ultrastructural features of the benthic dinoflagellate ostreopsis cf. ovata (dinophyceae)
TRANSCRIPT

This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and educational
use, including for instruction at the author’s institution and sharing with colleagues.
Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are
prohibited.
In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors
requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit:
http://www.elsevier.com/copyright

1
e
Protist, Vol. 165, 260–274, May 2014http://www.elsevier.de/protisPublished online date 13 March 2014
ORIGINAL PAPER
Ultrastructural Features of the BenthicDinoflagellate Ostreopsis cf. ovata(Dinophyceae)
Laura Escalera1, Giovanna Benvenuto, Eleonora Scalco, Adriana Zingone, andMarina Montresor
Stazione Zoologica Anton Dohrn, Villa Comunale, 80121 Napoli, Italy
Submitted June 19, 2013; Accepted March 5, 2014Monitoring Editor: Mona Hoppenrath
The toxic benthic dinoflagellate Ostreopsis cf. ovata has considerably expanded its distribution rangein the last decade, posing risks to human health. Several aspects of this species are still poorly known.We studied ultrastructural features of cultivated and natural populations of Ostreopsis cf. ovata from theGulf of Naples (Mediterranean Sea) using confocal laser scanning, and scanning and transmission elec-tron microscopy. New information on the morphology and location of several sulcal plates was gainedand a new plate designation is suggested that better fits the one applied to other Gonyaulacales. Themicrotubular component of the cytoskeleton, revealed using an anti-�-tubulin antibody, consisted of acortical layer of microtubules arranged asymmetrically in the episome and in the hyposome, comple-mented by a complex inner microtubular system running from the sulcal area towards the internal partof the cell. The conspicuous canal was delimited by two thick, burin-shaped lobes ending in a tubularventral opening. The canal was surrounded by mucocysts discharging their content into it. A similarstructure has been reported in other benthic and planktonic dinoflagellates and may be interpreted as
Author's Personal Copy
an example of convergent evolution in species producing large amounts of mucus.© 2014 Elsevier GmbH. All rights reserved.
Key words: Benthic dinoflagellate; confocal laser scanning microscopy; microtubular cytoskeleton; mucuscanal; Ostreopsis cf. ovata; ultrastructure.
Introduction
The genus Ostreopsis E.J. Schmidt includes ben-thic and epiphytic species initially found in tropicaland subtropical areas (Faust 1999; Parsons et al.2012). In the last decade, these species havebeen widely recorded also in more temperateregions (Mangialajo et al. 2011; Parsons et al.2012; Shears and Ross 2009). Several Ostreopsis
Corresponding author; fax: +39 081 7641355-mail [email protected] (L. Escalera).
species produce potent neurotoxins of the paly-toxin group (PLTXs) (Ciminiello et al. 2008, 2010;Lenoir et al. 2004; Mercado et al. 1994; Rossiet al. 2010; Ukena et al. 2002), which have beenassociated with serious human poisoning throughthe consumption of intoxicated seafood, mainly fish(Amzil et al. 2012; Taniyama et al. 2003). How-ever, the link between Ostreopsis-produced toxinsand clupeotoxism syndromes attributed to palytox-ins remains to be established (Tubaro et al. 2011).Ostreopsis blooms have also been associated withskin irritations and respiratory problems in humansthrough aerosolization of toxins (Ciminiello et al.
http://dx.doi.org/10.1016/j.protis.2014.03.0011434-4610/© 2014 Elsevier GmbH. All rights reserved.

Ostreopsis cf. ovata Ultrastructure 261
2008; Deeds and Schwartz 2010; Kermarec et al.2008; Tubaro et al. 2011). Toxins produced byOstreopsis spp. have been found in benthic marinebivalves, crustaceans and fishes, thus highlightingthe need for a careful monitoring of these ben-thic dinoflagellates in the affected areas as well asan effective management of the potential risks tohuman health (Aligizaki et al. 2011).
In the genus Ostreopsis, the species O. ovataFukuyo is apparently the most widely distributed(Parsons et al. 2012; Rhodes 2010). A consider-able genetic variability has been described for thistaxon (Penna et al. 2010; Sato et al. 2011), whichis presently considered a species-complex includ-ing several distinct cryptic species. Among them,the one deserving the name of O. ovata has notbeen established yet. Strains found in the Mediter-ranean and Atlantic regions belong to a singleclade, designated as Ostreopsis cf. ovata. The gen-eral morphology and plate pattern arrangement ofO. ovata has been illustrated in various investiga-tions (Besada et al. 1982; Faust et al. 1996; Fukuyo1981; Kang et al. 2013; Penna et al. 2005; Satoet al. 2011), where different designations were usedfor some of the thecal plates.
Living in close relationship with the substrateinvolves specific adaptations for benthic dinoflag-ellates as compared to planktonic ones, whichare reflected in both ecophysiological (Fragaet al. 2012) and morphological characteristics. Forinstance, benthic gonyaulacoid dinoflagellates aregenerally more flattened as compared to relatedspecies in the plankton. The production of highamounts of mucus is another aspect of this adap-tation. In the case of Ostreopsis, a high numberof trichocysts provide more mechanical resistanceto the mucus (Honsell et al. 2013), which at timesforms very thick layers on macroalgae and othersubstrates. The mucus may detach from the sub-strate and float, thus conferring an unpleasantaspect to seawater. Mucilage is not exclusive tobenthic species, as several planktonic dinoflagel-lates, e.g., Gonyaulax fragilis, produce it in largeamounts (Pistocchi et al. 2005). However, in thecase of those benthic species, mucus produc-tion is particularly relevant to the connection ofthe cells to the substrate, which is substantiallymodified, providing a clear example of ecosystemengineering. Several functions, such as dynamicstreamlining and defence against grazing, havebeen suggested for mucus production in planktonicspecies (Reynolds 2007), as well as in Ostreopsiscf. ovata (Barone 2007; Honsell et al. 2013).
The production of mucus has been associatedwith the presence of a conspicuous amount of
mucocysts, which are visible in ultrathin sectionsof Ostreopsis species (Besada et al. 1982; Honsellet al. 2013). In addition, a canal was illustratedwithin the cells, which was supposed to collect themucous material produced in the mucocysts andconvey it to a discharge opening localised in theventral area (Besada et al. 1982; Honsell et al.2013; Kang et al. 2013). Faust and Morton (1995)and Faust et al. (1996) associated the ventral open-ing with the ingestion of particles.
In this study, we investigated ultrastructural fea-tures of Ostreopsis cf. ovata through confocallaser scanning (CLSM) and electron microscopy(SEM and TEM) of cultured and natural popula-tions collected in the Gulf of Naples (TyrrhenianSea, Mediterranean Sea). We focused on the orga-nization of the peripheral and internal microtubularsystem, on the plate arrangement in the sulcal areaand on the ultrastructure of the mucus canal.
Results
In apical view, cells of Ostreopsis cf. ovata had anoval outline, more rounded dorsally, with a protru-sion often visible in the less pigmented ventral area(Fig. 1A). Numerous, elongated chloroplasts werepresent towards the cell periphery (Fig. 1B). Thesmall (20 �m long, 7 �m wide), elongated nucleuswas positioned obliquely from the dorsal area inthe episome to the central area in the hyposome(Fig. 1C, D).
