αv integrin subunit is predominantly located in nervous tissue and skeletal muscle during mouse...
TRANSCRIPT

DEVELOPMENTAL DYNAMICS 201:10%120 (1994)
a, Integrin Subunit Is Predominantly Located in Nervous Tissue and Skeletal Muscle During Mouse Development EMILIO HIRSCH, DONALD GULLBERG, FIORELLA BALZAC, FIORELLA ALTRUDA, LORENZO SILENGO, AND GUIDO TARONE Dipartimento di Genetica, Biologia e Chimica Medica, Uniuersita di Torino, 10126 Torino, Italy (E.H., F.B., F.A., L.S., G.T.); Department of Animal Physiology, Uppsala Uniuersity, S-75124 Uppsala, Sweden (D.G.)
ABSTRACT a, integrin subunit can dimerize with different p subunits to form receptors for several matrix proteins. The function of these re- ceptors in vivo is still largely unknown. We exam- ined the localization of a, integrin during mouse development and showed that its distribution is dynamically regulated in the glia of the central nervous system and in skeletal muscle. Immuno- reactivity in the neural tube was firstly localized at embryonic day 10.5 (E10.5) around cell bodies lining the lumen and along tiny fibres extending towards the outer margin. At E12.5 a, distribution follows the highly defined pattern of the radial glia: fascicles of immunoreactive fibres form par- allel palisades, in particular along the hindbrain and the spinal cord. At E15.5, although with weaker intensity, a, was still detectable in radial glia fibres, and it codistributed with glial fibrillary acidic protein positive fascicles. After birth (P8) a, immunoreactivity in the brain and spinal cord de- creased dramatically, but remained high in the ra- dial glia of the cerebellum. In adult mice a, reac- tivity in the central nervous system disappeared. During myogenesis a, appears at E10.5 in myoto- ma1 cells and from E12.5 a, was evident in myo- blasts and in myotubes. In the developing skeletal muscle of E 15.5 embryos, immunoreactivity be- came more concentrated in the apical portion of the myotubes. In adult striated muscle the amount of a, subunit dramatically declined and immuno- staining was no longer detectable. During devel- opment, a, was weakly evident in other sites in- cluding heart and endothelia of blood vessels, mesonephric tubula, smooth muscle of the diges- tive tract, and bronchia. Comparative analysis of the localization of a,, a3, and a5 integrin subunits indicated that a, has a unique and highly regu- lated distribution pattern. The distribution in the nervous system is consistent with a role of a, in neuron-glia interaction during the organization of the neuronal layers in the brain cortex and in the cerebellum. Moreover, a, is likely to be involved in the myotendinous junction during embryonic life, suggesting a dual functional role of this integ- rin in muscle and nervous tissue. 0 1994 Wiley-Liss, Inc.
0 1994 WILEY-LISS, INC.
Key words: a, integrin subunit, Mouse develop- ment, Muscle, Radial glia
INTRODUCTION a, belongs to a large family of transmembrane gly-
coproteins termed integrins. Integrins are heterodi- mers consisting of non-covalently associated a and p subunits that mediate cell adhesion between cells and to extracellular matrix components. Fourteen a sub- units and eight p subunits have been identified so far (Hynes, 1992; Albelda and Buck, 1990). a, associates with at least four different f3 subunits, namely PI, p3, p5, and ps (Vogel et al., 1990; Krissansen et al., 1990; Cheresh et al., 1989; Busk et al., 1992). The complex a,p3, also known as vitronectin receptor, is capable of binding multiple ligands including vitronectin, fibrin- ogen, thrombospondin, von Willebrand's factor, and fi- bronectin (Charo et al., 1990). a,pl and a,& bind fi- bronectin (Vogel et al., 1990; Busk et al., 1992); a,& binds both vitronectin and fibronectin (Cheresh et al., 1989; Pasqualini et al., 1993). These different associa- tions therefore result in a complex network of receptors with distinct specificities.
The functional role of these receptors in vivo during embryogenesis and in adult tissues is still poorly un- derstood. An approach to clarify integrin function is to locate their expression pattern during different em- bryonic stages and possibly correlate it to specific mor- phoregulatory events. Due to the scarcity of reagents suitable for analyzing integrins in embryos, the local- ization of only a few integrin subunits has been de- scribed so far (Muschler and Horwitz, 1991; Duband et al., 1992; Bronner-Fraser et al., 1992; Hirsch et al., 1993). A more extensive analysis is required to estab- lish the expression pattern of each a and p subunit during development. In this study we used an antibody directed to the cytoplasmic domain of a, to analyze its temporal and spatial distribution in the developing mouse embryo. We show that a, is expressed predom- inantly in the radial glia of the central nervous system and in the skeletal muscle. In both tissues expression is
Received May 3, 1994; accepted June 23, 1994. Address reprint requestsicorrespondence to Emilio Hirsch, Diparti-
mento di Genetica, Biologia e Chimica Medica, Sezione de Biologia, Universita di Torino, Via Santena 5 bis, 10126 Torino, Italy.
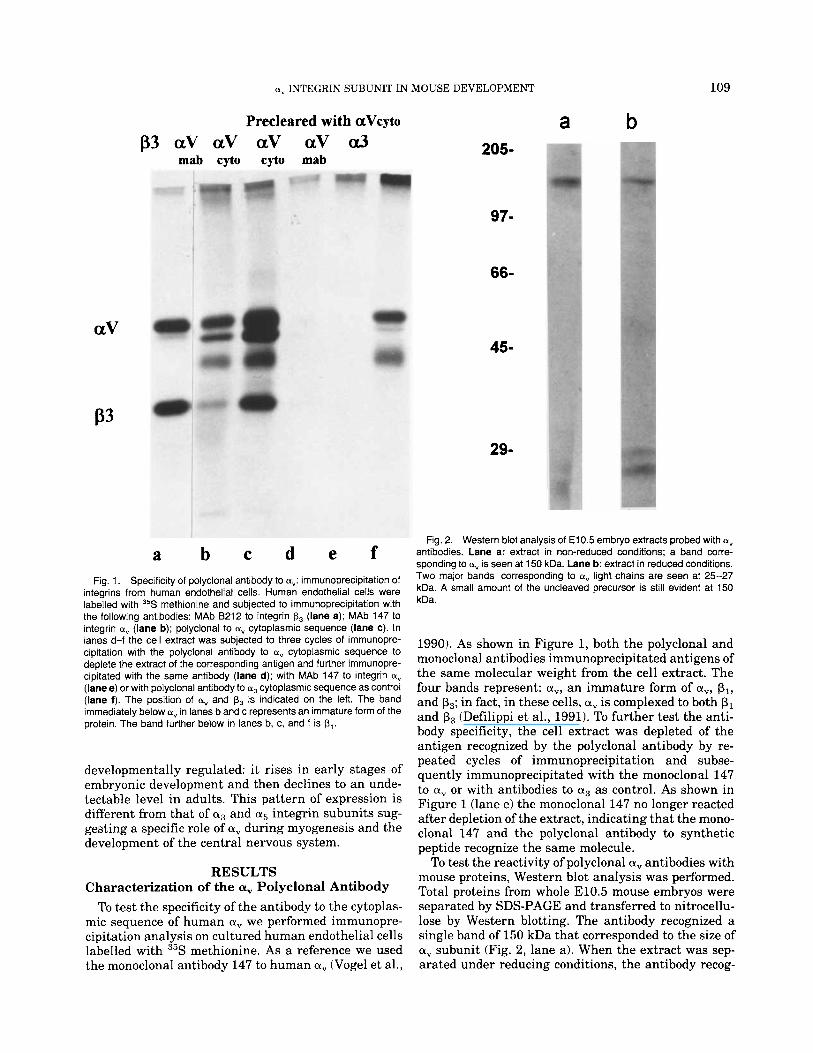
a, INTEGRIN SUBUNIT IN MOUSE DEVELOPMENT
Precleared with aVcyto p3 aV aV aV aV a3
mab cyto cyto mab 205-
97-
66-
aV 45-
P3
29-
Fig. 1. Specificity of polyclonal antibody to a": immunoprecipitation of integrins from human endothelial cells. Human endothelial cells were labelled with 35S methionine and subjected to immunoprecipitation with the following antibodies: MAb B212 to integrin p3 (lane a); MAb 147 to integrin a, (lane b); polyclonal to ct, cytoplasmic sequence (lane c). In lanes d-f the cell extract was subjected to three cycles of immunopre- cipitation with the polyclonal antibody to a, cytoplasmic sequence to deplete the extract of the corresponding antigen and further immunopre- cipitated with the same antibody (lane d); with MAb 147 to integrin ctv
(lane e) or with polyclonal antibody to a3 cytoplasmic sequence as control (lane f). The position of a, and p3 is indicated on the left. The band immediately below av in lanes b and c represents an immature form of the protein. The band further below in lanes b, c, and f is p,.
