the presence of dinogymnium (dinophyceae) in lower cretaceous sediments from the northwest tethys...
TRANSCRIPT

REVIEW OF
PALAEOBOTANY AND
PALYNOLOGY E L S E V I E R Review of Palaeobotany and Palynology 92 (1996) 367 382
The presence of Dinogymnium (Dinophyceae) in Lower Cretaceous sediments from the northwest Tethys (southeast France and
western Switzerland) and Gulf of Mexico areas: stratigraphic and systematic consequences
Laurent Londeix a, Dominique Pourtoy b, James P.G. Fenton c
a Dkpartement de Gkologie et OcOanographie, UniversitO Bordeaux L Avenue des Facultks, 33405 Talence Ckdex, France b S.N. Elf-Aquitaine, Laboratoire de Gbologie, C.S.T.C.S., 64018 Pau, France c Simon Petroleum Technology (SPT), Llandudno, Gwynedd, LL30 1SA, UK
Received 10 August 1994; revised and accepted 20 September 1995
Abstract
Palynological investigations of two Lower Cretaceous surface sections from the northwestern Tethyan area, Chgttel- Saint-Denis (western Switzerland) and Vergons (southeast France), have yielded specimens of the fossil dinoflagellate Dinogymnium. These established in-situ occurrences in Hauterivian ammonite-dated sediments represent the first formal record of this genus in the Lower Cretaceous in the Tethyan area. A re-examination of the assemblages studied by Riley and Fenton (1984) from DSDP Site 535 in the Gulf of Mexico, has confirmed the in-situ occurrence of Dinogymnium in Upper Berriasian, Valanginian and Hauterivian sediments. Their comparison with the Tethyan material establishes the occurrence of Dinogymnium in the Lower Cretaceous of the Tethys-Atlantic area and leads us to reconsider the earliest occurrence of Dinogymnium which was previously generally accepted to be at the base of the Upper Cretaceous. Two new species are erected: Dinogymnium aerlicum and Dinogymnium vergonsense. Morphological, stratigraphical and palaeogeographical comparisons are made with other rare Early Cretaceous records of Dinogymnium, as well as with other, younger species of the genus.
1. Introduction
Species of Dinogymnium Evitt et al., 1967 are an important component of Upper Cretaceous dinoflagellate cyst assemblages from marine paleo- environments worldwide. Their distribution has been documented f rom Lower Cretaceous (very rarely) to Lower Paleocene (rarely) deposits, but the stratigraphical range of the genus is so com- monly presumed to be highly characteristic of Upper Cretaceous strata (e.g. Evitt et al., 1967, p. 5; Millioud et al., 1975, p. 68, ch. 3; Tappan,
0034-6667/96/$15.00 © 1996 Elsevier Science B.V. All rights reserved SSDI 0034-6667 ( 95 ) 00097-6
1980, p. 386; Riley and Fenton, 1984, p. 685; Evitt, 1985, pp. 74, 88-89, 175, 266 267; Goodman, 1987, pp. 653, 667, 675, fig. 15.2) that its rare, non-Upper Cretaceous occurrences have been con- sidered as most probably being the result of con- tamination or reworking (Evitt et al., 1967, p. 5; Harker and Sarjeant, 1975, p. 228; Evitt, 1985, p. 267).
A summary of the published records of Dinogymnium is presented in Fig. 1, featuring the number of citations of the genus from Cretaceous and Cenozoic sediments. This diagram has been
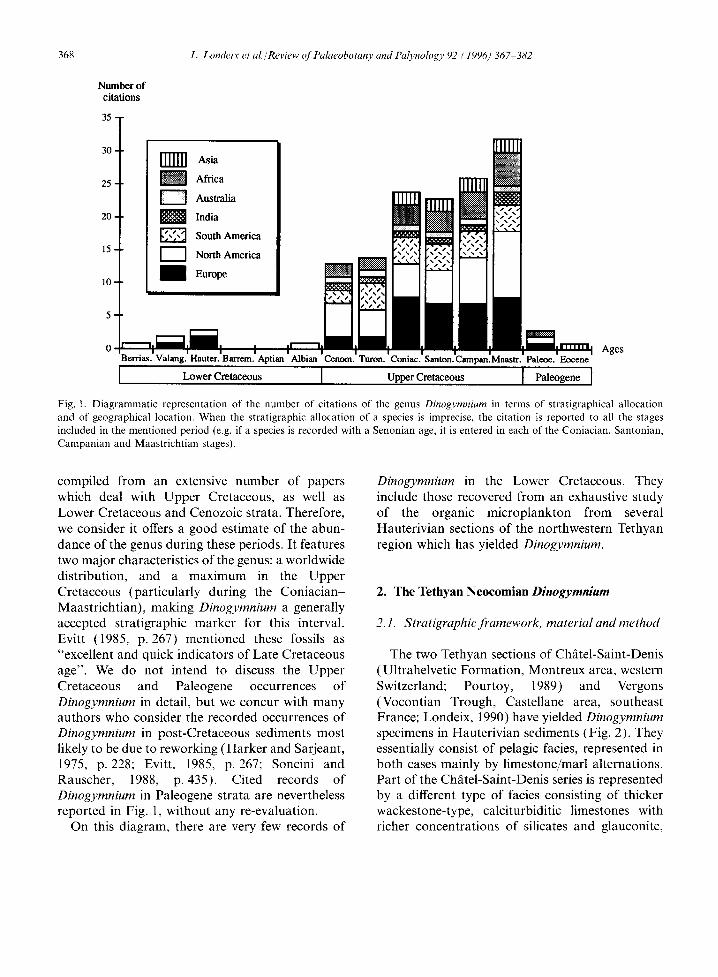
368 L. Londeix et al./Review of Palaeobotany and Palynology 92 (1996) 367 382
Number of citations
35
30-
25
20-
15-
10-
[Him Asia
i ~ Africa
Australia
India
South America
I I North America
Europe
5-
0 o
Berrias. Valang. Hauter. Barre~n. Aptian Albian Cenom. Turon. Coniac. Santon.Campan.Maastr. Paleoc. Eocene
[ Lower Cretaceous I Upper Cretaceous [ Paleogene I
Ages
Fig. 1. Diagrammatic representation of the number of citations of the genus Dinogymnium in terms of stratigraphical allocation and of geographical location. When the stratigraphic allocation of a species is imprecise, the citation is reported to all the stages included in the mentioned period (e.g. if a species is recorded with a Senonian age, it is entered in each of the Coniacian, Santonian, Campanian and Maastrichtian stages).
compiled from an extensive number of papers which deal with Upper Cretaceous, as well as Lower Cretaceous and Cenozoic strata. Therefore, we consider it offers a good estimate of the abun- dance of the genus during these periods. It features two major characteristics of the genus: a worldwide distribution, and a maximum in the Upper Cretaceous (particularly during the Coniacian- Maastrichtian), making Dinogymnium a generally accepted stratigraphic marker for this interval. Evitt (1985, p. 267) mentioned these fossils as "excellent and quick indicators of Late Cretaceous age". We do not intend to discuss the Upper Cretaceous and Paleogene occurrences of Dinogymnium in detail, but we concur with many authors who consider the recorded occurrences of Dinogymnium in post-Cretaceous sediments most likely to be due to reworking (Harker and Sarjeant, 1975, p. 228; Evitt, 1985, p. 267; Soncini and Rauscher, 1988, p. 435). Cited records of Dinogymnium in Paleogene strata are nevertheless reported in Fig. 1, without any re-evaluation.
On this diagram, there are very few records of
Dinogymnium in the Lower Cretaceous. They include those recovered from an exhaustive study of the organic microplankton from several Hauterivian sections of the northwestern Tethyan region which has yielded Dinogymnium.
