toxic dinoflagellate alexandrium catenella revealed with nuclear
TRANSCRIPT

1
Unexpected Genetic Diversity among and within Populations of the 1
Toxic Dinoflagellate Alexandrium catenella Revealed with Nuclear 2
Microsatellite Markers 3
4
Estelle Masseret,*1 Daniel Grzebyk,
1 Satoshi Nagai,
2 Benjamin Genovesi,
1,2 Bernard Lasserre,
1 5
Mohamed Laabir,1 Yves Collos,
1 André Vaquer,
1 Patrick Berrebi
3 6
7
Université Montpellier II, CNRS, Ifremer, UMR 5119 Ecosystèmes lagunaires 5119, 8
cc093, place E. Bataillon, 34095 Montpellier, France 1 9
National Research Institute of Fisheries and Environment of the Inland Sea, Maruishi 2-10
17-5, Hatsukaichi, Hiroshima 739-0452, Japan 2 11
Université Montpellier II, CNRS, UMR 5554 Institut des Sciences de l'Evolution, cc065, 12
place E. Bataillon, 34095 Montpellier, France 3 13
14
KEY WORDS: 15
Alexandrium catenella; dinoflagellates; microsatellites; population genetics; phylogenetic 16
diversity. 17
RUNNING TITLE: French-Japanese A. catenella population genetics 18
19
*Corresponding author. mailing address: UMR UM2-CNRS-Ifremer 5119 Ecosystèmes 20
lagunaires, Université Montpellier II, cc093, Place Eugène Bataillon, 34095 Montpellier, France. 21
Phone: 33 467144762. Fax: 33 467143719, E-mail: [email protected] 22
ACCEPTED
Copyright © 2009, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.Appl. Environ. Microbiol. doi:10.1128/AEM.01686-08 AEM Accepts, published online ahead of print on 5 February 2009
Copyright © 2009, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.Appl. Environ. Microbiol. doi:10.1128/AEM.01686-08 AEM Accepts, published online ahead of print on 5 February 2009

2
Since 1998, blooms of Alexandrium catenella associated with paralytic shellfish 23
poisoning have been repeatedly reported in Thau lagoon (French Mediterranean coastal area). 24
From data obtained on ribosomal DNA markers, it has been suggested that these strains could be 25
closely related to the Japanese Temperate Asian ribotype of the Temperate Asian clade. In order 26
to get more insights into their origin, we carried out a genetic analysis of 61 Mediterranean and 27
23 Japanese strains on both ribosomal and microsatellite markers. Whereas the phylogeny on 28
ribosomal markers tended to confirm the previous findings, the analysis of microsatellite 29
sequences revealed an unexpected partition between the French and Japanese populations. This 30
analysis also highlighted a large intra-specific diversity that was not detected with the classical 31
rDNA markers. The Japanese strains are divided into two differentiated A. catenella lineages: the 32
Sea of Japan lineage and the east coast lineage which includes populations from the Inland Sea 33
and the Pacific Ocean. A. catenella strains isolated in Thau lagoon would belong to another 34
lineage. These findings indicate that microsatellite markers are probably better suited to 35
investigate the population genetics of this worldwide distributed species. Finally, the application 36
of the population genetics concepts available for macroorganisms could support new paradigms 37
for speciation and migration in phytoplankton assemblages. 38
39
40
41
ACCEPTED

3
Toxic phytoplankton blooms, also known as harmful algae blooms (HABs), have long 42
been considered as sporadic phenomena. These world-wide distributed events occur particularly 43
in coastal and confined waters when HAB species bloom or rise up from their “hidden status” (3). 44
Their increasing occurrence is a significant and expanding threat to health, the fisheries and 45
shellfish industries (22). Many toxic phytoplankton species involved in these toxic events belong 46
to the class Dinophyceae (50). In French coastal waters, the genus Alexandrium Halim is 47
represented by A. minutum in the North Brittany coast and A. catenella (Whedon and Kofoid) 48
Balech in the NW Mediterranean coast; both produce neurotoxins responsible for paralytic 49
shellfish poisoning (PSP) (19). In the French Mediterranean coast, A. catenella was first observed 50
in 1998 in the Thau lagoon, and was responsible for a first toxic event. Since then, the species has 51
repeatedly produced toxic blooms (years 2001, 2003, 2004, 2005, 2007) particularly in autumn 52
and/or spring periods during which the shellfish trade, a significant economic activity in this area, 53
was forbidden (26, 30). Moreover, it is regularly detected along the neighbouring Catalan coast 54
(52). 55
Previous studies based on ribosomal RNA gene sequencing have highlighted the existence 56
of a complex of morphologically similar species named the “tamarense complex”. Within this 57
complex including A. tamarense, A. catenella and A. fundyense, the ribotypes were grouped into 58
phylogeographic clusters closely related to geographic zones defined by the main oceanic coastal 59
areas (19, 45, 47). These studies accredited the obviousness of A. catenella spread by human 60
activities such as shipping or shellfish aquaculture (7). Lilly et al (29) assumed that A. catenella 61
could have recently been introduced into Thau lagoon through ballast water from Japan. Indeed, 62
PSP events of A. catenella have frequently caused damage in Japanese coastal waters over the 63
last 25 years (55). This hypothesis was recently supported after a clear link was established 64
ACCEPTED

4
between NW Mediterranean and Japanese A. catenella strains based on phylogenetic analyses of 65
the 5.8S rDNA gene and ITS regions (40). 66
Microsatellite markers, commonly used to analyse population genetics of various 67
terrestrial and aquatic macroorganisms (25), have recently been developed on phytoplankton 68
species: diatoms Ditylum brightwellii (41-43) and Pseudo-nitzschia pungens (12), and 69
coccolithophore Emiliania huxleyi (24). They are recognized as valuable markers for better 70
understanding the genetic structure of phytoplankton populations over both temporal and spatial 71
scales. Among the Dinophyceae, many microsatellite markers have recently been developed in 72
order to gain more insight on the development and dynamics of blooms of the genus 73
Alexandrium: A. tamarense, North American and Temperate Asian ribotypes (2, 32), A. minutum 74
(34) and A. catenella (35). In a first study of the A. tamarense population, microsatellite markers 75
evidenced some correlation between genetic and geographic distances, and several segregated 76
populations, in coastal waters around Japan (33). 77
In the present survey, we investigated the inter- and intra- specific diversity of French 78
Mediterranean and Japanese strains using a comparative genetic analysis of 12 microsatellite 79
markers developed in A. catenella (35) with population genetics tools. Their phylogenetic 80
relationships were established using the ribosomal marker spanning the region ITS1 + 5.8S 81
rDNA + ITS2 + D1/D3 28S rDNA. This survey tries to measure the genetic diversity of French 82
and Japanese A. catenella strains, in order to explore the Japanese origin of the French 83
Mediterranean strains; it is also a first opportunity to check the ploidy level of the different 84
phases of this dinoflagellate’s life cycle. 85
86
87
88
ACCEPTED

5
MATERIALS AND METHODS 89
Sampling site and isolates collection. Thau lagoon is the deepest marine lagoon located 90
on the French Mediterranean coast (43° 24’ N-3° 36’ E) covering 75 km2 of which 1/5 is 91
occupied by shellfish farming structures (Fig.1). The lagoon is connected to the Mediterranean 92
Sea by three channels, one of them opening into the Sète international harbour. Water and 93
sediment samples were collected in the Angle Creek where the A. catenella blooms are usually 94
initiated. A. catenella cultures were developed from single vegetative cells (ACTEM) or single 95
resting cysts (ACTKM) (Table 1). Resting cysts were isolated from the first 3 cm of sediment 96
cores in April 2004. Their isolation and germination were performed as described in (17). 97
Planktonic vegetative cells were isolated from the bloom occurring in November 2004. A. 98
catenella cultures were grown in Enriched Sea Natural Water (23) at 20°C with a 12:12 h 99
light:dark photocycle at a photon flux density of ~100 µmol m-2
s-1
. After centrifugation at 1800 100
g for 3 minutes, cultures were frozen at -20°C. Japanese strains were isolated in four locations in 101
the Akasaki seaport in the Sea of Japan (35°51’ 11’’ N-133°65’69’’ E), Uchino-Umi 102
(34°22’03’’N-134° 60’ 57’’ E) and Kita Nada (34° 21’ 44’’ N-134° 44’ 86’’ E) in the Inland Sea 103
and Ago bay (34° 28’ 36’’ N-136°80’ 04’’E) on the Pacific coast (Fig.1 and Table 1). A 104
collection of 61 monoclonal cultures from Thau lagoon and 23 from Japanese waters and 105
sediment were analyzed in this study (Table 1). 106
Molecular methods. Total genomic DNA was extracted from monoclonal cultures 107
according to the classical phenol:chloroform method (44). Ribosomal DNA sequencing was 108
carried out on 23 selected A. catenella strains (11 from Thau Lagoon, 12 from Japan). A 109
ribosomal DNA fragment, spanning the region ITS1 + 5.8S rDNA + ITS2 + D1/D3 28S rDNA 110
(~1500 bp), was PCR-amplified with our novel primer 18S-ITS1-Ac-F 5’-111
CTTAGAGGAAGGAGAAGTCG-3’ (forward) specifically designed to cover the full length 112
ACCEPTED

