toxicity of five antidepressant drugs on embryo–larval development and metamorphosis success in...
TRANSCRIPT

1 23
Environmental Science and PollutionResearch ISSN 0944-1344 Environ Sci Pollut ResDOI 10.1007/s11356-013-2211-y
Toxicity of five antidepressant drugson embryo–larval development andmetamorphosis success in the Pacific oyster,Crassostrea gigas
C. Di Poi, L. Evariste, A. Serpentini,M. P. Halm-Lemeille, J. M. Lebel &K. Costil

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag Berlin Heidelberg. This e-offprint is
for personal use only and shall not be self-
archived in electronic repositories. If you wish
to self-archive your article, please use the
accepted manuscript version for posting on
your own website. You may further deposit
the accepted manuscript version in any
repository, provided it is only made publicly
available 12 months after official publication
or later and provided acknowledgement is
given to the original source of publication
and a link is inserted to the published article
on Springer's website. The link must be
accompanied by the following text: "The final
publication is available at link.springer.com”.

COMBINED EFFECTS OF ENVIRONMENTAL STRESSORS IN THE AQUATIC ENVIRONMENT
Toxicity of five antidepressant drugs on embryo–larvaldevelopment and metamorphosis success in the Pacific oyster,Crassostrea gigas
C. Di Poi & L. Evariste & A. Serpentini &M. P. Halm-Lemeille & J. M. Lebel & K. Costil
Received: 28 June 2013 /Accepted: 1 October 2013# Springer-Verlag Berlin Heidelberg 2013
Abstract Unlike conventional pollutants, pharmaceuticalresidues are continuously discharged at low levels (low tomid ng l−1 concentrations), which results in the chronic con-tamination of non-target organisms, but little is known aboutthe effects of these residues. The purpose of this study was toprovide the first assessment of the ecotoxicity of five antide-pressants (selective serotonin reuptake inhibitors [SSRIs] flu-oxetine and sertraline, tricyclic antidepressants [TCAs] clo-mipramine and amitriptyline, and serotonin norepinephrinereuptake inhibitor [SNRI] duloxetine) at a wide range ofconcentrations from 0.1 to 100,000 μg l−1 on two early lifestages in the Pacific oyster. The toxicity was quantified in D-shaped larvae after 36 h of exposure, and in 21-day-oldpediveliger larvae after 24 h of exposure using the percentageof normal larval development and the metamorphosis rate asendpoints, respectively. The embryotoxicity assays reportedthat the EC50 values were within the same range of concen-trations (67 to 192 μg l−1) for all of the tested molecules. Themetamorphosis tests revealed that the antidepressants can beranked along an increasing severity gradient: clomipramine <amitriptyline < duloxetine ~ fluoxetine. Sertraline appeared to
be the less toxic molecule on this endpoint; however, a differ-ent concentration range was used. The embryotoxicity testwas more sensitive than the metamorphosis bioassay for threeof the five molecules tested, but the latter test showed morepractical benefits. Overall, the obtained toxicity values were atleast 10,000-fold higher than the reported environmentalconcentrations.
Keywords Antidepressants . Selective serotonin reuptakeinhibitor (SSRI) . Serotonin norepinephrine reuptake inhibitor(SNRI) . Tricyclic antidepressant (TCA) .Crassostrea gigas .
Embryotoxicity . Metamorphosis
Introduction
Pharmaceuticals are recognised as emerging pollutants inaquatic environments due to their increased consumptionworldwide and to the recent development of more sensitiveanalytical tools (Vasskog et al. 2006; Quinn et al. 2008).Unlike conventional pollutants, pharmaceutical residues andtheir metabolites remain biologically active when releasedinto hydrosystems (Kümmerer 2001) via sewage treatmentplant pathways (Kolpin et al. 2002; Brooks et al. 2005). Thesecontaminants are continuously discharged at levels rangingfrom low-mid ng l−1 to low μg l−1 in effluents and surfacewaters (Kolpin et al. 2002; Brooks et al. 2005; Metcalfe et al.2010) resulting in the likely chronic contamination of aquaticorganisms over their entire life cycle (Brooks et al. 2003;Carlsson et al. 2006; Péry et al. 2008). However, due to theincreasing population and the lack of efficiency of wastewatertreatment plants, it can be expected that effluents will containincreasing amount of pharmaceutical residuals. As a conse-quence, there are growing concerns regarding the adverseeffects of these compounds on non-target aquatic organisms,particularly marine species inhabiting the ultimate receptacle
Responsible editor: Philippe Garrigues
C. Di Poi : L. Evariste :A. Serpentini :M. P. Halm-Lemeille :J. M. Lebel :K. CostilNormandie Université, Esplanade de la Paix, 14032 Caen cedex 5,France
C. Di Poi : L. Evariste :A. Serpentini : J. M. Lebel :K. Costil (*)CNRS INEE, FRE3484 BioMEA, 14032 Caen cedex 5, Francee-mail: [email protected]
C. Di PoiGMPc EA 4259, 14032 Caen cedex 5, France
M. P. Halm-LemeilleCERMN, UPRES EA4258 FR CNRS 3038 INC3M,14032 Caen cedex 5, France
Environ Sci Pollut ResDOI 10.1007/s11356-013-2211-y
Author's personal copy

for waste, which have not been studied much compared withfreshwater fauna.
There are very few data available regarding the level ofchemical contamination in marine waters. The levels of antide-pressant amitriptyline and the anxiolytics diazepam andnordiazepam have been measured to be approximately10 ng l−1 in French Mediterranean coastal waters (Togola andBudzinski 2008). Using polar organic contaminant integrativesamplers (POCIS), the levels of diazepam and nordiazepamhave been quantified at different sampling sites within the samearea from 1 to 4 ng g−1 of sorbent (Munaron et al. 2012). Thesepharmaceuticals are primarily prescribed for the treatment ofdepression, behavioural disorders, or anxiety (Wong et al. 1995;Pélissolo et al. 1996). Amongst antidepressants, three mainclasses can be cited: selective serotonin reuptake inhibitors(SSRIs), serotonin norepinephrine reuptake inhibitors(SNRIs), and tricyclic antidepressants (TCAs). In Europe,France is one of the largest consumers of antidepressants, and45 % of its antidepressant consumers take SSRIs, 39 % takeTCAs, and the remaining consume another class of antidepres-sants (Olié et al. 2002). The SSRI fluoxetine and the TCAsamitriptyline and clomipramine are three drugs available on theFrench market and rank as the most prescribed molecules (Oliéet al. 2002). Although the sub-cited three classes of antidepres-sants differ with respect to their chemical structure, they haveessentially the same mode of action, i.e., they exert monoaminereuptake inhibition in the human brain (DrugBank 2012).Briefly, SSRIs specifically block serotonin (5-HT) reuptaketransporters. SNRIs are dual-uptake inhibitors and inhibit thereuptake of both 5-HT and norepinephrine, and TCAs actprimarily as serotonin–norepinephrine reuptake inhibitors byblocking 5-HT and norepinephrine transporters, respectively.This results in an elevation in the concentrations of theseneurotransmitters in the synaptic cleft, and therefore an en-hancement of neurotransmission.
In mollusc bivalves, catecholamines (dopamine, epineph-rine, and norepinephrine) play essential roles in many biologi-cal endpoints including reproduction (Osada and Nomura1989; Martinez and Rivera 1994), the stress response(Lacoste et al. 2001a, b), the control of cilia on the velum oflarvae (Beiras andWiddows 1995a), and the triggering of larvalsettlement and metamorphosis (Coon et al. 1985; Coon andBonar 1987; Beiras and Widdows 1995b). Similarly to cate-cholamines, 5-HT appears to be implicated in the control ofnumerous physiological processes, such as respiration, feeding,digestion, excretion, movement and visual information process-ing in the sea scallop Placopecten magellanicus (Crolla et al.1995). Moreover, Gibbonsa and Castagna (1984) and Fong(1998) showed the role of 5-HT in reproduction by inducingspawning in several bivalve species. Serotonin also increasesciliary activity of the gills in the mussel Modiolus demissus(Catapane 1983) and induces metamorphosis in pediveligerlarvae of Mytilus galloprovincialis (Satuito et al. 1999).