2. The Tethyan Neocomian Dinogymnium
2.1. Stratigraphic framework, material and method
The two Tethyan sections of Chfitel-Saint-Denis (Ultrahelvetic Formation, Montreux area, western Switzerland; Pourtoy, 1989) and Vergons (Vocontian Trough, Castellane area, southeast France; Londeix, 1990) have yielded Dinogymnium specimens in Hauterivian sediments (Fig. 2). They essentially consist of pelagic facies, represented in both cases mainly by limestone/marl alternations. Part of the ChAttel-Saint-Denis series is represented by a different type of facies consisting of thicker wackestone-type, calciturbiditic limestones with richer concentrations of silicates and glauconite,

L. Londeix et al.lReview qf Palaeobotany and Palynology 92 (1996) 367-382 369
Fig. 2. Location of the outcrop sections of Chbtel-Saint-Denis
and Vergons.
referred to as the “Membre G&so-glauconieux”. The Chbtel-Saint-Denis Mesozoic section includes an incomplete sequence of Upper Berriasian- Lower Barremian strata; Upper Cretaceous sedi- ments are absent, tectonically removed by thrust- ing. In this section, the “Membre Greso- glauconieux” was dated as Early Hauterivian (equivalent to the Radiutus ammonite zone) using dinoflagellate cysts (Pourtoy, 1989) which have proved to be of great stratigraphical value in the absence of independent ammonite dating. The Vergons section includes Upper Valanginian- Lower Barremian strata. The stratigraphic control of these two sections was provided by a high-
resolution zonation based on ammonites studied by R. Busnardo (Lyon University, France).
The palynological analyses were done on outcrop samples taken at regular intervals in marly interbeds only. Samples were taken as deeply as possible in the beds in order to avoid sampling allochthonous material and thus to eliminate the risk of surface contamination. For each study, the samples were palynologically treated in different laboratories, following normal preparation pro- cedures with residues screened at 20 and 10 urn. In order to ensure that the sediments were not contaminated by Late Cretaceous material, either during sampling (although Late Cretaceous strata are not present near the outcrops) or palynological treatment, the samples which contained specimens of Dinogymnium were carefully reprocessed. Again, they yielded the same positive records of specimens of Dinogymnium.
2.2. Results
Considering the fact that Dinogymnium speci- mens were recovered from outcrop samples, the procedures used for sample processing, and the absence of any other palynomorphs of Upper Cretaceous age in preparations [cf. lists of taxa in Pourtoy (1989) and Londeix (1990); Upper Cretaceous sediments being furthermore absent at Chatel-Saint-Denis], we conclude that the presence of Dinogymnium specimens in these samples repre- sents in-situ occurrences of this genus in Tethyan Neocomian sediments.
Two main forms were distinguished in the Tethyan Hauterivian sediments:
( 1) Dinogymnium cf. acuminatum is recorded at Chatel-Saint-Denis (Fig. 3) in samples 152a, 156a and 161a in the Lower Hauterivian (Radiutus ammonite zone equivalent) sediments from the “Membre G&so-glauconieux”, as well as in sample 177 in the basal part of the Upper Hauterivian (Suyni ammonite zone) from the marl/limestone alternations facies. At Vergons, Dinogymnium cf. acuminatum was positively identified in sample H12 and questionably in sample H13, both in the Lower Hauterivian (Loryi ammonite zone).
(2) Dinogymnium vergonsense Londeix, Pourtoy and Fenton, sp. nov. was first recorded at Vergons

370 L. Londeix et al.IReview of Palaeobotany and Palynology 92 (1996) 367-382
AGE AMMONITE *s ZONES &@
Fig. 3. Records of Dinogymnium in the Tethyan sections of
ChPtel-Saint-Denis and Vergons, with the sample numbers
referred to in text. For the exact location of these samples, see
Pourtoy (1989) and Londeix ( 1990), respectively.
(Fig. 3) in sample H12 from the Lower Hauterivian (Lovyi ammonite zone). A further study of re-prepared samples yielded this species in sample 152a of the Chatel-Saint-Denis Lower Hauterivian @ad&us ammonite zone equivalent) sediments.
A single specimen of Dinogymnium sp. B was described by Londeix ( 1990, pp. 2 15-2 16, pl. XXVII,
2) from sample H12 at Vergons, and represents a potential third species. However, because of insufficient specimens and non-significant morpho- logical characteristics, we prefer to consider this specimen as falling within the range of variation of Dinogymnium cf. acuminatum, pending the dis- covery of more abundant material (see section Systematics below).
Considering the stratigraphic intervals studied in each section (i.e. mainly Hauterivian sediments), the observed occurrences of Dinogymnium in these two sections cannot yet be fully indicative of the actual stratigraphic range of this genus in the
Tethyan area and have, therefore, to be considered as partial, pending studies of more complete strati- graphic sections. It appears, however, that facies control does not interfere with the occurrence of the genus at Chatel-Saint-Denis.
3. Previous stratigraphic records of Dinogymnium in the Lower Cretaceous
Although extremely rare, Dinogymnium has been previously reported from Lower Cretaceous sedi- ments (see Table 1).
During a study on dinocyst assemblages from a Tethyan Neocomian section of the Villeneuve-de- Berg area (Ardeche, southeastern France), Monteil (pers. commun., 1985) found a specimen of Dinogymnium, referable to our Dinogymnium cf. acuminatum in an outcrop sample of Upper Valanginian hemipelagic facies sediments. Unfortunately, believing the occurrence was due to laboratory contamination, he did not report this discovery in his study.
The single specimen of Dinogymnium sp. A mentioned by Davey (1970, p. 360), originally reported from the Albian of International Yarbo Borehole No. 17 (Saskatchewan, Canada) is now reinterpreted as the result of caving from overlying Upper Cretaceous sediments in the well (Davey, pers. commun., 1992).
Habib ( 1972) reported the occurrence of Dinogymnium sp. A in core samples of Deep Sea Drilling Project Leg 11, offshore northwest Atlantic. This species is cited as a diagnostic species (Habib, 1972, p. 369) of Association B at Site 105 (cores 1 l-15), tentatively dated as Albian-Early Cenomanian on the basis of dinoflagellate assem- blages. The specimen illustrated (Habib, 1972, pl. 17, fig. 5) from Site lOlA, core sample 4-l is also dated as Albian-Early Cenomanian by foramini- fera and dinoflagellate assemblages. Unfortun- ately, no details are provided of the exact occur- rences of Dinogymnium sp. A within this strati- graphic interval at Sites 105 and 101A. It is therefore not possible to assess whether or not they represent a definite record of Dinogymnium in Lower Cretaceous (Albian) sediments of the northwest Atlantic.

