nutritional suitability of the dinoflagellate ceratium furcoides
TRANSCRIPT

Journal of Plankton Research Vol.18 no.3, pp.323-333,1996
Nutritional suitability of the dinoflagellate Ceratium furcoides forfour copepod species
Barbara SanterMax-Planck-Institute for Limnology, Postfach 165, D-24302 Plbn, FRG
Abstract The nutritional suitability of Ceratium furcoides for Eudiaplomus gracilis, Mesocyclopsleuckarti, Thermocyclops oithonoides and Cyclops abyssorum was studied by observing ingestion andassimilation. The dinoflagellate was ingested by female adults of all species. The calanoid copepodE.gracilis could not utilize Ceratium; mortality was high and no egg production was observed whenCeratium was the only food source. Mortality was low for the adult cyclopoids on this food and repro-duction indicated that Ceratium was assimilated. The first two copepodite instars of M. leuckarti werenot able to handle Ceratium, while older stages preyed on them. The dinoflagellate was not ingested byfemale M.leuckarti when its densities were low. Advanced copepodite stages of C.abyssorum devel-oped into adults on a diet of Ceratium only. Mesocyclops leuckarti females ingested Ceratium whenoffered a mixed food source of Ceratium and the rotifer Brachionus rubens, but the rotifer was posi-tively selected, even if its density was low. The results show that Ceratium provides a suitable foodsource for advanced copepodite instars and adult cyclopoid copepods, although it is not a preferredfood source.
Introduction
Dinoflagellates are generally believed to constitute poor food for most crus-taceans, due to their size and spiny shape (Pollingher, 1987). Conover (1978, citedex Nielsen, 1991), Marshall and Orr (1955), Elbrachter (1973), Graneli etal. (1989,cited ex Nielsen, 1991) and Hargrave and Geen (1970) found that marine copepodsdid not consume Ceratium spp. In a more recent study, the largest marine cope-pods were found to feed on Ceratium, while the smaller species could not ingestCeratium spp. (Nielsen, 1991).
In freshwater ecology, it is rare to find freshwater copepods grazing on dinofla-gellates. Brandl and Fernando (1979) found that the predatory stages of Meso-cyclops edax did not consume Ceratium. Karabin (1978) and Adrian (1988)reported that cyclopoid copepods ingest Ceratium occasionally. Hard to ingestdinoflagellates are believed to have little nutritional value (Williamson, 1984).
Recently, a growing number of studies have shown that soft-bodied flagellatesprovide a suitable food source for the survival, development and reproduction ofthe predaceous stages of freshwater cyclopoids (DeMott, 1990; Adrian and Frost,1993; Santer, 1993a; Santer and van den Bosch, 1994).
Owing to their ability to handle bulky prey items with a hard outer body cover-ing, like rotifers (Fryer, 1957a; Williamson, 1984,1986), copepods should be ableto feed on thecate dinoflagellates. In this study, the nutritional suitability of thedinoflagellate Ceratium furcoides for three cyclopoid and one calanoid copepodspecies was examined. Copepodite instars and adult cyclopoid and calanoid cope-pods were exposed to a Ceratium suspension, to test their ability to ingest andutilize the dinoflagellate. Copepodite development and reproduction were used asindicators of the assimilation of the dinoflagellate. To answer the question would
© Oxford University Press 323
Dow
nloaded from https://academ
ic.oup.com/plankt/article/18/3/323/1460996 by guest on 08 January 2022

B .Santer
Ceratium also be ingested in the presence of another prey, the dinoflagellate wasoffered to Mesocyclops together with the rotifer Brachionus rubens.
Method
Culturing and routine techniques
Ceratiumfurcoid.es (CCAP 1110/4, C.hirudinella fo. furcoides) came from the cul-ture collection of the Institute of Freshwater Ecology, The Windermere Lab-oratory, Ambleside, UK. Cryptomonas sp. (979-6) came from the culturecollection of G.Schlosser, University of Gdttingen, FRG. Both algae were culturedin batch cultures using modified WC medium (Guillard and Lorenzen, 1972).
To determine the carbon content of Ceratium cells, culture samples were filteredon pre-combusted glass fibre filters (Whatman GF/C) and dried at 60°C overnight.The carbon content was determined by combustion and IR measurement of CO2
(Krambeck et al., 1981). The number of cells in the culture was counted under aninverted microscope.
