the theoretical and empirical diversities of the concept of biodiversity
TRANSCRIPT
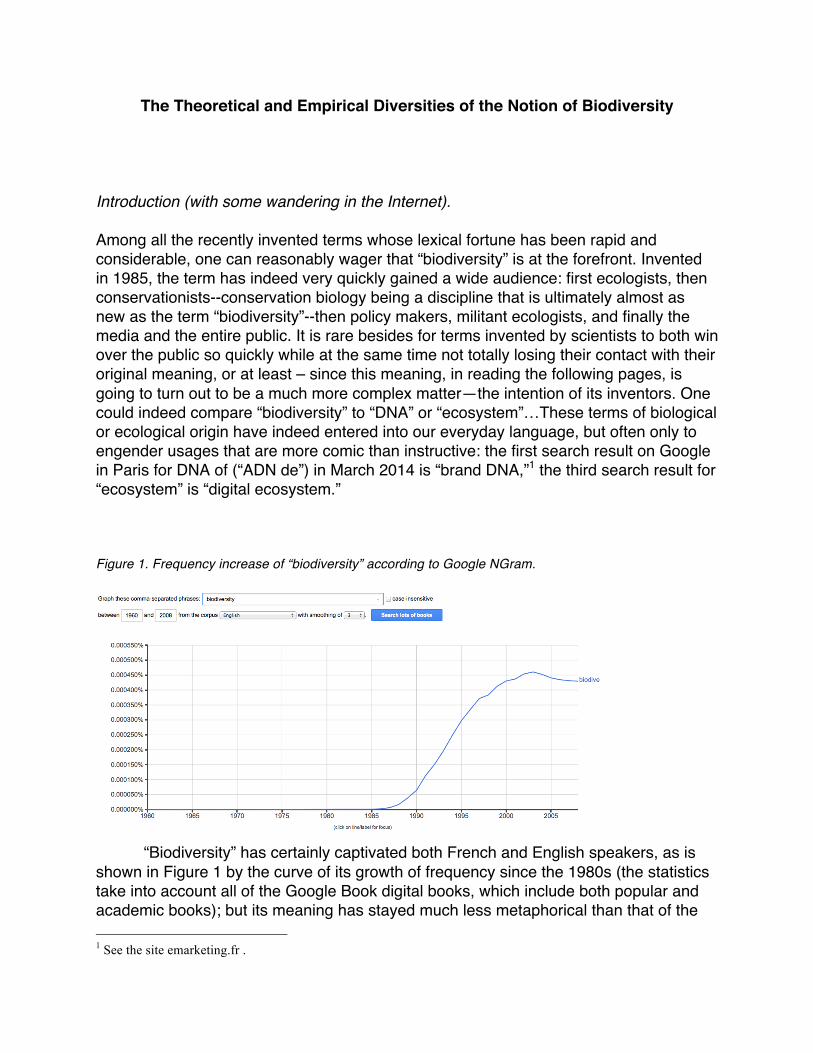
The Theoretical and Empirical Diversities of the Notion of Biodiversity Introduction (with some wandering in the Internet). Among all the recently invented terms whose lexical fortune has been rapid and considerable, one can reasonably wager that “biodiversity” is at the forefront. Invented in 1985, the term has indeed very quickly gained a wide audience: first ecologists, then conservationists--conservation biology being a discipline that is ultimately almost as new as the term “biodiversity”--then policy makers, militant ecologists, and finally the media and the entire public. It is rare besides for terms invented by scientists to both win over the public so quickly while at the same time not totally losing their contact with their original meaning, or at least – since this meaning, in reading the following pages, is going to turn out to be a much more complex matter—the intention of its inventors. One could indeed compare “biodiversity” to “DNA” or “ecosystem”…These terms of biological or ecological origin have indeed entered into our everyday language, but often only to engender usages that are more comic than instructive: the first search result on Google in Paris for DNA of (“ADN de”) in March 2014 is “brand DNA,”1 the third search result for “ecosystem” is “digital ecosystem.” Figure 1. Frequency increase of “biodiversity” according to Google NGram.
“Biodiversity” has certainly captivated both French and English speakers, as is shown in Figure 1 by the curve of its growth of frequency since the 1980s (the statistics take into account all of the Google Book digital books, which include both popular and academic books); but its meaning has stayed much less metaphorical than that of the 1 See the site emarketing.fr .
