the antibody response during amphibian ontogeny
TRANSCRIPT

Immunogenetics 3 : 381 --391, 1976 �9 1976 by Springer-Verlag New York Inc.
The Antibody Response During Amphibian Ontogeny
L. Du Pasquier 1 and J. Haimovich 2
1 Basel Institute for Immunology, Grenzacherstrafle 487, Postfach, 4005 Basel 5, Switzerland 2 The Weizmann Institute of Science, Rehovot, Israel
Received December 4, 1975; Revised version received January 15, 1976
Abstract. The maturation of the anti DNP-response in adult and larval amphibians has been compared in two species of anurans, Rana catesbeiana and Xenopus laevis. IgM responses were very similar in larvae and adults of both species. Specific IgG antibody synthesis, studied in Rana catesbeiana only, was not obvious in larval primary response but was easily detectable in adults. Although metamorphosis results in profound changes in the lymphoid system associated with the apparent acquisition of self tolerance to new antigens, immunological memory of antigens injected during larval life of Rana and Yenopus persists after the metamorphosis period.
Introduction
During the ontogeny of the amphibian lymphoid system two events may influence antibody diversity of the individual. First, as the immunocompetent larva develops into an adult, the number of its lymphocytes increases about a hundredfold; this increase may affect the heterogeneity of the antibody popula- tions detectable after immunization with one antigen. Second, profound lym- phoid changes occur in amphibians during metamorphosis. In Xenopus, for example, the thymus and spleen lose 90% and 50% of their lymphocytes, respec- tively. Two months later this depletion is followed by a new lymphopoiesis (Du Pasquier and Weiss 1973). These changes coincide with a period of immuno- depression. During this period it is easy to induce tolerance to weak histocompa- tibility antigens (Chardonnens and Du Pasquier 1973, Du Pasquier and Char- donnens 1975), an observation that may relate to the generation of self-tolerance to newly appearing adult specific antigens. It is conceivable, therefore, that the perimetamorphic period would influence antibody diversity and immunologi- cal memory.
Recent studies have shown that young bullfrog larvae could make anti- dinitrophenyl (DNP) antibodies that were apparently as good as mammalian ones (Haimovich and Du Pasquier 1973), and that the larval response was characterized by an increase in the affinity of IgM antibody during the three to four weeks following a single injection of antigen (Du Pasquier and Haimovich 1974). Such a maturation is usually interpreted as reflecting the existence of a heterogeneous population of lymphocytes producing anti-DNP antibodies of various affinities (Siskind and Benacerraf 1969). Therefore, a comparison of the maturation of immune responses in larvae and adults can be a simple

382 L. Du Pasquier and J. Haimovich
way of estimating the change in antibody heterogeneity caused by lymphocyte proliferation occurring during ontogeny.
The present study concerns the maturation (i.e., increase in binding affinity) of antibody response to DNP in larval and adult forms of two anuran species, the bullfrog (Rana catesbeiana) and the more primitive clawed toad (Xenopus laevis). In addition, the effect of metamorphosis on the immunological memory of this antigen was also investigated.
Materials and Methods
Animals. Larval and adult Rana catesbeiana were obtained commercially (Mogul-Ed., Oshkosh, Wisconsin). Adults were kept in a vivarium and fed living crickets and small frogs; larvae were maintained in aquaria and fed nettle powder. Adult Xenopus laevis, purchased from the South African Snake Farm (Fish Hoek, Cape Town), were bred in our laboratory (Elkan 1957). All animals were maintained at 20+ I~
Methods. Order of magnitude estimates of the total number of leucocytes in individual animals were determined by multiplying the total number of counted thymus and spleen cells by a factor of five to account for cells in other lymphoid organs and in blood 1.
Animals were injected intraperitoneally (i.p.) with various doses of 2,4-dinitrophenyl keyhole limpet hemocyanin (DNP-KLH) emulsified in Freund's complete adjuvant. The total volume injected was 10-20 Ixl.
Inactivation of DNP-bacteriophage Tr (DNP-T~) by peritoneal fluids (Rana, larval Xenopus) or serum (adult Xenopus) and inhibition of the inactivation by DNP-, 2,4,6-trinitrophenyl (TNP)- lysine and 4-mononitrophenyl (MNP)-lysine were performed in Terasaki plates according to the microtechnique already described (Haimovich and Du Pasquier 1973). The same preparation of DNP-T4, prepared according to the method of Segal and coworkers (1970) was used in all experi- ments.
To follow the maturation of anti-DNP response, animals were injected i.p. with 10 ~tg DNP-KLH per gram of body weight. Peritoneal fluids were withdrawn weekly during a two-month period. The antibody activity was titrated by determining the inactivation of the DNP-T4; the relative affinities of the antibody were estimated by determining the inhibition of inactivation of the modified bacteriophage with various concentrations of homologous and crossreacting ligands. (For discussion of the advantages and limits of the method see Du Pasquier and Haimovich 1974.)