Copepods were sampled 2 days before each experiment from Schierensee, asmall eutrophic Holstein lake located near Plon. Ceratium spp. occur in high num-bers in this lake during the summer months (Santer, 1990). Copepods were pre-conditioned for 48 h to the light and temperature conditions used in the exper-iments, and adapted to the WC medium. They were starved for 24 h before theexperiment.
The length of the copepods was measured from the anterior end of the ceph-alothorax to the end of the furcal rami at 50x magnification. The carbon content ofthe cyclopoid copepods was estimated using the length-carbon relationship givenby Santer and van den Bosch (1994). For Eudiaptomusgracilis, carbon content wasestimated from dry weight using the mean specific carbon content given by Hessenand Lyche (1991). Dry weight was estimated using the length-dry weight relation-ship given by Bottrell et al. (1976).
Feeding experiments
The general design was to add a certain number of copepods to small bottles con-taining a Ceratium suspension. Copepods were allowed to feed on Ceratium for 1or more days. Three or four replicates were performed for each experiment, threeor four replicates without copepods served as controls. The bottles were kept in atemperature- and light-controlled room at 20 ± 0.5°C and at a daily light period of16 h. The bottles were continuously turned over (0.5 r.p.m.) on a tissue cultureroller to avoid aggregation of Ceratium and copepods. The experiments werestopped by adding a few drops of Lugol's solution to each bottle. Numbers of Cer-atium cells were counted, undamaged cells with visible cell content were regardedas living cells; dead cells (empty thecae) and cell fragments were also counted.Three cell fragments were counted as one dead cell. Samples containing low num-bers of cells were counted completely; at high densities (>20 cells ml"1), at leastthree subsamples per bottle containing 2, 5 or 10 ml were counted.
The effect of copepods was tested by comparing the numbers of living and deadCeratium with copepods (treatment) and the numbers of Ceratium without
324
Dow
nloaded from https://academ
ic.oup.com/plankt/article/18/3/323/1460996 by guest on 08 January 2022

Ceratium feeding by copepods
Table L Experimental design of feeding experiments of copepods on Ceratium. M: Mesocyclops leuck-arti; T: Thermocyclops oithonoides; A: Cyclops abyssorum; E: Eudiapiomus gracitis; C: Ceratium fur-coides; B: Brachionus rubens; C2-C5: copepodite stages 2-5
Experiment
Feeding ofdifferentspecies onCeratium
Feeding ofdifferent stageson Ceratium
Feeding ondifferentdensities ofCeratium
Feeding on afood mixturecontainingCeratium andBrachionus
Volume(ml)
20
50
50
50
Ceratium andBrachionusdensity (ml1)
5 C
15 C
3C11C21 C43 C87C
162 C
5C + 0.1 B5 C + 0.2 B5C + 1.0B
Copepods(species, stageand numberper vial)
M, 9, 5T, 9, 10A, 9, 3E, 9, 5
M, C2,200 , 2 0C4.20C5,15
M, 9, 3358
1518
M. 9. 333
Durationofexperiment(days)
4
1
1
1
Number ofreplicates(number ofcontrols)
4(4)
3343
(5)
3(3)
333
(3)(3 controlswithout M)
copepods (control). Ingestion of Ceratium was calculated from the differences intotal cell numbers (living and dead) between treatments and controls. The detailson the different feeding experiments are summarized in Table I.
To determine changes in the Ceratium population during the experimentalperiod, which might have been caused by cell growth or natural mortality, the num-bers of living and dead Ceratium cells in the controls before and after the exper-iment were counted. No significant changes in cell numbers were found during1 day. In the first experiment (duration of 4 days), changes in the controls were notexamined, since the aim of this experiment was to detect differences betweencopepod species. Prior to the selectivity experiment, an experiment was performedto test the influence of the rotifer Brachionus on Ceratium. No effect of Brachionuson Ceratium was found.
One-way ANOVAs, followed by post hoc tests, were used for analysing the dif-ferences between treatments and controls. When the rotifer Brachionus wasoffered together with Ceratium, a two-way ANOVA was performed testing theeffect of Mesocyclops and the effect of Brachionus density on Ceratium.
