tuffreau et al 2013 grotte la adam dobrogea roumanie
TRANSCRIPT

Seediscussions,stats,andauthorprofilesforthispublicationat:https://www.researchgate.net/publication/260190830
Tuffreauetal2013GrotteLaAdamDobrogeaRoumanie
DATASET·FEBRUARY2014
READS
20
7AUTHORS,INCLUDING:
AlainTuffreau
UniversitédeLille,Scienceset…
93PUBLICATIONS518CITATIONS
SEEPROFILE
Availablefrom:SandaBalescu
Retrievedon:04February2016

75
LA GROTTE LA ADAM : UN REPAIRE DE CARNIVORES VISITÉ PAR LES CHASSEURS DU PALÉOLITHIQUE
Alain TUFFREAU1, Roxana DOBRESCU2, Alexandru PETCULESCU3, Emanoil ŞTIUCĂ3, Sanda BALESCU1, François LANOË4, Meredith WISMER5
1 Université Lille 1, UMR 8164, 59655 Villeneuve d’Ascq cedex, France,
2 Institutul de Arheologie « Vasile Pârvan », str. Henri Coandă 11, 010667 Bucureşti, România,
3 Institutul de Speleologie « Emil Racoviţă », Calea 13 Septembrie 13-15, Bucureşti, România,050711,
4 Museum National d’Histoire Naturelle, UMR 7194, Institut de Paléontologie Humaine, 1 rue René Panhard, 75013 Paris, France,
5 Department of Anthropology, University of Iowa, 114 MacBride Hall, Iowa City, IA 52242-1322,USA.
1. INTRODUCTION
La grotte La Adam se situe à moins d’une trentaine de kilomètres du rivage de la Mer Noire, sur la rive droite du ruisseau Vistorna, affluent de la Casimcea, qui entaille un massif calcaire rauracien reposant sur des schistes verts (44° 27’ 52 N, 28° 28’ 16 E). Cette partie de la Dobroudja recèle de nombreuses cavités karstiques dont certaines présentent un remplissage d’âge pléistocène. Cependant, le matériel lithique mis au jour dans les grottes de la Dobroudja est peu abondant [1]. Cette relative pauvreté est certainement le reflet d’occupations humaines peu nombreuses mais résulte aussi, pour une bonne part, des méthodes de fouilles qui ont été mises en œuvre.
Durant les années 50 et 60, les paléontologues C. Rădulescu et P. Samson ont effectué une tranchée d’une longueur de 42 m, d’une largeur de 2,25 m et d’une profondeur maximale de 9 m, qui a recoupé toute la séquence stratigraphique à l’intérieur de la grotte La Adam et à l’extérieur. Ces recherches ont surtout concerné les aspects biochronologiques [2, 3] et les questions concernant la domestication à l’Holocène [4]. La séquence stratigraphique a été datée par son contenu faunistique. Elle s’étend du début du «Riss» jusqu’à l’Holocène. Les restes d’un culte à Mithra ont été découverts au sommet du remplissage.