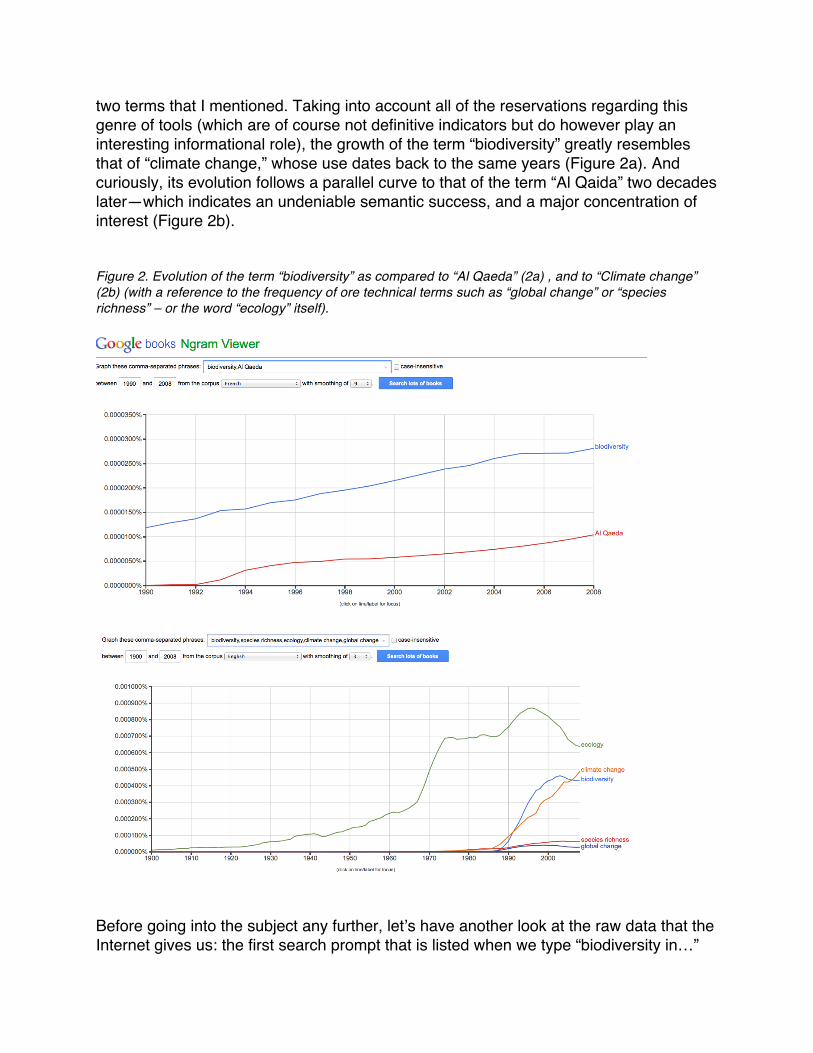
two terms that I mentioned. Taking into account all of the reservations regarding this genre of tools (which are of course not definitive indicators but do however play an interesting informational role), the growth of the term “biodiversity” greatly resembles that of “climate change,” whose use dates back to the same years (Figure 2a). And curiously, its evolution follows a parallel curve to that of the term “Al Qaida” two decades later—which indicates an undeniable semantic success, and a major concentration of interest (Figure 2b). Figure 2. Evolution of the term “biodiversity” as compared to “Al Qaeda” (2a) , and to “Climate change” (2b) (with a reference to the frequency of ore technical terms such as “global change” or “species richness” – or the word “ecology” itself).
Before going into the subject any further, let’s have another look at the raw data that the Internet gives us: the first search prompt that is listed when we type “biodiversity in…”

(thus the search prompt that is most popular that day, at least in Paris) is “biodiversity in danger”; it lists around 100,000 websites. In English, “biodiversity is…” brings up as its first result “biodiversity is important” (Fig.3); as for “biodiversity loss,” there are around seven million pages that are available, more than half of the 13 million results for “biodiversity.” Elsewhere, still staying in February 2014, the first search prompt we see when we type “biodiversity loss” is “biodiversity loss and its impact on humanity” (most of whose results send us back to the article of the same name in Nature (Cardinale et al 2012) ). Granted that this is very loose and empirical, two impressions stand out when we make this first overview: “biodiversity” is an extremely widespread term; and its usage is rarely metaphorical and involves significant negative connotations; in particular, it acts as a danger sign for the human species. “Biodiversity” and “climate change” are thus spread in a parallel manner, in scientific discussions as well as in public ones; we also have the impression that “biodiversity” leads us to contexts that are not emotionally neutral, which is also a clear property of the term “climate change”; in this sense, the two seem to share a burden that is in a manner of speaking axiological and deeply emotional. Figure 3. Ranking of searches for “biodiversity is”.
Having made these observations, I am first going to develop a sort of cartography of the different meanings and usages of “biodiversity,” which will emphasize a few leitmotivs. Next, to introduce some of these leitmotifs, I will highlight two or three important elements in the process through which the term came to form a decisive role both for scientists from different fields linked to ecology, and the politicians or lawyers involved with the policies that govern the consequences of man’s actions on nature. 1. The Meanings and the Usages of Biodiversity: A Preliminary Overview Why do we say biodiversity rather than biological diversity? After all, when William Rosen, the creator of the National Forum on Biological Diversity of the American National Research Council first used this word, Edward O. Wilson then used this term “Biodiversity” as the name of the forum that he coordinated (Wilson 1988); and destined

for the success which we have seen, it seems that it came about simply to replace “biological diversity”--which was originally a part of the title of the conference. Otherwise said, is there a real scientific necessity for “biodiversity?” Does the term “biodiversity” indicate properties, phenomena, or even a research program or a type of approach that is not indicated or taken into account when we simply talk about “diversity” in biology? Or rather, does the invention of “biodiversity” and the subsequent spreading of this term respond to extrascientific approaches, whether political, ethical, social, or others? After all, the first sentence of the princeps document about biodiversity--the Convention of Biological Diversity, signed in 1992--mentions the “intrinsic value of biological diversity,” without supplementary justification: clearly, from its beginnings the term had ethical sway. It is very plausible that there is no appropriate definitive answer to this question; and that the term “biodiversity” is simply overdetermined, carrying independent semantic weights which are sometimes in tension with each other. In essence, the term could reflect the flagrant ambiguity (at least in French or English) of the term “ecology:” scientific ecology on the one hand (for which we use the term “ecologist,” or in French we recently invented écologue,) which includes its disciplines, researchers, methods, and results; and political ecology on the other (that of “the environmentalist,” or l’écologiste in French now, which signifies political parties, schedules, worries about certain constraints and certain values being brought into the political arena, etc). For several decades, citizens have been aware of this ambiguity (which is much more than a homonymy), since scientific ecology and political ecology have been in constant interaction with each other since the creation of the latter--the political concerns of ecology needing to be, at least in principle, reasonably informed of the facts as well as of the possibilities for action in the short and long term. To whoever asks--naively and as such preliminarily--what is biodiversity, multiple answers appear. The range of their variation is striking. On the one hand, ecologists (of communities, functionalities, landscapes) and biologists (systematists, geneticists, molecular biologists) present different and sophisticated notions of biodiversity--which are attached to complex respective measurement types which concern specific aspects of living beings (structure or function, species or genes, ecosystems or populations2). On the other hand, the promoters of 2010: The Year of Biodiversity list at the top of their web page this formula : “Biodiversity is life.” There is something like a short circuit here between these two types of statements--between the striking phrase which tries to raise more awareness of the issue with greater audiences, and the elaborate theoretical reflections of systematists, ecologists, or conservationists. Maybe we should ask ourselves what might justify such a simplification of the term. Still using expressions destined for a large public--once again being theoretical as well as exoteric, so that it seems legitimate to mobilize academic work as well as popular interest around the topic--the Biodiversity Council of California writes: “Why is biodiversity important? Everything that lives in an ecosystem is part of the Web of life, including humans. Each species of vegetation and each creature has a place on the earth and plays a vital role in the circle of life. Plant, animal, and insect species interact 2 For example, see Noss (1990) .

