glycoconjugates for the recognition of bacillus spores
TRANSCRIPT

Seediscussions,stats,andauthorprofilesforthispublicationat:https://www.researchgate.net/publication/8144558
GlycoconjugatesfortherecognitionofBacillusspores
ARTICLEinCARBOHYDRATERESEARCH·JANUARY2005
ImpactFactor:1.93·DOI:10.1016/j.carres.2004.10.008·Source:PubMed
CITATIONS
19
READS
19
4AUTHORS,INCLUDING:
KalleLevon
PolytechnicInstituteofNewYorkUniversity
128PUBLICATIONS2,143CITATIONS
SEEPROFILE
Availablefrom:KalleLevon
Retrievedon:10February2016

Revie
w C
opy
1
GLYCOCONJUGATES FOR RECOGNITION OF BACILLUS SPORES
Olga Tarasenko, Sharmin Islam, David Paquiot, and Kalle Levon*
The Othmer Department of Chemical and Biological Sciences and Engineering,
Polytechnic University, Brooklyn, NY, USA.
* Corresponding author:
Kalle Levon, Ph.D., Professor
The Othmer Department of Chemical and
Biological Sciences and Engineering,
Polytechnic University
Six MetroTech Center
Brooklyn, NY, 11201
USA
Tel.: +1 718-260-3339
Fax: +1 718-260-3125
E-mail: [email protected]
1 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
2
Abstract
Carbohydrates act as ligands in many biological processes, including the folding and
secretion of proteins, cell-cell recognition, adhesion, and sporulation in the Bacillus genus. To
elucidate the mechanism of bacterial spore recognition, fluorescent labeled disaccharide
glycoconjugates as a model system have been applied to evaluate binding to bacterial spores,
thus addressing key issues in the cell surface carbohydrate recognition field. Our study has
shown that specific recognition of bacterial spores is based on interactions between units of the
disaccharide glycoconjugates that act as ligands and monosaccharide units expressed on the
exterior of bacterial spores exterior. Using fluorophore assisted carbohydrate electrophoresis
(FACE) the carbohydrates that comprised the receptor recognition sites on the spores’ exterior
were enumerated. The findings have an impact on how carbohydrates function as recognition
signals in nature, as well as offering a basis for sensor development or inhibition of the bacterial
spores.
Keywords: B. cereus, B. thuringiensis, B. pumilus, B. subtilis spores, glycoconjugate, FACE,
ELISA.
2 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
3
1. Introduction
The problems associated with the Bacilli genus are as diverse as the environments where
they are found. The bacterial spore is the cause of problems such as food spoilages and food-
borne illnesses 1. Spores can withstand a wide range of assaults including heat, UV radiation,
oxidizing agents, desiccation, and physical damage 2-6. This makes them suitable for
incorporation into bioweapons and for concealment in terrorist weapons 7.
Given the severity of the threat posed by biological weapons, detection of potentially
harmful pathogens needs to be rapid and extremely sensitive since the presence of a single
pathogen can lead to infection. Furthermore, very selective detection methods are required
because pathogenic bacteria exist alongside non-pathogenic organisms in their environment.
There are several detection systems currently on the market: (i) Anthrax Bio-Threat Alert
(BTA) test strips (Tetracore, Gaithersburg, MD), (ii) BioWarfare Agent Detection Devices
(BADD) (Osborne Scientific, Lakeside, AZ), and (iii) Anthrax (spore) SMART II (New
Horizons Diagnostics, Columbia, MD) 8; immunomagnetic assay system 9-10; PCR-based assays 11-14 or traditional phenotyping of cultured bacteria 15-17. These systems are complex, expensive,
time consuming, and require highly qualified technical staffs. Other less complex and more
portable systems are based on antibody binding to surface antigens 18-19. Unfortunately these
simpler systems are flawed. Current antibody-based detectors suffer from a short shelf life, lack
of accuracy, and inadequate sensitivity. These deficiencies results in an unacceptably high level
of false-positive and false-negative responses, according to federal government trials 20 and
independent tests 8.
This lack of accuracy and sensitivity in the antibody-based detectors is compounded by
the presence of other spore-forming bacteria in the environment. Particularly problematic are
spores of the B. cereus group, including both the human pathogen B. cereus and the insect
pathogen B. thuringiensis, species which, based on genome sequence comparisons, are the most
similar to B. anthracis 21. Due to the limitations previously cited, all currently available systems
for detecting B. anthracis are inadequate for frontline use by first response units: firefighters,
police officers, soldiers, hospital personnel, and hazardous materials teams [HAZMAT] 22-23.
There is an urgent need for a quick and accurate biosensor, especially one specific to B.
anthracis. While working toward that goal, it was discovered that short peptides ligands bind
3 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
4
selectively to spores of B. anthracis 24. The Asn-His-Phe-Leu-Pro peptide has been identified
that selectively binds to spores of closely related species of B. anthracis 25. We recall that B.
cereus, genetically almost indistinguishable from B. anthracis 24. Our group has focused on the
synthesis and development of enhanced ligands for selective recognition of closely related
species of the B. cereus group to be used for bacterial spore recognition. Enhanced ligands,
which include peptides, polypeptides, dendrimers 26-28, lectins, antibodies 29-30, liposomes 31, 32,
and glycoconjugates 33-35, have been designed and have proved to be effective tools for selective
recognition of bacterial spores.
In our previous publications, we have shown that macromolecular conjugates allow for
the simultaneous binding of multiple ligands to biological entities such as spores 26, 29, 31-33, 35.
This binding was confirmed by fluorescence-based assays and the results clearly showed that the
affinity of the biomolecules was enhanced to a great extent by the polyvalent effect 36-40. These
polyvalent interactions can be collectively stronger than the corresponding monovalent ligands
as previously reported 36-40. The main focus of our work has been an electrochemical sensor that
uses potentiometry for biorecognition of bacterial spores 42, 43. This approach has allowed the
identification of specific ionic interactions between the spore’s surface 42, 43 and selected ligands 26-28, 43 and enables quick selective recognition of spores of the B. cereus group 41.
Until recently, the explanation of cell recognition has focused mostly on proteins while
marginalizing the role played by carbohydrates. Although carbohydrates are ubiquitously present
both in prokaryotic and eukaryotic cells, an appreciation of their diverse biochemical roles has
began only recently 44, 45. The mammalian and bacterial cellular surfaces are littered with
complex carbohydrate structures such as glycoproteins, glycolipids, glycosaminoglycans and
proteoglycans.
In all biological systems, glycoconjugates are involved with the recognition and signaling
processes intrinsic to biochemical functions in both healthy and diseased cells. They play a
central role in cell-to-cell adhesion and in subsequent recognition, receptor activation, and in
growth and in the maturation of living organisms 46-49. The host receptors that play a role in
microbial recognition are in fact glycoconjugates 49, and the various antigenic structures
identified by the host receptors, are also constituted in part by glycoconjugates 50.
The difficulty in understanding the relationship between structure and function in
glycoconjugates is compounded not only by the diversity of carbohydrate structures, and also by
4 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
5
the number of different cellular environments in which they can be found. Furthermore,
glycoconjugates have been found to often exhibit relatively low binding affinities for their
binding partners 37. It is therefore is difficulty to locate the specific function in such a broad
range of structures.
The carbohydrate’s role in recognition has only been studied in the case of endogenous
lectins: proteins that form stable complexes upon binding carbohydrates 51-53. Carbohydrate
groups can assist in protein folding or improve its stability 54-56. It is now known that N-
glycosylation of an Fc fragment of immunoglobulin influences the conformation of the Fc
peptide backbone 54. Different glycoforms stabilize different conformations and can exhibit
significantly different functional behaviors in vivo 54-56. Biochemically, carbohydrates tend to
further the binding initially begun by protein structures. For example, in the case of
immunoglobulin binding, the protein structures form an initial complex with the antigen 54-56.
However, it is the carbohydrates present that allow binding into a tighter complex. Given the
numerous binding sites on a carbohydrate (a result of the many sites available for chain
elongation and their structural flexibility) it is not surprising that structural heterogeneity and
conformational multiplicity are characteristic features of glycoproteins 55. The complex
geometries of the immunoglobulin domains and of the oligosaccharides mean that even relatively
small changes in glycan structure can leads to functionally significant changes in protein
structure 56.
A model system was needed in order to better study the specifics of carbohydrate-
carbohydrate interactions. This model system needed to have ligands and receptors with
corresponding glycoconjugate counterparts that illustrate carbohydrate-carbohydrate interaction.
B. cereus, B. anthracis, B. thuringiensis species, along with B. mycoides, comprise the
phylogenetically similar B. cereus group 21. The different Bacilli are however, quite distinct in
terms of chemical composition and morphology 57, 58. For this study, the spore’s specific
composition and morphology dictate its use as a model system.
A spore’s morphological structure typically includes the following components:
appendages (hair- or pilus-like filaments located on the outer spore surface), an exosporium, an
outer coat, an inner coat, a cortex, and a core. For each species of spore, the morphology can be
slightly different. It has been reported that B. cereus and B. thuringiensis spores possess
appendages 59, originating from an exterior basal membrane of the exosporium 59-61.
