unusual glycoconjugates in the oesophagus of a tilapine polyhybrid
TRANSCRIPT

Journal of Fish Biology (1998) 53, 39–48Article No. jb980681
Unusual glycoconjugates in the oesophagus of a tilapinepolyhybrid
P. S*§, D. A†, G. M† P. C‡*Facolta di Medicina Veterinaria, †Dip. Sc. Morfol. Biochim. Comp., Universita di
Camerino, Italy and ‡Istituto di Anatomia degli Animali Domestici, Facolta di MedicinaVeterinaria Universita di Perugia, Italy
(Received 14 October 1997, Accepted 24 February 1998)
The aim of this work is to elucidate the glycoconjugate composition of the secretory productsof the oesophageal mucous cells in a tilapine polyhybrid. Lectin histochemistry gave evidenceof the presence of â-galactose, á-N-acetylgalactosamine and sialic acid residues in the terminalposition. The majority of sialic acid belongs to short side chains; a few sialic acid residues areacetylated at the C7 and/or C8 and/or C9 level. The heterogeneity of the carbohydrate chainsmay mask potential receptor sites for micro-organisms and hamper the formation of multiplebonds. ? 1998 The Fisheries Society of the British Isles
Key words: Tilapia spp.; oesophagus; lectins; sialic acids.
INTRODUCTION
The oesophageal wall structure in teleosts is well documented. The data in theliterature show a wide morphological heterogeneity of the oesophageal wallamong different species (Kapoor et al., 1975; Reifel & Travill, 1977; Hirji, 1983;Martin & Blaber, 1984; Elbal & Agulleiro, 1986; Grau et al., 1992). Theultrastructural features of the oesophagus of the Tilapia polyhybrid wereinvestigated by Gargiulo et al. (1996) who showed that the oesophageal liningconsists of a stratified, irregularly folded epithelium showing cuboidal basal cells,columnar cells in the intermediate layer and an outer layer of more flattenedcells. The dominant feature of epithelium throughout the oesophagus is theabundance of mucus-secreting cells.
The aim of this work was to characterize the tilapine oesophagus on the basisof glycoconjugate composition of secretion from mucous cells.
MATERIALS AND METHODS
TISSUE PREPARATIONSixteen adult Tilapia spp. (nilotica#mossambica#zilii) of both sexes, weighing
400–600 g, in fasting and feeding conditions, obtained from La Casella Acquacoltura(Piacenza, Italy) and Cooperativa Sorgeva (Ferrara, Italy) were used. The animals werekilled by decapitation and the oesophagus was removed quickly and fixed in Carnoy’sfluid for 24 h, post-fixed in 2% calcium acetate–4% paraformaldehyde (1 : 1) for 3 h
§Author to whom correspondence should be addressed: Facolta di Medicina Veterinaria, ViaCirconvallazione 93, 62024 Matelica (MC), Italy. Tel.: (39) 737 789 1; fax: (39) 737 789 321.
39
0022–1112/98/070039+10 $30.00/0 ? 1998 The Fisheries Society of the British Isles

40 . .
(Menghi, 1984; Spicer & Schulte, 1988), dehydrated and embedded in paraffin wax.Serial sections (5 ìm) were mounted on albumin coated slides.
CONVENTIONAL HISTOCHEMICAL STAININGSSamples were treated for carbohydrate conventional histochemistry with periodic acid
Schiff (PAS, to detect vicinal hydroxyls), Alcian blue-periodic acid Schiff (AB-PAS, todemonstrate acidic groups and vicinal hydroxyls), AB pH 2·5 (to reveal acidic groups)and AB pH 1 (to discriminate sulphate groups) (Spicer et al., 1967, Pearse, 1968).
Mild and strong periodate oxidation with periodic acid (PO) were performed byimmersing the specimens in 1 m or 44 m periodic acid solution respectively for 15 minat room temperature to reveal the presence of terminal sialic acid residues with O-acetylsubstituents (Reid et al., 1978; Schauer, 1978). Acyl groups were removed by immersingspecimens in 0·5% potassium hydroxide in 70% ethanol for 30 min at room temperature(Reid et al., 1978).