developmentally regulated: i t rises in early stages of embryonic development and then declines to an unde- tectable level in adults. This pattern of expression is different from that of as and a5 integrin subunits sug- gesting a specific role of a, during myogenesis and the development of the central nervous system.
RESULTS Characterization of the a, Polyclonal Antibody
To test the specificity of the antibody to the cytoplas- mic sequence of human a, we performed immunopre- cipitation analysis on cultured human endothelial cells labelled with 35S methionine. As a reference we used the monoclonal antibody 147 to human a, (Vogel et al.,
a 109
b
Fig. 2. Western blot analysis of El 0.5 embryo extracts probed with a, antibodies. Lane a: extract in non-reduced conditions; a band corre- sponding to ct" is seen at 150 kDa. Lane b: extract in reduced conditions. Two major bands corresponding to av light chains are seen at 25-27 kDa. A small amount of the uncleaved precursor is still evident at 150 kDa.
1990). As shown in Figure 1, both the polyclonal and monoclonal antibodies immunoprecipitated antigens of the same molecular weight from the cell extract. The four bands represent: a,, an immature form of a,, pl, and p3; in fact, in these cells, a, is complexed to both p1 and p3 (Defilippi et al., 1991). To further test the anti- body specificity, the cell extract was depleted of the antigen recognized by the polyclonal antibody by re- peated cycles of immunoprecipitation and subse- quently immunoprecipitated with the monoclonal 147 to a, or with antibodies to a3 as control. As shown in Figure 1 (lane e) the monoclonal 147 no longer reacted after depletion of the extract, indicating that the mono- clonal 147 and the polyclonal antibody to synthetic peptide recognize the same molecule.
To test the reactivity of polyclonal a, antibodies with mouse proteins, Western blot analysis was performed. Total proteins from whole E10.5 mouse embryos were separated by SDS-PAGE and transferred to nitrocellu- lose by Western blotting. The antibody recognized a single band of 150 kDa that corresponded to the size of a, subunit (Fig. 2, lane a). When the extract was sep- arated under reducing conditions, the antibody recog-
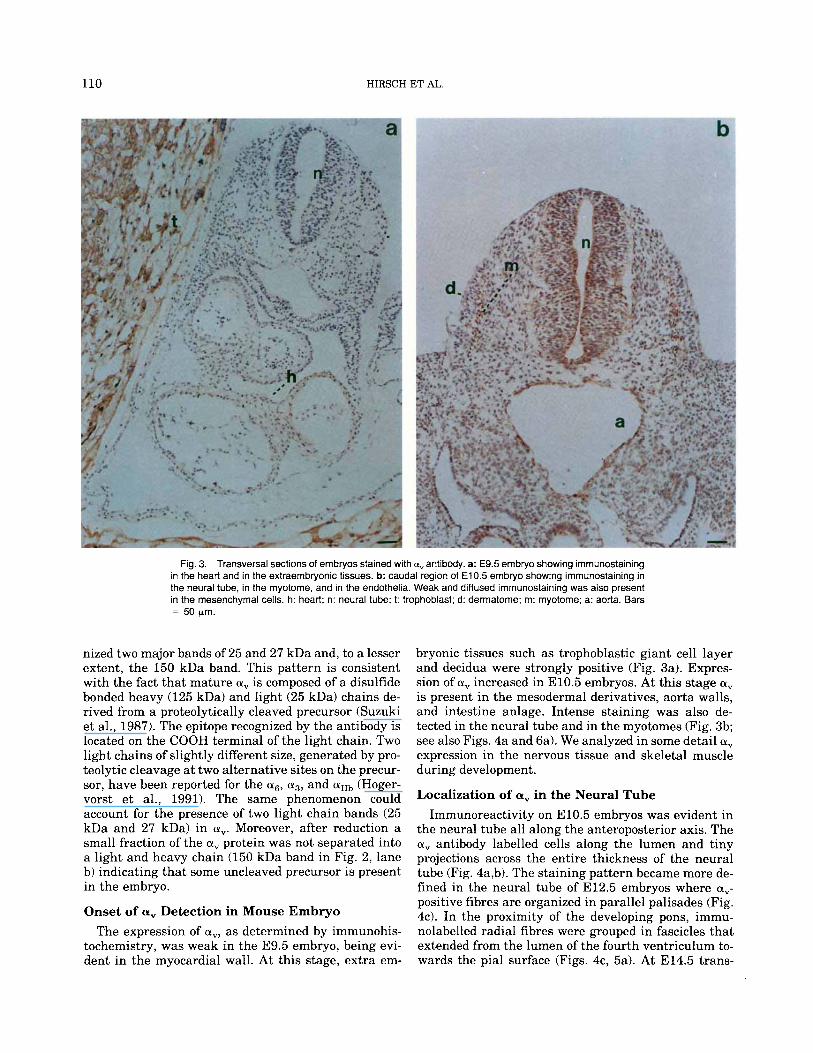
110 HIRSCH ET AL.
Fig. 3. Transversal sections of embryos stained with a, antibody. a: E9.5 embryo showing immunostaining in the heart and in the extraernbryonic tissues. b: caudal region of E10.5 embryo showing immunostaining in the neural tube, in the myotome, and in the endothelia. Weak and diffused irnrnunostaining was also present in the mesenchyrnal cells. h: heart; n: neural tube; 1: trophoblast; d: derrnatorne; m: rnyotorne; a: aorta. Bars = 50 prn.
nized two major bands of 25 and 27 kDa and, to a lesser extent, the 150 kDa band. This pattern is consistent with the fact that mature a, is composed of a disulfide bonded heavy (125 kDa) and light (25 kDa) chains de- rived from a proteolytically cleaved precursor (Suzuki et al., 1987). The epitope recognized by the antibody is located on the COOH terminal of the light chain. Two light chains of slightly different size, generated by pro- teolytic cleavage a t two alternative sites on the precur- sor, have been reported for the a6, a3, and aIIb (Hoger- vorst et al., 1991). The same phenomenon could account for the presence of two light chain bands (25 kDa and 27 kDa) in a". Moreover, after reduction a small fraction of the a, protein was not separated into a light and heavy chain (150 kDa band in Fig. 2, lane b) indicating that some uncleaved precursor is present in the embryo.
Onset of a, Detection in Mouse Embryo The expression of a,, as determined by immunohis-
tochemistry, was weak in the E9.5 embryo, being evi- dent in the myocardial wall. At this stage, extra em-
bryonic tissues such as trophoblastic giant cell layer and decidua were strongly positive (Fig. 3a). Expres- sion of a, increased in E10.5 embryos. At this stage a, is present in the mesodermal derivatives, aorta walls, and intestine anlage. Intense staining was also de- tected in the neural tube and in the myotomes (Fig. 3b; see also Figs. 4a and 6a). We analyzed in some detail a, expression in the nervous tissue and skeletal muscle during development.