5 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
6
Appendages, though present on a variety of Bacillus spores, have not been observed in B. subtilis 62, 63.
Biochemical analysis of appendages and of the exosporium of B. cereus spores has been
reported 57, 58. Since lacking appendages 62, 63, the exosporium of B. subtilis has been found to be
composed of mostly proteins with minor amounts of carbohydrates and lipids 64, 65. Several major
proteins in the exosporium of B. anthracis Sterne strain have been identified 18, 39. A collagen-
like glycoprotein BclA 39 was recently identified that appears to be a structural component of the
exosporium hair-like nap 18. Two other unique proteins, BxpA and BxpB, were also identified18.
In addition, BxpB appears to be glycosylated or associated with glycosylated material 18.
The exosporium and appendages might in fact be the regions where chemical recognition
occurs. Considerable efforts have been directed toward understanding the chemical recognition
process in spores and toward recreating it in-vitro. Biomolecules such as polysaccharides,
glycoproteins or lipoproteins embedded in the exosporium or the appendages could, in fact, play
a role in the recognition process.
Glycoconjugates have been extensively used with the purpose of studying carbohydrate-
binding sites in histochemical and cytochemical experiments 37, 66-72. Their ability to bind
carbohydrate epitopes was established for a number of important cell surface proteins, such as
clusters of differentiation and adhesion factors 37, 66- 72. Furthermore, certain cells are able to
selectively bind to mono- or oligosaccharides present on the glycoconjugate probes 73. Attempts
have been made to develop an effective system to mimic spore recognition 24-35. Receptor-
specific sites of Bacillus species have not yet been characterized, but identification of specific
sites is indispensable for understanding the recognition events.
For our studies, the glycoconjugate-polyacrylamide-fluorescein (glycoconjugate PAA-
flu) polymers proved to be the most convenient synthetic ligands because they could be
immobilized on a solid-phase 74. The glycoconjugate-PAA-flu probe binds non-specifically with
cellular components 67-72, 74, 75. Both the fluorescent label and the polymer backbone affect this
interaction minimally 45, 69, 70. PAA-flu has certain properties that make it advantageous as the
polymer backbone of this glycoconjugate probe. It is hydrophilic and shows low binding affinity
and flexibility. In addition, its structure allows for the attachment of shorter ligands. The function
of the fluorescent label is to be an efficient marker for different techniques: spectroscopic
6 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
7
measurements, fluorescence and confocal microscopy, microspectroscopy on cells and tissue
sections 67-69, 74, 75.
The aim of the present study is two-fold: first, to elucidate the mechanism of Bacillus
spore recognition based on the interaction between the glycoconjugate-PAA-flu and spores as a
model, and second, to identify receptors expressed on the spore surface that are involved in this
interaction.
2. Results and Discussions
Often, recognition can easily be accomplished using surface-immobilized molecules that
interact with other species such as analytes in solution. Specific interaction can lead to the
binding of analytes to the immobilized ligands. The function of an assay is to detect binding
events, to convert them to registration signals, and to extract information. In pursuit of the
recognition of bacterial spores (analyte), we have developed (i) a glycoconjugate assay using the
polystyrene microplates, (ii) a novel sensing concept based on a synthetic ligand like
glycoconjugates, and (iii) we introduce synthetic ligands into ELISA as described in the
Experimental section. Binding events were converted into registration signals to produce binding
curves based on the average signal intensity measured at optical density (OD) 405 nm.
In our system, fluoresceinated glycoconjugates served as the synthetic ligands of our
hypothetical system to recognize and potentially capture the spores of B. cereus, B. subtilis, B.
pumilus and finally B. thuringiensis. The glycoconjugate-PAA-flu group is comprised of a
soluble PAA backbone of approximately 30 kDA with various carbohydrate side groups and a
fluorescent molecule. All the ten glycoconjugates used: Fucα1-4 GlcNacβ–PAA-flu (1), Fucα1-3
GlcNacβ–PAA-flu (2), Galβ1-3 GalNacβ–PAA-flu (3),Galβ1-3 GalNacα –PAA-flu (4), Galβ1-3
Galβ–PAA-flu (5), Galβ1-4 Glcβ–PAA-flu (6), Galα1-3 GalNacα–PAA-flu (7), GalNacβ1-3
GalNacα–PAA-flu (8), GalNacβ1-6 GalNac-PAA-flu (9), and GlcNacβ1-4 GlcNacβ–PAA-flu
(10) have the same molecular weight. In the microtiter plate we utilized each well and coated it
with a specific glycoconjugate of the ten. After coating each well, glycoconjugates served as the
synthetic ligands of our hypothetical system to mimic the spore recognition used to capture
spores. Next, the spores were introduced to bind with glycoconjugates. An anti-mouse (IgM
+IgG) horseradish peroxidase (HRP) labeled conjugate was added, leading to the creation of the
spore-glycoconjugate complex formation. Concerning the complex formed in the wells, the more
7 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
8
enzyme present, the more the HRP-labeled conjugate attached itself to the ligand-target complex.
Therefore, the amount of HRP-labeled conjugate present is determined by the amount of target
available. Finally, because glycoconjugates bind to an antigen (spore), the more target antigen
that is accessible, the more the ligand-target complex will be retained. The measure of OD values
therefore, reflects the amount of ligand-target complex initially present. The formation of this
multivalent complex was a critical step in establishing the high avidity binding. The overall
strength of binding ligands to spores (antigens) is called its avidity 76, 37-40 and was determined by
finding the magnitude of the optical density of the complex in each well. The OD values are a
measure of how tightly the synthetic ligands bind spores. A greater OD value indicates both a
stronger overall binding and a greater number of spores attached to the studied glycoconjugates.
OD analysis was performed using a SpectraMax® Plus384 spectrophotometer.
We evaluated glycoconjugates as a tool to discriminate related Bacilli species including
B. cereus and B. subtilis (Graph 1), B. thuringiensis and B. pumilus (Graph 3); B. cereus and B.
thuringiensis (Graph 5); B. subtilis and B. pumilus (Graph 7); B. subtilis and B. thuringiensis
(Graph 9); B. cereus and B. pumilus (Graph 11). Graphs 2, 4, 6, 8, and 10 present the influence
of the concentration of the ligands during spore recognition. The Y-axis represents OD values for
different spotted glycoconjugate ligands (X-axis) obtained at a wavelength of 405nm. Two sets
of experiments were carried out for each of the 10 sets of glycoconjugates. The results of both
experiments were nearly identical.
Graph 1. Patterns of B. cereus and B. subtilis
spore recognition using glycoconjugates. The
glycoconjugates used are: Fucα1-4 GlcNacβ–
PAA-flu (1), Fucα1-3 GlcNacβ–PAA-flu (2),
Galβ1-3 GalNacβ–PAA-flu (3),Galβ1-3
GalNacα–PAA-flu (4), Galβ1-3Galβ–PAA-
flu (5), Galβ1-4 Glcβ–PAA-flu (6), Galα1-3
GalNacα–PAA-flu (7), GalNacβ1-3 GalNacα–PAA-flu (8), GalNacβ1-6 GalNac–PAA-flu (9),
and GlcNacβ1-4 GlcNacβ–PAA-flu (10). The Y-axis represents the OD binding values for each
glycoconjugate ligands (X-axis) obtained at a wavelength of 405nm (here and thereafter).
8 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
9
Graph 1 displays the signal intensity for each glycoconjugate-spore binding event of B.
cereus and B. subtilis spore recognition. The average signal intensity value for each
glycoconjugate was plotted to illustrate the glycoconjugates that bound more strongly to a
particular spore. It can be clearly seen that i) on the whole, the glycoconjugates utilized in this
experiment were binding to B. cereus with higher OD value (Graph 1); ii) glycoconjugates
Galβ1-4 Glcβ–PAA-flu (6), and Galα1-3 GalNacα–PAA-flu (7), have a marked affinity for B.
cereus; iii) all glycoconjugates bound to B. subtilis with extremely low OD values compare with
B. cereus spores.
In order to examine the concentration effects of the ligands during spore recognition,
solutions of the glycoconjugates used were prepared according to the manufacturer’s protocol.
Than solutions were diluted ×10, ×100, ×1000 and used in recognition of B. cereus and B.
subtilis spores (Graph 2A-C). In Graph 2 the
curves of both sets of experiments almost
completely overlap. Our results demonstrate
that the dilution of glycoconjugates does not
impede spore recognition as seen in Graph
2. We would like to point out that due to
dilution, the recognition ability of B. cereus
using Galβ1-4 Glcβ–PAA-flu (6) has
increased, while the recognition ability of B.
cereus using Galα1-3 GalNacα–PAA-flu (7)
has observed with approximately same OD
values (Graph 2A-C).
Graph 2. Role of dilution effect of
glycoconjugates on recognition abilities of
B. cereus and B. subtilis spores. There are
glycoconjugate dilution factors ×10 (A);
×100 (B); ×1000 (C).