Estuaries and coastal habitats are continuously subjected toimportant xenobiotic inputs resulting from heavy shippingtraffic, industrialisation, and high population density, and thiseffect is particularly observed in the Seine Basin (Cachot et al.2006; Schnitzler et al. 2011). Among important speciesinhabiting these zones, the Pacific oyster Crassostrea gigasis the leading aquaculture product at the worldwide level.France is a top producer of oysters in Europe and the fourthin the world. However, during the summer, French oysterbasins sporadically experience high mortalities (>30 %),which have important socioeconomic consequences (Royeret al. 2007; Samain et al. 2007; Soletchnik et al. 2007). Thesemortality outbreaks may result from multiple extrinsic andintrinsic factors including temperature variations, physiologi-cal stress and the presence of pathogens and contaminants inthe environment (Samain et al. 2007); in addition, pharma-ceutical pollution might represent another stress factor. Thereis a general assumption in ecotoxicology that the early lifestages (eggs, embryos and larvae) of fish and invertebrates aremore sensitive to xenobiotic agents than adult stages andtherefore represent the critical phases (Herkovits et al. 1997;Hutchinson et al. 1998; Mohammeda et al. 2009). Conse-quently, the early development stages of the Pacific oysterare commonly used as sentinel organisms to assess the toxicityof a large variety of pollutants in marine bioassays due to theirfilter-feeding mode of life and their high sensitivity to a largerange of pollutants (His et al. 1999; Geffard et al. 2002;Wessel et al. 2007; Mai et al. 2012; Mottier et al. 2013). Theembryo–larval toxicity test using C . gigas as a biologicalmodel is recognised as one of the most sensitive of all classi-cally used models (His et al. 1999; Poirier et al. 2007; Mottieret al. 2013). In comparison, the metamorphosis success ofbivalve larvae has started to be used as a biological endpointfor the monitoring of pollution in ecotoxicology (His et al.1997; Mottier et al. 2013).
The present study was designed to test the potential toxiceffects of five antidepressants at a wide range of concentrations(0.1–100,000 μg l−1) on two early life stages, i.e., D-shapedlarvae and pediveliger larvae, of the Pacific oysterC . gigas .Westudied five of the most prescribed antidepressant pharmaceu-ticals in France (Olié et al. 2002): the SSRIs fluoxetine andsertraline; the TCAs clomipramine and amitriptyline, and theSNRI duloxetine. The aim of the work was to provide the firstevaluation of the toxicity of these antidepressants in a marinebivalve using two endpoints, namely, embryo–larval develop-ment and metamorphosis success, and to highlight the useful-ness of these biomarkers in marine ecotoxicology.
Materials and methods
The effects of acute exposure to antidepressants were studiedusing the bioassay method described by Mottier et al. (2013)
Environ Sci Pollut Res
Author's personal copy

in accordance with the standardised AFNOR procedure XPT90-382 (AFNOR 2009).
Chemical compounds and ranges of concentrations
All of the tested concentrations were prepared from a stocksolution of the hydrochloride compound (KEMPROTECLimited®) fluoxetine (CAS No. 59333-67-4), sertraline(CAS No. 79559-97-0), duloxetine (CAS No. 136434-34-9),amitriptyline (CAS No. 549-18-8), and clomipramine (CASNo. 17321-77-6) dissolved in filtered and sterilised seawater(0.22 μm, Steritop®, Millipore).
Embryo–larval toxicity tests were first conducted using abroad range of concentrations from 0.1 to 100,000 μg l−1 (+control) with a factor of 10× between each two consecutiveconcentrations. The concentration range was then narrowed tothe range of 100 to 1,000 μg l−1 (100, 150, 200, 250, 300, 400,600, 800, and 1,000 μg l−1; no data available at 150 μg l−1 forduloxetine) to precisely determine the EC50 values. Eachconcentration was tested in triplicate. In addition, coppersulphate (CuSO4⋅5H2O) was used as a positive control to testthe sensitivity of larvae to a concentration range of 0 to100 μg l−1 CuSO4 (equivalent to 0–25 μg l
−1 Cu2+) accordingto the AFNOR procedure (AFNOR 2009).
Metamorphosis bioassays were first conducted using abroad range of concentrations from 0.1 to 10,000 μg l−1 (+control) with a factor of 10× between each two consecutivedoses. The concentrations used were then narrowed to therange of 100–1,000 μg l−1 (100, 200, 400, 600, 800, and 1,000 μg l−1) to precisely determine the EC50 values. However,because the effect of sertraline was different from that obtain-ed with the four other molecules, different concentrationswere tested. The first range-finding experiment revealed thatthe metamorphosis rate of pediveliger larvae reached 94 % onaverage after exposure to 1,000 μg l−1 sertraline. The concen-tration range was then narrowed to the range of 1,000–10,000 μg l−1 (1,000, 2,000, 4,000, 6,000, 8,000, and 10,000 μg l−1). Each concentration of each molecule was testedat least in sixplicate.
Embryotoxicity bioassay and experimental design
Conditioned oysters from the commercial hatchery GuernseySea Farm Ltd. (Guernsey, UK) were used in theembryotoxicity assays as soon as they arrived at the laborato-ry. Four bioassays using four different batches of genitorswere conducted on January and March 2012 and then, onJanuary and February 2013 (batch nos. 1, 2, 3, and 4, respec-tively; Fig. 1).
For each experiment, five male and five female oysterswere induced to spawn through thermal stimulation by alter-nating between 30-min baths in filtered seawater at 16 °C or28 °C. After spawning, gametes from the different genitor
pairs were observed under a light microscope to selectgenitors with the best cell quality: highly concentrated andmotile spermatozoa and a high number of even and pear-shaped oocytes. The sperm and eggs of the selected parentswere sieved separately through 40- and 100-μm meshes,respectively. The eggs were fertilised with sperm at an egg/sperm ratio of 1:6. The occurrence of fertilisation was verifiedthrough light microscope observations, and 20 min post-fertilisation, the embryos were distributed into glass pillboxescontaining 25 ml of natural filtered and sterilised seawater(0.22 μm; Steritop®Millipore). The embryos were exposed ata density of 60,000 l−1 (corresponding to 1,500 embryos perpillbox) without feeding, aeration, and light. After 36 h ofexposure at 22±1 °C, the organisms were fixed with 500 μl ofan 8 % formalin solution per pillbox. An average of onehundred individuals per replicate were observed under aninverted binocular microscope at 400× magnification (Leica®DM IRB) to determine the number of embryos and abnormal/normal D-shaped larvae. The observations allowed the calcu-lation of the rate of normality and the discrimination of fourtypes of abnormalities amongst the abnormal larvae: shell and/or hinge abnormalities, mantle abnormalities (hypertrophies),shell and/or hinge plus mantle abnormalities, and arresteddevelopment at the "old embryo" stage (Mottier et al. 2013).The results of the embryo–larval development assessment ofthe exposed organisms were expressed as net percentages ofthe normal development (NPNe) of D-shaped larvae adjustedfor the controls as calculated by Mottier et al. (2013). All ofthe bioassays were validated because the mean rates of normallarvae in the control groups was 83.75±1.64 % (ranging from75.5 % to 92.8 %) and/or the EC50 of Cu2+ fell within therequired range of 6–16 μg l−1 (mean EC50=14.87±1.88 μg l−1).
Fig. 1 Comparison of mean (±SEM, error standard and SD [standarddeviation]) percentages of normal D-shaped larvae in the control groupsbetween the four different batches of C . gigas genitors used in theembryo–larval bioassays (ANOVA followed by Newman–Keuls post-hoc tests; batches not sharing a same letter were significantly differentfrom each other)
Environ Sci Pollut Res
Author's personal copy

Metamorphosis bioassay and experimental design
To perform the metamorphosis bioassay, 21-day-oldpediveliger larvae were provided by the SATMAR (SociétéATlantique de MARiculture) hatchery (Barfleur, France). Theanimals were used in the metamorphosis assays as soon asthey arrived at the laboratory. Five bioassays were conductedusing five different batches of pediveliger larvae on January,February, March and May 2012 and March 2013 (batches no.1, 2, 3, 4, and 5, respectively; Fig. 4).
The larvae were exposed in multiwell plates (12 wells,NUNC®; Penfield, New York, USA) in a final volume of1.5 ml of natural filtered and sterilised seawater (0.22 μm;Steritop® Millipore). The larval density was set between 50and 80 larvae/well. Epinephrine at 10−4 M (Sigma Aldrich®)was added to promote larvae settlement and metamorphosis(Coon and Bonar 1987). The exposure to antidepressants wasconducted for 24 h at 22 °C without feeding, aeration, andlight. After 24 h, the exposed larvae were observed using aninverted binocular microscope at 100Xmagnification (Leica®DM IRB) to count the dead larvae that showed no movementand/or tissue degradation. After this first count, the larvaewere fixed with 8 % formalin solution. The metamorphosisrate (%) was evaluated by counting the number of metamor-phosed larvae versus the number of both dead larvae and non-metamorphosed larvae. A larva was considered metamor-phosed if it presented an obvious loss of its velum and itsfoot, new shell growth and well-developed gills. The results ofthe metamorphosis test for the exposed organisms wereexpressed as net percentages of metamorphosis (NPMe) ofpediveliger larvae adjusted for with the controls as calculatedby Mottier et al. (2013). For the five experiments, the meanmetamorphosis success of the control animals was found to be93.70±0.80 % (ranging from 88.3 % to 97.9 %).
