transcriptional profiling of androgen receptor (ar) mutants suggests instructive and permissive...
TRANSCRIPT

Transcriptional Profiling of Androgen Receptor (AR)Mutants Suggests Instructive and Permissive Rolesof AR Signaling in Germ Cell Development
Stephen M. Eacker, James E. Shima, Charles M. Connolly, Manju Sharma, Robert W. Holdcraft,Michael D. Griswold, and Robert E. Braun
Department of Genome Sciences (S.M.E., C.M.C., M.S., R.W.H., R.E.B.), School of Medicine,University of Washington, Seattle, Washington 98195-5065; and School of Molecular Biosciences(J.E.S., M.D.G.), Washington State University, Pullman, Washington 99164
The androgen receptor (AR) is a transcription fac-tor that plays a critical role in male sexual devel-opment, spermatogenesis, and maintenance ofhormonal homeostasis. Despite the extensiveknowledge of the phenotypic consequences ofmutations in Ar, very little is known about the tran-scriptional targets of AR within the testis. To iden-tify potential targets of androgen signaling in thetestis, we have analyzed the transcriptional profileof adult testes from Ar hypomorphs alone or incombination with Sertoli cell-specific Ar ablation.Using Affymetrix MOE430A mouse genome arrayswe interrogated more than 22,000 transcripts. Wefound the expression level of 62 transcripts in theAr mutants differed by greater than 2-fold com-pared with wild type. We also found that moretranscripts were up-regulated than down-regu-lated, highlighting AR’s role as a transcriptionalrepressor in the testis. Twelve transcripts wereuniquely affected, and 16 transcripts were more
severely affected in Sertoli cell-specific Ar ablationcompared with hypomorphic Ar mutants. Using acomparative genomic approach, we analyzed the 6kb around the transcriptional start sites of affectedtranscripts for conserved AREs (androgen re-sponse elements). We identified at least one con-served ARE in 65% of the genes misregulated inour microarray analysis where clear mouse-humanorthologs were available. We used a reporter assayin cell culture to functionally verify the AREs for thekallikrein 27 gene. This suggests that the majorityof the misregulated transcripts have a high proba-bility of being direct AR targets. The transcriptsaffected by these Ar mutations encode a diversearray of proteins whose molecular functions sup-port the contention that AR supports spermato-genesis in both a permissive and instructivefashion. (Molecular Endocrinology 21: 895–907,2007)
THE ANDROGEN RECEPTOR (AR) is a member ofthe nuclear hormone receptor family of ligand-
dependent transcription factors. AR plays a criticalrole in the development of the testis and in mainte-nance of spermatogenesis in adult mammals. Duringdevelopment, testosterone (T) and dihydrotestoster-one (DHT) act through AR to promote testicular de-scent and Leydig cell maturation. Extensive work inthe rat has demonstrated that T is the critical hormonethat supports spermatogenesis (1). A variety of models
have been used to study the effects of T withdrawal onspermatogenesis. Many studies have used hypophy-sectomy to ablate gonadotropin secretion, and con-sequently T production (2, 3). Other studies usingtestosterone-estrogen (TE) pellets and selective de-struction of Leydig cells by ethanedimethane sulfonate(EDS) also have been extremely useful in dissectingthe steps of spermatogenesis, which require T signal-ing. These studies have largely concluded that T isessential for maintenance of spermatogenesis, al-though FSH is required for maximal sperm output (4).
AR is expressed in the somatic cells of the testis: theSertoli cells, which make intimate contacts with germcells, Leydig cells located within the interstitium, andperitubular myoid cells, which encompass the semi-niferous tubule (5). Extensive data support a role for Tin meiotic progression, retention of round spermatids,the transition of round spermatids to elongating sper-matids, and the release of mature sperm (6–9). The roleof T in germ cell differentiation is mediated in large partby expression of AR in the Sertoli cell (10–12). Althoughthese studies have been crucial in the identification of thecell type where AR is required for germ cell maturation,they have not yet defined the mechanism by which AR
First Published Online January 23, 2007Abbreviations: AR or Ar, Androgen receptor; ARE, andro-
gen response element; 8-Br-cAMP, 8-bromo-cAMP; BTB,blood-testis-barrier; cAREs, conserved AREs; Cyp17a1, cy-tochrome P450 17a1; DHT, dihydrotestosterone; EDS,ethanedimethane sulfonate; Hsd17b3, hydroxysteroid 17�dehydrogenase 3; Lhcgr, LH/chorionic gonadotropin recep-tor; LLR, log-likelihood ratio; MMTV-luc, mouse mammarytumor virus-luciferase; PSA, prostate-specific antigen; RA,retinoic acid; T, testosterone; TE, testosterone-estrogen; TP,testosterone propionate; TSS, transcriptional start site.
Molecular Endocrinology is published monthly by TheEndocrine Society (http://www.endo-society.org), theforemost professional society serving the endocrinecommunity.
0888-8809/07/$15.00/0 Molecular Endocrinology 21(4):895–907Printed in U.S.A. Copyright © 2007 by The Endocrine Society
doi: 10.1210/me.2006-0113
895

supports germ cell development. One possibility mightbe that AR acts in an instructive fashion, affecting theexpression of signals that directly support germ cell mat-uration. Another possibility is that AR is required to cre-
ate a permissive microenvironment within the seminifer-ous epithelium that allows for the progression of germcell development. Identification of AR targets may helpdistinguish between these two models.
896 Mol Endocrinol, April 2007, 21(4):895–907 Eacker et al. • Testicular Gene Expression in Ar Mutants

The function of AR in the Leydig cell has been thetopic of a number of investigations. Work on the spon-taneous Ar mutant Artfm/Y has shown that AR is re-quired for the normal development of adult-type Ley-dig cells (13). The Artfm/Y mice fail to express keysteroidogenic genes such as cytochrome P450 17a1(Cyp17a1) and hydroxysteroid (17�) dehydrogenase 3(Hsd17b3). The absence of these enzymes in Leydigcells prevents high levels of T synthesis. Although anumber of genes encoding steroidogenic enzymes aredown-regulated in wild-type animals surgically ren-dered cryptorchid, it is clear that their expression ismuch lower in Artfm/Y mice. However, it is not clearwhether transcription of these steroidogenic genesrequires Ar function directly. In fact, a large body ofevidence has shown AR can act as a negative regu-lator of Cyp17a1 in Leydig cells (14, 15). It has beenshown that AR binds to the promoter of Cyp17a1 andcan repress its expression (16). These seemingly contra-dictory data can be resolved by the proposed dual role ofAR as a positive regulator of Leydig cell developmentand as a negative regulator of steroidogenesis (13, 17).
We have developed a unique model to investigateAR function in the testis of Mus musculus (10). Ahypomorphic conditional allele of Ar was created byplacing a neomycin resistance cassette within intron 1of Ar and flanking exon 1 with inverted loxP sites. Acryptic splice acceptor in the neomycin resistancecassette results in stochastic splicing with exon 1,thereby reducing the amount of full-length Ar mRNA.This hypomorphic allele, termed Arinvflox(ex1-neo), re-sults in a partial failure of terminal spermatid differen-tiation accompanied by elongating spermatid phago-cytosis and a 95% reduction in epididymal spermnumbers. Using Cre recombinase driven by the Sertolicell-specific Amh promoter [Tg(Amh-Cre)], we ablatedAr from Sertoli cells in an otherwise hypomorphic Arbackground. This Sertoli cell-specific ablation resultsin a more severe phenotype characterized by near-complete failure in elongating spermatid differentiationand severe round spermatid sloughing. In the hope ofidentifying the AR targets responsible for these phe-notypes, we compared the gene expression profiles ofadult whole testis mRNA from wild-type, hypomorphicAr mutants, and hypomorphic Ar mutants with Sertolicell-specific Ar ablation. Using a computational ap-proach, we determined which of the genes misregulatedin Ar mutants contain consensus androgen-responsiveelements (AREs) that are conserved between mice andhumans, and thus may be direct targets of AR action.
RESULTS
Identification of Genes Misregulated in Testesof Hypomorphic and Sertoli Cell-SpecificAr Mutants
In an effort to identify AR-regulated genes involved inspermatogenesis, we conducted an expression array
analysis of mRNA from testes of 8-wk-old wild-type,Arinvflox(ex1-neo)/Y, and Arinvflox(ex1-neo)/Y; Tg(Amh-Cre)mice. The hypomorphic nature of Arinvflox(ex1-neo)/Y leadsto a reduction of AR in all cells in the animal includingthose in the testis (10). Therefore, we expected that anal-ysis of gene expression in Arinvflox(ex1-neo)/Y testis wouldreveal AR targets in all of the somatic cell types in thetestis. We also expected that some gene expression ingerm cells may be affected because of changes inparacrine signaling between the soma and germ line.When AR expression is ablated from Sertoli cells bythe addition of Tg(Amh-Cre), we expected that somegenes that are misregulated in Arinvflox(ex1-neo)/Y wouldbe more severely affected. In many cases, this shouldreflect changes in gene expression in Sertoli cells.