Antibodies of the 19S and 7S classes were separated by sucrose gradient centrifugation. Peri- toneal fluids or sera were centrifuged in 10 33% linear gradients for 5 h at 55,000 rpm in a SW56 rotor (Beckman). Antibodies present in the 19S and 7S fractions were characterized by immunoelec- trophoresis as IgM- and IgGqike, respectively. Specific anti-Xenopus ~t and 7 antisera were originally used to characterize the precipitation lines. In Rana IgM- and IgG-like molecules have different electrophoretic mobilities and can be identified easily.
Radioimmunoelectrophoresis was performed as described by Rosenquist and Hoffmann (1972) using 131I-labelled ct-N-(3 5 diodo-4 hydroxyphenacetyl)-e-DNP-L-lysine as the ligand.
Results
Maturation of Antibody Response in Larval and Adult Rana catesbeiana. Maturation studies were conducted on 14 tadpoles weighing 2 to 3 g (stages 26~8, Witschi 1956) and on 6 adults (10 to 20 g) 3 months after they had
1 The factor of five was suggested by cell counts on control animals where blood lymphocytes numbers were estimated and where histological preparations of gut, liver, and kidneys were made to determine their lymphocytes content.
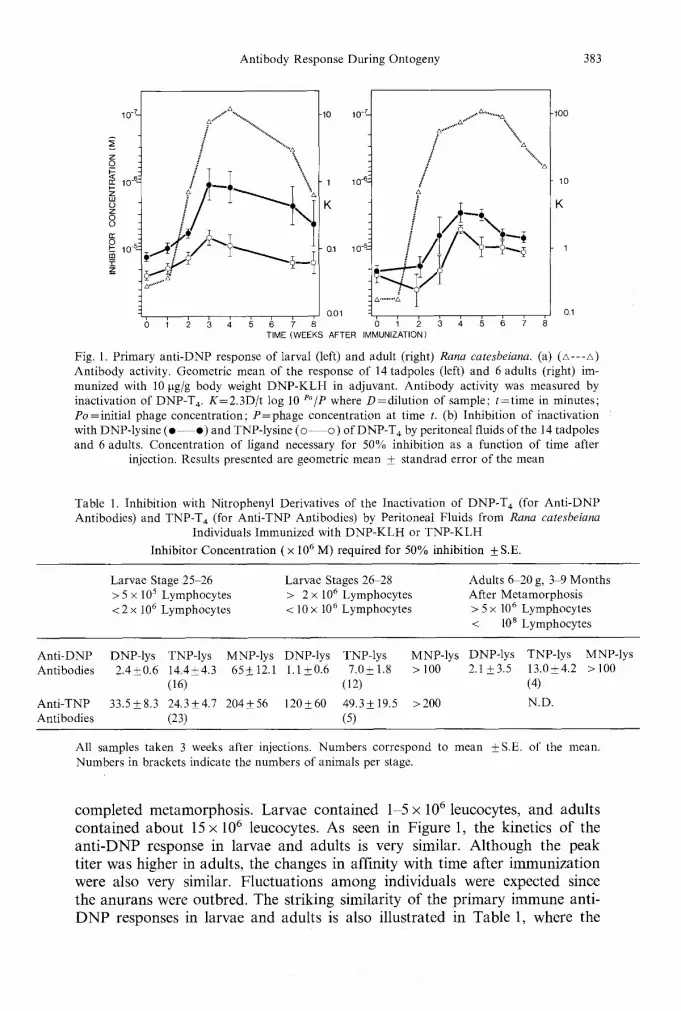
Antibody Response During Ontogeny 383
10-7.-
o_ z
10 -6-
, 0 _ 5 - co
........ 10 10- A.�9 �9149 %,%�9
.~ "%'A / x
2 I \ \ ,o-~4
A �9 .." ...... , A
.A" % A ........ ~�9
/ "z ,- X
/ 1 /
A
, i i i i , i i r
0 , 2 3 4 5 6 7
0 , , 0
o o 0 1 2 3 4 5 6 7
-100
1 0
K
0,1
TIME (WEEKS AFTER IMMUNIZATION)
Fig. I. Primary anti-DNP response of larval (left) and adult (right) Rana catesbeiana. (a) (A---A) Antibody activity. Geometric mean of the response of 14 tadpoles (left) and 6 adults (right) im- munized with 10 gg/g body weight DNP-KLH in adjuvant. Antibody activity was measured by inactivation of DNP-T 4. K=2.3D/t log 10 eo/p where D=dilution of sample; t=time in minutes; Po=initial phage concentration; P=phage concentration at time t. (b) Inhibition of inactivation with DNP-lysine ( e - - o ) and TNP-lysine ( � 9 of DNP-T 4 by peritoneal fluids of the 14 tadpoles and 6 adults. Concentration of ligand necessary for 50% inhibition as a function of time after
injection. Results presented are geometric mean _+ standrad error of the mean
Table 1. Inhibition with Nitrophenyl Derivatives of the Inactivation of DNP-T 4 (for Anti-DNP Antibodies) and TNP-T 4 (for Anti-TNP Antibodies) by Peritoneal Fluids from Rana catesbeiana
Individuals Immunized with DNP-KLH or TNP-KLH
Inhibitor Concentration (x 106 M) required for 50% inhibition _+S.E.
