assimilation of gelatin-acacia microencapsulated lipid by pacific oyster (crassostrea gigas) spat
TRANSCRIPT

Aquaculture
ELSEVIER Aquaculture 153 (I 997) 291-300
Assimilation of gelatin-acacia microencapsulated lipid by Pacific oyster ( Crassostrea gigas) spat
Jens Knauer * , Paul C. Southgate Deprtment of Zoology, James Cook l/nil,ersir?; of North Queensland, TowsrYlle. Qld. 481 I, Austrtrlicr
Accepted 18 December 1996
Abstract
Gelatin-acacia microcapsules (GAM) are commonly used as a delivery vehicle for lipids in nutritional studies with bivalves. In this study the efficiency with which lipid contained in GAM was assimilated by Pacific oyster (Crassostrea gi~ns) spat was determined. The marine diatom Ckaeroceros muelleri was cultured in the presence of “C-sodium bicarbonate for 24 h. The 14C-lipid fraction was extracted, mixed with corn oil and microencapsulated in GAM. Spat assimilated ‘“C-lipids from a mixed diet of the marine flagellate Dunalielln tertiolecta (80%) and GAM (20%) with an efficiency of 79.9 + 2.9%, while spat fed a 100% GAM diet assimilated “C-lipids at a rate of 57.1 + 5.2%. Addition of 100 mg 1-l of the antibiotic Thimerosal to both a mixed diet and GAM only significantly reduced bacterial degradation of GAM over 24 h. The high assimilation efficiencies underline the potential of GAM as carrier of lipids in nutritional studies with bivalves. 0 1997 Elsevier Science B.V.
Kewvrds: Assimilation: Microcapsule: Nutrition; Lipid; Oyster; Crassostwu gigas
1. Introduction
Microalgal diets have been used in many studies as tools to assess the nutritional requirements of bivalves (for review see Webb and Chu, 1983; Brown et al., 1989). However, it is probably impossible to offer either a number of microalgal species which differ in the level of one component only or, alternatively, to feed one species of microalgae and alter the quantity of a particular nutrient without affecting the levels of
* Corresponding author: Tel: +61-77-815547; fax: +61-77.251570; e-mail: [email protected].
0044-8486/97/$17.00 0 1997 Elsevier Science B.V. All rights reserved. PII SOO44-8486(97)00038-O

292 J. Knauer, P.C. Southgaie/Aquaculture 153 (1997) 291-300
others. The information derived from such studies is thus often inconclusive (Webb and Chu, 1983; Thompson and Harrison, 1992) and, consequently, relatively little is known of the nutritional requirements of bivalves. Currently, the major constraint to advances in this field of study is therefore the lack of satisfactory artificial diets whose composi- tion, in contrast to microalgae, can be precisely controlled.
Over recent years, microencapsulation of either specific nutrients or complete test diets has emerged as a promising technique to study the nutritional needs of bivalves. Microcapsules (MC) of the cross-linked protein-walled type have been used to examine aspects of protein (Langdon and Siegfried, 1984; Kreeger, 1993; Kreeger et al., 1995, 1996) and carbohydrate (Kreeger et al., 1996) metabolism of bivalves. Similarly, lipid nutrition has been investigated using gelatin-acacia microcapsules (GAM) (Langdon and Waldock, 198 1; Numaguchi and Nell, 199 1).
Lipids have been delivered to suspension-feeders in the form of liposomes (Parker and Selivonchick, 1986), lipid microspheres (Robinson, 1992a,b; Heras et al., 1994), emulsions (Coutteau et al., 1994, 1996), or various types of MC (Langdon and Bolton, 1984; Langdon and Siegfried, 1984; Chu et al., 1987; Southgate and Lou, 1995). GAM, however, have been the most widely used artificial delivery vehicle for lipids in nutritional studies with bivalves (Langdon and Waldock, 1981; Chu et al., 1982, 1987; Southgate, 1988; Numaguchi and Nell, 1991). Langdon and Waldock (1981) demon- strated the essentiality of eicosapentaenoic acid (EPA, 20:5n - 3) and/or docosahex- aenoic acid (DHA, 22:6n - 3) to Pacific oyster (Crussostrea gigas) spat by feeding a diet of the marine flagellate Dunaliella tertiolecta and lipid-containing GAM. Despite their potential, however, the assimilation efficiency (AE) of lipids presented to bivalves in GAM has not yet been determined. This information is of particular importance with regard to the quantification of nutritional requirements.