LECTIN HISTOCHEMISTRYLectin staining was performed as described by Menghi (1984). In brief, after inhibition
of endogenous peroxidase activity by 0·3% H2O2/methanol for 30 min, tissue sectionswere immersed in lectin horseradish peroxidase (HRP) conjugates (0·02–0·2 mg ml"1) in0·05 phosphate buffered saline (PBS), pH 7·2 for 30 min at room temperature. Afterrinsing in PBS and 0·5 Tris/HCl buffer, pH 7·4, sections were treated withdiaminobenzidine-H2O2 medium (Graham & Karnovsky, 1966) for 10 min at roomtemperature before dehydration and mounting in Permount. The following lectins wereused: PNA from Arachis hypogaea [terminal â-D-galactose(1-3)-N-acetylgalactosamine],DBA from Dolichos biflorus (terminal á-N-acetylgalactosamine), RCA I from Ricinuscommunis and ECA from Erytrina cristagalli [terminal â-D-galactose(1-4)-N-acetylglucosamine], GSA IB4 from Griffonia simplicifolia (terminal á-D-galactose), WGAfrom Triticum vulgaris (terminal and internal â-N-acetylglucosamine±sialic acid), GSAII from Griffonia simplicifolia (terminal N-acetylglucosamine), Con A from Canavaliaensiformis (terminal and internal á-D-mannose>á-D-glucose), UEA I from Ulexeuropaeus and LTA from Lotus tetragonolobus (terminal á--fucose).
Before lectin histochemical staining, some sections were incubated, at 37) C for 16 h, in0·86 U mg"1 protein of sialidase (Type V, from Clostridium perfringens) dissolved in0·1 sodium acetate buffer, pH 5·5, containing 10 m CaCl2. Other sections wereincubated, at 37) C for 14 h, with á-fucosidase from bovine epididymis (2·3 U mg"1
protein) dissolved in 0·1 sodium citrate buffer, pH 6·0, containing 25 m EDTA(Menghi et al., 1989). Some sections were incubated at 37) C for 3 h in 500 U mg"1
protein of â-galactosidase (from Escherichia coli) dissolved in 0·05 sodium citratebuffer, pH 3·8 containing 25 m EDTA. Lectins and enzymes were purchased fromSigma Chemical Co. (St Louis, MO, U.S.A.).
The respective peroxidase-conjugated lectins were omitted and/or their hapten sugars(0·2–0·4 ) were added in control sections. As controls for enzyme digestion, sectionswere incubated with the respective enzyme-free buffers under the same experimentalconditions, or with the lectins specific for sugar detached by enzyme pretreatment.
The following histochemical procedures were performed as sequential treatments:AB pH 2·5Sial/AB pH 2·5KOH/Sial/AB pH 2·5PNA1 m–44 m PO/PNAâ-galactosidase/PNASial/PNAâ-galactosidase/Sial/PNAKOH/Sial/PNAâ-galactosidase/KOH/Sial/PNAâ-galactosidase/1 m–44 m PO/Sial/PNAâ-galactosidase/1 m–44 m PO/KOH/Sial/PNAKOH/1 m–44 m PO/Sial/PNA.

41
RESULTS
No differences were revealed in the reactivity of mucous cells between fastingand feeding subjects.