Statistical analyses
The analyses were conducted using the STATISTICA 8.0software (Statsoft®, Tulsa, OK, USA). The percentages ofnormal development and metamorphosis were bothtransformed to meet the assumption of normality using theangular transformation arcsin√p , where p is the proportion.The distribution of residuals was then assessed using a Gen-eralised Linear model (GLM) and tested using the Kolmogo-rov–Smirnov normality test. The two endpoints were com-pared between the control groups from each bioassay toevaluate the inter-batch variability using one-way analysesof variance (ANOVAs) followed by Student–Newman–Keuls(SNK) post-hoc tests. ANOVAs followed by SNK post-hoctests were also used to assess the effects of the differentantidepressant concentrations on the percentage of normalD-shaped larvae and the metamorphosis rate.
Nonlinear regressions using the Hill equation wereconducted using the data obtained from the two endpoints(embryo-toxicity and metamorphosis) to calculate the EC50
values (effective concentration that affects 50 % of the individ-uals tested) for each molecule and for each experiment. Theseregressions were conducted using the Excel™ macroREGTOX (Vindimian 2012). Whenever possible, the NOEC(No Observed Effect Concentration) and LOEC (Lowest Ob-served Effect Concentration) values for the two endpoints werealso determined for each antidepressant. The mean EC50 werethen compared between the different antidepressant moleculesfor each endpoint (the EC50 values for the four embryotoxicitybioassays and for all five metamorphosis bioassays were aver-aged) through ANOVAs followed by SNK post-hoc tests be-cause the data were normally distributed. Finally, the EC50
values for each antidepressant were compared between thetwo endpoints (normality rate in D-shaped larvae vs. metamor-phosis rate in pediveliger larvae) using pairwise Mann–Whit-ney U-tests. The statistical threshold was set at p <0.05.
Results
Effects of antidepressants on embryo–larval development
The results revealed very high levels of fecundation becauseonly a few oocytes were observed, and 36 h post-fertilisation,the organisms were either embryos or D-shaped larvae. Fourdifferent batches of oyster genitors were used to perform theembryotoxicity bioassays, and the results showed an inter-genitor variability (ANOVA, p <0.0001; Fig. 1). The percent-age of normal D-shaped larvae observed in the controls fromgenitor batch no. 2 (studied in March 2012) was significantlylower than the rates obtained with the three other lots (SNK, p<0.05).
The results also showed that all the molecules exhibited aconcentration-dependent effect on embryo–larval develop-ment characterised by a decrease in the rates of normal D-shaped larvae with an increase in the antidepressant concen-trations (ANOVAs, p <0.0001 for each molecule; Fig. 2).Compared with the controls, all of the tested molecules, withthe exception of sertraline, had no effect on the normal devel-opment rate at concentrations between 0.1 and 100 μg l−1
(NOEC). A significant impact was observed at concentrationsof at least 150 μg l−1 of fluoxetine, amitriptyline, and clomip-ramine, and at concentration of at least 200 μg l−1 ofduloxetine (no data available at 150 μg l−1; SNK, p <0.001).No more normal D-larvae were observed after exposure toduloxetine at a concentration of 250 μg l−1 and the othermolecules at a concentration of 400 μg l−1, and the mortalityrates reached 100 % at higher concentrations of all of theantidepressants. For sertraline, a significant effect on em-bryo–larval development occurred at 100 μg l−1 (LOEC); at
Environ Sci Pollut Res
Author's personal copy
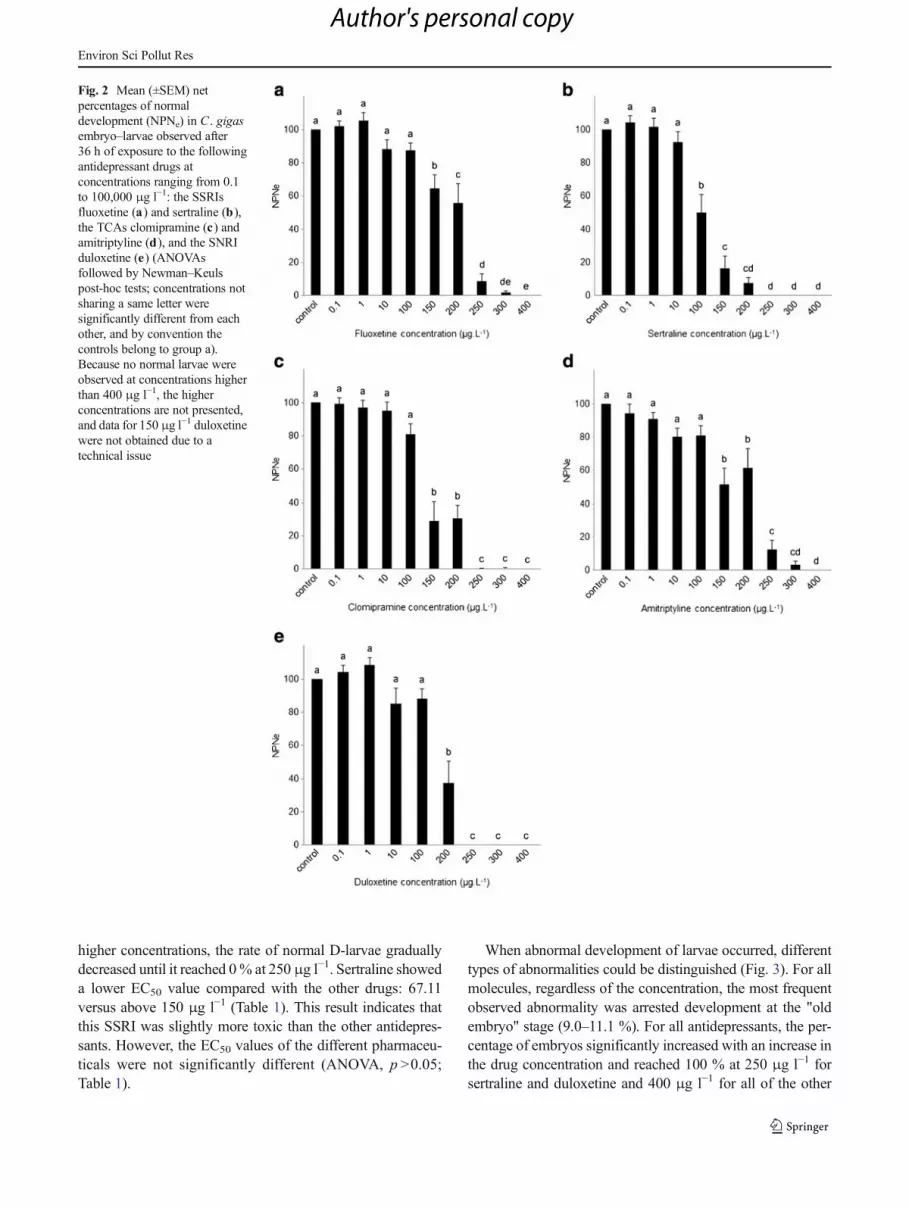
higher concentrations, the rate of normal D-larvae graduallydecreased until it reached 0% at 250 μg l−1. Sertraline showeda lower EC50 value compared with the other drugs: 67.11versus above 150 μg l−1 (Table 1). This result indicates thatthis SSRI was slightly more toxic than the other antidepres-sants. However, the EC50 values of the different pharmaceu-ticals were not significantly different (ANOVA, p >0.05;Table 1).