Localization of a, in the Neural Tube Immunoreactivity on E10.5 embryos was evident in
the neural tube all along the anteroposterior axis. The a, antibody labelled cells along the lumen and tiny projections across the entire thickness of the neural tube (Fig. 4a,b). The staining pattern became more de- fined in the neural tube of E12.5 embryos where a,- positive fibres are organized in parallel palisades (Fig. 4c). In the proximity of the developing pons, immu- nolabelled radial fibres were grouped in fascicles that extended from the lumen of the fourth ventriculum to- wards the pial surface (Figs. 4c, 5a). At E14.5 trans-

e., INTEGRIN SUBUNIT IN MOUSE DEVELOPMENT 111
versa1 sections of the spinal cord displayed a pattern of radial fibres interrupted in the raphe where variably oriented thick bundles were labelled (Fig. 4d). The specificity of this pattern was confirmed by the absence of staining when antibodies were preincubated with the peptide used for immunization (not shown). Neuro- filament staining indicated that reactivity for a, was restricted to a non-neuronal population of cells (Fig. 5a,b). Moreover, the pattern of a, staining described above suggests labelling of the radial glia (Dupouey et al., 1985). To further investigate this aspect we ana- lyzed the cerebellum of newborn mice where the radial glia has well-defined organization. As shown in Figure 5c, a, was present in fibres that run parallel to each other and extend through the thickness of the cerebel- lar cortex. These structures were also stained with an- tibodies to glial fibrillary acidic protein (GFAP), a well- characterized marker of radial glia (Dupouey et al., 1985; Misson et al., 1991) confirming that a, antibody labels glial cells (Fig. 5d).
After E15.5, a, expression in the brain and spinal cord rapidly decreased and by the time of birth, sec- tions of brain showed no detectable immunoreactivity. In adult brain (Fig. 5e,f) and cerebellum (not shown) a, reactivity disappeared even though some glial cells were still evidenced by GFAP antibodies.
Localization of a, in the Developing Skeletal Muscle
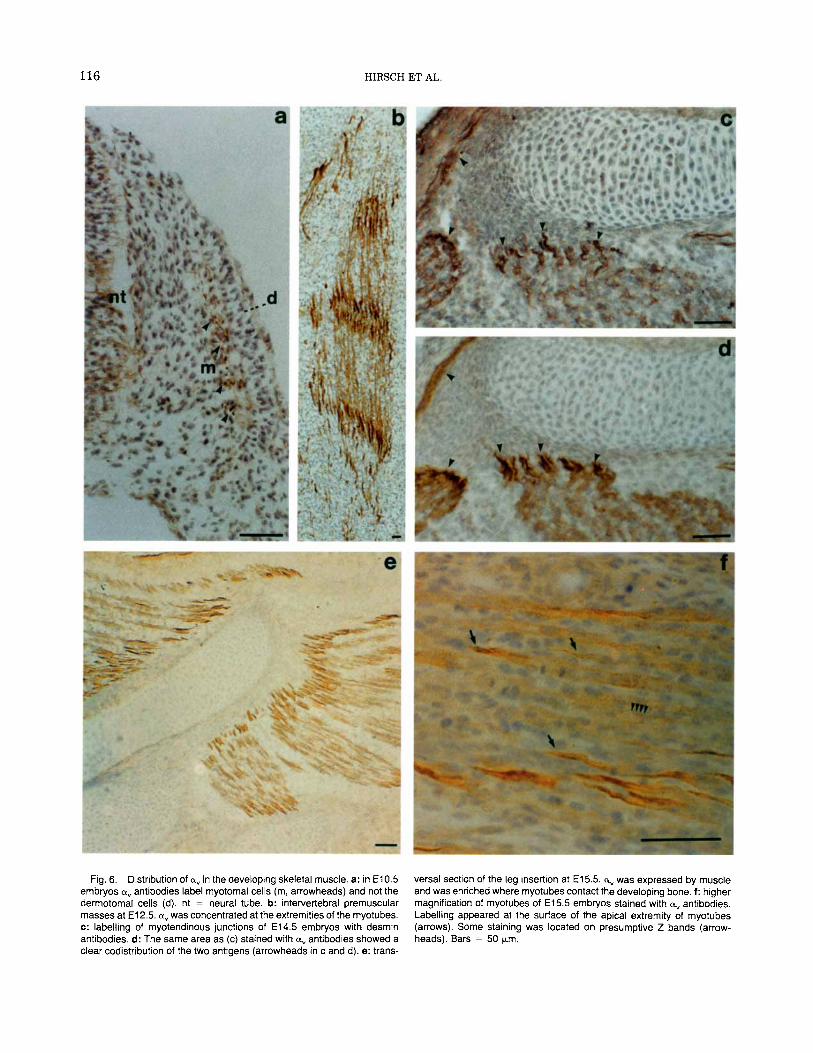
At E10.5 a, reactivity is firstly detectable in somites that have undergone their subdivision in the three compartments: dermatome, myotome, and sclerotome. Strongly immunoreactive cells were located in the re- stricted layer corresponding to the myotome (Fig. 6a). No positive staining for a, could be seen in the der- matome, whereas some expression was noted in the sclerotome.
At E12.5 premuscle masses located between verte- bras, in the body wall, and in the limbs showed a marked immunoreactivity for a, (Fig. 6b). Primary myotubes were strongly labelled and immunostaining is particularly intense a t their ends.
At E15.5 the skeletal muscles showed well-formed myotubes that still possessed high levels of a,. At this stage a clear immunoreactivity was seen on the perim- eter of the myotubes, consistently with the localization of the antigen on the plasma membrane (Fig. 6f). a, is more concentrated in the apical extremities of the myotubes, pointing out their spindle shape. The a,- positive areas were grouped to delimit the apex of each individual muscle and were distributed all around the developing bone (Fig. 6e). This pattern of staining sug- gests that a, was enriched at the myotendinous junc- tion. In fact the same districts were strongly stained also with antibodies that recognize desmin (Fig. 6c,d) which is a muscle specific intermediate filament pro- tein known to be concentrated in the myotendinous junction (Tidball, 1992).
During late muscular development, a, expression de-
creased and no reactivity could be detected on skeletal muscles of adult mice.
