the machinery for cell polarity, cell morphogenesis, and the cytoskeleton in the basidiomycete...
TRANSCRIPT

THE MACHINERY FOR CELL POLARITY, CELLMORPHOGENESIS, AND THE CYTOSKELETON IN THEBASIDIOMYCETE FUNGUS Ustilago maydis A SURVEY OF THEGENOME SEQUENCE#
Flora Banuett1,*, Rene H. Quintanilla Jr.1,&, and Cristina G. Reynaga-Peña2
1,*Department of Biological Sciences, California State University, 1250 Bellflower Boulevard, Long Beach,CA 90840, Phone: (562)-985-5535. email: [email protected]
1,&Department of Biological Sciences, California State University, 1250 Bellflower Boulevard, Long Beach,CA 90840, email: [email protected]
2Departamento de Ingeniería Genética, Centro de Investigación y de Estudios Avanzados del IPN, UnidadIrapuato, Campus, Guanajuato, Irapuato, Gto. CP 36821 MEXICO, Phone: +52 (462) 623-9653, e-mail:[email protected]
AbstractUstilago maydis, a Basidiomycete fungus that infects maize, exhibits two basic morphologies, ayeast-like and a filamentous form. The yeast-like cell is elongated, divides by budding, and the budgrows by tip extension. The filamentous form divides at the apical cell and grows by tip extension.The repertoire of morphologies is increased during interaction with its host, suggesting that plantsignals play an important role in generation of additional morphologies. We have used S.cerevisiae and S. pombe genes known to play a role in cell polarity and morphogenesis, and in thecytoskeleton as probes to survey the U. maydis genome. We have found that most of the yeastmachinery is conserved in U. maydis, albeit the degree of similarity varies from strong to weak. TheU. maydis genome contains the machinery for recognition and interpretation of the budding yeastaxial and bipolar landmarks; however, genes coding for some of the landmark proteins are absent.Genes coding for cell polarity establishment, exocytosis, actin and microtubule organization,microtubule plus-end associated proteins, kinesins, and myosins are also present. Genes not presentin Saccharomyces cerevisiae and Schizosaccharomyces pombe include a homologue of mammalianRac, a hybrid myosin-chitin synthase, and several kinesins that exhibit more similarity to theirmammalian counterparts. We also used the U. maydis genes identified in this analysis to search otherfungal and other eukaryotic genomes to identify the closest homologues. In most cases, notsurprisingly, the closest homologue is among filamentous fungi, not the yeasts, and in some cases itis among mammals.
Correspondence to: Flora Banuett.#In memoriam Ira HerskowitzPublisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customerswe are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resultingproof before it is published in its final citable form. Please note that during the production process errors may be discovered which couldaffect the content, and all legal disclaimers that apply to the journal pertain.
NIH Public AccessAuthor ManuscriptFungal Genet Biol. Author manuscript; available in PMC 2009 August 1.
Published in final edited form as:Fungal Genet Biol. 2008 August ; 45(Suppl 1): S3–S14. doi:10.1016/j.fgb.2008.05.012.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Keywordspolarized growth; cytoskeleton; polarisome; fungal morphogenesis
I. OVERVIEW OF CELL POLARITY AND CELL ORPHOGENESISCell polarity is a fundamental biological process by which asymmetry of cell components(cytoskeleton, cell organelles, and membrane domains) is generated. Polarization of cellcomponents occurs in diverse cells in response to endogenous programming and to externalcues. This cellular asymmetry is important in determining cell morphogenesis and the planeof cell division in a variety of eukaryotic cells, and is crucial for development of metazoans.Studies in budding yeast and fission yeast have been instrumental in identification of themachinery involved in cell polarity and morphogenesis. These studies indicate that recognitionof a spatial landmark triggers assembly of protein complexes that initiate assembly andpolarization of the actin cytoskeleton, which directs vesicle delivery to sites of growth.
Fungal filaments (hyphae) exhibit continuous polarized growth at their tips. New axes ofpolarized growth (branches) are established along the main axis of polarized growth. Currentwork in filamentous fungi aims to understand the molecular mechanisms that determine: (a)the axis of polarized growth, (b) new axes of cell polarity, and (c) the site of cytokinesis andseptum formation. The availability of the genome sequences of fungi from across the differentsubdivisions of the kingdom, together with improved tools for generation of knock out strains,regulatable gene expression, and sophisticated imaging techniques promise to reveal newinsights about polarized growth and cell morphogenesis in filamentous fungi, and will likelylead in the future to the generation of protein interaction maps for this machinery as has beendone in Saccharomyces cerevisiae (Drees et al., 2001).