9 of 33
Tuesday , August 17, 2004
Elsevier
Revie
w C
opy
10
The recognition ability of B. subtilis using Galβ1-3Galβ–PAA-flu (5), and GlcNacβ1-4
GlcNacβ–PAA-flu (10) has increased using ×10, ×100 dilutions (Graph 2A, B). Presumably,
different units of the glycoconjugates recognized different receptor units expressed on the
exterior (Figure 5) of B. cereus and B. subtilis spores (Figure 3).
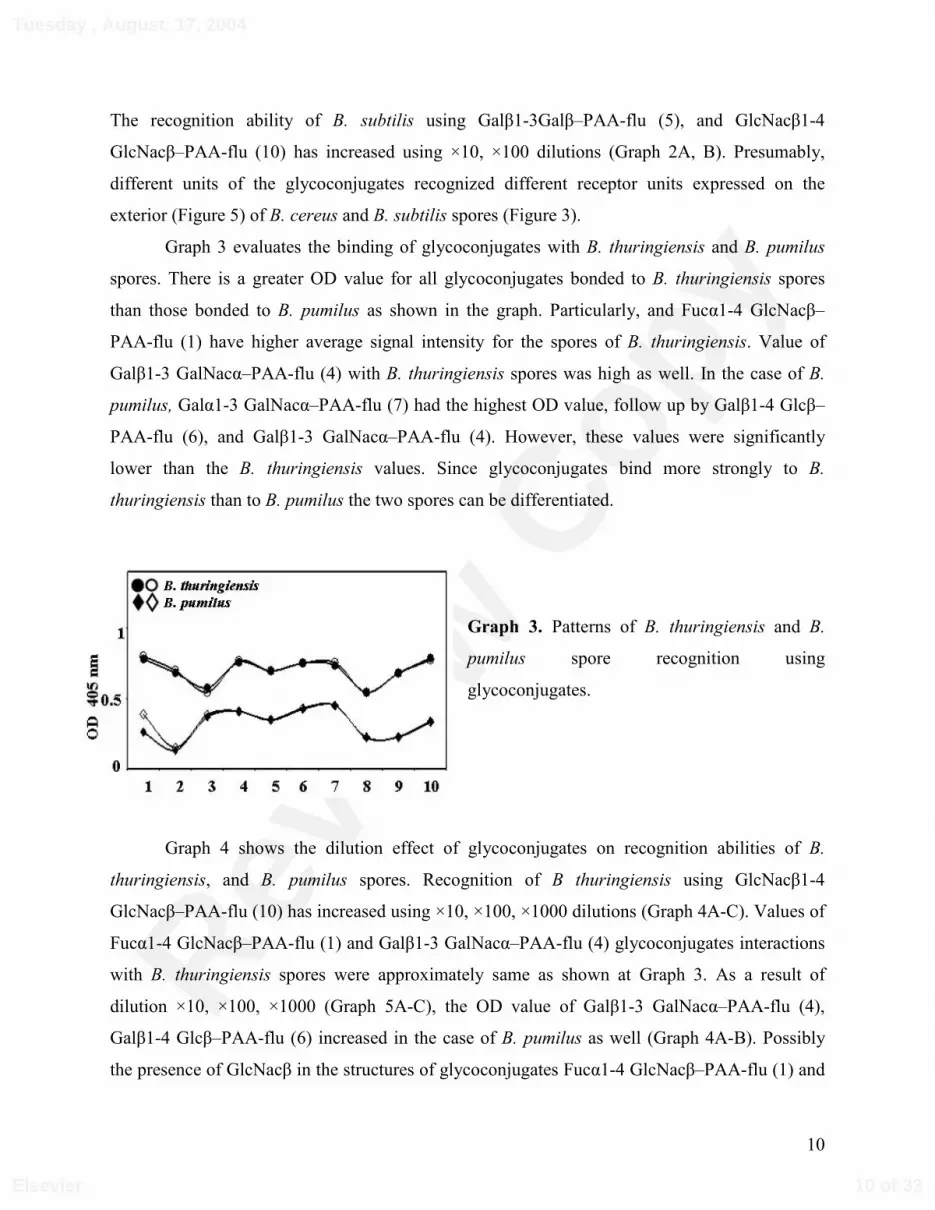
Graph 3 evaluates the binding of glycoconjugates with B. thuringiensis and B. pumilus
spores. There is a greater OD value for all glycoconjugates bonded to B. thuringiensis spores
than those bonded to B. pumilus as shown in the graph. Particularly, and Fucα1-4 GlcNacβ–
PAA-flu (1) have higher average signal intensity for the spores of B. thuringiensis. Value of
Galβ1-3 GalNacα–PAA-flu (4) with B. thuringiensis spores was high as well. In the case of B.
pumilus, Galα1-3 GalNacα–PAA-flu (7) had the highest OD value, follow up by Galβ1-4 Glcβ–
PAA-flu (6), and Galβ1-3 GalNacα–PAA-flu (4). However, these values were significantly
lower than the B. thuringiensis values. Since glycoconjugates bind more strongly to B.
thuringiensis than to B. pumilus the two spores can be differentiated.
Graph 3. Patterns of B. thuringiensis and B.
pumilus spore recognition using
glycoconjugates.
Graph 4 shows the dilution effect of glycoconjugates on recognition abilities of B.
thuringiensis, and B. pumilus spores. Recognition of B thuringiensis using GlcNacβ1-4
GlcNacβ–PAA-flu (10) has increased using ×10, ×100, ×1000 dilutions (Graph 4A-C). Values of
Fucα1-4 GlcNacβ–PAA-flu (1) and Galβ1-3 GalNacα–PAA-flu (4) glycoconjugates interactions
with B. thuringiensis spores were approximately same as shown at Graph 3. As a result of
dilution ×10, ×100, ×1000 (Graph 5A-C), the OD value of Galβ1-3 GalNacα–PAA-flu (4),
Galβ1-4 Glcβ–PAA-flu (6) increased in the case of B. pumilus as well (Graph 4A-B). Possibly
the presence of GlcNacβ in the structures of glycoconjugates Fucα1-4 GlcNacβ–PAA-flu (1) and
10 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
11
GlcNacβ1-4 GlcNacβ–PAA-flu (10)
facilitates binding to B. thuringiensis
spores. The presence of Galβ in the ligands
increases the binding ability of
glycoconjugates in the case of B. pumilus.
Diluted glycoconjugates bind more
strongly to B. thuringiensis than to B.
pumilus the two spores can be
differentiated
Graph 4. Role of dilution effect of
glycoconjugates on recognition abilities of
B. thuringiensis and B. pumilus spores.
There are glycoconjugate dilution factors
×10 (A); ×100 (B); ×1000 (C).
Graph 5. Patterns of B. cereus and
B. thuringiensis spore recognition.
11 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
12
Graph 5 illustrates the average signal intensity values for the spores that have had the
highest average signal intensity: B. cereus and B. thuringiensis. The data illustrates that i) despite
being the most similar species of spore morphology, with both appendages and a developed
exosporium 59, they can still be differentiated by these synthetic ligands; ii) glycoconjugates
Galβ1-4 Glcβ–PAA-flu (6) and Galα1-3 GalNacα–PAA-flu (7) have a marked affinity for both
species of spores. The OD values for glycoconjugates GlcNacβ1-4 GlcNacβ–PAA-flu (10), and
Fucα1-4 GlcNacβ–PAA-flu (1) which bound to both B. thuringiensis and B. cereus spores, were
greater in the case of B. thuringiensis than of B. cereus as shown by Graph 5.
As a result of dilution ×10, ×100, ×1000 (Graph 6A-C), the OD value for Galβ1-4 Glcβ–
PAA-flu (6) increased with respect to B.
cereus, while decreased in the case of B.
thuringiensis. The OD value for
glycoconjugate GlcNacβ1-4 GlcNacβ–
PAA-flu (10) increased with respect to B.
thuringiensis and decreased in the case of B.
cereus (Graph 6A-C). Diluted ×1000
glycoconjugates differentiated B. cereus and
B. thuringiensis (Graph 6C).
Graph 6. Role of dilution effect of
glycoconjugates on recognition abilities of
B. cereus and B. thuringiensis spores. There
are glycoconjugate dilution factors ×10 (A);
×100 (B); ×1000 (C).
12 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
13
Graph 7. Patterns of B. subtilis and B.
pumilus spores recognition.
We compared the average signal
intensity values for B. subtilis and B. pumilus spores as well (Graph 7). These species have a
different morphology (Fig.3 B-C). B. subtilis has only an exosporium and B. pumilus has only
appendages as previously reported 59, 61-63. Here the data shows that i) both B. pumilus and B.
subtilis have a low affinity binding with the ten glycoconjugates in question. No average signal
intensity is greater than 0.5. ii) Glycoconjugates Galα1-3 GalNacα–PAA-flu (7) Galβ1-4 Glcβ–
PAA-flu (6), Galβ1-3 GalNacα–PAA-flu (4), Fucα1-4 GlcNacβ–PAA-flu (1) and GlcNacβ1-4
GlcNacβ–PAA-flu (10) showed the highest average signal intensity for B. pumilus. iii) Used
glycoconjugates have noticeably lower average signal intensities with B. subtilis than with B.
pumilus (Graph 7). iv) Glycoconjugates Galα1-3 GalNacα–PAA-flu (7), Galβ1-3Galβ–PAA-flu
(5), Galβ1-4 Glcβ–PAA-flu (6), and GlcNacβ1-4 GlcNacβ–PAA-flu (10) have bound to both
spores, but stronger to B. pumilus than to B. subtilis spores. Graph 8 shows the effects of dilution
for recognition of B. subtilis and B. pumilus. On the whole dilutions decreased binding of
glycoconjugates to B. subtilis and B. pumilus (Graph 8A-C).
