation in mouse B16 melanoma cells cultivated in vitro were analyzed. Cell number assays, melanin, and protein mea- surements, a 3(4,5-dimethylthiazol-2-yl)-2,5 diphenyltetrazolium bro- mide reduction test (MTT survival test), and a clonal growth assay were performed. At 10-4M, metal ions such as As 3+, As 5+, Cd 2+,
H -~ + Co 2 + Cr 6+, Se 4+, V 5+, VO 2+, and, to a minor extent, Li +, g , and significantly reduced the number of the B16 melanoma cells. For the same molar concentration, the order of the levels of cell toxicity of the metal compounds to B16 cells as measured by the MTT test -was as fo l lows:Hg 2+ Cr 6+ Cd 2+ As 3+ As 5+ V 5+ VO2+ Se 4+ Ni 2+ = C o 2+ > = > , > , > = = Li + . An increased synthesis of melanin in B16 cells was noted after incubation with Co 2+, Ni 2+, Cd 2+, and Li § whereas Se 4+ had, on the contrary, an inhibiting effect on melanogenesis,
Index Entries: Trace metals, in vitro effects of; B16 melanoma ceils; cell differentiation; cytotoxicity; protein synthesis; melanin.
*Author to whom all correspondence and reprint requests should be addressed.
Biological Trace Element Research ] 91 VoL 36, 1993

192 Bahbouth et aL
INTRODUCTION
It is known that trace metals such as Cu, Fe, Se, and Zn are found in all living organisms and are essential for life (1). They participate in numerous biological processes, exhibiting a wide range of functions, e.g., as components of enzymes and redox systems, or structural func- tions. Other trace metals, e.g., As, Co, Cr, Mn, Mo, Sn, and V, are also suspected to play an essential biological role in mammals (2), whereas for others no biological function is so far known.
However , h u m a n activities such as fossil fuel combustion for electri- cal energy production, incineration of urban wastes, the use of agri- cultural chemicals, inorganic fertilizers, and solid wastes from sewage treatment, as well as mining, smelting, and metallurgy activities lead to an excessive mobilization of trace metals with consequent overexposure of the general populat ion and occupational workers and the induction of toxic effects (3,4). In particular, increased exposure to metals such as As, Be, Cd, Cr, and Ni (5,6) has been reported to cause either carcinogenic effects in humans and/or in laboratory animals and genotoxic or immu- notoxic effects in vitro and in vivo (6,7). Furthermore, metal ions such as As 3+, Cd 2+, Hg 2+, Se 4+, or V 5+ display cytotoxic properties on cultured cells in a dose-dependent manner (8). Recently, it has been observed that Cu R+ and Fe 2 § influence the proliferation rate and the degree of differen- tiation in B16 melanoma cells in vitro (9,10), whereas the action of other cations (Ca 2+, Cd 2+, Mg 2+, Li § and Zn 2+) has also been tested on melanoma cells (11,12). The amount of melanin formed from tyrosine can be increased by the presence of Co 2+ or Ni 2§ ions (13).
A program on the potential effects of metal ions on mouse B16 melanoma cells and their mechanism(s) of action has been undertaken. This paper is the first step of the program. It involves a systematic study on the effects of different metal compounds on the cell number and degree of differentiation in mouse B16 melanoma cells in order to identify what metals must receive priorities in the study of the mechanism(s) of toxicity.
MATERIALS AND METHODS
Cell Lines
B16 melanoma cells (25-30 passages), obtained from primitive tu- mors subcutaneously inoculated into C57BL/6J mouse (14), were culti- vated in vitro. Cells were cultured in T75 Falcon flasks containing 20 mL Eagle's min imum essential med ium (Gibco, UK) supplemented with 10% fetal calf serum (Gibco, UK), 100 IU penicillin/mL and 100 i~g/mL strep- tomycin under a 5% CO2 atmosphere, at 37~ Fresh culture med ium was supplied every 48 h.