6
sequence of the ITS1 region and the reverse primer 28S-D3B-R 5’-113
TCGGAGGGAACCAGCTACTA-3’ (reverse) (38). PCR amplification was performed using the 114
proof-reading DNA polymerase PrimeSTAR HS (Takara Bio Inc., Otsu, Shiga, Japan), with the 115
following conditions: 95°C for 2 min, 35 cycles (95°C, 30 s; 55°C, 15 s; 72°C, 2 min) and a final 116
extension at 72°C for 5 minutes. The PCR products checked on agarose electrophoresis gel and 117
revealing a single band were purified using the MinElute PCR Purification Kit (Qiagen, Hilden, 118
Germany). The purified PCR products were used as templates for direct sequencing using the two 119
PCR primers and an additional internal primer. Sequencing was performed by Macrogen (Seoul, 120
South Korea) using an ABI 3730XL Genetic Analyzer (Applied Biosystems). The sequence data 121
were assembled using the software package Vector NTI (Invitrogen, Carlsbad, CA, USA). 122
Nucleotide sequence alignments, with data retrieved from Genbank, were performed using 123
ClustalX (51). The maximum likelihood phylogenetic analysis was carried out using PHYML 124
v2.4.4 (21) as follows: alignment spanning 515 A. catenella positions (ITS1 + 5.8S rDNA + 125
ITS2), 1000 bootstraps, GTR substitution model and an “invariant + Γ” among-site substitution 126
rate distribution model approximated by 8 category rates with all the parameters estimated by the 127
program. The accession numbers of the deposited sequences used in this analysis are provided in 128
the phylogenetic tree. 129
The microsatellite analysis used the twelve loci described in (35). The PCR reactions 130
were carried out in a total volume of 10 µl, containing 1 µl of 10X buffer, 2.5 mM MgCl2 131
(Promega), 0.2 mM of each dNTP (Invitrogen), 0.5 µM of each primer labelled either with CY5 132
or fluorescein (MWG-Biotech AG), 0.3 U of Taq polymerase (Sigma), 0.1 mg/ml of BSA 133
(Roche), 2 µl of DNA template (at about 5 ng/µl) and completed up to 10 µl with DNA-free water 134
(Sigma). Thermal cycling conditions consisted of an initial denaturation at 94°C for 10 min, 135
followed by 38 cycles including denaturation at 94°C for 30 s, annealing at 60°C for 30 s, 136
ACCEPTED

7
extension at 72°C for 1 min, and a final extension at 72°C for 5 min. Two µl of PCR mixture 137
from each strain were electrophoresized onto an 8% denaturing polyacrylamide gel (Bio-Rad), 138
along with a fluorescently labelled DNA ladder 100–600 bp (Promega). Labelled DNA fragments 139
were visualized with a FMBIO II fluorescent imaging system (Hitachi), and the allele sizes were 140
estimated using the FMBIOAnalysis 8.0 image analyser program (Hitachi). 141
Statistical analyses of microsatellite markers. The statistical analyses were performed 142
using the programme GENETIX (4). A series of multidimensional analyses (Factorial 143
Correspondence Analysis or FCA, (5) provided the overall genetic structuring of the samples. 144
The objective of this analysis was to represent each individual according to its complete 145
genotype. For this, the genotypic data were first coded according to the presence of each allele 146
with value 0 (allele absent), 1 (heterozygous for the allele), 2 (homozygous for the allele) (49). 147
The computation then aimed at finding composite axes which were a combination of the 148
variables and optimised the differences between the individuals analysed. The relationships 149
among individuals can be visualized on two or three axes. In order to compare genotypes of 150
different ploidies in the same analysis, we had to weight the haploid genotypes as 2, meaning that 151
a haploid genotype was given the same weight as the two alleles of a diploid heterozygote 152
genotype. The inertia values (i.e. the proportion of the total information contained by an axis, 153
given as a percent) along each axis, were shown to be equivalent to linear combinations of the 154
monolocus Fixation Index (Fst) values (20). Standard parameters of population genetics were 155
calculated for each group of strains: allele frequencies, P(0.95) and P(0.99) (proportion of 156
polymorphic loci) and A (mean number of alleles per locus) parameters. The observed 157
heterozygosity (Hobs) and unbiased expected heterozygosity (Hnb) (unbiased according to the 158
sample sizes) (36) were estimated for the diploid samples (cyst cultures). Regarding the F 159
statistics of Wright, the Fis value (f parameter of Weir and Cockerham (54)) was used to test the 160
ACCEPTED

8
departures from Hardy-Weinberg equilibrium of the cyst collection (expected to be diploid), and 161
the Fst value (f of Weir and Cockerham (54)) was used to measure the differentiation between 162
samples using both cells and cysts. For these tests, the significance of departure from zero (null 163
hypothesis) was estimated by (5000) permutation tests, using GENETIX options. The ploidy 164
level of each cycle phase has seldom been tested and microsatellite data can be used to estimate 165
this level according to the heterozygote genotypes observed. However, apparent heterozygotes 166
can also result from false monoclonal cultures. Indeed, in spite of thorough verifications during 167
the cell isolation procedure, two swimming cells could sometimes be isolated, thus giving rise to 168
a false monoclonal culture. Here, we compared the proportion of heterozygote genotypes in 169
cultures of expected haploid cells and in cultures of expected diploid resting cysts. 170
Nucleotide sequence accession numbers. Sequence data have been deposited with 171
GenBank accession numbers FM211455 to FM211477. 172
ACCEPTED

9
RESULTS 173
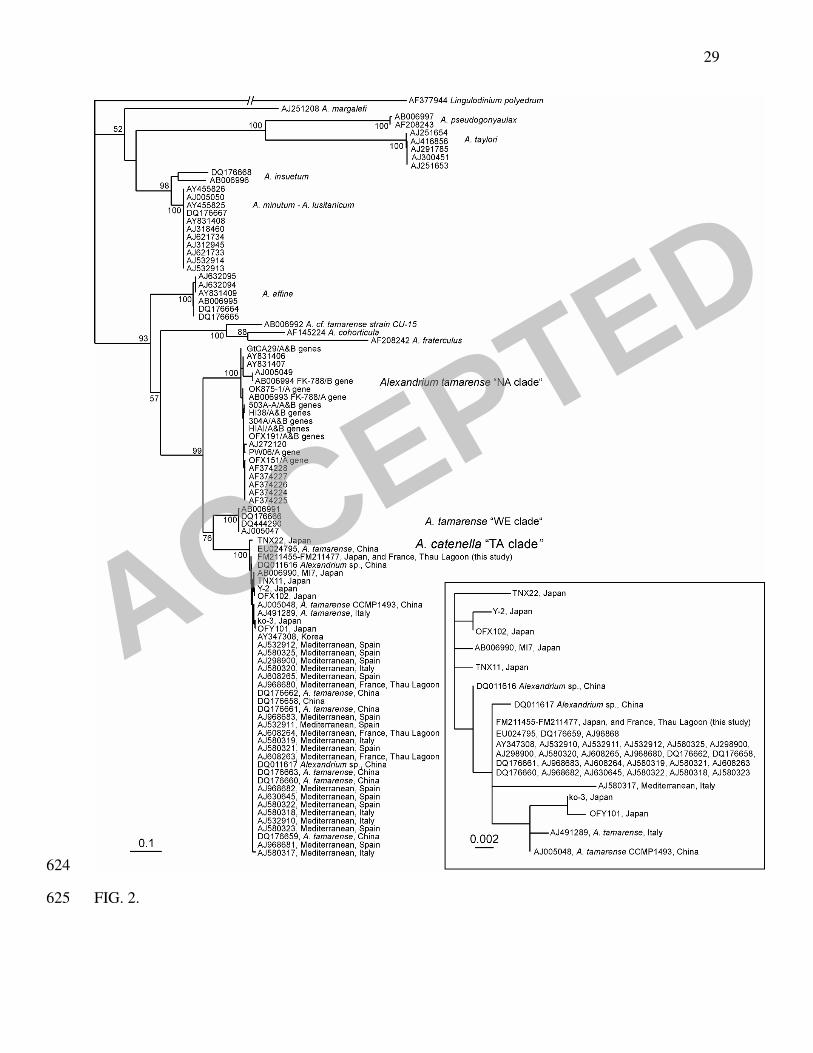
Ribotypes analysis. Though direct sequencing of rDNA amplicons revealed some 174
polymorphisms, as reported by Lilly et al (28), the ribosomal DNA sequences spanning the (ITS1 175
+ 5.8S rDNA + ITS2 + 28S rDNA D1/D3) region (~1500 bp) obtained from the 23 selected A. 176
catenella strains from Thau Lagoon and Japan were identical. Over the (ITS1 + 5.8S rDNA + 177
ITS2) region (515 bp), the consensus sequences were identical, or nearly, to most other A. cf. 178
catenella sequences deposited since 2000. This is featured in the phylogenetic tree of the 179
Alexandrium genus showing our new data among Genbank deposited sequences (Fig. 2). 180
Microsatellite analyses. The average proportion of missing genotypes per locus was 181
12.2% and varied from 4.5% (loci Acat02 and 49) to 24.7% (locus Acat10). Missing genotypes 182
represented 11.6% and 13.8% in the French and Japanese strains, respectively. This lack of 183
amplification is not likely to originate from cross-priming (the primers were designed from a 184
Japanese strain DNA) but rather from sample preservation problems and/or null alleles. 185
Microsatellite genotypes obtained in cultures originating from planktonic cells and germinated 186
resting cyst cultures exhibited few heterozygote patterns. Though Chi2 test was not significant, 187
the rate of heterozygous genotypes in the French Mediterranean isolates, was higher in cyst-188
derived cultures (5/114 genotypes = 4.4%) than in cultures obtained from planktonic cells (8/477 189
= 1.7%). The number of investigated Japanese strains was not large enough to make the same 190
comparison. In the following calculations, heterozygous cysts were considered as true 191
heterozygotes, whereas heterozygous planktonic/vegetative cells were assumed to be artefactual 192
(e.g. possibly due to polyclonal cultures mistakenly produced during isolation or to an occasional 193
duplication and mutation of a microsatellite target region). In the French strains, heterozygotes were 194
detected in loci Acat14, 49 and in loci Acat14, 16, 20, 34 and 50, in cyst- and vegetative cell-195
derived cultures, respectively. In the Japanese samples, only one heterozygous genotype was 196
ACCEPTED