The aim of this study was therefore firstly, to assess the AE of gelatin-acacia microencapsulated 14C-lipids by Pacific oyster (Crussostrea gigas) spat fed as either a 20% replacement of Dunaliella tertiolecta, or as a 100% ration. D. tertiolecta was chosen since it lacks fatty acids (FA) greater than C 18 and, in combination with GAM, is a useful tool to study the requirements for highly unsaturated FA by bivalves (Langdon and Waldock, 1981). Secondly, the effect of the antibiotic Thimerosal on the bacterial degradation of GAM was evaluated.
2. Materials and methods
2.1. Culture of microalgae
The marine flagellate Dunaliellu tertiolectu (Code CS 175) and the marine diatom Chaetoceros muelleri (Code CS 176) were obtained from CSIRO Marine Laboratories, Hobart, Australia. Microalgal cultures were grown in 5 1 glass flasks using f/2 medium (Guillard, 1975) at 24.6 + 1.7”C under a 12 h L:12 h D light regime. Cultures in the exponential growth phase were used to feed spat. The dry weights of D. tertiolecta and

.I. Knauer, P.C. Southgate/Aquaculture 153 (1997) 291-300 293
C. muelleri were determined as 115.62 + 3.87 pg cell-’ and 33.88 k 2.67 pg cell-‘,
respectively. One mCi of “C-labelled sodium bicarbonate (Amersham) was added to a 4 1 batch
culture of C. muelleri (3.02 X lo6 cells ml- ’ >, which was used because of its relatively high lipid content (Knauer and Southgate, 1996). The flask was then sealed and stirred using a magnetic stirrer for 24 h. Following that, the algae were centrifuged at 3000 g for 10 min and the lipid fraction extracted according to a modification (Dunstan et al., 1993) of the method of Bligh and Dyer (1959).
2.2. Microencapsulation of t4C-microalgal lipids
The extracted “C-microalgal lipids were mixed with 2 ml of corn oil and a total of 2.5 ml of the mixture was microencapsulated following the method of Southgate and Lou (1995). The resulting GAM were resuspended in 10 ml distilled water. The mean diameter of GAM was 4.5 f 1.0 pm (mean k SD, IZ = 100). To determine the specific activity (disintegration min ’ (dpm) ml- ’ ) of the GAM, 0.1 to 0.5 ml of a 1: 10 dilution of GAM in distilled water were mixed with 0.2 ml Soluene (Packard) and left for 2 h at 50°C. Five ml of a scintillation cocktail (Starscint, Packard) were added and the samples were counted in a scintillation counter (1217 Rackbeta, LKB Wallac) for 5 min. The specific activity of the GAM suspension was 11715 272 i 1 169 854 dpm ml-’ or 158315 f 15809 dpm mg-’ dry weight.
2.3. Feeding experiment
Crassostrea gigas spat were obtained from Shellfish Culture P/L, Tasmania, Aus- tralia. Triplicate groups of 50 spat (group wet weights ranging from 1.38 to 1.43 g) were kept in 5 1 beakers filled with 4 1 of seawater (30%0 salinity) filtered through a 5, 1, 0.45 Frn and an activated carbon cartridge followed by UV-sterilisation. Spat were condi- tioned at 22.4 i 1.2”C under a 12 h L: 12 h D light regime for one week. During that time, all spat were fed C. muelleri since it has a high nutritional value for oysters (Knauer and Southgate, 19961, but they were not fed one day prior to the experiment. Control beakers were set up in triplicate, containing the same number of baked (450°C for 24 h) oyster shells of the same initial wet weight as the experimental animals (Langdon and D e B evoise, 1990). In all treatments, seawater was exchanged every day. All beakers were covered by an airtight lid and gentle aeration was provided. Escaping 14C0, was trapped in GF/C filters soaked in 1 ml of 4 M NaOH.