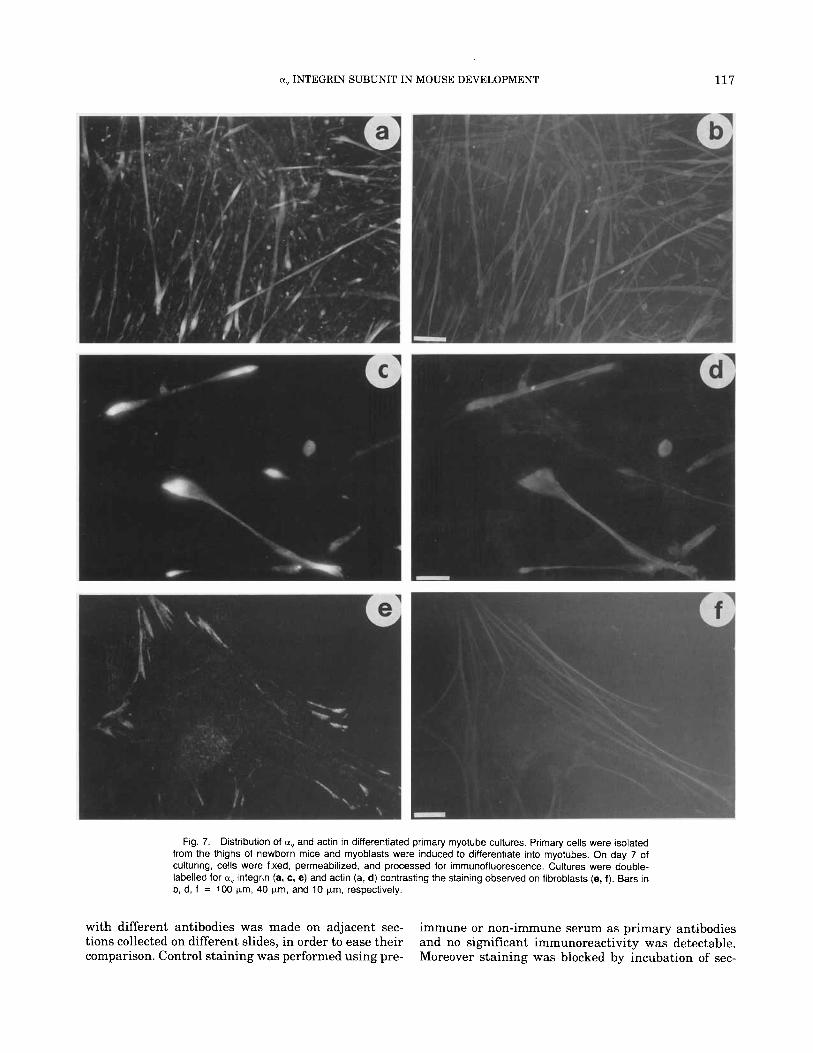
Localization of a, on Cultured Mouse Myotubes In order to compare the staining pattern obtained
with the a, antibody in immunohistochemistry of mus- cle tissue with the staining pattern on myotubes in vitro we established primary cultures from thighs of newborn mice. At the beginning these cultures con- sisted mainly of fibroblast and undifferentiated myo- blasts. With time in culture the fibroblasts produced a fibronectin rich matrix that coated the collagen coated coverslips. At 3 days of culture the first differentiated multinucleated myotubes began to appear. Myotube formation reached its maximum at 7 days. At 2 weeks the myotubes had deposited basement membrane com- ponents on their surface and organized their cytoskel- eta1 proteins into contractile sarcomeres (not shown). Figure 7 shows the staining pattern obtained on differ- entiated day 7 primary cultures containing mainly mononucleated fibroblasts and multinucleated skeletal myotubes. Figure 7a shows a low magnification of the myotube rich cultures stained with a". The immuno- staining of differentiated myotubes antibody showed that a, is enriched at myotube ends. No corresponding enrichment of actin at the end was evident (Fig. 7b). This was more apparent a t the higher magnification of a,/actin stained myotubes seen in Figure 7c,d. In con- trast to the bipolar staining on the elongated myo- tubes, the fibroblasts in these cultures showed a prom- inent staining of a, a t the ends of stress fibres terminating in focal contacts (Fig. 7e,f).
Localization of a, on Other Embryonic Tissues Although a, expression was evident in the develop-
ing nervous and muscular systems, other sites showed weaker immunoreactivity. Heart cells, for example, were firstly shown to possess a, in E9.5 embryos (see Fig. 3a). Myocardium continued to express a low level of the antigen throughout development (not shown). Endothelial cells appeared to be a,-positive from E10.5 embryos (see Fig. 3b) and then remained immunoreac- tive in all the later stages analyzed (i.e., Fig. 4c in E12.5 embryos).
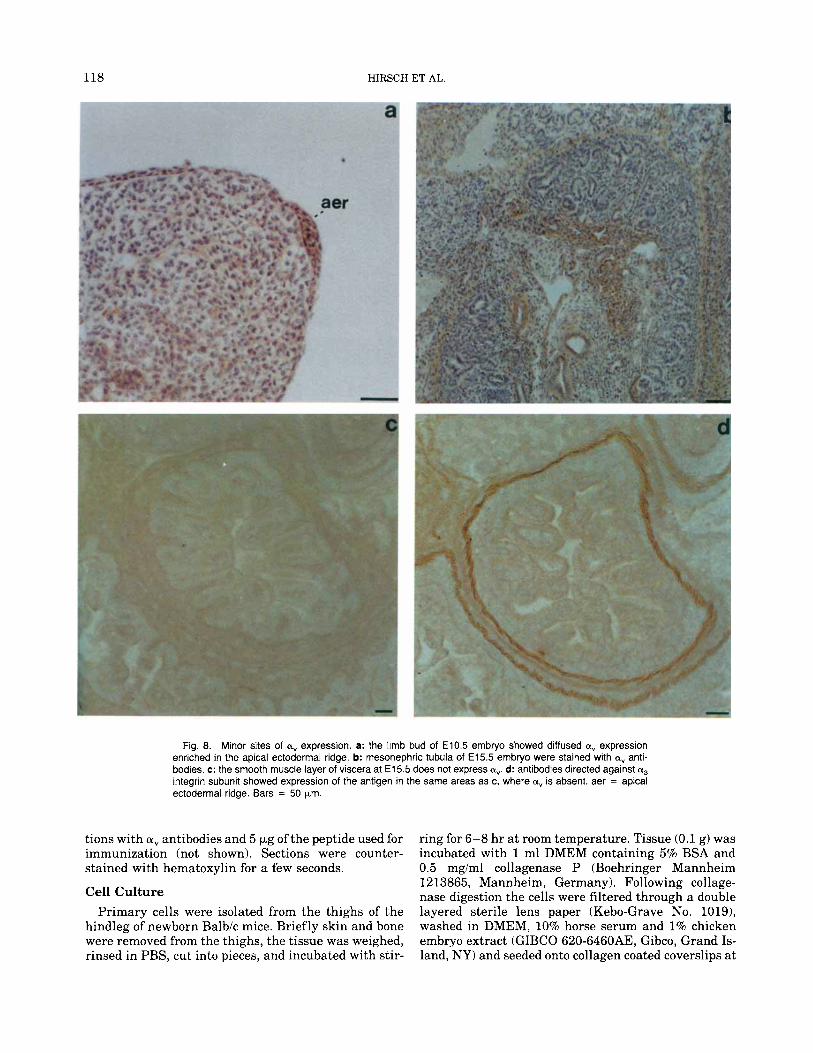
Other sites of expression in the early embryo (E10.5) included the apical ectodermal ridge, known to orga- nize limb development (Fig. 8a). Later, in E15.5 em- bryos, weak reactivity was evident in tissues such as mesonephric tubula (Fig. 8b) and bronchial smooth muscle (not shown). In adult mice the choroid plexus was a,-positive (not shown).
