effects of peptidase inhibitors and other additives on fermentation and nitrogen distribution in...
TRANSCRIPT

Journal of the Science of Food and Agriculture J Sci Food Agric 79 :679–686 (1999)
Effects of peptidase inhibitors and otheradditives on fermentation and nitrogendistribution in perennial ryegrass silageVictor L Ns erekoº and John A Rooke*Animal Biology Divis ion, SAC,Craibs tone Es tate, Aberdeen,AB21 9YA,UK
Abstract : The eþ ects of formic acid, three concentrations of formaldehyde in a formic acid/formaldehyde mixture and cysteine-peptidase inhibitors, 1-trans epoxysuccinyl-leucylamido-(4-
guanidino) butane (E-64), N-ethylmaleimide and cystamine on nitrogen (N) distribution during
ensilage of perennial ryegrass (Lolium perenne) were investigated. A third cut or perennial ryegrass
(163 g dry matter kg—1 and 61 g water-soluble carbohydrate kg—1 dry matter) was ensiled in two silo
sizes; formaldehyde-treated herbage was ensiled only in larger silos (500–550 g herbage) and cysteine-
peptidase inhibitor-treated herbage only in smaller silos (130–150 g herbage). Control silages were
poorly fermented but contained low concentrations of butyric acid and ammonia N indicating little
activity of spoilage bacteria. Formic acid increased peptide N concentrations (P Æ 0.01) in silage from
smaller silos but had little eþ ect on other N constituents; in the larger silos, formic acid reduced
soluble non protein nitrogen (NPN) and ammonia-N concentrations and increased peptide N concen-
trations. Increments in formaldehyde reduced silage soluble and ammonia N concentrations (linear
eþ ect ; P Æ 0.001). N-Ethylmaleimide and E-64 reduced soluble NPN concentrations (P Æ 0.05) but had
little eþ ect on other N constituents. Cystamine, however, increased silage peptide N concentrations.
Gel ültration on Sephadex G-25 of silage juice prepared from control and formic acid-treated silages
suggested that most silage peptides were small, with molecular weights of less than 520 Da.
1999 Society of Chemical Industry(
Keywords: grass silage proteolysis ; peptides ; cysteine peptidase inhibitors
INTRODUCTION
During ensilage, plant endopeptidases degradesoluble herbage protein to peptides and free aminoacids.1 Silage-soluble nitrogen (N), therefore, con-sists predominantly of non-protein-N (NPN). As aconsequence, the supplies of silage N and energy-yielding substrates to rumen microorganisms may bepoorly synchronised and this may result in rumenmicrobial protein synthesis being less efficient thanin animals fed fresh or dried forages.2 Inhibitingproteolysis during ensilage could improve the uti-lisation of silage N since the efficiency of rumenmicrobial N synthesis can be improved by supple-menting silage with protein N rather than NPN.3However, although formic acid4 and formaldehyde5restricted proteolysis during ensilage, only formalde-hyde subsequently improved silage N utilisation.6h8Recent attempts to characterise the enzymes involvedin proteolysis during ensilage have revealed thataspartic- (EC 3.4.23) and cysteine- (EC 3.4.22) endo-peptidases were present in perennial ryegrass (PRG)and that speciüc inhibitors of these classes of pepti-dases reduced proteolysis in ensiled PRG.9
Several studies have suggested that a considerableproportion of silage N can be present as peptideN.10h13 However, little is known about factors thatinýuence silage peptide N concentrations and therehave been no reported attempts to characterise silagepeptides. The objectives of the study were to investi-gate the eþects of a speciüc cysteine peptidase inhib-itor, 1-trans epoxysuccinyl-leucylamido-(4-guanidino) butane (E-64), and two general cysteinepeptidase inhibitors, N-ethylmaleimide14 andcystamine15 in comparison with formic acid, mix-tures of formic acid and formaldehyde and an inocu-lant of Lactobacillus plantarum on the chemicalcomposition of ensiled PRG, with particular empha-sis on the distribution of silage N. Secondly, gel ül-tration was used to characterise silage peptides.
MATERIALS AND METHODS
Herbage
Perennial ryegrass (Lolium perenne) was harvestedfrom experimental plots at MacRobert Farm, Craib-stone Estate, Aberdeen as a third cut in September1992, using a precision chop forage harvester. Grass
º Pres ent addres s : Res earch Center, Agriculture and Agri-Food
Canada, Lethbridge, AB, Canada, T1J 4B1.