Here we explore the genome sequence of Ustilago maydis in search of homologues of genesknown to play crucial roles in cell morphogenesis and cell polarity in the yeasts. We firstprovide a synopsis of the morphologies that characterize U. maydis growth in culture and inthe plant. We follow with a brief summary of key players of polarized growth in S.cerevisiae, and indicate which components of the machinery are conserved in U. maydis, andwhere known, describe their role in polarized growth. We conclude from our analysis that mostof the yeast machinery is conserved in U. maydis, as it is in other filamentous fungi (Harris,2006; Philippsen et al., 2005), and in many instances it is used in novel ways.
II. OVERVIEW OF THE MORPHOLOGICAL TRANSITIONS IN THE LIFE CYCLEOF U. maydis
Ustilago maydis is a Basidiomycete fungus that infects maize (Zea mays L) and teosinte (Zeamays spp. mexicana or spp. parviglumis). The fungus exhibits two basic morphologies and iscapable of switching from one form to the other (the dimorphic switch) (Fig. 1). This switchis crucial to pathogenicity. The yeast-like form is unicellular, haploid, divides by budding, andis nonpathogenic; the filamentous form is dikaryotic, grows by tip extension, and is pathogenic(reviewed in Banuett, 1995;2002;Christensen, 1963;Holliday, 1974;Klosterman et al., 2007).The dimorphic switch and other aspects of the life cycle are controlled by two unlinked matingtype loci, a and b. The a locus codes for components of a signal transduction pathway(pheromone precursor and receptor genes) and the b locus codes for a combinatorialhomeodomain protein. The a locus governs cell fusion of haploid cells and filamentous growthof the dikaryon in vitro but not in planta; the b locus is the major determinant of filamentousgrowth, in vitro and in planta, and of pathogenicity (reviewed in Banuett, 2002;2007;Klosterman et al., 2007). In order to complete the life cycle, haploids that fuse must differ
Banuett et al. Page 2
Fungal Genet Biol. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

at both a and b (for example, a1 b1 + a2 b2). The a locus has two alleles (a1 and a2) and theb locus 25 naturally occurring alleles (b1....b25), and any combination of different b allelesresults in an active b protein (reviewed in Banuett 2002;2007;Klosterman et al., 2007).
The repertoire of morphologies is expanded by interaction of U. maydis with its hosts (Fig. 1).This is evidenced by the formation of a specialized structure for penetration, the appressorium,(see for example, Brachmann et al., 2003; Snetselaar and Mims, 1993), by extensive branchingon the leaf surface and inside plant cells (Banuett and Herskowitz, 1996), and by formation ofclamp-like structures for nuclear distribution (Scherer et al., 2006). Appressoria, branchformation, and clamp-like structures are not observed in culture. Once tumors are formed,dikaryotic hyphae undergo a discrete developmental program characterized by distinctmorphologies (cylindrical cells, spherical cells, and other diverse shapes) that arise upon hyphalfragmentation and lead to formation of the teliospore, a round spore with a specialized cellwall (Fig. 1) (Banuett and Herskowitz, 1996). The distinct cell morphologies present in theplant are not observed in vitro. It has thus been proposed that the plant produces signals thattrigger fungal differentiation and reorganization of the machinery for polarized growth(Banuett and Herskowitz, 1996). The nature of these signals is not known. Interestingly, thefungus also elicits changes in cell morphology and nuclear position of the host (Banuett andHerskowitz, 1996;Ruiz-Herrera et al., 1999; reviewed in Banuett, 2002). Thus, the interactionof U. maydis with its host appears to be a reciprocal process of signal exchange that results inalterations in cell morphology of both host and fungus.
A. The yeast-like cell morphologyThe yeast-like cells are elongated with tapered ends (cigar-shaped). They bud once per cellcycle at one of the cell poles. The mechanism by which one of the cell poles is chosen versusthe other is not known. Cells are able to bud at new sites at the cell poles or use a previouslychosen site for budding as in apiculate yeasts (Jacobs et al., 1994). In some respects this patternresembles the bipolar budding pattern of S. cerevisiae. The U. maydis bud grows by tipextension (Banuett and Herskowitz, 2002). There is no phase of isotropic growth, whichcontrasts with bud growth in S. cerevisiae where there is a short phase of polarized growthearly in the cell cycle followed by a switch to isotropic growth in the G2/M phase of the cellcycle (reviewed in Pruyne and Bretscher, 2000a,b). Formation of the U. maydis bud entailsseveral morphological steps that result in the final shape of the bud (Banuett and Herskowitz,2002).