a, Has a Unique Distribution Pattern The pattern of immunoreactivity observed with a,
antibodies is peculiar to this integrin subunit. Compar- ison with the distribution of a5 and a3 revealed that these two subunits were either absent or poorly ex- pressed in the developing nervous tissue and skeletal muscle. Moreover a5 (not shown) and a3 (Fig. 8d) were
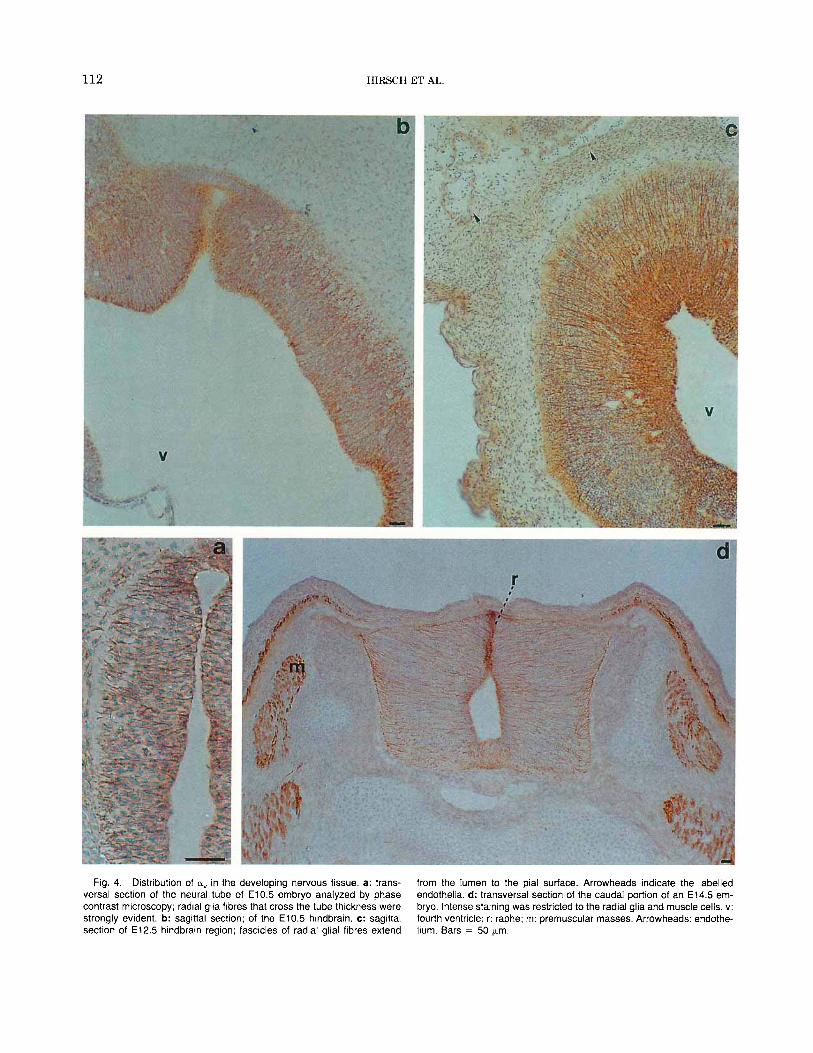
112 HIRSCH ET AL
Fig. 4. Distribution of aV in the developing nervous tissue. a: trans- versa1 section of the neural tube of E10.5 embryo analyzed by phase contrast microscopy; radial glia fibres that cross the tube thickness were strongly evident. b: sagittal section; of the E l 0.5 hindbrain. c: sagittal section of E12.5 hindbrain region; fascicles of radial glial fibres extend
from the lumen to the pial surface. Arrowheads indicate the labelled endothelia. d: transversal section of the caudal portion of an E14.5 em- bryo. Intense staining was restricted to the radial glia and muscle cells. v: fourth ventricle; r: raphe; m: premuscular masses. Arrowheads: endothe- hum. Bars = 50 pn.
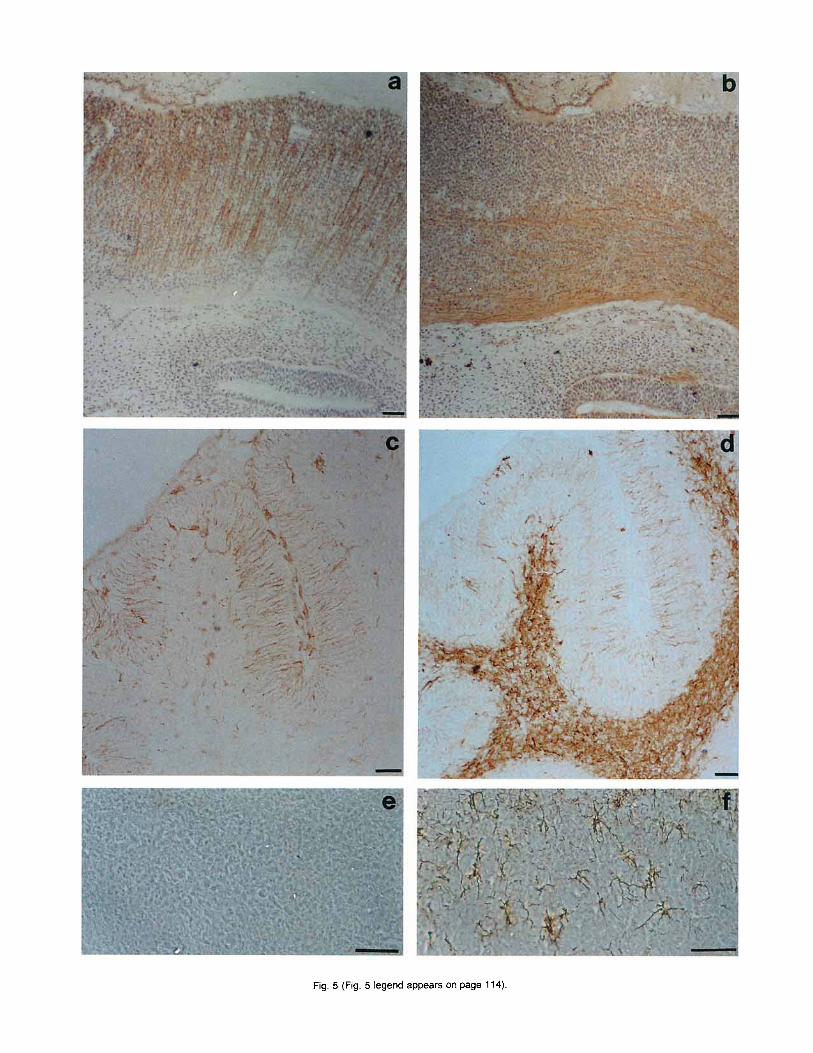
Fig. 5 (Fig. 5 legend appears on page 114).

114 HIRSCH ET AL.
located preferentially in areas where a, expression was weak (Fig. 8c). They were highly expressed in smooth muscle cells of the gut and in some other mesenchymal tissues.
DISCUSSION In this report we have shown a specific pattern of
modulated distribution of the a, integrin subunit in mouse development. Immunoreactivity was very low in E9.5 embryos and appeared in E10.5 embryos. At this stage a, was strongly expressed in the neural tube and in the myotomic portion of the differentiating somites. Later the reactivity distributed on the radial glia and forming myotubes. The a, subunit was down regulated in the adult nervous and muscular tissues.
During the histogenesis of the central nervous sys- tem, neuroblasts are generated in the ventricular zone and migrate radially to generate the neuronal layers present in brain cortex and cerebellum. The migration of neuroblasts and neurons is dependent on radial glial cells whose processes span the entire thickness of the wall of the neural tube, extending from the lumenal to the pial surface. Fascicles of radial glia fibres provide a permissive substrate for neuron migration and axonal guidance during embryonic life. After birth, when neu- ronal migration in brain and spinal cord is terminated, radial glia differentiates into astrocytes (Misson et al., 1991). In the postnatal cerebellum, where granular cells still have to reach their final position, radial glia persists for a short time. Our data show that in brain and cerebellum, a, integrin subunit was expressed in the radial glia cells when neuronal migration occurs and it was down regulated when this phenomenon had ceased. Thus, the presence of a, was linked to the abil- ity of glial cells to support neuronal migration, sug- gesting an important role of this integrin in the adhe- sive function of these cells.