* Corres pondence to : A Animal Biology Divis ion,John Rooke,
SAC, Craibs tone Es tate, Aberdeen, AB21 9YA, UK
(Received 3 March 1998; revis ed vers ion received 24 July 1998;
accepted 24 Augus t 1998)
( 1999 Society of Chemical Industry. J Sci Food Agric 0022-5142/99/$17.50 679

VL Nsereko, JA Rooke
Table 1. Treatments impos ed on gras s at ens ilage
Treatment Application rate Silo s ize
Control 20ml kgÉ1 150 g and 500 g
Formic acid 5.4 g formic acid kgÉ1 150 g and 500 g
Inoculant 106 cfu L plantarum gÉ1 150 g and 500 g
FFL 2.7 g formic acid and 0.51 g formaldehyde kgÉ1 500 g
FFM 2.7 g formic acid and 1.02 g formaldehyde kgÉ1 500 g
FFH 2.7 g formic acid and 1.53 g formaldehyde kgÉ1 500 g
E-64a 22.5mg kgÉ1 and 106 cfu L plantarum gÉ1 150 g
N-Ethylmaleimide 22 g kgÉ1 and 106 cfu L plantarum gÉ1 150 g
Cys tamine dihydrochloride 50 g kgÉ1 and 106 cfu L plantarum gÉ1 150 g
a E-64, 1-trans epoxys uccinyl-leucylamido-(4-guanidino) butane
All additives were applied with water at an application rate of 20ml kgÉ1 FW
was ensiled without wilting in two sizes of silo. Thesmaller silos consisted of plastic pipe, 32mm diam-eter by 250mm length, ütted with screw capsequipped with rubber O-rings and sealed withacetate-free ýexible sealant. Gas release was achievedthrough a 1mm hole in the screw cap, sealed withheavy-duty adhesive tape. Larger silos consisted ofplastic pipe, 50mm diameter by 0.5m length, sealedat each end with a rubber bung and acetate-free ýex-ible sealant. The upper bung was equipped with afermentation lock (Boots Pharmaceutical Company,UK) also sealed into place with ýexible sealant. Thelarge and smaller silos were packed with 500–550gand 130–150g of grass, respectively. Triplicate siloswere ülled for each treatment and size of silo.
Additives
Additives were prepared on the day of ensilage andapplied to grass in a total of 20ml kg~1 fresh weightof herbage (FW). Cystamine, as the dihydrochloride,and N-ethylmaleimide were applied as solids fol-lowed by application of water at 20ml kg~1 FW ;these additives were applied as solids in order toavoid addition of large volumes of water because ofthe large quantities of additive applied. Rates ofaddition were calculated from the amounts ofcystamine15 and N-ethylmaleimide14 reported toinhibit cysteine endopeptidases. The peptidaseinhibitors were obtained from Sigma Chemical Co(Poole, UK). Water alone was applied as a negativecontrol (Control). The inoculant (Ecosyl, EcosylProducts, Billingham, UK) was prepared accordingto the manufacturer’s instructions and diluted so thataddition at a rate of 20ml kg~1 resulted in the appli-cation of 106 L plantarum g~1 grass. The amounts ofadditives applied and silo sizes are given in Table 1.Additives were thoroughly mixed with grass beforepacking into silos.
Analytical procedures
Silos were opened after 100 days. Dry matter (DM)concentrations were determined by drying at 80¡Cfor 24h in a forced draft oven. Juice was extractedfrom the silages using a hand operated press and cen-
trifuged at 10000] g for 15min to remove particu-late material. Extracts were prepared from freshherbage by homogenising 10g in 100ml water in aWaring blender for 1min, followed by centrifugationat 10000] g for 15min. Water-soluble carbohydrate(WSC) concentrations were measured on dilutions offresh herbage extracts or silage juice using theanthrone procedure.16 Ethanol, lactic acid and vola-tile fatty acid concentrations (VFA) in silage juicewere measured by HPLC as described previously.17Total- and soluble-N concentrations in silage juiceand herbage extracts were determined by the Kjel-dahl procedure and ammonia-N by autoanalyserusing the phenol hypochlorite procedure (Techniconmethod 321-74A). Peptide-N concentrations weredetermined on silage juice using the ýuorescaminepeptide assay,18 except that ýuorescence intensitieswere determined 30min after addition of ýuores-camine to suitably diluted silage juice.
For gel ültration, Sephadex G-25 (medium grade,Sigma Chemical Company, Poole, UK) was packedinto a column (26mm diameter and 700mm length ;Pharmacia LKB Biotechnology, St Albans, UK).Equal volumes of juice from triplicate silos of controland formic acid-treated silages were pooled to givetwo samples. Trichloroacetic acid (TCA) was thenadded (ünal concentrations 40mg TCA ml~1) andthe extracts centrifuged at 18000] g for 15min.The supernatants were stored at [ 20¡C untilrequired. TCA-soluble silage juice (3.5ml) wasapplied in turn to the column using distilled water asthe eluant at a ýow rate of 60ml h~1 and 3.75mlfractions collected. Trypticase (1mg ml~1 ; 4ml) wasalso applied to the column as a peptide preparationof known characteristics.18 Absorbance was deter-mined at 206nm using a Uvikon 810 spectrophotom-eter (Kontron Instruments, Zurich, Switzerland) andýuorescamine-reactive peptide-N concentrations asdescribed above.