The amounts of food to be added were calculated according to the formula of Epifanio ( 1979):
QR = 0.01 x W-“.33
where QR is the dry weight of ration per g wet weight of oysters, and W is the g initial wet weight of oysters. Three groups of spat were fed mixtures of D. tertiolecta (80%) and “C-labelled GAM (20%), and the same diet was added to the three controls. Likewise, GAM (100%) were added to a further three batches of spat and three control

294 J. Kncruer, P.C. Southgate/AquacLllturee 1.53 (19971 291-300
beakers. The experimental diet was added once every 24 h for 3 days. Following this, spat were purged of “C-labelled GAM by feeding unlabelled C. muelleri for 2 days, followed by a day of no feeding.
2.4. Assimilation of microencapsulated “C-lipids
The AE of ‘“C-lipid contained in GAM was determined according to methods modified from Langdon and DeBevoise (1990). The ‘“C-activities of seawater at the beginning of each 24 h feeding period were determined by taking three 5 ml samples 1 min after addition of the diets. At the end of each 24 h feeding period and prior to seawater exchange, three 5 ml samples of seawater were taken again. All seawater samples were incubated with 0.2 ml Soluene at 50°C for 2 h, mixed with 10 ml scintillation cocktail (Starscint) and counted in a scintillation counter for 5 min. The ‘“C-activities of final seawater samples were corrected for the amount of respired 14C02 dissolved in seawater.
Each experimental beaker was checked for the presence of faeces after 6, 12 and 24 h to minimise leaching of 14C from faeces into seawater. Faeces were siphoned off the bottom, collected on a 25 pm mesh, and the seawater was returned to the beaker. On Day 2 and 3, some of the experimental beakers contained small amounts of pseudofae- ces. Faeces and pseudofaeces were separated using a dissecting microscope, and the latter were returned to the respective beaker. At the end of each 24 h feeding period all the seawater in a beaker was poured through a 25 p,m mesh to collect the faeces. All collected faeces were combined in a scintillation vial and, once the faeces settled, the supematant fraction of seawater was removed. One ml Soluene was added to the faeces and they were kept for 24 h at 50°C. Following that, the faecal samples were decolourised by incubation with 0.5 ml of 30% (w/v) hydrogen peroxide at 50°C for a further 12 h before addition of 5 ml scintillation fluid (Hionic-Fluor, Packard). Particu- late material from the control beakers was collected and processed in the same manner. The results of faecal “C-activity from live spat were corrected for non-faecal 14C-activ- ity by subtraction of particulate 14C-activity from the control beakers.
The amount of respired 14C02 was determined from a 1 1 seawater sample collected at the end of each 24 h feeding period, which was filtered through GF/C and 0.2 pm Whatman filters. The three seawater samples collected over the three days were then combined in sealed containers and preserved by additions of 1% v/v (final concentra- tion) formaldehyde and 4 M NaOH (final pH > 10). Prior to analysis, the acidity of the samples was increased to pH < 3 using 1 M citric acid, and the ‘“CO, escaping from the samples was trapped in GF/C filters soaked in 1 ml of 4 M NaOH. The 14C02 trapped by filters was then eluted with 5 ml of distilled water. This process removes over 95% of ‘“CO, from the filters (K. Anthony, James Cook University of North Queensland, pers. comm., 1996). Using “C-labelled sodium bicarbonate as a standard, the amount of radioactivity accounted for using this method was 41.3 f 3.5% (n = 3). The GF/C filters used to trap 14C02 diffusing out of seawater were processed in the same way, and the 14C-activity obtained was combined with that of the respired ‘“CO, fraction. Measurements of respired 14C02 were corrected for “CO2 produced by bacteria in control beakers in order to determine the amount of ‘“CO, produced by spat alone.

J. Knauer, P.C. Southgate/Aquacultlrre 153 (19971 291-300 295
The amount of “‘C incorporated by spat was determined by digesting the combined soft tissue of all spat in each beaker in 2 ml Soluene at 50°C for 24 h, followed by a further incubation with 0.5 ml of 30% (w/v> hydrogen peroxide at 50°C for 12 h. Finally, 5 ml scintillation fluid (Hionic-Fluor) was added.
The AE of ‘“C-labelled GAM was defined as the percentage of ingested 14C that was either incorporated or respired:
AE= respired14C + incorporated’“C
ingestedI C X 100
Ingestion was determined as the sum of 14C present in the faecal, respired and
incorporated fractions. Total 14C recovered, expressed as a percentage of the total “C added to each beaker, includes ingested 14C, faecal 14C and 14C remaining in the beakers after 24 h.