Plate Arrangement
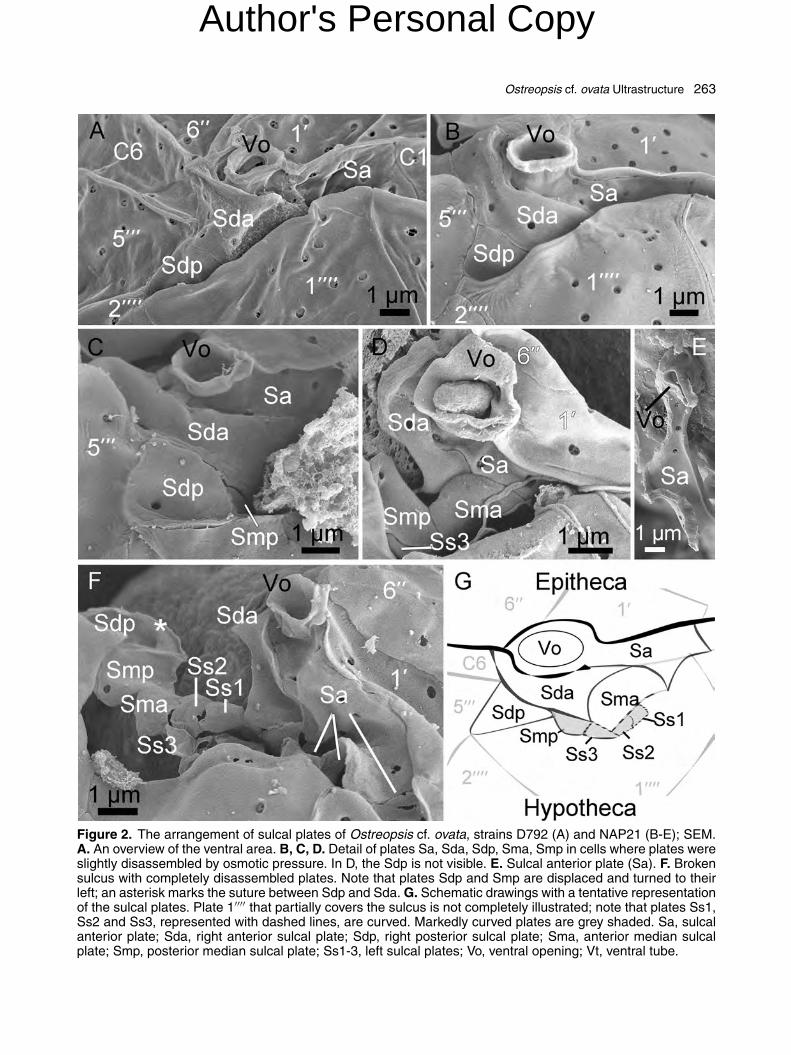
The thecal plate formula of O. cf. ovata was inter-preted as Po, 4′, 6′′, 6C, 8?S, 5′′′, 2′′′′ (Fig. 1E,F). Plates were smooth with numerous trichocystpores (ca. 250 nm diameter) which were scatteredon the epi- and hypothecal plates and lined up alongboth borders of the two cingular lists (Fig. 2A). Thecingulum was descending and displaced one timeits width (Fig. 2A, B). It consisted of six plates, ofwhich the two dorsal ones (C3 and C4) were slightlylonger (Fig. 1F). The sulcal groove was narrow andran obliquely from the left side of the ventral areainto the hypotheca. The sulcal area was partly cov-ered by the ‘wing’ constituted by plate 1′′′′ (Fig. 2A,B). In the sulcal area, five plates were clearly identi-fied: the anterior sulcal plate (Sa), the right anteriorsulcal plate (Sda), the right posterior sulcal plate(Sdp), an anterior sulcal median plate (Sma) anda posterior sulcal median plate (Smp) (Fig. 2). Incells with a broken sulcus, at least three additionalplatelets, provisionally labelled as Ss1, Ss2, Ss3,
Author's Personal Copy

262 L. Escalera et al.
Figure 1. Ostreopsis cf. ovata from field samples (A,B) and strain D792 (C, D). A. Cell with a tubular pro-trusion in the ventral area (arrow), LM (PC). B. Cellshowing the arrangement of chloroplasts, LM (EF). C,D. Cells with nuclei stained with Sybr-Green I; apical(C) and dorsal (D) views, CLSM. E, F. Line drawingsillustrating the plate pattern of epitheca (E), hypothecaand cingulum (F).
were seen in the inner left side of the sulcus (Fig. 2F,G). The Sa plate was wide, extending from theleft end of the 6′′ plate to about ¾ of the posteriorside of 1′ plate. The posterior margin of plate Sawas in contact with the anterior, sigmoid edge ofthe Sda plate (Fig. 2B-D). In the right part, the Saplate showed a conspicuous tube-like structure, theventral tube (Vt), ending with an opening (ventralopening, Vo) (Fig. 2E). The Sda plate surroundedthe posterior margin of plate Sa from its right sidetill about half of its length. The left portion of the
Sda plate was hidden by the 1′′′′ ‘wing’ (Fig. 2C). Aconcave Sdp plate was placed below the Sda plate,forming a small and closed pocket (Fig. 2C). On theleft side of the Sdp plate and posterior to the Sdaplate, a small platelet, here tentatively identifiedas median sulcal posterior (Smp), was observed(Fig. 2C). A sulcal median anterior plate (Sma) wasidentified anterior to Smp, in connection with the leftside of Sda and the posterior side of Sa (Fig. 2D).
Microtubular System
A layer of cortical microtubules lined the whole cell(Fig. 3A-C). In the episome (Fig. 3A, C; Supple-mentary Movie S1), the microtubules followed anoblique path, radiating from the apical pore plate(Po), in the left dorsal side, towards the cingulum.In the hyposome (Fig. 3B; Supplementary MovieS2), the cortical microtubules ran from the ven-tral to the dorsal part of the posterior edge of thecingulum, arranged in almost parallel lines. Tightlyadpressed longitudinal microtubules, delimited bytwo transversal microtubular bands, were presentin the cingular area (Fig. 3C). In the ventral area,a loose bundle of microtubules departed from theright side of the hypotheca forming a collar-likestructure that surrounded the sulcal area, borderedthe left side of the sulcus and continued along thedorsal surface of the cell as a long twisted strand(Fig. 3B, F, H, J; Supplementary Movie S2 and S3).In some cells, a ring of microtubules surroundingthe Vo was observed. In cells presenting a promi-nent Vt, a coiled bundle of microtubules wrappedit (Fig. 3E-J; Supplementary Movie S3). In somecells, the microtubules of the short longitudinal fla-gellum were observed emerging from the sulcus(Fig. 3K; Supplementary Movie S4).
Inside the cell, two thick adpressed strands, com-posed by several twisted microtubules, originatedfrom the sulcal area, near the emergence area ofthe flagella (Fig. 3D, J; Supplementary Movie S1and S3). One strand continued towards the cellcentre and the other reached the surface of theepisome, at times bifurcating (Fig. 3A, F, H, J; Sup-plementary Movie S1 and S3).
Ultrastructure of O. cf. ovata
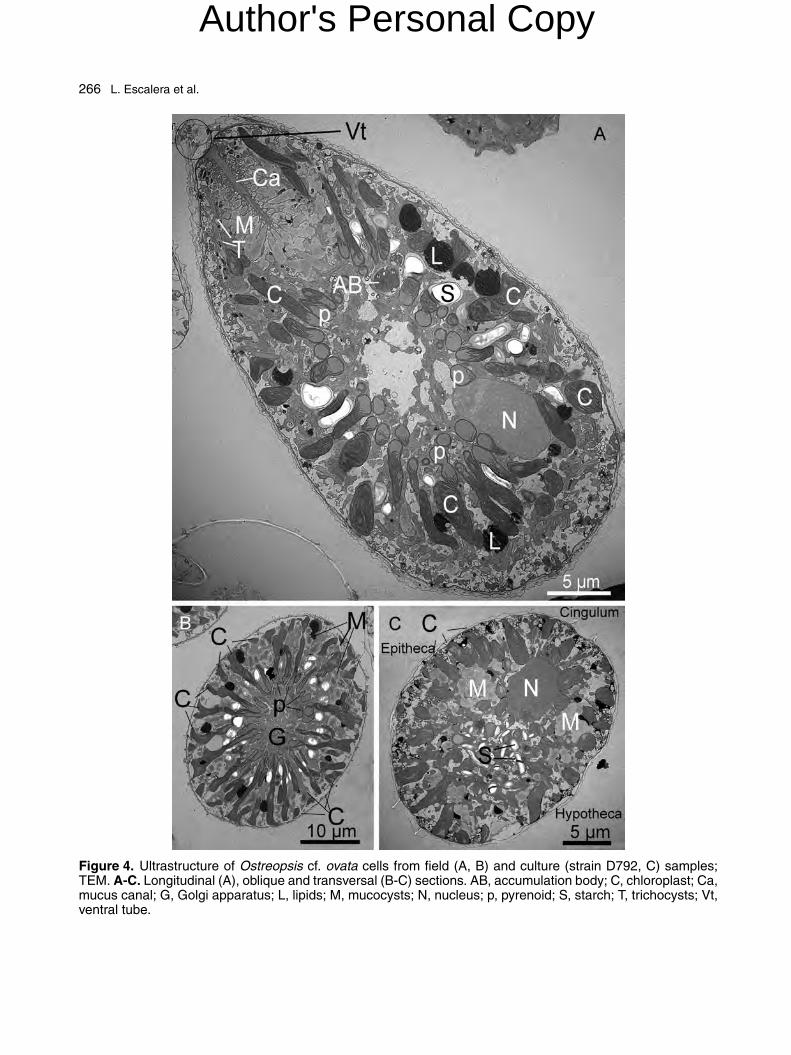
Numerous elongated chloroplasts radiating fromthe central area towards the periphery wereobserved in TEM sections of different orientation(Fig. 4A-C). The same sections also showed manymucocysts, more concentrated in the ventral por-tion of the cell, which converged towards a central,straight and elongated structure, the mucus canal.A small nucleus was located in the dorsal area.