L. Londeix et al./Review of Palaeobotany and Palynology 92 (1996) 367-382
Table 1 Previous known records of Lower Cretaceous Dinogymnium
371
Age Location References
Albian Canada (International Yarbo Borehole No. 17)
Albian/LowerCenomanian North American Basin (DSDP Sites 101 and 105)
Upper Berriasian- Gulf of Mexico (DSDP Site 535) Hauterivian Lower Hauterivian-Upper Hauterivian Lower Hauterivian
Western Switzerland
Southeast France
Davey (1970, p. 360, pl. 6, fig. 6)
Habib (1972, p. 369, pl. 17, fig. 5)
Riley and Fenton (1984, pp. 684-685, fig. 1, pl. 1, fig. 10) Pourtoy (1989, pp. 82-83, pl. 9, fig. 14 16)
Londeix (1990, pp. 214-217, fig. S.38, pl. xxvII, figs. 1 3)
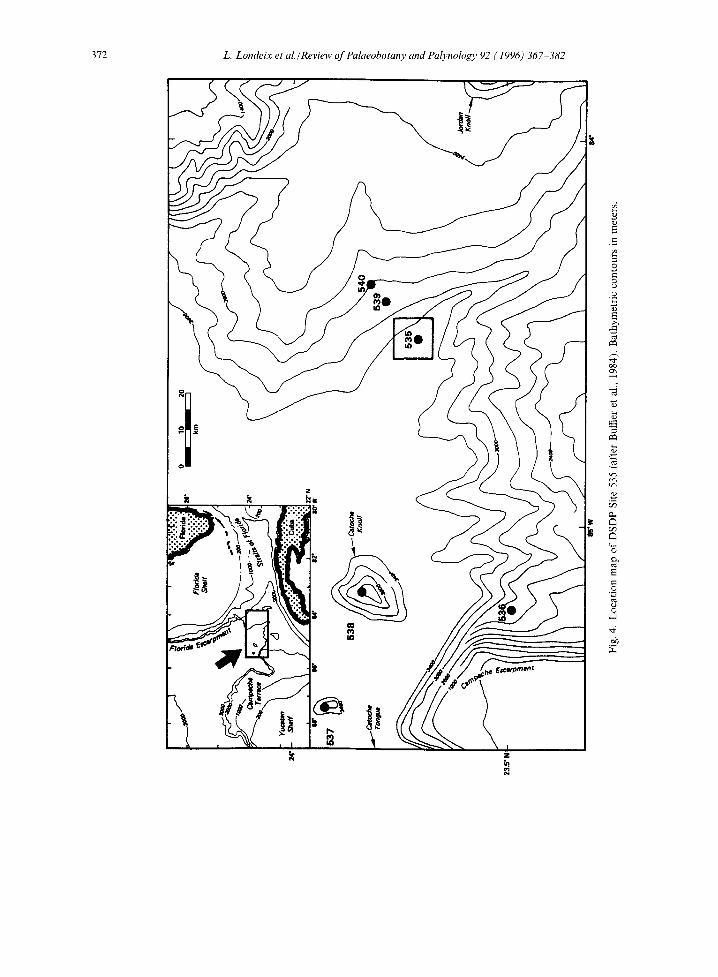
Riley and Fenton (1984) recorded Dinogymnium specimens in core samples from DSDP Site 535, offshore Gulf of Mexico (Fig. 4), in sediments representing a deep basinal depositional facies, dated as "late Berriasian-late Valanginian" and Hauterivian. Riley and Fenton (1984, p. 685) stated that "if not the result of contamination these occurrences may represent the earliest record of the genus". Further biostratigraphical and sedi- mentological analyses have proved that no Upper Cretaceous sediments are present in the well. Therefore, the possibility of contamination is excluded and the occurrences of Dinogymnium in Lower Cretaceous sediments in Site 535 are consid- ered in situ.
4. Revision of the specimens of Dinogymnium from D S D P site 535
Following the discovery of Dinogymnium in Tethyan Neocomian sediments, a revision of the material studied by Riley and Fenton (1984) has been undertaken. A review of the stratigraphy of the Lower Cretaceous at Site 535 based on the palaeontological synthesis of Leg 77 (Watkins and McNulty, 1984, fig. 5), and a palynological recon- sideration in the light of recent publications allows us to precisely date the Dinogymnium specimens (cf. Table 1). As a result of the comparison with the Tethyan material, it appears that two distinct species are present at Site 535. Although both forms were described as Dinogymnium sp. in the
original publication, Riley and Fenton (1984, p. 685) reported a variation in outline among the specimens, stating that "those from the late Berriasian-early Valanginian appear to possess pointed antapical poles, while those from the Hauterivian are generally subrounded".
The first type corresponds to Dinogymnium cf. acuminatum and is present at Site 535 in Valanginian (core sample 535-74-2/117-119cm) and Hauterivian (core samples 535-57-4/85-87 cm, 535-53-2/96-97 cm and 535-50-2/76-78 cm) sedi- ments. The specimens observed are closely compa- rable to the Tethyan material both in morphological features and size range, although there are slight intra-specific variations, which is characteristic of the genus due to the nature of the wall (Evitt et al., 1967, p. 6).
Despite the very low number of specimens observed, a new species, Dinogymnium aerlicum, is erected for the very distinctive second type. It seems to be restricted to Upper Berriasian sedi- ments at Site 535, where it is observed in core sample 535-79-1/52-53 cm only.
5. Conclusions
With the help of very well dated sections, and having established the in-situ character of its observed occurrences, we can state that the genus Dinogymnium was present in the northwestern Tethyan and northwestern Atlantic provinces during the Neocomian. Its new stratigraphic base
372 L. Londeix et al./Review of Palaeobotany and Palynology 92 (1996) 367-382
W
lID
E~ ,~ tnen e
.=_
o
3
.J
tg~
f ~
G
©
y.

L. Londeix et al./Review of Palaeobotany and Palynology 92 (1996) 367-382 373
is represented by the oldest occurrence recorded so far, in the Late Berriasian on the western side of the Nor th Atlantic (Gulf of Mexico). This also represents the earliest occurrence of the Family Gymnodiniaceae, in which we retain the genus Dinogymnium (see Systematics, Preliminary remarks below). Even though the occurrences of Dinogymnium in the Lower Cretaceous do not have a significant age-diagnostic value, its strati- graphical range can no longer be considered as exclusively Upper Cretaceous.
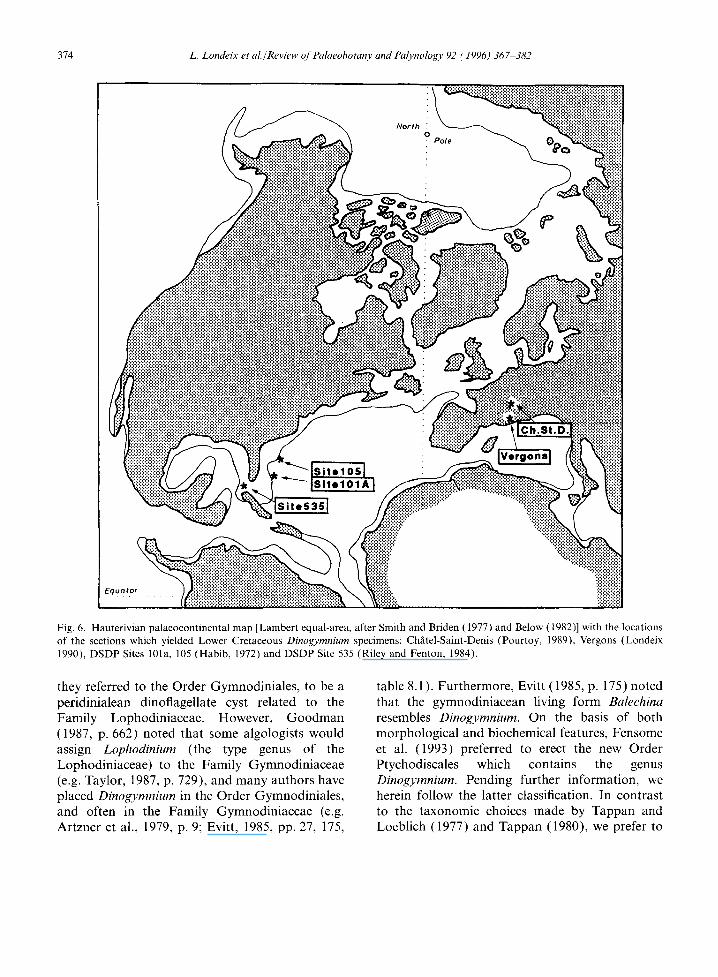
A summary of the proved or supposed in-situ early occurrences of Dinogymnium in the Lower Cretaceous known so far is presented in Fig. 5, showing an already widespread geographical pres- ence. Dinogymnium is recorded throughout the Neocomian in the Gulf of Mexico, including Upper Berriasian, Valanginian and Hauterivian sedi- ments. In the Tethyan area, it has been observed confidently in lower and basal Upper Hauterivian sediments only, but intensive studies of shallower and/or older and more complete Neocomian
~--. OI CENOMANIAN " ..a ual
! APTIAN "~
cz[ ,
> N E • ',
HAUTERVIAN - i Q
ua ~ , i _ s • i • i
VALANGINIAN, UPPER ~ ~' •° ~ ~-" e " i
BERRIASIAN
Fig. 5. Currently observed early stratigraphic occurrences of Dinogymnium in the Lower Cretaceous.
sequences may yield further records of this genus. Dinogymnium cf. acuminatum is present in all three sections and common to both provinces, with an overall stratigraphic range of Valanginian-Late Hauterivian, whereas Dinogymnium vergonsense is present in the Tethyan Early Hauterivian only, and Dinogymnium aerlicum is observed only in the Upper Berriasian of the Gulf of Mexico.