Transcripts that demonstrated greater than 2-foldchange compared with wild-type are listed in Table 1.In the testes of Arinvflox(ex1-neo)/Y, 31 transcripts wereup-regulated and 15 down-regulated compared withwild type. When Arinvflox(ex1-neo)/Y; Tg(Amh-Cre) testismRNA was profiled, the levels of 40 transcripts were in-creased and 17 decreased relative to wild type. As dis-cussed above, the differences between Arinvflox(ex1-neo)/Y
and Arinvflox(ex1-neo)/Y; Tg(Amh-Cre) likely reflect changesin Sertoli cell-specific gene expression. In all, nine tran-scripts were up-regulated and three transcripts down-regulated in Arinvflox(ex1-neo)/Y; Tg(Amh-Cre) but not inArinvflox(ex1-neo)/Y. Interestingly, there was one transcriptuniquely up-regulated and four uniquely down-regulated inArinvflox(ex1-neo)/Y but not Arinvflox(ex1-neo)/Y; Tg(Amh-Cre).
To validate results obtained from our microarrayanalysis, we performed northern analysis for selectedtranscripts. Both Arinvflox(ex-neo)/Y and Arinvflox(ex1-neo)/Y;Tg(Amh-Cre) animals demonstrate elevated serum T(�40� over wild type), LH (�25� over wild type), andFSH (�3� over wild type) (10); these hormones canhave significant impacts on transcription in the testis.Therefore, we analyzed transcripts known to be regu-lated by T (Rhox5, Star), LH (Ren1, Star, Inha), and FSH(Ren1, Klf4, Inha) in addition to transcripts regulatedby unknown factors (Adh1, Klk27, Lcn2). Of the tran-scripts that were misregulated in our microarray anal-ysis, all (7/7) demonstrated similar magnitude ofchange by Northern blot (Fig. 1). With this sample ofNorthern blots, we validated the results for 11% of thetranscripts misregulated in our microarray analysis. Asindicated by our analysis, the level of Rhox5 messagewas reduced in Arinvflox(ex1-neo)/Y and Arinvflox(ex1-neo)/Y;Tg(Amh-Cre), consistent with a reduction in AR sig-naling. T is known to be a negative regulator of Starexpression, whereas LH is a positive regulator. Ac-cordingly, Star mRNA was also up-regulated in bothmutants as predicted by our microarray analysis. Thelevels of neither Ren1 nor Klf4 appeared to be alteredin our microarray analysis. However, despite its ab-sence from the list of misregulated transcripts, Ren1was up-regulated by Northern blot (Fig. 1). The vari-ability observed in the levels of Ren1 message, asseen by Northern, could have resulted in exclusionfrom the list of 2-fold altered transcripts. In summary,
Eacker et al. • Testicular Gene Expression in Ar Mutants Mol Endocrinol, April 2007, 21(4):895–907 897

these data provide evidence for changes in LH and ARsignaling, but no clear perturbation in FSH signaling.The consistency between the results of the microarrayanalysis and Northern blot validation give us a highdegree of confidence in the results listed in Table 1.
The transcripts misregulated in the testes ofArinvflox(ex1-neo)/Y and Arinvflox(ex1-neo)/Y; Tg(Amh-Cre)encode proteins encompassing a broad range of mo-
lecular functions (Table 1). These molecular functionswere assigned based on their gene ontology annota-tions provided by The Jackson Laboratory Mouse Ge-nome Informatics (http://www.informatics.jax.org). Agraphical representation of the molecular functions ofmisregulated transcripts with known function clearlydemonstrates an overrepresentation of genes involvedin metabolic processes (Fig. 2). The transcripts encod-ing metabolic proteins are dominated by genes in-volved in cholesterol biosynthesis and steroidogene-sis. The next most abundant class of protein functionsis involved in signal transduction. Most striking amongthe signal transduction genes is the 14- to 17-foldup-regulation of the LH/chorionic gonadotropin recep-tor (Lhcgr, Table 1). Also of note is the significantup-regulation of inhibin � (Inha) mRNA. After signaltransduction-related transcripts, proteases were mostseverely affected. Among these transcripts are fourmRNAs encoding members of the kallikrein family ofproteases. Data from previous studies and the grow-ing information from cell type-specific microarray data(18) have allowed us to assign the cell types in which
Fig. 1. Verification of Microarray ResultsNorthern analysis of wild-type, Arinvflox(ex1-neo)/Y, Arinvflox(ex1-neo)/Y;
Tg(Amh-Cre) whole testis RNA from 8-wk-old animals. Se-lected transcripts that were up-regulated (Adh1, Inha, Star)and down-regulated (Rhox5, Lcn2, Klk27) were chosen fortheir known regulation patterns or for scientific interest.Rhox5 and Star are known to be under positive regulation byandrogen and LH signaling, respectively. Klf4 is a FSH-reg-ulated transcript and showed no change in expression level.Although not identified on the microarray analysis, Ren1, aFSH- and LH-regulated transcript was up-regulated in bothAr mutants. Actb was used as a loading control. These resultsare representative of results from Northern analyses of threeto six individuals of each genotype.
Fig. 2. Classification of Molecular Functions of Proteins En-coded by Misregulated Transcripts in Arinvflox(ex1-neo)/Y andArinvflox(ex1-neo)/Y; Tg(Amh-Cre)
A single molecular function was assigned for each tran-script based on its gene ontology annotation provided by theMouse Genome Informatics web site (http://www.informatics.jax.org). Transcripts associated with metabolic functions inthe testis, primarily steroidogenesis and cholesterol biosyn-thesis, dominate the collection of misregulated transcripts.The next most abundant classification of transcripts is thoseof unknown function. Addition of the Tg(Amh-Cre) to thehypomorphic Arinvflox(ex1-neo)/Y background results in an in-crease the proportion of misregulated transcripts involved incell adhesion, nucleic acid metabolism, and transcription.The transcripts comprising each functional classification arelisted in Table 1.
898 Mol Endocrinol, April 2007, 21(4):895–907 Eacker et al. • Testicular Gene Expression in Ar Mutants

the misregulated transcripts are normally expressed(Table 1).
Identification of Potential AREs
Identifying transcripts altered in our microarray analy-sis as being androgen regulated must be carefullyconsidered because of the complex endocrine pheno-type of Arinvflox(ex1-neo)/Y. High levels of FSH and LHcould obscure the relative contribution of the absenceof AR to the observed transcriptional phenotype. ARinteracts with DNA sequences termed AREs that con-sist of two 6-bp half-sites separated by a 3-bp spacer.The sequence of these AREs has been experimentallydetermined in a variety of androgen-responsive pro-moters, many of which are listed in Table 2. The highdegree of degeneracy at many of the sites within the6-bp half-sites has made de novo prediction of AREsin promoter regions a difficult task. Also, studies usingin vitro selection for sequences that bind the AR DNAbinding domain have identified high-affinity se-quences that are found rarely, if at all, in nature (19). Tohelp identify transcripts that were more likely to beandrogen-regulated, we examined 6000 bp encom-passing the transcriptional start site (TSS) of eachgene listed in Table 1. Of the 62 genes listed in Table1, 59 genes had good evidence identifying the TSSand were therefore retained for further analysis. Usingsequences listed in Table 2 we generated a weight-matrix model for AREs and used it to identify potentialAREs with a log-likelihood ratio (LLR) � 3. A graphicalrepresentation of the weight matrix used in this anal-ysis is shown in Fig. 3A. The results of this analysis anddetails of the method are supplied in SupplementalFig. 1 and Supplemental Table 1 (published as sup-plemental data on The Endocrine Society’s JournalsOnline web site http://mend.endojournals.org/). Using
these criteria, we identified 3689 potential AREs in the6 kb encompassing the TSS for 59 genes.