76
Les données concernant le matériel lithique sont faibles. Aucun préhistorien n’a été associé à l’équipe scientifique. Des pièces taillées ont été découvertes dans toute la séquence stratigraphique qui comprend des dépôts thermoclastiques, du loess, des limons et des argiles. Le matériel lithique, en silex, a été attribué à des industries «pré-moustériennes», moustériennes, aurignaciennes et gravettiennes. Le haut de la séquence, qui se présente sous la forme de plusieurs paliers, contient du Mésolithique et du Néolithique.
Une molaire supérieure humaine, appartenant à un enfant de moins de six ans a été trouvée. Les restes fauniques sont assez abondants avec des espèces froides (Renne, renard polaire, Saiga) ou tempérées (Cervus, Megaceros, Hydruntinus) selon les dépôts.
Nous ne disposons pas de levés détaillés de la stratigraphie de la grotte car la séquence a été décrite d’une manière synthétique qui ne rend pas compte de la géométrie des dépôts.
2. LES FOUILLES RÉALISÉES À LA ADAM DE 2007 À 2009
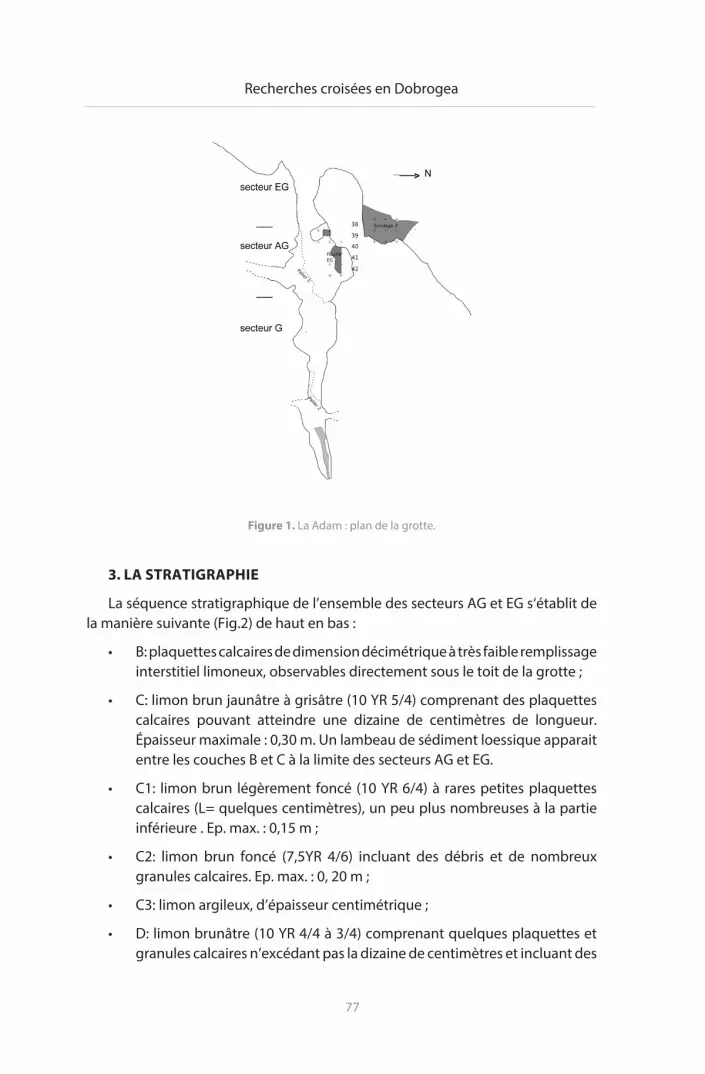
Les travaux ont concerné l’intérieur de la grotte en raison de la présence de gros blocs de calcaire et de déblais dans la partie de la tranchée située à l’extérieur (Fig. 1).
L’avant de la grotte (secteur AG = avant grotte), forme un couloir d’accès à la grotte ; il subsiste contre les parois calcaires des sédiments.
Les fouilles ont concerné plusieurs secteurs :
• à l’intérieur de la grotte, la banquette située dans le secteur EG (= entrée grotte), limitée vers l’intérieur par un palier d’un peu moins de 2 m de hauteur, a été en partie fouillée (m2 P40, P41 et P42 et Q39 sur une quarantaine de centimètres de profondeur). La séquence stratigraphique de la banquette se prolonge vers l’extérieur de la grotte (secteur AG). La partie profonde de la grotte (secteur G), dont la partie visible du remplissage est constituée de plaquettes calcaire, n’a pas fait l’objet de fouilles.
• à l’extérieur de la grotte, plusieurs sondages ont été ouverts. Un seul (S3), situé à 8 m au NE de l’entrée de la grotte, au pied de la paroi calcaire, a montré la présence de sédiments en place, d’âge pléistocène comme l’a confirmé la découverte d’un racloir en silex. Des ossements de petits mammifères et de micromammifères, en bon état de conservation, ont été mis au jour.
77
Recherches croisées en Dobrogea
Figure 1. La Adam : plan de la grotte.
3. LA STRATIGRAPHIE
La séquence stratigraphique de l’ensemble des secteurs AG et EG s‘établit de la manière suivante (Fig.2) de haut en bas :
• B: plaquettes calcaires de dimension décimétrique à très faible remplissage interstitiel limoneux, observables directement sous le toit de la grotte ;
• C: limon brun jaunâtre à grisâtre (10 YR 5/4) comprenant des plaquettes calcaires pouvant atteindre une dizaine de centimètres de longueur. Épaisseur maximale : 0,30 m. Un lambeau de sédiment loessique apparait entre les couches B et C à la limite des secteurs AG et EG.
• C1: limon brun légèrement foncé (10 YR 6/4) à rares petites plaquettes calcaires (L= quelques centimètres), un peu plus nombreuses à la partie inférieure . Ep. max. : 0,15 m ;
• C2: limon brun foncé (7,5YR 4/6) incluant des débris et de nombreux granules calcaires. Ep. max. : 0, 20 m ;
• C3: limon argileux, d’épaisseur centimétrique ;
• D: limon brunâtre (10 YR 4/4 à 3/4) comprenant quelques plaquettes et granules calcaires n’excédant pas la dizaine de centimètres et incluant des