Statistical analysis
Data were analysed by one-way analysis of variance.For the smaller silos, control means were comparedwith treatment means using Dunnett’s test.19
680 J Sci Food Agric 79 :679–686 (1999)

Fermentation and nitrogen distribution in ryegrass silage
Orthogonal contrasts were used to identify between-treatment diþerences for silage made in larger silos.Contrasts were: control vs other additives ; inoculantvs all formic acid containing additives ; formic acid vsformic acid/formaldehyde additives ; linear eþect offormaldehyde; quadratic eþect of formaldehyde.
RESULTS AND DISCUSSION
Herbage and fermentation characteristics
The herbage ensiled had a DM content of 163g kg~1FW and WSC and N concentrations of 61 and40g kg~1 DM, respectively. These low WSC andhigh N concentrations were typical of a vegetative
third cut PRG and represented material likely to bedifficult to ensile.
Silages made in the smaller silos (Tables 2 and 3were generally less well fermented than those madein larger silos, despite the fact that the same batch ofherbage was used for both silo sizes. This diþerencein fermentation between silo sizes may have been theresult of a larger surface area to volume ratio in thesmaller silos resulting in more oxygen being trappedin these silos and allowing the utilisation of WSC byrespiration rather than fermentation, In all silages,the recovery of fermentation products plus residualWSC was greater than the WSC concentration in theensiled herbage. This anomaly has been noted before
Table 2. The effects of additives on pH and DM (g kgÉ1 FW), WSC and fermentation products (g kgÉ1 DM) in s ilages made in larger s ilos
Treatment a SED c Significance of contras ts b
Control Formic Inoculant FFL FFM FFH 1 2 3 4 5
acid
pH 6.1 4.1 5.3 4.2 4.4 4.6 0.11 *** *** * ** NS
DM 161 176 167 181 180 181 3.5 *** *** NS NS NS
WSC 11 32 11 24 68 98 10.7 ** *** ** *** NS
TFP 81 47 71 96 51 34 10.9 * NS NS *** NS
Lactic acid 12 20 27 67 27 11 8.0 *** *** NS NS NS
Formic acid 2 12 2 10 11 14 1.6 *** *** NS * NS
Acetic acid 41 4 32 9 5 3 3.3 *** *** NS NS NS
Propionic acid 4 1 3 1 2 2 0.4 *** *** * * NS
Ethanol 17 10 6 8 7 4 2.4 *** NS NS NS NS
Butyric acid 5 NDd 1 1 1 ND 1.0 *** NS NS NS NS
a For details of treatments s ee Table 1
b Contras ts : 1, Control v additives ; 2, inoculant v other additives ; 3, Formic acid v FFL, FFM and FFH; 4, Linear effect of formaldehyde; 5,
quadratic effect of formaldehyde
c SED, Standard error of difference for triplicate s ilos ; NS, not s ignificant ; *, P\ 0.05 ; **, P\ 0.01 ; ***, P\ 0.001d ND, not detected. TFP, total fermentation products
Table 3. The effects of additives on pH, dry matter (DM, g kgÉ1 FW) and WSC and fermentation products (g kgÉ1 DM) in s ilages made in
s maller s ilos a
Control Formic acid L plantarum and SEDb
Alone E-64 N-ethyl maleimide Cys tamine
pH 5.4 5.3 5.3 5.4 5.0 5.0 0.22
DM 132 135 135 133 162** 170** 5.5
WSC 15 13 12 12 110** 17 2.8
TFP 137 136 107 125 17** 89 25.9
Lactic acid 20 24 29 22 2 24 15.7
Formic acid 2 23** 4 2 3 2 2.1
Acetic acid 92 40 60 73 12* 54 23.1
Propionic acid 8 4** 7 6 \ 1** 4** 1.0
Ethanol 10 35** 7 8 NDc 6 3.9
Butyric acid 6 11 ND 14 ND ND 7.3
a For details of treatment s ee Table 1
b SED, s tandard error of difference for triplicate s ilos ; *, P\ 0.05 ; **, P\ 0.01 s ignificantly different from the controlc ND, not detected. TFP, total fermentation products
J Sci Food Agric 79 :679–686 (1999) 681
VL Nsereko, JA Rooke
in silages made from low DM forage containing lowWSC concentrations9,20 and attributed to hydrolysisof structural carbohydrates during ensilage.