To identify AREs that have a higher probability ofbeing functional, we identified potential AREs con-served over the course of evolution. We examined6000 bp of sequence encompassing the TSS of thehuman orthologs of the genes identified in Table 1. Weused information provided in the “Mammalian Homol-ogy” section of Mouse Genome Informatics web site(http://www.informatics.jax.org) to identify the humanorthologs of genes affected in our microarray analysis.The total number of genes with clear human orthologsreduced the number of promoter regions to 49 fromour original 59 sequences. Using aligned mouse andhuman sequences, we used MONKEY (20) to identifyconserved AREs (cAREs). We identified 108 potentialcAREs within the 6 kb surrrounding the TSS (Supple-mental Table 2). The distribution of these cAREs rela-tive to TSS are shown in Fig. 3B. Most genes had atleast one cARE, but 35% had no identifiable cAREs(Fig. 3C). Positions of the cAREs in the promoter re-gions of validated transcripts with clear human or-thologs are illustrated in Fig. 3D. As a negative controlwe performed the same computational analysis on twogenes that are expressed in the testis but are notunder androgen regulation, Protamine 1 (Prm1), andtestis nuclear RNA binding protein (Tenr). Neither ofthese genes contained a conserved ARE.
Expression of Four Members of the KallikreinFamily of Proteases Is Disrupted by Loss of AR
The Kallikrein family of serine proteases reside in acluster on mouse chromosome 7 (Fig. 4A). This familyhas a dynamic evolutionary history that includes nu-merous species-specific expansions and contractions(21). Interestingly, two human Kallikrein family mem-
Table 2. Experimentally Validated Androgen Response Elements
Promoter Position (bp) Sequence Ref.
Probasin ARE1 �243 to �228 ATAGCATCTTGTTCT 53Probasin ARE2 �137 to �122 AGTACTCCAAGAACC 53PSA ARR �390 to �375 GGATCAGGGAGTCTC 54PSA ARE �167 to �154 AGAACAGCAAGTGCT 54KLK2 �157 to �144 GGAACAGCAAGTGCT 55C3-intron �1359 to �1374 AGTACGTGATGTTCT 56C1 �207 to 222 AGGACACAAAAATCC 56Slp ARE 1 �112 to �127 GTAATTATCTGTTCT 57Slp ARE 2 �128 to �143 TGGTCAGCCAGTTCT 57Slp ARE 3 �144 to �159 AGAACAGGCTGTTTC 57Factor IX �37 to �22 AGCTCAGCTTGTACT 58JRE �111 to �96 ATTACACCAAGTACC 59Aldose reductase �111 to �96 TGAAGTTCCTGTTCT 60GUS �7833 to �7848 AGTACTTGTTGTTCT 61AR-ARE1 �2378 to �2393 CTTTCTGAATGTCCT 62AR-ARE2 �2576 to �2591 AGTACTCCTGGATGG 62Pem ARE2 �247 to �233 AGCACATCGTGCTCA 63PSCA ARE1 �2796 to �2782 AGGACGGAAAGTTCC 64
Many of the AREs collected in this table were collected in Ref. 19.
Eacker et al. • Testicular Gene Expression in Ar Mutants Mol Endocrinol, April 2007, 21(4):895–907 899

bers, prostate-specific antigen (PSA) and KLK2, arewell studied for their tissue-specific androgen respon-siveness. Because androgen responsiveness is acommon feature of members of the Kallikrein family,and four Klks were misregulated in our microarrayanalysis, Klks seem to be a good candidate for directandrogenic regulation. Indeed, the 2-kb encompass-ing the TSS of the four Klks affected contain numerouspotential AREs (Fig. 4B). To test whether Klks respondto androgen signaling, we treated MA-10 Leydig tumorcells with DHT. MA-10 cells are an easily manipulatedcell culture model of murine Leydig cells. After treat-ment, total RNA from the cell lines was analyzed byNorthern blot (Fig. 4C). MA-10 cells alone or with 50�M 8-bromo-cAMP (8-Br-cAMP) did not exhibit signif-icant levels of Klk expression. However, when treatedwith 10 �M DHT, MA-10 cells expressed detectablemessage. Surprisingly, addition of 8-Br-cAMP to DHT-treated cells reduced the level of Klk27 mRNA. Al-though the probe used in this hybridization is from a
Klk27 cDNA, the high degree of sequence identitybetween Klk-family members makes it impossible todetermine which Klk members are being induced byDHT.
To determine whether the consensus AREs we iden-tified in the promoter region of Klk27 confer androgenresponsiveness, we fused 1.3 kb of sequence up-stream of the Klk27 TSS to a luciferase reporter (Klk27-luc). When MA-10 Leydig tumor cells transfected withKlk27-luc are stimulated with DHT, we observed a3.4-fold increase in reporter activation, demonstratingandrogen-dependent activation (Fig. 4D). MA-10 cellsexpress low levels of AR, as demonstrated by insig-nificant androgen-dependent activation of a mousemammary tumor virus-luciferase (MMTV-luc) reporter(Fig. 4D) (16). However, upon cotransfection of MMTV-luc with an AR expression construct, we observed an8-fold increase in luciferase activity. Similarly, whenMA-10 cells were cotransfected with Klk-luc and anAR expression vector and treated with DHT we ob-
Fig. 3. Computational Identification of cAREs in the 6 kb Surrounding the Transcriptional Start Site of Misregulated TranscriptsA, A logo representation of the frequencies of bases at each position in experimentally validated AREs used to generate the
weight matrix model of AR binding sites. The heights of the letters represent the relative frequency of a nucleotide at each positionin the ARE. The logo was generated using the sequences in Table 2 with WebLogo (http://weblogo.berkeley.edu/logo.cgi). Thedistribution of sequences conserved between mouse and human in the promoters of genes misregulated in Arinvflox(ex1-neo)/Y andArinvflox(ex1-neo)/Y; Tg(Amh-Cre) are illustrated in panel B. Details of method used to identify the cAREs are described in Materialsand Methods. C, The number of cAREs identified in the promoters of genes analyzed in (B). D, A diagram of the location andorientation of cAREs in misregulated genes validated in Fig. 2. Large black boxes indicate exons and a black arrow indicates thetranscriptional start site (TSS). The gray arrows indicate the position of cAREs as defined in panel A. Right-pointing arrows indicatethe cARE is on the (�) strand, whereas left-pointing arrows indicate the cARE is on the (�) strand.
900 Mol Endocrinol, April 2007, 21(4):895–907 Eacker et al. • Testicular Gene Expression in Ar Mutants
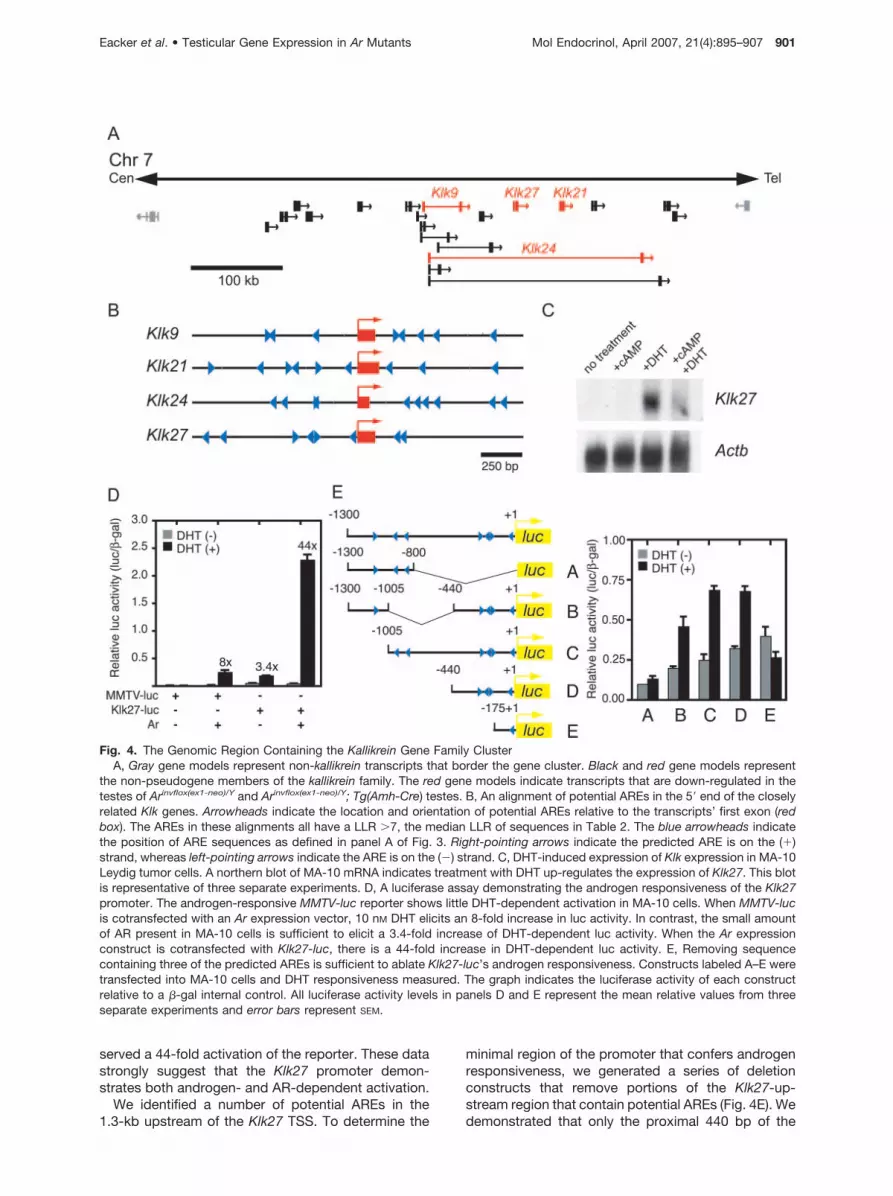
served a 44-fold activation of the reporter. These datastrongly suggest that the Klk27 promoter demon-strates both androgen- and AR-dependent activation.