Author's Personal Copy
Ostreopsis cf. ovata Ultrastructure 263
Figure 2. The arrangement of sulcal plates of Ostreopsis cf. ovata, strains D792 (A) and NAP21 (B-E); SEM.A. An overview of the ventral area. B, C, D. Detail of plates Sa, Sda, Sdp, Sma, Smp in cells where plates wereslightly disassembled by osmotic pressure. In D, the Sdp is not visible. E. Sulcal anterior plate (Sa). F. Brokensulcus with completely disassembled plates. Note that plates Sdp and Smp are displaced and turned to theirleft; an asterisk marks the suture between Sdp and Sda. G. Schematic drawings with a tentative representationof the sulcal plates. Plate 1′′′′ that partially covers the sulcus is not completely illustrated; note that plates Ss1,Ss2 and Ss3, represented with dashed lines, are curved. Markedly curved plates are grey shaded. Sa, sulcalanterior plate; Sda, right anterior sulcal plate; Sdp, right posterior sulcal plate; Sma, anterior median sulcalplate; Smp, posterior median sulcal plate; Ss1-3, left sulcal plates; Vo, ventral opening; Vt, ventral tube.
Author's Personal Copy

264 L. Escalera et al.
Figure 3. Cortical and internal microtubules of Ostreopsis cf. ovata cells stained with an anti-�-tubulin antibody;strain D483 (A, C), strain D792 (B, D, E-K); CLSM. A. Episome microtubules, flattened z-stack image. The Po ismarked with an arrow; the bifurcated, superficial tip of the inner microtubular strand is marked with an arrowhead.B. Hyposome microtubules with the collar-like strand running around the ventral area, flattened z-stack image.C. Cell in lateral view showing the cingular microtubules. D. Same cell as in A in a different focus, showing thetwo tightly adpressed inner strands. E-J. Serial optical sections of the cortical and internal microtubules in apicalview (the cell is slightly turned, with its left side up), BF (E, G, I), with the corresponding 488 nm Argon/2 laserexcitation images (F, H, J). Arrows indicate the sulcus, arrowheads indicate the coiled strand of microtubules.Encircled numbers correspond to the frames of a z-stack image from the surface to the inner part of the cell.
Author's Personal Copy

Ostreopsis cf. ovata Ultrastructure 265
Trichocysts, accumulation bodies, starch grains,lipid droplets, and mitochondria were scatteredthroughout the cytoplasm. The central portion ofthe cell was surrounded by the Golgi apparatus(Fig. 4B).
The small nucleus was surrounded by amembrane with numerous nuclear pores (Sup-plementary Fig. S1B, D). The chromosomespresented different levels of condensation, at timesshowing the typical condensed fibrous structure(Supplementary Fig. S1A, C). However, in sev-eral cells from both natural and culture samples,small fragments of chromatin were dispersed in alight, unstructured matrix, while a ring of condensedchromosomes delimited a darker portion at oneend of the nucleus (Fig. 4A; Supplementary Fig.S1B, D). A clear evidence of a nucleolus was notfound in many examined sections. Trichocysts wereobserved all over the cell (Fig. 4A; Supplemen-tary Fig. S2A). In the cell periphery, they presenteda square or rhomboid core, tightly wrapped bya membrane, and generally had one end closeto the amphiesma (Fig. 4A; Supplementary Fig.S2A). In the central area of the cell, in proximityto the Golgi apparatus, the trichocysts were onlyloosely wrapped by the membrane and had a gran-ular appearance, with electron-opaque bands lessmarked than in the peripheral ones (SupplementaryFig. S2B). Similar structures have been interpretedas ‘primordial trichocyst’ in Lingulodinium polye-drum (as Gonyaulax polyedra, Bouck and Sweeney1966). A few, round and relatively large (2.5-5 �mdiameter) accumulation bodies, containing darkamorphous material, were present in the centraland dorsal part of the cell (Fig. 4A). They wereoften surrounded by Golgi vesicles, mucocysts andchloroplasts apparently discharging material intotheir lumen (Supplementary Fig. S2C, D).
The mucus canal (4–6 �m long) was delimitedby two thick and elongated lobes, anterior and pos-terior, joined dorsally and along their whole length(Figs 4A, 5A–D). In longitudinal sections, the lobeshad a burin-shaped outline, thinner proximallyand swollen at the ventral end, in the proximity ofthe Vo (Fig. 5A, B), while they had a lip-shapedappearance in transversal sections (Fig. 5D, E). Inthe ventral area, a thick band of electron-opaquematerial was present at both sides of the lobes(Fig. 5A, B). Cortical microtubules were at timesdetected close to the proximal portion of thelobes (Fig. 5C). The inner portion of the lobes
was delimited by a membrane which invaginatedinto the canal (Fig. 5A, C). The mucocysts werearranged radially along the canal and showed afingerprint pattern inside (Fig. 5B, E). Most of themwere drop-shaped, with the thinner end towardsthe canal, at times opening into it (Fig. 5A, B, E).