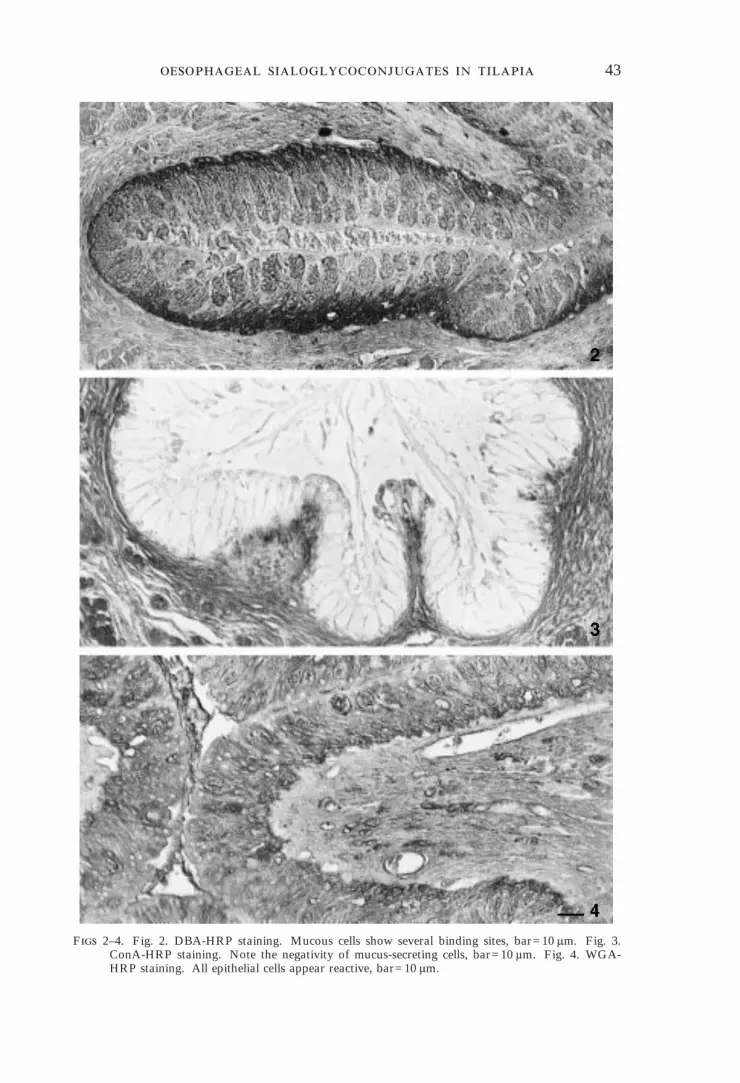
Mucous cells were PAS and AB/PAS positive [Fig. 1(a), (b)], negative to AB atpH 1 [Fig. 1(c)] but appeared strongly positive after the application of AB atpH 2·5 [Fig. 1(d)]. This last positive reaction was reduced greatly after sialidasedigestion [Fig. 1(e)] and abolished completely after the application of theKOH/Sialidase treatment [Fig. 1(f)]. The mucous cells displayed a large numberof binding sites for lectin DBA (Fig. 2) that appeared unaffected by sialidasedigestion. The reaction with the lectin ConA was negative (Fig. 3), but anintense staining was evident after WGA application (Fig. 4). Mucous cellreactivity to PNA was of variable intensity ranging from negative to moderatelypositive [Fig. 5(a)]. Reactivity was enhanced by sialidase digestion [Fig. 5(b)].When preceded by mild periodic oxidation the sequence sialidase/PNA gave onlya weak staining [Fig. 5(d)] that disappeared completely when the strong periodicoxidation was applied. Modifications in reactivity to PNA following theperiodic acid, KOH and sialidase treatments applied in sequence are summarizedin Table I. Some of the treatments reported included incubation withâ-galactosidase to abolish the background staining to PNA [Fig. 5(c)]. All otherlectins failed to bind to our sections; fucosidase digestion did not modify lectinreactivities.
No staining was revealed in sections exposed to unconjugated lectins and toincubation with hapten sugars in the conjugated lectin solutions. The immersionof sections in enzyme-free buffer solution resulted in unmodified binding.
DISCUSSION
Conventional histochemical procedures seem to suggest a codistribution ofneutral and acidic glycoconjugates with carboxylic groups attributable to sialicacid which may be acetylated occasionally at C4. Lectin staining revealed thepresence of á-N-acetylglucosamine in an internal location along the glucidicchains and of terminal á-N-acetylgalactosamine. These data, together with thelack of mannosal moieties, suggest that all the glycoconjugates are linkedO-glycosidically. The positive reaction to PNA staining suggests the presence ofmany terminal residues of â-galactose which is the unique sub-terminal acceptorfor sialic acid.
The mild and strong periodate oxidation in association with sialidase diges-tion, preceded or not by saponification (KOH treatment), revealed that almostall of the sialic acid residues linked to â-galactose were not acetylated. Only aminor quantity of the sialic acid residues seems to be acetylated at C7 and/or C8
and/or C9; in this last case the sialic acid is á-2-6 linked to the galactose(Montreuil, 1980) on the basis of the abolition of induced Sial/PNA staining with44 m PO.