We identified a number of potential AREs in the1.3-kb upstream of the Klk27 TSS. To determine the
minimal region of the promoter that confers androgenresponsiveness, we generated a series of deletionconstructs that remove portions of the Klk27-up-stream region that contain potential AREs (Fig. 4E). Wedemonstrated that only the proximal 440 bp of the
Fig. 4. The Genomic Region Containing the Kallikrein Gene Family ClusterA, Gray gene models represent non-kallikrein transcripts that border the gene cluster. Black and red gene models represent
the non-pseudogene members of the kallikrein family. The red gene models indicate transcripts that are down-regulated in thetestes of Arinvflox(ex1-neo)/Y and Arinvflox(ex1-neo)/Y; Tg(Amh-Cre) testes. B, An alignment of potential AREs in the 5� end of the closelyrelated Klk genes. Arrowheads indicate the location and orientation of potential AREs relative to the transcripts’ first exon (redbox). The AREs in these alignments all have a LLR �7, the median LLR of sequences in Table 2. The blue arrowheads indicatethe position of ARE sequences as defined in panel A of Fig. 3. Right-pointing arrows indicate the predicted ARE is on the (�)strand, whereas left-pointing arrows indicate the ARE is on the (�) strand. C, DHT-induced expression of Klk expression in MA-10Leydig tumor cells. A northern blot of MA-10 mRNA indicates treatment with DHT up-regulates the expression of Klk27. This blotis representative of three separate experiments. D, A luciferase assay demonstrating the androgen responsiveness of the Klk27promoter. The androgen-responsive MMTV-luc reporter shows little DHT-dependent activation in MA-10 cells. When MMTV-lucis cotransfected with an Ar expression vector, 10 nM DHT elicits an 8-fold increase in luc activity. In contrast, the small amountof AR present in MA-10 cells is sufficient to elicit a 3.4-fold increase of DHT-dependent luc activity. When the Ar expressionconstruct is cotransfected with Klk27-luc, there is a 44-fold increase in DHT-dependent luc activity. E, Removing sequencecontaining three of the predicted AREs is sufficient to ablate Klk27-luc’s androgen responsiveness. Constructs labeled A–E weretransfected into MA-10 cells and DHT responsiveness measured. The graph indicates the luciferase activity of each constructrelative to a �-gal internal control. All luciferase activity levels in panels D and E represent the mean relative values from threeseparate experiments and error bars represent SEM.
Eacker et al. • Testicular Gene Expression in Ar Mutants Mol Endocrinol, April 2007, 21(4):895–907 901

upstream sequence are required for androgen-depen-dent activation (Fig. 4E, construct D) and when thesequence containing the three predicted AREs is re-moved, the androgen responsiveness of the Klk27promoter is eliminated (Fig. 4E, construct E). This sug-gests that these computationally predicted AREs arefunctional in the context of the Klk27 promoter.
DISCUSSION
Despite the absolute requirement of androgen signal-ing for spermatogenesis, very few direct targets of ARin the testis are known. In our current study, we havecharacterized the transcriptional profile of the testes ofadult animals that bear a hypomorphic allele of Ar aloneor in combination with Sertoli cell-specific ablation of Ar.We identified a total of 62 transcripts that were alteredgreater than 2-fold compared with wild-type in the testesof Arinvflox(ex1-neo)/Y and Arinvflox(ex1-neo)/Y; Tg(Amh-Cre)mice. The majority of these transcripts contain evolution-arily conserved androgen response elements, suggest-ing that many of these transcripts are direct targets ofAR. For one of these transcripts, Klk27, we demon-strated that computationally predicted AREs in thepromoter region are functional in a Leydig-cell culturemodel. We have therefore greatly expanded the poolof potential AR targets in the testis, providing novelinsight into possible mechanisms by which T pro-motes spermatogenesis.
These animals provide significant advantages forthe identification of targets of AR signaling in the wholetestis as well as specifically in Sertoli cells. UnlikeArtfm/Y mice, Arinvflox(ex1-neo)/Y and Arinvflox(ex1-neo)/Y;Tg(Amh-Cre) mice have normally descended testes.Cryptorchidism alone has a strong effect on testiculargene expression making the use of Artfm/Y to identifytargets of androgen signaling problematic (13). Addi-tionally, unlike Artfm/Y and other Sertoli cell-specific Armutant mice, Arinvflox(ex1-neo)/Y and Arinvflox(ex1-neo)/Y;Tg(Amh-Cre) animals have a nearly intact complementof transcriptionally active germ cells. Therefore,changes in gene expression observed in our analysisare not the result of a lack of major classes of germcells within the testis. The presence of particularclasses of germ cells are also required for the produc-tion proteins synthesized in Sertoli cells, making thenearly normal complement of germ cells even moreimportant to our analysis (22).
The best-characterized target of AR signaling withinthe testis is Rhox5 (formerly known as Pem), a ho-meobox gene located on the X chromosome that isexpressed exclusively in Sertoli cells. Our microarrayanalysis demonstrated the expected down-regulationof Rhox5 (Table 1) that was confirmed by Northern blot(Fig. 1). It has been shown that Rhox5 is responsive toandrogens and that androgen-induced transcriptionrequires two AREs located �233 and �69 relative tothe transcriptional start site (23, 24). Male mice bearing
a null mutation of Rhox5 are subfertile but do not havesevere defects in spermatogenesis (25, 26). Becauseandrogen withdrawal has a far more severe phenotypethan mutation of Rhox5, this clearly suggests thatRhox5 is only one of several targets of AR required tosupport spermatogenesis.
Two recent investigations have used pharmacolog-ical treatment coupled with microarray expressionanalysis to identify transcripts that are regulated byandrogens in an in vivo system. To isolate the effectsof testosterone alone on testis gene expression, Sa-date-Ngatchou and colleagues (28) used the sponta-neous Gnrh mutant Gnrhhpg/hpg treated with testoster-one propionate (TP). This work on the Gnrhhpg/hpg
animal has the significant advantage of no confound-ing effects of LH and FSH on gene regulation duringthe treatments with TP. Total testis RNA was collectedafter short-term treatment with TP and microarray anal-ysis revealed a small number of transcripts (234) thatchange relative to controls. This is consistent with ourobservation that a relatively small number of transcriptswere significantly altered in both Arinvflox(ex1-neo)/Y andArinvflox(ex1-neo)/Y; Tg(Amh-Cre) testis. Also consistentwith our observations, more transcripts were down-regulated than up-regulated with testosterone treat-ment. Our data further support the suggestion that ARfunctions primarily as a mediator of transcriptional re-pression within the adult testis. A second study toidentify AR-regulated transcripts used wild-type pre-pubertal animals treated with TP (27). The prepubertalanimals lack a functional HPG axis, thus removing theconfounding effects of FSH and LH. In contrast to ourfindings and those in TP-treated adult Gnrhhpg/hpg
mice, prepubertal animals treated with TP show aroughly equal number of up- and down-regulated tran-scripts. One intriguing possibility is that the differencein response to androgens in the prepubertal animalreflects a developmental change in the profile of ARcoregulators in the testis. Rhox5 is also up-regu-lated by TP in prepubertal mice, making it the onlycommonly regulated gene between the TP-treatedGnrhhpg/hpg mice (28), TP-treated prepubertal mice,and the current study.