13 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
14
Graph 8. Role of dilution effect of
glycoconjugates on recognition abilities of
B. subtilis and B. pumilus pores. There are
glycoconjugate dilution factors ×10 (A);
×100 (B); ×1000 (C).
In trial A and B, only glycoconjugates
Galβ1-3 GalNacβ–PAA-flu (3), and Galβ1-4
Glcβ–PAA-flu (6) allowed both species of
spores to be distinguished (Graph 8A, B). At
×1000 with the exception of glycoconjugate
Galβ1-3 GalNacβ–PAA-flu (3) and Galβ1-
3Galβ–PAA-flu (5), the OD values for both
species of spores are quite similar (Graph
8C).
Graph 9. Patterns of B. subtilis and B.
thuringiensis spore recognition.
14 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
15
Graph 9 shows that glycoconjugates can recognize and distinguish B. subtilis and B.
thuringiensis spores. Even when diluted,
glycoconjugates were still able to recognize
and distinguish both B. subtilis and B.
thuringiensis spores as shown in Graph 10.
Graph 10. Role of dilution effect of
glycoconjugates on recognition abilities of
B. subtilis and B. thuringiensis
spores. There are glycoconjugate dilution
factors ×10 (A); ×100 (B); ×1000 (C).
Graph 11. Patterns of B. cereus and B. pumilus
spore recognition.
15 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
16
Next, we compared B. cereus and B. pumilus spore recognition (Graph 11). B. cereus and
B. pumilus possess appendages (Fig.3 A, C).
In Graph 11, with the exception of glycoconjugates Galβ1-3 GalNacβ–PAA-flu (3), and
GalNacβ1-3 GalNacα–PAA-flu (8), the OD values were significantly higher for B. cereus than
B. pumilus. The interaction of Galβ1-4 Glcβ–PAA-flu (6) and Galα1-3 GalNacα–PAA-flu (7)
with B. cereus and B. pumilus had the highest OD values.
Like Graph 11, Graph 12 shows that further dilution markedly increases the OD value for
glycoconjugate Galβ1-4 Glcβ–PAA-flu (6) in the case of B. cereus. B. pumilus showed lower
overall OD values.
Graph 12. Role of dilution effect of
glycoconjugates on recognition abilities of
B. cereus and B. pumilus spores
recognition. There are glycoconjugate
dilution factors ×10 (A); ×100 (B); ×1000
(C).
16 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
17
Figure 1. Glycoconjugate binding to B. subtilis, B. pumilus,
B. cereus, B. thuringiensis spores. There are stock glycoconjugate solutions
(A), ×10 (B), ×100 (C), ×1000 diluted (D). Interactions with higher OD values presented. There
are Galβ1-4 Glcβ–PAA-flu (6), Galα1-3 GalNacα–PAA-flu (7) for B. cereus; Galα1-3 GalNacα–
PAA-flu (7) for B. subtilis; Fucα1-4 GlcNacβ–PAA-flu (1), GlcNacβ1-4 GlcNacβ–PAA-flu (10)
for B. thuringiensis; Galβ1-4 Glcβ–PAA-flu (6), Galα1-3 GalNacα–PAA-flu (7), GlcNacβ1-4
GlcNacβ–PAA-flu (10) for B. pumilus.
Figure 1 shows glycoconjugate-spore interactions with higher OD values. Galβ1-4 Glcβ–
PAA-flu (6) was capable recognize of B. cereus (Fig. 1 A-D). Recognition ability of B. cereus
has increased with dilutions of glycoconjugate Galβ1-4 Glcβ–PAA-flu (6) (Fig. 1 B-D).
Although, Galβ1-4 Glcβ–PAA-flu (6) was among glycoconjugates with OD values that bound to
B. pumilus spores. ×10, ×100, ×1000 diluted glycoconjugates did not increase binding of Galβ1-
4 Glcβ–PAA-flu (6) with B. pumilus spores (Fig. 1 B-D). Galα1-3 GalNacα–PAA-flu (7) has
shown the binding with B. cereus, B. subtilis, and B. pumilus, but OD values were lower with B.
subtilis, and B. pumilus spores. Dilutions of Galα1-3 GalNacα–PAA-flu (7) did not change
binding ability (Fig. 1 B-D). A high OD value was observed for interaction of GlcNacβ1-4
GlcNacβ–PAA-flu (10) glycoconjugate with B. thuringiensis and with B. pumilus spores.
Recognition of B. thuringiensis using GlcNacβ1-4 GlcNacβ–PAA-flu (10) has increased using
17 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
18
×10, and ×100 dilutions (Fig. 1B-C), while the recognition ability of B. thuringiensis using
Fucα1-4 GlcNacβ–PAA-flu (1) has decreased. GlcNacβ1-4 GlcNacβ–PAA-flu (10) was capable
of binding to B. pumilus spores, but the OD value was lower than for B. thuringiensis. A high
OD value was observed for interaction of Fucα1-4 GlcNacβ–PAA-flu (1) glycoconjugate and B.
thuringiensis. The OD value of ×10, ×100 dilutions of Fucα1-4 GlcNacβ–PAA-flu (1)
glycoconjugate and B. thuringiensis interaction was approximately the same. In summary, using
glycoconjugates as synthetic ligands allowed us to recognize and distinguish four species of
bacteria spores from each other (Graphs 1, 3, 5, 7, 9, 11, Fig. 1). Changes in the concentration of
glycoconjugates did not hinder the recognition (Graphs 2, 4, 6, 8, 10, 12, Fig. 1 B-D). Diluted
glycoconjugates were still capable of recognizing and distinguishing individual spores.
According to recent research, the avidity of binding glycoconjugates to the surface of bacterial
spores that display multiple receptor sites depends on both the structure of the antigen and on
that of the ligands 77, 37-40. Though both the antigen and the ligands may display different
bimolecular units in various arrangements; glycoconjugates afford us the chance to mimic the
spores’ recognition.
Nikon Eclipse E400 Pol, a polarizing optical microscope, was used to visually confirm
the time-scale for ligand-target complex presence. Images were taken at constant intervals prior
to (Fig. 2A) and after ELISA completion (Fig. 2 B, C). Figure 2 A shows spores only. The
glycoconjugate-spore complexes have appeared under optical microscopy as shown at the Figure
2 B.
Figure 2. Spores as observed under optical microcopy as a control (A). Interaction of
glycoconjugate-spore complexes under optical microcopy (B) and fluorescent microscopy (C).
18 of 33
Tuesday , August 17, 2004
Elsevier
Revie
w C
opy
19
Figure 2 C depicts the changes in fluorescence intensities that characterize the interaction with
the help of fluorescein-labeled glycoconjugates. This method appears to be a convenient method
to reveal the presence of interacted glycoconjugate-spores complexes.