These results induce us to attribute some hypothetical structural models to theglycoconjugates secreted by the oesophageal mucous cells (Table II). Thelocation of sialic acid residues in the glucidic chains is very interesting because oftheir involvement in the formation of short lateral side chains. The strong
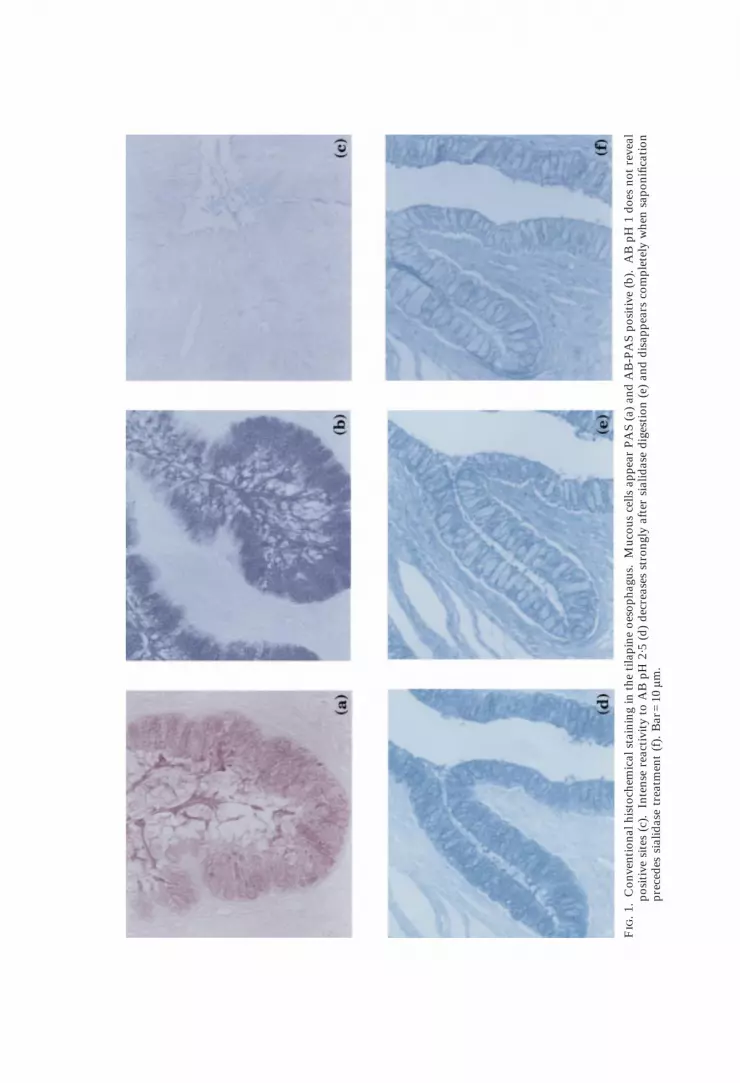
F
.1.
Con
vent
iona
lhis
toch
emic
alst
aini
ngin
the
tila
pine
oeso
phag
us.
Muc
ous
cells
appe
arP
AS
(a)
and
AB
-PA
Spo
siti
ve(b
).A
BpH
1do
esno
tre
veal
posi
tive
site
s(c
).In
tens
ere
acti
vity
toA
BpH
2·5
(d)
decr
ease
sst
rong
lyaf
ter
sial
idas
edi
gest
ion
(e)
and
disa
ppea
rsco
mpl
etel
yw
hen
sapo
nific
atio
npr
eced
essi
alid
ase
trea
tmen
t(f
).B
ar=
10ìm
.
43
F 2–4. Fig. 2. DBA-HRP staining. Mucous cells show several binding sites, bar=10 ìm. Fig. 3.ConA-HRP staining. Note the negativity of mucus-secreting cells, bar=10 ìm. Fig. 4. WGA-HRP staining. All epithelial cells appear reactive, bar=10 ìm.

44 . .