10
observed at locus Acat14 in a vegetative cell-derived culture. For cyst samples only, expected to 197
contain a diploid genetic reservoir, the Hardy-Weinberg Equilibrium (HWE, genotype 198
frequencies expected in panmixia) was tested by estimating the significance of Fis departure from 199
zero. The Fis values were between 0.87 and 1.00, and the permutation tests (5000 permutations) 200
showed that the heterozygote deficit was highly significant (p<0.001). The genetic diversity of 201
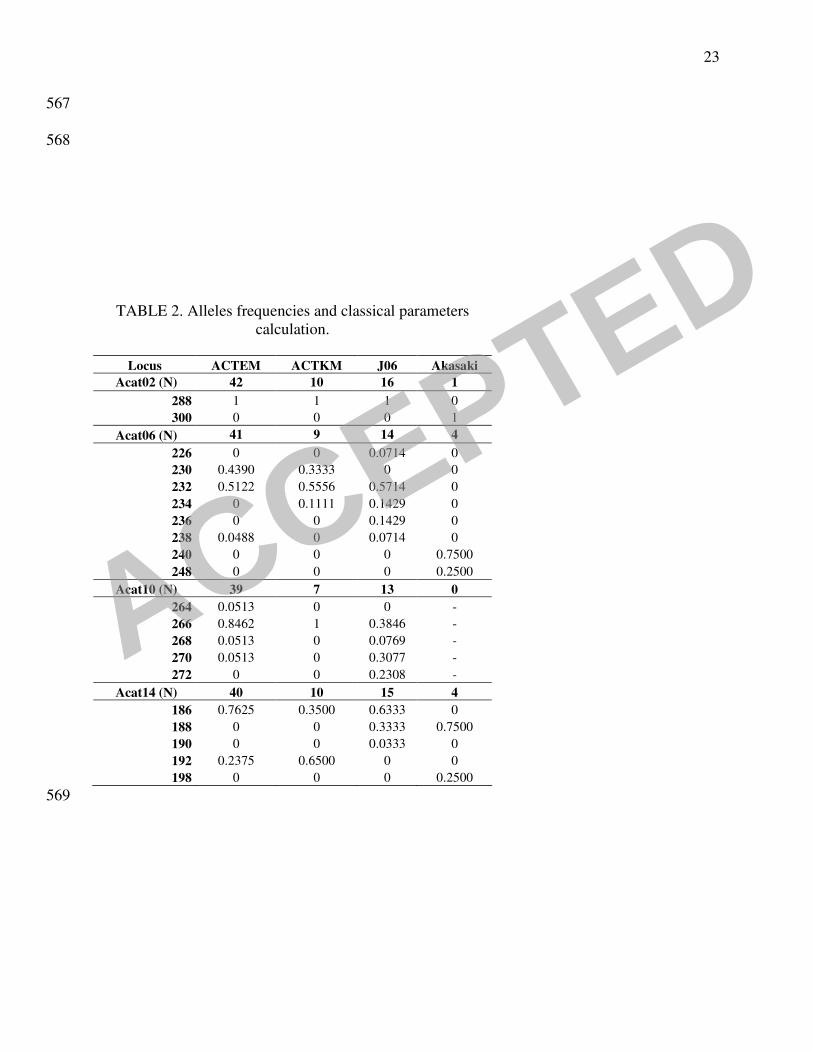
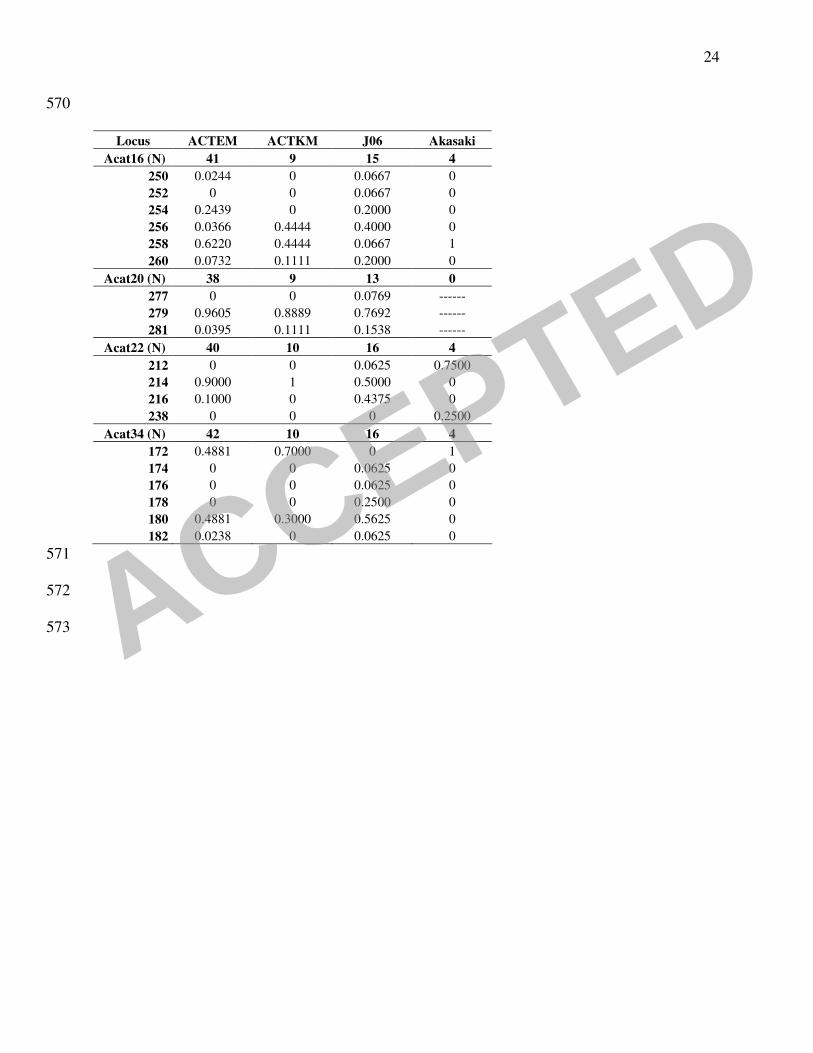
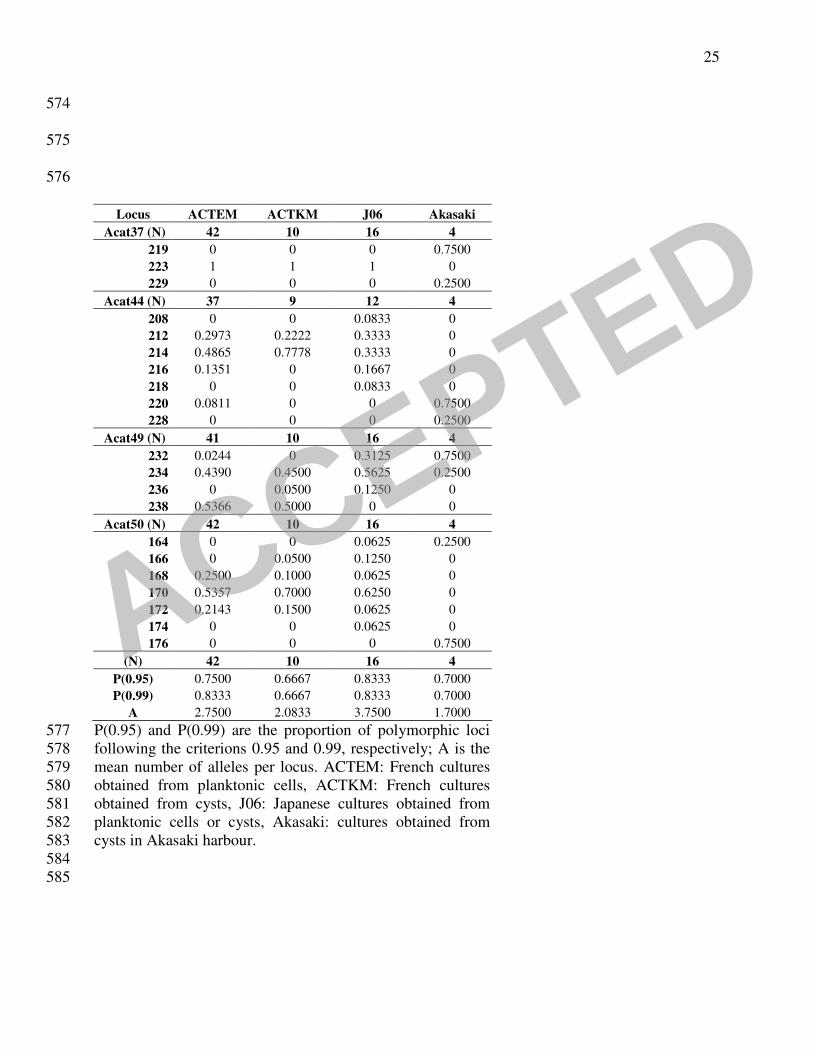
the A. catenella samples was investigated through several methods. The classical descriptive 202
parameters of population genetics showed that the genetic polymorphism was higher in Japan 203
than in Thau (Table 2). The Akasaki strains were considerably different from other Japanese and 204
Mediterranean strains, showing ten private alleles in seven loci (Acat02, Acat06, Acat14, Acat22, 205
Acat37, Acat 44 and Acat50). Parameter A (mean number of alleles per locus) which is 206
dependent on the sample size, reached 2.7 (42 strains) and 2.1 (10), respectively, for vegetative 207
cells and cysts sampled in 2004 in Thau, whereas the main Japanese sample (Akasaki excluded 208
because of its too small sample size) reached 3.7 with only 16 strains. The value obtained from 209
the strains isolated in Akasaki seaport could not be considered because of its too small sample 210
size (6 strains). This group of strains was thereafter separated from the other Japanese ones 211
because of its high differentiation (see FCA paragraph, below). In order to avoid strong 212
distortions, only nearly complete genotypes were taken into account in the FCA analyses, i.e. 213
with 3 (i.e. 25%) missing genotypes (either due to failed PCR or null alleles) at the most among 214
the 12 loci. As a result, the numbers of strains used in the analyses were as follows: 20 Japanese 215
strains, and 42 Thau 2004 vegetative cells strains (but due to the transformation of heterozygotes 216
by the creation of virtual haploid strains, this number rose up to 53), 10 Thau 2004 cyst strains 217
(heterozygote genotypes were retained since they were considered as true), plus ATTL01, ACT1 218
ACT2 and ACT3 (Table 1). The FCA of the whole dataset showed that the Japanese strains from 219
Akasaki were genetically very different from all the other strains (diagram not shown); the entire 220
ACCEPTED