Control silages from both silo sizes were poorlyfermented as indicated by high pH, high acetic acidconcentations and low lactic acid concentrations(Tables 2 and 3). These fermentation characteristicswere probably a result of low herbage WSC and DMconcentrations. Butyric acid concentrations in bothsilo sizes and ammonia-N concentrations, especiallyin the larger silos, were low (Tables 4 and 5), sug-gesting that the activity of spoilage organisms inthese silages was minimal. It is likely that the highacetic concentrations, particularly in the smallersilos, were caused by conversion of lactic acid toacetic acid, when hexose substrates became limiting,by lactic acid bacteria21 after an initial formation oflactic acid. The low DM concentrations of controlsilaes when compared with some additive-treatedsilages probably resulted from losses of volatilematerial, which were higher in the control silages,while determining oven DM concentrations.22
Inoculant treated silages were slightly better fer-mented than the control silages as indicated by lowerpH and lower concentrations of acetic, butyric andpropionic acids and higher concentrations of lacticacid (Tables 2 and 3). Despite inoculating theherbage with 106 cfu L plantarum g~1 grass, acetate
was the dominant acid produced in inoculated silagesand overall fermentation quality was unsatisfactory.Low herbage WSC and DM concentrations mayhave limited the scope for improvement due to inoc-ulation and it was possible for inoculant lactic acidbacteria to convert lactic to acetic acid when thehexose supply was limiting.21
When compared with control silages, formic acidtreated silages had higher WSC concentrations andmuch lower concentrations of total fermentation pro-ducts, particularly acetic acid in larger silos (Table2). In smaller silos (Table 3), the eþects of formicacid were less marked but acetic acid concentrationwas reduced and ethanol concentrations increased.Generally, when control silages are well preserved,formic acid has the eþect of restricting the pro-duction of lactic and acetic acids and conservingmore of the soluble carbohydrate component.23 Thiswas the case for silage made in larger silos, but insmaller silos, the most abundant acid was acetic acid.
When compared with the silages treated with onlyformic acid (Table 2), silages treated with the threeconcentrations of formaldehyde and with formic acidhad, on average, undergone a more restricted fer-mentation as indicated by higher average pH(P \ 0.05) and higher average WSC concentrations(P \ 0.01). As the amount of formaldehyde applied
Table 4. Total N (g kgÉ1 DM) and s oluble N components of s ilages (g kgÉ1 N) made in larger s ilos
Treatmenta SED c Significance of contras ts b
Control Formic L plantarum FFL FFM FFH 1 2 3 4 5
acid
Total N 42 42 41 45 46 44 1.1 NS ** ** NS NS
Soluble N 604 463 626 470 378 357 16.4 *** *** *** *** *Peptide N 92 331 113 177 154 147 30.6 ** ** *** NS NS
Ammonia N 72 26 80 33 25 23 2.1 *** *** NS *** NS
a For treatments s ee Table 1
b Contras ts , 1, Control v additives ; 2, L plantarum v other additives ; 3, Formic acid v FFL, FFM and FFH; 4, Linear effect of formaldehyde;
5, quadratic effect of formaldehyde
c SED, Standard error of difference between triplicate s ilos ; NS, not s ignificant ; *, P\ 0.05 ; **, P\ 0.01 ; ***, P\ 0.001
Table 5. Total N (g kgÉ1 DM) and s oluble N cons tituents of s ilages (g kgÉ1 N) made in s maller s ilos a
Control Formic acid L plantarum SEDb
Alone E-64 N-ethylmaleimide Cys tamine
Total N 43 44 45 44 46* 45 0.9
Soluble N 724 715 748 569* 565* 618 49.5
Peptide N 63 159** 90 66 40 159** 26.4
Ammonia N 89 96 102 92 50* 68 17.4
a For details of treatments s ee Table 1
b SED, s tandard error of difference for triplicate s ilos ; *, P\ 0.05 ; **, P\ 0.01.Significantly different from the control
682 J Sci Food Agric 79 :679–686 (1999)

Fermentation and nitrogen distribution in ryegrass silage
increased there was a progressive restriction of fer-mentation as noted by the increases in pH and inWSC concentrations (linear eþect, P \ 0.001) anddecreases in total fermentation products. Lactic,acetic and butyric acid concentrations declined witheach increase in formaldehyde concentration;however, these relationships were not statisticallysigniücant. Other studies24,25 have found that thebacteriostatic eþects of formaldehyde, which cause arestriction of fermentation, are enhanced by formicacid. The silages treated with the medium and highconcentrations of formaldehyde (Table 2) containedhigher concentrations of WSC than those measuredin the herbage prior to ensilage (68 and 99 vs61g kg~1 DM, respectively). Similar observationshave been noted by others10,26 and attributed tohydrolysis of hemicellulose and cellulose to yieldWSC.