B. The filamentous formIn culture, the filamentous form grows by tip extension, as occurs in other filamentous fungi,and divides at the apical cell to produce an actively growing tip cell and a subapical cell devoidof cytoplasm, but little is known about division of the apical cell. In planta, most hyphalcompartments, not just the tip cell, contain cytoplasm (Banuett and Herskowitz, 1996) andnuclear distribution involves clamp-like structures (Scherer et al., 2006).
C. The Spitzenkörper and filamentous growthFungal hyphae exhibit continuous polarized growth; there is no arrest of growth duringcytokinesis. Examples of cells that exhibit such highly polarized growth are pollen tubes androot hairs in plants, and neurons in animals.
In filamentous fungi, a fungal-specific phase-contrast opaque body, the Spitzenkörper (apicalbody), located at or just below the tip of the apical cell, is thought to drive hyphal growth(López-Franco, 1996; Reynaga-Peña et al., 1997). It consists of a heterogeneous population ofvesicles surrounding a core that contains polysomes, microtubules, and actin, and is proposedto serve as a supply center for the distribution of vesicles containing materials necessary for
Banuett et al. Page 3
Fungal Genet Biol. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

tip extension (reviewed in Bartnicki-García, 2002; Harris et al., 2005; Harris, 2006). TheSpitzenkörper can be visualized with the amphiphilic styryl dye FM4-64 (Fischer-Parton et al,2000). Long-range transport of vesicles from other parts of the hypha to the Spitzenkörper isproposed to occur along microtubules, and delivery from the Spitzenkörper to sites of growthis proposed to occur on actin tracks (reviewed in Harris et al., 2005; Harris, 2006). Bothmicrotubules and actin are necessary for hyphal morphogenesis. Disruption of microtubules,kinesin, dynein, and dynactin results in unstable axes of polarized growth but not in absenceof polarized growth per se, whereas disruption of actin causes complete loss of cell polarityand results in isotropic growth (Harris et al., 2005; Heath, 2000; Lee et al., 2001; Riquelme etal., 2000; Rupes et al., 1995; Seiler et al., 1999; Torralba et al., 1998a,b; Virag and Griffiths,2004). Some of these conditions also result in loss or alteration of the position or function ofthe Spitzenkörper (Crampin et al., 2005; Konzack et al., 2005; Riquelme et al., 2000; Seiler etal., 1999).
There is a dearth of information concerning the molecular composition of the Spitzenkörper.Recent work in different filamentous fungi indicates that formin (a multidomain, actin-nucleating protein), myosin light chain, Sec4 (a Rab GTPase), BemA (homologue of S.cerevisiae Bem1), TeaA and TeaR (homologues of S. pombe end cell markers Tea1 and Mod5,respectively), and two chitin synthases are components of the Spitzenkörper (see below)(Crampin et al., 2005; Leeder and Turner, 2007; Riquelme et al., 2007; Sharpless and Harris,2000; Takeshita et al., 2008). The presence of formin suggests that the Spitzenkörper may serveas an actin nucleation center. Little is known about the Spitzenkörper in U. maydis.
III. U. maydis HOMOLOGUES OF YEAST GENES INVOLVED IN CELLPOLARITY AND CELL MORPHOGENESIS
The yeasts, S. cerevisiae and Schizosaccharomyces pombe have been used extensively instudies of cell polarity and morphogenesis (Chang and Peter, 2003). These studies provide theframework in which to understand these processes in other organisms, in particular the fungi;therefore, we have used these two yeasts as a reference point in our analysis. Below we describesome of the components of the machinery for polarized growth in S. cerevisiae (for S.pombe see supplementary material), and identify components conserved in U. maydis. Wedescribe, where known, their function in U. maydis, and in some filamentous fungi.