Two other groups have published studies of adultSertoli-selective Ar knockouts (11, 12). With the ex-ception of Rhox5, there is no overlap in published geneexpression changes associated with these mutationsand Arinvflox(ex1-neo)/Y;Tg(Amh-Cre). Certainly, this ispartly due to the relatively small changes observed forthe transcripts these groups have selected for analy-sis; we have chosen a 2-fold or greater change cut-offfor the transcripts we have analyzed. In a recent studyof prepubertal Sertoli cell-specific Ar mutant(SCARKO) mice, the authors identified 12 up-regu-lated and 28 down-regulated transcripts at a 2-fold orgreater cut-off (29). Of these transcripts, only Rhox5was commonly regulated gene between our study andthat of Denolet et al. (29). There are several possiblereasons for the lack of common transcriptionalchanges between the two studies. Prepubertal Sertoli
902 Mol Endocrinol, April 2007, 21(4):895–907 Eacker et al. • Testicular Gene Expression in Ar Mutants

cells are mitotically active, whereas adult Sertoli cellsare mitotically quiescent, prepubertal Sertoli cells arenot yet surrounded by the full-complement of germ cells,and AR coactivator and corepressor expression may bedifferent between prepubertal and adult Sertoli cells.
Determining which of the misregulated transcriptsare regulated directly by AR is a challenging problem.To address this question we chose to take a bioinfor-matic approach to identify evolutionarily conservedsequences in the promoter regions of the genes, weidentified as misregulated in Ar mutants. We identified108 cAREs in the 6 kb encompassing the transcrip-tional start site (TSS) of 49 of the misregulated genes.These cAREs show a modest enrichment in the �500-to �500-bp region relative to the TSS (Fig. 3A). This issignificant because many experimentally validatedAREs were found within this region (Table 2). Interest-ingly, there is a spike in the number of cAREs from�2501 to �3000 bp. The significance of these poten-tial regulatory sequences is unclear, although thepresence of AREs in this region are not without pre-cedent. As is true for many androgen-responsivegenes, most ARE-containing promoters have morethan one cARE (Fig. 3B). In general, AREs in androgen-responsive promoters function in a cooperative man-ner where each binding site contributes to a portion ofthe promoter’s responsiveness. Although the pres-ence of cAREs in the promoters of genes misregulatedin Ar mutants is not proof that they are direct targets ofAR, the localization of cAREs does provide fertileground for hypothesis testing.
A potential complication to our analysis is the highlevel of gonadotropins in Arinvflox(ex1-neo)/Y andArinvflox(ex1-neo)/Y; Tg(Amh-Cre). In both Arinvflox(ex1-neo)/Y andArinvflox(ex1-neo)/Y; Tg(Amh-Cre), there is an approximately25� increase in serum LH and an approximately 3�increase in FSH. As discussed previously, we believethat these increases reflect a lack of feedback regulationwithin the hypothalamic-pituitary-gonadal axis as a resultof a reduction in AR signaling. Indeed, many knowntargets of LH signaling are up-regulated in our analysis ofAr mutants, including transcripts encoding steroidogen-esis-related proteins. It is not clear to what extent, how-ever, the high levels of steroidogenic gene expressionare due to elevated LH levels or to the reduction of ARsignaling. For instance, Star is a well characterized targetof LH signaling and is up-regulated in Arinvflox(ex1-neo)/Y
and Arinvflox(ex1-neo)/Y; Tg(Amh-Cre). However, it has beenshown that T can suppress LH-induced Star transcrip-tion (30). Therefore, both the presence of high level ofgonadotropin and the low levels of AR signaling couldcontribute to the levels of gene expression observed inArinvflox(ex1-neo)/Y and Arinvflox(ex1-neo)/Y; Tg(Amh-Cre). Inaddition to the strikingly high levels of serum LH, weobserved a 14- to 17-fold increase in Lhcgr transcriptlevels. Although we do not know whether this up-regu-lation of Lhcgr mRNA results in an increase in Lhcgrprotein, it does suggest that there is an as-yet-unchar-acterized role for AR or LH signaling in the transcriptionof Lhcgr.
Despite the 3-fold increase in FSH levels, we ob-served no clear transcriptional effect that could beascribed to increases in FSH signaling in the testis ofArinvflox(ex1-neo)/Y and Arinvflox(ex1-neo)/Y; Tg(Amh-Cre)animals. To confirm this observation made from themicroarray expression profile, we measured levels ofKlf4 and Ren1 message, both of which are targets ofFSH signaling in vivo (31). We found no significantchange in the levels of Klf4. Although not borne out inthe microarray analysis, there was modest and vari-able up-regulation of Ren1 as determined by Northernblot (Fig. 1). However, Ren1 can be up-regulated byboth LH and FSH signaling (31, 32). Transcription ofInha is also thought to be stimulated by FSH and isindeed up-regulated in both Arinvflox(ex1-neo)/Y andArinvflox(ex1-neo)/Y; Tg(Amh-Cre) testes. However, ouranalysis of serum Inhibin B levels thus far has dem-onstrated no significant difference betweenArinvflox(ex1-neo)/Y, Arinvflox(ex1-neo)/Y; Tg(Amh-Cre),and wild-type animals (Eacker, S. M., unpublishedobservations). Therefore, it seems more likely thatthe extraordinarily high level of LH is stimulating thetranscription of Inha in Leydig cells, which is a well-documented phenomenon (33).
Regardless of being direct or indirect targets of AR,there are a number of interesting misregulated tran-scripts that may shed light on gonadotropin and ste-roid action in the testis. The Leydig cell-specific tran-script insulin-like 3 (Insl3) was down-regulated nearly10-fold in Arinvflox(ex1-neo)/Y. Similar observations havebeen made in an investigation of Leydig cell develop-ment in Artfm/Y mice (13). The authors in this studyconcluded that the misregulation of Insl3 was due afailure in Leydig cell maturation in Artfm/Y. Treatmentwith GnRH antagonists can also down-regulate Insl3in the adult male rat, suggesting that its transcription ishormonally regulated (34). Late-stage germ cell apo-ptosis observed in GnRH antagonist-treated rats is par-tially relieved when exogenous INSL3 is provided, sug-gesting that INSL3 can function to support germ celldevelopment. We have also observed late-stage germcell apoptosis in Arinvflox(ex1-neo)/Y and Arinvflox(ex1-neo)/Y;Tg(Amh-Cre) mice, which could in part be due to thelack of INSL3 production by the Leydig cell (Meng, J.,and R. E. Braun, unpublished observations). WhenhCG is provided to Gnrhhpg/hpg mice or to rats treatedwith a GnRH antagonist, Insl3 levels return to normal,suggesting either direct effects through LHCGR orsecondary effects through androgen production (34,35). Because LH levels in Arinvflox(ex1-neo)/Y mice arevery high, AR signaling is reduced, and Insl3 expres-sion is low, we suggest that Insl3 may be a target ofAR. Alternatively, additional factors acquired duringLeydig cell maturation may be required for properhormonally induced Insl3 expression.
As previously noted, a number of steroidogenic genesare severely down-regulated in the Artfm/Y mouse. Inparticular, Hsd3b6, Hsd17b3, and Cyp17a1 were signif-icantly down-regulated in adult Artfm/Y Leydig cells (17).This trend holds true in Arinvflox(ex1-neo)/Y for Hsd3b6 and
Eacker et al. • Testicular Gene Expression in Ar Mutants Mol Endocrinol, April 2007, 21(4):895–907 903

Hsd17b3 but not for Cyp17a1. Surprisingly, despite thehigh levels of LH in Arinvflox(ex1-neo)/Y, Cyp17a1 expres-sion was not significantly affected. Previous work dem-onstrated that reduction of 17-� hydroxylase activity as-sociated with CYP17A1 is a significant contributor to theblock in steroidogenesis observed in Artfm/Y testis (36,37). The normal levels of Cyp17a1 in Arinvflox(ex1-neo)/Y
may therefore be the reason that Arinvflox(ex1-neo)/Y miceare capable of synthesizing vast amounts of testosteronedespite the apparent down-regulation of other steroido-genic enzymes.
The four members of the kallikrein family of serineproteases, Klk9, Klk21, Klk24, and Klk27, are presentamong the down-regulated transcripts. Within the tes-tis, expression of Klk21, 24, and 27 is limited to theLeydig cells (38–40). The genes encoding the Kal-likreins reside in a large cluster on mouse chromo-some 7 (Fig. 4) (21). The best-studied member of thekallikrein gene family is the human PSA (41). We havedemonstrated that one or more members of the Klkgene family are under T regulation in the Leydig tumorline MA-10. Previous investigations have shown thatKlk21 is up-regulated by testosterone in immatureLeydig cells in culture (39). Taken together with ourdata showing a severe down-regulation in the Klk tran-scripts under conditions of reduced androgen signal-ing, we propose that the four Klks down-regulated inour array analysis represent good markers for andro-gen signaling within the Leydig cell. It is also worthnoting that the expression of all the Klk transcriptsfurther decreased in the presence of Tg(Amh-Cre).This further reduction may represent Sertoli-Leydigcell interaction, a phenomenon described in a study ofanother Sertoli cell-specific Ar mutant (42).