Application of E-64 (Table 3) had little or noeþect on the silage fermentation. N-ethylmaleimideand cystamine treatment reduced silage pH andresulted in higher silage DM concentrations whencompared with the control (P \ 0.01) and otheradditive-treated silages. The higher DM concentra-tions observed in N-ethylmaleimide-treated silagesmay be explained by the fact that very little fermen-tation had taken place in these silages and hencethere were minimal losses of volatile componentssuch as fatty acids and ethanol during oven drying.The higher DM concentrations in cystamine-treatedsilages may be attributed to the weight of additiveapplied. Both N-ethylmaleimide and cystamine treat-ments produced silages with trace concentrations ofbutyric acid. N-ethylmaleimide and cystamine treat-ments reduced the concentrations of total fermenta-tion products (P \ 0.01 and P [ 0.05, respectively).In addition, N-ethylmaleimide-treated silages con-tained higher WSC concentrations (P \ 0.01) thanthe control but a similar total concentration of fer-mentation products plus residual WSC. Despite thelow concentration of fermentation products, N-ethylmaleimide-treated silages had a lower pH value(P [ 0.05) than the controls. Unpublished resultsfrom our laboratory have demonstrated that N-ethylmaleimide inhibits the growth of the silagemicroorganisms, L plantarum and Pediococcus pento-saceus in vitro. The inhibitory eþect of N-ethylmaleimide on silage fermentation may thereforebe attributed to both its slightly acidic nature and itstoxicity to silage microorganisms.
N components of silages
Soluble- and ammonia-NThe N components of the large and smaller silos arepresented in Tables 4 and 5, respectively. Thesilages resulting from treatment with cystamine con-tained signiücantly greater concentrations of N thanthe control silage because of cystamine-N. For pur-poses of comparison, the data in Table 5 for
cystamine-treated silages are corrected forcystamine-N assuming that all cystamine-N wassoluble. The poorer fermentation which had takenplace in the smaller silos was also apparent in the Ncomponents of the smaller silos, in that soluble-Nconcentrations were higher in the small than largersilos indicating more extensive proteolysis had takenplace. Inoculant-treatment had no eþect on silagesoluble- or ammonia-N concentrations in either silosize nor did formic acid-treatment in the smallersilos. However in the larger silos, formic acid-treatment reduced soluble- and ammonia-N concen-trations. The rate of fall of pH is important indetermining the extent of proteolysis in the silo. IfpH is slow to fall, more protein will be brokendown27 as the optimum pH for plant peptidase activ-ity is between 5 and 7.28 In the present experiment,therefore, the lack of any reduction in proteolysiswhen the inoculant and formic acid were applied tosilage made in the smaller silos was consistent withthe inability of the additives to reduce silage pHbelow 5.28
Increments in formaldehyde concentration (20,41and 61g kg~1 crude protein) decreased total soluble-N (linear eþect (P \ 0.001; quadratic P \ 0.05) andammonia-N concentrations (linear eþect P \ 0.001,Table 4). These observations were in accordancewith previous ones.5 Formaldehyde is more eþectiveat reducing proteolysis in the presence of formicacid.25,29,30 The increase in protein preservationoften observed when a combination of formic acidand formaldehyde is applied to grass at ensilage,using commercial application rates, may be attrib-uted to the diþering mechanisms by which thesecompounds restrict protein breakdown. Whenapplied together, formaldehyde binds herbage pro-teins and in doing so, renders the protein unde-gradable by endopeptidases,25 whereas formic acidrapidly reduces pH to a value at which proteolysis islimited.4 In silage, ammonia is predominantly gener-ated by deamination of amino acids by micro-organisms.11 Therefore, intensive proteolysis mayoccur without any signiücant increase in ammoniaconcentrations.23 In the present study, the reductionin ammonia N concentrations (Table 4) in formicacid and formaldehyde/formic acid treated silagesmay reýect either a reduction in soluble precursorsfor ammonia production or of bacterial proteolyticactivity.
E-64 (22.5mg kg~1 FW) reduced soluble-N con-centrations to 79% (P \ 0.05) of those in untreatedcontrols (Table 5). Wetherall31 ensiled ryegrass withE-64 (22.5mg kg~1 FW) and, as a result, soluble-and ammonia-N concentrations were reduced to 79%and 78% of control concentrations. In another studyby the same author, PRG was treated with a higherconcentration of E-64 (44.5mg kg~1 FW) andsoluble N concentrations were reduced to 86% of thecontrol with no reduction in ammonia-N concentra-tions. In the present study, E-64 had no eþect on
J Sci Food Agric 79 :679–686 (1999) 683
VL Nsereko, JA Rooke
ammonia-N. Wetherall et al9 concluded that thereduction in soluble N concentration on applicationof E-64 suggested that cysteine-endopeptidases werepresent in PRG and involved in proteolysis duringensilage. The present study supports these conclu-sions.