and depend upon one another for what each offers, such as food, shelter, oxygen, and soil enrichment.” Here, Biodiversity is life, because it maintains the “great network of life;” and we note that the writers here make reference to a general idea of balance in nature, the evidence of which has been rendered more fragile by the appearance of evolutionary biology (Pimm 2001) , and then by the multiple theoretical elaborations of ecology, of which chaotic dynamics models form a part (May 1974; Solé and Goodwin 1988) . Such contrast is a bit striking: whereas the acknowledgement of the value of biodiversity and, accordingly, the duty for human beings to take care of it is supposedly supported by the most recent science, the most public claims tend to call for ancient schemes of thought that have been precisely challenged by this science.
The same holds with the idea of “biodiversity loss”, actually: people are asked to help conserving species, or at least be sympathetic to the idea of preventing them from getting extinct – even though evolutionary theory has established for a long time that extinctions are an ordinary part of biological nature. Of course, what goes on with the “6th extinction”, as one calls it often, or the idea of “biodiversity loss”, is not the fact that some species go extinct, but the pace of this extinction, which by any estimation exceeds what could be considered as the normal rhythm of extinction – as well as the causal role human industry is playing in this process, be it directly, via illegal hunting or deforestation, or more indirectly, via climate change. Yet, this is not trivial to demonstrate: the incidence on human life and activities on biodiversity is not easy to estimate, and, regarding the pace of biodiversity loss, a skeptic mind could always question our estimations, given that it’s not even decided how many species exist in nature, controversies raging about several orders of magnitude. In any case, communicating about “biodiversity loss” seems to involve a drastic kind of simplification of what science actually unravels.
This example perhaps raises some difficulties to the idea of biodiversity itself: by trying to speak to a large number of people--non-scientists, in any case--it appeals to references, echoes, meanings that might be regarded as disputable or archaic within the scientific world. This would be a reason for the continual uneasiness which many ecologists and biologists show in regard to the rapid development of the concept, as is shown in the spirit of the already ancient remarks of Stuart Hurlbert regarding specific diversity (Hurlbert 1971). It could be that nothing ultimately justifies the unlikely jump from the multiple theoretical usages of “biological diversity” to this unique and concise phrase “Biodiversity is life” --whose essence and vital interest for the layman rests in its identification of life and biodiversity.
This inclines us to have a more precisely look on politic discourses on biodiversity. The 1992 Convention on Biological Diversity defined “biological diversity” in its Second Article as follows: “‘Biological diversity’ means the variability among living organisms from all sources including inter alia, terrestrial, marine and other aquatic ecosystems, and the ecological complexes of which they are a part; this includes diversity within species, between species and of ecosystems.” And indeed, if we look a

little closer in detail, it seems that we describe and evaluate biodiversity both in terms of multiple levels and multiple scales. Levels. In addition to a focus on species that was salient in what precedes, biodiversity is also concerned with genes and ecosystems. The different scientific disciplines are engaged at different levels--but it seems that species are the primary level of concern. This would reflect the fact that biodiversity was originally invented by ecologists, and above all reminds us that diversity in general has been the business of ecologists since the origins of the field3; in particular this has been the case for community ecology, population ecology (Kingsland 1995), and landscape ecology--even if there is tension throughout all ecological disciplines between an emphasis placed on living species in their diversity and the emphasis placed on ecosystems as complex and linked systems, potentially leading to an approach analogous to one of the physical sciences.
Obviously, as Lewontin (1974) wrote, all of biology tries to explain both adaptation and diversity; and, to keep our focus on biologists who have left their mark on the field, the great fresco of biology that Ernst Mayr wrote when he turned to the history and the philosophy of his field, The Growth of Biological Thought, is divided into three themes of which one is “Diversity” (Mayr 1982)... But, as much as it is an object of systematics and phylogenetic, diversity is very clearly a fundamental subject of ecology--particularly of community ecology – though from a very different angle. This science considers biological diversity on Earth – that is, on various places and landscapes on Earth - to be among its chief objects of explanation. It studies the patterns of diversity--which one can find at all levels--and the great regularities that govern these patterns : the species-area law, the specificities of ‘islands’ as significantly isolated ecosystems, the most frequent forms of species abundance distributions, the kinds of biodiversity patterns and dynamics proper to distinct types of ecological systems such as mangroves, deserts, tropical forests, etc.; it considers in a general manner the rules of species assembly in communities and assembly successions; it also aims to create measurements of this diversity and indexes to evaluate them; ultimately, it examines the processes responsible for these patterns--and in doing so, has been formulating rival theories for many decades. On the other hand, ecology also studies the effects of diversity on crucial properties of communities and ecosystems: their stability (in a number of different ways (Pimm 1984; Ives and Carpenter 2007), their resilience (Holling 1973), their functioning (Tilman 1996), etc. In most cases, this diversity is above all a diversity conceived at the level of species, even if recent programs of research in ecology combine this diversity at a specific level with a phylogenetic diversity conceived in a more genetic manner (e.g. Mouquet et al 2012). Scales. On the other hand, biodiversity can be described and evaluated on different scales: the biodiversity of a landscape, of an ecosystem--or more simply--of the Earth. While ecologists often focus on the first two, the focus is placed on the maximum scale when there is interaction with the general public. The “erosion of biodiversity” -- “biodiversity loss” -- is frequently presented in connection with what has been named the “sixth extinction”, which is evaluated on a scale that is temporally geological and 3.