Two different scintillation cocktails were used since the use of Starscint is recom- mended for water samples, and Hionic-Fluor for samples dissolved in Soluene (manufac- turer’s information). An aliquot of 0.2 ml Soluene was added to all seawater samples to disrupt any particulate matter, but since the samples were mainly water, Starscint was used as the scintillant. Initial controls showed that the background readings of both scintillation cocktails did not differ significantly (P I 0.05).
2.5. Bacterial degradation of gelatin-acacia microcapsules
The effect of bacteria on the degradation of GAM and subsequent release of ‘“C-lipids from GAM was quantified in the absence of spat but under conditions identical to the AE study. The method used was modified from Langdon and DeBevoise (1990). The appropriate amounts of either D. tertiolectu and GAM or GAM alone (see above) plus 100 mg l- ’ of the antibiotic Thimerosal (SIGMA) were added to triplicate 5 1 beakers. Control suspensions, without the antibiotic, were also prepared in triplicate. All beakers were covered by an airtight lid and aeration was provided over the 24 h incubation period (22.9”C, 12 h L:12 h D light regime). Any escaping ‘“CO, was trapped as described above.
The initial and final specific activity of each suspension was determined in triplicate 5 ml samples taken 1 min after the addition of GAM and after 24 h, respectively. Furthermore, initial and final 100 ml samples were filtered through 0.8 pm cellulose nitrate filters and the filters were subsequently washed in 5 ml distilled water. In a preliminary study this method was shown to remove more than 95% of particulate matter from the filters. The specific activities of both the distilled water and a 5 ml sample of the filtrates were determined. After addition of 0.2 ml Soluene, the water samples were incubated for 2 h at 50°C before adding 10 ml scintillation cocktail (Starscint) and counting for 5 min. The ‘“CO, trapped in GF/C filters over the 24 h period was processed as detailed above, and the proportion of ‘“C detected in the filtrate.
14C-activity obtained was added to the
The retention rate of “C-particulate matter on filters in beakers containing Thimerosal was compared with control beakers using a Student t-test. Percentage values were arcsin-transformed prior to analysis. Results were considered to be significantly different at the P 5 0.05 level of probability.
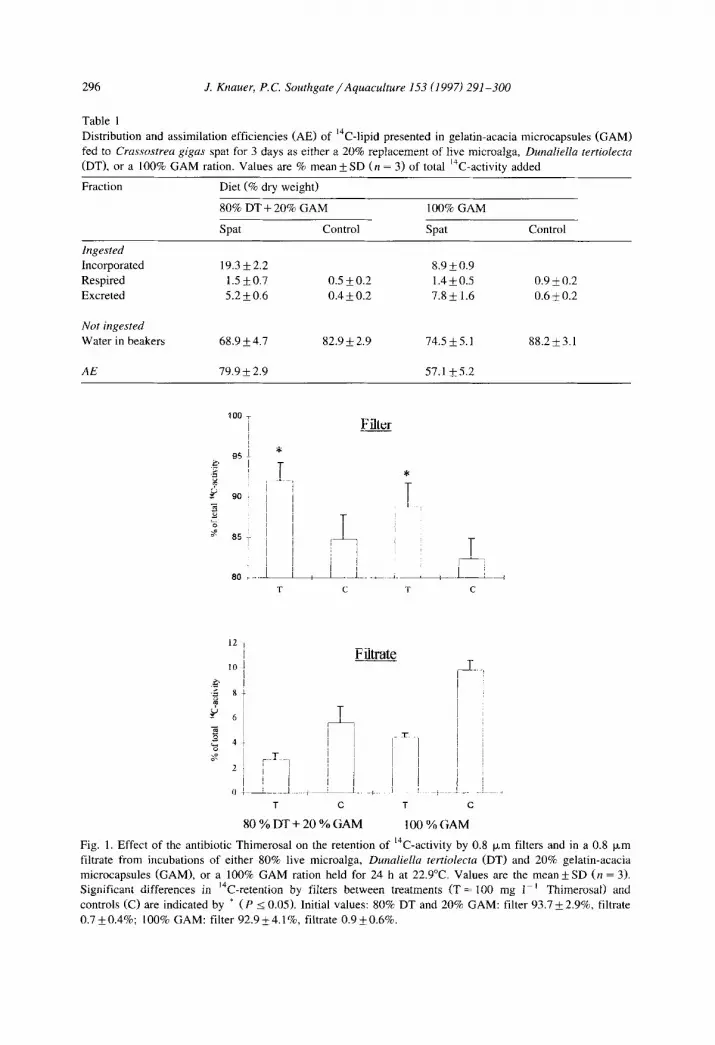
296
Table 1
.I. Knauer, P.C. Southgate/Aquaculture 153 (1997) 291-300
Distribution and assimilation efficiencies (AE) of 14C-lipid presented in gelatin-acacia microcapsules (GAM)
fed to Crassmtrea gigas spat for 3 days as either a 20% replacement of live microalga, Dunaliella tertiolectn
CDT), or a 100% GAM ration. Values are % mean f SD (n = 3) of total ‘“C-activity added
Fraction Diet (% dry weight)