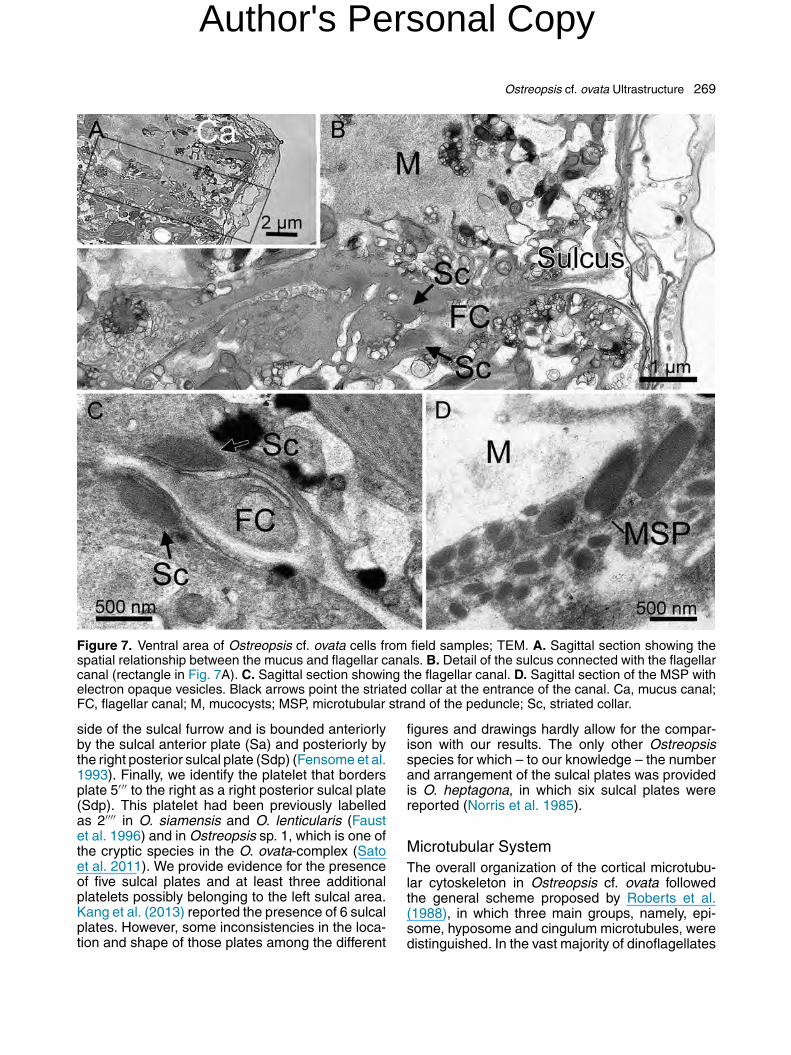
The sulcus was visible in several transversaland sagittal sections as a narrow (ca. 2 �m wide)groove (Figs 6, 7). The relative position of the sulcalgroove, mucus canal and Vo is shown in Figures6D and 7A, C. A line of microtubules ran alongthe anterior side of the sulcal groove (Fig. 6A).Several small platelets were located along theinner portion of the sulcus, across its anterior side(Fig. 6C). The short longitudinal flagellum was visi-ble in some sections (Fig. 6B). The sulcus internallycontinued with the flagellar canal (Fig. 7A-C). Twoelectron-opaque structures, here interpreted as thestriated collars of the flagella, were observed at theentrance of the canal (Fig. 7B, C). A strand of tightlyadpressed microtubules, interpreted as the micro-tubular strand of the peduncle (MSP) was detectedwithin the sulcal area, but on a different plane withrespect to the flagellar canal. The MSP was in closeconnection with elongated electron-opaque vesi-cles lying obliquely to it (Fig. 7D).
Discussion
Plate Pattern
The thecal plates of Ostreopsis ovata have changeddesignation several times over the years. The orig-inal description (Fukuyo 1981) included a partialplate formula of Po, 3′, 7′′, 5′′′, 1′′′′. Besada et al.(1982) reinterpreted the apical plate pattern andprovided a complete plate formula of Po, 4′, 6′′, 6C,8S, 5′′′, 2′′′′, which was closer to that of gonyaula-coid dinoflagellates. Faust et al. (1996) adoptedthe original interpretation of the epitheca (Fukuyo1981) but labelled the large 1′′′′ plate as a poste-rior intercalary plate. Nonetheless, their hypothecalplate formula was the same as in the originaldescription because they considered as 2′′′′ theplatelet identified as sulcal posterior (ps) by Besada(1982). The plate pattern interpretation of Faustet al. (1996) was also followed, with little variations,by other authors (Faust et al. 1996; Kang et al.2013; Penna et al. 2005; Sato et al. 2011).
➛
K. Cell in ventral lateral view showing microtubules of the sulcal area and the longitudinal flagellum (LF). The sulcus is marked withan arrow and the inner strand of microtubules with a white arrowhead. Frames shown in A and D, B, and E-J can be visualized asthree-dimensional projections in Supplementary Movies 1, 2, and 3, respectively.
Author's Personal Copy
266 L. Escalera et al.
Figure 4. Ultrastructure of Ostreopsis cf. ovata cells from field (A, B) and culture (strain D792, C) samples;TEM. A-C. Longitudinal (A), oblique and transversal (B-C) sections. AB, accumulation body; C, chloroplast; Ca,mucus canal; G, Golgi apparatus; L, lipids; M, mucocysts; N, nucleus; p, pyrenoid; S, starch; T, trichocysts; Vt,ventral tube.
Author's Personal Copy

Ostreopsis cf. ovata Ultrastructure 267
Figure 5. The mucus canal of Ostreopsis cf. ovata cells from field (A-C) and culture (strain D792, D, E) samples;TEM. A-C. (A and C) Sagittal and (B) longitudinal sections. The black rectangle, in C, marks cortical microtubulesshown in the inset. D. Transversal section, showing the lip-shaped canal lobes. E. Transversal section showingthe mucocysts containing fibrous material connected to the lumen of the canal. The electron-opaque bands inthe proximal portion of the canal lobes are marked with white circles (A, B); the lumen of the canal is markedwith an asterisk. C, chloroplast; Ca, canal; M, mucocysts; T, trichocysts; Vo, ventral opening; Vt, ventral tube.
In this paper, we adopt the interpretation of thethecal plates introduced by Besada et al. (1982)and recently applied to Gambierdiscus and Coolia(Fraga et al. 2011; Mohammad-Noor et al. 2013).As discussed by Fraga et al. (2011), this interpre-tation reflects a modification of the Kofoidean platenomenclature that takes into account the homol-ogy of the plates rather than their strict relationshipswith the other plates (Balech 1980; Fensome et al.
1993). In this view, the plate considered as 1′′ inFukuyo (1981) and Faust et al. (1996), based onthe lack of contact with the apical pore, is actuallyhomologous to plate 1′. Similarly, the large hypothe-cal plate at the antapex, at times interpreted asan intercalary (1p) plate (Faust et al. 1996; Kanget al. 2013; Penna et al. 2005; Sato et al. 2011), ishomologous to the antapical plate 2′′′′ (Besada et al.1982; Fukuyo 1981) although the contact between
Author's Personal Copy

268 L. Escalera et al.
Figure 6. Sulcal area of Ostreopsis cf. ovata cells from field samples; TEM. A. Sagittal section of the sulcus; highmagnification of the cortical microtubules in the inset. B. Sagittal section of the sulcus showing the longitudinalflagellum. C. Transversal section of the sulcus; the white circle highlights the canal. D. Sagittal section of theventral area showing the spatial relationship between mucus canal, ventral opening and sulcus. The enlargeddistal part of the canal lobes are marked with an arrow. C, chloroplasts; Ca, canal; LF, longitudinal flagellum;M, mucocysts; T, trichocyst; Vo, ventral opening.
this plate and the sulcus may not be visible (fig. 7Lin Sato et al. 2011; Fig. 1E, F, this paper).
Besada et al. (1982) identified 8 platelets in thesulcal area, which they illustrated in LM micro-graphs (their figs 6-9) that are rather difficult tointerpret. These authors also first reported the pres-ence of a ventral opening, which however wasthought to be laying in the epitheca, between plate1′ and 6′′, and hence interpreted as homologousto the ventral pore present in other gonyaulacoiddinoflagellates. Faust et al. (1996) reported thepresence of 6 sulcal platelets but neither describednor illustrated them. These latter authors alsoshowed that the ventral opening, Vo, was on a dis-tinct plate, which they named ‘ventral plate (Vp)’,
and reported another platelet, labelled as ‘ridgedplate (Rp)’, in the sulcal area posterior to ‘Vp’.In our opinion, the ‘Vp’ in Ostreopsis rather fitsthe definition of the sulcal anterior plate (Sa), fol-lowing the modified Kofoidean tabulation system(Balech 1980), widely adopted in the literature (e.g.,Fensome et al. 1993). The Sa plate is indeed‘lying close to the apex [of the sulcus] and is usu-ally aligned with the proximal end of the cingulum’(Fensome et al. 1993). Naming that plate as Saalso solves the problem that the acronym ‘Vp’ isused in other gonyaulacales to designate the ven-tral pore often found on plate 1′. We also suggestinterpreting the ‘ridged plate (Rp)’ as the right ante-rior sulcal plate (Sda), as it is placed on the right
Author's Personal Copy
Ostreopsis cf. ovata Ultrastructure 269
Figure 7. Ventral area of Ostreopsis cf. ovata cells from field samples; TEM. A. Sagittal section showing thespatial relationship between the mucus and flagellar canals. B. Detail of the sulcus connected with the flagellarcanal (rectangle in Fig. 7A). C. Sagittal section showing the flagellar canal. D. Sagittal section of the MSP withelectron opaque vesicles. Black arrows point the striated collar at the entrance of the canal. Ca, mucus canal;FC, flagellar canal; M, mucocysts; MSP, microtubular strand of the peduncle; Sc, striated collar.