The similarities and probable communication between the two provinces during the Early Cretaceous is well known, bearing in mind that the North Atlantic Basin was then less than half of its present width (Fig. 6). Furthermore, the presence of Dinogymnium in the Early Cretaceous is unknown in the Boreal and Arctic areas, at least from published records. This tends to favour the hypothesis of a warmer water, if not tropical, affinity for the genus Dinogymnium.
The absence of records of Dinogymnium from Barremian and Aptian strata represents an impor- tant stratigraphic gap between the Lower Cretaceous occurrences that have been observed in this study and the subsequent ones reported from the Cenomanian onwards. It is a possibility that Dinogymnium species have hardly ever been reliably recorded in the Lower Cretaceous before because of their poorly preservable test [gymnodi- nioids have a mostly mucoid wall (Goodman, 1987, p. 676), or non acid-resistant wall], their small size, and their occurrence being commonly expected to be restricted to the Upper Cretaceous.
In both the northwestern Tethyan and north- western Atlantic (Gulf of Mexico) provinces, Dinogymnium specimens have been found in sedi- ments reflecting distal (basinal) environments, as opposed to a proximal, shallow environmental preference reported for this genus in the Upper Cretaceous (e.g. May, 1976, p. 110). Whether or not this might represent yet another characterisitic of Neocomian Dinogymnium in these areas needs to be determined by further studies.
6. Systematics
Preliminary remarks Tappan and Loeblich (1977) and Tappan (1980,
pp. 292, 302, 354) considered Dinogymnium, which
374 L. Londeix et al./Review of Palaeobotany and Palynology 92 (1996) 367 382
North 0
Pore
. . . . / !iiii!i!i!!i!iiiiii!i!!i!i!iiiiiii!i i i!ii! ii!!!!iii \\ i!iii !!iiii i i i!il . . . . . . . . . ~i~i~::~i!~.:.::!i:
Fig. 6. Hauterivian palaeocontinental map [Lambert equal-area, after Smith and Briden (1977) and Below (1982)] with the locations of the sections which yielded Lower Cretaceous Dinogymnium specimens: Chfitel-Saint-Denis (Pourtoy, 1989), Vergons (Londeix 1990), DSDP Sites 101a, 105 (Habib, 1972) and DSDP Site 535 (Riley and Fenton, 1984).
they referred to the Order Gymnodiniales, to be a peridinialean dinoflagellate cyst related to the Family Lophodiniaceae. However, Goodman (1987, p. 662) noted that some algologists would assign Lophodinium (the type genus of the Lophodiniaceae) to the Family Gymnodiniaceae (e.g. Taylor, 1987, p. 729), and many authors have placed Dinogymnium in the Order Gymnodiniales, and often in the Family Gymnodiniaceae (e.g. Artzner et al., 1979, p. 9; Evitt, 1985, pp. 27, 175,
table 8.1 ). Furthermore, Evitt ( 1985, p. 175) noted that the gymnodiniacean living form Balechina resembles Dinogymnium. On the basis of both morphological and biochemical features, Fensome et al. (1993) preferred to erect the new Order Ptychodiscales which contains the genus Dinogymnium. Pending further information, we herein follow the latter classification. In contrast to the taxonomic choices made by Tappan and Loeblich (1977) and Tappan (1980), we prefer to

L. Londeix et al./Review of Palaeobotany and Palynology 92 (1996) 367-382 375
retain Dinogymnium in the Family Gymnodinia- ceae.
May (1976) reported Dinogymnium from the Upper Cretaceous of New Jersey with a flagellar pore in the ventral region, thus making Dinogymnium an exceptional case of fossilized motile dinoflagellate cells rather than cysts (Evitt, 1985, pp. 89, 174; Sournia, 1986, p. 27). Consequently, and as suggested by Evitt (1985, pp. 174, 266), we shall use the term "test" in the following descriptions in preference to "cyst". For the same reason, we follow the recommendations stated by Evitt (1985, p. 266) that "the para- terminology applied to features on the cyst is not used. However, with admitted inconsistency but in the interest of simplicity, the opening at the apex will be called the 'archeopyle', using quotation marks to indicate that this may be an extension of the customary connotation". As a result, the classi- fication followed here is not the one commonly used to describe dinocysts, but the one regularly used by algologists.
Several authors have mentioned the occasionally great variability of morphologic features within an intra-specific population of Dinogymnium, such as size, ambitus and ornamentation (ridges and folds) and, therefore, the subsequent problems for estab- lishing and recognising specific criteria. In particu- lar, the observed differences in ambitus and size are probably mechanically affected by distortion of the tests due to the nature of the wall in Dinogymnium, in addition to normal inter-specific gradation. Nevertheless, some morphologic fea- tures such as "the cingulum index, the distribution of trichocyst pores and surface granulation, and general pattern of ribbing appear to be less vari- able" according to Evitt (1985, p. 267), as well as probably the wall thickness.
These morphological criteria have been used to define the new species recovered from the Tethyan and Gulf of Mexico material. The cingulum index sensu Evitt (1985, p. 267) is defined as "the decimal quotient of the distance from apex to midwidth of cingulum divided by the total length of the polar axis".
All the slides containing the type specimens of the new species described hereafter are housed in the D6partement de G6ologie et Oc6anographie
(D.G.O.), Universit6 Bordeaux I, Avenue des Facult6s, 33405, Talence C6dex, France.
Division DrNOFLAGELLATA (Biitschli, 1885 ) Fensome et al., 1993
Class DINOPHYCEAE Pascher, 1914 Order PTYCHODISCALES Fensome et al., 1993 Family PTYCHODISCACEAE Willey and Hickson,
1909 Sub-Family OINOCYMNIOIOEAE (Sarjeant and
Downie, 1974) Fensome et al., 1993
Genus Dinogymnium Evitt, Clarke and Verdier, 1967 emend. Lentin and Vozzhennikova, 1990
Type: Dinogymnium acuminatum Evitt, Clarke and Verdier, 1967. Remarks: Part of the emendation of Dinogymnium made by Lentin and Vozzhennikova (1990, pp. 14-15) excludes from this genus a number of species showing a bowl-shaped hypotest, in which folds or ridges are faintly expressed or absent. They transrerred these species to Alisogymnium Lentin and Vozzhennikova, 1990. Considering the variability in shape, ambitus and development of ornamental features observed in the present mate- rial, we prefer not to follow such a restricted definition of Dinogymnium and we, therefore, con- sider the new species erected herein to belong to Dinogymnium emended by Lentin and Vozzhennikova (1990) sensu lato.