80% DT + 20% GAM
Spat Control
100% GAM
Spat Control
Ingested
Incorporated
Respired
Excreted
Not ingested
Water in beakers
AE
19.3 +2.2 8.9 * 0.9
1.5f0.7 0.5 f 0.2 1.4+0.5 0.9 i 0.2
5.2 f 0.6 0.4 f 0.2 7.8+ 1.6 0.6 i 0.2
68.9k4.7 82.9 + 2.9 74.5 f 5.1 88.2i3.1
79.9 k 2.9 57.1 +5.2
1
1. c I : -.---I
C T
8O%DT+2O%GAM 100 % GAM
Fig. 1. Effect of the antibiotic Thimerosal on the retention of 14C-activity by 0.8 p,rn filters and in a 0.8 pm
filtrate from incubations of either 80% live microalga, Dunaliella tertiolecta (DT) and 20% gelatin-acacia
microcapsules (GAM), or a 100% GAM ration held for 24 h at 22.9”C. Values are the mean f SD (n = 3). Significant differences in “C-retention by filters between treatments (T = 100 mg I-’ Thimerosat) and
controls CC) are indicated by * (P 5 0.05). Initial values: 80% DT and 20% GAM: filter 93.7 + 2.9%, filtrate
0.7*0.4%; 100% GAM: filter 92.9*4.1%, filtrate 0.9*0.6%.
F&ate
i
i i
I I

J. Knauer, P.C. Sourhgate/Aquaculture 153 (1997) 291-300 297
3. Results
3.1. Assimilation of microencapsulated ‘“C-lipids
The amount of 14C-activity accounted for in treatment beakers and controls varied from 83 to 95%. Spat fed the 80% D. tertiolecta and 20% GAM diet ingested 26.0 f 1.7% of the total 14C presented. Expressed as a percentage of the total 14C added, 19.3 * 2.2% was incorporated into tissue, 1.5 f 0.7% respired and 5.2 f 0.6% excreted as faeces. The AE of 14C-lipid contained in GAM was 79.9 + 2.9% (Table 1).
Spat fed the 100% GAM diet ingested 18.2 k 1.8% of the total 14C-label; 8.9 + 0.9% of the total 14C added was incorporated, 1.4 &- 0.5% respired and 7.8 f 1.6% excreted as faeces. This resulted in an AE of 57.1 f 5.2% for the 100% GAM diet (Table 1).
3.2. Bacterial degradation of gelatin-acacia microcapsules
A total of 86 to 97% of the 14C-activity added to the treatment and control beakers was accounted for. Thimerosal produced a significant increase in the retention of “C-particulate matter by the filters in the 80% D. tertiolecta and 20% GAM-treatment (92.0 k 2.3%) compared with the control (84.8 f 3.0%, P 5 0.05) (Fig. 1). Similarly, Thimerosal also produced a significantly better retention in the 100% GAM diet (86.8 + 2.9%) when compared with the control (80.6 + 3.0%, P I 0.05) (Fig. 1).
4. Discussion
The antibiotic Thimerosal had a small but significant effect on the rate of bacterial breakdown of GAM, even though the seawater used was filtered to 0.45 pm and LJV-sterilised. The beneficial effect of using antibiotics to inhibit bacterial degradation of MC has been shown in previous studies (Langdon and Bolton, 1984; Langdon and DeBevoise, 1990), and the results of this study support these findings. The retention of 87% of “C-activity (100% GAM) on filters is very similar to the findings of Langdon and DeBevoise (19901, who retained 90 to 94% of 14C-activity in protein-walled MC using the same antibiotic.
