the ontogeny of nasal floor shape variation in extant humans
TRANSCRIPT

The Ontogeny of Nasal Floor Shape Variation in ExtantHumans
Christina L. Nicholas,1* and Robert G. Franciscus1,2
1Department of Anthropology, University of Iowa, Iowa City, IA 52242-13222Department of Orthodontics, University of Iowa, Iowa City, IA 52242-1322
KEY WORDS internal nasal floor topography; oronasal cavity; palatal growth; piriformaperture entrance; modern humans; Neandertals; fossil Homo
ABSTRACT Variation in nasal floor topography hasgenerated both neontological and paleontological inter-est. Three categories of nasal floor shape (Franciscus: JHum Evol 44 (2003) 699–727) have been used when ana-lyzing this trait in extant humans and fossil Homo: flat,sloped, and depressed (or “bi-level”). Variation in the fre-quency of these configurations within and among extantand fossil humans has been well-documented (Francis-cus: J Hum Evol 44 (2003) 699–727; Wu et al.: AnthropolSci 120 (2012) 217–226). However, variation in this traitin Homo has been observed primarily in adults, withcomparatively small subadult sample sizes and/or largeage gradients that may not sufficiently track key ontoge-netic changes. In this study, we investigate the ontogenyof nasal floor shape in a relatively large cross-sectionalage sample of extant humans (n 5 382) ranging from4.0 months fetal to 21 years post-natal. Results indicate
that no fetal or young infant individuals possess adepressed nasal floor, and that a depressed nasal floor,when present (ca. 21% of the sample), does not occuruntil 3.0 years postnatal. A canonical variates analysisof maxillary shape revealed that individuals withdepressed nasal floors were also characterized by rela-tively taller anterior alveolar regions. This suggests thatpalate remodeling at about 3.0–3.5 years after birth,under the influence of tooth development, strongly influ-ences nasal floor variation, and that various aspects ofdental development, including larger crown/root size,may contribute to the development of a depressed nasalfloor. These results in extant humans may help explainthe high frequency of this trait found in Neandertal andother archaic Homo maxillae. Am J Phys Anthropol000:000–000, 2014. VC 2014 Wiley Periodicals, Inc.
A depressed nasal floor that is positioned significantlybelow the level of the inferior border of the nasal aper-ture was originally noted by T. Dale Stewart in his ini-tial descriptions of the Shanidar 1 and Shanidar 2Neandertal crania (Stewart, 1959:479, 1961). The fea-ture was also described in the Amud 1 Neandertal cra-nium by Suzuki (1970), who pointed out that it was alsopresent in the La Chapelle 1, La Quina 5, and La Fer-rassie 1 Neandertals. Stewart (1977) subsequently sug-gested an explanation for this trait by linking it to thelarge facial size of Neandertals in comparison to subse-quent modern humans whose reduced facial size, heargued, precluded the characteristic Neandertal nasalfloor depression. Since then, this trait has been fre-quently mentioned as a Neandertal autapomorphy(Stringer and Trinkaus, 1981; Stringer, 1983; Trinkaus,1983; Stringer et al., 1984; Franciscus and Trinkaus,1988; Hublin, 1991; Arsuaga et al., 1997; Churchillet al., 1999; Arsuaga et al., 1999). More recently, in anextensive survey, Franciscus (2003) demonstrated thatwhile 80% of Neandertals possess a depressed nasalfloor, this configuration is also found, albeit in low fre-quencies, in a wide range of western Old World extanthuman populations. Thus, while the idea that adepressed nasal floor configuration represented a Nean-dertal autapomorphy was rejected, the question of whyNeandertals show such a high frequency of depressednasal floors still remains unanswered. More recently, Wuet al. (2012) added to the Franciscus (2003) survey byincorporating, in particular, the nasal floor assessmentsof the available and sufficiently intact archaic Homomaxillae from East Asia along with extant comparative
data from this region. The East Asian archaic Homosample while comparatively small, nonetheless exhibiteda prevalence of the depressed nasal floor pattern similarto that seen in the western Eurasian Neandertals andraises additional questions regarding precisely what thehigh frequency of the depressed nasal floor in Neander-tals and other archaic humans signifies. Furthermore,nasal floor depression appears in differing frequencies,albeit low ones, across modern human populations(Franciscus, 2003; Wu et al., 2012), with extant sub-Saharan Africans showing nasal floor depression at sig-nificantly higher frequencies than other geographic
Additional Supporting Information may be found in the onlineversion of this article.
Grant sponsors: National Science Foundation SBR-9312567(R.G.F.), Leakey Foundation (R.G.F.), UI Old Gold Fellowship(R.G.F.), UI Stanley Graduate Award for International Research(C.L.N.), and UI Department of Anthropology Summer Funding(C.L.N.).
*Correspondence to: Christina L. Nicholas, Department ofAnthropology, 114 Macbride Hall, University of Iowa, Iowa City, IA52242-1322, USA. E-mail: [email protected]
Received 10 January 2014; accepted 3 July 2014
DOI: 10.1002/ajpa.22570Published online 00 Month 2014 in Wiley Online Library
(wileyonlinelibrary.com).
� 2014 WILEY PERIODICALS, INC.
AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 00:00–00 (2014)
region that all show comparatively low levels ofdepressed nasal floors (Franciscus, 2003). As a result,the actual significance of nasal floor shape variationremains unresolved at both inter- and intraspecificlevels.
Additionally, previous work (McCollum, 2000) hasinvestigated nasal floor configuration variation specifi-cally among earlier hominins (mainly the australopithe-cines) and has demonstrated that the “robust”australopithecines tend to have a flat nasal floor while“gracile” australopithecines have a higher frequency ofdepressed nasal floors. The McCollum et al. (1993) studyalso coded nasal floor shape in some early Homo speci-mens that originally yielded a mix of flat, sloped, anddepressed configurations, but upon reanalysis McCollum(2000) determined that only three early Homo/AfricanH. erectus specimens could be coded (OH62; AL 666-1;KNM-WT 15000), and all were found to be “continuous-smooth,” or flat. Franciscus (2003) has argued based onhis coding results on the original fossils for Paranthro-pus (OH 5), Early Homo (KNM-ER 1470; KNM-ER 1813;OH 24; OH 62), and early Homo erectus (KNM-ER 3733and KNM-WT 15000), that a level or sloped internalfloor is in fact the primitive condition for Homo.
In addition to such taxonomic considerations, functionalinterpretations for variation in nasal floor shape havealso been proffered. Early hypotheses explaining nasalfloor depression in Neandertals posited that this traitserved to enlarge the internal nasal cavity, as well asincrease turbulent airflow and thereby enhanced warm-ing of inspired air [see review in Franciscus (2003)]. How-ever, based upon the fact that the extant humanpopulations with highest frequencies of depressed nasalfloors are actually warm-adapted sub-Saharan Africans,with Europeans and other living cold-adapted populationsshowing low frequencies of depressed nasal floors, Fran-ciscus (2003) disputed functional hypotheses and positedinstead that a depressed nasal floor may be the result ofstochastically derived populational differences in fetalnasofacial growth patterns. Moreover, the literature onextant human midfacial development has suggested thatmany traits that vary in frequency between populationssuch as nasal margin configuration, anterior nasal spinedevelopment, and piriform aperture shape develop beforebirth, by the second trimester of fetal development(Schultz, 1918, 1920; Mooney and Siegel, 1986a, b;Mooney et al., 1992).