We may speculate that a, on radial glial cells can interact directly with neuronal surface proteins pro- viding a recognition signal necessary for migratory events. Alternatively, the interaction can be indirect: a, can bind an intermediate matrix protein recognized by neuronal receptors. This second mechanism is sug- gested by the fact that fibronectin is located in close association with the radial glia processes and delin-
Fig. 5 (Fig. 5 appears on page 113). Comparative analysis of a,, neurofilament and glial fibrillary acidic protein (GFAP) distribution in em- bryonic and adult tissues. a: sagittal section of the pons region of E12.5 embryo, showing radial glia expressing av. The lumen of the fourth ven- tricle is visible in the upper part, while the cochlea is at the bottom. b: the same region as (a) stained with anti-neurofilament 200 kDa antibodies. Only axons orthogonally oriented with respect to the radial glia were labelled. c: cerebellum at 8 days after birth. Labelling for IY, appeared on the radial glia and capillaries. d: the same area as (c) labelled with GFAP antibody, showing staining of radial glial cells and other glial structures. e: adult olfactory bulb stained with CY, antibodies and photographed in phase contrast. No labelled cells could be detected. f: the same region as (e) stained with GFAP antibodies. Labelling of astrocytes was evident. In c, d, e, 1, counter staining with hematoxylin was omitted. Bars = 50 pm.
eates the pathway for migrating neurons and axonal extensions (Sheppard et al., 1991). Moreover, migrat- ing neurons require p1 integrin subunit, known to form fibronectin receptors (Galileo et al., 1992). Deposition of fibronectin around radial glial fibres may be regu- lated by a, integrins on these cells (Vogel et al., 1990; Conforti et al., 1990).
Other possible explanations could account for the presence of a, integrins on radial glia. a, antigen ap- pears very early (stage E10.5) when radial glial cells are not fully differentiated and do not yet express GFAP. We therefore suggest that a, may be implicated in the formation and orientation of the glial fibres that extend from the ventricular zone to the cortical plate.
The second major site of a, localization was on the skeletal muscles. The presence of a, in premuscular cells was detectable together with the first signs of muscle differentiation (Buckingham, 1992). The immu- noreactivity observed in the myotome appeared at E10.5, 2 days later than the appearance of my%, a transcription factor that is the earliest marker of mus- cular differentiation (Ott et al., 1991). The number of a,-positive myotomal cells increases, however, a t the time when another transcription factor of the same family of myf5, MyoD, is first detected (Sassoon et al., 1989). As myotomal cells reach their definitive places and give rise to the first fusion products, immunoreac- tivity for a, was particularly enriched in the extremi- ties of the primary myotubes. The distribution detected in embryos was consistent with the analysis of in vitro cultured murine myotubes, showing that it was not due to artifactual coartation of fixed tissues. This distribu- tion pattern corresponds remarkably to the staining observed with antibodies to desmin, a cytoskeletal in- termediate filament. Desmin is specifically concen- trated at myotendinous junction (Tidball, 1992), a spe- cialized structure that transmits force generated by muscle contraction to the tendon. Integrins are known to be critical for the adhesive muscle-tendon interac- tions: the Drosophila integrin apsBpps on larval mus- cles is concentrated on muscle insertions (Volk et al., 19901, and mutants lacking pps are unable to form the sarcomere and die as muscles retract from their attach- ment sites (MacKrell et al., 1988; Volk et al., 1990). Recently, vertebrate vitronectin has been found to bind the aps2pps (Hirano et al., 1991; Bunch and Brower, 1992) and promote differentiation of Drosophila myo- blasts in vitro (Gullberg et al., 1994). Moreover aps2pps was found to bind tiggrin, a novel matrix protein spe- cifically localized in muscle attachments in the devel- oping Drosophila embryo (Fogerty et al., 1994).
Other integrin alpha subunits such as alr a4, (Yg, and a7 have been reported to be expressed during myogen- esis (Bronner-Fraser et al., 1992; Duband et al., 1992; Rosen et al., 1992; Bao et al., 1993). ag and a5 have also been detected in cultured myoblasts (von der Mark et al., 1991; Lakonishok et al., 1992). Both a6 and a1 are expressed in the myotome and in early myotubes. a1 disappears long before the hatching, whereas ffg re-

a, INTEGRIN SUBUNIT IN MOUSE DEVELOPMENT 115
mains present, although its expression in adult muscle is ambiguous. The a4p1 receptor is present during mu- rinc foetal myogenesis and its binding the counter re- ceptor VCAM-1 appears to be essential €or the forma- tion of secondary myotubes (Rosen et al., 1992). Recently a7 integrin subunit has been localized in skel- etal muscle. This molecule appears in primary myo- tubes only after their differentiation and it is present in the precursors of the secondary myoblasts (George- Weinstein et al., 1993). In contrast with a, that disap- pears in adult muscle, a7 persists after birth and be- comes concentrated at the myotendinous junction (Bao et al., 1993). Thus the presence of two different integrin subunits in the foetal and adult myotendinous junction suggests important functional changes in this struc- ture during development. The newest addition to the p1 integrin family, a&, has also been reported to be ex- pressed on skeletal muscle although its developmental pattern is not yet known (Palmer et al., 1993). It is interesting to note that most of the integrins that are highly expressed in the developing muscle are known to bind laminins or collagens, from in vitro experi- ments. This is so for al, &g, and a7 (Hynes, 1992). On the other hand, a, forms receptors that can bind differ- ent ligands, vitronectin and fibronectin being the most well defined. a, is known to bind several distinct p subunits such
as pl, P3, p5, ps (Hyncs, 1992). A previous attempt to map the distribution of the a,p3 vitronectin receptor in murine embryos was made with antibodies raised against the complex. The presence of a, is reported in myocytes but not in the nervous tissues (Corless et al., 1992). These differences, however, can be explained when considering that in this study we used an anti- body specifically directed against the a, subunit, a re- agent that broadens the range of detectable a, contain- ing heterodimers. Comparison with the distribution of p1 integrin subunit, which is known to bind a, (Vogel et al., 1990), shows that a, does not codistribute with in the nervous system (Hirsch et al., 1993). is, how- ever, evenly expressed on the membrane of myotubes including myotendinous junctions (Bozyczko et al., 1989), suggesting that i t could be bound to a, in that area. Due to the lack of available reagents, the pattern of expression of the murine p5 or p6 has not been de- scribed so far. It could not be excluded that in the radial glia and in the myotubes a, is bound either to those or to new p subunits.
In conclusion, our results show that a, has a specific tissue distribution, distinct from that of a3 and a5 and it is regulated during mouse development. It is dynam- ically expressed in myotubes and in the radial glia, suggesting possible roles in the formation of the skel- etal muscles and of the nervous system.
EXPERIMENTAL PROCEDURES Embryos and Mice
Donor mice were F1 progeny of C57B and DBA in- bred strains. After mating, noon on the day of vaginal
plug was considered day 0.5 of gestation. Mice were killed by cervical dislocation; uteri were dissected and washed in PBS (10 mM phosphate, pH 7.4, 150 mM NaC1). Embryos were collected under a Wild M7 ste- reomicroscope.
Antibodies Rabbit polyclonal antibodies to synthetic peptide de-
rived from the COOH terminal sequence of a, integrin subunits (KKRVRPPQEEQERE) were generated as described previously (Defilippi e t al., 1992; Tarone et al., 1993). The antibodies were purified by affinity chromatography on the relevant peptide coupled to Sepharose as described (Hirsch et al., 1993). Antibodies to synthetic peptides from a3 a5 integrins were also used (Defilippi et al., 1992). Due to the highly con- served sequence of the COOH terminal region of integ- rin subunits these antibodies react across the animal species. The monoclonal antibody (MAb) 147 to human a, (Vogel et al., 1990), a generous gift of E. Ruoslahti, and the monoclonal B212 to human p3 (Defilippi et al., 1991) were used as a reference. The specificity of anti- bodies was tested by immunoprecipitation experiments from 35S methionine labelled endothelial cells, as de- scribed by Defilippi et al. (1991).