Of the total 14C-activity added to the experimental beakers in the AE study, 83 to 95% was accounted for. This is higher than the 74 to 89% of 14C-protein encapsulated in protein-walled and glyceride-coated nylon-protein-walled MC accounted for by Langdon and DeBevoise (1990). The difference in recovery of 14C could be due to the trapping of “C0, diffusing out of seawater over the experimental period, even though Langdon and DeBevoise (1990) stated that such losses were negligible if the seawater was alkaline (pH 8.3). This was not supported in the present study (pH 8.1 to 8.3) where diffusional losses of 14C0, accounted for up to 11% of the total 14C-activity of the respired fraction.
The data obtained for AE show that GAM are a suitable vehicle to deliver lipids to oyster spat. C. gigas spat readily assimilated lipid presented in GAM supplied either as

298 J. Knauer, P.C. Southgate/Aquaculture 153 (1997) 291-300
dietary supplement to microalgae or as a 100% ration. The ingestion rate of the 100% GAM diet (18%) falls within the 17 to 21% range reported for a 100% diet of protein-walled MC ingested by juvenile C. gigas (Langdon and DeBevoise, 1990). On a percentage basis, more of the GAM were ingested when added as a 20% replacement of a microalgal diet (26%) than when fed as a 100% ration (18%). This may have been due to the presence of microalgae and their dissolved metabolites in the experimental beakers, which have been shown to affect the feeding rate of bivalves on both natural diets (Ward and Targett, 1989; Ward et al., 1992) and MC (Kreeger, 1993).
MC whose walls are made up of protein and/or carbohydrates are readily digested by bivalves (Gabbott et al., 1976; Langdon, 1989; Langdon and DeBevoise, 1990; Kreeger and Langdon, 1994) due to the presence of a wide variety of digestive enzymes in the gastric juices (Gabbott et al., 1976). Similarly, GAM have been shown to be readily digested by larval bivalves (Chu et al., 1982; Southgate, 19881, and the results of the present study indicate that C. gigas spat are also capable of digesting GAM. Incorpora- tion of “C-cholesterol contained in GAM by bivalve larvae has previously been demonstrated (Southgate, 19881. However, the present study is the first to put forward AE values for microencapsulated lipids. Microencapsulated nutrients have been shown to be incorporated by bivalves with AE ranging from 6 (Kreeger and Langdon, 1994) to 40% (Langdon, 1989) for protein, and from 16 to 20% (Kreeger et al., 1995) for carbohydrate. In contrast, the AE of 14C-lipid from GAM offered as a 20% supplement to a microalgal diet (80%) falls within the 77 (Kiorboe et al., 1981) to 90% (Widdows and Bayne, 1971; Peirson, 1983) range of AE reported for microalgae of good food value. This is not surprising since bivalves possess a wide variety of lipid-digesting enzymes in their digestive tract (Gabbott et al., 1976). The AE of encapsulated ‘“C-protein has been shown to be influenced by reproductive seasonality in the mussels Mytilus trossulus (Kreeger, 1993) and M. edulis (Kreeger et al., 1995). It is possible that, similar to protein, the AE of dietary lipids also varies according to the reproductive cycle of adults as well as during the early ontogeny of bivalves.
GAM are not a suitable vehicle to deliver complete artificial diets to bivalves since only water-insoluble nutrients can be encapsulated using this method. However, they may prove to be a very useful tool in studies addressing aspects of the lipid nutrition of bivalves. For example, although the essentiality of the fatty acids EPA and/or DHA has been shown for C. gigas spat (Langdon and Waldock, 1981), the requirements for these essential nutrients have yet to be quantified. GAM offer a suitable means for assessing these requirements as well as those for other fatty acids and lipid components. The data generated in such studies will facilitate the further development of suitable artificial diets for bivalves.
Acknowledgements
This study was supported by the Aquaculture Cooperative Research Centre Ltd. and the Department of Zoology, James Cook University of North Queensland.

J. Knauer, P.C. Sourhgare/Aquaculture 153 (1997) 291-300 299
References
Bligh, E.G. and Dyer, W.F., 1959. A rapid method of total lipid extraction and purification. Can. J. Biochem.
Physiol., 47: 91 l-917.