spatially planetary (and which one has reasons to think that the effect of human activity precipitates it). This last scale characterizes a fundamental aspect of the usage of the concept of biodiversity: its merging of political and ethical questions, whose range involves all of humanity to some degree. It is here that the solidarity between the notions of “biodiversity” and of “climate change” -- which emerged at the same time and spread in a parallel fashion--is striking. In both cases, they give rise to a scientific double question which hinges on planetary political stakes. Thus, for biodiversity: is there an erosion of biodiversity, and what are its dimensions? If yes, how much is human activity responsible for it (and in what regards)? The general response is this: “Human actions [are] dismantling Earth’s ecosystems, eliminating genes, species, and biological traits at an alarming rate” (Cardinale et al 2012). This leads us to another theoretical question, which is more prospective: how much will this erosion affect us humans? As we indicated at the beginning of the essay, the discourse around biodiversity is focused on what can happen to us--as this statement by the one who more than anyone else popularized the term, the biologist Edward O. Wilson, illustrates in his book on the diversity of life: “It is reckless to suppose that biodiversity can be diminished indefinitely without threatening humanity itself.” (Wilson 1992). From here on out, scientific inquiry joins practical and political questioning: what can we do to prevent this declared erosion and disappearance? It thus seems that even the term biodiversity--by authorizing the usage of “diversity” on a global scale--allows for a simple conceptual link between specific theoretical questioning and the formulation of the practical and political stakes and options at play. In order to challenge the existence of such a simple conceptual link I am going to try - through an exploratory reflection on the theoretical concept of biodiversity - to characterize the status of the discourse surrounding biodiversity within the general framework of scientific ecology. 2. Biodiversity and the Diversities: The Contrast 2.1. Vague reference There are multiple meanings and multiple usages of “biodiversity;” but these meanings and these usages seem to gravitate around a sort of semantic attractor that is excessively rhetorical and made of a constellation of phrases, which easily translate one into the other: “biodiversity and life”, “biodiversity is life”, “biodiversity is an essential property of the Earth”, “living Earth”, etc. Because this term essentially speaks of a certain variability, “biodiversity” thus denotes the variation of life in general: “the degree of variation of life” on the English Wikipedia site, “the natural diversity of living organisms” on the French one (we note that this last formulation ultimately excludes ecosystems and possibly bacteria and viruses). As is shown, these formulations are very much faithful to the definition given by the Convention on Biological Diversity, which begins (as we have seen) with this sentence: “‘Biological diversity‘ means the variability among living organisms from all sources.” Otherwise said, the term biodiversity includes, among its other usages, a fundamental usage that confers a holistic tonality to life on Earth. It is in some way an extremely general concept--and ultimately a unifying one: whether the diversity is

specific, ecosystemic, genetic, allelic, structural, or functional, it still remains an avatar of biodiversity. We thus have here a concept whose meaning is extremely general; and at the same time, it is quite difficult to specify in a non-controversial manner what it is referring to exactly.
To make this latter point salient, think of the imagery of species extinctions: biodiversity loss evokes for many of us the poor polar bear alone on a shrinking banquise, the Orang-Outang losing their native forests in Sumatra, or the kakapo parrot (Strigops habroptila) who underwent a fascinating rescue operation in the 80s – all of them being so to say descendants of the Dodo, this first well known victim of human greediness. Yet, many conservationists and ecologists had warned us against focusing too much on these “extraordinary species” on the verge of extinction because of side-effects of our social or economic system; as Couvet and Vandevelde (2014) argued, rare species are only one sole aspect of biodiversity, next to which one must allow for what is called the “ordinary biodiversity” of more common, less-threatened organisms--these two kinds of biodiversity calling for very different discursive practices. Especially, whereas rare species trigger a sort of aesthetic understanding of biodiversity, the intrication of the less visible and more ordinary species with our ways of life implies that their drop in abundance would be less spectacular, and should be grasped by another kind of receptivity from our part. The aesthetic understanding of biodiversity is especially called upon when some economic valuations of biodiversity and ecosystem services, such as the ones that arose in the wake of the Millenium Ecosystems Assessment, asking people about whether they would pay for getting to see some specimen of a rare species in its natural environment4. Clearly, this contrasts with the kind of valuation of biodiversity that would support an action in favor of the brown squirrel or the red trout. So here, the vagueness of what “biodiversity loss” refers to is manifest in a split between various ways of experiencing biodiversity, and possibly, empathizing with the species or individuals at stake. It may even be the case that decisions regarding biodiversity conservation have to be taken in a context where rival valuations, based on distinct grasps of the value of biodiversity – i.e. an aesthetic grasp vs an instrumental grasp – don’t allow for a single most reasonable choice. Here, the lack of a clear reference of “biodiversity loss” resonates with the general trouble of environmental ethics, which are torn between valuations centered either on ecosystems, or on living beings, or on sentient beings in general (Millstein 2013). 2.2 Opposite pathways in semantics A general concept with an unstable reference point, biodiversity now seems to enter into striking contrast with what we call “diversity”--if one compares its semantic use in the evolution of ecological science and conservation biology over the past three decades. Indeed, at the same time that the term “biodiversity” was popularized in denoting the variation of life in general, ecology--in its diverse branches--built concepts that were more and more precise, rich and formalizable, that were able to capture a maximum of features (sometimes inter-contradictory) of what we intuitively understand as diversity. Of course, as Blandin (2014) or Gosselin (2014) describe with acuity, 4 See Maris (2014), for an overview of these economic evaluations.

ecologists from the start spoke of “diversity” along with other concepts that were both ambiguous and hardly operationalizable --capable of giving rise to measurements and diagnostics - which were against what one would spontaneously imagine with the word diversity. Species richness and then species evenness had already appeared in the middle of the Twentieth century as two components of what would eventually be called diversity--whether that of an ecosystem or of a community. Indeed, let’s compare a territory with 10,000 butterflies of species A, 10 insects of species B, 10 birds of species D and 10 fish of species E, with a territory with 10,000 organisms from each of these species: the second seems much more diverse, even , when one abstracts away from the compared abundances of each species (evenness), they would have the same value of diversity. The abundance of each species had also be taken into account, as much for the role it plays in intuitive judgments about the ordinary diversity of a community as for its effects on community dynamics: if all the species are greatly reduced (keeping fixed a high degree of species evenness), it is more difficult to imagine the beneficial and very frequently studied effect of diversity on community persistence--in particular, persistence in regard to stochastic fluctuations - since each species can quickly disappear following stochastic fluctuations, with the effect of drastically diminishing diversity.