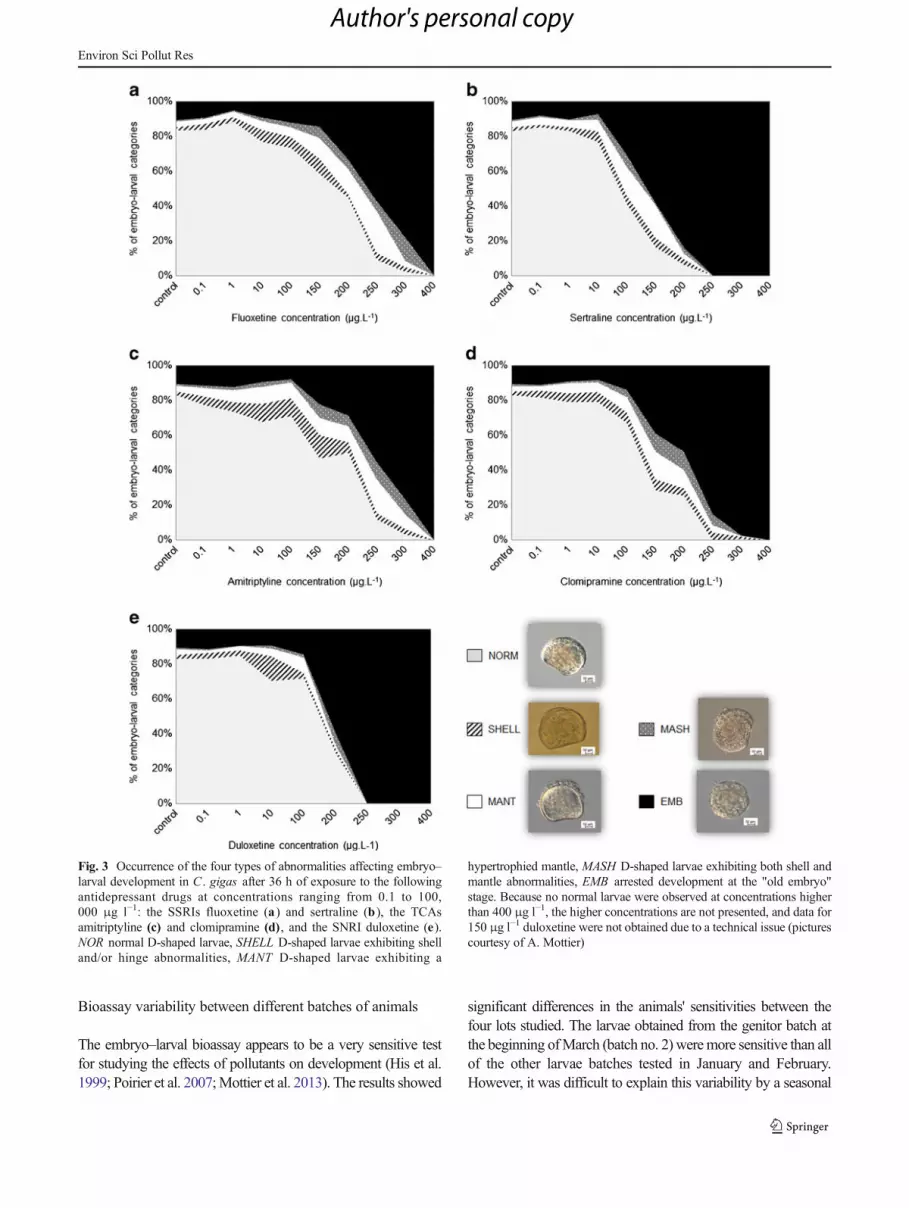
When abnormal development of larvae occurred, differenttypes of abnormalities could be distinguished (Fig. 3). For allmolecules, regardless of the concentration, the most frequentobserved abnormality was arrested development at the "oldembryo" stage (9.0–11.1 %). For all antidepressants, the per-centage of embryos significantly increased with an increase inthe drug concentration and reached 100 % at 250 μg l−1 forsertraline and duloxetine and 400 μg l−1 for all of the other
Fig. 2 Mean (±SEM) netpercentages of normaldevelopment (NPNe) in C . gigasembryo–larvae observed after36 h of exposure to the followingantidepressant drugs atconcentrations ranging from 0.1to 100,000 μg l−1: the SSRIsfluoxetine (a) and sertraline (b),the TCAs clomipramine (c) andamitriptyline (d), and the SNRIduloxetine (e) (ANOVAsfollowed by Newman–Keulspost-hoc tests; concentrations notsharing a same letter weresignificantly different from eachother, and by convention thecontrols belong to group a).Because no normal larvae wereobserved at concentrations higherthan 400 μg l−1, the higherconcentrations are not presented,and data for 150 μg l−1 duloxetinewere not obtained due to atechnical issue
Environ Sci Pollut Res
Author's personal copy
molecules. Overall, the abnormalities affecting the mantlewere more frequent (3.8–6.6 %) than the shell and/or hingeabnormalities (3.0–6.6 %) and both the shell and the mantleabnormalities (1.1–1.8 %). The occurrence of these threeabnormality types was stable regardless of the molecule andthe concentration.
Effects of antidepressants on pediveliger metamorphosis
Five different batches of oyster pediveliger larvae were usedto perform the metamorphosis bioassays. The results indicatedvariability between the lots (ANOVA, p <0.0001; Fig. 4). Themetamorphosis rates of batch nos. 2 and 3 (studied in Febru-ary and March 2012, respectively) were significantly lowerthan those calculated from the other lots (SNK, p <0.05).
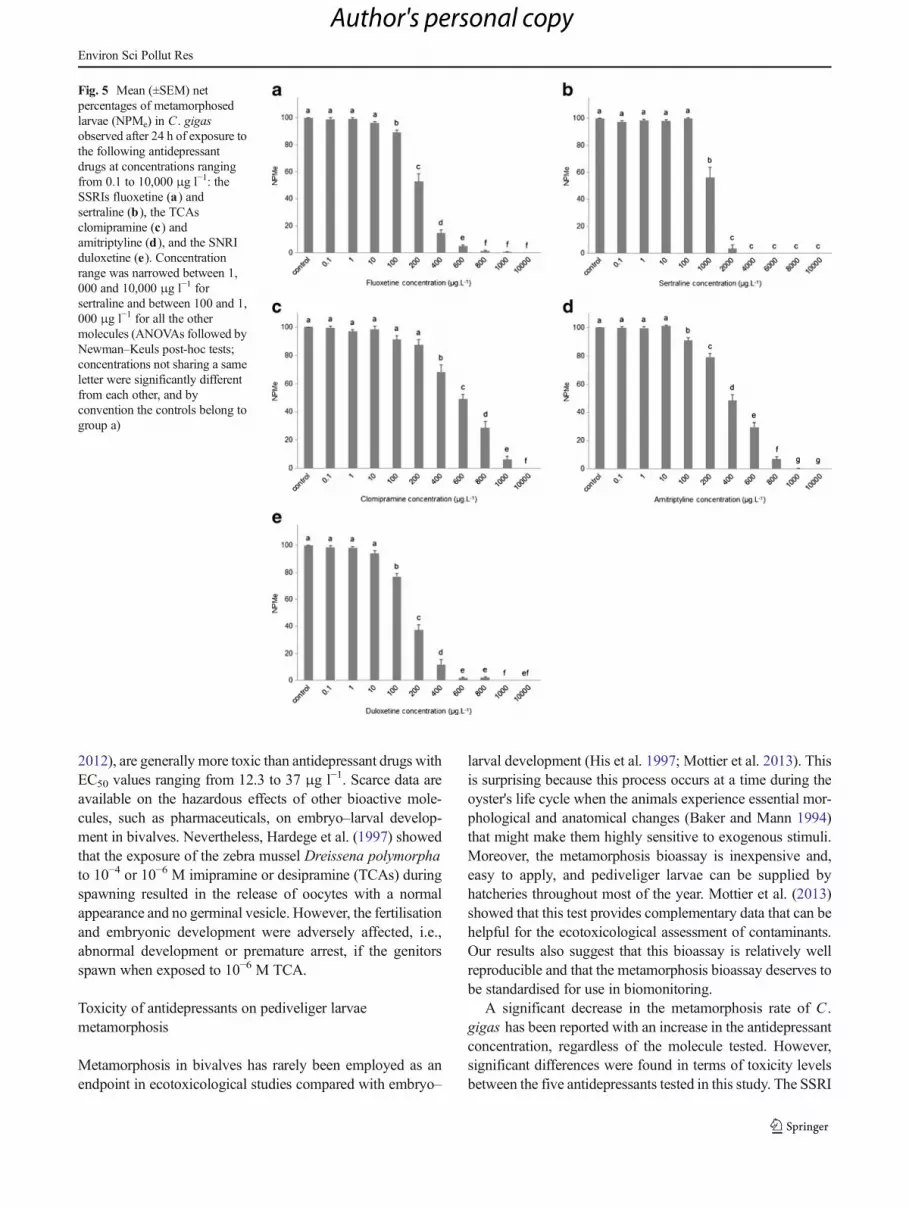
The results showed a significant concentration-dependentdecrease in the metamorphosis rate for all of the molecules(ANOVAs; p <0.0001; Fig. 5). Compared to the controls, 0.1to 10 μg l−1 fluoxetine, duloxetine, and amitriptyline, 0.1 to100 μg l−1 sertraline, and 0.1 to 200 μg l−1 clomipramine hadno effect on the pediveliger larvae metamorphosis rate. Athigher concentrations, a significant decrease in the metamor-phosis rate was observed with an increase in drug concentra-tions until the rate reached 0 % (SNK, p <0.001). A 100 %mortality rate was obtained at 10,000 μg l−1 for all of thetested drugs. Finally, the EC50 values for the different antide-pressants were significantly different (ANOVA, p <0.0001;Table 1). Based on the EC50, NOEC, and LOEC values, theantidepressants can be ranked along a severity gradient (low-est to highest): clomipramine<amitriptyline<duloxetine~flu-oxetine.With caution due to its different concentration ranges,sertraline appeared to be significantly less toxic than the other
molecules as supported by its EC50 value of 1,099 μg l−1
versus that of 155–515 μg l−1 obtained for the four other drugs(SNK, p <0.05; Table 1).