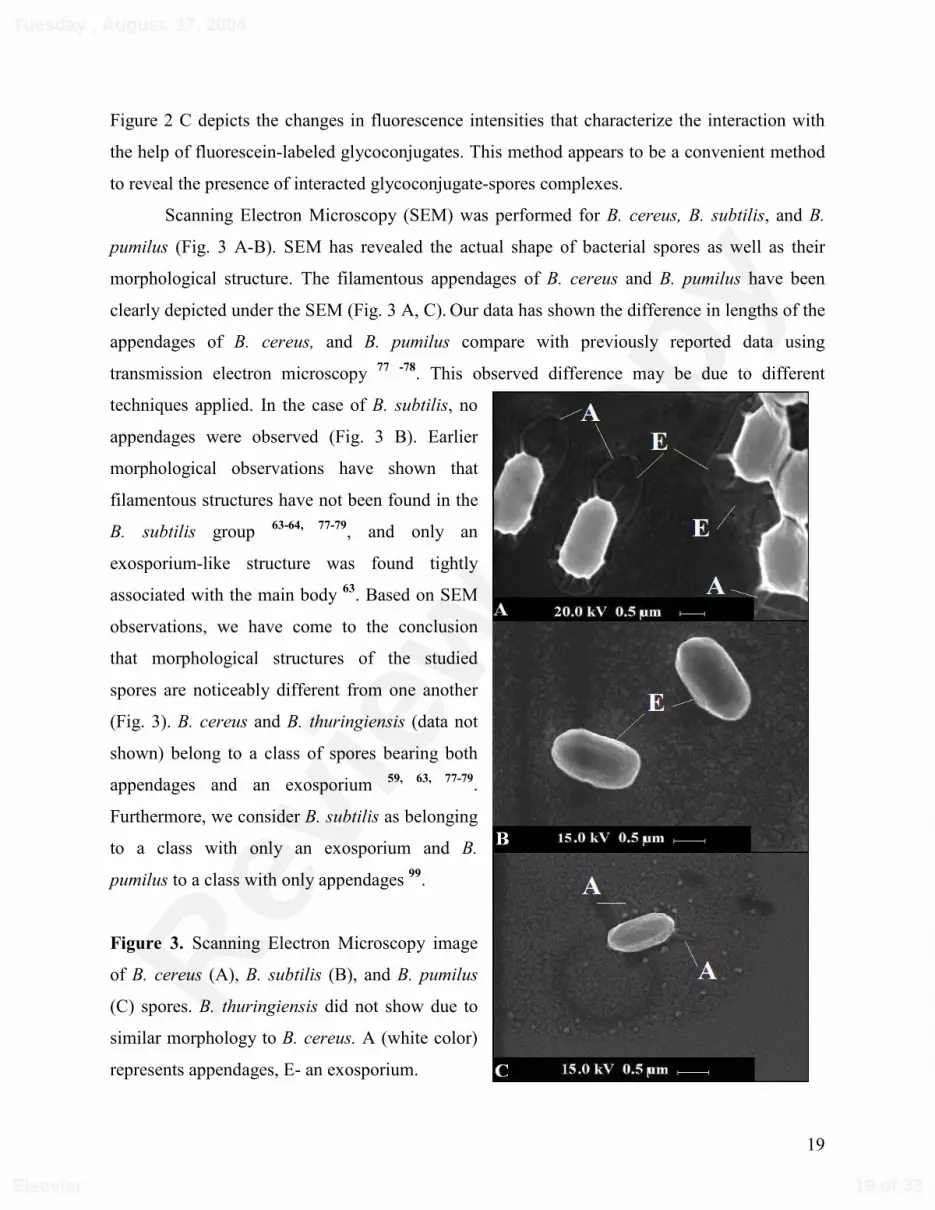
Scanning Electron Microscopy (SEM) was performed for B. cereus, B. subtilis, and B.
pumilus (Fig. 3 A-B). SEM has revealed the actual shape of bacterial spores as well as their
morphological structure. The filamentous appendages of B. cereus and B. pumilus have been
clearly depicted under the SEM (Fig. 3 A, C). Our data has shown the difference in lengths of the
appendages of B. cereus, and B. pumilus compare with previously reported data using
transmission electron microscopy 77 -78. This observed difference may be due to different
techniques applied. In the case of B. subtilis, no
appendages were observed (Fig. 3 B). Earlier
morphological observations have shown that
filamentous structures have not been found in the
B. subtilis group 63-64, 77-79, and only an
exosporium-like structure was found tightly
associated with the main body 63. Based on SEM
observations, we have come to the conclusion
that morphological structures of the studied
spores are noticeably different from one another
(Fig. 3). B. cereus and B. thuringiensis (data not
shown) belong to a class of spores bearing both
appendages and an exosporium 59, 63, 77-79.
Furthermore, we consider B. subtilis as belonging
to a class with only an exosporium and B.
pumilus to a class with only appendages 99.
Figure 3. Scanning Electron Microscopy image
of B. cereus (A), B. subtilis (B), and B. pumilus
(C) spores. B. thuringiensis did not show due to
similar morphology to B. cereus. A (white color)
represents appendages, E- an exosporium.
19 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
20
Given the varied structure present in all four spores used in this experiment, it is our belief that
glycoconjugates can account for their varied morphology (Fig. 3). Varied morphology here not
only refers to the many different receptors
expressed on the spore surface and their
arrangement (Fig. 3) but it also refers to the
existence of filamentous appendages on B.
cereus, B. thuringiensis (data not shown)
and B. pumilus (Fig. 3 A, C), and the lack of
an exosporium in the case of B. subtilis (Fig.
3B) 99. These results, at first glance, allude to
the role played by structure in avidity.
Furthermore, for a number of
bacterial genera, including Bacillus, the
presence of extracellular polysaccharides in
bacterial and fungal spores has previously
been demonstrated 57-58, 64-66, 81.
In order to identify receptors
expressed on the spore surface that involved
glycoconjugate-spore interaction, the
exterior layers of bacterial spore strains have
been isolated using physico-chemical
treatment as described in the Experimental
section.
Figure 4. Tapping Mode AFM images
showing spore prior to (A) and after
treatment (B, C). Mixture of isolated spore’s
exterior (B) and remaining inner shell (C) of
B. cereus ATCC 11778 spore: a- height and
b- amplitude.
20 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
21
The process of isolation consists of reducing the disulfide bonds in a protein, by using a
sulfhydryl agent such as 2% 2-mercaptoethanol. Successful isolation of the exterior layer was
confirmed by AFM (Fig. 4). Figure 3 shows B. cereus ATCC 11778 spore prior to (Fig. 3 A) and
after physico-chemical treatment (Fig. B, C).
Then, the content of the exterior of four Bacillus spore strains was analyzed using FACE
(Fig. 5). FACE allows the examination of monosaccharides from neutral and amine sugars
hydrolysis. FACE electrophoresis analysis reveals the presence of several monosaccharides,
namely N-acetylglucosamine (GlcNAc), N-acetylgalactosamine (GalNAc), galactose, mannose,
fucose as well as glucose.
Figure 5. Monosaccharide composition of (2- 9) spores’ exterior and AMAC–labeled
monosaccharide standard (1, 10); lines 2, 4, 6, 8 represent monosaccharides from neutral sugar
hydrolysis; lines 3, 5, 7, 9 represent monosaccharides from amino sugar hydrolysis. Lines 2, 3
represent monosaccharide composition of B. cereus; lines 4, 5 represent monosaccharide
21 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
22
composition of B. thuringiensis spores; lines 6, 7 represent monosaccharide composition of B.
subtilis; lines 8, 9 represent monosaccharide composition of B. pumilus spores.
Both, B. cereus and B. thuringiensis contained neutral and amine profiles. In contrast, B.
subtilis and B. pumilus spore’s exterior exclusively exhibit neutral sugar profiles. Galactose has
been identified in the B. cereus spores exterior through neutral and amine sugar profiles. A
neutral sugar profile of B. thuringiensis appendages exhibited two monosaccharides: glucose and
galactose, whereas the amine sugar profile contained only galactose. Mannose, fucose, and
galactose were detected on the B. subtilis spore’s exterior. B. pumilus spores contained galactose
and GlcNAc. FACE has proved to be a powerful tool in identification of monosaccharide units of
studied spores. B. cereus 81, as well B. subtilis typically contain high levels of galactose 81, 82.
Glucose is considered the principal neutral carbohydrate found in B. anthracis, B. thuringiensis
and B. subtilis spores 58. Using FACE, the carbohydrates that comprised the receptor recognition
sites on the spore’ exterior were enumerated 33. It was shown that BxpB protein appears to be
glycosylated or associated with glycosylated material 18.
In addition, carbohydrates are recognized as differentiation markers and as antigenic
determinants. Recent evidence suggests that carbohydrate determinants may serve as binding
molecules and are essential for recognition through the interaction between their carbohydrate
moieties 83-88, 91-94. The crucial biological processes are either mediated by oligosaccharide
sequences, by common terminal sequences, or even by further modifications of the sugars
themselves. Such monosaccharide units of oligosaccharides/ glycoproteins/ glycolipids are also
more likely to be targets for recognition by pathogenic toxins and microorganisms 44, 80 or other
molecules. Being used primarily as a substrate for recognition, fluoresceinated glycoconjugates
played a major role in our study. As synthetic ligands they helped to establish the relevancy of
carbohydrate-carbohydrate interaction. The carbohydrates located on the spores’ exterior surface
created the potential multivalent displays of receptor/s (Fig. 5) to be recognized by carbohydrates
units of disaccharide glycoconjugates (Graphs 1-12, Fig. 1). The technique presented in this
study may be valuable in finding and isolating future spore receptor recognition sites and may be
valuable in discovering of the inhibitors that are essential for the bacterial spores’ survival.
Detecting carbohydrate-binding sites is a suitable application since most pathogens possess
unique cell-surface carbohydrates. The specific carbohydrate structures differentially expressed
22 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
23
in each cell type at each stage of development are assumed to be recognized by complementary
molecule(s) expressed on the surface of the counterpart interacting cell 89, 90. Previously reported
experimental data directly shows complex carbohydrates in the recognition processes, including
adhesion between cells, adhesion of cells to the extracellular matrix, and specific recognition of
cells by one another 46-49, 67-72. It was shown that carbohydrate-carbohydrate interactions play an
important role for complimentary binding of glycosphingolipids 91- 93.
The major advantage of our approach is based on glycoconjugates binding capabilities to
the studied bacterial spores. Furthermore, we applied our method to detect carbohydrate binding
and to investigate the selectivity of glycoconjugates for various Bacillus spores. Our results
demonstrated that disaccharide glycoconjugates were able to recognize and to distinguish the
Bacilli group including B. cereus, B. thuringiensis, B. pumilus, and B. subtilis spores (Graphs 1-
12, Fig. 1). Furthermore, our results demonstrated that the dilution of glycoconjugates does not
impede spore recognition. The advantage associated with using glycoconjugates as recognition
tools is they contain a full array of carbohydrate determinants that are essential for recognition.