However, specific abundance, specific richness and species evenness are insufficient if one wants to grasp the role that what we like to call diversity plays in the variation of certain variables which macroscopically describe the ecosystem (biomass, productivity – see Tilman (1996), or to take account of the evolutionary history of species and ecosystems (and therefore allelic diversity, which can be very high within one same species, whatever is its notion or its measurement). This is surely in issue regarding diversity measurements and the indices which determine this diversity (which relates back to the statistical tools that allow us to move from simple surveys of the individual presence or absence of different species within a sample to statistical descriptors of diversity), but it also, above all, defines theoretical problem concerning what can count as diversity in biology. Here, the sophistication and the thoroughness of diversity concepts go as far back as the history of theoretical ecology, to the emergence of the sectors and sub-disciplines within it. Functional diversity (as it is defined by the roles that species play in a community: herbivores, carnivores, predators, etc) has thus been regarded as something fundamental for two decades--from the moment that, as with functional ecology, we asked how an ecosystem could persist in time, could be resilient, could be the seat of chemical processes on grand scales, such as the flux of matter and energy governed by thermodynamic principles and so forth. In the 1950s, authors such as Eugene Odum insisted on the importance of an ecology whose base unit would be ecosystems rather than species (Odum 1953); this tradition gave rise to many problems within functional ecology, which are of exceptional interest to those who examine the sustainability of ecosystems and the consequences of human intervention on this sustainability--or, inversely, the usage of ecosystems to promote sustainable economic activities (such as those which ecological engineering aims toward (Jorgensen et al. (2008)).

Functional ecology has developed spectacularly over the past two decades: if diversity seems to be primarily a major concept for community ecology, it is however also important within functional ecology--although it is considered as “functional diversity,” which is instantiated by “functional types.” The theoretical question that arises now is how one can link the problem agendas of community ecology with those of functional ecology--and thus, among other issues, how to link the different types of diversity. In a famous paper, Ricklefs (2008) called for the “disintegration of ecological community,” arguing that placing emphasis on the ecological scale of the community seemed to make it blind to fundamental processes at other scales like region or metacommunity--these processes being responsible for important ecological patterns (such as the stability of communities). In a similar manner, it seems here that scientific ecology also invites a “disintegration” of diversity, in the sense that there is a proliferation of diversity concepts whose meanings do not seem to be able to be easily identified, juxtaposed or reconciled. The advent of the general term “biodiversity,” which denotes--less like a theoretical term than like the title of a problem set--the diversity of life as a whole seems to symmetrically respond to such disintegration--or the multiplication, if one prefers--of diversities in ecology between functional ecology and community ecology (but also between ecology, “macroecology” (ecology on the scale of meta-communities and beyond), and the theories of “eco-phylogenetics” (Mouquet et al. (2012), etc.). And this term would say as much or even more outside of scientific ecology as within it. This theoretical proliferation or disintegration of biodiversity is also scientifically important because the diversities do not play the same roles. Recently, Stéphanie d’Agata and colleagues (d’Agata et al. 2014) examined the loss of diversity in coral reefs. Like many studies, they indicated the importance of the anthropic factor on this loss of diversity. But, in relation to the topic at hand, they established three diversities: functional, phylogenetic, and specific diversity. The third--which has long been the central diversity of ecology, and which is that towards which the term “biodiversity” is most closely linked--declines slowly and linearly in relation to the growth of human density. Yet, when we look at phylogenetic and functional diversity, the authors show that these decline drastically and non-linearly as human densities rise. Thus, by distinguishing between the different concepts of diversity, which generally clot together to give a general concept of “biological diversity” or biodiversity, one can bring to light the real scale of the effects of human existence and activity on a certain type of ecosystem. 2.3. The diversity of explanatory models of biodiversity. Granted, all this is not about opposing biodiversity as both a political or ethical term and as a scientific term. In fact, in a number of scientific and academic contexts, “biodiversity” is used today in a manner that is comparable to “diversity”--the other term that was used to say the same things more than thirty years ago. Ecologists and conservationists invented and spread the term biodiversity and continue to use it in their publications. In fact, one of the books which loudly resonated among ecologists (Hubbell

2001) was called The Unified Neutral Theory of Biodiversity and Biogeography; even though the title Biodiversity, Biological Diversity, or simply Diversity, would have been enough to describe the contents of the book. But specifically, this contrast between biodiversity and the diversities which ecologists or biologists examine extends still further if we consider now not only the concepts, measures or indices of biodiversity, but also the theories and models themselves--where diversity is either an explanans or an explanandum. Thus, with his “unified theory,” Stephen Hubbell proposed a relatively new conception of the processes that generate diversity patterns that one can see in communities and metacommunities by arguing that the principles responsible for them are “neutral” processes—namely, processes analogues to what genetic drift is within evolutionary biology. (The neutralist theory of Hubbell is moreover explicitly built in correspondence with the neutralist theory of Motoo Kimura’s molecular evolution). Here, diversity is clearly the explanandum of the theory – unlike other theories in community ecology that question the consequences of some kinds of biodiversity (e.g. stability, productivity, any parameter of ecosystem functioning, etc.). More precisely, by “biodiversity”, Hubbell is mostly concerned with several key patterns of species diversity : the distribution of species abundances (which ones includes many individuals, which ones have few, etc.), sometimes modeled as species ranking (Fig. 4), less saliently the “species-area law” (which ties species richness to the variation in species area), etc. In addition to approaches that were already more stochastic than selective in community ecology, such as the island model of Mac Arthur and Wilson (1967), Hubbell’s theory confers a crucial role to the regional scale of the metacommunity, and its articulation with local processes within a focal community. Biodiversity emergence is modeled as a zero sum game in which at each time step a random individual dies and is randomly replaced by another individual from the metacommunity, birth and death rates being therefore equal for all individuals of all species which defines the “ecological equivalence assumption”, definitive of neutral models. As a result of this game, ecological drift in a community leads stochastically to the fixation of a single species. The neutral process as such is thereby defined as the articulation of ecological drift at the level of the community and of the metacommunity, where also events of speciation and colonization take place. Over the long term the metacommunity composition appears as the result of the balance between local ecological drift and speciation. Two parameters are therefore conditioning the neutral dynamics of biodiversity – I, the number of immigrants competing with local offspring to replace an individual in a focal community, and the parameter θ, called the “biodiversity number”, which captures the number of new species competing with regional offspring to replace a dead individual in the community. Figure 4. Examples of Species Abundance Distribution (SAD) , and its presentation as Species ranking.