Brown, M.R., Jeffrey, S.W. and Garland. C.D., 1989. Nutritional aspects of microalgae used in mariculture; a
literature review. CSIRO Mar. Lab. Rep. Ser., No. 205, 44 pp.
Chu, F.-L.E., Webb, K.L., Hepworth, D. and Roberts. M., 1982. The acceptability and digestibility of
microcapsules by larvae of Crassostreu ~,irginica. J. Shellfish Res., 2: 29-34.
Chu, F.-L.E., Webb, K.L., Hepworth, D. and Casey, B.B., 1987. Metamorphosis of larvae of Crassosrrea
kginica fed microencapsulated diets. Aquaculture, 64: 185-197.
Coutteau, P.. Caers, M., Mallet. A., Moore, W., Manzi, J. and Sorgeloos, P., 1994. Effect of lipid
supplementation on growth, survival and fatty acid composition of bivalve larvae (Osrrea edulis L. and
Mrrcenariu mercenaria L.). In: P. Kestemont, J. Muir. F. Seviila and P. Williot (Editors). Measures for
Success. Proceedings of Bordeaux Aquaculture 1994, March 23-25, 1994, France. CEMAGREF, pp.
213-218.
Coutteau, P., Castell, J.D., Ackman. R.G. and Sorgeloos, P.. 1996. The use of lipid emulsions as carriers for
essential fatty acids in bivalves: a test case with juvenile Plncopecten magrllunicus. J. Shellfish Res., 15:
259-264.
Dunstan, G.A., Volkman, J.K. and Barrett, S.M., 1993. The effect of lyophilization on the solvent extraction of
lipid classes, fatty acids and sterols from the oyster Crassostren gigtrs. Lipids, 28: 937-944.
Epifanio, C.E., 1979. Growth in bivalve molluscs: nutritional effects of two or more species of algae in diets
fed to the American oyster Crmsostrecr r,irginica (Gmelin) and the hard clam Mrrcenariu mwrcenariu.
Aquaculture, 18: I - 12.
Gabbott, P.A., Jones. D.A. and Nichols. D.H., 1976. Studies on the design and acceptability of micro-en-
capsulated diets for marine particle feeders. II. Bivalve mollusca. In: G. Persoone and E. Jaspers (Editors).
Proceedings of the 10th European Symposium on Marine Biology, @tend. Belgium, Sept. 17-23. 1975,
Universa Press, Wetteren, pp. 127- 14 I. Guillard, R.R., 1975. Culture of phytoplankton for feeding marine invertebrates. In: W.L. Smith and M.H.
Chanley (Editors). Culture of Marine Invertebrate Animals. Plenum Press, NY, pp. 29-60.
Heras, H., Kean-Howie, J. and Ackman, R.G., 1994. The potential use of lipid microspheres as nutritional
supplements for adult Osrretr e&/is. Aquaculture. 123: 309-322.
Kigrboe. T., Mghlenberg, F. and Npihr, 0.. 1981. Effect of suspended bottom material on growth and
energetics in MyIilrr.7 edulis. Mar. Biol., 61 : 283-288.
Knauer, J. and Southgate, P.C., 1996. Nutritional value of a spray-dried freshwater alga, Spongiococcum
e.wwnrricun~. for Pacific oyster (Crtrssostrea gigus) spat. Aquaculture 146: I35- 146.
Kreeger, D.A.. 1993. Seasonal patterns in utilization of dietary protein by the mussel A4ytilrr.s mmulus. Mar.
Ecol. Prog. Ser.. 95: 215-232.
Kreeger, D.A. and Langdon. C.J.. 1994. Digestion and assimilation of protein by MpiIu.7 ~rmsdu.~ (Bivalvia: Mollusca) fed mixed carbohydrate/protein microcapsules. Mar. Biol., I1 8: 479-488.
Kreeger. D.A.. Hawkins, A.J.S.. Bayne, B.L. and Lowe. D.M., 1995. Seasonal variation in the relative
utilization of dietary protein for energy and biosynthesis by the mussel Myrilus ~&/is. Mar. Ecol. Prog. Ser., 126: 177-184.
Kreeger, D.A.. Hawkins. A.J.S. and Bayne. B.L.. 1996. Use of dual-labeled microcapsules to discern the
physiological fates of assimilated carbohydrate, protein carbon. and protein nitrogen in suspension-feeding
organisms. Limnol. Oceanogr., 41: 208-215.