If ontogenetic differences were responsible for differen-ces in the frequency of depressed nasal floors, we might
reasonably expect that these differences also developearly. Franciscus (2003) hypothesized that this trait, likeother midfacial traits, might develop prenatally. Inter-estingly, in a previous study of subadults we foundinstead that no individuals exhibited a depressed config-uration prior to 3.0 years of age (Nicholas and Francis-cus, 2009), the time at which the permanent anteriordentition is beginning to develop in the tooth cryptsabove the deciduous incisors (Demirjian and Goldstein,1976). Our sample for the prior study, however, lackedsufficient numbers of individuals across all age classes,and was populationally limited.
This study was undertaken to address two questions.First, we test whether there is, in fact, a relatively latedevelopmental onset of nasal floor depression in extanthumans (i.e., after 3.0 years of age commensurate withthe well-established onset of permanent anterior denti-tion above the deciduous incisors) with enhanced sam-pling across a broader range of subadult age categoriesand geographic regions. Second, we use Canonical Vari-ates Analysis of maxillary shape to test the hypothesisthat individuals with depressed nasal floors are charac-terized by a particular form of lower midfacial shape(e.g., alveolar dimensions) that differs from individualswith non-depressed nasal floors.
MATERIALS AND METHODS
Samples
Visual scoring of nasal floor configuration was col-lected on 382 individuals ranging in age from 4 monthsprenatal to 21 years post-birth representing a rangeof geographic diversity including Euro-Americans,Europeans, African-Americans, Africans, Asians, and arelatively small sample of aboriginal Australian/Mela-nesians (Table 1). Historical time periods for this aggre-gate sample include the 20th century (Fetal–InfantCollection, Johns Hopkins Collection, Hamann–ToddCollection), the 19th Century (Cambridge, Lisbon), the17th Century (Spitalfields), and the 13th Century(Abingdon). Preservation of material varied by collec-tion, with those from archaeological contexts (Spital-fields, Abingdon, and Lisbon) tending to be morefragmentary. Only individuals for whom the entirenasal floor was well-preserved were included in thestudy. All individuals were grouped into one of eightdevelopmental classes (Table 1) using available docu-mented ages and/or age estimates taken from collectionrecords, with the exception of the material from the
TABLE 1. Sample composition and number of individuals in each age class
FetalBirth to2.9 years
3.0–5.9years
6.0–8.9years
9.0–11.9years
12.0–14.9years
15.0–17.9years
18.0–21.0years
European-deriveda,b,c,d,e,f 42 53 24 12 12 13 10 22African-derivedd,e,f,g 89 16 9 8 6 2 7 17Asian-derivedf,g 1 – 5 7 10 3 1 2Australian/Melanesiang – – 1 6 – – – 4Total 132 69 39 33 28 18 18 45
a Spitalfields Collection, Natural History Museum (London).b Abingdon Collection, Natural History Museum (London).c Luis Lopes Collection, Natural History Museum (Lisbon).d Johns Hopkins Collection, Cleveland Museum of Natural History.e Hamann–Todd Collection, Cleveland Museum of Natural History.f Fetal–Infant Collection, National Museum of Natural History (Smithsonian).g Duckworth Collection, Cambridge University Leverhulme Center.
2 C.L. NICHOLAS AND R.G. FRANCISCUS
American Journal of Physical Anthropology

Cambridge Duckworth Collection (13.1% of the totalsample) for which only broad age ranges were given inthe document records. For these specimens, CLN esti-mated age based on dental eruption standards (Ube-laker, 1989; AlQahtani, 2008). These standards wereaugmented by confirming the presence of still patentspheno-occipital sutures in the highest age category(i.e., 18.0–21.0 years) to guard against the inclusion ofolder adults with third molar impaction or agenesis.Age groupings were structured to produce categorieswhich segregated ages in which the complete range ofnasal floor variation was documented from those inwhich it was not, based on our preliminary study(Nicholas and Franciscus, 2009). Hence, birth to 2.9years is the first post-natal group. To then keep agegroups consistent, we chose 3-year intervals for theremaining age categories. This strategy had the addedbenefit of allowing age divisions that roughly corre-sponded with key stages of dental development: that is,birth to 2.9 and 3–5.9 both representing a period domi-nated by deciduous dentition; 6–8.9 and 9–11.9 charac-terized by mixed dentition; and all subsequentcategories consisting of adult dentition (Ubelaker, 1989;AlQahtani, 2008).
Data collection
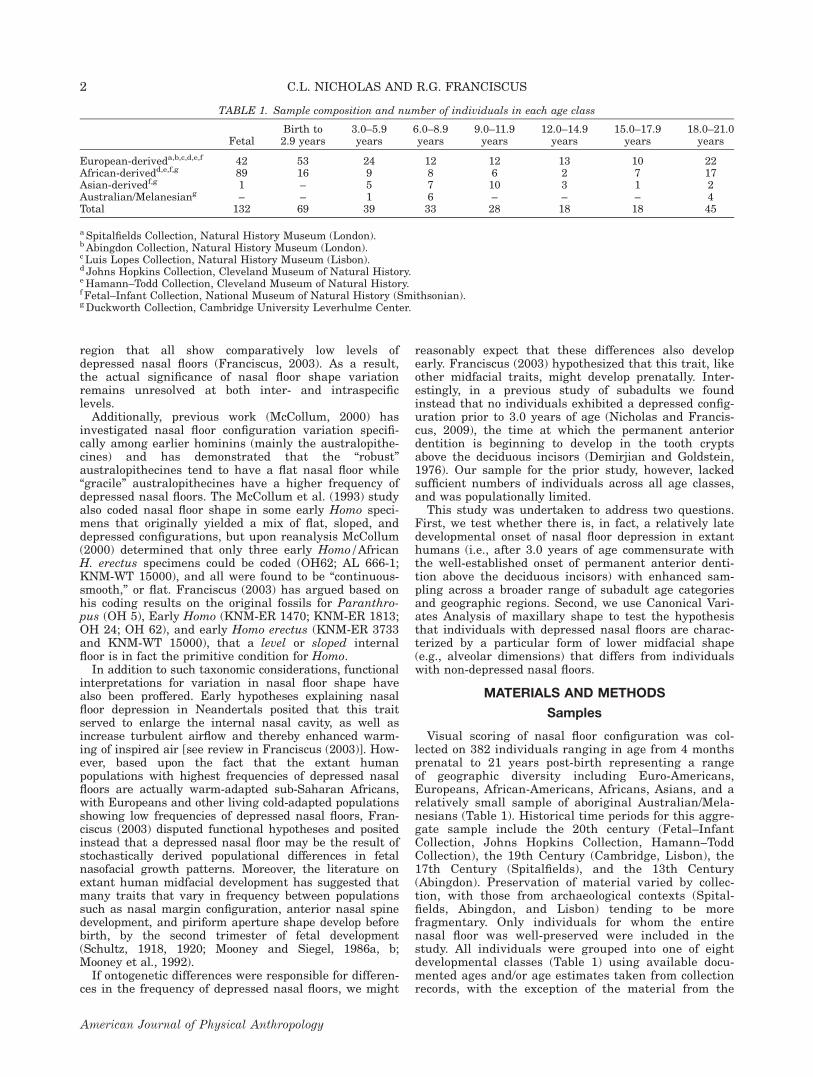
The nasal floor, as considered here, is comprised of themaxilla (and premaxilla) anteriorly and the palatinesposteriorly. Although nasal floor shape variation maywell exist along a continuous gradient, to date it hasonly been visually scored as a two- or three-categorytrait (McCollum, 2000; Franciscus, 2003; Wu et al.,2012), with the three-category system (Fig. 1) havingbeen argued by Franciscus (2003) to best capture therange of variation in Homo. A flat nasal floor is a config-uration where the inferior, interior surface of the nose islevel from the inferior border of the piriform aperture tothe maximum posterior extent of the palatines. A slopednasal floor is when the nasal floor is at a slight gradient,being more raised anteriorly. Lastly a depressed, or “bi-level,” nasal floor is one in which the plane of the nasalfloor exhibits a marked descent just posterior to the piri-form aperture; the area which thus appears raised rela-tive to the rest of the palate is the portion of the maxillawhich exists as the premaxilla during early development(Fig. 1).