78
lentilles limono-argileuses pouvant atteindre une dizaine de centimètres d’épaisseur. Ep. max. : 0,50 m.
• D base: lentille limono-argileuse brunâtre présentant des traces de bioturbation ;
• D0: limon jaune brunâtre à rares granules de craie, d’épaisseur centimétrique localement présent dans le secteur EG ;
• D1: limon à revêtements argileux, rouge jaunâtre (5 YR 5/6) à brun foncé (7,5 YR 5/8). Ep. max. : 0, 25 m ;
• E: loess jaune brunâtre (10 YR 6/6) incluant de nombreux blocs calcaires et présentant à la partie supérieure des traces de manganèse (Em) ; ép. : max. : 0,80 m ;
• E1: loess jaune rougeâtre présentant quelques revêtements argileux, observable dans le secteur AG sur plus d’un demi mètre (profondeur maximale observable dans la banquette de la fouille ;
• F: épaisse couche de plaquettes calcaires à très faible remplissage interstitiel, observable dans le secteur EG. L’unité F remonte latéralement vers l’intérieur de la grotte à partir du m2 P42.
La géométrie des dépôts présente des différences importantes entre les secteurs AG et EG. Dans le secteur AG, le limon jaune brunâtre E à nombreux blocs de calcaire recélant un paléosol (D1) à la partie sommitale est particulièrement développé. A sa partie inférieure, un autre paléosol (E1) apparaît. Dans le secteur EG, le limon E est nettement moins épais et s’adosse à d’importants dépôts thermoclastiques en direction du secteur G (partie profonde de la grotte). Les dépôts sus-jacents s’épaississent avec différentes unités (C, C1, C2, C3). Ils colmatent une cuvette qui doit constituer l’extrémité d’un chenal d’érosion correspondant à une faille se situant dans l’axe de la galerie adjacente observable du côté sud du secteur EG (Fig. 3). La partie inférieure du remplissage de cette cuvette (D, D base, D0) présente les caractéristiques d’un complexe pédologique correspondant à une pédogenèse différente de celle (D1) que l’on observe au sommet de l’unité E.
Par contre, les corrélations avec les unités observables sur le côté sud du secteur AG demeurent problématiques en raison de la présence de la tranchée des anciennes fouilles.
Le secteur grotte (secteur G) se poursuit au-delà du palier limitant le secteur EG. D’importants témoins du remplissage sont visibles de part et d’autre de la tranchée. L’essentiel du remplissage est constitué de plaquettes calcaires dont la longueur peut atteindre une vingtaine de centimètres.