This recognition may be critical to optimal binding. In the future, the experimental evidence can
shed some light on the function of complex carbohydrates in these recognition processes and
adhesion between cells, adhesion of cells to the extracellular matrix, and specific recognition of
cells by one another 83-88. Another advantage of the proposed platform becomes clearly evident
when the chromogenic reaction product is quantitatively determined using a plate reader upon
completion of the immuno- and enzymatic reactions. This approach has also dramatically
increased the potential of rapidly determining the presence of specific carbohydrate epitopes.
Most importantly, there is a high surface-area to-volume ratio in the immunosorbent.
Furthermore, it was shown that molecular interaction based on carbohydrate-carbohydrate
interaction is a rapid process (1-5 min) 95. While a single molecular force between a pair of
carbohydrates is weak, the strength of interaction between clusters of carbohydrates is
comparable to that of protein-protein interactions 45, 88. Secondly, the U-shaped wells ensure
improved contact between the sample and solid-phase glycoconjugates, resulting in an increased
antigen-antibody encounter rate. Moreover, immunobinding is quantitatively achieved during the
relatively short time of immunoassays.
Moreover, glycoconjugates were observed to interact with a monosaccharide epitope(s)
expressed on the exterior layer of spores (Graphs 1-12, Fig. 5). This observation also suggests
23 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
24
that our approach is of value when it comes to discovering unexpected, but biologically relevant,
bacterial species. The detection of carbohydrate-binding epitopes is considered useful for this
type of application since most pathogens possess unique cell-surface carbohydrates. Taken
together, the major advantage of our approach lies in the glycoconjugate-specificity and
selectivity to spore species, reproducibility, and the magnitude of interaction. The multiple
aspects of carbohydrate-carbohydrate recognition lead to applications like the detection of
specific bacterial spores. In conclusion, the this study provides further evidence that specific
recognition of bacterial spores is based on multivalent carbohydrate-carbohydrate interactions
between disaccharide glycoconjugates that act as ligands and monosaccharide units expressed on
the exterior of bacterial spores.
3. Experimental
3.1 Materials.
Multivalent disaccharide-PAA-flu glycoconjugates were purchased from GlycoTech Inc.
(Rockville, MD, USA). Tween 20, 30% Albumin bovine solution, water W3500 tissue culture
grade, 2,2’–azino–bis(3–ethylbenzthiazoline–6–sulfonic acid) liquid substrate system for ELISA,
proteins standards were purchased from Sigma- Aldrich (St. Louis, MO, USA). Anti-mouse
(IgG+IgM)-horseradish peroxidase (HRP) conjugate was purchased Roche (Indianapolis, IN,
USA). Sterile bacterial spore suspensions, namely B. cereus ATCC 11778, B. thuringiensis
ATCC 29730, B. subtilis ATCC 9372, and B. pumilus ATCC 27142 were purchased from Raven
Biological Laboratories Inc. (Omaha, Nebraska, USA). The fluorophore assisted carbohydrate
electrophoresis FACE® monosaccharides composition kit was obtained from Glyko Inc.,
(Novato, CA, USA). Monosaccharides were obtained from Aldrich (Milwaukee, WI, USA).
3.2 Isolation of Spores’ Exterior Layer
The isolation of the exterior layer of spores was performed as described earlier 62. Briefly, a 500
µl spore suspension with a concentration of 2×106/100µl was mixed with 2% solution of 2-
mercaptoethanol (1mM carbonate - bicarbonate buffer, pH 10.0), vortexed for 5 min, and
incubated at 37oC for 120 min. After exposure to the reagent, the mixture was centrifuged at
24 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
25
4,000 × g for 20 min. The fractions, containing appendages (supernatant) and spore inner
constituents (pellets), were washed four times with deionized water. Successful isolation of
fractions was confirmed by AFM (Fig. 4) prior to further use.
3.3 Scanning Electron Microscopy (SEM)
Spore samples were deposited on the AFM mica disks for SEM observations and desiccated
during seven days. The surface of the specimen for SEM must be electron conducting. This
requires, in most cases, metal layer coating by sputtering of the specimen surface 96. Normally, in
order to satisfyingly scan spore samples using SEM, each sample is to be coated individually by
conductive materials 97- 98 such as gold and/or palladium, due to transparency. Samples were
coated with a thin film of evaporated gold, approximately 10 nm in thickness, for 60 sec and then
examined with a scanning electron microscope S-570 by Hitachi (44 Old Ridenburg Road,
Danbury, CT 06810) at an accelerating voltage of 15-20 kV.
3.4 Atomic Force Microscopy (AFM)
Solutions of untreated spores and purified spore’ exterior layer were immobilized on mica discs
(Digital Instruments, Inc., Santa Barbara, CA, USA) using sterile syringes, then dried in air at
room temperature. The prepared samples were later mounted on an AFM sample holder for
imaging. All AFM observations were carried out at room temperature (20°C), using a Nano
Scope® IIIa controller as well as a MultiModeTM microscope (Digital Instruments, Inc., Santa
Barbara, CA, USA) operating in tapping mode (amplitude) together with an E scanner. A 125
µm silicon Nanoprobe (Digital Instruments, Inc.) was employed. The calculated spring constant
was 0.3 N/m, the resonance frequency remained in the range of 240-280 kHz, the radius of
curvature was approx. 10 nm, and the scan rate was of 1 µm/s. Flattening and high-pass filtering
of the image data were performed in order to remove the substrate slope from images as well as
high-frequency noise strikes, otherwise more pronounced in the high-resolution tapping mode
imaging.
3.5 FACE of Bacterial Spores’ Exterior Layer
FACE was performed with the FACE® monosaccharide composition kit according to
manufacturer’s instructions (Glyko, Inc., Novato, CA, USA). The kit allows for the analysis of
25 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
26
constituent neutral and amine monosaccharides. Briefly, monosaccharides originating in the
exterior layers of the spore’ strains were hydrolyzed. Neutral monosaccharides were hydrolyzed
by dissolving in 2 M trifluoroacetic acid (TFA) at 100°C for 5 h, whereas amino sugars were
hydrolyzed by dissolving isolated mixture of the exterior layers in 100 µl of 4 M HCl at 100°C
for 3 h. Following hydrolysis, the mixture was dried under reduced pressure. The dried
monosaccharides from the amino hydrolysis reaction were re-N-acetylated by the addition of a
re-acetylation reagent M3 according to the manufacturer’s protocol. The dried monosaccharide
from both hydrolyses were labeled with a fluorescent tag (AMAC, see Scheme 1 for labeling
reaction) and incubated overnight at 37°C.
Scheme 1. FACE labeling chemistry. Fluorophore labeling reaction was carried out with AMAC
forming the Schiff base. Reduction occurred with NaBH3CN.
Separation of the fluorophore labeled monosaccharides was done through polyacrylamide gel
electrophoresis. Upon comparison with the standard (the standard mixture of monosaccharides,
consists of 100 pmol each of AMAC-labeled N-acetylglucosamine (GlcNAc), N-
acetylgalactosamine (GalNAc), galactose, mannose, fucose, and glucose, was then loaded into a
lane adjacent to the lane of amine and neutral hydrolysis reaction) the resulting band patterns
illustrated the monosaccharide composition of the starting material which. Electrophoresis was
performed at 5°C with a constant electric current per gel for 75 min.
3. 6 Enzyme Linked Immunosorbent Assay (ELISA)
The utilization of ELISA for the study bacterial spores with glycoconjugates was performed as
follows: each of the wells of microplate (Nunc, MaxiSorp) was coated with 20 µl disaccharide
glycoconjugate-PAA-flu overnight at 4 oC. The plates were washed three times with 50 µl per
well phosphate-buffered saline (PBS) containing 0.1% Tween 20. Then the plates blocked with
26 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
27
100 µl /well of 3% BSA in PBS at 37 oC for 1 h or 2 hrs at room temperature followed by a
washing step. Later, spores were added at a 7.5 µl /well rate and incubated for 1-2 hrs at room
temperature with continuous shaking. The plate was washed three times with PBS containing
0.1% Tween-20, after which 12.5 µl anti-mouse (IgG + IgM) - horseradish peroxidase(HRP)-
labeled conjugate (1:4000 dilution) (Roche Diagnostics Corp., Indianapolis, IN) was added and
incubated at 37 oC for 1 h. After washing, a substrate solution 2,2’–azino–bis(3–
ethylbenzthiazoline–6–sulfonic acid) was added and the colorimetric reaction was terminated by
addition of 50 µl /well of 1 M H2SO4. Spectrophotometric measurements were carried out with a
microplate reader SpectraMax® Plus384 (Molecular Devices Corp., Sunnyvale, CA, USA) at 405
nm. SpectraMax® Plus384 which allows carrying out a standard spectrophotometric measurement
in seconds. PathCheck Sensor of SpectraMax® Plus384 was used to measure the depth (optical
pathlength) of samples within the microplate. SOFTmax® PRO software was used to
automatically normalize the well absorbance with an equivalent path length of 1 cm 100. Blank
readings were subtracted from the optical density of the final reaction in order to obtain the
corrected absorbance value.