Among the most severely affected transcripts iden-tified in our microarray analysis is alcohol dehydroge-nase 1 (Adh1), which was up-regulated �13-fold inboth Arinvflox(ex1-neo)/Y and Arinvflox(ex1-neo)/Y; Tg(Amh-Cre). Aside from catalyzing the conversion of ethanolto acetaldehyde for detoxification, ADH1 is the rate-limiting enzyme in the conversion of vitamin A to thepotent signaling molecule retinoic acid (RA) (43). De-pletion of vitamin A from the diets of mammals hassevere consequences for spermatogenesis (44). Alsoof interest, two candidate retinoid carrier molecules,prostaglandin D2 synthase and lipocalin 2, are amongthe most severely down-regulated transcripts (45, 46).These changes in gene expression offer the intriguingpossibility of cross talk between the androgenic sig-naling and the RA pathway. Recently, RA signaling hasbeen implicated in promoting meiotic progression inthe murine gonad (47, 48). Although no meiotic phe-notype has been observed in our Ar mutants, ourresults suggest that regulation of RA synthesis andtransport could be the mechanism by which T pro-motes meiotic progression in other models of ARsignaling.
In both Arinvflox(ex1-neo)/Y and Arinvflox(ex1-neo)/Y;Tg(Amh-Cre) we observed a severe decrease in epi-didymal sperm. However, we observed sloughing of
round spermatids from the seminiferous epitheliumand observed round spermatids in the epididymis onlyin Arinvflox(ex1-neo)Y;Tg(Amh-Cre) males. A number ofgenes with putative roles in cell adhesion are either ex-clusively or more severely affected in Arinvflox(ex1-neo)/Y;Tg(Amh-Cre) compared with Arinvflox(ex1-neo)/Y. The tran-script encoding the tight junction component Cldn3 isexclusively down-regulated when the Tg(Amh-Cre) ispresent. CLDN3 is a component of newly forming tightjunctions that make up the blood-testis-barrier (BTB)(49). Loss of CLDN3 correlates with a failure of the BTB,which could lead to the disorganization and disruption ofgerm-Sertoli cell contacts. It is possible that the dysfunc-tion of the cell adhesion in the absence of CLDN3 leadsdirectly to the sloughing of round spermatids. However,the improper establishment of the BTB could also lead toa disruption of a permissive microenvironment neces-sary for germ cell development.
Together, the alterations in the expression of tran-scripts identified in this study contribute to the over-all phenotype observed in Arinvflox(ex1-neo)/Y andArinvflox(ex1-neo)/Y; Tg(Amh-Cre). Identification ofcAREs in the promoters of affected genes will alsobe useful in the identification of genes under directAR regulation. The potential targets of AR signalingidentified in this analysis suggest that AR couldpromote germ cell development by providing bothinstructive signals (i.e. RA signaling) as well as bycreating a permissive environment (i.e. maintenanceof the BTB). The relative contribution of individualtargets of AR identified in this study will provideinsight into the mechanisms by which T supports theongoing developmental process of spermatogenesis.
Materials and Methods
Animals
All animals were housed in a specific pathogen-free environ-ment and cared for under Institutional Animal Care and UseCommittee guidelines. Generation of the hypomorphic Arallele Arinvflox(ex1-neo) has been previously described (10). Thegeneration and characterization of Tg(Amh-Cre) mice will bedescribed in a forthcoming publication. Animals bearing Arin
-
vflox(ex1-neo) and/or Tg(Amh-Cre) were identified by PCR fromtail DNA using methods described previously (10). Animalswere euthanized by CO2 asphyxiation in accordance withInstitutional Animal Care and Use Committee guidelines. Allanimals used in this study were maintained on a 129Sv/JaeSor background.
Preparation of Total Testis mRNA and MicroarrayAnalysis
The testes from wild-type, Arinvflox(ex1-neo)/Y, and Arinvflox(ex1-neo)/Y;Tg(Amh-Cre) 8-wk-old adults were dissected for RNA isolation.Total RNA was extracted from whole testis that was homoge-nized in 1 ml Trizol (Invitrogen, Carlsbad, CA) using a Dounce(Wheaton, Milville, NJ) homogenizer and prepared in accor-dance with the manufacturer’s protocol. RNA from two animalsof each genotype were used in separate array experiments. Thequality of the RNA was assessed by gel electrophoresis and
904 Mol Endocrinol, April 2007, 21(4):895–907 Eacker et al. • Testicular Gene Expression in Ar Mutants

measuring the A260:A280 ratio. Biotinylated cRNA was generatedfrom an oligo-deoxythymidine-primed reverse transcription re-action with 10 �g of total RNA using MEGAscript (Ambion,Austin, TX). Labeled cRNA was hybridized to Affymetrix (SantaClara, CA) MOE430A microarrays following the manufacturer’sprotocol. Microarrays were stained and washed using theGeneChip Fluidics Station 400. The arrays were then scannedwith a GeneArray Scanner 2500A (Agilent, Palo Alto, CA) andanalyzed using Microarray Suite 5.0 (Affymetrix) and Genespring6.1 (Silicon Genetics, Redwood City, CA). Transcripts demon-strating at least a 2-fold change in expression and a minimalsignal strength greater than 50 in any of the comparisons wereconsidered for further analysis. All resulting transcripts demon-strated a statistically significant difference in at least onecomparison.
Northern Analysis
For Northern analysis, 15 �g of total RNA was run on a 1.5%agarose formaldehyde gel. The RNA was then transferredovernight in 20� SSC to Genescreen Hybridization Mem-brane (PerkinElmer, Foster City, CA) and then UV cross-linked. The membranes were hybridized overnight withprobes generated using randomly incorporated �32P-deoxy-ATP using standard methods. Membranes were washed for30 min in 2� SSC, 0.5% sodium dodecyl sulfate, 0.1%sodium pyrophosphate at 65 C followed by a 30-min wash in0.5� SET (10 mM Tris, pH 7.5; 5 mM EDTA; 1% sodiumdodecyl sulfate), 0.1% sodium pyrophosphate at 42 C.
Computational Prediction of AREs
Conserved AREs were found using MONKEY, a program thatuses a sequence alignment, the phylogeny of the sequences,and a weight matrix describing a transcription factor bindingsite to identify conserved transcription factor binding sites(20). The frequencies of the bases at each position in the AREwere determined from the AREs listed in Table 2. Mouse-human alignments were determined using sequences fromthe November 2003 mouse genome assembly (build 32) andthe November 2003 human genome assembly (build 34).Sequences encompassing the 6 kb around transcriptionalstart sites were obtained using EZ-Retrieve (50). Alignmentswere performed using ClustalW (51). We considered a con-served site to be significant if the LLR of the ARE was greaterthan 3 in both mouse and human sequences, and the P valueproduced by MONKEY was less than 0.01. We chose an LLRcutoff of 3 because the lowest LLR for experimentally vali-dated AREs listed in Table 2 was 3.81.
MA-10 Leydig Tumor Cell Culture and cAMP/AndrogenResponsiveness
An isolate of mouse Leydig tumor line MA-10 was a generousgift of Dr. Mario Ascoli (University of Iowa, Iowa City, IA) (52).The MA-10 cells were cultured in RPMI 1640 supplementedwith 25 mM HEPES and 15% normal horse serum (Invitrogen)at 37 C and 5% CO2. To test responsiveness of the cells toDHT and cAMP, 107 cells were transfected with 25 �g ofmouse Ar expression vector (a gift of Dr. Donald Tindall,Mayo Clinic, Rochester, MN) using a Gene Pulser electropo-rator (Bio-Rad, Hercules, CA). The transfected cells weredivided four ways and plated on 60-mm tissue culture plates.Cells were allowed to recover for 16 h and then were exposedto normal media or media supplemented with DHT, 8-Br-cAMP, or both. Cells to be exposed to DHT were pretreatedwith 10 �M DHT (Sigma, St. Louis, MO) for 2 h. After pre-treatment, cells were then exposed to 50 �M 8-Br-cAMP for24 h. Afterward cells were rinsed twice with PBS and totalRNA was harvested using Trizol as described above.