Langdon, C.J., 1989. Preparation and evaluation of protein microcapsules for a marine suspension-feeder, the Pacific oyster Crasaostrrn gigus. Mar. Biol., 102: 217-224.
Langdon. C.J. and Bolton, E.T.. 1984. A microparticulate diet for a buspension-feeding bivalve mollusc, Crussos~recr rirginicrr (Gmelin). J. Exp. Mar. Biol. Ecol., 82: 239-258.
Langdon, C.J. and DeBevoise, A.E., 1990. Effect of microcapsule type on delivery of dietary protein to a marine suspension-feeder, the oyster Cras.rostrea gigm. Mar. Biol., 105: 437-443.
Langdon, C.J. and Siegfried, C.A., 1984. Progress in the development of artificial diets for bivalve filter feeders. Aquaculture, 39: I35- 153.

300 J. Knauer, P.C. Southgate/Aquaculturee 153 (1997) 291-300
Langdon, C.J. and Waldock, M.J., 1981. The effect of algal and artificial diets on the growth and fatty acid
composition of Crassostrea gigas spat. J. Mar. Biol. Assoc. UK, 61: 431-448.
Numaguchi, K. and Nell, J.A., 1991. Effects of gelatin-acacia microcapsule and algal meal supplementation of
algal diets on growth rates of Sydney rock oyster, Saccostrea commercialis (Iredale and Roughley), larvae. Aquaculture, 94: 65-78.
Parker, R.S. and Selivonchick, D.P., 1986. Uptake and metabolism of lipid vesicles from seawater by juvenile
Pacific oysters (Crassostrea gigas). Aquaculture, 53: 215-228.
Peirson, W.M., 1983. Utilization of eight algal species by the bay scallop, Argopecten irradians concentricus
(Say). J. Exp. Mar. Biol. Ecol., 68: l-1 1.
Robinson, A., 1992a. Dietary supplements for the reproductive conditioning of Crassostrea gigas kumamoto
(Thunbergl. I. Effects of gonadal development, quality of ova and larvae through metamorphosis. J.
Shellfish Res., 11: 437-441.
Robinson, A., 1992b. Dietary supplements for the reproductive conditioning of Crassostrea gigas kumamoto
(Thunbergl. II. Effects on glycogen, lipid and fatty acid content of broodstock oysters and eggs. J. Shellfish Res., 1 I: 443-447.
Southgate, P.C., 1988. Use of microencapsulated diets in the culture of giant clam larvae. In: J.W. Copland
and J.S. Lucas (Editors). Giant Clams in Asia and the Pacific, ACIAR Monograph No. 9, Canberra, pp.
155-160.
Southgate. P.C. and Lou, D.C., 1995. Improving the n-3 HUFA composition of Artemia using microcapsules
containing marine oils. Aquaculture, 134: 91-99.
Thompson, P.A. and Harrison, P.J., 1992. Effects of monospecific algal diets of varying biochemical
composition on the growth and survival of Pacific oyster (Crassostrea gigas) larvae. Mar. Biol., 113:
645-654.
Ward, J.E. and Targett, N.M., 1989. Influence of marine microalgal metabolites on the feeding behaviour of
the blue mussel Mytilus edulis. Mar. Biol., 101: 313-321.
Ward, J.E., Cassell, H.K., MacDonald, B.A., 1992. Chemoreception in the sea scallop Placopecten magellani-
cus (Gmelin). I. Stimulatory effects of phytoplankton metabolites on clearance and ingestion rates. J. Exp.
Mar. Biol. Ecol., 163: 235-250.
Webb, K.L. and Chu, F.-L.E., 1983. Phytoplankton as a food source for bivalve larvae. In: G.D. Pruder, C.J.
Langdon and D. Conklin (Editors). Proceedings of the Second International Conference on Aquaculture
Nutrition: Biochemical and Physiological Approaches to Shellfish Nutrition. October 27-29, 1981,
Rehoboth Beach, Delaware. World Mariculture Society, Special Publication No. 2, Louisiana State
University, Baton Rouge, LA, pp. 272-291.
Widdows, J. and Bayne, B.L., 1971. Temperature acclimation of Mytilus edulis with reference to its energy
budget. J. Mar. Biol. Assoc. UK, 51: 827-843.