11
analyse was thus devoted to the Akasaki/non-Akasaki differences. After excluding Akasaki, the 221
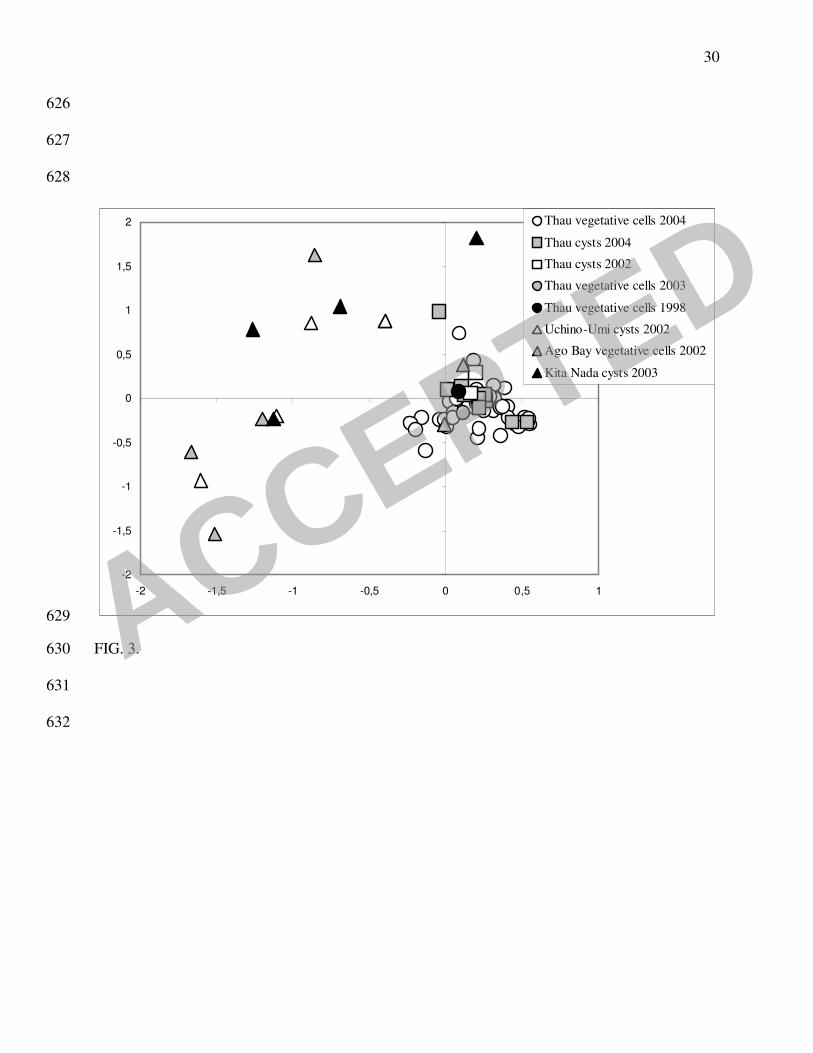
FCA showed a clear separation between the French Mediterranean and Japanese strains in the 222
first FCA axis. The dispersion was much larger in the Japanese isolates, indicating a higher 223
polymorphism (Fig. 3). However, a cyst strain from Thau seemed to be closer to the Japanese 224
strains, and one strain from Ago Bay fell among the Thau population (a second one was 225
discriminated along the third axis - not shown). When analyzed alone, the 61 Thau strains 226
showed no heterogeneity in the population structure (diagram not shown). 227
Fst estimations were calculated and the statistical evaluation of the significance of the 228
departure from zero tested by 5000 permutations (Table 3). These values indicated that (i) there 229
was no differentiation between vegetative cells and cysts from Thau in 2004 (θ=0.04, ns), (ii) the 230
differentiation was significant to highly significant between France and the Pacific coast of Japan 231
(0.15<θ<0.29), (iii) the differentiation was higher between France and the Sea of Japan 232
(0.55<θ<0.58), (iv) the differentiation between the two Japanese coasts was significant only 233
between Akasaki and Ago Bay (θ=0.48). The fact that this significant value was lower that non 234
significant values of θ between Akasaki and the other Pacific stations (0.52< θ <0.54) indicated 235
that this distortion was probably due to very small sample sizes. Finally, it was clear that there 236
was no differentiation among Pacific stations in Japan (-0.02< θ<0.05, ns). 237
238
DISCUSSION 239
Phylogenetic relationships among A. catenella ribotypes. Until now, the differentiation 240
among Alexandrium lineages had mainly been investigated using ribosomal sequences (19, 39, 241
40, 45-48). These analyses showed that the Alexandrium lineages are notably related to their 242
geographic origin defined as the North American, Western European, Mediterranean, Temperate 243
Asian, Tropical Asian and Tasmanian ribotypes. Since 1998, when a toxic event of A. catenella 244
ACCEPTED