45
F. 5. PNA-HRP staining. The mucous cells located in the basal portion of the oesophageal plicae arenegative (asterisks), whereas those in the apical zone show a discrete reactivity (arrows) (a).Sialidase pretreatment enhanced strongly mucous cell reactivity in both apical and basal portionsof the plicae (b). Pretreatment with â-galactosidase removed PNA reactivity (inset) and made thePNA reactivity imparted by sialidase digestion evident (c); 1 m periodate oxidation (d) decreasedPNA reactivity strongly after sialidase, indicating the presence of a large amount of periodate labilesialic acid; only few cells remained moderately reactive (arrows) indicating the occurrence of sialicacid with acyl substitutes on C7 and/or C8 and/or C9. Bar=10 ìm.
T I. Histochemical treatments and PNAlabelling in the tilapine oesophageal mucous
cells
PNA "/+1 m PO/PNA "/+
44 m PO/PNA "â-gal/PNA "Sial/PNA +/+ +â-gal/Sial/PNA &/+KOH/Sial/PNA +/+ +â-gal/1 m PO/Sial/PNA "/&â-gal/44 m PO/Sial/PNA "KOH/1 m PO/Sial/PNA "/+KOH/44 m PO/Sial/PNA "
Results are given in arbitrary units as follows: ",negative; &, weak; +, moderate; + +, strong staining.
positive reaction to the AB pH 2·5 staining and its marked reduction aftersialidase digestion do not agree with the low amount of sialic acid linked to theâ-galactose, but can be explained well by the presence of lateral chains. Thenegative staining of AB preceded by sialidase digestion and saponificationsuggests that only a few residues of sialic acid are acetylated at the C4 position,and that perhaps they make up the lateral chains. This hypothesis seems to besupported by the observation that sialic acid residues acetylated at the C4 are notlinked to â-galactose as demonstrated by the unmodified reactivity of Sial/PNApreceded by saponification. The large amount of mucus secreted does not seemto be explicable simply as a lubricant (Reifel & Travill, 1977; Hirji, 1983). It isimportant to note that salivary glands are lacking in fish and so the oesophagealmucous cells could carry the same functions of mammalian saliva in protectingthe mucosa of the entire alimentary canal. This hypothesis could, in our opinion,be maintained even if the sialoglycoconjugates from the tilapia oesophagusdisplay an organization which is unusual in mammals (Klein et al., 1992; Wuet al., 1994). The presence of sialic acid residues acetylated at C4 prevents virusesfrom recognizing their receptor determinants and preserves the mucosae fromthe attack of sialidase producing bacteria (Zimmer et al., 1992; Hanaoka et al.,1989). Finally, the variability and the location of the sialic acid residues in theglucidic chains could hamper the bindings of virus and bacteria to the receptorsites in the mucosal surface (Nieuw-Amerongen et al., 1995).

T
II.
Stru
ctur
alm
odel
spr
opos
edfo
rth
esi
alog
lyco
conj
ugat
espr
oduc
edby
the
oeso
phag
eal
muc
ous
cells
inT
ilapi
asp
p.
Per
iphe
ral
bran
ches
Rep
eati
ngsa
ccha
ride
stru
ctur
eC
ore
stru
ctur
e
Sial
icac
id(2
-3)â
-gal
acto
seN
-ace
tylg
alac
tosa
min
eâ-
gala
ctos
e(1-
3)-N
-ace
tylg
alac
tosa
min
e|
sial
icac
id
(1-3
)-ga
lact
ose-
(1-4
)-N
-ace
tylg
luco
sam
ine
Ser
(1-3
)-N
-ace
tylg
alac
tosa
min
e/ \|
Thr
sial
icac
id

47
This study was supported by a grant (60%) from M.U.R.S.T., Italy. We thank P.Melotti, A. Roncarati, La Casella Acquacoltura and Cooperativa Sorgeva, Italy for theconstructive collaboration.
References
Elbal, M. T. & Agulleiro, B. (1986). A histochemical and ultrastructural study of the gutof Sparus auratus (Teleostei). Journal of Submicroscopic Cytology 18, 335–347.
Gargiulo, A. M., Dall’Aglio, C., Tsoku, Z., Ceccarelli, P. & Pedini, V. (1996).Morphology and histology of the oesophagus in a warmwater tilapiine fish(Teleostei). Journal of Applied Ichthyology 12, 121–124.