Development and reproduction
Advanced copepodite stages and adult copepods were kept in small glass bottles
325
Dow
nloaded from https://academ
ic.oup.com/plankt/article/18/3/323/1460996 by guest on 08 January 2022

B-Santer
(100 ml); the Ceratium density was ~16 cells ml"1, the numbers of copepods aregiven in Table III. Three replicates were performed for each copepod species. Oneadditional bottle of each species was kept without food and served as a control.Temperature and light conditions were the same as for the feeding experiment.After 4 days, the contents of the bottles were filtered on a 50 ̂ .m net and rinsed in aPetri dish. Dead animals were removed. Living copepods were narcotized with afew drops of carbonated water, and developmental stage and ovaries were moni-tored. Egg sacs were removed with a thin needle and copepods were transferred tofresh food media. After 7 days, developmental stages and egg production weremonitored again. Numbers of eggs per egg sac were counted and a few egg sacsfrom each species were pipetted into 50 ml glass bottles containing WC mediumand Cryptomonas sp. at a density of ~1 mg C I1. These bottles were checked forhatching nauplii and viability of nauplii.
Results
Species-specific feeding
Significant reductions (ANOVA, Bonferroni P < 0.001) in the total numbers ofCeratium (= living and dead cells) were observed for all tested copepods after the4 day experimental period. This indicates that Ceratium was ingested by cyclopoidand calanoid copepods (Figure 1). Cyclops abyssorum preyed on significantlymore cells per day than all other copepods, while Thermocyclops killed signifi-cantly fewer cells than all other species (ANOVA, Bonferroni P < 0.001). No sig-nificant differences were found between the cyclopoid copepod Mesocyclops andthe calanoid copepod Eudiaptomus concerning Ceratium mortality (difference inliving cells).
Significant differences were found between all copepod species when comparingonly the numbers of ingested Ceratium cells (ANOVA, Bonferroni P < 0.05). Thepresence of the smaller cyclopoid copepods Mesocyclops and Thermocyclopscaused a significant increase (ANOVA, Bonferroni P < 0.05) in the number ofdead cells and cell fragments during the 4 day period. Some Ceratium cells wereobviously only destroyed, but not ingested, by the small copepods. Only —75% ofthe destroyed cells were eaten by the Mesocyclops. Thermocyclops ingested lessthan half of the destroyed cells. This phenomenon was not found for the largecyclopoid copepod Cyclops abyssorum and the calanoid Eudiaptomus. Theyseemed to be able to ingest whole cells or they destroyed them, but also ingestedthe fragments.
Considering the average carbon content of one Ceratium cell (5.3 ng) and thecarbon content of the copepods (Table II), the carbon ingestion via Ceratium isonly 0.2-0.4% of the body carbon of the copepods. Since only ingested cells wereincluded in these estimations, the values might be slightly higher for Mesocyclopsand Thermocyclops, which might have sucked out some cells without ingesting thecell walls. On the other hand, cytoplasm might have been lost when breaking apartthe cells (sloppy feeding).
326
Dow
nloaded from https://academ
ic.oup.com/plankt/article/18/3/323/1460996 by guest on 08 January 2022
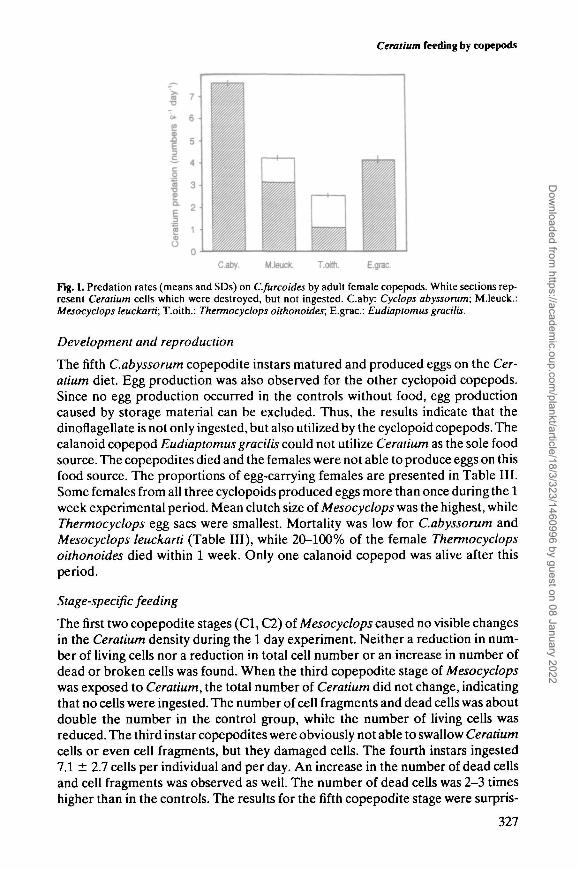
Ceratium feeding by copepods
Caby. M.leuck. T.oith. E.grac.