Dinogymnium acuminatum Evitt, Clarke and Verdier, 1967
1967 Dinogymnium acuminatum sp. nov. Evitt, Clarke and Verdier, pp. 8-16, pl. 1, pl. 2, pl. 3, figs. 1-8, 10, 12, 20; text-fig. 11 23
Dinogymnium cf. acuminatum (Fig. 7b-d; Plate I, 4-9)
1984 Dinogymnium sp., Riley and Fenton, pp. 684-685, pl. 1, fig. 11
1989 Dinogymnium sp., Pourtoy, pp. 82-83, pl. 9, figs. 14-16. ?1990 Dinogymnium sp. B, Londeix, pp. 215-216, fig. S.38B,
pl. XXVll, 2 ?1990 Dinogymnium sp. C, Londeix, pp. 216-217, fig. S.38c,
pl. xxvn, 3
376 L. Londeix et al./Review of Palaeobotany and Palynology 92 (1996) 36~382
20 ,urn
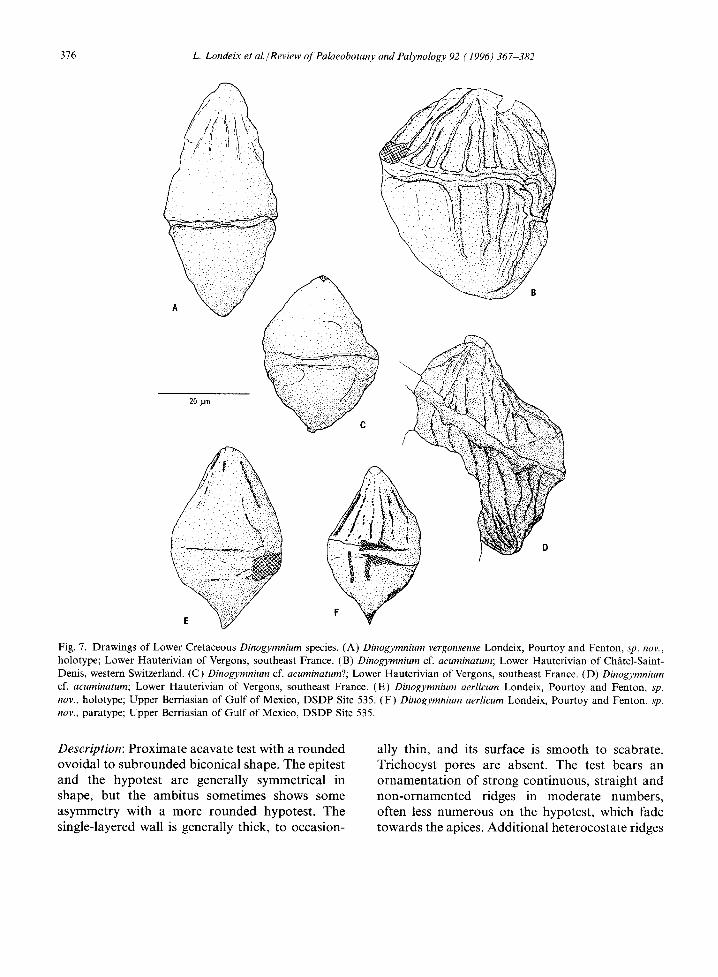
Fig. 7. Drawings of Lower Cretaceous Dinogymnium species. (A) Dinogymnium vergonsense Londeix, Pourtoy and Fenton, sp. nov., holotype; Lower Hauterivian of Vergons, southeast France. (B) Dinogymnium cf. acuminatum; Lower Hauterivian of Chgttel-Saint- Denis, western Switzerland. (C) Dinogymnium cf. acuminatum?; Lower Hauterivian of Vergons, southeast France. (D) Dinogymnium cf. acuminatum; Lower Hauterivian of Vergons, southeast France. (E) Dinogymnium aerlicum Londeix, Pourtoy and Fenton, sp. nov., holotype; Upper Berriasian of Gulf of Mexico, DSDP Site 535. (F) Dinogymnium aerHcum Londeix, Pourtoy and Fenton, sp. nov., paratype; Upper Berriasian of Gulf of Mexico, DSDP Site 535.
Description: Proximate acavate test with a rounded ovoidal to subrounded biconical shape. The epitest and the hypotest are generally symmetrical in shape, but the ambitus sometimes shows some asymmetry with a more rounded hypotest. The single-layered wall is generally thick, to occasion-
ally thin, and its surface is smooth to scabrate. Trichocyst pores are absent. The test bears an ornamentation of strong continuous, straight and non-ornamented ridges in moderate numbers, often less numerous on the hypotest, which fade towards the apices. Additional heterocostate ridges

L. L~ndeix et al./Review of Palaeobotany and Palynology 92 (1996) 367-382 377
are absent. The cingulum is relatively wide, well marked and variably indented. The "archeopyle" is apical, recognizable as a triangular to circular opening on the apex. The operculum is attached or missing.
Dimensions Chdtel-Saint-Denis section: Test length=35-(46)551am;
epitestal length = 17.5-(22.5) 25 I~m; hypotestal length = 15-(21.5)-30 ~tm; test breadth=30-(39)-47.5 ~tm; cingulum height=3-(5.5)-7.5 Ixm; cingulum index=0.45-(0.52)-0.64; number of measured specimens = 6. Vergons section: Test length = 50-( 56)-62 ~tm; epitestal length = 27-(30) 32gm; hypotestal length=281am; test breadth= 34-(36)-37 ~tm; cingulum height=2-(5)-8 gm; cingulum index = 0.50; number of measured specimens = 2. DSDP Site 535: Test length = 33-(38)-45 ~tm; epitestal length = 12-(15) 21~rn; hypotestal length=14-(18.5)-22~n; test breadth = 30-(36)-40 ~tm; cingulum height = 3-(6)-9 ~tm; cin- gulum index=0.38-(0.46)-0.60; number of measured speci- mens = 5. Occurrence: Chgttel-Saint-Denis: Lower Haute- rivian, Radiatus ammonite zone equivalent (samples 152a, 156a and 161a)-basal Upper Hauterivian, Sayni ammonite zone (sample 177); Vergons: Lower Hauterivian, Loryi ammonite zone (samples H12 and ?H13); DSDP Site 535: Valanginian-Hauterivian (core samples 535-57- 4/85-87 cm, 535-53-2/96-97 cm and 535-50- 2/76-78 cm). Remarks: Dinogymnium cf. acuminatum is best represented of the three types of Dinogymnium encountered in all three Lower Cretaceous sections studied here. It shows variability in a number of morphological features with minor differences observable both within a given population and between different populations (i.e. from the west- ern Tethys and Gulf of Mexico). Thus, this vari- ability as such does not represent a geographical identity for a given population. It involves mainly test shape (more or less subrounded), symmetry, and the number of ridges on the surface and how strongly they are marked. All these features are affected by preservation and the degree of compres- sion of the tests (see above). A relative symmetry of the test with an average cingulum index of 0.50, a moderate number of straight, non-ornamented and well-marked ridges, often less numerous on the hypotest, seem to be characteristic of D. cf. acuminatum. The wall of the test appears to be
generally thick, although the much thinner wall exhibited by some specimens from Site 535 could be due to preservational factors. Also characteristic of this form are the absence of trichocyst pores, of additional heterocostate ridges and of any sur- face ornamentation or sculpture.
The single specimen of Dinogymnium sp. B reported at Vergons from sample H12 (Loryi ammonite zone) by Londeix (1990, p. 215-216) and described as being devoid of any ribs or ridges, is thought to fall within the range of variability of D. cf. acuminatum, pending the study of more specimens. Comparisons: Dinogymnium cf. acuminatum clearly belongs to the morphological group ofD. acumina- turn Evitt et al., 1967/D. heterocostatum (Deflandre, 1935) Evitt et al., 1967, from which it is distinguished by the absence of additional heter- ocostate ridges and of trichocyst pores, by a more rounded shape and a thicker wall. The well- rounded specimens are similar in shape to D. albertii Clarke and Verdier, 1967, but differ from this species by the absence of micronegative sculp- ture on both the grooves and ridges.