79
Recherches croisées en Dobrogea
Figure 2. La Adam : profil stratigraphique synthétique.
Le sédiment loessique au sommet de la couche E a donné un âge de 151 + 13 ka (datation par luminescence : IRSL, cf. Balescu, ce volume).
Figure 3. La Adam : stratigraphie dans le secteur EG (côté nord).

80
4. LE MATÉRIEL LITHIQUE
Figure 4. La Adam. 1 : nucleus à une surface de débitage, silex local (couche C). 2 : lame retouchée, schiste vert (couche C). 3 : galet présentant un tranchant distal, schiste vert (couche C2). 4 : nucleus
sur plaquette de schiste vert (couche Em). 5: racloir simple, silex brun allochtone (couche E). 6 : racloir simple, silex brun allochtone (déblais des anciennes fouilles, S3).
Le matériel lithique, peu abondant, provient essentiellement de la partie inférieure du niveau C, de C1 et de C2 dans le secteur EG de la grotte. La matière première utilisée est un silex local. Cependant, dans quelques rares cas, un schiste vert affleurant à quelques kilomètres de La Adam a été utilisé.
Le débitage est atypique et ne saurait permettre une diagnose facilitant une attribution culturelle.
Le sondage S3, à l’extérieur de la grotte, n’a livré que quelques pièces. Un racloir en silex de facture paléolithique moyen a été découvert dans les anciens déblais du sondage S3.
5. LES MICROMAMMIFÈRES
5.1. Les espèces présentes
En ce qui concerne les micromammifères, 179 dents appartenant à 10 espèces ont été déterminées.

81
Recherches croisées en Dobrogea
Figure 5. La Adam : dents de micromammifères.
Les espèces présentes sont caractéristiques de celles qui ont été décrites en Dobroudja [2, 5, 6].

82
5.2. Répartition par couche
Couche B
L’association faunistique témoigne d’un climat assez froid (présence de Microtus gregalis) dans un paysage se situant entre la toundra et la sylvo-steppe.
Couche C
Prédominance des espèces caractéristiques des steppes arides (Eolagurus luteus, Lagurus lagurus et Microtus arvalis, plus de 85% du total des micromammifères) avec des températures moyennes annuelles et des valeurs des précipitations nettement inférieures à ce que l’on observe actuellement en Dobroudja centrale. La présence du rat taupier ou campagnol terrestre (Arvicola terrestris) indique la proximité de l’eau, même si le climat est très aride.
Couche C1
La croissance significative de la fréquence de L. lagurus, la disparition d’Arvicola et l’apparition de Spalax leucodon (rat taupier) montrent que le climat devient plus aride, ayant les caractéristiques d’une steppe semi-désertique sans cours d’eau permanent.
Couche C2
La fréquence d’Eolagurus luteus (45%) témoigne d’une aridité qui augmente accompagnée d‘un léger réchauffement. Le paysage de la Dobroudja ressemble à celui du Kazakhstan et de la Mongolie.
Couche D
Prédominance de Lagurus lagurus et de Microtus arvalis (85%) et présence significative d’Eolagurus luteus (14%), caractéristiques d‘un milieu continental avec une aridité comparable aux steppes actuelles à l’Est de la Mer Noire.
Couche E
Lagurus lagurus reste dominant (65%) mais l’apparition significative de Microtus gregalis (17%) indique un refroidissement accentué du climat et une aridité plus importante qu’actuellement. Cette couche correspond à l’épisode climatique le plus rude de toute la séquence analysée avec, cependant, une courte période de réchauffement modéré comme le suggère la présence de Clethrionomys glaréoles (2 %) caractéristique d’un environnement forestier ou arbustif.
Couche F
Il représente l’épisode le plus chaud et le plus humide de toute la séquence analysée. On remarque que Lagurus lagurus reste toujours dominant (62%) mais en légère diminution par rapport à ce que l’on observe dans la couche E. La