Biological Trace Element Research Vol. 36, 1993

Effects of Metals on ~elanorna Cells 193
Metal Salts Used
Na2HAsO4"7H20, NaAsO2, NH4VO3, VOSO4~ O, were purchased from Fluka (Switzerland); CdC12.H20, COC12"6H20, Hg(NO3)2, LiC1, MnC12"4H20, NiCI2-6H20 from Merck (FRG); CrCly6H20, Na2CrO 4 from Riedel-de-Ha6n (FRG); MgSO4, Na2SeO 3 from Sigma (St. Louis, MO). All metal salts were dissolved in bidistilled water.
Cell Number Assay
Two types of experiments were carried out here called " three-day" and "seven-day" experiments.
Three-day experiments: 50 • 104 cells were seeded into Petri dishes (6 cm diameter) and incubated with 3 mL Eagle's min imum essential medi- um supplemented with 10% fetal calf serum (MEM-FCS) for 24 h.
Fresh aqueous solutions of different metal salts were then added in aliquots of 300 wE/dish to obtain the required final ion concentration (from 10 -4 to 10-6M). An equal volume of sterile bidistilled water was added to the control cell dishes. After 24 h of incubation with one of these metal ions, the med ium was eliminated from the dishes and cell plates were trypsinated (trypsin 0.1%, EDTA 0.02%). The cells were taken out with 1 mL of PBS and counted in a hemocytometer (Thoma slide).
Seven-day experiments: 15 x 10 3 cells were seeded into Petri dishes under the same experimental conditions. After 24 h of treatment with one of these metals, the med ium was replaced by fresh culture med ium and cell growth was allowed to occur for 5 more d. Then these cells were trypsinated and counted.
Cell Differentiation (Melanin Assay) and Protein Measurements Cells previously recovered in PBS were centrifuged and NaOH was
added in order to achieve a concentration of 106 cells/mL. Total cell proteins were measured with the Bradford method using bovine serum albumin as a standard (15). For melanin assay (16), absorbance was read at 450 nm.
Cell Survival Assay (MTT Test) B16 melanoma cells were plated into 96-well microplates (6 m m
Falcon; 15 • 103 cells/well). These cells were incubated for 24 h in 90 wE MEM supplemented with 10% fetal calf serum in order to allow cell attachment. Aqueous solutions of metal salts or sterile bidistilled water were added (10 wE/well) in order to obtain the required concentrations. The amount of living cells in these two-dimensional cultures was mea- sured by the colorimetric microassay described by Mosmann (17). After
Biological Trace Element Research VoL 36, 199.3

194 Bahbouth et at
24 h of treatment, 10 b~L of a solution of tetrazolium salt (MTT: 3(4,5- dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide, 3 mg/mL) was added to each well for I h. Under these conditions, MTT was reduced by living cells into an insoluble blue formazan product. The latter was then solubilized by the addition of 50 ILL isopropanol-0.08M HCl/well. After homogenizing, the plates were immediately read with a multiwell scan- ning spectrophotometer (Titertek Elisa reader: wavelength 540 nm, refe- rence wavelength 630 nm). The relative absorbances measured in the treated cells were converted into percentages, considering the control values as 100%.
Clonal Growth
Nine hundred cells were seeded into 6-cm diam Petri dishes under the same conditions of culture and treatment as described above. Cells were fixed and stained on d 8 and the number of colonies (>30 cells)/dish was scored.
Statistical Analysis
The results (MTT test, cell and colony counts, protein, and melanin measurements) were compared to the control values by the non parame- tric Mann-Whitney test. The data represent the mean + SE of 3 to 8 replicates for each experimental condition, p Values lower than 0.01 were considered significant.
RESULTS
Effects on Cell Humber
Table 1 shows the effects of metal ions on the number of Bt6 melanoma ceils after 1-d of exposure to the metal as determined by cell count assay. The following conclusions can be drawn:
1. At 10-4M, the number of cells was drastically reduced by V 5+, VO 2+, As 3+, As 5+, Cr 6+, Se 4+, and Hg 2+ (>94% reduc- tion, control assumed as 100%); Cd 2+, Li +, and Co ~+ were also significantly cytotoxic, with rate of toxicity ranging from 34% (Co 2§ to 73% (Cd2+);
2. At 10 5M, VO 2+, V ~+, Cr 6+ were still strongly cytotoxic (more than 77% of toxicity rate). As 3+ as well as Se 4+, Hg 2+, Co 2§ and Cd 2+ displayed a lower but still significant cyto- toxic effect (from 35% (Co 2§ to 43% (As 3+) of the control);
3. At 10-6M, none of the metals tested significantly influenced the B16 melanoma cell populations as far as the number of cells is concerned.