Its straight, continuous and unornamented ridges distinguish D. cf. acuminatum from a number of species. Dinogymnium westralium (Cookson and Eisenack, 1958) Evitt et al., 1967 differs from D. cf. acuminatum in bearing pustulate ribs. Dinogymnium microgranulosum Clarke and Verdier, 1967 has ribs sculptured with scabrae and a microgranular surface. This latter species was considered to be a junior synonym of D. acumina- turn by Lentin and Vozzhennikova (1990, p. 15).
The Lower Cretaceous specimen encountered and illustrated by Habib (1972, pl. 17, fig. 15) as Dinogymnium sp. A may be referable to D. cf. acuminatum, but the lack of description for this taxon prevents a closer comparison.
Dinogymnium vergonsense Londeix, Pourtoy and Fenton, sp. nov. (Fig. 7a; Plate I, 1-3)
1990 Dinogymnium sp. A, Londeix, pp. 214-215, fig. S.38a; pl. xxvn, 1
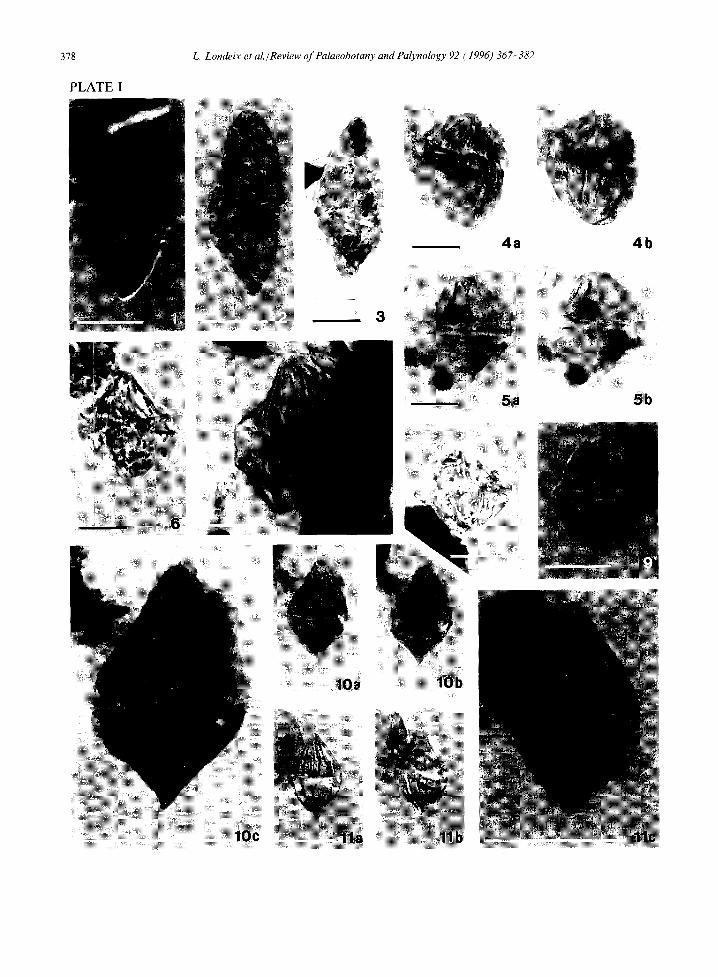
Holotype: Plate I, 1; Fig. 7a. Slide H12-20g-1 (England Finder reference: M 47/0).
378
PLATE I
L. Londeix et al./Review of Palaeobotany and Palynology 92 (1996) 367-382
4 8
~i l ! ~iii i ii! ~II
4 b
3
i~!ii~ ~ ~ ~!i~!~!~ii~!~!~i~!~i~i~i~i~i~!~i~Ji~!~i~i~!!iiii ~ ili!iiiii~ i ~i~J ~!~i~i~:!~i~i~i~i~i~i~i~!~i~i~i~i~i~i~i~i~i~i~i~i~i~ii~ii~ii~ii~
............ ~iii~i!!!!ii~!!i! ~'~ i ~ !!~ii~i~iiiii~i~,~i~iiii!~!i!iii!i!iiii!ii!iiiiiiii
~ii~ii~ii!iiiiiiiiiiiiiiiiiiiU~i~7111!!iiiiiiiill ̧ ~
~ i ~ i i ~ i i i ~ 5 ¸

L. Londeix et al./Review of Palaeobotany and Palynology 92 (1996) 367-382 379
Paratype: Pla te I, 2. Slide H12-101a-1 ( E n g l a n d F i n d e r reference: Q 50/1). Repository: D.G.O . , Univers i t6 B o r d e a u x I. Type locality: Vergons sect ion ( A l p e s - d e - H a u t e - Provence , sou theas t France) . Occurrence: Vergons: L o w e r Hau te r iv ian , Loryi
a m m o n i t e zone (sample H 12); Chgttel- S a in t -Denis : L o w e r Hau te r iv ian , Radiatus a m m o n i t e zone equiva lent (sample 152a). Derivation o f name: F r o m the name o f the type section, Vergons. Diagnosis: Prox ima te , acava te a symmet r i ca l test wi th an e longa te b iconica l shape and r o u n d e d poles. The s ingle- layered wall is th ick and its surface is s m o o t h to scabrate . Tr ichocys t pores are absent . A re la t ively wel l -depressed n a r r o w cingu- lum divides the test in to two dis t inct ive par ts . The epi test is s o m e w h a t longer than the hypotes t . I t bears a few fine r ibs or folds converg ing t owards the u n o r n a m e n t e d apex, which, however , is no t r ibbed. A d d i t i o n a l he te rocos ta t e r idges are absent . The hypo te s t is u n o r n a m e n t e d . The test seems to be wider in the p rec ingu la r a rea c o m p a r e d with the pos tc ingu la r area. The " a r c h e o p y l e " is apical , recognizable as a t r i angu la r to c i rcular open ing at the apex, wi th the ope rcu lum being genera l ly a t tached .
Dimensions
Holotype: Test length=54 ~tm; epitestal length=33 Ixm; hypo-
testal length = 20 ~n; test breadth = 26 ~tm; cingulum height = 2 ~-n; cingulum index = 0.61. Paratype: Test length= 59 p_m; epitestal length =33 ~tm; hypo- testal length = 25 ~tm; test breadth = 23 txm; cingulum height = 5 ~tm; cingulum index=0.57. Ch(ttel-Saint-Denis section: Test length = 70 txm; epitestal length = 42 ~m; hypotestal length = 25 Ixm; test breadth = 30 ~tm; cingulum height =2 ~tm; cingulum index=0.60; number of measured specimens = 1.
Comparisons: I ts dis t inct ive e longate b iconica l shape, c ingu lum index and scarce to absen t r idges on the epicyst only different iate D. vergonsense
f rom the o ther Lower Cre taceous species o f Dinogymnium.
Dinogymnium sp. A Ja in and Mil lepied, 1975 is morpho log i c a l l y very close to D. vergonsense
Londe ix , P o u r t o y and Fen ton , sp. nov. in terms o f shape, o r n a m e n t a t i o n , and c ingu lum index. Never theless , Dinogymnium sp. A seems to have a much th inner wall and bears folds and r idges on bo th the epi- and the hypo- tes t , wi th a size near ly doub le to tha t o f the Te thyan mater ia l . Dinogymnium digitus (Def landre , 1935) Evi t t et al., 1967, erected on the basis o f a single specimen, seems to have a more e longate shape, an aproxi - ma te ly equi la ten t epi- and hypotes t , and was descr ibed by Def tandre (1936, p. 12) as hav ing long i tud ina l ly a r r anged " punc tua t i ons " . Dinogymnium vergonsense Londeix , P o u r t o y and Fen ton , sp. nov. is somewha t s imilar to D. nelso-
nense (Cookson , 1956) Evi t t et. al., 1967 in the
PLATEI
Scale bars = 20 ~tm. 1. Dinogymnium vergonsense Londeix, Pourtoy and Fenton, sp. nov. Holotype. H12-20 ix-1. Lower Hauterivian, Vergons.