Cystamine and N-ethylmaleimide treated silagescontained lower concentrations of soluble-N (Table5), however this diþerence was only signiücant forthe N-ethylmaleimide treated silages (P \ 0.05). Inaddition, treatment with N-ethylmaleimide reducedsilage ammonia-N concentrations. The reductions insoluble-N concentrations suggest that generalcysteine-peptidase inhibitors may be as eþective asthe speciüc inhibitor, E-64, in reducing proteolysisduring ensilage.
Peptide NThe control silages contained low concentrations ofpeptide-N (Tables 4 and 5). These values are in thesame range as reported values (11–207g kg~1 N)11,32and similar to concentrations measured in com-mercial farm silages.13 Of the additive used in thepresent study, only the formic acid containing addi-tives and cystamine increased peptide-N concentra-tions. The lower concentrations of peptide N(377–443g peptide-N kg~1 soluble-N) observed inthe formaldehyde-treated silages, when comparedwith the silage treated with formic acid alone (715gpeptide-N kg~1 soluble-N) may be attributed to thelower concentration of formic acid (2.7 vs 5.4g kg~1)applied in conjunction with formaldehyde. It there-fore appears that rapid acidiücation of herbage, fol-lowing the application of formic acid, not onlyreduced the breakdown of protein but also that ofpeptides. Alternatively, formic acid itself, and notthe H` ions, may have increased peptide N concen-trations. A direct eþect of formic acid may explainwhy peptide concentrations were increased in thesmaller silos (Table 5) when there was no eþect ofadditive on soluble N concentrations. Unlike E-64and N-ethylmaleimide, which only inhibited proteol-ysis, cystamine inhibited the breakdown of bothprotein and peptides. One explanation for this couldbe the presence of both proteolytic and peptidolyticenzymes in PRG. It has been suggested33 that thepresence of several peptidases would be an advantageto the senescing plant given the range of structuralcomplexity of proteins to be degraded.
Gel filtration of silage juice
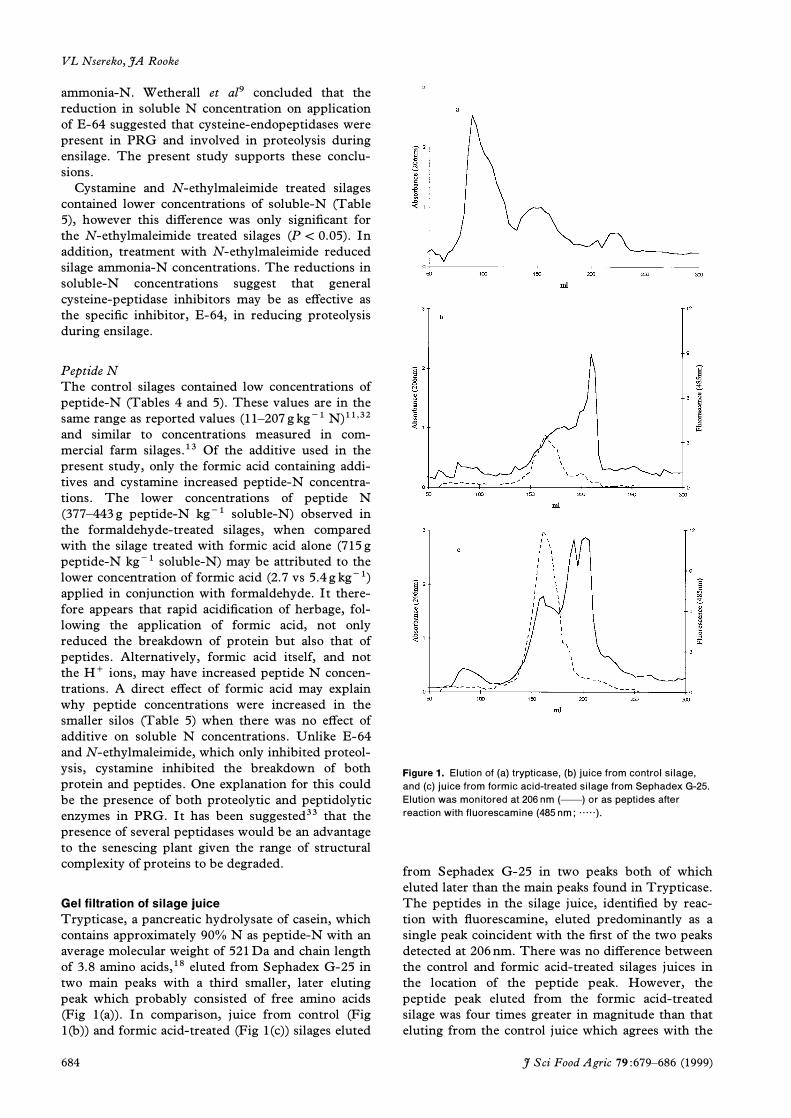
Trypticase, a pancreatic hydrolysate of casein, whichcontains approximately 90% N as peptide-N with anaverage molecular weight of 521Da and chain lengthof 3.8 amino acids,18 eluted from Sephadex G-25 intwo main peaks with a third smaller, later elutingpeak which probably consisted of free amino acids(Fig 1(a)). In comparison, juice from control (Fig1(b)) and formic acid-treated (Fig 1(c)) silages eluted
Figure 1. Elution of (a) trypticas e, (b) juice from control s ilage,
and (c) juice from formic acid-treated s ilage from Sephadex G-25.