side of the sulcal furrow and is bounded anteriorlyby the sulcal anterior plate (Sa) and posteriorly bythe right posterior sulcal plate (Sdp) (Fensome et al.1993). Finally, we identify the platelet that bordersplate 5′′′ to the right as a right posterior sulcal plate(Sdp). This platelet had been previously labelledas 2′′′′ in O. siamensis and O. lenticularis (Faustet al. 1996) and in Ostreopsis sp. 1, which is one ofthe cryptic species in the O. ovata-complex (Satoet al. 2011). We provide evidence for the presenceof five sulcal plates and at least three additionalplatelets possibly belonging to the left sulcal area.Kang et al. (2013) reported the presence of 6 sulcalplates. However, some inconsistencies in the loca-tion and shape of those plates among the different
figures and drawings hardly allow for the compar-ison with our results. The only other Ostreopsisspecies for which – to our knowledge – the numberand arrangement of the sulcal plates was providedis O. heptagona, in which six sulcal plates werereported (Norris et al. 1985).
Microtubular SystemThe overall organization of the cortical microtubu-lar cytoskeleton in Ostreopsis cf. ovata followedthe general scheme proposed by Roberts et al.(1988), in which three main groups, namely, epi-some, hyposome and cingulum microtubules, weredistinguished. In the vast majority of dinoflagellates
Author's Personal Copy

270 L. Escalera et al.
with a symmetrical dinokont morphology (Robertset al. 1988; Roberts and Roberts 1991; Sekidaet al. 2012), the episome microtubules have sim-ilar size and radiate from the Po plate towards thecingulum in a regular pattern, like umbrella ribs. Inthe hyposome, the microtubules depart from theposterior transversal band of the cingulum and con-verge towards the posterior end of the sulcul atthe cell antapex, mirroring those of the episome. InO. cf. ovata, the episome microtubules also departfrom the Po but they are of unequal length due tothe dorsal-left displacement of the Po. The almostparallel arrangement of the microtubules runningdorsoventrally along the hyposome is due to thevery short sulcus which does not reach the cellantapex. In the sulcal area, the cortical micro-tubules of O. cf. ovata follow an independent patternwith respect to the hyposome microtubules, asdescribed for Scrippsiella hexapraecingula (Sekidaet al. 2012). They form a collar-like structure alongthe sulcal area, which continues with a long strandalong the hyposome surface. Cortical microtubulesseem to regulate the organization and shape of the-cal plates in armoured dinoflagellates (Sekida et al.2012).
In CLSM, we could also visualize two thick andlong inner microtubular bundles departing from thesulcal area. We could not reconstruct the arrange-ment of the internal microtubular system in TEMsections. However, based on the data available forother dinoflagellates, these bundles could be inter-preted as the longitudinal microtubular root (LMR)and the transversal microtubular root (TMR) withits extension (TMRE). Microtubular roots extendfrom the basal bodies of the flagella, which mighthave different orientations in their proximal portion.Usually, basal bodies diverge from 45◦ to antiparal-lel but they are parallel in the case of species witha strong anterior/posterior compression (Robertsand Roberts 1991), like in O. cf. ovata (presentstudy). The LMR originates on the left side ofthe longitudinal basal body and runs towards thecell antapex (Calado et al. 1998; Craveiro et al.2009; Roberts and Roberts 1991). The TMR origi-nates as a single microtubule from the transversalflagellar roots and subsequently nucleates numer-ous microtubules (TMRE), which extend into thecell interior at right angles (Calado et al. 1998;Calado and Moestrup 2002; Roberts and Roberts1991). An alternative interpretation is that the innermicrotubular bundles also include the microtubu-lar strand of the peduncle (MSP). In fact, in theventral area near the flagellar apparatus of O. cf.ovata, a structure has been detected closely resem-bling the MSP described for Peridinopsis borgei
(Calado and Moestrup 2002), Amphidinium lacus-tre (Calado et al. 1998) and Esoptrodinium gemma(Calado et al. 2006). In these species, the MSP iscomposed by several microtubules that run nearlyparallel to part of the TMRE (Calado et al. 2006).A peduncle was never observed in culture or fieldsamples of O. cf. ovata, nor was it detected in anyTEM sections. However, a peduncle-homologousmicrotubular strand was reported in Palatinus apic-ulatus (Craveiro et al. 2009), which also lacks apeduncle.
Cell Ultrastructure
The main ultrastructural features of Ostreopsiscf. ovata do not present important differencescompared with other dinoflagellates, with someexceptions. In addition to the internal mucocystswith a fingerprint-like content, Honsell et al. (2013)reported flask-shaped peripheral mucocysts. Thelatter are quite common in dinoflagellates (Lee1999) but were not observed in our materialfrom field or culture samples. The accumulationbodies found in O. cf. ovata, also called PAS bod-ies due the positive reaction with the periodicacid-Schiff reagent, have been reported for manydinoflagellates, e.g. Prorocentrum lima, P. macu-losum (Zhou and Fritz 1994) and Lingulodiniumpolyedrum (Schmitter and Jurkiewicz 1981). Thesestructures have been related to autophagy or stor-age of waste material (e.g., Taylor 1968; Schmitter1971) and considered analogous to animal lyso-somes (Dodge and Greuet 1987).
The nucleus of Ostreopsis cf. ovata is insteadpeculiar. In addition to the small size, elongatedshape and oblique position in the cell, severalaspects contrast with the typical dinokaryon (Lee1999), including the absence a clear nucleolus, thering of condensed chromosomes at one end of thenucleus and the fragments of chromatin dispersedin an unstructured matrix. The degree of chromatincondensation has been related with the nucleardivision (Lee 1999). However, in cells of O. cf. ovataundergoing mitotic division the chromatin organiza-tion was similar to that of non-dividing cells.
The most peculiar feature in the microanatomyof Ostreopsis cf. ovata is the presence of thecanal delimited by two thick lobes, surroundedby mucocysts that discharge their content into itand ending with a ventral tubular opening. Thepresence of this structure in Ostreopsis was dis-covered by Besada et al. (1982), who interpretedit as a pusule and hypothesized that the vesiclescontained mucous material extruded into the canaland externally through the ventral pore. Mucocysts
Author's Personal Copy

Ostreopsis cf. ovata Ultrastructure 271
discharging into a canal were observed in otherbenthic gonyaulacoid dinoflagellates that producehigh amounts of mucus, such as Gambierdiscustoxicus and Coolia monotis (Besada et al. 1982),which however apparently lack a ventral opening.A mucus canal was also observed in some benthicProrocentrum species, such as P. lima, P. mac-ulosum (Zhou and Fritz 1993) and P. concavum(Mohammad-Noor et al. 2007), as well as in theplanktonic P. micans (Dodge and Bibby 1973). Inall these species, an accessory pore, also calledauxiliary pore or apical pore, is located close tothe large pore from which the flagella emerge. Theconnection between this pore and a vesicular struc-ture identified as a pusule was clearly describedfor P. concavum, where a canal containing fibrousmaterial expands into two small chambers sur-rounded by mucocysts (Mohammad-Noor et al.2007). The upper chamber has a thick collar at thebase, which was interpreted as a sphincter thatregulates excretion. A similar structure, consistingof two thick and electron-opaque lobes at the apicalend of the canal, was found in P. maculosum and inP. lima (see figs 9 and 11 in Zhou and Fritz 1993).