Nasal floor shape was scored lateral to the incisivecanal by visual assessment in anterior view into thenasal cavity and observation of its topographic reliefarrayed along the transverse plane (McCollum andWard, 1997; McCollum, 2000; Franciscus, 2003) closelyfollowing the three category system from Franciscus(2003) described above. The preceding studies havefocused either largely, or exclusively, on coding adults.In this study of subadult individuals, in a flat configura-tion (Fig. 1a), the nasal floor lies even with the inferioraspect of the piriform aperture along its entire length(ignoring any localized depression occurring only in theimmediate vicinity of the incisive foramen). In a slopedconfiguration, the nasal floor is level with the inferioraspect of the piriform aperture in the anterior aspect,but gradually slopes inferiorly toward the posterioraspect (Fig. 1b). In a depressed configuration, the planeof the nasal floor is positioned much deeper (inferiorly)than the lower border of the piriform aperture in bothits anterior and posterior aspects (Fig. 1c). The youngest
individuals often had disarticulated cranial bones inwhich an actual lateral (i.e., sagittal) view of the nasalfloor was possible; nonetheless, even for these specimensthe maxillae were coded for nasal floor configuration byviewing them in the same orientation (i.e., the anteriorview) as for intact crania found in older subadult ages tomaintain consistency in visual coding across all agegroups. Nasal floor category frequency was tabulated ina single regionally aggregated sample across sex-combined age groups (Table 2).
Craniofacial coordinate landmark data were collectedusing an Immersion Co. Microscribe. The landmarksincluded in this study (Fig. 2) are standard landmarkswhich capture aspects of maxillary morphologyinvolved in palatal shape, subnasal morphology, andnasal aperture shape. Subnasal and nasal aperturemorphology in particular were especially important tocapture as these are the aspects of the midface thatare highly variable between populations (e.g., anteriornasal spine prominence, subnasal alveolar progna-thism, lateral nasal wall position related to nasalbreadth) in adults (Franciscus, 1995) and subadults(Schultz, 1918, 1920; Mooney and Siegel, 1986a, b,1991; Mooney et al., 1992). To create a homologouslandmark set across all age ranges, only right sidemaxillary landmarks were included in this study. Someindividuals were missing one or more of these land-marks (e.g., prosthion), and this yielded a reducednumber of individuals for inclusion in our shape analy-ses. Landmarks were not collected on the nasal flooritself for two reasons. First, we wanted to make com-parisons to all previous studies which used a visuallycoded rather than digitized metric approach, and sec-ondly, because many of the individuals in the oldersubadult age categories in this study were representedby complete, intact crania, and thus the internal nasalfloor was not sufficiently exposed for accurate andrepeatable landmarking using the Microscribe. How-ever, landmarks along the palatal midline, that is, theoral roof (Fig. 2), were recorded to augment the othermidfacial points of comparison.
Geometric morphometrics and statisticalanalyses
Generalized procrustes analysis (GPA) was conductedon the landmark data. This analysis translates, rotates,and scales data such that size is removed while shapeinformation is retained (Zelditch et al., 2012). Toaccount for the potential effects of allometry in oursample, we performed a regression of shape on centroidsize. We found evidence for allometry, and so chose toconduct all further analyses on the regression residuals(although results were very similar when run on thenon-residual data). Relative warps analysis (RWA), atype of factor analysis (Zelditch et al., 2012), was runon the regression residuals to determine what aspectsof shape made the greatest contribution to variationwithin the sample. To examine variation across sub-groups within our sample, canonical variates analysis(CVA) was used. A CVA can be used to highlight inter-group variation, especially when intragroup variabilityis high (as in this sample, where individuals from dif-ferent ancestral populations are included in each nasalfloor grouping) (Albrecht, 1980). Analyses of shape vari-ation (e.g., relative warps analysis, canonical variatesanalysis) were conducted on the landmark data using