Comparison between the sensitivities of the two life stagesto antidepressants
The variation coefficients of the percentages of normal D-larvae and the metamorphosis rate between the differentbatches of animals fluctuated by 12.7 % and 7.2 %, respec-tively. The D-shaped larvae development was significantlymore sensitive than the pediveliger larvae metamorphosis(lower EC50 values) to three of the five molecules tested: theEC50 values were 16-fold lower for sertraline and 2- to 3-foldlower for the TCAs amitriptyline and clomipramine, respec-tively (Mann–Whitney U -tests; p =0.049, p =0.034, p =0.049, respectively; Table 1). In contrast, this difference wasnot observed with fluoxetine and duloxetine (the EC50 valueswere not significantly different between the two endpoints;Mann–Whitney U -tests, p >0.05; Table 1).
Discussion
We performed the first assessment of the effects of five anti-depressants on the sentinel organism C . gigas using a stan-dard embryotoxicity bioassay and a metamorphosis test tocomplement the lack of ecotoxicological data on their hazard-ous impacts on the environment and non-target marineorganisms.
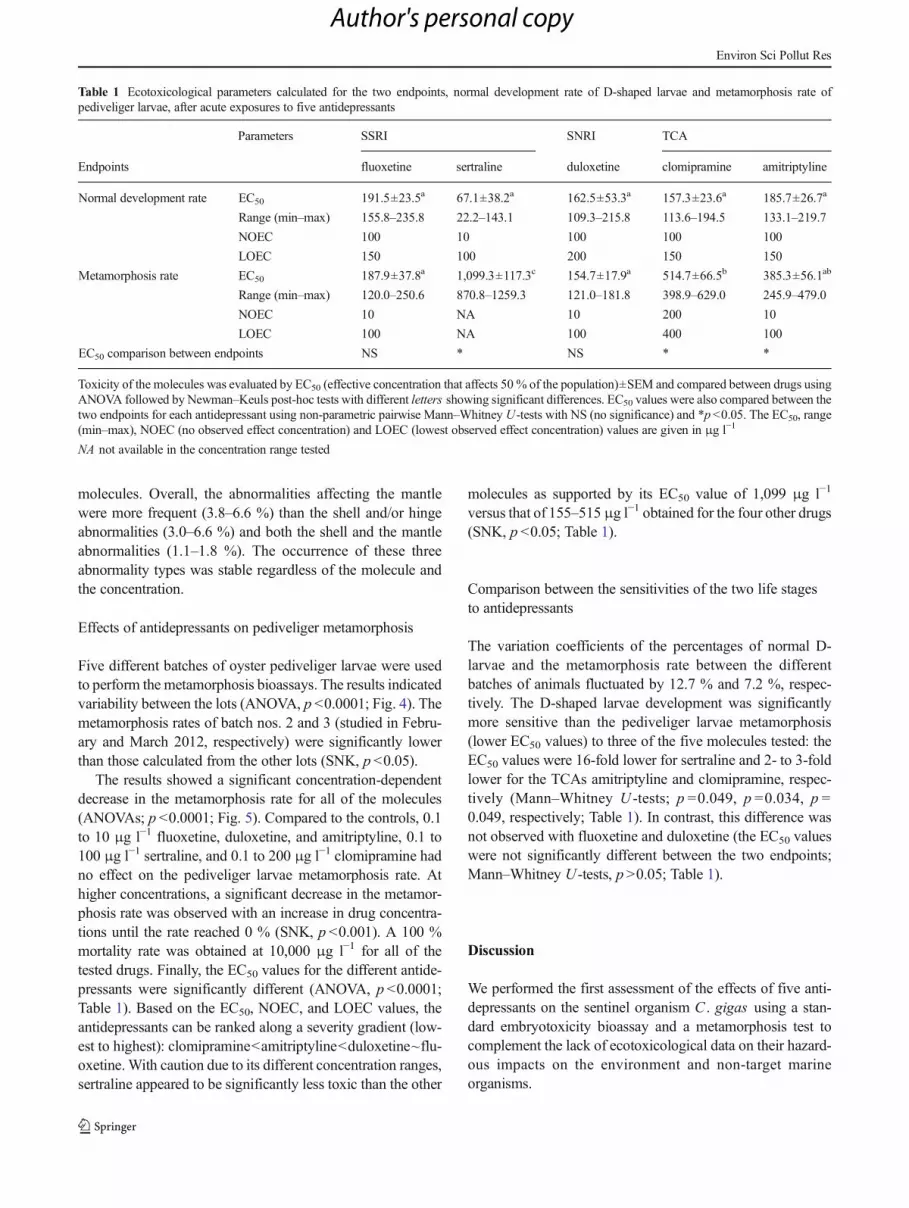
Table 1 Ecotoxicological parameters calculated for the two endpoints, normal development rate of D-shaped larvae and metamorphosis rate ofpediveliger larvae, after acute exposures to five antidepressants
Parameters SSRI SNRI TCA
Endpoints fluoxetine sertraline duloxetine clomipramine amitriptyline
Normal development rate EC50 191.5±23.5a 67.1±38.2a 162.5±53.3a 157.3±23.6a 185.7±26.7a
Range (min–max) 155.8–235.8 22.2–143.1 109.3–215.8 113.6–194.5 133.1–219.7
NOEC 100 10 100 100 100
LOEC 150 100 200 150 150
Metamorphosis rate EC50 187.9±37.8a 1,099.3±117.3c 154.7±17.9a 514.7±66.5b 385.3±56.1ab
Range (min–max) 120.0–250.6 870.8–1259.3 121.0–181.8 398.9–629.0 245.9–479.0
NOEC 10 NA 10 200 10
LOEC 100 NA 100 400 100
EC50 comparison between endpoints NS * NS * *
Toxicity of the molecules was evaluated by EC50 (effective concentration that affects 50 % of the population)±SEM and compared between drugs usingANOVA followed by Newman–Keuls post-hoc tests with different letters showing significant differences. EC50 values were also compared between thetwo endpoints for each antidepressant using non-parametric pairwise Mann–Whitney U-tests with NS (no significance) and *p<0.05. The EC50, range(min–max), NOEC (no observed effect concentration) and LOEC (lowest observed effect concentration) values are given in μg l−1
NA not available in the concentration range tested
Environ Sci Pollut Res
Author's personal copy
Bioassay variability between different batches of animals
The embryo–larval bioassay appears to be a very sensitive testfor studying the effects of pollutants on development (His et al.1999; Poirier et al. 2007;Mottier et al. 2013). The results showed
significant differences in the animals' sensitivities between thefour lots studied. The larvae obtained from the genitor batch atthe beginning ofMarch (batch no. 2) weremore sensitive than allof the other larvae batches tested in January and February.However, it was difficult to explain this variability by a seasonal
Fig. 3 Occurrence of the four types of abnormalities affecting embryo–larval development in C . gigas after 36 h of exposure to the followingantidepressant drugs at concentrations ranging from 0.1 to 100,000 μg l−1: the SSRIs fluoxetine (a) and sertraline (b ), the TCAsamitriptyline (c) and clomipramine (d) , and the SNRI duloxetine (e).NOR normal D-shaped larvae, SHELL D-shaped larvae exhibiting shelland/or hinge abnormalities, MANT D-shaped larvae exhibiting a
hypertrophied mantle, MASH D-shaped larvae exhibiting both shell andmantle abnormalities, EMB arrested development at the "old embryo"stage. Because no normal larvae were observed at concentrations higherthan 400 μg l−1, the higher concentrations are not presented, and data for150 μg l−1 duloxetine were not obtained due to a technical issue (picturescourtesy of A. Mottier)
Environ Sci Pollut Res
Author's personal copy

effect because the genitors were conditioned at the same periodto provide gametes between mid-January and the beginning ofMarch (in 2012 and 2013). However, taken together, the datashowed a relatively low dispersion (the values of the variationcoefficients were less than 13 %) in the percentage of normaldevelopment among the different batches used.
In bioassays using pediveliger larvae, natural differences insensitivity between the different lots of animals were also ob-served. The percentage of metamorphosis obtained for the larvaestudied in February and March 2012 was particularly low com-pared to the batches obtained in January and May 2012, andMarch 2013, whichwere similar to each other. However, the datadispersion was low (the values of the variation coefficients wereless than 8%) and even lower compared with the embryotoxicitytest validated by the AFNOR procedure (AFNOR 2009), whichshows that this bioassay is relatively reproducible.
Several authors have noted variability between several C .gigas larvae bioassays (Stronkhorst et al. 2004; Akcha et al.2012). As suggested byAkcha et al. (2012), this variability maybe explained by the different sensitivities of the genitors topollutants, which may underlie differences in the physiologicalstatus of the larvae batches and/or may be due to geneticdifferences (Damiens et al. 2004). Thus, it appeared particularlyimportant to consider the rates of normal development andmetamorphosis as net percentages standardised to the controlswhen compiling the results from different animal lots instead ofusing the raw rates, as is sometimes used in the literature (e.g.,Wessel et al. 2007).