Fig. 1. Predation rates (means and SDs) on C.furcoides by adult female copepods. White sections rep-resent Ceratium cells which were destroyed, but not ingested. Caby: Cyclops abyssorum; M.leuck.:Mesocyclops leuckarti; T.oith.: Thcrmocyclops oithonoides; E.grac: Eudiaptomus gracilis.
Development and reproduction
The fifth C.abyssorum copepodite instars matured and produced eggs on the Cer-atium diet. Egg production was also observed for the other cyclopoid copepods.Since no egg production occurred in the controls without food, egg productioncaused by storage material can be excluded. Thus, the results indicate that thedinoflagellate is not only ingested, but also utilized by the cyclopoid copepods. Thecalanoid copepod Eudiaptomus gracilis could not utilize Ceratium as the sole foodsource. The copepodites died and the females were not able to produce eggs on thisfood source. The proportions of egg-carrying females are presented in Table III.Some females from all three cyclopoids produced eggs more than once during the 1week experimental period. Mean clutch size of Mesocyclops was the highest, whileThermocyclops egg sacs were smallest. Mortality was low for C.abyssorum andMesocyclops leuckarti (Table III), while 20-100% of the female Thermocyclopsoithonoides died within 1 week. Only one calanoid copepod was alive after thisperiod.
Stage-specific feeding
The first two copepodite stages (Cl, C2) of Mesocyclops caused no visible changesin the Ceratium density during the 1 day experiment. Neither a reduction in num-ber of living cells nor a reduction in total cell number or an increase in number ofdead or broken cells was found. When the third copepodite stage of Mesocyclopswas exposed to Ceratium, the total number of Ceratium did not change, indicatingthat no cells were ingested. The number of cell fragments and dead cells was aboutdouble the number in the control group, while the number of living cells wasreduced. The third instar copepodites were obviously not able to swallow Ceratiumcells or even cell fragments, but they damaged cells. The fourth instars ingested7.1 ± 2.7 cells per individual and per day. An increase in the number of dead cellsand cell fragments was observed as well. The number of dead cells was 2-3 timeshigher than in the controls. The results for the fifth copepodite stage were surpris-
327
Dow
nloaded from https://academ
ic.oup.com/plankt/article/18/3/323/1460996 by guest on 08 January 2022

B-Santer
Table II. Length (mean and SD) and mean carbon content of the female copepods and amount ofingested carbon (mean and SD). expressed also as mean percentage of body carbon
Copepod Length (mm) Carbon content Ingestion (ng C %species (jig) female1 day-')
Mesocyclops 1.1 ±0.08 3.6 16.6 ± 3.5 0 4Thermocyclops 0.9 ± 0.07 2.4 5.7 ± 1.0 0.2Cyclops 1.9 ±0.12 11.5 40.3 + 0.4 0.4Eudiaptomus 1.4 ±0.21 7 3 21.9 ± 0.9 0.3
ing. In contrast to the fourth copepodite instar, the total number of Ceratium didnot change, indicating no ingestion of cells. The number of empty and broken cellsincreased, thus the C5 copepodites might have sucked out Ceratium cells (seeDiscussion).
Feeding on different densities of Ceratium
At low Ceratium densities (2.6 and 11.3 cells ml"'), no influence of copepod feedingwas found. Neither the number of living cells nor the total cell numbers (living cells+ dead cells + cell fragments) were significantly different from the controls afterMesocyclops females were present for 1 day. When the density was a bit higher(21.6 cells ml"1), the total number of Ceratium was not significantly different fromthe controls when Mesocyclops was present, but a significant difference betweenthe treatments with copepods and the controls was found when only the number ofliving cells was considered. This indicates that cells were killed by the copepodsand the cell content might have been ingested. At higher densities (>43.5 cellsml"1), total cell number was reduced compared to the total cell number of the con-trols, indicating an uptake by the copepods. In addition, destroyed cells wereobserved (Figure 2). The amount of empty cells and cell fragments ranged between4 and 13% of the Ceratium mortality. At the highest Ceratium density (161 cellsml"'), one Mesocyclops female ingested an average of 209 cells during 1 day. Thisresults in a carbon uptake of 1.1 ng or 31 % of its body carbon per day.