Western Blot Analysis E10.5 embryos were frozen in liquid nitrogen,
thawed to room temperature, and solubilized in Laem- mli's buffer (Laemmli, 1970). The extract was boiled 5 min, passed several times through a 25 gauge needle to fragment genomic DNA, and centrifuged 10,OOOg for 10 min to remove insoluble material. Soluble proteins were separated on a 10% polyacrylamide gel in the presence of SDS and subsequently blotted to a nitrocel- lulose membrane. a, was detected by incubation in PBS, 0.1 % gelatine containing 5 pg/ml a, antibody, followed by peroxidase conjugated secondary antibody and chemiluminescent substrate (ECL, Amersham, U.K.).
Histochemical Reactions Embryos and adult organs were processed according
to Bronner Fraser et al. (1992) and 10 pm thick cryo- sections were made at -25°C and collected on polyly- sine-subbed slides. Sections were then incubated 10 min in 50% acetone/50% chloroform; gelatine was re- moved by incubation in PBS at 37"C, and endogenous peroxidase were inhibited with 0.3% H,02. Cryosec- tions were incubated overnight a t 4°C with 5 pglml affinity purified integrin antibodies, washed and then incubated for 1 hr a t room temperature with peroxi- dase-conjugate secondary antibodies (the Binding Site, Birmingham, UK). Staining was obtained incubating in 0.04% diaminobenzidine, 0.003% H202, 30 mM Tris HCl, pH 7.6, for the required time. Antibodies to glial fibrillary acidic protein, to neurofilaments 200 kDa, and to desmin were purchased from Sigma (St. Louis, MO) and used a t the recommended dilution. Labelling
116 HIRSCH ET AL.
Fig. 6. Distribution of a, in the developing skeletal muscle. a: in E10.5 embryos a, antibodies label myotomal cells (m, arrowheads) and not the dermotomal cells (d). nt = neural tube. b: intervertebral premuscular masses at El 2.5. a, was concentrated at the extremities of the myotubes. c: labelling of myotendinous junctions of E14.5 embryos with desmin antibodies. d: The same area as (c) stained with a, antibodies showed a clear codistribution of the two antigens (arrowheads in c and d). e: trans-
versa1 section of the leg insertion at E15.5. Q, was expressed by muscle and was enriched where myotubes contact the developing bone. f: higher magnification of myotubes of E15.5 embryos stained with a, antibodies. Labelling appeared at the surface of the apical extremity of myotubes (arrows). Some staining was located on presumptive Z bands (arrow- heads). Bars = 50 pm.
a, INTEGRIN SUBUNIT IN MOUSE DEVELOPMENT 117
Fig 7 Distribution of a, and actin in differentiated primary rnyotube cultures Primary cells were isolated from the thighs of newborn mice and rnyoblasts were induced to differentiate into rnyotubes On day 7 of culturing, cells were fixed, perrneabilized, and processed for irnmunofluorescence Cultures were double- labelled for a, integrin (a, c, e) and acttn (a, d) contrasting the staining observed on fibroblasts (e, f ) Bars in b, d, f = 100 pn. 40 yrn, and 10 yrn, respectively
with different antibodies was made on adjacent sec- tions collected on different slides, in order to ease their comparison. Control staining was performed using pre-
immune or non-immune serum as primary antibodies and no significant immunoreactivity was detectable. Moreover staining was blocked by incubation of sec-
118 HIRSCH ET AL.
Fig. 8. Minor sites of a, expression. a: the limb bud of E10.5 embryo showed diffused a, expression enriched in the apical ectodermal ridge. b: mesonephric tubula of E15.5 embryo were stained with av anti- bodies. c: the smooth muscle layer of viscera at E l 5.5 does not express a,. d: antibodies directed against a3 integrin subunit showed expression of the antigen in the same areas as c, where ay is absent. aer = apical ectodermal ridge. Bars = 50 pm.
tions with a, antibodies and 5 pg of the peptide used for immunization (not shown). Sections were counter- stained with hematoxylin for a few seconds.
Cell Culture Primary cells were isolated from the thighs of the
hindleg of newborn Balbic mice. Briefly skin and bone were removed from the thighs, the tissue was weighed, rinsed in PBS, cut into pieces, and incubated with stir-
ring for 6-8 hr a t room temperature. Tissue (0.1 g) was incubated with 1 ml DMEM containing 5% BSA and 0.5 mg/ml collagenase P (Boehringer Mannheim 1213865, Mannheim, Germany). Following collage- nase digestion the cells were filtered through a double layered sterile lens paper (Kebo-Grave No. 1019), washed in DMEM, 10% horse serum and 1% chicken embryo extract (GIBCO 620-6460AE, Gibco, Grand Is- land, NY) and seeded onto collagen coated coverslips at

a, INTEGRIN SUBUNIT IN MOUSE DEVELOPMENT 119
a density of 3 x lo6 cells/ml in the above medium. Collagen coating was performed by coating ethanol washed and sterilised coverslips with 0.1% poly-L- lysine/H,O for 2 hr at room temperature, washing three times with H,O and coating for 2 hr at 37°C with 0.1 mg/ml of collagen type I (Vitrogen) in PBS.
Immunofluorescence Cultured cells grown on coverslips were washed in
PBS, fixed in 2% paraformaldehyde in PBS at room temperature for 10 min, permeabilized in 0.5% Triton X-100 in PBS for 20 min, and rinsed in PBS. Non- specific binding sites were blocked by a 1-hr incubation with 5% BSA in PBS. Fixed cells were incubated with 10 p,g/ml affinity purified polyclonal antibodies to a, peptide for 1 h r at room temperature. After washing with PBS, cells were incubated with biotinylated sec- ondary goat-anti-rabbit antibodies (Jackson Laborato- ries, Bar Harbor, ME) for 1 hr. Bound antibodies were visualized by a 1-hr incubation with Texas red conju- gated avidin D (Vector Laboratories, Burlingame, CAI. Actin was visualized using bodipy phallacidin (Molec- ular Probes, Eugene, OR).
ACKNOWLEDGMENTS We thank Prof. Guido Filogamo and Drs. Ferdinand0
Rossi and Antonio Bertolotto for their advice in the interpretation of the embryonic histological features, and Immacolata Carfora for the help in embryo collec- tion and manipulation. This work was supported by Italian Minister0 della Pubblica Istruzione (40%), by Progetto Finalizzato CNR “Ingegneria Genetica,” and by Telethon.
REFERENCES Albelda, S.M., and Buck, C.A. (1990) Integrins and other cell adhesion
molecules. FASEB J . 4:2868-2880. Bao, Z.Z., Lakonishok, M., Kaufman, S., and Horwitz, A.F. (1993)
a7pI integrin is a component of the myotendinous junction on skel- etal muscle. J . Cell Sci. 106579-590.
Bozyczko, D., Decker, C., Muschler, J., and Horwitz, A.F. (1989) In- tegrin on developing and adult skeletal muscle. Exp. Cell. Res. 183: 72-91.
Bronner-Fraser, M., Artinger, M., Muschler, J., and Horwitz A.F. (1992) Developmentally regulated expression of as integrin in avian embryos. Development. 115:197-211.
Buckingham, M. (1992) Making muscle in mammals. Trends Genet. 8144-149.
Bunch, T.A., and Brower, D.L. (1992) Drosophila PS2 integrin medi- ates RGD-dependent cell-matrix interactions. Development 116: 239-247.
Busk, M., Pytela, R., and Sheppard, D. (1992) Characterization of the integrin a,& as a fibronectin-binding protein. J . Biol. Chem. 267: 5790-5796.
Charo, I.F., Nannizzi, L., Smith, J.W., and Cheresh, D.A. (1990) The vitronectin receptor a& binds fibronectin and acts in concert with a& in promoting cellular attachment and spreading on fibronec- tin. J . Cell. Biol. 111:2795-2800.
Cheresh, D., Smith, J., Cooper, H., and Quaranta, V. (1989) A novel vitronectin receptor integrin (a,p,) is responsible for distinct adhe- sive urouerties of carcinoma cells. Cell 5759-69.