Biological Trace Element Research Vol. 36, 1993
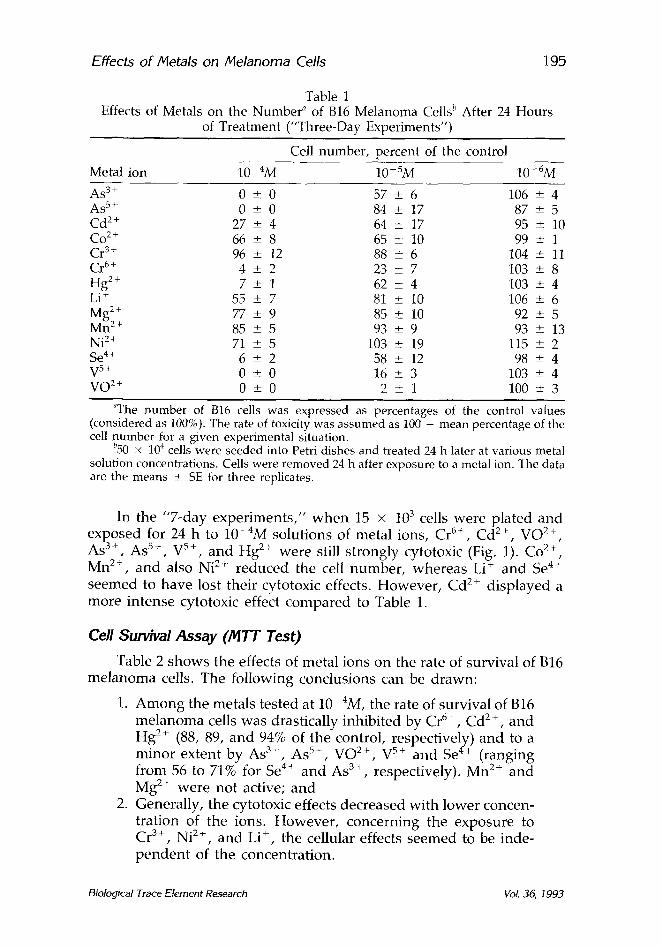
Effects of/~etals on Melanoma Cells 19.5
Table 1 Effects of Metals on the Number ~ of B16 Melanoma Cells l' After 24 Hours
of Treatment ("Three-Day Experiments")
Cell number, percent of the control
Metal ion 10 4M 10-SM 10-6M
As 3+ 0 + 0 57 _+ 6 106 --+ 4 As 5+ 0 -+ 0 84 _+ 17 87 -+ 5 Cd 2+ 27 + 4 64 _+ 17 95 + 10 Co 2§ 66 _+ 8 65 _+_ 10 99 + 1 Cr 3+ 96 _+ 12 88 _+ 6 104 +_ 11 Cr 6+ 4 _+ 2 23 _+ 7 103 _+ 8 Hg 2+ 7 + 1 62 _+_ 4 103 _+ 4 Li + 55 _+ 7 81 +_ 10 106 + 6 Mg 2+ 77 + 9 85 + 10 92 _+ 5 Mn 2+ 85 -+ 5 93 + 9 93 + 13 Ni 2+ 71 _+ 5 103 + 19 115 + 2 Se 4+ 6 -+ 2 58 _+ 12 98 -+ 4 V 5+ 0 + 0 16 ___ 3 103 + 4 VO 2+ 0 -+ 0 2 _+ 1 100 + 3
~The number of B16 cells was expressed as percentages of the control values (considered as 100%). The rate of toxicity was assumed as 100 - mean percentage of the cell number for a given experimental situation.
b50 x 104 cells were seeded into Petri dishes and treated 24 h later at various metal solution concentrations. Cells were removed 24 h after exposure to a metal ion. The data are the means + SE for three replicates.