Since the curve in Figure 2 showed no saturation plateau within the tested foodconcentration, the copepods should be able to feed on even more Ceratium cellsper day at higher densities. The clearance rates show a slight decrease from 1.8 mlday1 at a density of 43.5 cells ml1 to 1.3 ml day1 at a density of 161 cells ml1.
Feeding on Ceratium in the presence of Brachionus
When the rotifer B.rubens was offered to the Mesocyclops females together withCeratium, the dinoflagellates were still eaten by the copepods (P = 0.0036). Brach-ionus was always preferred to Ceratium, even at its lowest density (Table IV). TheCeratium ingestion fluctuated strongly between the replicates in this experiment.The mean numbers of Ceratium eaten by the copepods were not influenced statisti-cally by the rotifer density (P = 0.13). The numbers of Brachionus eaten by onecopepod female increased as their density increased.
328
Dow
nloaded from https://academ
ic.oup.com/plankt/article/18/3/323/1460996 by guest on 08 January 2022

Ceratium feeding by copepods
Table III. Egg production and mortality of copepods on a diet of Ceratium after 1 week. M: Meso-cyclops leuckarti; T: Thermocyclops oithonoider. A: Cyclops abyssorum; E: Eudiaptomus gracilis. C,:nth copepodite instars. In the controls (C), copepods were not fed
Copepodspecies
MMMM(C)
TTTT(C)
AAAA(C)
EEEE(C)
Number andins tar
1 91 91 91 9
1 91 91 91 9
6 97 97 97 9
10 910 910 910 9
1(5,4Q16\4C,16\4C,1(5,4C,
1 d, 4O,_,16, 4C,.,16, 4C,_j1<5,4CV,
Proportion ofegg-carryingfemales
0.831.430.570
1.10.50.40
1.02.01.00
0000
Proportion of deadanimals
0000.29
0.20.50.60.6
0000
1.01.00.81.0
Clutch size(mean ± SD)
13.5 d1 3 J d11.1 d-
3.8 d4.4 d3.9 d
-
12.012.012.5-
_---
t2.8t 2.4t 5.7
t 1.7t 1.4t 1.6
Discussion
The data presented here show that C.furcoides can be utilized by all three fresh-water cyclopoids. The dinoflagellate was ingested and assimilated by cyclopoidfemales and older copepodites.
These results are contradictory to the findings of Brandl and Fernando (1979)who reported that the predatory stages of the freshwater copepod M.edax did notingest Ceratium when fed an almost pure fraction of this alga. In contrast, Karabin(1978) reported that M.leuckarti consumed C.hirudinella at low rates, when fedwith phytoplankton containing 90% Ceratium. Dinoflagellates are believed to behard to ingest and to have little nutritional value. Williamson (1984) found that thedinoflagellate Peridinum was ingested by M.edax, but not digested. In the presentstudy, the cyclopoids were able to metabolize Ceratium, which is indicated bydevelopment and reproduction of the cyclopoids.
To my knowledge, no information is available in the literature regarding theability of freshwater calanoid copepods to utilize dinoflagellates. Since only onecalanoid species was tested in this study, no general assessment on the food valueof Ceratium for freshwater calanoid copepods is possible.