12
was observed for the first time in Thau lagoon, many studies have been carried out in order to 245
understand its ecophysiology and growth strategies (8-10, 26). Among the causation theories 246
advanced by (22) concerning HAB expansions, the appearance of A. catenella in Thau lagoon 247
through dispersal of resting cysts in ballast water (29) and in transplanted shellfish stocks from 248
aquaculture cultivation (26) has been considered as a putative source of this new and recurrent 249
outbreak. Lilly et al (29) first described the strains isolated in 1998 as belonging to the Japanese 250
ribotype of the Temperate Asian clade, and recently renamed them as Group IV in the A. 251
tamarense-complex phylogeny (28). According to the phylogenetic analysis, isolates from 252
November 2004 are similar to those isolated in 1998, 2002 and 2003. The phylogenetic tree (Fig. 253
3), gathering most of the published ITS1-5.8S rDNA-ITS2 sequences from the A. 254
tamarense/catenella/fundyense species complex including the 23 new French and Japanese A. 255
catenella strains, pointed out that the ribosomal operon could not elucidate the intraspecific 256
variability within and between A. catenella populations from a common geographic origin. 257
Furthermore, the A. catenella cluster contains some A. tamarense morphotypes from China. The 258
phylogenic grouping of isolates from this complex is consistent with the ribotype clades but not 259
with the morpho-species that form the complex (39). They are in accordance with the recently 260
developed trends (28, 39, 53) which question the phylogenetic, proteomic, biological and 261
morphological species definition within the A. tamarense complex. Because ribosomal genes are 262
ubiquitous, orthologous (15) and display little polymorphism due to a limited mutation rate (27), 263
they should still be useful and valuable markers for the species level. However, they may not 264
represent the most judicious and accurate molecular markers to analyse the intraspecific diversity 265
essential to understand migration and speciation phenomena within A. catenella assemblages. 266
Genetic diversity within the Temperate Asian A. catenella clade evidenced with 267
microsatellite markers. In contrast, it appears that microsatellite markers may be appropriate 268
ACCEPTED

13
genetic markers to investigate the intraspecific diversity of the A. catenella ribotype. For the first 269
time, Japanese and Mediterranean A. catenella isolates belonging to the Japanese Temperate 270
Asian ribotype of the Temperate Asian clade, were compared with these highly polymorphic 271
genetic markers designed from Japanese A. catenella strains. Thus, the analysis distinctly showed 272
that the studied Japanese samples were divided into two differentiated “catenella” lineages: the 273
Sea of Japan lineage (including the differentiated sample from Akasaki seaport), and the east 274
coast lineage which included populations from the Inland sea (Uchino-Umi, Kita Nada) and the 275
Pacific Ocean (Ago Bay) (Fig.1). This is the first time such heterogeneity within Japanese A. 276
catenella populations is described. It also appeared that A. catenella strains isolated in Thau 277
lagoon could belong to another lineage. Several discordances (private alleles) and Fst values were 278
always significant between France and Japan (Table 3). 279
Genetic diversity within the A. catenella lineage from Thau lagoon. Microsatellite 280
markers revealed a relatively low genetic diversity in whole Mediterranean isolates from Thau 281
lagoon (ATTL01, ACT1-2-3, ACTEM, ACTKM) as compared to Japanese populations. 282
According to these markers, these Thau strains could belong to the same lineage. This could be 283
explained by several causes: i) the introduction of a small exogenous population which would 284
have found its ecological niche; ii) the specific environmental characteristics of Thau lagoon (i.e. 285
high temperature (up to 26°C) and high salinity (up to 39-40 PSU in the summer) could have 286
selected adapted ecotypes. Despite a weak recorded diversity with microsatellites, the A. 287
catenella individuals within the population would not homogeneously participate in the formation 288
of the following generation because no panmixia was apparent according to Fis values. When the 289
following generation is not the product of a random pooling in the initial gametic pool (16) pre-290
zygotic selection should be considered. Such a low polymorphism and absence of panmixia in 291
cysts could be explained by a differential reproductive success. (i) If very few cells breed resting 292
ACCEPTED

14
cysts during the bloom, a founding effect would be generated at each reproduction. (ii) The 293
original settlement of the Thau population would stem from very few individuals of unknown 294
origin. (iii) Resting cysts sampled in 2004, which can settle for several years in the sediment, 295
could not represent a population at equilibrium. Thus, more accurate genetic markers are required 296
for the description of the diversity within A. catenella population and consequently the 297
understanding of its adaptative strategy and the success of its recurrent dominancy. 298
Ploidy level and artefacts. Many dinoflagellates, including the genus Alexandrium, have 299
a complex life cycle with a predominant vegetative haploid (n) phase with asexual replication and 300
a short sexual diploid (2n) phase (6, 18, 31). Haploid cells divide mitotically in the water column, 301
and some cells differentiate into a gametic stage. Compatible gametes (n) fuse in the water 302
column to form a short-living diploid zygote called planozygote (2n) which afterward transforms 303
into a resting cyst (2n). After a dormancy period, the cyst germinates and regenerates a 304
planomeiocyte (2n) which undergoes the meiotic division to give new haploid cells (n) that 305
develop as vegetative cells (n). The distinction between haploid and diploid cells through 306
population genetics can be made by examining the genotypes of polymorphic nuclear markers. A 307
polymorphic nuclear marker will show heterozygous genotypes in a diploid individual but not in 308
a haploid one. In our study, the genotypes were obtained from a monoclonal culture of a 309
“swimming cell” or a resting cyst. Isolating a single cell or cyst is a critical step where false 310
heterozygotes can be obtained if two cells (or more) are inadvertently isolated together, or a 311
planomeiocyte (2n) or planozygote (2n) is isolated rather than a haploid vegetative cell. 312
In the present study, the rate of heterozygote genotypes was higher in cyst cultures (4.4%) 313
than in planktonic cell cultures (1.7%) where the number of vegetative cells genotypes obtained 314
was more than four times higher than that of cyst genotypes. Despite the fact that this difference 315
in heterozygote genotypes is was not significant, we could attribute the 1.7% vegetative cell 316
ACCEPTED

15
heterozygote genotypes to mishandlings in the long process of monoclonal culture and/or a 317
percentage of planomeiocytes and planozygotes isolated in situ among the “swimming cells”. We 318
also considered that within the estimated 4.4% of cyst heterozygote genotypes, false 319
heterozygotes also occurred in the same proportion as in cell samples. This could show that the 320
analysed global diploid phase presents a low heterozygosity which has been overestimated (Ho < 321
0.04). This preliminary result is encouraging, but it is obvious that new highly polymorphic 322
microsatellites are necessary. This is why new markers were designed by the same team (37). 323
When analysing the expected Hardy-Weinberg equilibrium in the three cyst samples large 324
enough to allow the test, we observed an extreme Fis disequilibrium between 0.9 and 1 325
(p<0.001), meaning that only a few expected heterozygous genotypes were detected. Panmixia 326
(free sexuality) was not observed, highlighting the particular sexual crossing (i.e. heterothallism) 327
which occurred in the Alexandrium catenella populations. The fusion of compatible gametes 328
from the same clone (selfing, homothallism) results in the lack of effective recombination, whereas 329
the fusion of gametes from different
clones (outcrossing, heterothallism) may result in 330
recombination. Therefore, A. catenella from Thau lagoon may exhibit a heterothallic behaviour 331
as described by Yoshimatsu (56) for Japanese strains. The life cycle of A. catenella must be 332
studied in more details because sexual stages still remain unclear, contrasting with A. excavatum 333
(11), A. tamutum, A. minutum (14) and A. peruvianum (13), for which they are well described. 334
This knowledge is essential for the good understanding of the origin and occurrence of the toxic 335
events due to A. catenella. 336
Is the recent Mediterranean invasion of Japanese origin? We are far from having a 337
complete description of Japanese A. catenella polymorphism. The higher polymorphism and 338
structure of the Japanese isolates confirm that east-Asia is the natural origin of distribution of A. 339
catenella. In contrast, the weak genetic diversity among the Mediterranean strains from Thau 340
ACCEPTED

16
Lagoon could be related to a recent introduction of genotypes whose exact origin remains to be 341
determined. These preliminary results suggest that the hypothesis of the Japanese origin of A. 342
catenella blooming in Thau lagoon (French Mediterranean) should be precisely explored. 343
According to the observed and potential differentiation of the species in Asian coasts, more 344
strains from both regions must be analysed before drawing a conclusion. This study urges one to 345
compare other A. catenella isolates from the Mediterranean area with the same markers in order 346
to (i) determine if these populations belong to the same lineage and (ii) understand the genetic 347
distribution of the species in the Mediterranean basin. The genetic analysis will have to use more 348
microsatellite markers (37) and more samples, not limited to Japan. This genetic study must be 349
extended to other Asiatic sites and geographic areas in the world in order to clarify the origin of 350
A. catenella established in Thau lagoon. We consider the present survey as the first step of a 351
multi-year project prospecting A. catenella all around the world in order to understand the long 352
distance exchanges related to human activity. 353
These findings should bring insights into other environmental factors affecting the niche 354
adaptations of these microorganisms in situ. Large variations in single-cell characteristics and 355
physiological capabilities (adaptation to temperature, salinity, nutrient assimilation, allelopathic 356
activity) can be expected within and between lineages, which could have ecological 357
consequences, such as HAB events. Such a genetic approach could underpin a new paradigm for 358
speciation and migration in phytoplankton assemblages. 359
360
ACKNOWLEDGEMENTS 361
We thank Pierre-Alexandre Gagnaire for his statistical help; Annie Pastoureaud (Ifremer 362
LER/LR-Sète) for her helpful collaboration, Patrick Gentien (Ifremer DYNECO-Brest) for his 363
ACCEPTED