Phase contrast. 2. Dinogymnium vergonsense Londeix, Pourtoy and Fenton, sp. nov. Paratype. H12-10 I~-1. Lower Hauterivian, Vergons.
Phase contrast. 3. Dinogymnium vergonsense Londeix, Pourtoy and Fenton, ap. nov. CSD 152a-3. Lower Hauterivian, Chgttel-Saint-Denis.
Interference contrast. 4. Dinogymnium cf. acuminatum. CSD 152a-1. Lower Hauterivian, Chgttel-Saint-Denis. Interference contrast. 5. Dinogyrnnium cf. acuminatum. DSDP Site 538, 73-3 (117-119). Upper Berriasian, Gulf of Mexico. Interference contrast. 6. Dinogymnium cf. acuminatum. DSDP Site 535, 53-2 (96-97). Upper Berriasian, Gulf of Mexico. Phase contrast. 7. Dinogymnium cf. acuminatum. H12-20 ~t-1. Lower Hauterivian, Vergons. Phase contrast. 8. Dinogymnium cf. acuminatum. CSD 152a-1. Lower Hauterivian, Ch~tel-Saint-Denis. Phase contrast. 9. Dinogyrnnium cf. acuminatum? H12-10 Ix-1. Lower Hauterivian, Vergons. Phase contrast. 10. Dinogymnium aerlicum Londeix, Pourtoy and Fenton, sp. nov. Paratype. DSDP Site 535, 79-1 (52 53). Upper Berriasian, Gulf
of Mexico. a,b. Interference contrast, c. Phase contrast. 11. Dinogymnium aerlicum Londeix, Pourtoy and Fenton, sp. nov. Holotype. DSDP Site 535, 79-1 (52 53). Upper Berriasian,
Gulf of Mexico. a,b. Interference contrast, c. Phase contrast.

380 L. Londeix et al./Review of Palaeobotany and Palynology 92 (1996) 367-382
shape of the asymmetrical test, and its cingulum index, in particular the paratype (Cookson, 1956, pl. 1, fig. 9). However, D. nelsonense has a much thinner wall, longitudinally arranged granulations and has folds on both the epi- and hypotest (note: the holotype and the two other designated para- types are illustrated upside down; Cookson, 1956, pl. 1, figs. 8, 11).
Species formerly attributed to Dinogymnium are possessing a faintly ribbed or unornamented epitest and a typically unribbed hypotest which are further characterised by a bowl-shaped hypotest have been transferred to the genus Alisogymnium Lentin and Vozzhennikova, 1990: A. assamicum (Jain et al., 1975), A. deflandrei (Boltenhagen, 1977), A. evittii (Boltenhagen, 1977), and A. euclaense (Cookson and Eisenack, 1970).
Amphigymnium mitratum (Vozzhennikova, 1967) Lentin and Vozzhennikova, t990 has a very thin wall, a rounded antapex and a very short epitest (cingulum index = 0.24).
Dinogymnium aerlicum Londeix, Pourtoy and Fenton, sp. nov. (Fig. 7e, f; Plate I, 10a-c, 1 l ~ c )
1984 Dinogyrnnium sp., Riley and Fenton, pp. 684 685, pl. 1, fig. 10
Holotype: PlateI , 11; Fig. 7e. Slide 79-1, 52-53 (England Finder reference: M 34/0). Paratype: Plate I, 10; Fig. 7f. Slide 79-1, 52-53. (England Finder reference: N 39/2) Repository: D.G.O., Universit6 Bordeaux I. Type locality: DSDP Site 535, Gulf of Mexico. Occurrence: DSDP Site 535: Upper Berriasian (core sample 535-79-1/52-53 cm.) Derivation o f name: From the latin aerlicus, in reference to its stratigraphic occurrence represent- ing the earliest of the genus. Diagnosis: Proximate acavate asymmetrical test with a biconical shape. The single-layered wall is thick and its surface is smooth to scabrate. Trichocyst pores are absent. The conical epitest has a rounded apex and is similar in size to the more rounded hypotest which exhibits a distinctly pointed antapex. Ornamentation is more devel- oped on the epitest which bears a few strongly to faintly marked ridges. Ridges are faintly expressed
or absent on the hypotest. Additional heterocostate ridges are absent. The cingulum is well depressed and relatively wide. The 'archeopyle' is apical, recognizable as a triangular to circular opening on the apex. The operculum is either attached or missing.
Dimensions Holotype: Test length=38 ~tm; epitestal length=18 gm;
hypotestal length=141am; test breadth=23gm; cingulum height = 2 gm; cingulum index = 0.50. Paratype: Test length-43 gm; epitestal length=23 grn; hypo- testal length = 19 ~tm; test breadth = 25 gm; cingulum height = 3 ~tm; cingulum index =0.53. Overall size range: Test length=35 (38)43gm; epitestal length= 18 (19)-23 p.m; hypotestal length= 13-(15) 19 gm; test breadth=22 (23.5)-25 p.m; cingulum height= 3 (3) 3 gm; cingulum index=0.50-(0.52) 0.54; number of measured specimens = 3.
Remarks': Although very few specimens of this species have been found, its morphological features are regarded as sufficiently characteristic for the erection of a new species. One of the specimens exhibits an antapical bulge instead of a distinctively pointed antapex and bears very faintly expressed ridges compared to the other two. This probably reflects the intra-specific variability typical of the genus or possibly variations in quality of preserva- tion. Dinogymnium aerlicum Londeix, Pourtoy and Fenton, sp. nov. represents the earliest known record of the genus Dinogymnium. Comparisons: Its asymmetrical biconical shape and distinctively pointed antapex, in addition to its very small size distinguishes D. aerlicum Londeix, Pourtoy and Fenton, sp. nov. from the other Lower Cretaceous species of Dinogymnium although it is otherwise very close to D. cf. acuminaturn.
Dinogymnium biconicum Jain and Millepied, 1975 has a slightly pointed apex and antapex, but its size range is between 5 and 8 times greater than that of D. aerlicum Londeix, Pourtoy and Fenton, sp. nov.
Acknowledgements
We wish to thank Simon Petroleum Technology (SPT) for their technical and financial support, and permission to publish this paper. Our gratitude

L. Londeix et al./Review of Palaeobotany and Palynology 92 (1996) 36~382 381
also goes to Dr. Roger Davey (SPT) and Dr. Eric Monteil (IKU Petrol. Res., Norway) for their kind help, discussions and corrections of this manu- script, Dr. Nicos Ioannides (Esso-REP, France) and Dr. Jean-Louis Turon (Bordeaux University, France) for their substantial assistance.
References
Artzner, D., Davies, E.H., D6rh6fer, G., Fasola, A., Norris, G. and Poplawski, S., 1979. A systematic illustrated guide to fossil organic-walled dinoflagellate genera. R. Ont. Mus. Life Sci. Misc. Publ., pp. 1 119.
Below, R., 1982. Dinoflagellate cysts from Valanginian to lower Hauterivian sections near Ait Hamouch, Morocco. Rev. Esp. Micropaleontol., 14: 23-32.
Boltenhagen, E., 1977. Microplancton du Cr6tac6 sup6rieur du Gabon. Cah. Pal6ontol., pp. 1-50.
Burlier, R.T., Schlager, W. and Pisciotto, K.A., 1984. Introduction and explanatory notes, knit. Rep. DSDP, 77: 5-22.