Larvae Stage 25-26 > 5 x l0 s Lymphocytes <2 x 106 Lymphocytes
Larvae Stages 26 28 > 2 x 106 Lymphocytes < 10 x 106 Lymphocytes
Adults 6 20 g, 3-9 Months After Metamorphosis > 5 x 106 Lymphocytes < 108 Lymphocytes
Anti-DNP Antibodies
Anti-TNP Antibodies
DNP-lys TNP-lys MNP-lys DNP-lys TNP-lys MNP-lys DNP-lys TNP-lys MNP-lys 2.4_+0.6 14.4_+4.3 65_+12.1 1.1_+0.6 7.0_+1.8 >100 2.1_+3.5 13.0+4.2 >100
(16) (12) (4)
33.5_+8.3 24.3+4.7 204_+56 120_+60 49.3_+19.5 >200 N.D. (23) (5)
All samples taken 3 weeks after injections. Numbers correspond to mean _+S.E. of the mean. Numbers in brackets indicate the numbers of animals per stage.
completed metamorphosis. Larvae contained 1-5 x 106 leucocytes, and adults contained about 15 x 10 6 leucocytes. As seen in Figure 1, the kinetics of the anti-DNP response in larvae and adults is very similar. Although the peak titer was higher in adults, the changes in affinity with time after immunization were also very similar�9 Fluctuations among individuals were expected since the anurans were outbred. The striking similarity of the primary immune anti- DNP responses in larvae and adults is also illustrated in Table 1, where the

384 L. Du Pasquier and J. Haimovich
affinities recorded 3 to 1 weeks after immunization in this study are presented with results from previous work on smaller tadpoles (Haimovich and Du Pas- quier 1973).
Estimates of antibody activity in unfractionated peritoneal fluids are based only on IgM antibody activity (Du Pasquier and Haimovich 1974). To determine whether the humoral responses of larvae and adults might be different with respect to the titers and the affinities of IgG antibodies, peritoneal fluids were fractionated into IgM- and IgG-like fractions by sucrose gradient centrifugation. In peritoneal fluids from 8 of 10 tadpoles, virtually all (90-99%) of the antibody activity was localized in the 19S (IgM) fraction. For the remaining 2 animals, about 30% of the antibody activity in the peritoneal fluids was due to IgG. This poor IgG synthesis during the primary response of tadpoles is well known (Gecsy et al. 1973, Du Pasquier and Haimovich 1974) and has been further documented by radioimmunoelectrophoresis (Fig. 2).
The relative activity of IgM and IgG antibody in adult and larval Rana was different. At the peak of the primary response in each of the 6 adults studied, 30 to 50% of the antibody activity was in the IgG fraction (see also Fig. 2). When the IgG and IgM anti-DNP fractions were subjected to inhibition tests, another difference between larvae and adults became evident. Eight of 10 tadpoles (those showing 1 to 10% of the activity in the IgG fraction) had IgG of lower affinity than IgM. The two other tadpoles had IgG and IgM of similar affinities for the ligand. In all adults, the affinity of IgG for DNP was equal to that of IgM. In no instance, however, was affinity of frog IgG similar to that found in mammals (Haimovich et al. 1969). At best, frog IgG required 2 to 3 times less concentrated ligand than IgM for 50% inhibition. The inhibition curves (homologous hapten only) of IgG and IgM anti-DNP antibodies of an adult and larval frog as well as of a goat (control) are presented in Figure 3. It is clear that, whereas the affinities of the goat and larval or adult frog IgM anti-DNP are similar, the affinity of the goat IgG is much higher than that of the amphibian IgG.
In summary, the maturation of the primary antibody responses (titer and affinity) of larval and adult Rana catesbeiana is similar for IgM but dissimilar for IgG. In the adult, specific anti-DNP IgG antibodies are not only present in higher titers but have a greater affinity than larval antibodies. Their affinity however, is still much lower than that of mammalian IgG.
The Effect of Metamorphosis on Immunological Memory of Rana catesbeiana and the More Primitive Xenopus.