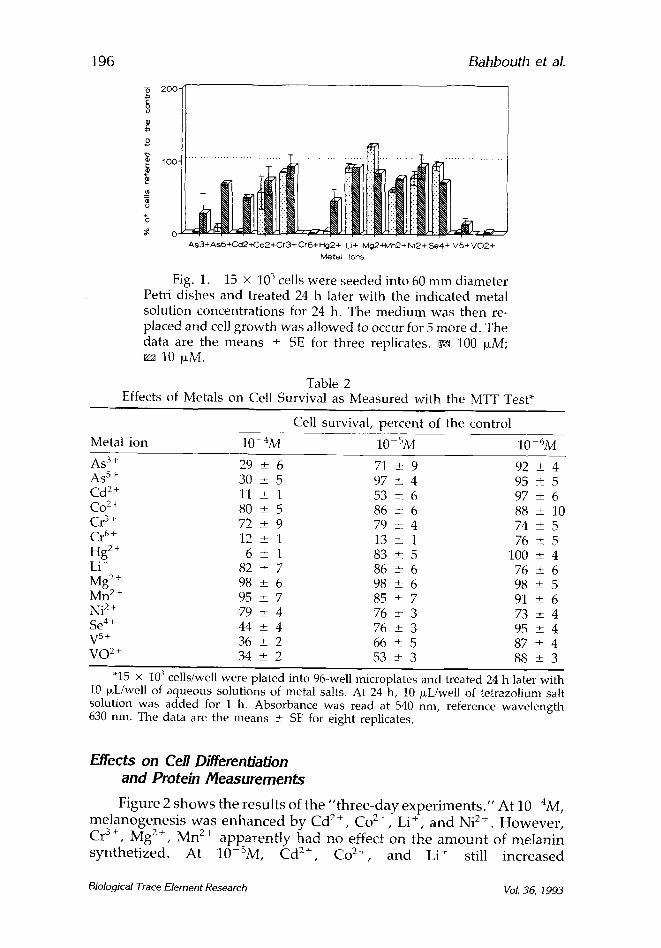
In the "7-day exper imen t s , " w h e n 15 x 103 cells were plated and exposed for 24 h to 1 0 - 4 M solut ions of metal ions, C r 6+, C d 2+, V O 2+, As 3+, As 5+, V 5+, and Hg 2+ were still s t rongly cytotoxic (Fig. 1). Co 2+, Mn 2+, and also Ni 2§ reduced the cell number , whereas Li + and Se 4+ seemed to have lost their cytotoxic effects. However , Cd 2+ d isp layed a more in tense cytotoxic effect compared to Table 1.
Cell Survival Assay (MTT Test) Table 2 shows the effects of metal ions on the rate of survival of
m e l a n o m a cells. The fol lowing conclusions can be drawn:
1. A m o n g the metals tested at 10-4M, the rate of survival of B16 m e l a n o m a cells was drastically inhibi ted by C r 6+, C d 2+, and Hg 2+ (88, 89, and 94% of the control, respectively) and to a minor extent by A s 3+, AS 5+, V O 2+, V 5+ and Se 4+ (ranging f rom 56 to 71% for Se 4+ a n d As 3+, respectively). Mn 2+ and Mg 2+ were not active; and
2. General ly, the cytotoxic effects decreased wi th lower concen- trat ion of the ions. However , concern ing the exposure to Cr 3+, Ni 2+, and Li +, the cellular effects seemed to be inde- p e n d e n t of the concentra t ion.
B16
Biological Trace Element Research VoL 36, t993
196 Bahbouth et aL
As3+AsS+Cd2-~-Co2+Cr3+ Cr6+ Pig24- L.i+ ivlg2+lvln2+ Ni24- Se4+ V5+ V 0 2 § Mete/ ions
Fig. 1. 15 • 103 cells were seeded into 60 m m diameter Petri dishes and treated 24 h later with the indicated metal solution concentrat ions for 24 h. The med ium was then re- placed and cell g rowth was allowed to occur for 5 more d. The data are the means • SE for three replicates, ca 100 laM;
10 ~tM.