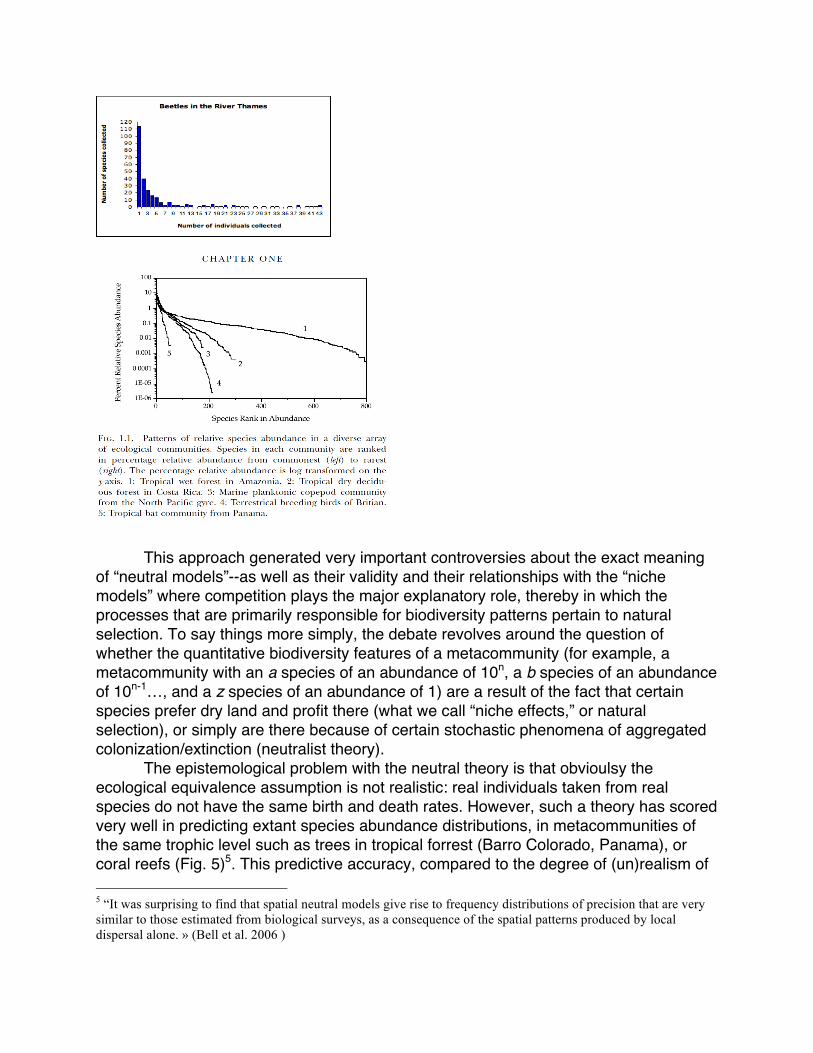
This approach generated very important controversies about the exact meaning of “neutral models”--as well as their validity and their relationships with the “niche models” where competition plays the major explanatory role, thereby in which the processes that are primarily responsible for biodiversity patterns pertain to natural selection. To say things more simply, the debate revolves around the question of whether the quantitative biodiversity features of a metacommunity (for example, a metacommunity with an a species of an abundance of 10n, a b species of an abundance of 10n-1…, and a z species of an abundance of 1) are a result of the fact that certain species prefer dry land and profit there (what we call “niche effects,” or natural selection), or simply are there because of certain stochastic phenomena of aggregated colonization/extinction (neutralist theory). The epistemological problem with the neutral theory is that obvioulsy the ecological equivalence assumption is not realistic: real individuals taken from real species do not have the same birth and death rates. However, such a theory has scored very well in predicting extant species abundance distributions, in metacommunities of the same trophic level such as trees in tropical forrest (Barro Colorado, Panama), or coral reefs (Fig. 5)5. This predictive accuracy, compared to the degree of (un)realism of 5 “It was surprising to find that spatial neutral models give rise to frequency distributions of precision that are very similar to those estimated from biological surveys, as a consequence of the spatial patterns produced by local dispersal alone. » (Bell et al. 2006 )
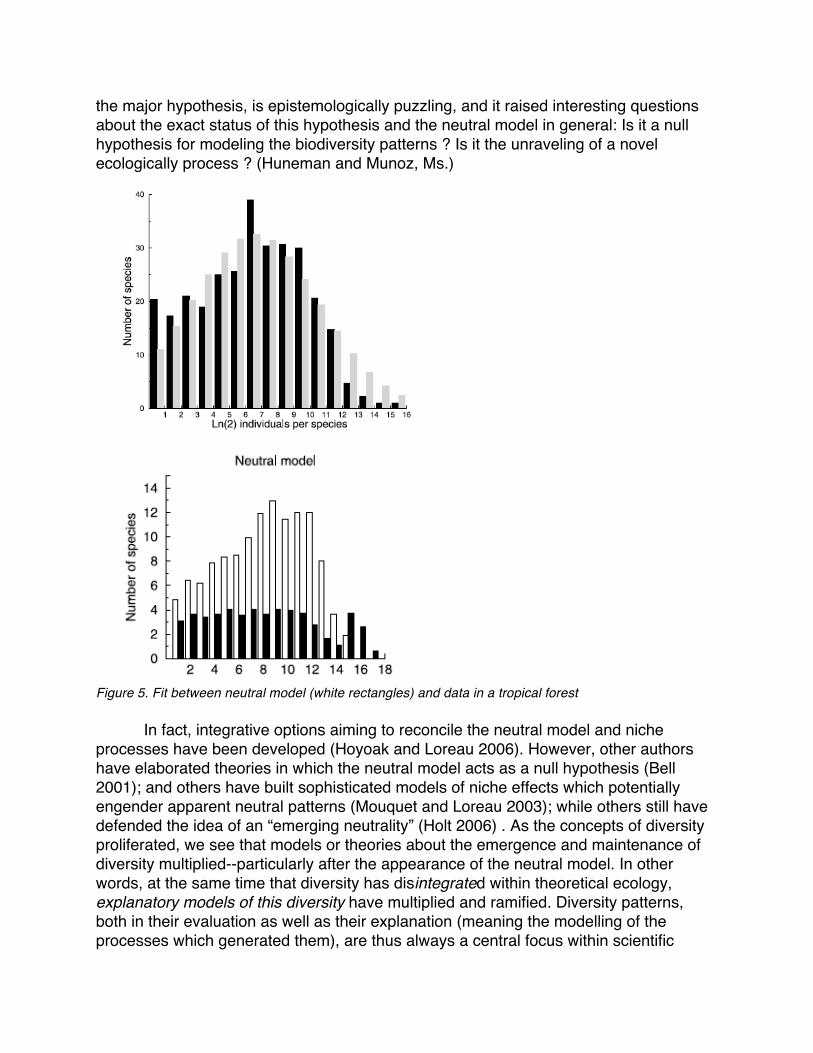
the major hypothesis, is epistemologically puzzling, and it raised interesting questions about the exact status of this hypothesis and the neutral model in general: Is it a null hypothesis for modeling the biodiversity patterns ? Is it the unraveling of a novel ecologically process ? (Huneman and Munoz, Ms.)
Figure 5. Fit between neutral model (white rectangles) and data in a tropical forest
In fact, integrative options aiming to reconcile the neutral model and niche processes have been developed (Hoyoak and Loreau 2006). However, other authors have elaborated theories in which the neutral model acts as a null hypothesis (Bell 2001); and others have built sophisticated models of niche effects which potentially engender apparent neutral patterns (Mouquet and Loreau 2003); while others still have defended the idea of an “emerging neutrality” (Holt 2006) . As the concepts of diversity proliferated, we see that models or theories about the emergence and maintenance of diversity multiplied--particularly after the appearance of the neutral model. In other words, at the same time that diversity has disintegrated within theoretical ecology, explanatory models of this diversity have multiplied and ramified. Diversity patterns, both in their evaluation as well as their explanation (meaning the modelling of the processes which generated them), are thus always a central focus within scientific

ecology. But the theoretical framework within which these ideas have appeared seems now to be too complex to be gathered within one unique and univocal conceptual structure. It is without a doubt significant that this double process was developed at the very moment where the term biodiversity began to rapidly develop--with its holistic ambition, and its all-encompassing global nature.