Most marine calanoid copepods did not ingest dinoflagellates (Marshall andOrr, 1955; Hargrave and Geen, 1970, Elbrachter, 1973; Conover, 1978, cited exNielsen, 1991; Graneli etal, 1989, cited ex Nielsen, 1991; Nielsen, 1991). Only a fewcalanoid copepods, i.e. Temora longicomis (Schnack, 1976) and the large speciesCentropages hamatus and Centropages typicus (Nielsen, 1991), fed on Ceratiumspp. Calculated from data from Peterson et al. (1991), Nielsen (1991) concludedthat C.hamatus and C.typicus are able to exploit the dinoflagellate and to increase
329
Dow
nloaded from https://academ
ic.oup.com/plankt/article/18/3/323/1460996 by guest on 08 January 2022
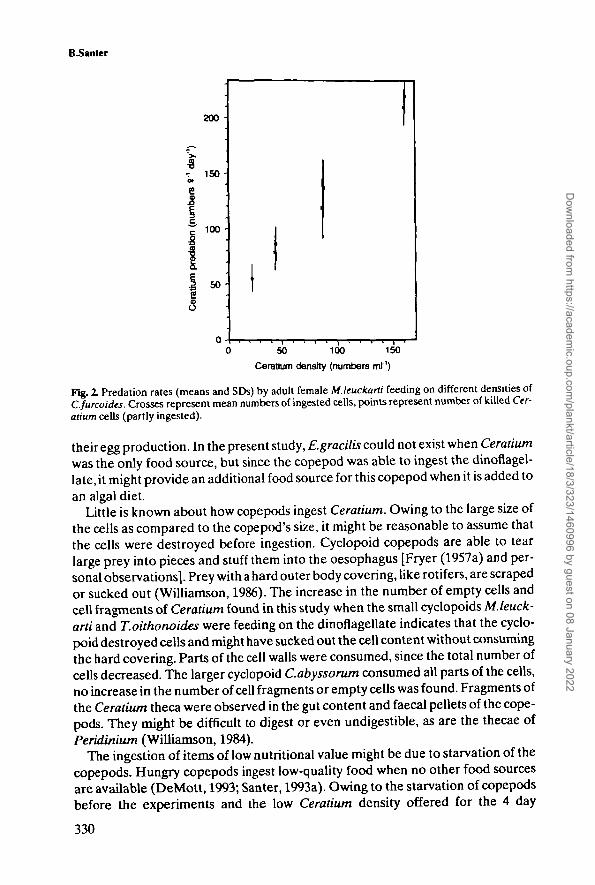
B-Santer
50 100 150Ceratium density (numbers ml"1)
Fig. 2. Predation rates (means and SDs) by adult female M.leuckarti feeding on different densities ofC.furcoides. Crosses represent mean numbers of ingested cells, points represent number of killed Cer-atium cells (partly ingested).
their egg production. In the present study, E.gracilis could not exist when Ceratiumwas the only food source, but since the copepod was able to ingest the dinoflagel-late, it might provide an additional food source for this copepod when it is added toan algal diet.
Little is known about how copepods ingest Ceratium. Owing to the large size ofthe cells as compared to the copepod's size, it might be reasonable to assume thatthe cells were destroyed before ingestion. Cyclopoid copepods are able to tearlarge prey into pieces and stuff them into the oesophagus [Fryer (1957a) and per-sonal observations]. Prey with a hard outer body covering, like rotifers, are scrapedor sucked out (Williamson, 1986). The increase in the number of empty cells andcell fragments of Ceratium found in this study when the small cyclopoids M.leuck-arti and T.oithonoides were feeding on the dinoflagellate indicates that the cyclo-poid destroyed cells and might have sucked out the cell content without consumingthe hard covering. Parts of the cell walls were consumed, since the total number ofcells decreased. The larger cyclopoid C.abyssorum consumed all parts of the cells,no increase in the number of cell fragments or empty cells was found. Fragments ofthe Ceratium theca were observed in the gut content and faecal pellets of the cope-pods. They might be difficult to digest or even undigestible, as are the thecae ofPeridinium (Williamson, 1984).
The ingestion of items of low nutritional value might be due to starvation of thecopepods. Hungry copepods ingest low-quality food when no other food sourcesare available (DeMott, 1993; Santer, 1993a). Owing to the starvation of copepodsbefore the experiments and the low Ceratium density offered for the 4 day
330
Dow
nloaded from https://academ
ic.oup.com/plankt/article/18/3/323/1460996 by guest on 08 January 2022

Ceratium feeding by copepods
Table IV. Ingestion rates (means and SDs) on Ceratiumand selectivity indices (Jakobs, 1974) for Brachionus, D&
Food density of Ceratiumand Brachionus (numberml-1)
5C -5C + 0.1 B5 C + 0.2 B5C + 1.0B
and Brachionus by adult female M.leuckani,- C: Ceratium furcoides; B: Brachionus nibens
Ingestion rates (numbers female1
day1)
Ceratium
49.2 ± 18.614.3 ± 7.645.0 ± 33.421.7 ± 4.2
Brachionus
13 ±0.52.7 ± 0.8
14.7 ± 1.2
Selectivity index
0.620.210.56
experimental period, the copepods might have been hungry and thus ingested theCeratium thecae.
Mesocyclops ingested Ceratium even in the presence of an animal prey (Brach-ionus). The availability of algae and invertebrate prey was shown to be advan-tageous for the carnivorous stages of cyclopoids (Adrian and Frost, 1993; Santer,1993a; Hansen and Santer, 1995).