In O. cf. ovata, Honsell et al. (2013) describedmucocysts converging into the canal as well as in alarge chamber. However that chamber, illustrated intheir figure 4D, rather resembles an accumulationbody, which in our images collects the dischargeof mucocysts and chloroplasts, probably in rela-tion with autolytic processes. These organelles,also visible in LM as bright orange granules, areindeed often located in the central and dorsal partof the cell, quite far from the mucus canal. Kanget al. (2013) described the presence of a singleelongated canal connected with the Vo, which cor-responds to the mucus canal that we describe indetail. However, in their figure 7F, they misinter-preted the sulcus as the elongated canal. Thereforewe propose that only one structure, the canal, col-lects the material produced in the mucocysts.
Despite the above-mentioned differences, thestructural and functional homology between thepusule-like organelle of Prorocentrum and themucus canal of Ostreopsis is quite evident, andmay be interpreted as an example of convergentevolution.
The ventral opening of Ostreopsis species wassuggested to be involved in the ingestion of preys(Faust and Morton 1995; Faust et al. 1996). Thishypothesis was further supported by the presenceof large vacuoles, supposed to contain ciliate prey,seen in light microscopy within the cells (Faust andMorton 1995). Mixotrophy cannot be excluded inOstreopsis cf. ovata. Cells of this species were
shown to aggregate around small invertebrates,suggesting they were able to feed on them (Barone2007). However, a full evidence of mixotrophy,through the detection of food vacuoles in TEM sec-tions or feeding experiments, is still lacking. We didnotice coloured vacuoles in some O. cf. ovata cellsfrom field samples (data not shown), but no suchvacuoles were detected in numerous thin sectionson natural material. The detection of a MSP, whichsuggests the presence of a peduncle, may indi-cate mixotrophy, but requires further investigations.Our study confirms that the ventral opening ratherplays a role in mucus excretion, which is related tothe species adaptation to the benthic environment.Indeed mucus production and excretion apparentlyinvolved relevant changes in the external and inter-nal morphology of the species. The high amountof mucocysts within the cell and the macroscopicmucilaginous net in the field suggest a great ener-getic investment, which points at the pivotal role ofmucus production in the colonisation of the benthichabitat.
Methods
Sampling and culture methods: Ostreopsis cf. ovata was iso-lated from macroalgal samples collected at the Gaiola station(Gulf of Naples, Tyrrhenian Sea, Mediterranean Sea, 40◦ 47′31′′ N 14◦ 11′ 17′′ E) in September 2008 (strain D483), in July2011 (strain D792) and in 2012 (strain NAP21). Specimens ofthe red alga Asparagopsis taxiformis were collected between3 and 7 m depth by scuba diving, put in plastic bottles filledwith seawater and kept in cool conditions until arrival in thelaboratory. Single Ostreopsis cells were isolated with a glassmicropipette under a light microscope and placed in culture-plate wells filled with K/2 medium (Keller et al. 1987). Cultureswere kept at 22 ◦C, under a photoperiod of 12L:12D h anda photon fluence rate of 100 �mol photons m-2 s-1. The ITSsequences of the strain D792 used in part of this study indi-cate that it belongs to the Mediterranean O. cf. ovata clade asdefined in Sato et al. (2011) (A. Z., unpublished data). A naturalsample with a very high concentration of Ostreopsis cf. ovata,collected in Cala San Basilio (Gulf of Naples, Tyrrhenian Sea,Italy: 40◦ 47′ 34′′ N 14◦ 11′ 18′′ E) on July 6th 2012, was used forCLSM and TEM observations. Strains D792 and NAP21 havebeen deposited in the Culture Collection of Algae at the Univer-sity of Cologne (CCAC; http://www.ccac.uni-koeln.de/) with thestrain numbers CCAC 3892 B and CCAC 3893 B, respectively.
Light and epifluorescence microscopy: Live and fixedsamples were examined with an inverted microscope ZeissAxiovert 200 (Carl Zeiss AG, Oberkochen, Germany) equippedwith bright field (BF), phase contrast (PC) and epifluorescence(EF) illumination and with a Zeiss AxioCam digital camera.Chloroplasts were observed with a DAPI filter set (excitationat 365/12 nm and emission at LP 397 nm).
Confocal laser scanning microscopy (CLSM): Formalde-hyde fixed (1.6%) cells of strain D792 were stained with 1%Sybr-Green I (Molecular Probes, Oregon, USA) and observedwith a ZEISS LSM 510META (Carl Zeiss, Oberkochen, Ger-many) (excitation with a 488 nm Argon laser and emission filter
Author's Personal Copy

272 L. Escalera et al.
BP 500-550 nm). Cortical and subcortical microtubules of O. cf.ovata strains D483 and D792 were visualized with an anti-�-tubulin antibody. To remove the mucous material, the samplewas first treated with 0.1 M Na-EDTA (Scalco et al. 2012) andplaced in an ultrasonic bath for 5 min. After two washes withPBS 1x, each followed by centrifugation, cells were fixed for20 min in 80% MeOH, 50 mM EGTA (pH 7) and 10% dimethylsulfoxide (DMSO) (von Dassow 2003). Cells were washedtwice in PBS-T (PBS 1x, 1% Triton X-100). The pellet wasfirst incubated for 30 min in 5% Goat serum diluted in PBS-T, centrifuged, and subsequently incubated overnight with amouse monoclonal anti-�-tubulin antibody (Sigma-Aldrich, St.Louis, Missouri, US) (1:1000) in 5% Goat serum and PBS-T at4 ◦C on a rotating wheel agitator. After three washes in 1 mL ofPBS-T, the pellet was incubated in a 1:400 dilution of CF488Aconjugated goat anti-mouse IgG (Biotium, SIC Società ItalianaChimici, Rome, IT) in 5% Goat serum/PBS-T on a rotating wheelagitator at room temperature in the dark for 2 h. The pellet wasthen washed three times with 1 mL of PBS-T, twice with 1 mLof PBS, 5 min each, and finally resuspended in 100-200 �L ofPBS. All the centrifugation steps were performed with a handcentrifuge (Hettich, DJB Labcare Ltd, Newport Pagnell UK).Stained microtubules were excited at 488 nm with an Argon/2laser and observed with a LP 505 nm emission wavelength filter.
Scanning electron microscopy (SEM): Cells (strain D792)were treated with 10% Triton X-100 in PBS and placed in anultrasonic bath for 5 min to remove the mucus material from theirsurface (Litaker et al. 2009). The sample was fixed overnightwith 5% glutaraldehyde at 4 ◦C and subsequently placed ona membrane filter of 5 �m-pore size in a Swinnex filter holder(Millipore, Billerica, Massachusetts, USA). Cells were washedwith 20 mL of fresh PBS 1x for approximately 40 min and dehy-drated with a serial acetone concentration. The sample wascritical point dried, gold sputter coated (Polaron SC7640, Quo-rum Technologies Ltd., Ashford, UK) and observed with a JEOLJSM 6700F (JEOL Ltd, Tokyo, Japan). To visualize the numberand relative position of the sulcal plates, cells of strain NAP21were swollen by treating them with 10% Triton X-100 in tapwater for 45 min before fixation (1 h in 2% glutaraldehyde atroom temperature) and acetone dehydration.