ONTOGENY OF NASAL FLOOR SHAPE 3
American Journal of Physical Anthropology
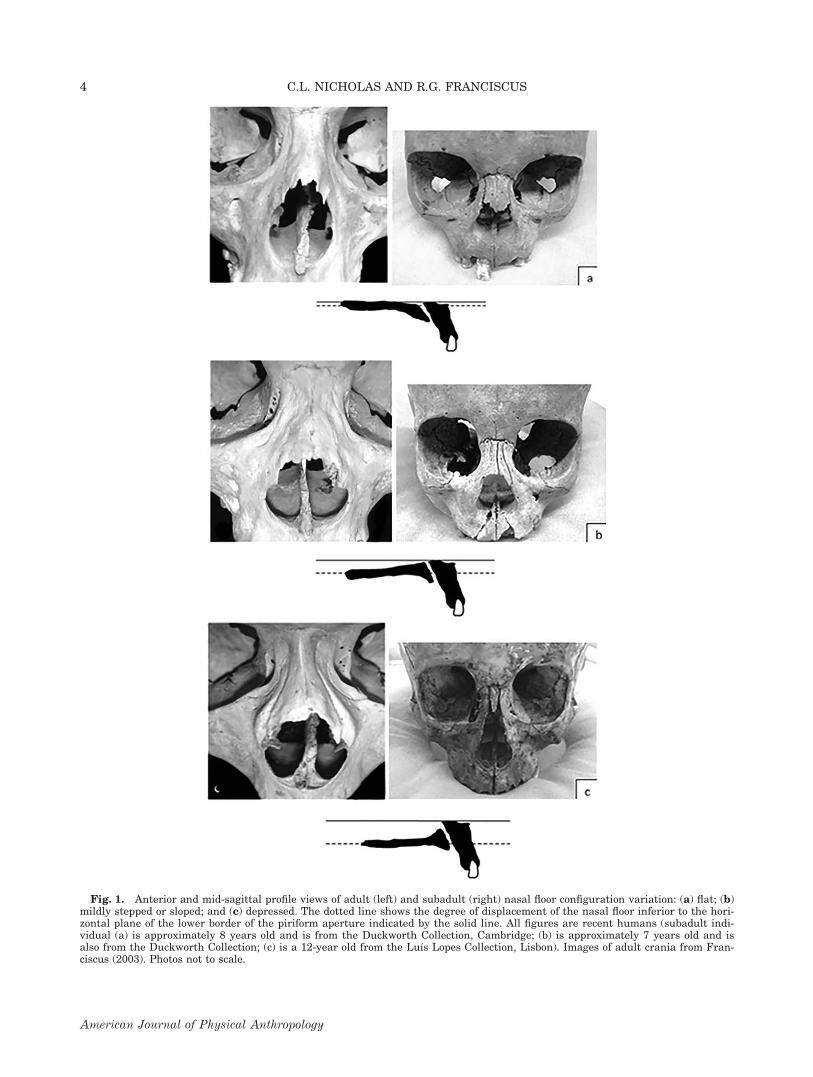
Fig. 1. Anterior and mid-sagittal profile views of adult (left) and subadult (right) nasal floor configuration variation: (a) flat; (b)mildly stepped or sloped; and (c) depressed. The dotted line shows the degree of displacement of the nasal floor inferior to the hori-zontal plane of the lower border of the piriform aperture indicated by the solid line. All figures are recent humans (subadult indi-vidual (a) is approximately 8 years old and is from the Duckworth Collection, Cambridge; (b) is approximately 7 years old and isalso from the Duckworth Collection; (c) is a 12-year old from the Lu�ıs Lopes Collection, Lisbon). Images of adult crania from Fran-ciscus (2003). Photos not to scale.
4 C.L. NICHOLAS AND R.G. FRANCISCUS
American Journal of Physical Anthropology
MorphoJ (Klingenberg, 2011) and the statistical soft-ware R (http://cran.r-project.org/).
To examine the relationship between nasal floor shapeand age, a Cross-tab Chi-Square analysis was run. Aftertesting for equal variances and normality in our sam-ples, we used an analysis of variance (ANOVA) to testwhether any of our relative warps (RWs) were signifi-
cantly correlated with our visual coding of nasal floortopography.
RESULTS
Nasal floor configuration frequencies across agegroups
As predicted by the observations in our earlier pilotstudy, a depressed nasal floor does not develop in utero,and in fact is entirely absent until 3.0–5.9 years of age(Table 2). The youngest individuals to show a depressedconfiguration are 3.0 years old. Although a depressednasal floor is completely absent prior to 3.0 years of age,it occurs in varying frequencies in all the subsequentolder age categories (Table 2; see Supporting Informa-tion Table S1 for a raw count of nasal floor score by age/ancestry groupings). When we combine the frequenciesof these depressed nasal floor occurrences in all ageswhere they appear (ages 3–21; n 5 181, Table 3), 20.9%
TABLE 2. Nasal floor configuration frequencies by age groupa
FetalBirth to 2.9
years3.0–5.9years
6.0–8.9years
9.0–11.9years
12.0–14.9years
15.0–17.9years
18.0–21.0years
Flat 2 15 7 11 11 7 6 24Sloped 130 54 21 16 12 6 8 14Depressed – – 11 6 5 5 4 7Total n 132 69 39 33 28 18 18 45% Depressed 0% 0% 28.21% 18.18% 17.86% 27.78% 22.2% 15.56%
a Frequencies for age groups are combined values across all geographic samples.
Fig. 2. Subadult maxillae illustrating the 12 coordinate landmarks included in this study.
TABLE 3. Nasal floor frequencies in combined pre- and post-3.0year age classesa
Age Flat Sloped DepressedTotal
n % Depressed
Fetal to2.9 years
17 184 0 201 0%
3–21 years 65 78 38 181 20.99%
a The two combined-age categories used here are based on thecontrast in presence/absence of nasal floor depression develop-ment before and after 3.0 years of age; see text for elaboration.
ONTOGENY OF NASAL FLOOR SHAPE 5
American Journal of Physical Anthropology

possess the depressed nasal floor configuration. This is ahigher frequency than found by Franciscus (2003) in anentirely adult, western Old World extant sample (11.7%depressed nasal floors out of n 5 522). Moreover, the fre-quency of nasal floor depression did not vary much fromone age bracket to the next starting with the 3–5.9 yearcategory onwards (ranging between 15.56 and 28.21%).
Cross-tab Chi-square analysis between these two com-bined age groups (fetal to 2.9 years and 3–21 years) andnasal floor configuration (Table 4) revealed a highly sig-nificant difference in the frequency of nasal floor config-uration (Chi-square 5 109.14; P 5 0.000001). Thelargest contribution to the total Chi-square results isfrom the complete absence of depressed nasal floors inthe fetal to 2.9-year group, and the greater thanexpected occurrence of this configuration in 3–21-yearage group. We consider this result to be paramount overthe differing frequencies in flat versus sloped betweenthe two combined age groups because the scale of differ-ence in these two latter categories is more nuanced bothin terms of coding in subadults compared to adults, andin terms of inferred developmental alterations (see Dis-cussion section for details).