1. Rana catesbeiana. Six of the tadpoles, whose primary response to DNP- KLH was described in the previous section, underwent metamorphosis. Three months later, their peritoneal fluids were sampled for baseline antibody activity determination. The animals then received a secondary intraperitoneal challenge with 0.1 gg DNP-KLH per g of body weight. Six adult unimmunized controls received the same dose of DNP-KLH as primary immunization. The immune response of control and experimental animals was followed for 8 months. The results presented in Figure 4 may be summarized as follows.
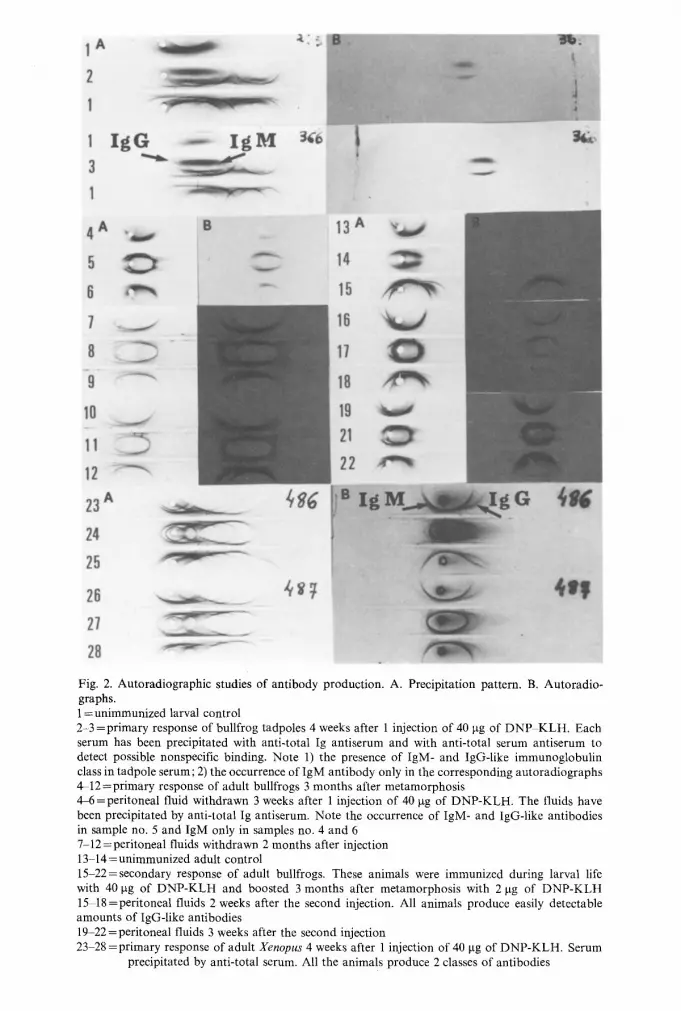
Fig. 2. Autoradiographic studies of antibody production. A. Precipitation pattern. B. Autoradio- graphs. 1 = unimmunized larval control 2-3 =primary response of bullfrog tadpoles 4 weeks after 1 injection of 40 lag of DNP KLH. Each serum has been precipitated with anti-total Ig antiserum and with anti-total serum antiserum to detect possible nonspecific binding. Note 1) the presence of IgM- and IgG-like immunoglobulin class in tadpole serum; 2) the occurrence of IgM antibody only in the corresponding autoradiographs 4-12=primary response of adult bullfrogs 3 months after metamorphosis 4-6 =peritoneal fluid withdrawn 3 weeks after 1 injection of 40 lag of DNP-KLH. The fluids have been precipitated by anti-total Ig antiserum. Note the occurrence of IgM- and IgG-like antibodies in sample no. 5 and IgM only in samples no. 4 and 6 7-12=peritoneal fluids withdrawn 2 months after injection 13 14=unimmunized adult control 15-22=secondary response of adult bullfrogs. These animals were immunized during larval life with 40 lag of DNP-KLH and boosted 3 months after metamorphosis with 2 lag of DNP-KLH 15 18 =peritoneal fluids 2 weeks after the second injection. All animals produce easily detectable amounts of IgG-like antibodies 19-22=peritoneal fluids 3 weeks after the second injection 23 28 =primary response of adult Xenopus 4 weeks after 1 injection of 40 lag of DNP-KLH. Serum
precipitated by anti-total serum. All the animals produce 2 classes of antibodies

386 L. Du Pasquier and J. Haimovich
100"
8o o /o _~ 60 -1- z o~ 40
o / Ill ~ m
ld -9 ld -a ld -z 10 o ld-S INHIBITOR CONCENTRATION (M)
Fig. 3. Inhibition with DNP-lysine of the inactivation of DNP-T~ by 1. Adult Rana catesbeiana IgM ( i - - A ) 1:450 2. Adult Rana catesbeiana IgG-like ( o - - I ) 1:120 3. Larval Rana catesbeiana IgM ( z ~ - - a ) 1 : 180 4. Larval Rana catesbeiana IgG-like (Ez- -m) 1:9 5. Goat IgM ( O - - o ) 1 : 105 6. Goat IgG ( o - - o ) 1 : 10 s The dilutions correspond to the dilutions of unfractionnated serum giving 90% phage inactivation
9 " . , - . , : % . Aee.~'"*~ A ,~ LX "*"A .......... ,,
1 O- J l l ~ l " ............... n
i/ i \ :~'--"-----'-" l O-~4 ~ ~1.o-o.ol \ _ _ o . . . . . ~ o ~
~o ~ .........