3. 7 Study of the Dilution Effect
Solutions of disaccharide glycoconjugates were prepared according to the manufacture’s
protocol. The solutions were then diluted ×10, ×100, ×1000 times into a phosphate-buffered
saline (PBS) / 0.2% NaH3 buffer. The ELISA platform was used in same manner as described
above (3.6 of Experimental section). Recognition was accomplished using surface-immobilized
molecules such as native and diluted glycoconjugates as ligands, which interact with other
species such as analytes (spores) in solution based on binding curves (Graphs 2, 4, 6, 8, 10, 12).
3. 8 Conventional Optical and Fluorescent Microscopy
Visual confirmation of the time-scale of glycoconjugate-spore complexes presence was
performed prior and after completion of ELISA. The glycoconjugate-spores complexes were
visually examined under fluorescent microscopy using a Nikon Eclipse E400 POL, at a
magnification of 400 times. Digital pictures of spores and of interactions of glycoconjugate-
spore complex were taken in real-time (Fig. 2).
27 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
28
Acknowledgments
This research was supported by grant 0660076225, originating from the Defense Advanced
Research Projects Agency (DARPA). We express our deepest gratitude to Joany Jackman,
Senior Scientist, Applied Physics Laboratory, Johns Hopkins University, Laurel, MD for
valuable discussions during this study. We would also like to thank Prof. Mary Cowman, Othmer
Department of Chemical and Biological Sciences and Engineering, Polytechnic University, NY
for allowing us to use her atomic force microscope. We express our appreciation to Professor
Said Nourbakhsh, Department of Mechanical Engineering for technical assistance during
scanning electron microscopy, Prof. Arnost Reiser and Prof. Edward Weil, Othmer Department
of Chemical and Biological Sciences and Engineering, Polytechnic University, NY for critical
reading of the manuscript.
References
1. Doyle, M.P.; Beuchat, L.P.; Montville, T.J. In Food Microbiology. Fundamentals and
Frontiers. Doyle, M. P., Beuchat, L. R., and T. J. Montville, Eds; American Society of
Microbiology Press: Washington D.C., 1997.
2. Barnaby, W. In The Plague Markers: The secret World of Biological Warfare, Barnaby, W.
Ed; Vision Paperbacks: London. 1997.
3. Gerhardt, P.; Ribi, E. J. Bacteriol. 1964, 88, 1774-1789.
4. Gerhardt, P.; Marquis, RE. In Regulation of prokaryotic development, Smith, I., Slepecky,
R., Setlow P., Eds; American Society for Microbiology: Washington, D.C., 1989; pp17-63.
5. Setlow, P. Annu. Rev. Microbiol. 1995, 49, 29-54.
6. Setlow, P. J. Appl. Bacteriol., 1994,176, 49S-60S.
7. Nicholson, W.L.; Munakata, N.; Horneck, G.; Melosh, H.J.; Setlow, P. Microbiol. Mol. Biol.
Rev. 2000, 64, 548-572.
8. King, D.; Luna, V.; Cannons, A.; Cattani, J.; Amuso, Ph. J. Clin. Microbiol. 2003, 41, 3454–
3455.
9. Yu, H., Stopa, P.J. In Environmental Immunochemical Methods: Perspectives and
Applications. Van Emon, J.M., Gerlach, C.L., Johnson J.C., Eds; ACS: Washington, DC,
1995; pp 297-306.
28 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
29
10. Vernozy-Rozand, C.; Mazuy, C.; Ray-Gueniot, S.; Boutrand-Loei, S.; Mayrand, A.; Richard,
Y. Lett. Appl. Microbiol. 1997, 25, 442-446.
11. http://www.asc2004.com/23rdASC/summaries/k/KP-12.pdf
12. http://www.internationalpbi.it/default.php?t=site&pgid=508&lang=EN
13. Bell, C.A.; Uhl, J.R.; Hadfield, T.L.; David, J.C.; Meyer, R.F.; Smith, T.F.; and Cockerill, F.
R. III J. Clin. Microbiol. 2002, 40, 2897-2902.
14. Bell, C.A.; Uhl, J.R.; Hadfield, T.L.; David, J.C.; Meyer, R.F.; Smith, T.F.; and Cockerill, F.
R. III J. Clin. Microbiol. 2003, 41, 524-525.
15. Higgins, J.A.; Cooper, M.; Schroeder-Tucker, L.; Black, S.; Miller, D.; Karns, J. S.;
Manthey, E.; Breeze, R.; Perdue, M.L. Appl. Environ. Microbiol. 2003, 69, 593-599.
16. Swartz, M.N. N. Engl. J. Med. 2001, 345,1621-1626.
17. Hartley, H.A.; Baeumner, A.J. Anal Bioanal Chem. 2003, 376, 319-327.
18. Steichen, C.; Chen, P.; Kearney, J.F.; Turnbough, C.L.Jr. J. Bacteriol. 2003, 185, 1903-1910.
19. Harris, P.C.; Cloney, L.; Fong, W.; Fulton, R.E. Paper #359
http://www.responsebio.com/pdf/ASM_poster.pdf
20. www.gsa/gov/mailpolicy
21. Read, T. D.; Peterson, S. N.; Tourasse, N.; Baillie, L. W.; Paulsen I. T., Nelson K. E.,
Tettelin, H.; Fouts, D. E.; Eisen, J. A.; Gill, S. R.; Holtzapple, E. K.; Okstad, O. A.;
Helgason, E. Rilstone, J.; Wu, M.; Kolonay, J. F.; Beanan, M. J.; Dodson, R. J.; Brinkac, L.
M.; Gwinn, M.; DeBoy, R. T.; Madpu, R.; Daugherty, S. C.; Durkin, A. S.; Haft, D. H.;
Nelson, W. C.; Peterson, J. D.; Pop, M.; Khouri, H. M.; Radune, D.; Benton, J. L.;
Mahamoud, Y.; Jiang, L.; Hance, I. R.; Weidman, J. F.; Berry, K. J.; Plaut, R. D.; Wolf, A.
M.; Watkins, K. L.; Nierman, W. C.; Hazen, A.; Cline, R.; Redmond, C.; Thwaite, J. E.;
White, O.; Salzberg, S. L.; Thomason, B.; Friedlander, A. M.; Koehler, T. M.; Hanna, P. C.;
Kolsto, A. B.; Fraser, C. M.. Nature. 2003, 423, 81-86.
22. Logan, N. A.; Turnbull P. C. B. In Bacillus and recently derived genera. Manual of clinical
microbiology. Murray, P. R. E., Baron, J., Pfaller, M. A., Tenover, F. C., Yolken R. H. Eds,
7th ed. American Society for Microbiology, Washington, D.C., 1999; pp 357–369.
23. Tengelsen, L.; Hudson, R.; Barnes, S.; Hahn, C. Emerg. Infect. Dis. 2002, 8, 1093–1095.
24. Williams, D.D.; Benedek, O.; Turnbough, Ch. L. Jr. Appl. Envir. Microbiol. 2003, 69, 6288-
6293.
29 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
30
25. Knurr, J.; Benedek, O.; Heslop, J.; Vinson, R. B.; Boydston, J. A.; McAndrew, J.; Kearney,
J. F.; Turnbough, Ch. L. Jr. Appl. Envir. Microbiol. 2003, 69, 6841-6847.
26. Ji, T.; Levon, K. MMC-9 Poster session, August 19-23rd Brooklyn, NY. 2001.
27. Ji ,T.; Tarasenko, O.; Paquiot, D.; Levon, K. (unpublished data), 2004.
28. Ji, T.; Tarasenko, O.; Paquiot, D.; Turnbough, Ch. L. Jr.; Levon, K. (unpublished data), 2004.
29. Yu, B.; Zhou, Y.; Ji, T.; Turnbough, Ch. L. Jr.; Heslop, J.; Kearney, J.; Jackman, J.; Levon,
K. Open House of Polytechnic University, NY, October 25-26th, 2002.
30. Yu, B.; Tarasenko, O.; Paquiot, D.; Zhou, Y.; Levon, K. (unpublished data). 2004.
31. Kazakov, S.; Kaholek, M.; Ji, T.; Turnbough, Ch. L. Jr.; Levon K. Chem. Commun. 2004,
430-431.
32. Ji, T.; Kaholek, M.; Kazakov, S.; Tarasenko, O. M.; Sharma, N. K.; Zhou, Y. Yu, B.; Levon,
K. 226th ACS National Meeting, New York, NY, September 7-11th, 2003, Polymer Preprint
44 (2) 483-484.
33. Tarasenko, O.; Kudasheva, D.; Alusta, P.; Paquiot, D.; Cowman, M.; Levon, K. 224th ACS
National Meeting, Boston, MA, August 18-22th, 2002, pp. 48-tech.
34. Levon, K.; Tarasenko, O.; Yu, B. U.S. Patent Application Serial No.: 10/719,688, filed:
November 21st, 2003; PCT Application: PCT/US03/37401 filed: November 21st, 2003.
35. Sharma, N.; Levon K. 224th ACS National meeting, Boston, MA, August 18-22nd, 2002,
pp.141.