All cyclopoid copepods reproduced when Ceratium was the sole food source.The suitability of algae for egg production was also found for other cyclopoid cope-pods, but reproductive success was low (Adrian and Frost, 1993; Santer, 1993a;Hansen and Santer, 1995). When Ceratium was the only food source, the clutchsizes of all three cyclopoids were within the range found in their natural environ-ment (see Santer, 1990) and eggs were fertile.
Under natural conditions, Ceratium might serve as an additional food source forcopepods, which might be important especially when convenient food sources, i.e.soft-bodied flagellates, rotifers, calanoid and cyclopoid nauplii or cladoceran juv-eniles and eggs (Fryer, 1957b; Gophen, 1977; Karabin, 1978; Williamson, 1991;Gliwicz and Stibor, 1993; Santer, 1993a,b; van den Bosch and Santer, 1993; Gliwiczand Lampert 1994; Santer and van den Bosch, 1994), are deficient. This might hap-pen in early spring, when the number of predaceous copepods is high and suitableprey is scarce (Santer, 1990). Ceratium appears in the pelagial at this time and thehigh mortality of Ceratium which can be observed in spring (B.Meyer, personalcommunication) might be due to copepod predation. The decrease in cyclopoidnumbers in early summer due to the beginning of the summer diapause mightfavour the population increase of the dinoflagellate. Ceratium is not grazed bycladocerans or by most of the rotifers which dominate the zooplankton in summer(Sommer et al., 1986; Pollingher, 1987); it can develop high population densitiesduring this season. Cyclopoid numbers are low during this period and predationdoes not have a significant influence on Ceratium densities (B.Santer, unpublishedresults).
Acknowledgements
B.Meyer provided the Ceratium cultures, G.FuBmann cultured the Brachionus,H.Buhtz counted most of the samples and H.Mumm gave me helpful statistical
331
Dow
nloaded from https://academ
ic.oup.com/plankt/article/18/3/323/1460996 by guest on 08 January 2022

B.Santer
advice. M.Gliwicz, B.Meyer and an anonymous reviewer gave important advice onthe manuscript. N.Zehrbach corrected the English text. I am grateful to all of them.
ReferencesAdrian,R. (1988) Untersuchungen zur herbivoren und carnivoren Ernahrungsweise von Cyclops
kotensis und C.vicinus (Crustacea, Copepoda). PhD Thesis, University of Berlin, FRG, 188 pp.Adrian,R. and Frost.T.M. (1993) Omnivory in cyclopoid copepods: comparison of algae and invert-
ebrates as food for three, differently sized species. J Plankton Res., 15,643-658.Bottrell.H.H., Duncan,A., Gliwicz,Z.M., Grygierek.E., Herzig,A., Hillbncht-Ilkowska.A., Kur-
asawa.H., Larsson,P. and Weglenska,T. (1976) A review of some problems in zooplankton pro-duction studies. Norw. J. Zool, 24,419—456.
Brandl.Z. and Fernando.C.H. (1979) The impact of predation by the copepod Mesocydops edax(Forbes) on zooplankton in three lakes in Ontario, Canada. Can. J. Zool., 57,940-942.
DeMott.W.R. (1990) Retention efficiency, perceptual bias, and active choice as mechanisms of foodselection by suspension-feeding zooplankton. In Hughes.R.N. (ed.). Behavioural Mechanism ofFood Selection. NA TO ASI Series G, Ecological Sciences. Springer, Heidelberg, pp. 569-594.
DeMott.W.R. (1993) Hunger-dependent diet selection in suspension feeding zooplankton. InHughes.R.N. (ed.). Diet Selection: An Interdisciplinary Approach to Foraging Behaviour. BlackwellScientific, Oxford, pp. 102-123.
Elbr8chter,M. (1973) Population dynamics of Ceratium in coastal waters of the Kiel Bay. Oikos, 15(Suppl.), 43-48.
Fryer.G. (1957a) The feeding mechanism of some freshwater copepods. Proc Zool Soc. London, 129,1-25.
Fryer,G. (1957b) The food of some freshwater cyclopoid copepods and its ecological significance. J.Anim.Ecol, 26,263-286.
Gliwicz,Z.M. and Lampert.W. (1994) Clutch-size variability in Daphnia: Body-size related effects ofegg predation by cyclopoid copepods. Limnol. Oceanogr., 39,485-497.
Gliwicz^.M. and Stibor,H. (1993) Egg predation by copepods in Daphnia brood cavities. Oecologia(Berlin), 95, 295-298.