Transmission electron microscopy (TEM): Two protocolswere applied to study the ultrastructure of Ostreopsis cf. ovata.A standard protocol (protocol 1) was used for culture samples.Strain D792 was fixed with 2.5% glutaraldehyde at room tem-perature for 2 h and post-fixed with 1% osmium tetroxide at4 ◦C for 1 h. Cells were dehydrated in an ethanol series, fur-ther substituted by propylene oxide and embedded in Epon812 (TAAB, TAAB Laboratories Equipment Ltd, Berkshire, UK)at room temperature for 1 d and polymerized at 60 ◦C for 2d. Resin blocks were sectioned with a MT X ultramicrotome(RMC products, Boeckeler, Tucson, AZ, USA). Sections werecontrasted with 4% aqueous uranyl acetate for 30 min, rinsedonce with a mix (1:1) of methanol and bidistilled water, twice withbidistilled water, placed on formvar-coated grids and observedwith a Zeiss LEO 912AB TEM (Zeiss, Oberkochen, Germany).For the observation of the natural sample collected during theOstreopsis bloom, a modification of the protocol described inEikrem and Moestrup (1998) was followed (protocol 2). A vol-ume of 15 mL of natural sample of O. cf. ovata was fixed for30 min with 4% glutaraldehyde at room temperature. The pel-let was then rinsed four times with 0.1 M cacodylate buffer (pH7.5), centrifuging with a hand centrifuge for 1-2 min. After thelast wash, 1 mL of a mix of 0.5% ferricyanide and 0.5% osmiumtetroxide in 0.1 M cacodylate buffer (pH 7.5) was added to thepellet to increase the contrast. The sample was kept overnightat 4 ◦C, rinsed five times with 0.1 M cacodylate buffer (pH 7.5)
and centrifuged as mentioned above. The dehydration, poly-merization, sectioning and staining steps were the same as inprotocol 1.
Acknowledgements
We thank G. Zazo and M. Cannavacciuolo (ServizioPesca, SZN) for sampling, A. Italiano and D.Di Cioccio for culture provision and F. Iamunno(Electron Microscopy Service, SZN) for techni-cal support. L. Escalera’s postdoc fellowship wasfunded by the Spanish Ministry of Education(MEC) within ‘Programa Nacional de Movilidad deRecursos Humanos del Plan Nacional de I+D+i2008-2011’. This investigation was supported bySZN and by the Ostreopsis project funded by theHealth Office of the Regione Campania. We thankthree anonymous reviewers for their constructivecomments.
Appendix A. Supplementary Data
Supplementary data associated with this arti-cle can be found, in the online version, athttp://dx.doi.org/10.1016/j.protis.2014.03.001.
References
Aligizaki K, Katikou P, Milandri A, Diogène J (2011) Occur-rence of palytoxin-group toxins in seafood and future strategiesto complement the present state of the art. Toxicon 57:390–399
Amzil Z, Sibat M, Chomerat N, Grossel H, Marco-MirallesF, Lemee R, Nezan E, Sechet V (2012) Ovatoxin-a and paly-toxin accumulation in seafood in relation to Ostreopsis cf.ovata blooms on the French Mediterranean coast. Mar Drugs10:477–496
Balech E (1980) On thecal morphology of dinoflagellates withspecial emphasis on cingular and sulcal plates. An CentroCienc del Mar y Limnol Univ Nal Auton México 7:57–68
Barone R (2007) Behavioural trait of Ostreopsis ovata (Dinoph-iceae) in Mediterranean rock pools: the spider’s strategy.Harmful Algae News 33:1–3
Besada EG, Loeblich LA, Loeblich AR III (1982)Observations on tropical, benthic dinoflagellates fromciguatera-endemic areas: Coolia, Gambierdiscus, andOstreopsis. Bull Mar Sci 32:723–735
Bouck GB, Sweeney BM (1966) The fine structure andontogeny of trichocysts in marine dinoflagellates. Protoplasma61:205–223
Calado AJ, Moestrup Ø (2002) Ultrastructural study of thetype species of Peridiniopsis borgei (Dinophyceae), with specialreference to the peduncle and flagellar apparatus. Phycologia41:567–584
Author's Personal Copy

Ostreopsis cf. ovata Ultrastructure 273
Calado AJ, Craveiro SC, Moestrup Ø (1998) Taxonomyand ultrastructure of a freshwater, heterotrophic Amphidinium(Dinophyceae) that feeds on unicellular protist. J Phycol34:536–554
Calado AJ, Craveiro SC, Daugbjerg N, Moestrup Ø (2006)Ultrastructure and LSU RDNA-based phylogeny of Esoptro-dinium gemma (Dinophyceae), with notes on feeding behaviorand the description of the flagellar base area of a planozygote.J Phycol 42:434–452
Ciminiello P, Dell’Aversano C, Fattorusso E, Forino M,Grauso L, Tartaglione L (2010) A 4-decade-long (and stillongoing) hunt for palytoxins chemical architecture. Toxicon57:362–367
Ciminiello P, Dell’Aversano E, Fattorusso E, Fiorino M,Tartaglione L, Grillo C, Melchiorre N (2008) Putative palytoxinand its new analogue, ovatoxin-a, in Ostreopsis ovata collectedalong the Ligurian coasts during the 2006 toxic outbreak. J AmSoc Mass Spetr 19:111–120
Craveiro SC, Calado AJ, Daugbjerg N, Moestrup Ø (2009)Ultrastructure and LSU rDNA-based revision of Peridiniumgroup palatinum (Dinophyceae) with the description of Palatinusgen. nov. J Phycol 45:1175–1194
Deeds JR, Schwartz MD (2010) Human risk associated withpalytoxin exposure. Toxicon 56:150–162
Dodge JD, Bibby BT (1973) The Prorocentrales (Dino-phyceae). I. A comparative account of fine structure inthe genera Prorocentrum and Exuviaella. Bot J Linn Soc67:175–187
Dodge JD, Greuet C (1987) Dinoflagellate Ultrastructure andComplex Organelles. In Taylor FJR (ed) The Biology of Dinoflag-ellates. Blackwell Sci Publ, London, pp 92–119
Eikrem W, Moestrup Ø (1998) Structural analysis of the fla-gellar apparatus and the scaly periplast in Chrysochromulinascutellum sp. nov. (Prymnesiophyceae, Haptophyta) from theSkagerrak and the Baltic. Phycologia 37:132–153
Faust MA (1999) Three new Ostreopsis species (Dino-phyceae): O. marinus sp. nov., O. belizeanus sp. nov., and O.caribbeanus sp. nov. Phycologia. 38:92–99
Faust MA, Morton S (1995) Morphology and ecology ofthe marine dinoflagellate Ostreopsis labens sp. nov. (Dino-phyceae). J Phycol 31:456–463
Faust MA, Morton SL, Quod JP (1996) Further study of marinedinoflagellates: the genus Ostreopsis (Dinophyceae). J Phycol32:1053–1065
Fensome RA, Taylor FJR, Norris G, Sarjeant WAS, Whar-ton DJ, Williams GL (1993) A Classification of Living andFossil Dinoflagellates. Micropaleontology, Special PublicationNumber 7. Sheridan Press, Hanover
Fraga S, Rodriguez F, Bravo I, Zapata M, Maranon E(2012) Review of the main ecological features affecting benthicdinoflagellate blooms. Cryptog Algol 33:171–179
Fraga S, Rodríguez F, Caillaud A, Diogène J, Raho N, ZapataM (2011) Gambierdiscus excentricus sp. nov. (Dinophyceae), abenthic toxic dinoflagellate from the Canary Islands (NE AtlanticOcean). Harmful Algae 11:10–20
Fukuyo Y (1981) Taxonomical study on benthic dinoflagellatescollected in coral reefs. B Jpn Soc Sci Fish 47:967–978
Honsell G, Bonifacio A, De Bortoli M, Penna A, Battoc-chi C, Ciminiello P, Dell’Aversano C, Fattorusso E, SosaS, Yasumoto T, Tubaro A (2013) New insights on cytologicaland metabolic features of Ostreopsis cf. ovata Fukuyo (Dino-phyceae): a multidisciplinary approach. PLoS ONE 8:e57291