83
Recherches croisées en Dobrogea
relative abondance de Microtus arvalis (25%) et de Spalax leucodon indique un climat assez comparable à l’actuel.
6. LES GRANDS MAMMIFÈRES
6.1. Taxonomie
On a identifié 15 espèces de grands mammifères: Ursus spelaeus, U. arctos, Capra/Ovis, Megaceros giganteus, Saiga tatarica, Rangifer tarandus, Bos primigenius, Alopex lagopus, Vulpes corsac, Coelodonta antiquitatis, Cevus elaphus, Equus przewalskii, Hydruntinus hydruntinus, Martes martes, Mustela nivalis. Le matériel prélevé est riche (plus de 1000 fragments) mais mal conservé. La fragmentation des pièces osseuses est due à la présence de l’Hyène. Par contre, aucun reste appartenant à l’Hyène n’a été trouvé.
Les taxons présents sont représentatifs d’un écosystème de climat froid et sec, de type steppe-toundra notamment avec la présence de Saiga tatarica, Equus sp., Rangifer tarandus, Coelodonta antiquitatis [7, 8]. L’échantillon est cependant faible et il est difficile de séparer les niveaux sur la base d’associations de faune.
6.2. Action des carnivores
La présence de traces de carnivores sur de nombreux ossements de mammifères de taille moyenne à grande exclut l’action des mustélidés et des renards identifiés dans l‘assemblage osseux. En revanche, le loup et l’ours des cavernes sont des agents potentiels d’accumulation d’individus mammaliens. Les stigmates de modification ne sont pas seulement localisés sur les extrémités d’os longs mais incluent aussi des rayures et des perforations sur les diaphyses d’os longs. Avec la présence de bords crénelés et de traces de digestion, ces stigmates laissent penser à une action de hyénidés ou de canidés [9, 10].
6.3. Traces d’activités anthropiques
Un seul spécimen osseux a livré des traces d’activité humaine, sous forme de stries de boucherie. Il s’agit de l’extrémité proximale, incomplète, d’une côte droite, attribuée à un taxon de mammifère de taille moyenne, issue de la couche E. Les stries sont localisées sur la surface externe de la côte, dans une région à forte densité d’attaches musculaires et ligamentaires. Elles correspondent probablement à une action de désarticulation des côtes et des organes associés (muscles, graisses) depuis le rachis.
Aucun spécimen osseux ne montre de traces de combustion.

84
6.4. Discussion
La présence de restes d’ours des cavernes de tous âges indique vraisemblablement l’utilisation de la grotte comme un lieu d’hivernage et d’élevage des oursons. Les ossements d’ours des cavernes ont ainsi dû être accumulés par mort naturelle, sans processus de prédation, d’autant que peu de traces de modification des surfaces osseuses par des carnivores ont été observées.
La présence d’ongulés divers est sans doute à mettre au crédit des loups et / ou des hyènes. Si les hyènes n’ont laissé aucun vestige osseux, elles sont présentes sur le site selon les études anciennes [3].
L’homme est donc un agent d’accumulation mineur.
7. INTERPRÉTATION CHRONOLOGIQUE ET DISCUSSIONS
La présence de plusieurs paléosols dont la pédogénese témoigne de conditions interglaciaires (D1, et E1 ; cf. Balescu, ce volume) constitue des repères chronostratigraphiques. Le paléosol D1, observable au sommet du loess E qui a donné un âge IRSL de 151 + 13 ka est attribuable au Dernier Interglaciaire (MIS 5.5, 128 à 115 ka). Le complexe pédologique observable dans une cuvette (C3 à D0) représente le bilan du début du Dernier Glaciaire marqué par l’alternance de plusieurs améliorations et péjorations climatiques. L’âge de la partie supérieure de la séquence pourra être précisé lorsque la datation IRSL du lambeau de sédiment loessique observable entre la couche C et la couche B (plaquettes calcaires) sera achevée.
L’étendue chronologique des sédiments de la Grotte La Adam, suggérée par son contenu faunistique [2, 3], est confirmée et précisée.
La grotte ne domine actuellement le ruisseau Vistorna que de quelques mètres en raison d’un colmatage de la vallée à la fin du Dernier Glaciaire ou au début de l’Holocène. Il en était tout autrement durant les derniers 300 millénaires où l’accès de l’entrée de la grotte était plus difficile ce qui explique qu’elle servit de repère pour les carnivores. Cependant, il existe de nombreux exemples dans les Carpates, notament, où les grottes furent fréquentées à la fois par les carnivores et par les Hommes [11]. Le caractère très discret de la présence humaine à La Adam est peut-être en partie imputable aux méthodes de fouilles des paléontologues qui travaillèrent dans la grotte sans se focaliser sur les témoignages laissés par les Hommes. Les recherches récentes, qui ne constituent qu’un test de réévaluation de cette grotte, n’ont pas concerné un volume suffisant de sédiments pour permettre d’avancer des explications satisfaisantes.