Gophen,M. (1977) Food and feeding habits of Mesocydops leuckarti (Claus) in Lake Kinneret (Israel).Freshwater Bioi, 7,513-518.
Guillard,R.R. and Lorenzen.CJ. (1972) Yellow-green algae with chlorophyllide c.J. Phycol.,%, 10-14.Hansen.A.M. and Santer.B. (1995) The influence of food resources on the development, survival and
reproduction of two cyclopoid copepods: Cyclops vicinus and Mesocydops leuckarti. J. PlanktonRes., 17, 631-646.
Hargrave.B.T. andGeen.G.H. (1970) Effects of grazing on two natural phytoplankton populations. J.Fish. Res. Board Can., 27, 1395-1403.
Hessen.D.O. and Lyche.A. (1991) Inter- and intraspecific variations in zooplankton element compo-sition. Arch. Hydrobioi, 121, 343-353.
JakobsJ. (1974) Quantitative measurement of food selection. A modification of the forage ratio andIvlev's electivity index. Oecologia (Berlin), 14, 413-417.
Karabin.A. (1978) The pressure of pelagic predators of the genus Mesocydops (Copepoda, Crustacea)on small zooplankton. EkoL Pol, 26,241-257.
Krambeck,HJ., Lampert.W. and Brede.H. (1981) Messungen geringer Mengen von partikulSremKohlenstoff in natUrlichen Gewassern. GIT Fachz. Lab., 25,1009-1012.
Marshall.S.M. and Orr.A.P. (1955) On the biology of Calanus finmarchicus. 8. Food uptake, assimi-lation and excretion in adult and stage V Calanus. J. Mar. Biol. Assoc UK, 34,495-529.
Nielsen.T.G. (1991) Contribution of zooplankton grazing to the decline of a Ceratium bloom. Limnol.Occanogr.,36, 1091-1106.
Peterson.W.T.. Tiselius.P. and Ki0rboe.T. (1991) Copepod egg production, moulting and growth rates,and secondary production, in the Skagerrak in August 1988. J. Plankton Res., 13,131-154.
Pollingher.U. (1987) Ecology of dinoflagellates: B. Freshwater ecosystems. In Taylor,FJ.R. (ed.), TheBiology of Dinoflagellates. Blackwell, Oxford, pp. 502-529.
Santer.B. (1990) Lebenszyklusstrategien cyclopoider Copepoden. PhD Thesis. University of Kiel.FRG, 224 pp.
Santer,B. (1993a) Potential importance of algae in the diet of adult Cyclops vicinus. Freshwater BioL,30.269-278.
Santer.B. (1993b) Do cyclopoid copepods control Daphnia populations in early spring, thereby pro-tecting their juvenile instar stages from food limitation. Verh, Int. Ver. Limnol, 25, 634-637.
332
Dow
nloaded from https://academ
ic.oup.com/plankt/article/18/3/323/1460996 by guest on 08 January 2022

Ccratlum feeding by copepods
Santer.B. and van den Bosch,F. (1994) Herbivorous nutrition of Cyclops vicinus: the effect of a purealgal diet on feeding, development, reproduction and life cycle./. Plankton Res., 16,171-195.
Schnack.S.B. (1976) Preliminary studies on the importance of Ceratium as food source for copepods.Comm. Int. Counc Explor. Sea C. M.-ICES/L, 30.
Sommer.U., GliwiczXM., Lampert.W. and Duncan.A. (1986) The PEG-Model of seasonal successionof planktonic events in fresh waters. Arch. Hydrobiol., 106.433-471.
Van den Bosch.F. and Santer.B. (1993) Cannibalism in Cyclops abyssorum. Oikos, 67,19-28.Williamson.C.E. (1984) Laboratory and field experiments on feeding ecology of the cyclopoid cope-
pod, Mesocyclops edax. Freshwater BioL, 14, 575-585.Williamson.C.E. (1986) The swimming and feeding behaviour of Mesocyclops. Hydrobiologia, 134,
11-19.Williamson.C.E. (1991) Copepoda. In Thorp, J. (ed.). Ecology and Classification of North American
Freshwater Invertebrates. Academic Press, San Diego, pp. 787-822.
Received on March 26, 1995; accepted on September 15, 1995
333
Dow
nloaded from https://academ
ic.oup.com/plankt/article/18/3/323/1460996 by guest on 08 January 2022

Dow
nloaded from https://academ
ic.oup.com/plankt/article/18/3/323/1460996 by guest on 08 January 2022