Klk27-luc was generated by introducing 1.3 kb of se-quence proximal to the Klk27 TSS into pGl3-Basic (Promega,
Madison, WI). For luciferase assay, MA-10 cells were trans-fected at 70% confluence in 24-well plates using Lipo-fectamine 2000 (Invitrogen). Reporter constructs (200 ngMMTV-luc or Klk27-luc) were cotransfected with pSV-�gal(20 ng; Promega) alone or in combination with 10 ng of Arexpression vector. Cells were allowed to recover overnight inRPMI1640 supplemented with charcoal-dextran-stripped fe-tal bovine serum. The next day, cells were treated with 10 nM
DHT for 24 h before luciferase assay. All assay were per-formed in triplicate using Bright-Glo and Beta-Glo systems(Promega).
Acknowledgments
We thank Drs. Martin Tompa and William Noble for helpfuldiscussion and comments during analysis of cAREs and De-rek Pouchnik for microarray hybridization and staining. Wealso thank Debra Sprague for assistance in preparation of thismanuscript and Kelly Tysseling for technical assistance.
Received March 7, 2006. Accepted January 16, 2007.Address all correspondence and requests for reprints to:
Robert E. Braun, University of Washington School of Medi-cine, Department of Genome Sciences, Box 355065, 1705NE Pacific, Foege Building, Room 133C, Seattle, Washington98195-5065. E-mail: [email protected].
This work was funded by the Specialized CooperativeCenters Program in Reproductive Research (U54 HD12629)(to R.E.B.) and the Contraceptive Centers Program (U54HD42454) (to M.D.G.).
Current address for J.E.S.: Department of Biopharmaceu-tical Sciences, University of California, San Francisco, SanFrancisco, California 94143
Current address for R.H.W.: Department of Surgery, Col-lege of Medicine (R.H.W.), University of Cincinnati, Cincinnati,Ohio 45267.
The authors have nothing to disclose.
REFERENCES
1. Sharpe RM 1994 Regulation of spermatogenesis. In:Knobil E, Neil JD, eds. The physiology of reproduction.2nd ed. New York: Raven Press; 1363–1434
2. Ghosh S, Sinha-Hikim AP, Russell LD 1991 Further ob-servations of stage-specific effects seen after short-termhypophysectomy in the rat. Tissue Cell 23:613–630
3. Russell LD, Clermont Y 1977 Degeneration of germ cellsin normal, hypophysectomized and hormone treated hy-pophysectomized rats. Anat Rec 187:347–366
4. Kerr JB, Maddocks S, Sharpe RM 1992 Testosteroneand FSH have independent, synergistic and stage-de-pendent effects upon spermatogenesis in the rat testis.Cell Tissue Res 268:179–189
5. Zhou Q, Nie R, Prins GS, Saunders PT, KatzenellenbogenBS, Hess RA 2002 Localization of androgen and estro-gen receptors in adult male mouse reproductive tract. JAndrol 23:870–881
6. Franca LR, Parreira GG, Gates RJ, Russell LD 1998 Hor-monal regulation of spermatogenesis in the hypophysec-tomized rat: quantitation of germ-cell population and ef-fect of elimination of residual testosterone after long-termhypophysectomy. J Androl 19:335–340; discussion341–342
7. O’Donnell L, McLachlan RI, Wreford NG, de Kretser DM,Robertson DM 1996 Testosterone withdrawal promotesstage-specific detachment of round spermatids from therat seminiferous epithelium. Biol Reprod 55:895–901
Eacker et al. • Testicular Gene Expression in Ar Mutants Mol Endocrinol, April 2007, 21(4):895–907 905

8. Saito K, O’Donnell L, McLachlan RI, Robertson DM 2000Spermiation failure is a major contributor to early sper-matogenic suppression caused by hormone withdrawalin adult rats. Endocrinology 141:2779–2785
9. McLachlan RI, O’Donnell L, Meachem SJ, Stanton PG,de Kretser DM, Pratis K, Robertson DM 2002 Identifica-tion of specific sites of hormonal regulation in spermat-ogenesis in rats, monkeys, and man. Recent Prog HormRes 57:149–179
10. Holdcraft RW, Braun RE 2004 Androgen receptor func-tion is required in Sertoli cells for the terminal differenti-ation of haploid spermatids. Development 131:459–467
11. De Gendt K, Swinnen JV, Saunders PT, Schoonjans L,Dewerchin M, Devos A, Tan K, Atanassova N, ClaessensF, Lecureuil C, Heyns W, Carmeliet P, Guillou F, SharpeRM, Verhoeven G 2004 A Sertoli cell-selective knockoutof the androgen receptor causes spermatogenic arrest inmeiosis. Proc Natl Acad Sci USA 101:1327–1332
12. Chang C, Chen YT, Yeh SD, Xu Q, Wang RS, Guillou F,Lardy H, Yeh S 2004 Infertility with defective spermato-genesis and hypotestosteronemia in male mice lackingthe androgen receptor in Sertoli cells. Proc Natl Acad SciUSA 101:6876–6881
13. O’Shaughnessy PJ, Johnston H, Willerton L, Baker PJ2002 Failure of normal adult Leydig cell development inandrogen-receptor-deficient mice. J Cell Sci 115:3491–3496
14. Hales DB, Sha LL, Payne AH 1987 Testosterone inhibitscAMP-induced de novo synthesis of Leydig cell cyto-chrome P-450(17�) by an androgen receptor-mediatedmechanism. J Biol Chem 262:11200–11206
15. Payne AH, Youngblood GL 1995 Regulation of expres-sion of steroidogenic enzymes in Leydig cells. Biol Re-prod 52:217–225
16. Burgos-Trinidad M, Youngblood GL, Maroto MR,Scheller A, Robins DM, Payne AH 1997 Repression ofcAMP-induced expression of the mouse P450 17�-hydroxylase/C17–20 lyase gene (Cyp17) by androgens.Mol Endocrinol 11:87–96
17. O’Shaughnessy PJ, Willerton L, Baker PJ 2002 Changesin Leydig cell gene expression during development in themouse. Biol Reprod 66:966–975
18. Shima JE, McLean DJ, McCarrey JR, Griswold MD 2004The murine testicular transcriptome: characterizing geneexpression in the testis during the progression of sper-matogenesis. Biol Reprod 71:319–330
19. Nelson CC, Hendy SC, Shukin RJ, Cheng H, BruchovskyN, Koop BF, Rennie PS 1999 Determinants of DNA se-quence specificity of the androgen, progesterone, andglucocorticoid receptors: evidence for differential steroidreceptor response elements. Mol Endocrinol 13:2090–2107
20. Moses AM, Chiang DY, Pollard DA, Iyer VN, Eisen MB2004 MONKEY: identifying conserved transcription-fac-tor binding sites in multiple alignments using a bindingsite-specific evolutionary model. Genome Biol 5:R98
21. Olsson AY, Lundwall A 2002 Organization and evolutionof the glandular kallikrein locus in Mus musculus. Bio-chem Biophys Res Commun 299:305–311
22. McKinnell C, Sharpe RM 1992 The role of specific germcell types in modulation of the secretion of androgen-regulated proteins (ARPs) by stage VI-VIII seminiferoustubules from the adult rat. Mol Cell Endocrinol 83:219–231
23. Barbulescu K, Geserick C, Schuttke I, Schleuning WD,Haendler B 2001 New androgen response elements inthe murine pem promoter mediate selective transactiva-tion. Mol Endocrinol 15:1803–1816
24. Rao MK, Wayne CM, Meistrich ML, Wilkinson MF 2003Pem homeobox gene promoter sequences that directtranscription in a Sertoli cell-specific, stage-specific, andandrogen-dependent manner in the testis in vivo. MolEndocrinol 17:223–233
25. Pitman JL, Lin TP, Kleeman JE, Erickson GF, MacLeodCL 1998 Normal reproductive and macrophage functionin Pem homeobox gene-deficient mice. Dev Biol 202:196–214
26. Maclean 2nd JA, Chen MA, Wayne CM, Bruce SR, RaoM, Meistrich ML, Macleod C, Wilkinson MF 2005 Rhox: anew homeobox gene cluster. Cell 120:369–382
27. Zhou Q, Shima JE, Nie R, Friel PJ, Griswold MD 2005Androgen-regulated transcripts in the neonatal mousetestis as determined through microarray analysis. BiolReprod 72:1010–1019
28. Sadate-Ngatchou PI, Pouchnik DJ, Griswold MD 2004Identification of testosterone-regulated genes in testes ofhypogonadal mice using oligonucleotide microarray. MolEndocrinol 18:422–433