o o
10 -~
"10
4 6 8 10 1'2 1'4 1'6 1'8 2'0 2'2 2'4 26 2'8 3'0 TIME (WEEKS AFTER IMMUNIZATION)
Fig. 4. Top." kinetics of anti-DNP primary response in adult Rana catesbeiana following 1 injection of 0.1 gg/g DNP-KLH in adjuvant. Same symbols as for Figure 1 Bottom: kinetics of anti-DNP response in adult Rana catesbeiana which were primed as tadpoles. Adults were boosted with 0.1 p,g/g DNP-KLH in adjuvant. The 6 adults used in this experiment
were members of the group presented in Figure 1. Same symbols as those for Figure 1
0.1
/%
i ~..,,zx,..~ .z '% : ",,, ,,~" % : �9 : "W "n..,:S" : ~- \ ] ". %& ....... , .....
�9 : l . . i o
,(i,J ~~ o
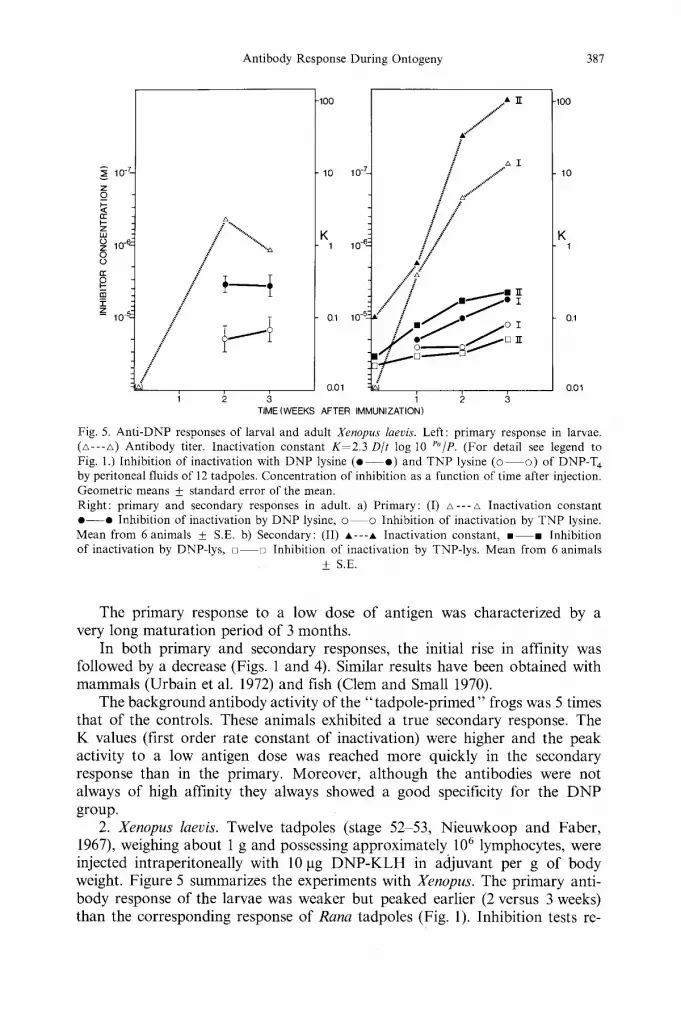
Antibody Response During Ontogeny 387
10_7 -
E
~ 10 -~ 0 o n."
"l-
Z 10.,
A.
/
/ /
/
-100 .,,,,,,,,,,,,,,,,,,,,, A Ir
~..., /
- io io-7 / ............." A""
- 1 10 -4
x / / z
/ / - ol lOq,," ../ �9
~ � 9 I
/ 0.01
-I00
10
K I
0.1
0.01
TIME(WEEKS AFTER IMMUNIZATION)
Fig. 5. Ant i -DNP responses of larval and adult Xenopus laevis. Left: primary response in larvae. (A---A) Antibody titer. Inactivation constant K - 2 . 3 D/t log 10 P~ (For detail see legend to Fig. 1.) Inhibition of inactivation with D N P lysine ( e - - o ) and TNP lysine (o �9 of DNP-T 4 by peritoneal fluids of 12 tadpoles. Concentration of inhibition as a function of time after injection. Geometric means _+ standard error of the mean. Right: primary and secondary responses in adult, a) Primary: (I) z~---A Inactivation constant o - - e Inhibition of inactivation by D N P lysine, �9 o inhibition of inactivation by TNP lysine. Mean from 6 animals + S.E. b) Secondary: (II) A---A Inactivation constant, �9 �9 Inhibition of inactivation by DNP-Iys, [ ] - - [ ] Inhibition of inactivation by TNP-lys. Mean from 6 animals
_+ S.E.