Shape analysis of midfacial landmark data andnasal floor configuration
Because the first two age category samples (fetal andbirth to 2.9 years) were devoid of individuals withdepressed nasal floors, a GPA was conducted on the coor-dinate landmark data using only individuals aged 3.0 orolder (n 5 72; Tables 5 and 6). It is only at this age thatthe first appearance of nasal floor depression occurs, andmoreover, all three configurations appear in some fre-quency in each of the post-3.0-year age groups (Table 2).A RWA of the regression residuals showed that the firstsix RWs captured >5% of the total variation each
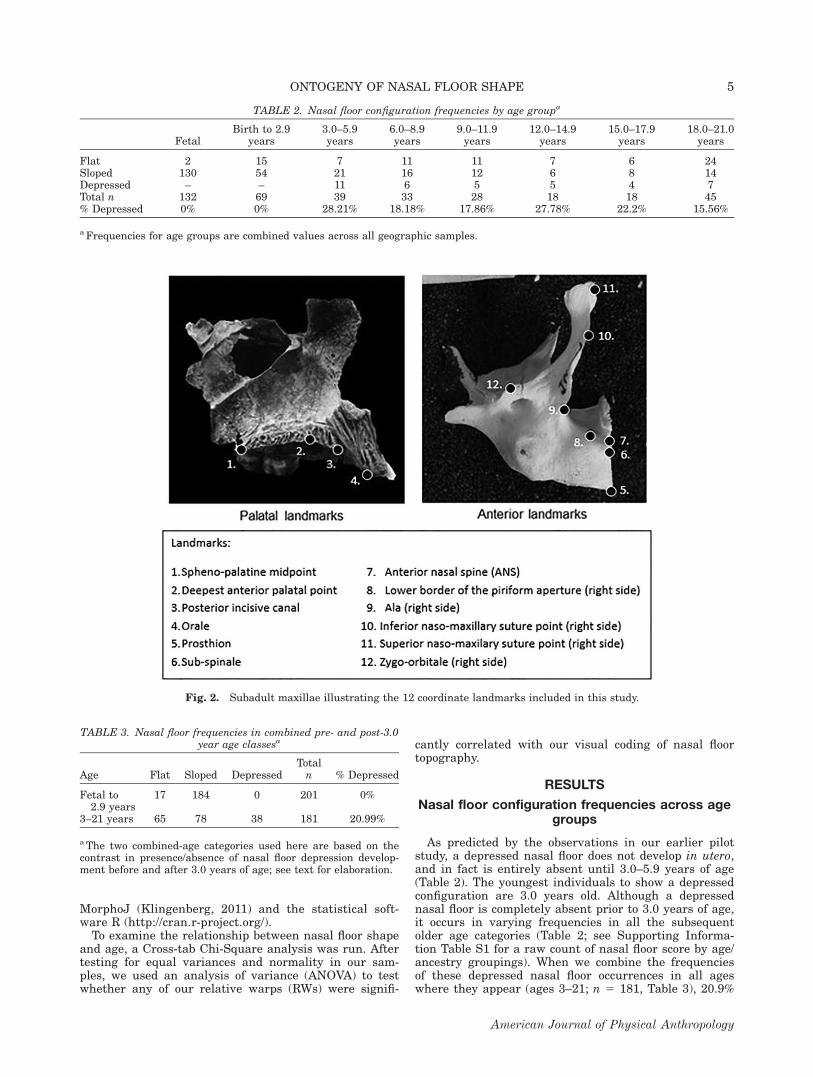
(72.56% total) and thus were selected for further analy-sis following the recommendations of Zelditch et al.(2012). An ANOVA was conducted on the RW scores ofmaxillary shape using nasal floor configuration as thegrouping variable. RW1 (Fig. 3) represents 18.28% of thetotal variation and is associated with nasal floor configu-ration differentiation (P 5 0.0071), capturing, amongother aspects of lower midfacial shape differences, varia-tion in the extent of superior-inferior height expansionin the alveolar region (Fig. 4). RW6, which represents7.55% of the total variation, also showed a significantdifference among the groups (P 5 0.0166), depicting avery similar pattern of variation to RW1, but with nochange in A-P breadth and added variation in the orien-tation of the anterior nasal spine (Fig. 5).
As neither RW1 nor RW6 showed a particularly clearpattern of relationship between nasal floor configurationand maxillary shape, with a considerable degree of over-lap between the three categories along both RWs, a CVAwas conducted on the Procrustes residuals from theGPA. The CVA results provide further informationregarding overall maxillary morphology and its associa-tion with the different nasal floor configurations, andallowed explicit examination of the intergroup variabili-ty relationship between nasal floor configuration andmaxillary shape. CV1 shows variation in maxillaryshape related to nasal floor shape, with a flat configura-tion at one extreme and a depressed configuration at theother (Fig. 6). When the variation along CV1 is exam-ined, we see that a depressed nasal floor is generallyassociated with a taller alveolar region (the same pat-tern found in the RW6 analysis noted earlier) and anantero-posteriorly narrower inferior border of the alveo-lus (Fig. 7). In contrast, a flat nasal floor is typified by ashorter and A-P broader alveolus and a more inferiorlyangled posterior palate. Lastly, a sloped nasal floor isgenerally characterized by morphology intermediate tothat seen in individuals with flat or depressed nasalfloors. Mahalanobis distances indicate that a flat nasalfloor shape is more similar in morphology to a slopedone (1.710 vs. 2.6831; P < 0.0001), and a sloped nasalfloor is more similar to a flat one than a depressed one(2.585 vs. 2.3786; P 5 0.0019). These same patterns areseen in the Procrustes distances, however, in this analy-sis only the variation between flat and depressedreaches significance (P 5 0.0020). Interestingly, a slopedconfiguration groups distinctly along CV2. This appearsto be related to variation in degree of palatal curvatureand anterior nasal spine prominence, where sloped nasalfloor individuals show somewhat smaller anterior nasalspines and somewhat less steeply superiorly curvedpalates.
DISCUSSION AND CONCLUSIONS
This study confirms that a depressed nasal floor doesnot appear before about 3.0 years of age in extanthumans. The fact that variation in nasal floor shape
TABLE 4. Cross tabs Chi-square results of nasal floor configu-ration frequency in two age classesa
Flat Sloped Depressed Totals
Fetal to 2.9 yearsRaw counts 17 184 0 201Percentages 8.5% 91.5% 0% 100%Standard residuals 24.02 3.97 24.47 0Chi-squared contribution 16.2 15.73 19.94 51.86
3–21 yearsRaw counts 66 78 38 182Percentages 36.3% 42.9% 20.9% 100%Standard residuals 4.23 24.17 4.69 0Chi-squared contribution 17.9 17.37 22.02 57.27
Total Chi-square 5 109.139; degrees of freedom 5 2; Prob 5
0.0000001.a The two combined-age categories used here are based on thecontrast in presence/absence in depressed nasal floor develop-ment before and after 3.0 years of age; see text for elaboration.
TABLE 5. Age composition of landmark data subsample
3.0–5.9years
6.0–8.9years
9.0–11.9years
12.0–14.9years
15.0–17.9years
18.0–21.0years Total
Flat – 7 5 4 3 15 34Sloped 6 2 5 3 2 5 23Depressed 3 2 1 2 3 4 15Total n 9 11 11 9 8 24 72
6 C.L. NICHOLAS AND R.G. FRANCISCUS
American Journal of Physical Anthropology
develops later than variation in many other key nasaltraits that manifest significant populational differencesby the second trimester of fetal development such as
nasal margin configuration, anterior nasal spine devel-opment, and piriform aperture shape (Schultz, 1918,1920; Mooney and Siegel, 1986a, b, 1991; Mooney et al.,1992) seems significant, and may indicate that nasalfloor shape is responding to different developmentalmechanisms and/or selective pressures. Our results sug-gest that the period of growth at this stage may be a keydevelopmental window for a depressed nasal floor amongindividuals that evince this trait as adults as the fre-quency of its occurrence remains more or less unchangedacross the later subadult age periods. We do not know,however, whether nasal floor configuration is stablethrough development post-3 years of age; futureresearch using a longitudinal sample will be needed totest this. This point in development (ca. 3.0 years) iswhen deciduous incisor root lengths have reached, or arereaching, their full extent and permanent anterior denti-tion development has begun (Demirjian and Goldstein,1976; Ubelaker, 1989). It is possible that a combinationof nasal floor (palate) drift and anterior palate remodel-ing (the latter under the influence of tooth developmentand tooth crypt expansion in a limited available space;see Fig. 8) is tied to nasal floor variation. Therefore, therate of tooth growth or length of roots may contribute tothe development of a depressed nasal floor.