Graham, R. C., Jr & Karnovsky, M. J. (1966). The early stages of absorption of injectedhorseradish peroxidase in the proximal tubules of mouse kidney: ultrastructuralcytochemistry by a new technique. Journal of Histochemistry and Cytochemistry14, 291–302.
Grau, A., Crespo, S., Sarasquete, M. C. & Gonzales de Canales, M. L. (1992). Thedigestive tract of the amberjack Seriola dumerili Risso: a light and scanningelectron microscope study. Journal of Fish Biology 41, 287–303.
Hanaoka, K., Pritchett, T. J., Tokasaki, S., Kochibe, N., Sabesan, S., Paulson, J. C. &Kobata, A. (1989). 4-O-acetyl-N-acetylneuraminic acid in the N-linked carbohy-drate structures of equine and guinea pig á2-macroglobulins, potent inhibitors ofinfluenza virus infection. Journal of Biological Chemistry 264, 9842–9849.
Hirji, K. N. (1983). Observations on the histology and histochemistry of the oesophagusof the perch Perca fluviatilis L. Journal of Fish Biology 22, 145–152.
Kapoor, B. G., Smith, H. & Verighina, I. A. (1975). The alimentary canal and digestionin teleosts. Advances in Marine Biology 13, 109–139.
Klein, A., Carnoy, C., Wieruszeski, J. M., Strecker, G., Strang, A. M., van Halblek, H.,Roussel, P. & Lamblin, G. (1992). The broad diversity of neutral and sialylatedoligosaccharides derived from human salivary mucins. Biochemistry 31, 6152–6165.
Martin, T. J. & Blaber, S. J. M. (1984). Morphology and histology of the alimentarytracts of Ambassidae (Cuvier) (Teleostei) in relation to feeding. Journal ofMorphology 182, 295–305.
Menghi, G. (1984). Reactivity of peroxidase-labeled lectins in rabbit submandibular andsublingual glands. Acta Histochemica 75, 27–35.
Menghi, G., Accili, D., Bondi, A. M. & Gabrielli, M. G. (1989). Enzymatic degradationand quantitative lectin labeling for characterizing glycoconjugates which act aslectin acceptors in cat SM gland. Histochemistry 90, 331–338.
Montreuil, J. (1980). Primary structure of glycoprotein-glycans: basis for the molecularbiology of glycoproteins. Advances in Carbohydrate Chemistry and Biochemistry37, 157–213.
Nieuw-Amerongen, A. V., Bolscher, J. G. M. & Veerman, E. C. I. (1995). Salivarymucins: protective functions in relation to their diversity. Glycobiology 5, 733–740.
Pearse, A. G. E. (1968). Histochemistry Theoretical and Applied. 3rd edn. Vol. I.London: Churchill.
Reid, P. F., Culling, C. F. & Dunn, W. L. (1978). A histochemical method for theidentification of 9-O-acyl sialic acids. An investigation of bovine submaxillarygland and intestinal mucins. Journal of Histochemistry and Cytochemistry 26,187–192.
Reifel, C. W. & Travill, A. A. (1977). Structure and carbohydrate histochemistry of theoesophagus in ten teleostean species. Journal of Morphology 152, 303–314.
Schauer, R. (1978). Characterization of sialic acids. In Methods in Enzymology, Vol. 50(Ginzburg, V., ed.), pp. 65–89. New York: Academic Press.
Spicer, S. S. & Schulte, B. A. (1988). Detection and differentiation of glycoconjugates invarious cell types by lectin histochemistry. Basic and Applied Histochemistry 32,307–320.

48 . .
Spicer, S. S., Horn, R. G. & Leppi, T. J. (1967). Histochemistry of connective tissuemucopolysaccharides. In The Connective Tissue (Wagner, B. M. & Smith, D. E.,eds), pp. 251–303. Baltimore, MD: Williams & Wilkins.
Wu, A. M., Csako, G. & Herp, A. (1994). Structure, biosynthesis, and function ofsalivary mucins. Molecular and Cellular Biochemistry 137, 39–55.
Zimmer, G., Reuter, G. & Schauer, R. (1992). Use of influenza c-virus for detectionof 9-O-acetylated sialic acids on immobilized conjugates by esterase activity.European Journal of Biochemistry 204, 209–215.