36. Bertozzi, C. R.; Kiessling, L. L. Science. 2001, 291, 2357- 2364.
37. Kiessling, L.L.; Pohl, N.L. Chem. and Biol. 1996, 3, 71-77.
38. Kiessling, L.L.; Gestwicki, J.E.; Strong, L.E. Curr. Opin. Chem. Biol. 2000, 6, 696-703.
39. Sylvestre, P.; Couture-Tosi, E.; Mock, M. J. Bacteriol. 2003, 185, 1555-1563.
40. Roy, R. Curr. Opin. Struct. Biol.1996, 6,692-702.
41. Zhou, Y.; Yu, B.; Levon, K. Chem. Mater. 2003, 15, 2774-2779.
42. Zhou, Y.; Yu, B.; Levon, K.; Nagaoka, T. Electroanalysis. 2004, 16, 955-960.
43. Zhou, Y.; Yu, B.; Levon, K. Open House of Polytechnic University, NY, October 25-26th,
2002.
44. Varki, A. Glycobiology. 1993, 3, 97-130.
45. Bovin, N.V. In Glycosciences: Status and Perspectives. Gabius H.J., and Gabius S., Eds;
Chapman & Hall, London, 1997, pp. 277- 289.
30 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
31
46. Crocker, P.E.R.; Feizi, T. Curr. Opin. Struct. Biol., 1996, 6, 679-691.
47. Feizi, T. Immunol. Rev. 2000, 173, 79-88.
48. Feizi, T. Glycoconj. J, 2000, 17, 553-565.
49. Karlsson, K.A.; Angstrom, J.; Bergstrom, J.; Lanne, B. APMIS Suppl., 1992, 27, 71-83.
50. Wang, D.; Kabat, E.A. In Structure of antigens. Regenmortal M. H. V. Van., Ed.; CRC Press,
Boca Raton New York London Tokyo, 1996. pp.247-276,
51. Frazier, W.; Glaser, L. Annu. Reu. Biochem. 1979, 48, 491-523.
52. Barondes, S. H. Annu. Rev. Biochem. 1981, 50, 207-231.
53. Sharon, N. Biol. Cell. 1984, 51, 239-246.
54. Mimura, Y.; Sondermann , P.; Ghirlando, R.; Lund, J.; Young, S.P.; Goodall , M.; Jefferis,
R. J. Biol. Chem. 2001, 276, 45539-45547
55. Harris, L.J.; Skaletsky ,E.; McPherson, A. J. Mol. Biol. 1998, 275, 861-872.
56. Burmeister W.P.; Huber A.H. Bjorkman P.J. 1994. Nature. 372:379-383.
57. Marz, L.L.; Beaman, T.C.; Gerhard, P. J. Bacteriol. 1970, 101, 196-201.
58. Scherer, P.S.; Somerville, H.J. Eur. J. Biochem., 1977, 72, 479-90.
59. Hachizuka, Y.; Kojima, K.; Sato, T. J. Bacteriol. 1966, 91, 2382-2384.
60. Sousa, J.C.; Silva, M.T.; Balassa, G. Nature, 1976, 263, 53-54.
61. Hultgren, S.J.; Abraham, S.; Caparon, M.; Falk, P.; St Geme, J.W.D.; Normark, S. Cell.
1993, 73, 887-901.
62. Kozuka, S.; Tochikubo, K. Microbiol. Immunol. 1985, 29, 21-37.
63. Driks, A. Microbiol. Molecul. Biol. Rev. 1999, 63, 1-20.
64. Warth, A.D.; Ohey, D.F.; Murrell, W.G. J. Cell Biol., 1963, 16, 578-592.
65. Kornberg, A.; Spudish, J.A.; Nelson, D.L.; Deutscher, M. Ann. Rev. Biochem. 1968, 37, 51-
78.
66. Hiragi, Y. J. Gen. Microbiol.1972, 72, 87-99.
67. Kayser, K.; Bovin, N.V.; Korchagina, E.Y.; Zelinger, C.; Zeng, F.Y.; Gabius H.-J. Eur. J.
Cancer. 1994, 30A, 653-657.
68. Kayser, K.; Bubenzer, J.; Kayser, G.; Eichhorn, S.; Zemlyanukhina, T.V.; Bovin, N.V.;
Andre, S.; Koopmann, J.; Gabius H. -J. Analyt. Quant. Cytol. Histol. 1995, 17, 135- 142.
69. Bovin, N.V.; Shiyan, S.D.; Mikhalchik, E.V. Glycocon. J. 1995, 12, 427.
70. Rye, P.D.; Bovin, N.V. Glycobiology. 1997, 7, 179-82.
31 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
32
71. Leteux, C.; Childs, R.A.; Chai, W.; Stoll, M.S.; Kogelberg, H.; Feizi, T. Glycobiology. 1998,
8, 227-236.
72. Houseman, B.T.; Mrksich, M. Topics Current Chem. 2002, 218, 1-44.
73. Danguy, A.; Kayser, K.; Bovin, N.V.; Gabius H.-J. Trends Glycosci. Glycotechnol.1995, 36,
261-275.
74. Bovin, N.V. Glycocon. J., 1998, 15, 431- 446.
75. Galanina, O.; Feofanov, A.; Tuzikov, A.B.; Papoport, E.; Crocker, P.R.; Grichine, A.; Egret-
Charlier, M.; Vigny, P.; Le Pendu, J.; Bovin, N.V. Spectrochimica Acta Part A. 2001, 57,
2285-2296.
76. Wang, D.; Liu, S.; Trummer, B. J.; Deng, C.; Wang, A. Nat. Biotechnol. 2002, 20, 275-281.
77. Hachisuka, Y.; Kuno, T. Jpn J Microbiol. 1976, 20, 555-558.
78. Hachisuka, Y.; Kozuka, S. Microbiol. Immunol. 1981, 25, 1201-1207.
79. Hachisuka, Y.; Kozuka, S.; Tsujikawa, M.. Microbiol. Immunol. 1984, 28, 619-624.
80. Hirmo, S.; Artursson, E.; Puu, G.; Wadström, T.; Nilsson, B. Analytical Biochem. 1998, 257,
63-66.
81. Fox, A.; Black, G.E.; Fox, K.; Rostovtseva, S. J. Clin. Microbiol. 1993, 31, 887- 94.
82. Tarasenko, O.; Islam, Sh.; Levon, K. 226th ACS National Meeting, New York, NY,
September 7-11th, 2003, Targeting chemical and biological warfare agents, Preprints Vol.43
(2), pp.1-6.
83. Siegelman, M.H.; Cheng, I.C.; Weissman, I.L.; Wakeland, E.K. Cell, 1990, 61, 611.
84. Iwabuchi, K.; Yamamura, S.; Prinetti, A.; Handa, K.; Hakomori, S. J. Biol. Chem. 1998, 273,
9130- 9138.
85. Handa, K.; Kojima, N.; Hakomori, S. Methods Enzymol. 2000, 312, 447-458.
86. Zheng, M.; Hakomori, S. Arch. Biochem. Biophys., 2000, 374, 93-99.
87. Fenderson, B. A.; Zehavi, U.; Hakomori, S. J. Exp. Med. 1984, 160, 1591-1596.
88. Bird, J. M.; Kimber, S. J. Dev. Biol. 1984, 104, 449-460.
89. Janeway, C. A.; Travers, P.; Walport, M.; Capra, J.D. In Immunobiology. The Immune
System in Health and Disease. 4th Edition. Current Biology Publications, London, UK. 1999.
90. Hakomori, S. Annu. Rev. Biochem. 1981, 50, 733-764.
91. Kojima, N.; Hakomori, S. J. Biol. Chem. 1989, 264, 20159-20162.
92. Kojima, N.; Hakomori, S. J. Biol. Chem. 1991, 266, 17552-17558.
32 of 33
Tuesday , August 17, 2004
Elsevier

Revie
w C
opy
33
93. Stewart, R. J.; Boggs, J. M. Biochemistry 1993, 32, 10666-10674.
94. Eggens, I.; Fenderson, B. A.; Toyokuni, T.; Dean, B.; Stroud, M. R.; Hakomori, S. J . Brol.
Chem. 1989, 264, 9476-9484.
95. Kojima, N.; Shiota, M.; Sadahira, Y.; Handa, K.; Hakomori, S. J. Biol. Chem. 1992, 267,
17264-17270.
96. Bergeret, G.; Gallezot, P. In Handbook of heterogeneous catalysis. vol. 2, Ertl, G.,
Knözinger, H., Weitkamp, J. Eds, Wiley-VCH, Weinheim, Germany. 1997.
97. Brady, D., Duncan, J. R. Enzyme Microb. Technol. 1994,16, 633- 638.
98. Pereira, R. S.; Durán, N.; Teschke, O. Probe Microsc. 1998, 1, 277-282.
99. Tarasenko, O.; Kudasheva, D.; Nourbakhsh, S.; Cowman, M.; Levon, K. 2004, (unpublished
data)
100. http://www.moleculardevices.com
33 of 33
Tuesday , August 17, 2004
Elsevier