17
continuous support and the Ifremer staff (LER/LR-Sète) for its technical assistance. We would 364
also like to thank the anonymous reviewers for their helpful comments and suggestions. 365
This research was supported by grants from the French National Programme “Ecosphère 366
Continentale et Côtière” (EC2CO-PNEC) and the Agence Nationale de la Recherche (ANR -05-367
BLAN-0219 XPressFloral), and by the program ALCAT of Ifremer. 368
REFERENCES 369
1. Adachi, M., Y. Sako, and Y. Ishida. 1996. Analysis of Alexandrium (Dinophyceae) 370
species using sequences of the 5.8S ribosomal DNA and internaltranscribed spacer 371
regions. Journal of Phycology 32:424-432. 372
2. Alpermann, T. J., U. E. John, L. K. Medlin, K. J. Edwards, P. K. Hayes, and K. M. 373
Evans. 2006. Six new microsatellite markers for the toxic marine dinoflagellate 374
Alexandrium tamarense. Molecular Ecology Notes 6:1057-1059. 375
3. Anderson, D. M. 1989. Toxic algal blooms and red tides: a global perspective, p. 11-16. 376
In T. Okaichi, D. M. Anderson, and T. Nemoto (ed.), Red Tides: Biology, Environmental 377
Science and Toxicology. Elsevier. 378
4. Belkhir, K., P. Borsa, J. Goudet, L. Chikhi, and F. Bonhomme 1998, posting date. 379
GENETIX, logiciel sous Windows TM pour la génétique des populations. Laboratoire 380
Génome et Populations. [Online.] 381
5. Benzecri, J. P. 1982. L'analyse des données. II. L'analyse des correspondances. Dunod, 382
Paris. 383
6. Blackburn, S. I., and N. Parker. 2005. Microalgal life cycles: encystment and 384
excystment, p. 399-417. In R. A. Anderson (ed.), Algal Culturing Techniques. Elsevier 385
Academic Press, Amsterdam. 386
7. Bolch, C. J. S., and M. F. de Salas. 2007. A review of the molecular evidence for ballast 387
water introduction of the toxic dinoflagellates Gymnodinium catenatum and the 388
Alexandrium "tamarensis complex" to Australasia. Harmful Algae 6:465-485. 389
8. Collos, Y., C. Gagne, M. Laabir, A. Vaquer, P. Cecchi, and P. Souchu. 2004. 390
Nitrogenous nutrition of Alexandrium catenella (Dinophyceae) in cultures and in Thau 391
lagoon, southern France. Journal of Phycology 40:96-103. 392
9. Collos, Y., M. Lespilette, A. Vaquer, M. Laabir, and A. Pastoureaud. 2006. Uptake 393
and accumulation of ammonium by Alexandrium catenella during nutrient pulses. African 394
Journal of Marine Science 28:313-318. 395
10. Collos, Y., A. Vaquer, M. Laabir, E. Abadie, T. Laugier, A. Pastoureaud, and P. 396
Souchu. 2007. Contribution of several nitrogen sources to growth of Alexandrium 397
catenella during blooms in Thau lagoon, southern France. Harmful Algae 6:781-789. 398
11. Destombe, C., and A. Cembella. 1990. Mating-type determination, gametic recognition 399
and reproductive success in Alexandrium excavatum (Gonyaulacales, Dinophyta), a toxic 400
red-tide dinoflagellate. Phycologia 29:316-325. 401
12. Evans, K. M., S. F. Kuhn, and P. K. Hayes. 2005. High levels of genetic diversity and 402
low levels of genetic differentiation in North Sea Pseudo-nitzschia pungens 403
(Bacillariophyceae) populations. Journal of Phycology 41:506-514. 404
ACCEPTED

18
13. Figueroa, R. I., I. Bravo, and E. Garces. 2008. The significance of sexual versus 405
asexual cyst formation in the life cycle of the noxious dinoflagellate Alexandrium 406
peruvianum. Harmful Algae 7:653-663. 407
14. Figueroa, R. I., E. Garces, and I. Bravo. 2007. Comparative study of the life cycles of 408
Alexandrium tamutum and Alexandrium minutum (Gonyaulacales, Dinophyceae) in 409
culture. Journal of Phycology 43:1039-1053. 410
15. Fitch, W. M. 1970. Toward defining the tree of maximum parsimony, p. 160-178. In G. 411
F. Estabrook (ed.), Eighth International conference on numerical taxonomy, Freeman: San 412
Francisco. 413
16. Genovesi-Giunti, B. 2006. Initiation, maintien et récurrences des efflorescences toxiques 414
d'Alexandrium catenella (Dinophyceae) dans une lagune méditerranéenne (Thau, France): 415
rôle du kyste dormant. Ph.D. thesis. Montpellier, France, Montpellier, France. 416
17. Genovesi-Giunti, B., M. Laabir, and A. Vaquer. 2006. The benthic resting cyst: A key 417
actor in harmful dinoflagellate blooms - A review. Vie Et Milieu-Life and Environment 418
56:327-337. 419
18. Giacobbe, M. G., and X. M. Yang. 1999. The life history of Alexandrium taylori 420
(Dinophyceae). Journal of Phycology 35:331-338. 421
19. Guillou, L., E. Nezan, V. Cueff, E. E. L. Denn, M. A. Cambon-Bonavita, P. Gentien, 422
and G. Barbier. 2002. Genetic diversity and molecular detection of three toxic 423
dinoflagellate genera (Alexandrium, Dinophysis, and Karenia) from French coasts. Protist 424
153:223-238. 425
20. Guinand, B. 1996. Use of a multivariate model using allele frequency distributions to 426
analyse patterns of genetic differentiation among populations. Biological Journal of the 427
Linnean Society 58:173-195. 428
21. Guindon, S., and O. Gascuel. 2003. A simple, fast, and accurate algorithm to estimate 429
large phylogenies by maximum likelihood. Systematic Biology 52:696-704. 430
22. Hallegraeff, G. M. 1993. A review of harmful algal blooms and their apparent global 431
increase. Phycologia 32:79-99. 432
23. Harrison, P. J., R. E. Waters, and F. J. R. Taylor. 1980. A broad spectrum artificial 433
seawater medium for coastal and open ocean phytoplankton. Journal of Phycology 16:28-434
35. 435
24. Iglesias-Rodriguez, M. D., O. M. Schofield, J. Batley, L. K. Medlin, and P. K. Hayes. 436
2006. Intraspecific genetic diversity in the marine coccolithophore Emiliania huxleyi 437
(Prymnesiophyceae): The use of microsatellite analysis in marine phytoplankton 438
population studies. Journal of Phycology 42:526-536. 439
25. Jarne, P., and P. J. L. Lagoda. 1996. Microsatellites, from molecules to populations and 440
back. Trends in Ecology & Evolution 11:424-429. 441
26. Laabir, M., Z. Amzil, P. Lassus, E. Masseret, Y. Tapilatu, R. De Vargas, and D. 442
Grzebyk. 2007. Viability, growth and toxicity of Alexandrium catenella and 443
Alexandrium minutum (Dinophyceae) following ingestion and gut passage in the oyster 444
Crassostrea gigas. Aquatic Living Resources 20:51-57. 445
27. Lecointre, G., H. Philippe, H. L. Van Le, and H. Le Guyader. 1993. Species sampling 446
has a major impact on phylogenetic inference. Molecular Phylogenetics and Evolution 447
2:205-24. 448
28. Lilly, E. L., K. M. Halanych, and D. M. Anderson. 2007. Species boundaries and 449
global biogeography of the Alexandrium tamarense complex (Dinophyceae). Journal of 450
Phycology 43:1329-1338. 451
ACCEPTED