These remarks tend to draw a strange parallelism or even an antagonism between biodiversity and the diversity(ies) of biologists, considered in themselves and in the course of their diffusion and ramification within different discourses. On the one hand, we understand biological diversity such as it is defined by the 1992 Convention on Biological Diversity, which gave rise to the large concept (of perhaps even global and all-inclusive import) of “biodiversity”--which allowed people to establish conservation practices (and even to lobby policy makers), as well as ethical conversations (particularly in regard to the wilderness, the central subject of ecocentric environmental ethics), and even economic strategies of valorization (with all three emerging in a conceptual triangle defined by the notions of “sustainability,” “ecosystemic services,” and “biodiversity”)6. On the other hand, there are many specific ecological concepts of diversity, sometimes contradictory, and not to say anything of other peculiar diversity concepts to be found in other branches of biology such as molecular phylogeny, etc.: all of them fragmented and impossible to assemble as the potentially scattered features of one same concept (as they can give rise to contradictory measurements or evaluations) but also calling scientists for integrations and disciplinary syntheses (as is seen now between community ecology and functional ecology); yet above all giving rise to multiple formal modelized constructions whose elaboration and confrontation define in some sort the theoretical life of ecology. This paradox may not be unfamiliar to who studies the history of science. After all, “energy” is a term of daily use, that one will meet in tons of advertisements or TV programs, yet since its inception in natural philosophy by Leibniz’s investigation about “forces vives” (Guéroult 1967) it ramified into many specific terms in various theories in physics. However, what is striking in the case of “biodiversity” is that the term has been coined long after the thing – or, rather, some particular aspects of the thing - was already a crucial object of inquiry in many ecology or biology departments. And moreover, the coining of the term was in no respect independent from political and social contexts, where the concern for biodiversity preservation almost wholly overlapped with a critique of aspects of the capitalist modes of production or of social relationships. Hence, even if some dimensions of the process I just highlighted may correspond to very classical features of the life of theoretical terms in science once they get in contact with the concerns of a wider audience, there is still something singular in the case of biodiversity, which may boil down in the end to the normative or pejorative semantic load of the theoretical investigations in ecology, and, moreover to the emotional tone that surrounded the coining and use of the word.
The fact that “biodiversity” is wedded to the phrase “biodiversity loss” ultimately would explain that the pragmatics of this word may not exactly overlap with the one of 6 In regard to the these questions see Maris (2010), Meinard (2011).

specific concepts of biological diversity that have been elaborated in various subfields of ecology. This eventually suggests that a complete understanding of biological diversity should take into account both the plethora of often conflicting and sometimes articulated theoretical concepts of diversity (together with the models intended to explain biodiversity patterns) and the all-encompassing keyword “biodiversity” as a starting point for making sense of various acquaintances with the value of biological diversity. In the same way as cognition is now seen as “situated cognition” because the agent is not detached from the world where its cognition takes place and has effect (Shapiro, 2010), biodiversity is something like a “situated concept”, a concept that does not get its legitimacy from models, theoretical elaborations and empirical data, but from the realization that something is altered in the biological world as such – within which humans are living and evolving – even if positive science by its own means is not capable of uncontroversially and exactly capturing what is going on and pinpointing its causes.