Toxicity of antidepressants on embryo–larval development
The embryo–larval development has been widely used as anendpoint in marine ecotoxicology in different bivalve species
including the Pacific oyster (His et al. 1999; Lyons et al. 2002;Cachot et al. 2006; Wessel et al. 2007; Libralato et al. 2008;Akcha et al. 2012; Mottier et al. 2013). However, this toxicitytest has always been performed by comparing the number ofnormal larvae to the number of abnormal larvae and is rarelygoing extended to a study of the nature of the abnormalitiesaffecting the larval development. For the first time, Mottieret al. (2013) showed the different types of abnormalities thataffect the embryo–larval development of C . gigas exposed toherbicides. Our results corroborate the previous observationsthat the most frequent abnormality is arrested development atthe "old embryo" stage when these organisms are exposed topesticides (Mottier et al. 2013) or antidepressants (our study)and that the percentage of embryos affected increased with anincrease in the pollutant concentration. In contrast, we dem-onstrated that the abnormalities affecting the mantle were onaverage more frequent than shell abnormalities after exposureto antidepressants, whereas pesticides appeared to more oftenaffect the shell (Mottier et al. 2013). As suggested by theseauthors, it will be interesting to conduct long-term studies todetermine whether shell and/or mantle abnormalities at differ-ent levels of severity are viable or lethal by observing theaffected organisms during their entire larval development.
The results demonstrated similar concentration-dependentresponses in the percentage of normal development of D-shaped C . gigas larvae for the five antidepressants tested.There was no significant difference in EC50 values betweenthe molecules, which ranged from 67 μg l−1 (sertraline) to192 μg l−1 (fluoxetine). This finding suggests that the antide-pressants tested exhibited the same toxicity level on embryo–larval development regardless of their class (TCA, SSRI, andSNRI). However, the comparison of the lowest concentrationthat induces a significant embryotoxic effect (LOEC) revealedthat sertraline was relatively more toxic than the other drugs.Indeed, exposure to sertraline at a concentration of at least100 μg l−1 for sertraline resulted in a significant decrease inthe percentage of normal development, whereas this effectwas only observed with fluoxetine, clomipramine, and ami-triptyline at concentrations of at least 150 μg l−1, and withduloxetine at concentrations of at least 200 μg l−1 (no dataavailable at 150 μg l−1).
The comparison of the endpoints evaluated in our studywith data obtained for other types of pollutants revealed thatpharmaceuticals appear to be more toxic on embryo–larvaldevelopment than several herbicide substances (commercialRoundup® herbicides, glyphosate and AMPA with an EC50
value higher than 1,000 μg l−1; Mottier et al. 2013), but lessthan the herbicide diuron (significant effects on developmentat 0.05 μg l−1; Akcha et al. 2012). Antidepressants are as toxicas pyrene, which belongs to the family of polycyclic aromatichydrocarbons (Lyons et al. 2002), and the organochlorineinsecticide endosulfan (Wessel et al. 2007). However, metals,such as Cl2Hg, CuSO4 (His et al. 1999), and copper (Mai et al.
Fig. 4 Comparison of mean (±SEM, error standard and SD [standarddeviation]) percentages of metamorphosed larvae in the control groupsbetween the five different batches of C . gigas pediveliger larvae used inthe metamorphosis bioassays (ANOVA followed by Newman–Keulspost-hoc tests; batches not sharing a same letter were significantly differ-ent from each other)
Environ Sci Pollut Res
Author's personal copy
2012), are generally more toxic than antidepressant drugs withEC50 values ranging from 12.3 to 37 μg l−1. Scarce data areavailable on the hazardous effects of other bioactive mole-cules, such as pharmaceuticals, on embryo–larval develop-ment in bivalves. Nevertheless, Hardege et al. (1997) showedthat the exposure of the zebra mussel Dreissena polymorphato 10−4 or 10−6 M imipramine or desipramine (TCAs) duringspawning resulted in the release of oocytes with a normalappearance and no germinal vesicle. However, the fertilisationand embryonic development were adversely affected, i.e.,abnormal development or premature arrest, if the genitorsspawn when exposed to 10−6 M TCA.
Toxicity of antidepressants on pediveliger larvaemetamorphosis
Metamorphosis in bivalves has rarely been employed as anendpoint in ecotoxicological studies compared with embryo–
larval development (His et al. 1997; Mottier et al. 2013). Thisis surprising because this process occurs at a time during theoyster's life cycle when the animals experience essential mor-phological and anatomical changes (Baker and Mann 1994)that might make them highly sensitive to exogenous stimuli.Moreover, the metamorphosis bioassay is inexpensive and,easy to apply, and pediveliger larvae can be supplied byhatcheries throughout most of the year. Mottier et al. (2013)showed that this test provides complementary data that can behelpful for the ecotoxicological assessment of contaminants.Our results also suggest that this bioassay is relatively wellreproducible and that the metamorphosis bioassay deserves tobe standardised for use in biomonitoring.
A significant decrease in the metamorphosis rate of C .gigas has been reported with an increase in the antidepressantconcentration, regardless of the molecule tested. However,significant differences were found in terms of toxicity levelsbetween the five antidepressants tested in this study. The SSRI
Fig. 5 Mean (±SEM) netpercentages of metamorphosedlarvae (NPMe) in C . gigasobserved after 24 h of exposure tothe following antidepressantdrugs at concentrations rangingfrom 0.1 to 10,000 μg l−1: theSSRIs fluoxetine (a) andsertraline (b), the TCAsclomipramine (c) andamitriptyline (d), and the SNRIduloxetine (e). Concentrationrange was narrowed between 1,000 and 10,000 μg l−1 forsertraline and between 100 and 1,000 μg l−1 for all the othermolecules (ANOVAs followed byNewman–Keuls post-hoc tests;concentrations not sharing a sameletter were significantly differentfrom each other, and byconvention the controls belong togroup a)
Environ Sci Pollut Res
Author's personal copy

fluoxetine and the SNRI duloxetine appeared to be the mosttoxic molecules on pediveliger larvae metamorphosis (EC50=188 and 155 μg l−1, respectively), whereas the SSRI sertralineappeared to have the least hazardous effect (EC50>1,000 μg l−1), which is contrary to its effect on embryotoxicity.Nevertheless, because a different range of concentrations wastested, further tests are needed to draw a definitive conclusionregarding sertraline toxicity. In contrast, the TCAs amitripty-line and clomipramine had an intermediate effect. As ob-served, antidepressants exerted greater effects on metamor-phosis than several herbicide substances (glyphosate, AMPA,and two commercial formulations), which exhibited EC50
values from approximately 6,000 μg l−1 to higher than 100,000μg l−1 in the same bivalve species using the same bioassay(Mottier et al. 2013).
Effects of antidepressants on neuroendocrine systemsunderlying metamorphosis
Studies on the Pacific oyster and other mollusc species haveindicated that commonly occurring neurotransmitters,neuromodulators, and secondary messenger pathways playimportant roles in the control of metamorphosis. For example,neuroactive compounds, such as GABA and epinephrine,have been shown to induce both settlement and metamorpho-sis in several marine bivalves, including the mussel Mytilusgalloprovincialis , the clams Venerupis pullastra andRuditapes philippinarum , and the oyster Ostrea edulis(García-Lavandeira et al. 2005). Coon et al. (1985) and Coonand Bonar (1987) also demonstrated the implication of theneurotransmitters epinephrine, norepinephrine, acetylcholine,and L-DOPA into this process in the Pacific oyster C . gigas .In contrast, serotonin induced less metamorphosis, whichsuggests that this neurotransmitter does not play a preponder-ant role in this species (Beiras and Widdows 1995b). Inaddition, Coon and Bonar (1987) demonstrated the existenceof receptors similar to vertebrate-type alpha1-adrenoreceptorsin the Pacific oyster that mediate metamorphosis. These re-searchers also suggested that the addition of 10−4 M epineph-rine artificially promotes metamorphosis in this bivalve. As aconsequence, 10−4 M epinephrine has been used to promotesettlement on an artificial substrate (multiwell plates), and thusmetamorphosis (Mottier et al. 2013). Thus, we cannot excludethe possibility that an interaction between artificially addedepinephrine and the antidepressants tested may have resultedin the negative effects observed on the metamorphosis rate.According to the DrugBank database (DrugBank 2012), TCAand SNRI molecules can have an antagonistic action onadrenergic receptors in mammals. The molecules tested in thisstudy may potentially interact with the alpha1-receptors pres-ent in C . gigas by blocking the receptors to which bothendogenous and exogenous epinephrine, which was addedin our culture medium, normally bind and thereby ultimately
suppress the induction of metamorphosis. However, we ob-served significant decreases in metamorphosis success with anincrease in the antidepressant concentration even for theSSRIs fluoxetine and sertraline, which are not antagonists ofthis type of receptors, at least in vertebrates (DrugBank 2012).However, the negative effects of antidepressants on metamor-phosis cannot be generalised to other marine molluscs. In fact,the injection of 10−6 M fluoxetine (i.e., 3,458 μg l−1), which isa higher concentration than the EC50 calculated in the Pacificoyster in the current study, induced metamorphosis in compe-tent larvae in the marine snail Ilyanassa obsoleta . A similareffect was observed with α-methyl-5-HT, which is a serotoninagonist. These effects could be species-dependent but mayalso be due to differences in the experimental design (e.g.,mode and time of exposure, tested doses).