19
29. Lilly, E. L., D. M. Kulis, P. Gentien, and D. M. Anderson. 2002. Paralytic shellfish 452
poisoning toxins in France linked to a human-introduced strain of Alexandrium catenella 453
from the western Pacific: evidence from DNA and toxin analysis. Journal of Plankton 454
Research 24:443-452. 455
30. Masselin, P., A. Z., A. E., E. Nézan, C. Le Bec, A. Carreras, C. Chiantella, and P. 456
Truquet. 2001. Presented at the Harmful Algal blooms 2000, Proceedings of the IXth 457
international Conference Harmful Algal Blooms, Hobart, Tasmania. 458
31. Montresor, M. 1995. The life history of Alexandrium pseudogonyaulax (Gonyaulacales, 459
Dinophyceae). Phycologia 34:444-448. 460
32. Nagai, S., C. Lian, M. Hamaguchi, Y. Matsuyama, S. Itakura, and T. Hogetsu. 2004. 461
Development of microsatellite markers in the toxic dinoflagellate Alexandrium tamarense 462
(Dinophyceae). Molecular Ecology Notes 4:83-85. 463
33. Nagai, S., C. Lian, S. Yamaguchi, M. Hamaguchi, Y. Matsuyama, S. Itakura, H. 464
Shimada, S. Kaga, H. Yamauchi, Y. Sonda, T. Nishikawa, C. H. Kim, and T. 465 Hogetsu. 2007. Microsatellite markers reveal population genetic structure of the toxic 466
dinoflagellate Alexandrium tamarense (Dinophyceae) in Japanese coastal waters. Journal 467
of Phycology 43:43-54. 468
34. Nagai, S., L. McCauley, N. Yasuda, D. L. Erdner, D. M. Kulis, Y. Matsuyama, S. 469
Itakura, and D. M. Anderson. 2006. Development of microsatellite markers in the toxic 470
dinoflagellate Alexandrium minutum (Dinophyceae). Molecular Ecology Notes 6:756-471
758. 472
35. Nagai, S., M. Sekino, Y. Matsuyama, and S. Itakura. 2006. Development of 473
microsatellite markers in the toxic dinoflagellate Alexandrium catenella (Dinophyceae). 474
Molecular Ecology Notes 6:120-122. 475
36. Nei, M. 1978. Estimation of average heterozygosity and genetic distance from a small 476
number of individuals. Genetics 89:583-590. 477
37. Nishitani, G., S. Nagai, E. Masseret, C. Lian, S. Yamaguchi, N. Yasuda, S. Itakura, 478
D. Grzebyk, P. Berrebi, and M. Sekino. 2007. Development of compound microsatellite 479
markers in the toxic dinoflagellate Alexandrium catenella (Dinophyceae). Plankton and 480
Benthos Research. 481
38. Nunn, G., B. Theisen, B. Christensen, and P. Arctander. 1996. Simplicity-correlated 482
size growth of the nuclear 28S ribosomal RNA D3 expansion segment in the crustacean 483
order isopoda. Journal of Molecular Evolution 42:211-223. 484
39. Penna, A., S. Fraga, M. Maso, M. G. Giacobbe, I. Bravo, E. Garces, M. Vila, E. 485
Bertozzini, F. Andreoni, A. Luglie, and C. Vernesi. 2008. Phylogenetic relationships 486
among the Mediterranean Alexandrium (Dinophyceae) species based on sequences of 487
5.8S gene and Internal Transcript Spacers of the rRNA operon. European Journal of 488
Phycology 43:163-178. 489
40. Penna, A., E. Garces, M. Vila, M. G. Giacobbe, S. Fraga, A. Luglie, I. Bravo, E. 490
Bertozzini, and C. Vernesi. 2005. Alexandrium catenella (Dinophyceae), a toxic 491
ribotype expanding in the NW Mediterranean Sea. Marine Biology 148:13-23. 492
41. Rynearson, T. A., and E. V. Armbrust. 2000. DNA fingerprinting reveals extensive 493
genetic diversity in a field population of the centric diatom Ditylum brightwellii. 494
Limnology and Oceanography 45:1329-1340. 495
42. Rynearson, T. A., and E. V. Armbrust. 2004. Genetic differentiation among 496
populations of the planktonic marine diatom Ditylum brightwellii (Bacillariophyceae). 497
Journal of Phycology 40:34-43. 498
ACCEPTED

20
43. Rynearson, T. A., and E. V. Armbrust. 2005. Maintenance of clonal diversity during a 499
spring bloom of the centric diatom Ditylum brightwellii. Molecular Ecology 14:1631-500
1640. 501
44. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: a laboratory 502
manual. Cold Spring Harbor Laboratory Press, New York. 503
45. Scholin, C. A., and D. M. Anderson. 1994. Identification of group-specific and strain-504
specific genetic-markers for globally distributed Alexandrium (dinophyceae) .1. RFLP 505
analysis of SSU ribosomal-RNA genes. Journal of Phycology 30:744-754. 506
46. Scholin, C. A., and D. M. Anderson. 1996. LSU rDNA-based RFLP assays for 507
discriminating species and strains of Alexandrium (Dinophyceae). Journal of Phycology 508
32:1022-1035. 509
47. Scholin, C. A., M. Herzog, M. Sogin, and D. M. Anderson. 1994. Identification of 510
group-specific and strain-specific genetic-markers for globally distributed Alexandrium 511
(dinophyceae) .2. Sequence-analysis of a fragment of the LSU ribosomal-RNA gene. 512
Journal of Phycology 30:999-1011. 513
48. Sebastian, C. R., S. M. Etheridge, P. A. Cook, C. O'Ryan, and G. C. Pitcher. 2005. 514
Phylogenetic analysis of toxic Alexandrium (Dinophyceae) isolates from South Africa: 515
implications for the global phylogeography of the Alexandrium tamarense species 516
complex. Phycologia 44:49-60. 517
49. She, J. X., M. Autem, G. Kotoulas, N. Pasteur, and F. Bonhomme. 1987. Multivariate 518
analysis of genetic exchanges between Solea aegyptiaca and Solea senegalensis (Teleosts, 519
Soleidae). Biological Journal of the Linnean Society 32:357-371. 520
50. Sournia, A. 1995. Red tide and toxic marine phytoplankton of the world ocean: an 521
inquiry into biodiversity, p. 103-112. In P. Lassus, G. Arzul, E. Erard-Le Denn, P. 522
Gentien, and C. Marcaillou-Le Baut (ed.), Harmful Marine Algal Blooms. Lavoisier, 523
Intercept Ltd, Paris. 524
51. Thompson, J. D., T. J. Gibson, F. Plewniak, F. Jeanmougin, and D. G. Higgins. 1997. 525
The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment 526
aided by quality analysis tools. Nucleic Acids Research 25:4876-4882. 527
52. Vila, M., E. Garces, M. Maso, and J. Camp. 2001. Is the distribution of the toxic 528
dinoflagellate Alexandrium catenella expanding along the NW Mediterranean coast. 529
Marine Ecology-Progress Series 222:73-83. 530
53. Wang, D. Z., L. Lin, H. F. Gu, L. L. Chan, and H. S. Hong. 2008. Comparative studies 531
on morphology, ITS sequence and protein profile of Alexandrium tamarense and A. 532
catenella isolated from the China Sea. Harmful Algae 7:106-113. 533
54. Weir, B. S., and C. C. Cockerham. 1984. Estimating F-statistics for the analysis of 534
population-structure. Evolution 38:1358-1370. 535
55. Yoshida, T., Y. Sako, and A. Uchida. 2001. Geographic differences in paralytic shellfish 536
poisoning toxin profiles among Japanese populations of Alexandrium tamarense and 537
Alexandrium catenella (Dinophyceae) Phycological Research 29:13-21. 538
56. Yoshimatsu, S. 1981. Sexual reproduction of Protogonyaulax catenella in culture I. 539
Heterothallism. Bulletin of Plankton Society of Japan 28 131-139. 540
541
542
ACCEPTED