29. Denolet E, De Gendt K, Allemeersch J, Engelen K, Mar-chal K, Van Hummelen P, Tan KA, Sharpe RM, SaundersPT, Swinnen JV, Verhoeven G 2005 The effect of a Sertolicell-selective knockout of the androgen receptor on tes-ticular gene expression in prepubertal mice. Mol Endo-crinol 20:321–334
30. Houk CP, Pearson EJ, Martinelle N, Donahoe PK, Teix-eira J 2004 Feedback inhibition of steroidogenic acuteregulatory protein expression in vitro and in vivo by an-drogens. Endocrinology 145:1269–1275
31. Sadate-Ngatchou PI, Pouchnik DJ, Griswold MD 2004Follicle-stimulating hormone induced changes in geneexpression of murine testis. Mol Endocrinol 18:2805–2816
32. Pandey KN, Ascoli M, Inagami T 1985 Induction of reninactivity by gonadotropic hormones in cultured Leydigtumor cells. Endocrinology 117:2120–2126
33. Drummond AE, Risbridger GP, de Kretser DM 1989 Theinvolvement of Leydig cells in the regulation of inhibinsecretion by the testis. Endocrinology 125:510–515
34. Kawamura K, Kumagai J, Sudo S, Chun SY, Pisarska M,Morita H, Toppari J, Fu P, Wade JD, Bathgate RA, HsuehAJ 2004 Paracrine regulation of mammalian oocyte mat-uration and male germ cell survival. Proc Natl Acad SciUSA 101:7323–7328
35. Balvers M, Spiess AN, Domagalski R, Hunt N, Kilic E,Mukhopadhyay AK, Hanks E, Charlton HM, Ivell R 1998Relaxin-like factor expression as a marker of differentia-tion in the mouse testis and ovary. Endocrinology 139:2960–2970
36. Murphy L, O’Shaughnessy PJ 1991 Testicular steroido-genesis in the testicular feminized (Tfm) mouse: loss of17�-hydroxylase activity. J Endocrinol 131:443–449
37. Murphy L, Jeffcoate IA, O’Shaughnessy PJ 1994 Abnor-mal Leydig cell development at puberty in the androgen-resistant Tfm mouse. Endocrinology 135:1372–1377
38. Matsui H, Moriyama A, Takahashi T 2000 Cloning andcharacterization of mouse klk27, a novel tissue kallikreinexpressed in testicular Leydig cells and exhibiting chy-motrypsin-like specificity. Eur J Biochem 267:6858–6865
39. Matsui H, Takahashi T 2001 Mouse testicular Leydigcells express Klk21, a tissue kallikrein that cleaves fi-bronectin and IGF-binding protein-3. Endocrinology 142:4918–4929
40. Matsui H, Takano N, Takahashi T 2005 Characterizationof mouse glandular kallikrein 24 expressed in testicularLeydig cells. Int J Biochem Cell Biol 37:2333–2343
41. Hsieh JT, Wu HC, Gleave ME, von Eschenbach AC,Chung LW 1993 Autocrine regulation of prostate-specificantigen gene expression in a human prostatic cancer(LNCaP) subline. Cancer Res 53:2852–2857
42. De Gendt K, Atanassova N, Tan KA, de Franca LR, Par-reira GG, McKinnell C, Sharpe RM, Saunders PT, MasonJI, Hartung S, Ivell R, Denolet E, Verhoeven G 2005Development and function of the adult generation ofLeydig cells in mice with Sertoli cell-selective or totalablation of the androgen receptor. Endocrinology 146:4117–4126
906 Mol Endocrinol, April 2007, 21(4):895–907 Eacker et al. • Testicular Gene Expression in Ar Mutants

43. Connor MJ, Smit MH 1987 Terminal-group oxidation ofretinol by mouse epidermis. Inhibition in vitro and in vivo.Biochem J 244:489–492
44. Chung SS, Wolgemuth DJ 2004 Role of retinoid signalingin the regulation of spermatogenesis. Cytogenet Ge-nome Res 105:189–202
45. Tanaka T, Urade Y, Kimura H, Eguchi N, Nishikawa A,Hayaishi O 1997 Lipocalin-type prostaglandin D syn-thase (�-trace) is a newly recognized type of retinoidtransporter. J Biol Chem 272:15789–95
46. Chu ST, Lin HJ, Huang HL, Chen YH 1998 The hydro-phobic pocket of 24p3 protein from mouse uterine lumi-nal fluid: fatty acid and retinol binding activity and pre-dicted structural similarity to lipocalins. J Pept Res 52:390–397
47. Menke DB, Koubova J, Page DC 2003 Sexual differen-tiation of germ cells in XX mouse gonads occurs in ananterior-to-posterior wave. Dev Biol 262:303–312
48. Koubova J, Menke DB, Zhou Q, Capel B, Griswold MD,Page DC 2006 Retinoic acid regulates sex-specific tim-ing of meiotic initiation in mice. Proc Natl Acad Sci USA103:2474–2479
49. Meng J, Holdcraft RW, Shima JE, Griswold MD, BraunRE 2005 Androgens regulate the permeability of theblood-testis barrier. Proc Natl Acad Sci USA 102:16696–16700
50. Zhang H, Ramanathan Y, Soteropoulos P, Recce ML,Tolias PP 2002 EZ-Retrieve: a web-server for batch re-trieval of coordinate-specified human DNA sequencesand underscoring putative transcription factor-bindingsites. Nucleic Acids Res 30:e121
51. Thompson JD, Higgins DG, Gibson TJ 1994 CLUSTALW: improving the sensitivity of progressive multiple se-quence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. NucleicAcids Res 22:4673–4680
52. Ascoli M 1981 Characterization of several clonal lines ofcultured Leydig tumor cells: gonadotropin receptors andsteroidogenic responses. Endocrinology 108:88–95
53. PS Rennie, N Bruchovsky, KJ Leco, PC Sheppard, SAMcQueen, H Cheng, R Snoek, A Hamel, ME Bock, andBS MacDonald 1993 Characterization of two cis-actingDNA elements involved in the androgen regulation of theprobasin gene. Mol Endocrinol 7:23-36
54. Cleutjens KB, van Eekelen CC, van der Korput HA, Brink-mann AO, Trapman J 1996 Two androgen response re-gions cooperate in steroid hormone regulated activity of
the prostate-specific antigen promoter. J Biol Chem 271:6379-6388
55. Murtha P, Tindall DJ, Young CY 1993 Androgen induc-tion of a human prostate-specific kallikrein, hKLK2: char-acterization of an androgen response element in the 5�promoter region of the gene. Biochemistry 32:6459-6464
56. Claessens F, Celis L, De Vos P, Peeters B, Heyns W,Verhoeven G, Rombauts W 1993 Intronic androgen re-sponse elements of prostatic binding protein genes. Bio-chem Biophys Res Commun 191:688-694
57. Adler AJ, Scheller A, Robins DM 1993 The stringency andmagnitude of androgen-specific gene activation arecombinatorial functions of receptor and nonreceptorbinding site sequences. Mol Cell Biol 13:6326-6335
58. Kurachi S, Furukawa M, Salier JP, Wu CT, Wilson EJ,French FS, Kurachi K 1994 Regulatory mechanism ofhuman factor IX gene: protein binding at the Leyden-specific region. Biochemistry 33:1580-1591
59. Clay CM, Keri RA, Finicle AB, Heckert LL, Hamernik DL,Marschke KM, Wilson EM, French FS, Nilson JH 1993Transcriptional repression of the glycoprotein hormone �subunit gene by androgen may involve direct binding ofandrogen receptor to the proximal promoter. J BiolChem 268:13556-13564
60. Fabre S, Manin M, Pailhoux E, Veyssiere G, Jean C 1994Identification of a functional androgen response elementin the promoter of the gene for the androgen-regulatedaldose reductase-like protein specific to the mouse vasdeferens. J Biol Chem 269:5857-5864
61. Lund SD, Gallagher PM, Wang B, Porter SC, GanschowRE 1991 Androgen responsiveness of the murine �-glu-curonidase gene is associated with nuclease hypersen-sitivity, protein binding, and haplotype-specific se-quence diversity within intron 9. Mol Cell Biol 11:5426-5434
62. Dai JL, Burnstein KL 1996 Two androgen response ele-ments in the androgen receptor coding region are re-quired for cell-specific up-regulation of receptor messen-ger RNA. Mol Endocrinology 10:1582-1594
63. Barbulescu K, Geserick C, Schuttke I, Schleuning WD,Haendler B 2001 New androgen response elements inthe murine pem promoter mediate selective transactiva-tion. Mol Endocrinol 15:1803-1816
64. Jain A, Lam A, Vivanco I, Carey MF, Reiter RE 2002Identification of an androgen-dependent enhancer withinthe prostate stem cell antigen gene. Mol Endocrinol 16:2323-2337
Molecular Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremostprofessional society serving the endocrine community.
Eacker et al. • Testicular Gene Expression in Ar Mutants Mol Endocrinol, April 2007, 21(4):895–907 907