85
Recherches croisées en Dobrogea
REMERCIEMENTS
Les recherches effectuées à La Adam on été menées dans le cadre d’une collaboration entre la mission archéologique « Le Paléolithique de Roumanie » (Ministère des Affaires étrangères et européennes) et l’Institut d’archéologie « Vasile Pârvan ».
RÉFÉRENCES
1. Păunescu, Al., 1999, Paleoliticul şi mezoliticul de pe teritoriul Dobrogei. Bucureşti: Satya Sai, 241 p.
2. Petculescu, A., Samson, P. M., 2001, Aspecte climatice ale ultimuliu ciclu glaciar, bazate pe asociaţiile de micromamifere din carstul Dobrogei, Ecocarst, 2.
3. Samson, P. M., 1971, La stratigraphie du Quaternaire supérieur en Roumanie. Etudes sur le Quaternaire dans le Monde, VIIIe Cong. INQUA, Paris, 1969, 629-636.
4. Rădulescu, C., Samson, P. M., 1962, Sur un centre de domestication du Mouton dans la grotte La Adam en Dobroudja. Z. f. Tierzücht. u. Züchtungsbiol., 76, 282-320.
5. Petculescu, A., 2003. New data on the evolution of the Arvicola (Rodentia, Mammalia) from the karst of Romania. Theoretikal and Applied Karstology, 16, 99-103.
6. Terzea, E., 2000-2001, La faune de mammifères du Würm final de Peştera Bursucilor (comm. Târguşor, Dép. Constanţa, Roumanie). Remarques sur les espèces de lagurini et de Microtus. Trav. Inst. Spéol. E. Racovitza, Bucharest, 39-40, 155-192.
7. Griggo, G., 1996, Etablissement de courbes climatiques quantifiées à partir des communautés animales pléistocènes suivi d’une application aux gisements de l’abri Suard (Charente) et la grotte de Bois-Ragot (Vienne). Paléo, 8, 81-97.
8. Petculescu, A., Stiucă, E., 2008, Peculiarity of the mammal associations from the Upper Pleistocene (Dobrodgea, Romania). Quaternary International, 179, 79-82.
9. Haynes, G., 1983, A guide for differiating carnivore mammalian taxa responsible for gnow damage to herbivore limb bones. Paleobiology, 9, 2, 164-172.
10. Quiles, J., Petrea, C., Moldovan, O., Zilhao J., Rodrigo, R., Rougier, H., Constantin, S., Milota, S., Gherase, M., Sarcină, L., Trinkaus, E., 2006, Cave bears (Ursus spelaeus) from the Peștera cu Oase (Banat, România): paleobiology and taphonomy. C.-R. Paleovol., 5, 927-934.
11. Cârciumaru, M., 1999, Le Paléolithique en Roumanie, Grenoble, 330 p.

Planche 1. A. Grotte La Adam. B. Grotte La Adam. Fouille du secteur « entrée de grotte » (EG).