Conclusion. Not only is the biodiversity that we experiment (and that which different discourses and practices introduce) volatile, fragile, and always capable of dwindling or of slipping between our fingers; but also, the notion itself of biodiversity is labile and almost intangible the moment we want to go beyond our intuitions of what it denotes to understand its contents in a distinct and articulated manner. This versatility, analyzed and illustrated throughout this paper, is however the reverse side of the fact that, less than an empirical concept or a theoretical term (sensu Hempel) biodiversity is a situated concept, whose content cannot be detached from the idea of a call for action that was inspirational when it has first been coined. As ecologists questioning biodiversity patterns or biodiversity loss, or as philosophers of ecology meditating on biodiversity concepts, we are in any sense the heirs of this coining, and of this context7. References. BELL G. (2001) “Neutral Macroecology”, Science, 293 (5539): 2413-2418 . BELL, G., LECHOWICZ, M.J., WATERWAY M.J. (2006) “The comparative evidence relating to the neutral theory of community ecology.” Ecology 87: 1378-1386 BLANDIN P (2014) “La diversité du vivant avant (et après) la biodiversité: repères historiques et épistémologique.” In Casetta E., Delord J. (eds.) La biodiversité en question. Paris: Matériologiques, pp.31-68. CARDINALE BJ et al., “Biodiversity Loss and its Impact on Humanity,” Nature, 486 (7401), 2012, 59-67
7 This paper uses extensive sections from the introduction of Casetta E., Delord J. (eds.) La biodiversité en question. (Paris: Matériologiques, 2014) written by the author, and translated by Adam Hocker. Thanks to Les Editions Materiologiques for authorizing to use them.

COUVET D, VANDEVELDE JC (2014) ”Biodiversité ordinaire: des enjeux écologiques au consensus social” In Casetta E., Delord J. (eds.) La biodiversité en question. Paris: Matériologiques, pp.183-208. D’AGATA S. ET AL. (2014) “Human-Mediated Loss of Phylogenetic and Functional Diversity in Coral Reef Fishes”, Current Biology, February 20 2014 GOSSELIN F (2014) “Diversité du vivant et crise d’extinction: des ambiguïtés persistantes.” In Casetta E., Delord J. (eds.) La biodiversité en question. Paris: Matériologiques, pp.119-138. GUÉROULT M. (1967) Leibniz, Dynamique et métaphysique. Paris: Aubier. HOLLING C. S. (1973) “Resilience and Stability of Ecological Systems,” Annual Review of Ecology and Systematics, 4: 1-23. HOLT ED (2006) “Emergent Neutrality”, Trends in Ecology & Evolution, 21(10): 531-533. HOLYOAK M. LOREAU M. (2006) “Reconciling Empirical Ecology with Neutral Community Models”, Ecology, 87(6): 1370-1377. HUBBELL S.P. (2001) The Unified Neutral Theory of Biodiversity and Biogeography, Princeton, Princeton University Press. HURLBERT S. H. (1971) “The non-concept of species diversity: a critique and alternative parameters,” Ecology, 52, 1971, 577-586 IVES R., J. CARPENTER J. (2007) “Stability and Diversity of Ecosystems,” Science, 317 (5834): 58-62 JORGENSEN S., GRANT, W., LEGOVIC, T., NIELSEN S. (2008) “New initiative for thematic issues: An invitation: Brief overview of the five "hot" themes.” Ecological modeling: 272-275 HUNEMAN P, MUNOZ F (MS) “The neutral theory of community ecology: is it null?” KINGSLAND S (1995) Modeling Nature: Episodes in the History of Population Ecology. 2nd edition, University of Chicago Press, 1995 LEWONTIN R (1974) The Genetic Basis of Evolutionary Change, New York, Columbia University Press, 1974 LOREAU M. (2010) “Linking Biodiversity and Ecosystems: Towards a Unifying Ecological Theory”, Philosophical Transactions of the Royal Society B, (365)1537: 49-60 . MAC ARTHUR R, WILSON E (1967). The Theory of Island Biogeography. Princeton, New Jersey: Princeton University Press. MARIS V (2014) Nature à vendre. MAY R. M. (1974) Stability and Complexity in Model Ecosystems. Princeton: Princeton University Press. MAYR E (1982) The Growth of Biological Thought. Diversity, Evolution, and Inheritance, Cambridge (Mass.): Belknap Press. MOUQUET N. ET AL. (2012) “Ecophylogenetics: Advances and Perspectives,” Biological Reviews, 87: 769-785 MOUQUET N., LOREAU M. (2003) “Community Patterns in Source-sink Metacommunities”, The American Naturalist, 162(5): 544-557. NOSS RF (1990) “Indicators for Monitoring Biodiversity: A Hierarchical Approach,” Conservation Biology, (4)4: 355-364. ODUM E (1953) Fundamentals of Ecology, Cambridge: Cambridge University Press. PIMM S. L. (1984) “The Complexity and Stability of Ecosystems,” Nature, 307, 1984, 321-326 PIMM SL The Balance of Nature? Ecological Issues in the Conservation of Species and Communities. Chicago: University of Chicago Press.

RICKLEFS RE (2008) “Disintegration of the Ecological Community”, American Naturalist, 172(6): 741-750. SHAPIRO L. (2010) Embodied cognition. London: Routledge. SOLÉ R., GOODWIN B. (1988) Signs of Life: How Complexity Pervades Biology. New York: Basic Book. TILMAN D. (1996) “Biodiversity: Population Versus Ecosystem Stability,” Ecology, 77: 350-363. WILSON E.O. (ed.), Biodiversity, Washington, National Academy Press, 1988 WILSON EO (1992) The Diversity of Life. Cambridge (Mass.): Belknap Press.