Biitschli, O., 1885. Erster Band. Protozoa. In: Dr. H.G. Bronn's Klassen und Ordnungen des Thier-Reichs, wis- senschaftlich dargestellt in Wort und Bild. Winter, Leipzig, pp. 865-1088.
Clarke, R.F.A. and Verdier, J.-P., 1967. An investigation of microplankton assemblages from the chalk of the Isle of Wight, England. Verh. K. Ned. Akad. Wet. Afd. Natuurk. Eerste Reeks, 24(3): 1-96.
Cookson, 1.C., 1956. Additional microplankton from Australian late Mesozoic and Tertiary sediments. Aust. J. Mar. Freshwater Res., 7(1): 183-191.
Cookson, I.C. and Eisenack, A, 1958. Microplankton from Australian and New Guinea Upper Mesozoic sediments. Proc. R. Soc. Victoria, 70(1): 19 79.
Cookson, I.C. and Eisenack, A., 1970. Cretaceous microplank- ton from the Eucla Basin, Western Australia. Proc. R. Soc. Victoria, 83: 137-157.
Davey, R.J., 1970. Non calcareous microplankton from the Cenomanian of England, northern France and North America, pt. II. Bull. Br. Mus. Nat. Hist. Geol., 18(8): 333 398.
Deflandre, G., 1935. Consid6rations biologiques sur les microorganismes d'origine planctonique conservds dans le silex de la craie. Bull. Biol. Fr. Belg., 69(2): 213-244.
Deflandre, G., 1936. Les flagell6s fossiles; apperqu biologique et pal6ontologique. R61e g6ologique. Hermann, Paris, Actual. Sci. Ind., 335, 98 pp.
Evitt, W.R., 1985. Sporopollenin dinoflagellate cysts. Their morphology and interpretation. Am. Assoc. Stratigr. Palynol. Found., 333 pp.
Evitt, W.R., Clarke, R.F.A. and Verdier, J.-P., 1967. Dinoflagellate studies. Ill. Dinogymnium acuminatum n. gen.,
n. sp. (Maastrichtian) and other fossils formerly referable to Gymnodinium Stein. Geol. Sci., 10(4): 3-35.
Fensome, R.A., Taylor, F.J.R., Norris, G., Sarjeant, W.A.S., Wharton, D.I. and Williams, G.L., 1993: A classification of living and fossil dinoflagellates. Micropaleontology Spec. Publ., 7: 1-351.
Goodman, D.K., 1987. Dinoflagellate cysts in ancient and modern sediments. In: F.J.R. Taylor (Editor), The Biology of Dinoflagellates. Blackwell, Oxford, Bot. Monogr., 21, pp. 649-722.
Habib, D., 1972. Dinoflagellate stratigraphy Leg 11, Deep Sea Drilling Project. Init. Rep, DSDP, 11: 367-425.
Harker, S.D. and Sarjeant, W.A.S., 1975. The stratigraphic distribution of organic-walled dinoflagellate cysts in the Cretaceous and Tertiary. Rev. Palaeobot. Palynol., 20: 217-315.
Jain, K.P. and Millepied, P., 1975. Cretaceous microplankton from Senegal Basin, W. Africa, Pt. II, Systematics and biostratigraphy. Geophytology, 5(2): 126 171.
Jain, K.P., Sah, S.C.D. and Singh, R.Y., 1975. Fossil dinoflagellates across Maestrichtia~Danian boundary in Lower Assam, India. Paleobotanist, 22 ( 1 ): 1-18.
Lentin, J.K. and Vozzhennikova, T.F., 1990. Fossil dinoflagel- lates from the Jurassic, Cretaceous and Paleogene deposits of the USSR: A re-study. Am. Assoc. Stratigr. Palynol. Contrib. Ser., 23: 1-221.
Londeix, L., 1990. La distribution des kystes de Dinoflagell6s dans les s6diments h~mip61agiques (Ard6che) et p61agiques (Arc de Castellane, S.E. de la France) en domaine vocontien, du Valanginien terminal au Barr6mien inf6rieu~ Biostratigraphie et relations avec la Stratigraphie s6quentielle. Thesis. Univ. Bordeaux I, Bordeaux, 478, 324 + 279 pp.
May, F.E., 1976. Dinoflagellate: Fossil motile-stage tests from the Upper Cretaceous of the Northern New Jersey coastal plain. Science, 193: 1128-1130.
Millioud, M.E., Williams, G.L. and Lentin, J.K., 1975. Stratigraphical range charts of selected fossil dinoflagellates: Selected Cretaceous dinoflagellates. Am. Assoc. Stratigr. Palynol. Contrib. Ser., 4:65 71.
Monteil, E., 1985. Les dinokystes du Valanginien du bassin du Sud-Est (Ard6che, France). Thesis. Univ. Paris VI, Paris, 85-46, 311 pp.
Pascher, A., 1914. Uber Flagellaten und Algen. Ber. Dtsch. Bot. Ges., 32(2): 136 160.
Pourtoy, D., 1989. Les kystes de dinoflagell6s du Cr~tac6 inf6rieur de la Veveyse de Chfitel-St-Denis (Suisse): Biostratigraphie et stratigraphie sdquentielle. Thesis. Univ. Bordeaux I, Bordeaux, 2245, 168 +214 pp.
Riley, L.A. and Fenton, J.P.G., 1984. Palynostratigraphy of the Berriasian to Cenomanian sequence at Deep Sea Drilling Project Site 535, Leg 77, Southeastern Gulf of Mexico. Init. Rep. DSDP, 77: 675-690.
Sarjeant, W.A.S. and Downie, C., 1974. The classification of dinoflagellate cysts above generic level; a discussion and revision. Birbal Sahni Inst. Palaeobot. Spec. Publ., 3:19 32.
Smith, A.G. and Briden, J.C., 1977. Mesozoic and Cenozoic

382 L. Londeix et al./Review of Palaeobotany and Palynology 92 (1996) 367-382
Paleocontinental Maps. Cambridge Univ. Press, Cambridge, 63 pp.
Soncini, M.-J. and Rauscher, R., 1988. Associations de dinokystes du Maastrichtian-Paldocene phosphat6 au Maroc. Bull. Cent. Rech. Explor.-Prod. Elf-Aquitaine, 12(1): 427 450.
Sournia, A., 1986. Atlas du phytoplancton marin. Volume I: introduction, cyanophyc6es, dictyochophyc6es, dinophyc6es et raphidophycbes. CNRS, Paris, 219 pp.
Tappan, H., 1980. The Paleobiology of Plant Protists. Freeman, San Fransisco, CA, 1028 pp.
Tappan, H. and Loeblich, A.R. Jr., 1977. Peridinialean cyst affinity, rather than gymnodinialean motile stage, of the Late Cretaceous dinoflagellate Dinogymnium. Trans. Am. Microsc. Soc., 9(4): 498 505.
Taylor, F.J.R., 1987. Appendix. Taxonomy and classifications. In: F.J.R. Taylor (Editor), The Biology of DinoflageUates. Blackwell, Oxford, Bot. Monogr., 21, pp. 723 731.
Vozzhennikova, T.F., 1967. Iskopaemye peridinei yurskikh, melovykh i paleogenovykh otlozheniy SSSR (Fossil peridini- ans of the Jurassic, Cretaceous and Paleogene deposits of the USSR). Tr. Akad. Nauk SSSR Sib. Otd. Inst. Geol. Geofiz., 347 pp. (in Russian).
Watkins, D.K. and McNulty, C.L., 1984. Paleontological synthesis, Leg 77. lnit. Rep. DSDP, 77: 703-714.
Willey, A. and Hickson, S.J., 1909. The Protozoa (continued). Section F.- The Mastigophora. In: R. Lankester (Editor), A Treatise on Zoology. Part 1. Introduction and Protozoa. First fascicle. Black, London, pp. 154 192 [reprinted by Asher, Amsterdam, 1964].