In addition to the absence of nasal floor depression infetal and infant individuals, we note that there is a com-paratively small degree of visible nasal floor morphologi-cal variation between individuals in this pre-3.0-year oldage group, and it can reasonably be asked whether theflat and sloped categories could actually be collapsedinto a single category. In young subadults, differentiat-ing between flat and sloped morphologies is more diffi-cult than in adults. First, there is the fact that a smallmaxilla (and hence a small nasal floor) makes visualobservations between what might be gently sloped andwhat might be considered flat more difficult. Also, themain difference between flat and sloped nasal floors isoften one of orientation, and proper orientation in theFrankfurt Horizontal in isolated young subadult maxil-lae is more difficult than in older subadults and adults
TABLE 6. Ancestry composition of landmark data subsample
European-derived
African-derived
Asian-derived
NativeAustralian/Melanesian Total
Flat 12 5 11 6 34Sloped 6 11 5 1 23Depressed 7 6 2 – 15Total n 25 22 18 7 72
Fig. 3. Plot of Relative Warp 1 and RW6. Nasal floor configu-ration scores are indicated by dot coloration (red 5 flat; green 5
sloped; blue 5 depressed). The ellipses represent the 90% confi-dence intervals around the means for each nasal floor configura-tion. Although depressed and flat nasal floors overlap on bothRWs, they have statistically significantly different mean values.
Fig. 4. Lateral view wireframes showing the range of varia-tion along RW1. Relative Warp 1 depicts variation in alveolarheight, A-P alveolar breadth at the inferior border of the alveo-lus, and palatal curvature: (a) shows the negative extreme ofvariation, represented by a tall, narrow alveolar region with amore superiorly oriented posterior palate; (b) shows the positiveextreme of variation, represented by a shorter, broader alveolarregion with a more inferiorly oriented posterior palate, all indi-cated by the black lines. The light gray lines illustrate themean shapes (see Fig. 3 for landmark information).
Fig. 5. Lateral view wireframes showing the range of varia-tion along Relative Warp 6. This RW illustrates variation inalveolar height and orientation of the anterior nasal spine: (a)shows the negative extreme of variation, with a tall alveolarheight and a more superiorly oriented nasal spine; (b) showsthe positive extreme of variation, with a relatively short alveo-lar height and an inferiorly angled nasal spine (see Fig. 3 forlandmark information).
ONTOGENY OF NASAL FLOOR SHAPE 7
American Journal of Physical Anthropology

with complete crania. A combination of these factorsmay be responsible for the elevated percentage of speci-mens showing a sloped nasal floor relative to a flat nasalfloor in our fetal-2.9 year combined categories (Table 3).
Furthermore, a potential explanation for why fetaland infant individuals might show a consistent configu-ration that is actually more of a continuum betweenflat or sloped can be found in earlier research on nasalfloor variation across hominoids. McCollum et al. (1993)investigated nasal floor configuration in fossil homi-nins, comparing morphology between extant apes(gorillas, chimpanzees, and orangutans), early hominins(A. afarensis, A. africanus, P. robustus, and P. boisei),Homo (“early Homo,” H. erectus) and extant H. sapiensand they concluded that the presence of a depressednasal floor, when present, is the result of the location ofthe attachment of the vomer to the nasal floor. Anattachment on the hard palate surface (and not the“pre-maxilla”) would be more likely to result in adepressed nasal floor. In modern humans, as the vomerattaches more anteriorly (McCollum et al., 1993), thesetwo units (hard palate and pre-maxilla) are essentiallyconstrained to function as one unit. Thus, they developthe same flat morphology (McCollum et al., 1993). Thisflat/sloped configuration would thus be the typical formof nasal floor shape in extant modern humans, barringfurther developmental pressures causing a relative supe-rior elevation of the anterior portion of the nasal flooramong the smaller number of adult individuals that showa depressed nasal configuration (Franciscus, 2003; Wuet al., 2012). Ultimately, there may, in fact, be only oneextant human fetal/infant (pre �3.0 years post-natal)nasal floor configuration, but a more refined quantifica-tion of nasal floor shape using geometric morphometricswould be necessary to fully resolve this issue.
The results of our Relative Warps analysis indicatethat the majority of maxillary shape variation in thesample is not related to nasal floor configuration. Therelative warps that were associated with nasal floorshape, RW1 (Fig. 4) and RW6 (Fig. 5), showed a verysimilar pattern of shape variation as that seen arrayedlong the first axis of our CVA (Fig. 7), although CV1 also
Fig. 6. Bivariate plot of the Canonical Variates scores,shown with 90% confidence ellipses around individuals, con-ducted on maxillary Procrustes shape residuals using nasalfloor score as the grouping variable (red 5 flat; green 5 sloped;blue 5 depressed). Flat and depressed configurations show sep-aration along CV1, with CV2 separating out individuals scoredas sloped from both of the two other groups.
Fig. 7. Lateral view wireframes showing the range of varia-tion along Canonical Variate 1 (see Fig. 5) illustrating variationin maxillary shape that arrays along a spectrum from flat todepressed. (a) Represents the negative extreme of variation flatnasal floors, (b) the positive extreme. The black lines representdeviations from the overall mean (the gray lines). This CV cap-tures variation in alveolar height, A-P alveolar breadth, andsubnasal alveolar prognathism: (a) shows a relatively short,inferiorly broad alveolar region and a moderate degree of sub-nasal alveolar prognathism and tends to be associated with aflat nasal floor configuration; (b) depicts a relatively tall, narrowalveolar region that is orthognathic and tends to be associatedwith a depressed nasal floor configuration.
Fig. 8. Anterior view of 4.0-year-old subadult midface. Post-mortem damage to the anterior maxilla illustrates how toothdevelopment and location (in this case the right central perma-nent incisor crown) occurs immediately inferior to the wall ofthe piriform aperture (specimen from the Abingdon Collection,Natural History Museum, London).
8 C.L. NICHOLAS AND R.G. FRANCISCUS
American Journal of Physical Anthropology

depicts variation in subnasal alveolar prognathism. Wetherefore argue that although the relationship betweennasal floor shape and overall maxillary shape is notstrongly deterministic, there are nonetheless perceptiblepatterns of maxillary shape variation that are related tonasal floor morphology. Alveolar height and A-P breadthappear to be the key difference depicted along RW1 andCV1 (Figs. 4 and 7), suggesting that this aspect of mor-phology has a developmental relationship to nasal floorshape. Specifically, nasal floor depression is associatedwith a relatively taller and narrower anterior alveolarregion. We posit that, in accordance with ideas put forthby Wu et al. (2012) and McCollum (2000), a more verti-cally expanded premaxilla may be the ultimate cause ofnasal floor depression when present in Homo.