The primary response to a low dose of antigen was characterized by a very long maturation period of 3 months.
In both primary and secondary responses, the initial rise in affinity was followed by a decrease (Figs. 1 and 4). Similar results have been obtained with mammals (Urbain et al. 1972) and fish (Clem and Small 1970).
The background antibody activity of the "tadpole-primed" frogs was 5 times that of the controls. These animals exhibited a true secondary response. The K values (first order rate constant of inactivation) were higher and the peak activity to a low antigen dose was reached more quickly in the secondary response than in the primary. Moreover, although the antibodies were not always of high affinity they always showed a good specificity for the DNP group.
2. Xenopus laevis. Twelve tadpoles (stage 52 53, Nieuwkoop and Faber, 1967), weighing about 1 g and possessing approximately 106 lymphocytes, were injected intraperitoneally with 10 gg DNP-KLH in adjuvant per g of body weight. Figure 5 summarizes the experiments with Xenopus. The primary anti- body response of the larvae was weaker but peaked earlier (2 versus 3 weeks) than the corresponding response of Rana tadpoles (Fig. 1). Inhibition tests re-

388 L. Du Pasquier and J. Haimovich
vealed that the affinity of anti-DNP antibodies in Xenopus and Rana tadpoles were of the same order of magnitude. The antibody titers one week after injection were still too low to permit relative affinities measurements. Thus, precise maturation of the response could not be followed in all specimens of Xenopus. However, the mean values reported in Figure 5 mask the fact that 3 of the 12 tadpoles used in this experiment clearly showed a progressive increase in the affinity of their anti-DNP antibodies with time after immunization. Unlike that of larval forms, the general pattern of the anti-DNP response in adult Xenopus and Rana (Figs. 1, 4, and 5) was very similar.
Nine months after the first injection (6 months after metamorphosis), 6 of the "tadpole-primed" frogs were boosted with 20 ~tg of DNP-KLH emulsified in Freund's complete adjuvant. These animals responded in a typical secondary manner (Fig. 5), since K values were 10 times higher than in the primary re- sponse. Antibodies were specific for the DNP group, as reflected by the con- centration of ligand necessary for their inhibition in the assay of the inactivation of DNP-T 4. In control primary responses, 1-week sera did not contain antibodies with clearcut anti-DNP specificity, i.e., the affinity of the antibodies was low and the crossreactivity with the TNP group was high.
Discussion
Our data on anti-DNP antibody production during anuran ontogeny indicate that regardless of the state of their lymphoid systems, young larvae and adults Show similar maturation of the immune response following one injection of antigen. Such a maturation is currently thought to reflect the heterogeneity of the lymphocyte population at the level of specific receptors. Thus, in amphi- bians the increase in the total number of lymphocytes with age does not result in an increase in lymphocyte heterogeneity. The antibody repertoire dose not seem to differ markedly in larvae and adults, meaning that the generation of antibody diversity has taken place earlier in ontogeny. This was already suggested by previous studies using sheep red blood cells as antigen (Du Pasquier 1970), and by comparison between responses in larval amphibians and mammals (Haimovich and Du Pasquier 1973). These data complement those obtained by detecting antigen-binding cells to various antigens in fetal mice (Spear et al. 1973) and in mice, rabbits, Chicken, and sheep (Decker and Sercarz 1975); both suggest that diversification of the antibody repertoire takes place very early in ontogeny. In tadpoles the antibody response apparently matures more rapidly in Xenopus laevis than in Rana catesbeiana. It may be suggested that in Xenopus the larval response reflects less heterogeneity than in Rana cates- beiana, where selection between several potential anti-DNP antibody-forming clones would take more time. This suggestion must be related to the phylogeneti- cally primitive characteristics of the species Xenopus as compared to Rana.
While larval and adult amphibian IgM appears to be potentially very similar for the IgM primary response, such is not the case for the IgG response. In response to one injection of DNP-KLH, Rana catesbeiana tadpoles synthesize very little IgG antibody. Moreover, these antibodies are usually of low affinity.