Table 2 Effects of Metals on Cell Survival as Measured with the MTT Test*
Cell survival, percent of the control
Metal ion 10-4M 10-SM 10 6M
As 3§ 29 + 6 71 + 9 92 + 4 As 5§ 30 + 5 97 + 4 95 • 5 Cd 2+ 11 + 1 53 • 6 97 _+ 6 Co 2+ 80 _+ 5 86 + 6 88 • 10 Cr 3§ 72 • 9 79 • 4 74 _+ 5 C r 6+ 12 _+ 1 13 + 1 76 • 5 Hg 2+ 6 _+ 1 83 • 5 100 • 4 Li + 82 • 7 86 + 6 76 • 6 Mg 2§ 98 • 6 98 + 6 98 _+ 5 Mn 2+ 95 • 7 85 • 7 91 • 6 Ni 2+ 79 _+ 4 76 • 3 73 • 4 Se 4+ 44 _+ 4 76 • 3 95 _+ 4 V 5+ 36 + 2 66 • 5 87 • 4 VO 2§ 34 • 2 53 + 3 88 • 3
"15 x 10 3 cells/well were plated into 96-well microplates and treated 24 h later with 10 p~Uwell of aqueous solutions of metal salts, At 24 h, 10 b~L/well of tetrazolium salt solution was added for 1 h. Absorbance was read at 540 nm, reference wavelength 630 nm. The data are the means • SE for eight replicates.
Effects on Ce//Differenb'ation and Protein Measurements
Figure 2 s h o w s the r e su l t s of the " t h r e e - d a y e x p e r i m e n t s . " A t 10 4M, m e l a n o g e n e s i s w a s e n h a n c e d b y C d 2+, Co p+, Li +, a n d Ni 2+ . H o w e v e r ,
M - 2 + + Cr 3+, 8 , M n 2 a p p a r e n t l y h a d n o effect o n the a m o u n t of m e l a n i n s y n t h e t i z e d . A t 10 SM, C d 2+, Co 2 . , a n d Li ~- still i n c r e a s e d
Biological Trace Element Research Vol. 36, 1993

Effects of Metals on Melanoma Cells
-2 3oc *E o o
2 200
c_ 10C
E
C AS3+ AsS+ Cot,2,+ Co2+ C~3+ He4- Li+ Mg2+ Mn2+ Ni2+ Se44-
Metal ions
Fig. 2. The melanin contents were measured according to the procedure described by Prasad et al., as mentioned in Materials and Methods. The assay could not be performed for the following metal salts: As 3+ (100 ~M), As s+ (100 ~M), Cr 6+ (10 ~M and 100 ~M), Hg 2+ (100 ~M), Se 4+ (100 ~M), V s+ (10 ~M and 100 p~M), and VO 2+ (10 ~M and 100 ~M) (insufficient number of cells recovered after the treatment). The data are expressed as percentages of the control values, considered as 100% (means _+ SE for three replicates).
100 ~M; ~ 10 p,M.
197
melanogenesis , whereas Se 4+ acted as an inhibitor. When 15 x 10 3 cells were plated for 7 d (figure not shown) , the same effects on mela- nogenesis were noted , but the effects were less intense.
In Figure 3 (" three-day exper iments") at 10-SM, As 3+, As 5+, C d 2+,
Co 2+, Hg 2§ and Li + are s h o w n to increase the protein content in B16 me lanoma cells. On the o ther hand , Se 4+ decreased this content. In the "seven-day exper iments ," for the same molar concentrat ion, As 5+, Cd 2+, Co 2+, Hg 2+, Li § and Mg 2+ as well as Se 4+ increased more or less the prote in content in B16 cells.
Colony Formation
Table 3 reports the results obtained by the colony formation assay. 5 6+ 3+ 2+ 2+ 5+ 5+ 4+ 2+ A t l 0 M, Cr , A s , V O , H g , V , A s , S e , a n d C o had an
inhibit ing effect on the n u m b e r of colonies (more than 33% inhibition as referred to the control). At a h igher concentrat ion (10 4M), nearly no me lanoma cells survived after incubat ion wi th A s 3+, A s 5 +, C d 2+, C r 6+,
Hg 2+, V s +, and VO 2+ . The colony formation efficiency of B16 me lanoma cells u n d e r the control condi t ions was always be tween 10-15%.