Recently, Wu et al. (2012) hypothesized that variationin adult nasal floor morphology could be related to dif-ferences in premaxillary and maxillary growth patterns.The premaxillary suture divides the alveolar portion ofthe maxilla (the premaxilla) from the majority of thepalate in early development, and studies have demon-strated that patency of this suture varies between mod-ern human populations (Mooney and Siegel, 1986a, b)and between Neandertals and extant humans (Maureilleand Bar, 1999). The premaxilla houses the maxillaryanterior dentition, and makes up the anterior portion ofthe nasal floor; this is also the region of the nasal floorwhich appears raised relative to the rest of the palate inindividuals with a depressed nasal floor. Our resultslend support to the hypothesis (McCollum, 2000; Wuet al., 2012) that differential growth patterns of the pre-maxilla are responsible for nasal floor depression. Addi-tionally, we hypothesize that these differences in growthare caused by variation in anterior dental size and/ortiming of the development of the permanent maxillaryincisors (Nicholas and Franciscus, 2010). Vertical expan-sion of the adult anterior dentition, and thereby the pre-maxillary region, would thus be the cause for this laterdevelopmental change in nasal floor configuration in cer-tain individuals.
There are several possible scenarios for how premaxil-lary expansion and elevation relative to the palate mightoccur. The literature on maxillary growth indicates thatduring facial growth (which, broadly speaking, is inferiorand anterior in orientation), the nasal floor and nasalclivus are resorptive, while the palate is depositional(Kurihara et al., 1980; McCollum, 2008). Additionally,recent research by Martinez-Maza et al. (2013) indicatesthat more localized resorptive versus depositional pat-terns may show inter-individual variation. We thereforesuggest that it is less likely that the premaxillary regionis expanding superiorly, and more likely that the palateis descending inferiorly at a greater rate than the pre-maxilla in individuals who develop nasal floor depres-sion. Broadly speaking, nasal floor depression is likely tobe the result of the need to maintain palatal shape formasticatory dynamics as well as speech functions, andnasal aperture shape for respiration, while also allowingenough space for the developing dentition. We envisiontwo scenarios which might lead to variation in howmuch space in the premaxilla different individualsrequire to accommodate the developing dentition. First,incisor root length is known to vary across populations(Bailey, 2005; Walker et al., 2008), and larger incisorroots may necessitate a premaxillary region that haslarger vertical dimensions. Second, timing of tooth erup-tion (precocious vs. delayed) relative to overall facial size
may play a role, in that individuals with precocioustooth development relative to their facial size may needa relatively expanded premaxillary region. However, it isclear that more work needs to be done in this area tobetter understand the mechanisms which lead to varia-tion in nasal floor topography.
Based on our results for extant humans, it seemslikely that a depressed nasal floor in archaic Homowould also not develop until about 3.0 years after birth.It is interesting to note that the youngest Neandertalspecimens with published nasal floor codings (Roc deMarsal 1 and Subalyuk 2) both have depressed nasalfloors (Franciscus, 2003), and both are estimated to beapproximately 3.0 years of age. Given that this is theage at which we see nasal floor depression develop inextant humans, it seems most parsimonious to infer thatnasal floor development in Neandertals follows a similarontogenetic pattern. The precise details of nasal floorconfiguration in those Neandertal specimens that areunambiguously younger than 3.0 years of age (e.g., LeMoustier 2, Mezmaiskaya, and Dederiyeh 1) have notyet been published, but such data would go a long waytowards either confirming or rejecting this hypothesis.
The results of this study also have broader implica-tions extending to the clinical sciences. Many pediatricmaxillo-facial surgeries rely upon current assumptionsabout maxillary ontogeny and expected spatial relation-ships between traits. Nasal septoplasty, for example, isoften performed at young ages, sometimes as early as afew weeks post-natal (Gray, 1965, 1967; Emami et al.,1996). This procedure surgically severs the juncturebetween the vomer/nasal septal cartilage and the nasalfloor, and then realigns the structure to the right or leftto fix a deviated septum (Gray, 1967). Severely deviatednasal septa can lead to many adverse effects in infantssuch as respiratory distress, snoring, and increased inci-dence of respiratory infections (Blahova, 1985; Emamiet al., 1996). Also, for many children nasal septal devia-tion worsens through development. For these reasons,many clinicians advocate early surgical intervention(Emami et al., 1996). Nasal septoplasty assumes thatthe spatial relationship between the nasal floor and thevomer are developmentally stable. The effects of nasalseptoplasty on midfacial growth are still not whollyunderstood (Verwoerd and Verwoerd-Verhoef, 2010).Understanding what ontogenetic shape changes thenasal floor and other aspects of the mid-face undergoduring development is important to deciding at what ageit is most efficacious to perform such procedures. Thefact that certain individuals will undergo spatialchanges to this portion of their maxilla subsequent toearly post-natal procedures may affect surgical out-comes, and thus the conventional idea that the patternsof change between the nasal floor and vomer are invari-antly predictable, should be taken under advisement.
ACKNOWLEDGMENTS
The authors are grateful to all the individuals andinstitutions who granted access to the subadult materi-als used in this study: Robert Kruszynski (Natural His-tory Museum, London); Hugo Cardoso (Natural HistoryMuseum, Lisbon); Mercedes Okumura (Cambridge Uni-versity Leverhulme Center); Lyman Jellema (ClevelandMuseum of Natural History); and David Hunt (NationalMuseum of Natural History, Smithsonian). Steve Millerand Sarah Trabert provided helpful comments on the
ONTOGENY OF NASAL FLOOR SHAPE 9
American Journal of Physical Anthropology

initial manuscript, and the authors are grateful to theAE and two anonymous reviewers for their many helpfulsuggestions.
LITERATURE CITED
Albrecht GH. 1980. Multivariate analysis and the study of formwith special reference to canonical variate analysis. Am Zool20:679–693.
AlQahtani SJ. 2008. Atlas of tooth development and eruption.Barts and the London School of Medicine and Dentistry. Lon-don, Queen Mary University of London: MClinDent.
Arsuaga JL, Martinez I, Gracia A, Lorenzo C. 1997. The Simade los Huesos crania (Sierra de Atapuerca, Spain). A compar-ative study. J Hum Evol 33:219–281.
Arsuaga JL, Mart�ınez I, Lorenzo C, Gracia A, Mu~noz A, AlonsoO, Gallego J. 1999. The human cranial remains from GranDolina Lower Pleistocene site (Sierra de Atapuerca, Spain).J Hum Evol 37:431–457.
Bailey SE. 2005. Diagnostic dental differences between Nean-dertals and Upper Paleolithic modern humans: getting to theroot of the matter. In: Zadzinska E, editor. Current trends indental morphology research. Lodz (Poland): University ofLodz Press. p 201–210.