Antibody Response During Ontogeny 389
In adults, however, IgG antibodies can easily be detected in the primary response. However, their affinity for the DNP group is much lower than that of mammals (Fig. 3). In this context it is worth mentionning that neither Rana catesbeiana IgG nor Xenopus laevis IgG is a true homologue to mammalian IgG, since the amphibian "7" heavy chain is heavier than its mammalian counterpart (Hadji-Azimi 1971, Marchalonis and AtweiI 1973). Their functional properties may also differ from their mammalian equivalent.
It is not known why the expression of IgG antibodies is easily detected in adults but not in larvae. Normal tadpole serum contains IgG (Gecsy et al. 1973, Du Pasquier 1973), and, given the diversity of the IgM repertoire, there is a piori no reason why tadpoles should not produce as good IgG antibodies as those produced by adults. The differences between larval and adult immune responses may depend on some characteristics of the immune system other than the presence of structural genes of the antibody molecules. Such differences exist at the level of the thymus. For instance, the expression of Ig molecules on thymus cells is stronger in larvae than in adults (Du Pasquier and Weiss 1973). More primitive vertebrates-fish, for example-which do not synthesize IgG exhibit apparently equal amounts of Ig on thymus and spleen cells (Emmrich et al. 1975). It is tempting therefore to consider the larval lymphoid system of the frog as being more primitive than that of the adult for some aspects of the T function. Phylogenetically and ontogenetically, it seems that the more detectable thymocyte Ig there is, the less IgG is produced. Therefore, the lack of mature T cells could be one of the reasons for the difference between larval and adult IgG responses.
In mammals this hypothesis has been ruled out (Goidl and Siskind 1975); the immune capacities of fetal and neonatal B cell precursors transferred in adult mice in the presence of an excess of T cells were compared to those of adult spleen or bone marrow cells. The response of adult cells, as measured essentially by indirect plaques (IgG), was more heterogeneous than that of fetal or neonatal cells. However, the cells which were transferred came from an organism submitted to maternofetal interactions, and some environmental repressing factor could have remained on the surface of the lymphocytes. Moreo- ver, the proportion of potential B cell precursors in each inoculum was probably not known, and taking into account the homing efficiency of the system, one could argue that fewer competent precursors were actually transferred from fetuses than from adults. This could explain the lower degree of heterogeneity recorded with fetal cells. For tadpoles living in water, free from maternofetal interaction, we favor the possibility that the difference in IgG antibody synthesis reflects a lack of maturity of some aspects of the T cell function. Since anuran amphibian larvae can reject grafts acutely and show mixed leucocyte reaction with thymus cells (Du Pasquier and Weiss 1973), this hypothesis implies that the various thymus functions mature at different times: the helper function would appear later than the mixed leucocyte reaction capacity.
The transmission of immunological memory to an antigen was already known in anuran amphibians for transplantation antigens (Volpe 1970). Its occurrence for an antigen like DNP-KLH suggests that the lack of IgG response in larvae was due to a difference in regulation processes. Indeed, rapid and active IgG

390 L. Du Pasquier and J. Haimovich
synthesis observed in boosted adults which were primed during larval life is better explained if one admits that IgG anti-DNP clones were present before the secondary stimulation. Under our experimental conditions, they necessarily came from the larvae, although their expression in larvae was not obvious. The persistence of immunological memory after metamorphosis must also be related to the immunodepression state which exists during the metamorphosis of anurans. This depression may affect antibody synthesis (Moticka et al., 1973) or graft rejection (Bernardini el al. 1969, Chardonnens and Du Pasquier 1973). It coincides with changes off the lymphoid system: larval organs disappear and are replaced by adult specific ones in Rana catesbeiana (Baculi and Cooper 1968). In Xenopus the thymus loses 80 to 90% of its lymphocytes, and the spleen 30 to 40% (Du Pasquier and Weiss 1973). If, according to the simplest assumption, the lymphocyte depletion or suppression occurring at metamor- phosis is due to nonspecific hormonal factors, it is likely that only large clones of cells will see a few of their cells go through metamorphosis. This applies to anti-DNP clones which expand during larval stimulation. On the other hand, small clones, not yet expanded, could either disappear or become highly sensitive to tolerance induction because of the reduction of their size. Such a change in the balance of lymphocyte population could be one of the mechanisms permit- ting the establishment of tolerance to allografts at metamorphosis and the gener- ation of self tolerance to the adult specific antigens appearing during this period.
Acknowledgments. We thank Chantal Guiet for her excellent technical assistance and Dr. N. Cohen for reviewing the manuscript.