I .
Conforti, G., Zanetti, A., Pasquali-Ronchetti, I., Quaglino, D., Neyroz, P. Jr., and Dejana, E. (1990) Modulation of vitronectin receptor
- (1991) Early expression of the myogenic regulatory gene, myf5, in
binding by membrane lipid composition. J . Biol. Chem. 265:4011- 4019.
Corless, C.L., Mendoza, A., Collins, T., and Lawler, J . (1992) Colocal- ization of thrombospondin and syndecan during murine develop- ment. Dev. Dyn. 193:346-358.
Defilippi, P., Truffa, G., Stefanuto, G., Altruda F., Silengo, L., and Tarone, G. (1991) Tumor necrosis factor a and interferon gamma modulate the expression of the vitronectin receptor (integrin p3) in human endothelial cells. J . Biol. Chem. 266:7638-7645.
Defilippi, P., Silengo, L., and Tarone, G. (1992) asp1 integrin (laminin receptor) is down-regulated by tumor necrosis factor a and inter- leukin-lp in human endothelial cells. J. Biol. Chem. 267:18303- 18307.
Duband, J.-L., Belkin, A,, Syfrig, J., Thiery J.P., and Koteliansky, V.E. (1992) Expression of a, integrin, a laminin-collagen receptor, during myogenesis and neurogenesis in the avian embryo. Devel- opment 116585-600.
Dupouey, P., Benjelloun, S., and Gomes, D. (1985). Immunohisto- chemical demonstration of an organized cytoarchitecture of the ra- dial glia in the CNS of the embryonic mouse. Dev. Neurosci. 7:81- 93.
Fogerty F.J., Fessler, L.I., Bunch, T.A., Yarin, Y., Parker, C.G., Nel- son, R.E., Brower, D.L., Gullberg, D., and Fessler, J.H. (1994) Tig- grin, a novel Drosophila extracellular matrix protein that functions as a ligand for Drosophila aps2pps3 integrins. Development 120: 1747-1758.
Galileo, D.S., Majors, J., Horwitz, A.F., and Sanes, J.R. (1992) Retro- virally introduced antisense integrin RNA inhibits neuroblast mi- gration in vivo. Neuron 9:1117-1131.
George-Weinstein, M., Foster, R.F., Gerhart, J.V., and Kaufman, S.J. (1993) In vitro and in vivo expression of a7 integrin and desmin define the primary and secondary myogenic lineages. Dev. Biol. 156:209-229.
Gullberg, D., Fessler, L.I., and Fessler, J.H. (1994) Differentiation, extracellular matrix synthesis, and integrin assembly by Drosoph- ila embryo cells cultured on vitronectin and laminin substrates. Dev. Dyn. 199116-128.
Hirano, S., Ui, K., Miyake, T., Uemura, T., and Takeichi, M. (1991) Drosophila PS integrins recognize vertebrate vitronectin and func- tion as cell-substratum adhesion receptors in vitro. Development 113:1007-1016.
Hirsch, E., Balzac, F., Pastore, C., Tarone, G., Silengo, L., and Al- truda, F. (1993) The p l integrin distal promoter is developmentally regulated in transgenic mice. Cell Adhesion Commun. 1:203-212.
Hogervorst, F., Kuikman, I., Van Kessel, A.G., and Sonnenberg, A. (1991) Molecular cloning of the human a 6 integrin subunit. Alter- native splicing of a6 mRNA and chromosomal localization of the a6 and p4 genes. Eur. J . Biochem. 199:425-433.
Hynes, R.O. (1992) Integrins: versatility, modulation and signalling in cell adhesion. Cell 6911-25.
Krissansen, G.W., Elliot, M.J., Lucas, C.M., Stomski, F.C., Berndt, M.C., Cheresh, D.A., Lopez, A.F., and Burns, G.F. (1990) Identifi- cation of a novel integrin p subunit expressed on cultured mono- cytes (macrophages). J. Biol. Chem. 265823-830.
Laemmli, U. (1970) Cleavage of structural protein during the assem- bly of the head of bacteriophage T4. Nature 227:680-685.
Lakonishok, M., Muschler, J., and Horwitz, A.F. (1992) The a5bl integrin associates with a dystrophin-containing lattice during muscle development. Dev. Biol. 152:209-220.
MacKrell, A,, Blumberg, B., Haynes, S.R., and Fessler, J.H. (1988) The lethal myospheroid gene of drosophila encodes a membrane protein homologous to vertebrate integrin p subunits. Proc. Natl. Acad. Sci. U.S.A. 85:2633-2637.
Misson, J.P., Takahashi, T., and Caviness, V.S. J r . (1991) Ontogeny of radial and other astroglial cells in murine cerebral cortex. Glia 4:138-148.
Muschler, J.L., and Horwitz, A.F. (1991) Down regulation of the chicken aspl integrin fibronectin receptor during development. De- velopment 113:327-337.
Ott, M.-0.. Bober, E., Lyons, G., Arnold, H.H., and Buckingham, M.

120 HIRSCH ET AL.
precursor cells of skeletal muscle in the mouse embryo. Develop- ment 114:1097-1107.
Palmer, E.L., Ruegg, C., Ferrando, R., Pytela, R., and Sheppard, D. (1993) Sequence and tissue distribution of the integrin a9 subunit, a novel partner of that is widely distributed in epithelia and muscle. J . Cell Biol. 123:1289-1297.
Pasqualini, R., Bodorova, J . , Song, Y., and Hemler, M.E.. (1993) A study of the structure, function and distribution of p5 integrin using novel anti-p, monoclonal antibodies. J. Cell Sci. 105:lOl-111.
Rosen, G., Sanes, J., Lachance, R., Cunningham, J., Roman, J., and Dean D.C. (1992) Roles for the integrin VLA-4 and its counter re- ceptor VCAM-1 in myogenesis. Cell 69:1107-1119.
Sassoon, D., Lyons, G., Wright, W.E., Lin, V., Lassar, A,, Weintraub, H., and Buckingham, M. (1989) Expression of two myogenic regu- latory factors myogenin and MyoD during mouse embryogenesis. Nature 314:303-307.
Sheppard, A.M., Hamilton, S.K., and Pearlman, A.L. (1991) Changes in the distribution of extracellular matrix components accompany early morphogenetic events of mammalialn cortical development. J . Neurosci. 11:3928-3942.
Suzuki, S., Argraves, W.S., Arai, H., Languino, L.R., Pierschbacher, M.D., and Ruoslahti, E. (1987) Amino acid sequence of the vitronec- tin receptor a subunit and comparative expression of adhesion re- ceptor mRNAs. J . Biol. Chem. 262:14080-14085.
Tarone, G., Russo, M., Hirsch, E., Odorisio, T., Altruda, F., Silengo, L., and Siracusa, G. (1993) Expression of p l integrin complexes on the surface of unfertilized mouse oocyte. Development 117:1369-1375.
Tidball, J.G. (1992) Desmin at myotendinous junctions. Exp. Cell Res. 199:206-212.
Vogel, B., Tarone, G., Giancotti, F.G., Gailit, J., and Ruoslahti, E. (1990) A novel fibronectin receptor with an unexpected subunit composition (a$,). J . Biol. Chem. 265:5934-5937.
Volk, T., Fessler, L.I., and Fessler, J.H. (1990). A role for integrin in the formation of sarcomeric cytoarchitecture. Cell 63:525-536.
von der Mark, H.J., Durr, A., Sonnenberg, K., von der Mark, K., Deutzmann, R., and Goodman, S.L. (1991) Skeletal myoblasts uti- lize a novel pl-series integrin and not a6pl for binding to the E8 and T8 fragments of laminin. J. Biol. Chem. 26623593-23601.