DISCUSSION
The effects of four teen metal c o m p o u n d s were invest igated in vitro, on a B16 tumorigenic and melan in-produc ing cell line. At 10-4M, metal
Biological Trace Element Research Vol. 36, 1993

198 Bahbouth et al.
3 0 0 -
2 O O
tO0-
AS3+ ASS+ Cd2+ Co2+ Hg2+ Li+ Mg2+ Ni2+ Se4+ Metal i ons
Fig. 3. The protein contents of B16 cells were measured by the Bradford method (15) and are graphically represented as percentages of the controls (assumed as 100%). After an exposure to 10 ~M solutions of As 3+ (only in the "seven-day experiments"), Cr 6+, V 5+, and VO 2+, the number of cells recovered was too low to allow the assay. The data are ex- pressed as percentages of the control values, considered as 100% (means + SE for three replicates). ~ 3-day exp.;
7-day exp.
ions such as As 3+, As 5+, C d 2+, Cr ~+, Hg 2+, Se 4+, V 5+, and VO 2§ but also Li § and Co 2§ were s h o w n to be able to lower the n u m b e r of B16 cells. The results concerning the inhibitory effects of Cd 2+ and Li + agree with previous observat ions in the literature (11,12). At 10 ~M, the cyto- toxicity of AS 3+, C d 2+, Co 2+ , Cr 6+ , Hg 2+, Se 4+ , V 5§ , and VO 2§ was still significant.
In cells exposed to lower concentrat ions, no significant effects on the n u m b e r of B16 cells were observed. However , the presence of se rum in the culture m e d i u m could well mask the specific effects of some of these metal ions in the cells (18). Metals could bind to proteins present in the serum, thereby decreasing their bioavailability.
At present , we cannot explain the exact mechanism(s) of the ob- served cytotoxic effects; we can only make supposi t ions. Damage to cell membranes could be involved, as in the case of Cd 2+ and Hg 2+ (19), allowing an increased uptake of these metal ions. Initiation of free radical reactions including lipid peroxidat ion by Cd 2+ and Hg 2§ or b inding to thiol g roups may also be involved. Metal ions such as Co 2+ and Cr 6+ are k n o w n to p romote the oxidation of glutathione, thus al ter ing--as in the case of Cr6+-- the reactivity of this ion with cellular componen t s (19). Moreover, another toxicity mechan i sm could be related to the b inding of metal ions such as C d 2+, Hg 2+, and Ni 2+ with glutathione, so that the removal of reactive oxygen species wou ld be impaired (19). However , direct effects on enzymes cannot be excluded. Vanad ium is k n o w n to affect several enzymes involved in the synthesis of DNA and proteins (20), m e m b r a n e phospho l ip ids (21), and t ransport ATPases (22).
Biological T r a c e Element Research Vol. 36, 1993

Effects of Netals on Melanoma Cells 199
TaMe 3 Effects of Metals on the Number of Colonies of B16 Melanoma Cells*
Number of colonies (percent of the control)
Metal ion 10 4M 10-SM
As 3+ 0 • 0 5 -+ ] As 5+ 3 • 0 40 +_ 6 Cd 2+ 0 + 0 82 + 15 Co x+ 32 + 1 67 _+ 7 Cr 3+ 100 +_ 16 86 + 3 Cr 6+ 0 _+ 0 0 + 0 Hg a+ 0 _+ 0 27 + 3 Li + 76 _+ 2 87 + 15 Mg 2+ 104 _+ 3 86 -+ 8 M n 2+ 85 -+ 2 93 +- 8 Ni 2+ 90 • 20 84 ----- 16 Se 4§ 89 +_ 2 63 + 10 V 5+ 0 + 0 35 + 6 VO 2+ 0 _+ 0 23 +- 4
*B16 cells were plated (900 cells per 60 mm diameter dish) and treated 24 h later with metal solutions, as specified in Materials and Methods. Cells were fixed and stained on d 8. The data represent the means -+ SE for three replicates.
Mercury inhibits ATPases and interferes with mitosis and induces D N A damage , w h e r e a s se len ium can slow d o w n cell g rowth in colon cancer by inhibit ing D N A synthesis , w i thou t affecting cell viability; at h igher doses, it is cytotoxic (18).