Elution was monitored at 206 nm (ÈÈ) or as peptides afterreaction with fluores camine (485 nm; ·····).
from Sephadex G-25 in two peaks both of whicheluted later than the main peaks found in Trypticase.The peptides in the silage juice, identiüed by reac-tion with ýuorescamine, eluted predominantly as asingle peak coincident with the ürst of the two peaksdetected at 206nm. There was no diþerence betweenthe control and formic acid-treated silages juices inthe location of the peptide peak. However, thepeptide peak eluted from the formic acid-treatedsilage was four times greater in magnitude than thateluting from the control juice which agrees with the
684 J Sci Food Agric 79 :679–686 (1999)

Fermentation and nitrogen distribution in ryegrass silage
concentrations of peptide-N measured in the silagejuice. These data suggest that the peptides in silagewere smaller than those found in trypticase (averagemolecular weight, 521Da) and therefore di- and tri-peptides and that the eþect of formic acid treatmentwas to inhibit the peptidase(s) responsible for furtherhydrolysis of these small peptides.
CONCLUSIONS
Proteolysis during ensilage was reduced by treatmentwith formic acid, formaldehyde or peptidase inhibi-tors. The breakdown of peptides to amino acids wasonly reduced by high concentrations of formic acidor cystamine-treatment. Gel ültration of silageextracts using Sephadex G-25 showed that silagepeptides were di- and tri-peptides.
ACKNOWLEDGMENTS
Financial support from the Ministry of AgricultureFisheries and Foods and Zeneca Bio-Products isgratefully acknowledged. SAC receives ünancialsupport from The Scottish Office Agriculture,Environment and Fisheries Department.
REFERENCES1 Ohshima M and McDonald P, A review of changes in nitroge-
nous compounds of herbage during ensilage. J Sci Food
Agric 29:497–505 (1978).2 Siddons RE, Nolan JV, Beever DE and Macrae JC, Nitrogen
digestion and metabolism in sheep consuming diets contain-ing contrasting forms and levels of N. Br J Nutr 54:175–187(1985).
3 Rooke JA and Armstrong DG, The importance of the form ofnitrogen on microbial protein synthesis in the rumen ofcattle receiving grass silage and continuous intraruminalinfusions of sucrose. Br J Nutr 61:113–121 (1989).
4 Carpintero CM, Henderson AR and McDonald P, The eþectof some pre-treatments on proteolysis during the ensilage ofherbage. Grass Forage Sci 34:311–315 (1979).
5 Kaiser AG, Osbourn DF and England P, The inýuence offormaldehyde application at ensilage and of supplementationwith urea or maize starch on the utilisation by calves of rye-grass and red clover silages. Anim Prod 36:371–381 (1983).
6 Beever DE, Thomson DJ, Cammell SB and Harrison DG,The digestion by sheep of silages made with and without theaddition of formaldehyde. J Agric Sci Camb 88:61–70(1977).
7 Siddons RC, Evans RT and Beever DE, The eþect of formal-dehyde treatment before ensiling on the digestion of wiltedgrass silage by sheep. Br J Nutr 42:535–548 (1979).
8 Henderson AR, McDonald P and Anderson DH, The eþect of
silage additives containing formaldehyde on the fermentationof ryegrass ensiled at diþerent dry matter levels and on thenutritive value of direct cut silage. Anim Feed Sci Technol
7:303–311 (1982).9 Wetherall JA, Armstrong DG, Finlayson HJ and Rooke JA,
Reduction of proteolysis during ensilage of perennial rye-grass by protease inhibitors. J Sci Food Agric 68:497–505(1995).