Kang NS, Jeong HJ, Lee SY, Lim AS, Lee MJ, Kim HS, YihW (2013) Morphology and molecular characterization of theepiphytic benthic dinoflagellate Ostreopsis cf. ovata in the tem-perate waters off Jeju Island, Korea. Harmful Algae 27:98–112
Keller MD, Selvin RC, Claus W, Guillard RRL (1987) Media forthe culture of oceanic ultraphytoplankton. J Phycol 23:633–638
Kermarec F, Dor F, Armengaud A, Charlet F, Kantin R,Sauzade D, De Haro L (2008) Health risks related to Ostreopsisovata in recreational waters. Envir Risque Sante 7:357–363
Lee RE (1999) Phycology. 3rd ed, Cambridge University Press,Cambridge
Lenoir S, Ten-Hage L, Turquet J, Quod JP, Bernard C, Hen-nion MC (2004) First evidence of palytoxin analogues from anOstreopsis mascarenensis (Dinophyceae) benthic bloom in thesouthwestern Indian Ocean. J Phycol 40:1042–1051
Litaker RW, Vandersea MW, Faust MA, Kibler SR, ChinainM, Holmes MJ, Holland WC, Tester PA (2009) Taxonomyof Gambierdiscus including four new species, Gambierdis-cus caribeanus, Gambierdiscus carolinianus, Gambierdiscuscarpenteri and Gambierdiscus ruetzleri (Gonyaulacales, Dino-phyceae). Phycologia 48:344–390
Mangialajo L, Ganzin N, Accoroni S, Asnaghi V, BlanfunéA, Cabrini M, Cattaneo-Vietti R, Chavanon F, Chiantore M,Cohu S, Costa E, Fornasaro D, Grossel H, Marco-Miralles F,Masó M, René A, Rossi AM, Sala MM, Thibaut T, Totti C, VilaM, Lemée R (2011) Trends in Ostreopsis proliferation along theNorthern Mediterranean coasts. Toxicon 57:408–420
Mercado JA, Viera M, Escalona de Motta G, TostesonTR, Gonzàlez I, Silva W (1994) An extraction proceduremodification changes the toxicity, chromatographic profile andpharmacologic action of Ostreopsis lenticularis extracts. Toxi-con 32:256
Mohammad-Noor N, Moestrup Ø, Daugbjerg N (2007) Light,electron microscopy and DNA sequences of the dinoflagel-late Prorocentrum concavum (syn. P. arabianum) with specialemphasis on the periflagellar area. Phycologia 46:549–564
Mohammad-Noor N, Moestrup Ø, Lundholm N, FragaS, Adam A, Holmes MJ, Saleh E (2013) Autecology andphylogeny of Coolia tropicalis and Coolia malayensis (Dino-phyceae), with emphasis on taxonomy of C. tropicalis based onlight microscopy, scanning electron microscopy and LSU rDNA.J Phycol 49:536–545
Norris DR, Bomber JW, Balech E (1985) Benthic Dinoflagel-lates Associated with Ciguatera from Florida Keys. I. Ostreopsisheptagona sp. nov. In Anderson DM, White AW, Baden DG (eds)Toxic Dinoflagellates. Elsevier, Amsterdam, pp 39–44
Parsons ML, Aligizaki K, Bottein M-YD, Fraga S, Mor-ton SL, Penna A, Rhodes L (2012) Gambierdiscus andOstreopsis: Reassessment of the state of knowledge of theirtaxonomy, geography, ecophysiology, and toxicology. HarmfulAlgae 14:107–129
Penna A, Fraga S, Battocchi C, Casabianca C, GiacobbeMG, Riobò P, Vernesi C (2010) A phylogeographical study of
Author's Personal Copy

274 L. Escalera et al.
the toxic benthic dinoflagellate genus Ostreopsis Schmidt. JBiogeogr 37:830–841
Penna A, Vila M, Fraga S, Giacobbe MG, Andreoni F, RiobóP, Vernesi C (2005) Characterization of Ostreopsis and Coo-lia (Dinophyceae) isolates in the Western Mediterranean Seabased on morphology, toxicity and internal transcribed spacer5. 8S rDNA sequences. J Phycol 41:212–225
Pistocchi R, Cangini M, Totti C, Urbani R, Guerrini F,Romagnoli T, Sist P, Palamidesi S, Boni L, Pompei M(2005) Relevance of the dinoflagellate Gonyaulax fragilis inmucilage formations of the Adriatic Sea. Sci Total Environ 353:307–316
Reynolds C (2007) Variability in the provision and function ofmucilage in phytoplankton: facultative responses to the envi-ronment. Hydrobiologia 578:37–45
Rhodes L (2010) World-wide occurrence of the toxic dinoflag-ellate genus Ostreopsis Schmidt. Toxicon 57:400–407
Roberts KR, Roberts JE (1991) The flagellar apparatus andcytoskeleton of the dinoflagellates: a comparative overview.Protoplasma 164:105–122
Roberts KR, Farmer MA, Schneider RM, Lemoine JE(1988) The microtubular cytoskeleton of Amphidinium rhyncho-cephalum (Dinophyceae). J Phycol 24:544–553
Rossi R, Castellano V, Scalco E, Serpe L, ZingoneA, Soprano V (2010) New palytoxin-like molecules inMediterranean Ostreopsis cf. ovata (dinoflagellates) and inPalythoa tuberculosa detected by liquid chromatography-electrospray ionization time-of-flight mass spectrometry. Toxi-con 56:1381–1387
Sato S, Nishimura T, Uehara K, Sakanari H, Tawong W, Hari-ganeya N, Smith K, Rhodes L, Yasumoto T, Taira Y, Suda S,Yamaguchi H, Adachi M (2011) Phylogeography of Ostreop-sis along West Pacific coast, with special reference to a novelclade from Japan. PLoS ONE 6:e27983
Scalco E, Brunet C, Marino F, Rossi R, Soprano V, Zin-gone A, Montresor M (2012) Growth and toxicity responsesof Mediterranean Ostreopsis cf. ovata to seasonal irradianceand temperature conditions. Harmful Algae 17:25–34
Schmitter RE (1971) The fine structure of Gonyaulax polyedra,a bioluminescent marine dinoflagellate. J Cell Sci 9:147–173
Schmitter RE, Jurkiewicz AJ (1981) Acid phosphatase local-ization in PAS-bodies of Gonyaulax. J Cell Sci 51:15–23
Sekida S, Takahira M, Horiguchi T, Okuda K (2012)Effects of high pressure in the armored dinoflagellate Scrip-siella hexapraencingula (Peridiniales, Dinophyceae): changesin thecal plate pattern and microtubule assembly. J Phycol48:163–173
Shears NT, Ross PM (2009) Blooms of benthic dinoflagel-lates of the genus Ostreopsis; an increasing and ecologicallyimportant phenomenon on temperate reefs in New Zealand andworldwide. Harmful Algae 8:916–925
Taniyama S, Arakawa O, Terada M, Nishio S, Takatani T,Mahmud Y, Noguchi T (2003) Ostreopsis sp. , a possible ori-gin of palytoxin (PTX) in parrotfish Scarus ovifrons. Toxicon42:29–33
Taylor DL (1968) In situ studies on the cytochemistry and ultra-structure of a symbiotic marine dinoflagellate. J Mar Biol AssocUK 48:349–366
Tubaro A, Durando P, Del Favero G, Ansaldi F, Icardi G,Deeds JR, Sosa S (2011) Case definitions for human poison-ings postulated to palytoxins exposure. Toxicon 57:478–495
Ukena T, Satake M, Usami M, Oshima Y, Fujita T, Naoki H,Yasumoto T (2002) Structural confirmation of ostreocin-D byapplication of negative-ion fast-atom bombardment collision-induced dissociation tandem mass spectrometric methods.Rapid Commun Mass Spectrom 16:2387–2393
von Dassow P (2003) Regulation of Bioluminescence in theDinoflagellate Lingulodinium polyedrum. PhD Thesis, Univer-sity of California
Zhou J, Fritz L (1993) Ultrastructure of two toxic marinedinoflagellates, Prorocentrum lima and Prorocentrum maculo-sum. Phycologia 32:444–450
Zhou J, Fritz L (1994) The PAS accumulation bodies in Pro-rocentrum lima and Prorocentrum maculosum (Dinophyceae)are dinoflagellate lysosomes. J Phycol 30:39–44
Available online at www.sciencedirect.com
ScienceDirect
Author's Personal Copy