Our s tudy conf i rms the inf luence of the chemical form of the metals on the rate of their cytotoxicity. At 10-SM, A s 3 § was more cytotoxic than As s+, and VO 2§ was more cytotoxic than Vs+; the rate of toxicity of C r 6+
was about 77%, w h e r e a s Cr ~* did not seem to exert any cytotoxic effect (Table 1). These f indings are in a g r e e m e n t wi th data of the l i terature concern ing the toxic proper t ies of A s 3+ and Cr 6+ (23,24). Surprisingly, VO 2+ was m o r e toxic than V 5+. However , it has recent ly been repor ted that VO 2+ can be rapidly oxidated to V 5+ in cell cul ture media (25). Thus, fur ther exper iments in wh ich the oxidat ion state is r igorously control led are necessary . The relative cytotoxicity and the cellular up take of A s 3+ in BALB/3T3 cells is fourfold h igher than that of As 5+ (23). Fur thermore , cell m e m b r a n e s are part icularly permeable to Cr 6+, w herea s Cr 3+ enters the cells only ve ry slowly (26). For arsenic, ch romium, and vanad ium, a m e c h a n i s m related to toxicity is intracellular g l u t a t h i o n e - d e p e n d e n t re- duct ion, wh ich is necessa ry for their detoxification to less toxic forms, e.g., C r 3 + and VO 2+.
In o rde r to explore w h e t h e r the cytotoxic effects of metal ions could be reversed , cells were also a l lowed to g row for 5 d after exposure ("7- day expe r imen t s" ) (Fig. 1). Compar ing the data in Fig. 1 wi th those in Table 1, it can be obse rved that in the case of se len ium and l i thium, the
Biological Trace Element Research Vol. 36, 1993

200 Bahbouth et aL
cytostatic effects were lost when cells, after exposure to these metal ions, were reincubated in a fresh medium.
At 10-4M, the MTT survival test (Table 2) suggested that the degree of cytotoxicity of the metal ions was either very high (Cr 6+, Cd 2+, and Hg 2+) or high (As 3+, As 5+, VO 2+, V 5+, and Se4+). No apparent inhibi- ting effects were observed for Li +, Mg 2+, and Mn 2+ under our experi- mental conditions. The apparent discrepancy between the results ob- tained by the cell survival assay (Table 2) and those obtained by cell number assay (Table 1) for metal ions such as As 3+, As 5§ Li +, Se 4+, V 5+, and VO 2+ could be caused by the different experimental pro- cedures. Moreover, MTT reduction is based on the activity of a mito- chondrial dehydrogenase. So this test does not allow a clear distinction to be made between an inhibition of a cell function and a real decrease in the number of viable cells.
The relative rate of cellular toxicity of the metal compounds tested on B16 melanoma cells was as follows: Hg 2+ > Cr 6+ = Cd 2+ > Aft 3 ~, As 5+
vB+, VO 2+ ) Se 4+ ) Cr 3+ = Ni 2+ = Co 2+ = Li +. The colony growth assay (Table 3) confirmed these results.
In this study, the mean protein and melanin contents in B16 mela- noma cells were affected by metal ion exposure. In the "three-day experi- ments" (Fig. 3), at 10 5M, As 3+, As 5+, Cd 2+, Co 2+, Hg 2+, and Li + increased (whereas Se 4 § reduced) the protein content of the treated cells. In the "seven-day experiments," at the same molar concentration, we observed an increased amount of cellular proteins, especially in the case of As 5+, Hg 2+, and Se 4+.
The effects of metal exposure on the cellular melanin content was particularly obvious for Co 2+, Ni 2+, Cd 2+, and Li +, which increased this content, and Se 4+, which reduced it (Fig. 2).
Data reported in the literature on the effects of metals on melano- genesis are related to Cu and Fe studied in B16 melanoma cells (9,10). Recently, further evidence of an increase of melanin synthesis by Ni 2+ and Co L§ has been given by Jara et al. (13), who carried out their study on isolated enzymatic systems: Ni 2§ and Co 2§ acted as activators of tyrosine hydroxylation, which is the rate-limiting step in m,~lanogenesis.
Melanin has the capacity to bind metal ions with various degrees of affinity. Among the divalent cations, Pb 2+ has the strongest affinity for melanin, followed by Cu 2§ Ni 2§ Co 2+, and Mn 2+ (27). On the other hand, metal ions like Co 2 + and Ni 2 + are shown to be able to promote the production of hydroxyl radicals (28,29) in vitro. The potential role of melanin interaction with metal ions in melanoma cells is largely un- known. Further studies are in progress to s tudy the significance of such interactions at a cellular level.