10 Ohshima M, McDonald P and Acamovic T, Changes duringensilage in the nitrogenous components of fresh and additivetreated ryegrass and lucerne. J Sci Food Agric 30:97–106(1979).
11 Heron SJE, Edwards RA and McDonald P, Changes in thenitrogenous components of gamma-irradiated and inoculatedensiled ryegrass. J Sci Food Agric 37:979–985 (1986).
12 Fairbairn RL, Alli I and Phillip LE, Proteolysis and aminoacid degradation during ensilage of untreated or formic acidtreated lucerne and maize. Grass Forage Sci 47:382–390(1992).
13 Rooke JA and Nsereko VL, Peptides in grass silages, in Proc
Tenth Int Silage Conf, Ed by O’Kiely P, O’Connell M andMurphy J, Dublin City University, Dublin, Republic ofIreland, pp 62–63 (1993).
14 Skupin J and Warchalewski J , Isolation and Properties of Pro-tease A from wheat grain. J Sci Food Agric 22:11–15 (1970).
15 Sanner T and Phil A, Studies on the active-SH group ofpapain and on the mechanism of papain activation by thiols.J Biol Chem 238:165–171 (1963).
16 MAFF, The Analysis of Agricultural Materials (2nd edn), HerMajesty’s Stationary Office, London, UK (1982).
17 Rooke JA, Borman A and Armstrong DG, The eþect of inocu-lation with Lactobacillus plantarum on fermentation in labor-atory silos of herbage low in water soluble carbohydrate.Grass Forage Sci 45:143–152 (1990).
18 Wallace RJ, Gel ültration studies of peptide metabolism byrumen microorganisms. J Sci Food Agric 58:177–184 (1992).
19 Dunnett CWA, Multiple comparison procedure for comparingseveral treatments with a control. J Am Stat Assoc 50:1096–1121 (1955).
20 Chamberlain DG and Quig J, The eþects of the rate of addi-tion of formic acid and sulphuric acid on the ensilage of per-ennial ryegrass in laboratory silos. J Sci Food Agric
38:217–228 (1987).21 Lindgren SE, Axelsson LT and McFeeters RF, Anaerobic L-
lactate degradation by Lactobacillus plantarum. FEMS
Microbiol Letts 66:209–214 (1990).22 McDonald P and Dewar WA, Determination of dry matter
and volatiles in silage. J Sci Food Agric 11:566–570 (1960).23 McDonald P, Henderson AR and Heron SJE, The Biochem-
istry of Silage (2nd edn), Chalcombe Publications, Marlow,UK (1991).
24 Brown DC and Valentine SC, Formaldehyde as a silage addi-tive. I. The chemical composition and nutritive value offrozen lucerne, lucerne silage and formaldehyde-treatedlucerne silage. Aust J Agric Res 23:1093–1100 (1972).
25 Barry TN, Eþect of treatment with formaldehyde, formic acidand formaldehyde-acid mixtures on the chemical composi-tion and nutritive value of silage. II. Mature herbage. NZ J
Agric 19:185–191 (1976).26 Morrison IM, Inýuence of some chemical and biological addi-
tives on the übre fraction of lucerne on ensilage in laboratorysilos. J Agric Sci Camb 111:35–39 (1988).
27 Brady CJ, Redistribution of nitrogen in grass and leguminousfodder plants during wilting and ensilage. J Sci Food Agric
11:276–284 (1960).28 Heron SJE, Edwards RA and Phillips P, Eþect of pH on the
activity of Lolium multiýorum proteases. J Sci Food Agric
46:267–277 (1989).29 Siddons RC, Beever DE and Kaiser AG, Evaluation of the
eþect of formic acid and level of formic acid applicationbefore ensilage on silage protein degradability. J Sci Food
Agric 33:609–613 (1982).
J Sci Food Agric 79 :679–686 (1999) 685

VL Nsereko, JA Rooke
30 Valentine SC and Brown DC, Formaldehyde as a silage addi-tive II. The chemical composition and nutritive value oflucerne hay, lucerne silage, and formaldehyde and formicacid treated lucerne silage. Aust J Agric Res 24:939–946(1973).
31 Wetherall J , The characterisation of perennial ryegrass pro-teases and their inhibition during ensilage. PhD Thesis,University of Newcastle Upon Tyne, Newcastle, UK (1993).
32 Hughes AD, The non-protein nitrogen composition of grasssilage II. The changes occurring during the storage of silage.J Agric Sci Camb 75:421–431 (1970).
33 Drivdahl RH and Thimann KV, Proteases of senescing oatleaves 1. Puriücation and general properties. Plant Physiol
59:1059–1063 (1977).
686 J Sci Food Agric 79 :679–686 (1999)