References
Baculi, B.S. and Cooper, E.L.: Lymphomyeloid organs of Amphibia. IV. Normal histology in larval and adult Rana catesbeiana. J. Morphol. 126:467476, 1968
Bernardini, N., Chardonnens, X., and Simon, D. : D~veloppement apr& la m6tamorphose de comp6- tences immunologiques envers les homogreffes cutan~es chez Xenopus laevis Daudin. C.R. Acad. Sci. (Paris) 269:1011 1014, 1969
Chardonnens, X. and Du Pasquier, L. : Induction of skin allograft tolerance during metamorphosis of the toad Xenopus laevis: a possible model of studying the generation of self tolerance to histocompatibility antigens. Eur. J. immunol. 3:569-573, 1973
Clem, L.W. and Small, P.A.: Phylogeny of immunoglobulin structure and function. V. Valences and association constants of teleost antibodies to a haptenic determinant. J. Exp. Med. 138:385 400, 1970
Decker, J. and Sercarz, E.: Antigen binding cells and the generation of diversity. Amer. Zool. 15:189 197, 1975
Du Pasquier, L. : Ontogeny of the immune response in animals having less than one million lympho- cytes: the larvae of the toad Alytes obstetricans. Immunology 19: 353~62, 1970
Du Pasquier, L. : Ontogeny of the immune response in cold-blooded vertebrates. Curr. Top. MTcro- biol. lmmunol. 61: 37-88; 1973
Du Pasquier, L. and Chardonnens, X.: Genetic aspect of the tolerance to allografts induced at metamorphosis in the toad Xenopus laevis. Immunogenetics 2:431 440, 1975
Du Pasquier, L. and Haimovich, J.: Changes in affinity of IgM antibodies in amphibian larvae. Eur. J. Immunol. 4:580 583, 1974
Du Pasquier, L., and Weiss, L.: The thymus during the ontogeny of the toad Xenopus laevis: growth, membrane bound immunoglobulins and mixed lymphocyte reaction. Eur. J. Immunol. 3:773-777, 1973

Antibody Response During Ontogeny 391
Elkan, E.: Amphibians. In: A.N. Worden and W. Lane-Petter (eds.): UFAW Handbook q] the Care and Managment of Laboratory Animals' (2rid Ed.), pp. 804~815. Livingstone, Edinburgh, 1957
Emmrich, F., Richter, R.F., and Ambrosius, H.: Immunoglobulin determinants on the surface of lymphoid cells of carps. Eur. J. Immunol. 5:76 78, 1975
Gecsy, C.L., Green, P.C., and Steiner, L.A.: Immunoglobulins in the developing amphibian Rana catesbeiana. J. Immunol. 111:1261 1267, 1973
Goidl, E.A. and Siskind, G.W.: Ontogeny of B lymphocyte function, i. Restricted heterogeneity of the antibody response of B lymphocytes from neonatal and fetal mice. J. Exp. Med. 140:1285 1302, 1974
Hadji-Azimi, I. : Studies on Xenopus laevis immunoglobulins. Immunology 21:463-474, 1971 Haimovich, J. and Du Pasquier, L.: Specificity of antibodies in amphibian larvae possessing a
small number of lymphocytes. Proc. Nat. Acad. Sei. USA. 70:1898 1902, 1973 Haimovich, J., Novik, N., and Sela, M. : Inhibition of the inactivation of modified phage. Isr.
J. Med. Sei. 5:438, 1969 Marchalonis, J.J. and Atwell, J.L.: Phylogenetic emergence of distinct immunogloublin classes.
In L'Etude phylogknique et ontogknique de la rOsponse immunitaire et son apport & la thkorie immunologique, pp. 153 162. Inserm., Paris, 1973
Moticka, E.J., Brown, B.A., and Cooper, E.L. : Immunoglobulin synthesis in bullfrog larvae. J. Im- munol. 110:855 861, 1973
Nieuwkoop, P.D. and Faber, J. : Normal Table ofXenopus laevis (Daudin). North Holland, Amster- dam, 1967
Rosenquist, G.L. and Hoffman, R.Z. : The production of anti-DNP antibody in the bullfrog Rana catesbeiana. J. Immunol. 108:1499 1505, 1972
Segal, S., Globerson, A., Feldman, M., Haimovich, J., and Sela, M.: In vitro induction of a primary response to the dinitrophenyl determinant. J. Exp. Med. 132:93-99, 1970
Siskind, G.W. and Benacerraf, B. : Cell selection by antigen in the immune response. Adv. Immunol. 10:1 50, 1969
Spear, P.G., Wang, A.L., Rutishauser, U., and Edelman, G.M. : Characterization of splenic lymphoid cells in fetal and newborn mice. J. Exp. Med. 138:557-573, 1973
Urbain, J., Van Acker, A., de Vos-Cloetens, C.H., and Urbain-Vansanten, G. : Increase and decrease in binding affinity of antibodies during the immune response. Immunochemistry 9:121 136, 1972
Volpe, E.P.: Transplantation immunity and tolerance in Anurans. Transplant. Proc. 2:286 292, 1970
Witschi, E. : Development of Vertebrates, pp. 78-92, Saunders, Philadelphia, 1956