Comparison of life stage sensitivity
It has been reported that early life stages of the Pacific oysterare more sensitive to toxicological injuries due to exposure toseveral herbicides than older organisms (Mottier et al. 2013).The present study suggests that 36-h-old D-shaped larvae aremore sensitive to exposure to the SSRI sertraline and theTCAs amitriptyline and clomipramine than 21-day-oldpediveliger larvae; this effect was not observed with the SSRIfluoxetine and the SNRI duloxetine. It thus appears that thetwo endpoints provide different and non-redundant results.The sensitivity of early life stages can be linked with essentialprocesses occurring during this time of life, i.e., morphologi-cal axes are being formed, cells are being rearranged, cell fateis being specified, and multilayered axes are being formed(Gilbert 2003). In comparison, morphological and anatomicalchanges also occur during metamorphosis in C . gigas , butthese cannot be considered as important. The main changesare the progressive degeneration of the velum, the dis-appearance of the eyespots and then the foot, the grad-ual development of the gills, and the growth of the shellbeyond the margin of the prodissoconch (Baker andMann 1994). These processes involve histolysis of lar-val tissues, differentiation and proliferation of adulttissues, and associated biochemical and physiologicalmodifications. However, their sensitivities were not dif-ferent for the SSRI fluoxetine and the SNRI duloxetine,which suggests that the animals are more sensitive tothe specific molecule than to the antidepressant class.This observation is quite surprising because, despite distinctstructural differences between the compounds, each antide-pressant class has essentially the same mode of action throughmonoaminergic neurotransmitters, even if different moleculeswithin the same class can bind to different secondary targets(DrugBank 2012) or act unspecifically, e.g., by narcosis(Cleuvers 2003).
Environ Sci Pollut Res
Author's personal copy

Antidepressant mode of actions in the Pacific oyster:hypotheses
Nothing is known regarding the neurotransmission systems inthe early life stages in the Pacific oyster. As a consequence, itcan only be hypothesised whether antidepressants bind spe-cifically to protein-like transporters that are homologues of themammalian monoaminergic transporters, and/or whether thetoxic effects observed are driven by indirect mechanisms dueto the penetration of the drug into the cells in the absence oftargets. Fong et al. (2003) further elucidated the pharmacologyof SSRI-induced spawning and parturition by blocking thesereproductive processes with two mammalian 5-HT2 receptorantagonists in the freshwater zebra mussel Dreissenapolymorpha and the fingernail clam Sphaerium spp. Thesetwo antagonists were potent inhibitors of both processes in-duced by the SSRIs fluvoxamine, fluoxetine, and zimelidine,and the results thus provide indirect evidence of the existenceof alternative SSRI targets in molluscs. In contrast, Raineyet al. (2010) demonstrated that, in the absence of its proteintarget, the antidepressant sertraline and, more globally, manypsychoactive cationic amphiphiles target phospholipid mem-branes in the yeast strain Saccharomyces cerevisiae . In addi-tion, El-Bassat et al. (2011) showed that fluoxetine causes cellrupture and cell deformations at low concentrations in algae(30–50 μg l−1) and at higher concentrations in several zoo-plankton species. During their acute experiment (72-hduration), all of the tested aquatic organisms suffered fromoxidative stress to different degrees depending on the fluoxe-tine concentration used. The symptoms were cell wall rupture,dialysis of the cell contents, deformation of the cell bodyshape, inhibition of antioxidant enzyme activities and elevatedlevels of lipid peroxidation. Moreover, the fact that molecules,such as fluoxetine and sertraline, belonging to the same SSRIclass of antidepressants showed different degrees of toxicityon the metamorphosis success raises the question of theirmode of action. Cleuvers (2003) showed that two anti-inflammatory drugs, diclofenac and ibuprofen, had oppositelevels of toxicity within the same aquatic species, and actunspecifically by non-polar narcosis. As a consequence, thisauthor suggested that their toxicity may be associated with thelog Kow of the drugs rather than any specific action. All of themolecules studied in our study have a relatively high log Kow
(between 4.09 and 5.19), which makes them highly lipophilicand more readily absorbed across membranes. As a conse-quence, the effects observed may also be the result of a non-specific narcotic action.
Conclusion
The objective of the study was to establish more comprehen-sive data on the ecotoxicity of antidepressants in a non-target
marine organism. We highlight innovative and relevant eco-toxicological data for five antidepressant drugs, and comple-ment the evident gap in knowledge. The results provide thefirst demonstration that all of the antidepressants studied,regardless of their class (SSRI, TCA, and SNRI), adverselyaffect the embryo–larval development and the metamorphosissuccess of the Pacific oyster C . gigas . Nevertheless, theconcentrations that resulted in significant effects were severalorders (10,000- to 100,000-fold) of magnitude higher than theconcentrations of antidepressants actually measured in coastalwaters (i.e., ~10 ng l−1). However, adverse effects may resultfrom chronic exposure to lower concentrations, and the puta-tive hazardous effects of pharmaceutical mixtures cannot beexcluded. In addition, both embryotoxicity and metamorpho-sis bioassays can be considered relevant and complementarybiomonitoring tools for the assessment of contaminants, in-cluding antidepressant pharmaceuticals. In C . gigas , theembryotoxicity test was found to be more sensitive for threemolecules of the five molecules tested, i.e., 16 times moresensitive for sertraline and two to three times more for theTCAs, than the metamorphosis bioassay, but the latter testshowed more practical benefits.
Acknowledgments This work is a contribution to the Pharm@Ecotoxproject funded by the French National Research Agency (ANR, fr:Agence Nationale de la Recherche). The authors gratefully acknowledgethe personnel of the SATMAR (Société ATlantique de MARiculture)hatchery (Barfleur, France) for providing pediveliger larvae, and RebeccaOlley for editing the English form.
References
AFNOR (2009) Bio indicateur de la toxicité potentielle de milieuxaqueux. XP T90-382. 19
Akcha F, Spagnol C, Rouxel J (2012) Genotoxicity of diuron and glyph-osate in oyster spermatozoa and embryos. Aquat Toxicol 106–107:104–13
Baker SM, Mann R (1994) Feeding ability during settlement and meta-morphosis in the oyster Crassostrea virginica (Gmelin, 1791) andthe effects of hypoxia on post-settlement ingestion rates. J Exp MarBiol Ecol 181:239–253
Beiras R, Widdows J (1995a) Effect of the neurotransmitters dopamine,serotonin and norepinephrine on the ciliary activity of mussel(Mytilus edulis) larvae. Mar Biol 122:597–603
Beiras R,Widdows J (1995b) Induction of metamorphosis in larvae of theoyster Crassostrea gigas using neuroactive compounds. Mar Biol123:327–334
Brooks BW, Chambliss CK, Stanley JK et al (2005) Determination ofselect antidepressants in fish from an effluent-dominated stream.Environ Toxicol Chem 24:464–469
Brooks BW, Foran CM, Richards SM et al (2003) Aquatic ecotoxicologyof fluoxetine. Toxicol Lett 142:169–183
Cachot J, Geffard O, Augagneur S et al (2006) Evidence of genotoxicityrelated to high PAH content of sediments in the upper part of theSeine estuary (Normandy, France). Aquat Toxicol 79:257–267
Carlsson C, Johansson A-K, Alvan G et al (2006) Are pharmaceuticalspotent environmental pollutants? Part I. Environmental risk
Environ Sci Pollut Res
Author's personal copy

assessments of selected active pharmaceutical ingredients. Sci TotalEnviron 364:67–87
Catapane EJ (1983) Comparative study of the control of lateral ciliaryactivity in marine bivalves. Comp Biochem Physiol Part C: CompPharmacol 75:403–405
Cleuvers M (2003) Aquatic ecotoxicity of pharmaceuticals including theassessment of combination effects. Toxicol Lett 142:185–194