Blahova O. 1985. Late results of nasal septum injury in chil-dren. Int J Pediatr Otorhinolatyngol 10:137–141.
Churchill SE, Shackleford LL, Georgi JN, Black MT. 1999. Air-flow dynamics in the Neandertal nose. J Hum Evol 36:A5.
Demirjian A, Goldstein H. 1976. New systems for dental matu-rity based on seven and four teeth. Ann Hum Biol 3:411–421.
Emami AJ, Brodsky L, Pizzuto M. 1996. Nasal septoplasty: acase report and review of the literature. Int J Pediatr Otorhi-nolaryngol 35:271–275.
Franciscus RG. 1995. Later pleistocene nasofacial variation inWestern Eurasia and Africa and modern human origins. PhDThesis, University of New Mexico.
Franciscus RG. 2003. Internal nasal floor configuration inHomo with special reference to the evolution of Neandertalfacial form. J Hum Evol 44:699–727.
Franciscus RG, Trinkaus E. 1988. Nasal morphology and theemergence of Homo erectus. Am J Phys Anthropol 75:517–527.
Gray LP. 1965. The deviated nasal septum. II. Prevention andtreatment. J Laryngol 79:806–816.
Gray LP. 1967. The deviated nasal septum. III. Its influence onthe physiology and disease of the nose and ear. J Laryngol81:953–986.
Hublin J-J. 1991. L’e�mergence des Homo sapiens archaiques:Afrique de Nord-Ouest et Europe occidentale. The�se d’Etat,Universite�de Bordeaux 1.
Klingenberg CP. 2011. MorphoJ: an integrated software packagefor geometric morphometrics. Mol Ecol Resour 11:353–357.
Kurihara S, Enlow DH, Rangel RD. 1980. Remodelling rever-sals in anterior parts of the human mandible and maxilla.Angle Orthod 5:98–106.
Martinez-Maza C, Rosas A, Nieto-Diaz M. 2013. Postnatalchanges in the growth dynamics of the human face revealedfrom bone modelling patterns. J Anat 223:228–241.
Maureille B, Bar D. 1999. The premaxilla in Neandertal andearly modern children: ontogeny and morphology. J HumEvol 37:137–152.
McCollum MA. 2000. Subnasal morphological variation in fossilhominins: a reassessment based on new observations andrecent developmental findings. Am J Phys Anthropol 112:275–283.
McCollum MA. 2008. Nasomaxillary remodeling and facial form inrobust Australopithecus: a reassessment. J Hum Evol 54:2–14.
McCollum MA, Grine FE, Ward SC, Kimbel WH. 1993. Sub-nasal morphological variation in extant hominoids and fossilhominids. J Hum Evol 24:87–111.
McCollum MA, Ward SC. 1997. Subnasoalveolar anatomy andhominoid phylogeny: evidence from comparative ontogeny.Am J Phys Anthropol 102:377–405.
Mooney MP, Siegel MI. 1986a. Racial variation in anterior nasalspine (ANS) formation: testing the traction hypothesis. Am JPhys Anthropol 69:242.
Mooney MP, Siegel MI. 1986b. Developmental relationshipbetween premaxillary-maxillary suture patency and anteriornasal spine morphology. Cleft Palate J 23:101–107.
Mooney MP, Siegel MI. 1991. Premaxillary-maxillary suturefusion and anterior nasal tubercle morphology in the chim-panzee. Am J Phys Anthropol 85:451–456.
Mooney MP, Siegel MI, Kimes KR, Todhunter J, Janosky J.1992. Multivariate analysis of second trimester midfacialmorphology in normal and cleft lip and palate human fetalspecimens. Am J Phys Anthropol 88:203–209.
Nicholas CL, Franciscus RG. 2009. Internal nasal floor configu-ration in contemporary fetal and postnatal subadult H. sapi-ens. Am J Phys Anthropol Suppl 139:199.
Nicholas CL, Franciscus RG. 2010. The ontogeny of variation ininternal nasal floor configuration in extant H. sapiens. Am JPhys Anthropol Suppl 141:177.
Schultz AH. 1918. Relation of the external nose to the bonynose and nasal cartilages in whites and negroes. Am J PhysAnthropol 1:329–341.
Schultz AH. 1920. The development of the external nose inWhites and Negroes. Contrib Embryol 34:175–190.
Stewart TD. 1959. The restored Shanidar I skull. Ann. Rep.Smithsonian Inst. for 1958, 479.
Stewart TD. 1961. The skull of Shanidar II. Ann. Rep. Smithso-nian Inst., 521–533.
Stewart TD. 1977. The Neanderthal skeletal remains from Sha-nidar Cave, Iraq: a summary of findings to date. Proc AmPhil Soc 121:121–165.
Stringer CB. 1983. Some further notes on the morphology anddating of the Petralona hominid. J Hum Evol 12:731–742.
Stringer CB, Hublin J-J, Vandermeersch B. 1984. The origins ofanatomically modern humans in Western Europe.In: SmithFH, Spencer F, editors. The origins of modern humans: aworld survey of the fossil evidence. New York: Alan R. Liss. p51–135.
Stringer CB, Trinkaus E. 1981. The Shanidar Neandertal cra-nia.In: Stringer CB, editor. Aspects of human evolution. Lon-don: Taylor & Francis. p 129–165.
Suzuki H. 1970. Skull of the Amud man.In: Suzuki H, Takai F,editors. The Amud man and his cave site. Tokyo: AcademicPress. p 123–206.
Trinkaus E. 1983. The Comprehensive R Archive Network.Institute for Statistics and Mathematics, UW Wein, n.d. Web.The Shanidar Neandertals. New York: Academic Press. 12Nov, 2011. <http://cran.r-project.org/>.
Ubelaker DH. 1989. Human skeletal remains. Excavation, anal-ysis, interpretation, 2nd ed. Washington, DC: Taraxacum.
Verwoerd CD, Verwoerd-Verhoef HL. 2010. Rhinosurgery inChildren: developmental and surgical aspects of the growingnose. GMS Curr Top Otorhinolaryngol Head Neck Surg Suppl89:S46–71.
Walker MJ, Gibert J, L�opez MV, Lombardi AV, P�erez-P�erez A,Zapata J, Ortega J. 2005. Late Neandertals in SoutheasternIberia: Sima de las Palomas del Cabezo Gordo, Murcia, Spain.Proc Natl Acad Sci USA 52:20631–20636.
Wu X-J, Maddux SD, Pan L, Trinkaus E. 2012. Nasal floor vari-ation among eastern Eurasian Pleistocene Homo. AnthropolSci 120:217–226.
Zelditch ML, Swiderski DL, Sheets HD. 2012. Geometric mor-phometrics for biologists: a primer, 2nd ed. London: ElsevierScience.
10 C.L. NICHOLAS AND R.G. FRANCISCUS
American Journal of Physical Anthropology