21
TABLE CAPTIONS 543
544
TABLE 1. Designation and location from which the French and Japanese A.catenella strains 545
analysed in the study were isolated. 546
547
TABLE 2. Alleles frequencies and classical parameters calculation. P(0.95) and P(0.99) are the 548
proportion of polymorphic loci following the criterions 0.95 and 0.99 respectively; A is the mean 549
number of alleles by locus. ACTEM: French cultures obtained from planktonic cells, ACTKM: 550
French cultures obtained from cysts, J06: Japanese cultures obtained from planktonic cells or 551
cysts, Akasaki: cultures obtained from cysts in Akasaki harbour. 552
553
TABLE 3. Fst values calculated according to the Weir and Cockerham (1984) θ estimator. Using 554
GENETIX software, 5000 permutations allowed to calculate the p values which were not 555
significant (ns), significant at 0.05 level (*) or at 0.01 level (**). 556 ACCEPTED
22
557
558
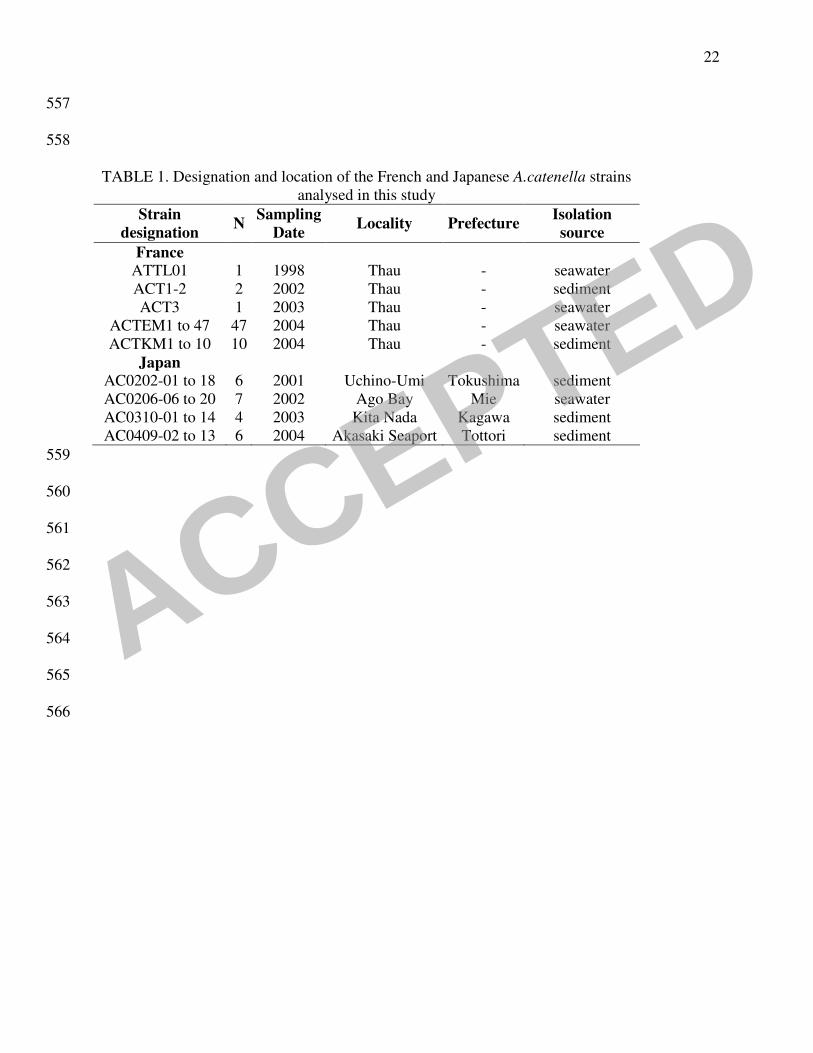
TABLE 1. Designation and location of the French and Japanese A.catenella strains
analysed in this study
Strain
designation N
Sampling
Date Locality Prefecture
Isolation
source
France
ATTL01 1 1998 Thau - seawater
ACT1-2 2 2002 Thau - sediment
ACT3 1 2003 Thau - seawater
ACTEM1 to 47 47 2004 Thau - seawater
ACTKM1 to 10 10 2004 Thau - sediment
Japan
AC0202-01 to 18 6 2001 Uchino-Umi Tokushima sediment
AC0206-06 to 20 7 2002 Ago Bay Mie seawater
AC0310-01 to 14 4 2003 Kita Nada Kagawa sediment
AC0409-02 to 13 6 2004 Akasaki Seaport Tottori sediment
559
560
561
562
563
564
565
566
ACCEPTED
23
567
568
TABLE 2. Alleles frequencies and classical parameters
calculation.
Locus ACTEM ACTKM J06 Akasaki
Acat02 (N) 42 10 16 1
288 1 1 1 0
300 0 0 0 1
Acat06 (N) 41 9 14 4
226 0 0 0.0714 0
230 0.4390 0.3333 0 0
232 0.5122 0.5556 0.5714 0
234 0 0.1111 0.1429 0
236 0 0 0.1429 0
238 0.0488 0 0.0714 0
240 0 0 0 0.7500
248 0 0 0 0.2500
Acat10 (N) 39 7 13 0
264 0.0513 0 0 -
266 0.8462 1 0.3846 -
268 0.0513 0 0.0769 -
270 0.0513 0 0.3077 -
272 0 0 0.2308 -
Acat14 (N) 40 10 15 4
186 0.7625 0.3500 0.6333 0
188 0 0 0.3333 0.7500
190 0 0 0.0333 0
192 0.2375 0.6500 0 0
198 0 0 0 0.2500
569
ACCEPTED
24
570
Locus ACTEM ACTKM J06 Akasaki
Acat16 (N) 41 9 15 4
250 0.0244 0 0.0667 0
252 0 0 0.0667 0
254 0.2439 0 0.2000 0
256 0.0366 0.4444 0.4000 0
258 0.6220 0.4444 0.0667 1
260 0.0732 0.1111 0.2000 0
Acat20 (N) 38 9 13 0
277 0 0 0.0769 ------
279 0.9605 0.8889 0.7692 ------
281 0.0395 0.1111 0.1538 ------
Acat22 (N) 40 10 16 4
212 0 0 0.0625 0.7500
214 0.9000 1 0.5000 0
216 0.1000 0 0.4375 0
238 0 0 0 0.2500
Acat34 (N) 42 10 16 4
172 0.4881 0.7000 0 1
174 0 0 0.0625 0
176 0 0 0.0625 0
178 0 0 0.2500 0
180 0.4881 0.3000 0.5625 0
182 0.0238 0 0.0625 0
571
572
573 ACCEPTED
25
574
575
576
Locus ACTEM ACTKM J06 Akasaki
Acat37 (N) 42 10 16 4
219 0 0 0 0.7500
223 1 1 1 0
229 0 0 0 0.2500
Acat44 (N) 37 9 12 4
208 0 0 0.0833 0
212 0.2973 0.2222 0.3333 0
214 0.4865 0.7778 0.3333 0
216 0.1351 0 0.1667 0
218 0 0 0.0833 0
220 0.0811 0 0 0.7500
228 0 0 0 0.2500
Acat49 (N) 41 10 16 4
232 0.0244 0 0.3125 0.7500
234 0.4390 0.4500 0.5625 0.2500
236 0 0.0500 0.1250 0
238 0.5366 0.5000 0 0
Acat50 (N) 42 10 16 4
164 0 0 0.0625 0.2500
166 0 0.0500 0.1250 0
168 0.2500 0.1000 0.0625 0
170 0.5357 0.7000 0.6250 0
172 0.2143 0.1500 0.0625 0
174 0 0 0.0625 0
176 0 0 0 0.7500
(N) 42 10 16 4
P(0.95) 0.7500 0.6667 0.8333 0.7000
P(0.99) 0.8333 0.6667 0.8333 0.7000
A 2.7500 2.0833 3.7500 1.7000
P(0.95) and P(0.99) are the proportion of polymorphic loci 577
following the criterions 0.95 and 0.99, respectively; A is the 578
mean number of alleles per locus. ACTEM: French cultures 579
obtained from planktonic cells, ACTKM: French cultures 580
obtained from cysts, J06: Japanese cultures obtained from 581
planktonic cells or cysts, Akasaki: cultures obtained from 582
cysts in Akasaki harbour. 583
584
585
ACCEPTED

26
586
587
588
TABLE 3. Fst values calculated according to the Weir and Cockerham
(1984) θ estimator. Using GENETIX software, 5000 permutations
allowed to calculate the p values which were not significant (ns),
significant at 0.05 level (*) or at 0.01 level (**).
ACTKM Uchino-Umi Ago Bay Kita Nada Akasaki
ACTEM 0.04326ns 0.26852** 0.14961** 0.26801** 0.55077**
ACTKM 0.27977** 0.17752** 0.29159* 0.58133*
Uchino-Umi -0.01687ns 0.05485ns 0.53721ns
Ago Bay 0.02373ns 0.48041ns
Kita Nada 0.52256ns
589
590
591
592
593
594 ACCEPTED

27
FIGURE CAPTIONS 595
596
597
598
599
600
601
602
603
604
605
606
607
608
609
610
611
612
613
614
FIGURE 1. Sampling locations in France and in Japan. (Top panel reprinted from Souchu et al.
[Mar. Ecol. Prog. Ser. 218:141-152, 2001] with permission of the publisher. Bottom panel
courtesy of Satoshi Nagai.)
FIGURE 2. Maximum likelihood phylogenetic tree of genus Alexandrium, with L. polyedrum as
an outgroup, using ribosomal DNA sequences (ITS1 + 5.8S rDNA + ITS2), spanning 515 A.
catenella nucleotide positions. The scale bar (bottom left corner) represent 0.1 nucleotide
substitution per site. Identical sequences have been obtained from 23 selected A. catenella strains
(12 from Japan, 11 from Thau Lagoon), which are also identical to most other sequences in this
clade. The tree displayed in the small frame (bottom right corner) represents the relative genetic
distance between the sequences belonging to the A. catenella clade; the scale bar (0.002
nucleotide substitution per site) is equal to one nucleotide difference for the analyzed sequences.
The sequence data for A. catenella strains TNX22, TNY11, Y-2, OFX102, OFY101, ko-3 and
MI7 (accession number AB006990) are from Adachi et al (1996) (1).
FIGURE 3. Diagram of the first plan of the CA (axis 1 and axis 2) including all samples
excepted those from Akasaki, considered as outliers due to their high genetic difference. The two
axes inertia reached 16.83% for a total of 59 axes.
ACCEPTED

28
613
614
615
616
617
618
619
620
FIG.1. 621
622
623
Ago Bay
UchinoUmi
Akasaki
seaport
450 km Kita Nada
ACCEPTED
29
624
FIG. 2. 625
ACCEPTED
30
626
627
628
-2
-1,5
-1
-0,5
0
0,5
1
1,5
2
-2 -1,5 -1 -0,5 0 0,5 1
Thau vegetative cells 2004
Thau cysts 2004
Thau cysts 2002
Thau vegetative cells 2003
Thau vegetative cells 1998
Uchino-Umi cysts 2002
Ago Bay vegetative cells 2002
Kita Nada cysts 2003
629
FIG. 3. 630
631
632
ACCEPTED