Coon SL, Bonar DB (1987) Pharmacological evidence that alpha 1-adrenoceptors mediate metamorphosis of the Pacific oyster,Crassostrea gigas. Neuroscience 23:1169–1174
Coon SL, Bonar DB, Weiner RM (1985) Induction of settlement andmetamorphosis of the pacific oyster, Crassostrea gigas (Thunberg),by L-DOPA and catecholamines. J Exp Mar Biol Ecol 94:211–221
Crolla RP, Too CK, Pani AK, Nason J (1995) Distribution of serotonin in thesea scallop Placopecten magellanicus. Invert Reprod Dev 28:125–135
Damiens G, His E, Gnassia-Barelli M et al (2004) Evaluation of bio-markers in oyster larvae in natural and polluted conditions. CompBiochem Physiol Toxicol Pharmacol CBP 138:121–128
DrugBank (2012) DrugBank. http://www.drugbank.ca. Accessed 26 June2013
El-Bassat RA, Touliabah HE, Harisa GI, Sayegh FAQ (2011) Aquatictoxicity of various pharmaceuticals on some isolated plankton spe-cies. Int J Med Sci 3:170–180
Fong PP (1998) Zebra mussel spawning is induced in low concentrationsof putative serotonin reuptake inhibitors. Biol Bull 194:143–149
Fong PP, Philbert CM, Roberts BJ (2003) Putative serotonin reuptakeinhibitor-induced spawning and parturition in freshwater bivalves isinhibited by mammalian 5-HT2 receptor antagonists. J Exp ZoolPart A Comp Exp Biol 298:67–72
García-LavandeiraM, Silva A, AbadM et al (2005) Effects of GABA andepinephrine on the settlement and metamorphosis of the larvae offour species of bivalve molluscs. J Exp Mar Biol Ecol 316:149–156
Geffard O, Budzinski H, His E (2002) The effects of elutriates from PAHand heavy metal polluted sediments onCrassostrea gigas (Thunberg)embryogenesis, larval growth and bio-accumulation by the larvae ofpollutants from sedimentary origin. Ecotoxicology 11:403–416
Gibbonsa M, Castagna M (1984) Serotonin as an inducer of spawning insix bivalve species. Aquaculture 40:189–191
Gilbert SF (2003) Developmental biology. Sinauer AsHardege JD, Duncan J, Ram JL (1997) Tricyclic antidepressants suppress
spawning and fertilization in the zebra mussel, Dreissenapolymorpha . Comp Biochem Physiol Part C Pharmacol ToxicolEndocrinol 118:59–64
Herkovits J, Cardellini P, Pavanati C, Perez-Coll CS (1997) Susceptibilityof early life stages of Xenopus laevis to cadmium. Environ ToxicolChem 16:312–316
His E, Beiras R, Silhouette C (1997) Research note a simplification thebivalve embryogenesis and larval development bioassay method forwater quality assessment. Water Res 31:351–355
His E, Heyvang I, Geffard O et al (1999) A comparison between oyster(Crassostrea gigas) and sea urchin (Paracentrotus lividus) larvalbioassays for toxicological studies. Water Res 33:1706–1718
Hutchinson TH, Solbé J, Kloepper-Sams PJ (1998) Analysis of the ecetocaquatic toxicity (EAT) database III— comparative toxicity of chem-ical substances to different life stages of aquatic organisms.Chemosphere 36:129–142
Kolpin DW, Furlong ET, Meyer MT et al (2002) Pharmaceuticals, hor-mones, and other organic wastewater contaminants in U.S. streams,1999–2000: a national reconnaissance. Environ Sci Technol 36:1202–1211
Kümmerer K (2001) Drugs in the environment: emission of drugs,diagnostic aids and disinfectants into wastewater by hospitals inrelation to other sources — a review. Chemosphere 45:957–969
Lacoste A, Malham SK, Cueff A et al (2001a) Evidence for a form ofadrenergic response to stress in the mollusc Crassostrea gigas . JExp Biol 204:1247–1255
Lacoste A, Malham SK, Cueff A, Poulet SA (2001b) Stress-inducedcatecholamine changes in the hemolymph of the oyster Crassostreagigas . Gen Comp Endocrinol 122:181–188
Libralato G, Losso C, Arizzi Novelli A et al (2008) Ecotoxicologicalevaluation of industrial port of Venice (Italy) sediment samples aftera decontamination treatment. Environ Poll 156:644–650
Lyons BP, Pascoe CK, McFadzen IRB (2002) Phototoxicity of pyreneand benzo[a]pyrene to embryo–larval stages of the Pacific oysterCrassostrea gigas. Mar Environ Res 54:627–631
Mai H, Cachot J, Brune J et al (2012) Embryotoxic and genotoxic effectsof heavy metals and pesticides on early life stages of Pacific oyster(Crassostrea gigas). Mar Poll Bull 64:2663–2670
Martinez G, Rivera A (1994) Role of monoamines in the reproductiveprocess of Argopecten pupuratus . Invert Reprod Dev 25:167–174
Metcalfe CD, Chu S, Judt C et al (2010) Antidepressants and theirmetabolites in municipal wastewater, and downstream exposure inan urban watershed. Environ Toxicol Chem 29:79–89
Mohammeda A, Halfhidea T, Elias-Samlalsingha N (2009) Comparativesensitivity of three life stages of the tropical mysid,Metamysidopsisinsularis to six toxicants. Toxicol Environ Chem 91:1331–1337
Mottier A, Kientz-Bouchart V, Serpentini A et al (2013) Effects ofglyphosate-based herbicides on embryo–larval development andmetamorphosis in the Pacific oyster, Crassostrea gigas . AquatToxicol 128–129:67–78
Munaron D, Tapie N, Budzinski H et al (2012) Pharmaceuticals,alkylphenols and pesticides inMediterranean coastal waters: Resultsfrom a pilot survey using passive samplers. Estuar Coast Shelf Sci114:82–92
Olié J, Omari FEL, Spadone C, Lépine J (2002) Résultats d'une enquêtesur l'usage des antidépresseurs en population générale française.L'Encéphale 28:411–417
Osada M, Nomura T (1989) Seasonal variations of catecholamine levelsin the tissues of the Japanese oyster, Crassostrea gigas . CompBiochem Physiol Part C Comp Pharmacol 93:171–173
Pélissolo A, Boyer P, Lépine JP, Bisserbe JC (1996) Epidemiology of theuse of anxiolytic and hypnotic drugs in France and in the world.L'Encéphale 22:187–196
Péry ARR, GustM, Vollat B et al (2008) Fluoxetine effects assessment onthe life cycle of aquatic invertebrates. Chemosphere 73:300–304
Poirier L, Quiniou F, Ruiz N et al (2007) Toxicity assessment ofpeptaibols and contaminated sediments on Crassostrea gigas em-bryos. Aquat Toxicol 83:254–262
Quinn B, Gagné F, Blaise C (2008) The effects of pharmaceuticals on theregeneration of the cnidarian,Hydra attenuata . Sci Tot Environ 402:62–69
Rainey MM, Korostyshevsky D, Lee S, Perlstein EO (2010) The antide-pressant sertraline targets intracellular vesiculogenic membranes inyeast. Genetics 185:1221–1233
Royer J, Ropert M, Costil K et al (2007) Spatio-temporal changes inmortality, growth and condition of the Pacific oyster, Crassostreagigas , in Normandy (France). J Shellfish Res 26:973–984
Samain JF, Dégremont L, Soletchnik P et al (2007) Genetically basedresistance to summer mortality in the Pacific oyster (Crassostreagigas) and its relationship with physiological, immunological char-acteristics and infection processes. Aquaculture 268:227–243
Satuito CG, Natoyama K, Yamazaki M et al (1999) Induction of meta-morphosis in the pediveliger larvae of the mussel Mytilusgalloprovincialis by neuroactive compounds. Fish Sci 65:384–389
Schnitzler JG, Thomé JP, Lepage M, Das K (2011) Organochlorinepesticides, polychlorinated biphenyls and trace elements in wildEuropean sea bass (Dicentrarchus labrax) off European estuaries.Sci Tot Environ 409:3680–3686
Soletchnik P, Ropert M, Mazurié J et al (2007) Relationships betweenoyster mortality patterns and environmental data from monitoringdatabases along the coasts of France. Aquaculture 271:384–400
Environ Sci Pollut Res
Author's personal copy

Stronkhorst J, Ciarelli S, Schipper CA et al (2004) Inter-laboratorycomparison of five marine bioassays for evaluating the toxicity ofdredged material. Aquat Ecosyst Health Manag Soc 7:147–159
Togola A, Budzinski H (2008) Multu-residue analysis of pharmaceuticalcompounds in aqueous samples. J Chromatogr A 1177:150–158
Vasskog T, Berger U, Samuelsen P et al (2006) Selective serotoninreuptake inhibitors in sewage influents and effluents from Tromsø,Norway. J Chromatogr A 1115:187–195
Vindimian E (2012) MSExcel Macro Regtox.7.0.6 Freely available fromEric Vindimian. IRSTEA, France
Wessel N, Rousseau S, Caisey X et al (2007) Investigating the relation-ship between embryotoxic and genotoxic effects of benzo[a]pyrene,17alpha-ethinylestradiol and endosulfan on Crassostrea gigas em-bryos. Aquat Toxicol 85:133–142
Wong DT, Bymaster FP, Engleman EA (1995) Prozac (fluoxetine, Lilly110140), the first selective serotonin uptake inhibitor and an antidepres-sant drug: twenty years since its first publication. Life Sci 57:411–441
Environ Sci Pollut Res
Author's personal copy