REFERENCES
1. C. T. Horovitz, J. Trace Elem. Electrolytes Dis. 2, 135 (1988). 2. W. Mertz, Science 213, 1332 (1981).
Biological Trace Element Research Vol. 36, 1993

Effects of/Vletals on Melanoma Cells 201
3. E. Sabbioni, L. Goetz, and G. Bignoli, Sci. Total Environ. 40, 141 (1984). 4. P. Baccini, Circulation of Metals in the Environment, vol. 18, Siegel, H., ed.,
Dekker, New York, pp. 359-361 (1984). 5. J. D. Heck and M. Costa, Biol. Trace Elem. Res. 4, 71 (1982). 6. J. D. Heck and M. Costa, Biol. Trace Elem. Res. 4, 319 (1982). 7. R. J. Smialowicz, R. R. Rogers, M. M. Riddle, R. J. Garner, D. G. Rowe, and
R. W. Luebke, Environ. Res. 36, 56 (1985). 8. J. Inmon, A. Stead, M. D. Waters, and J. Lewtas, In Vitro 17, 1004 (1981). 9. M. C1. De Pauw-Gillet, B. Siwek, G. Pozzi, E. Sabbioni, and R. J. B. Bassleer,
Anticancer Res. 10, 391 (1990). 10. M. C1. De Pauw-Gillet, B. Siwek, G. Pozzi, E. Sabbioni, and R. J. B. Bassleer,
Anticancer Res. 10, 1029 (I990). 11. J. Borovansky and P. A. Riley, Eur. J. Cancer Clin. Oncol. 19, 91 (1983). 12. J. Nordenberg, H. Paneth, L. Wasserman, A. Fuchs, and A. Novogrodsky,
Isr. J. Med. Sci. 21, 867 (1985). 13. J. R. Jara, F. Solano, J. C. Garcia-Borron, P. Aroca, and J. A. Lozano,
Biochim. Biophys. Acta Gen. Subj. (Netherlands) 1035, 276 (1990). 14. M. C1. De Pauw-Gillet, J. Hennet, and R. J. B. Bassleer, Eur. J. Cancer Clin.
Oncol. 21, 951 (1985). 15. M. M. Bradford, Anal. Biochem. 72, 248 (1976). 16. K. N. Prasad and J. Edwards-Prasad, Cancer Res. 42, 550 (1982). 17. T. Mosmann, J. Immunol. Meth. 65, 55 (1983). 18. J. L. Nano, D. Czerucka, F. Menguy, and P. Rampal, Biol. Trace Elem. Res.
20, 31 (1989). 19. N. T. Christie and M. Costa, Biol. Trace Elem. Res. 6, 139 (1984). 20. W. M. Bracken and R. P. Sharma, Biochem. Pharmacol. 34, 2465 (1985). 21. N. H. Stacey and C. D. Klaassen, Toxicol. Appl. Pharmacol. 58, 8 (1981). 22. K. M. Chart and K. D. Junger, J. Biol. Chem. 258, 4404 (1983). 23. F. Bertolero, G. Pozzi, E. Sabbioni, and U. Saffiotti, Carcinogenesis 8, 803
(1987). 24. S. De Flora, D. Serra, A. Camoirano, and P. Zanacchi, Biol. Trace Elem. Res.
21, 179 (1989). 25. G. Pozzi, E. Sabbioni, A. Pintar, A. Rossi, and L. Casella, in preparation. 26. K. E. Wetterhahn and J. W. Hamilton, Sci. Total Environ. 86, 113 (1989). 27. B. Larsson and H. Tj~ilve, Acta Physiol. Scand. 104, 479 (1978). 28. S. Kawanishi, S. Inoue, and K. Yamamoto, Biol. Trace Elem. Res. 21, 367
(1989). 29. R. Shainkin-Kestenbaum, C. Caruso, and G. M. Berlyne, Biol. Trace Elem.
Res. 28, 213 (1991).
Biological Trace Element Research Vol. 36, 1993