A. A hierarchy of GTPase modules controls bud site selection and establishment of polarizedgrowth in S. cerevisiae
1. Recognition of the spatial landmark—S. cerevisiae cells divide by budding andexhibit two budding patterns, axial and bipolar, which are genetically determined by theMAT locus. a or α cells exhibit the axial budding pattern, in which mother and daughter cellsbud immediately adjacent to the previous budding site. a/α cells exhibit a bipolar buddingpattern, in which daughters bud distally to the site of their birth, whereas the mother can buddistally or proximally to the previous budding site (reviewed in Casamayor and Snyder,2002; Chant, 1999). A GTPase module consisting of a Ras-like GTPase Rsr1/Bud1, its guaninenucleotide exchange factor (GEF) Bud5, and its GTPase-activating protein (GAP) Bud2determine the site of budding in response to a spatial landmark for axial or bipolar budding(reviewed in Casamayor and Snyder, 2002; Chant, 1999). The landmark for axial buddingconsists of septins, Bud3, Bud4, Axl1, Bud10/Axl2, and for bipolar budding of Bud8, Bud9,Bud7, Rax1, and Rax2. The Rsr1/Bud1 GTPase module is not essential for viability; in itsabsence cells are still capable of polarized growth but in a random manner (reviewed inCasamayor and Snyder, 2002; Chant, 1999; Park and Bi, 2007).
Banuett et al. Page 4
Fungal Genet Biol. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

2. Polarity establishment and the Cdc42 GTPase module—The Rsr1/Bud1 GTPasemodule interprets the spatial landmark (axial or bipolar) and recruits another GTPase modulethat consists of the Rho GTPase Cdc42, its guanine nucleotide exchange factor (GEF) Cdc24,and its GTPase-activating proteins (GAPs) Bem3 and Rga1/2 (reviewed in Casamayor andSnyder, 2002; Chant, 1999; Park and Bi, 2007). This module in turn recruits the actincytoskeleton, which polarizes secretion towards the site of growth (reviewed in Pruyne andBretscher, 2000a,b).
B. U. maydis homologues of landmark proteins and the BUD site selection and polarityestablishment GTPase modules
The U. maydis genome lacks coding information for homologues of Bud3 and Bud4 (axiallandmark), and Bud8 and Bud9 (bipolar landmark) (Table 1), but contains the information forBud10/Axl2 (axial landmark), and Rax1 and Rax2 (bipolar landmark). Thus, U. maydiscontains a subset of the genes that code for the axial and bipolar landmark proteins. Their rolein budding and cell morphogenesis remains to be determined. Interestingly, in Ashbyagossypii, a filamentous fungus closely related to S. cereviasae, AgBud3 appears to act as alandmark for septation (Wendland, 2003). Coding information for components of the Rsr1/Bud1 GTPase module (Rsr1/Bud1, Bud2, and Bud5) and many other recently identified genesrequired for bipolar budding (BUD13-32;Ni and Snyder, 2001) is present in the U. maydisgenome (Table 1). The function of these genes remains to be determined. In A. gossypii,AgRsr1/Bud1 is required for actin organization, for normal hyphal growth and branching, andfor position of the polarisome component AgSpa2. Control of the position of a polarisomecomponent by the Rsr1/Bud1 GTPase is a novel function of this GTPase. The Agrsr1 nullmutant is characterized by false branch initiation, pausing and reinitiation of growth resultingin bulges along the hypha, and transient appearance and disappearance of AgSpa2 thatcorrelates with phases of pausing and growth (Bauer et al., 2004). In wild type hyphae, AgSpa2(a homologue of S. cerevisiae Spa2, see below) is present at the hyphal tip continuously. InCandida albicans, another fungus closely related to S. cerevisiae, CaRsr1/Bud1 defects resultin random budding in yeast cells and in defects in germ tube emergence (Yaar et al., 1997).
The U. maydis genome contains coding information for homologues of the Rho Cdc42 GTPasemodule (Cdc42, Cdc24, Bem3, Rga1/2) (Table 1). In addition, U. maydis, like otherfilamentous fungi, contains a homologue of mammalian Rac, another Rho GTPase, which isabsent from the genomes of S. cerevisiae and S. pombe. In mammals, Cdc42 and Rac1 playpartially overlapping roles in morphogenesis, cell division, and migration (Jaffe and Hall,2005). Likewise in filamentous fungi, Cdc42 and Rac appear to have partially overlappingfunctions in control of cell polarity (see for example, Bassilana and Arkowitz, 2006;Boyce etal., 2001,2003;Chen and Dickman, 2004;2006;Rolke and Tudzynski, 2008;Scheffer et al.,2005;Virag et al., 2007).
In U. maydis, Cdc42 is required for cell separation but not for polarized growth, and is notessential for viability (Mahlert et al., 2006; F. Banuett, unpublished results), whereas UmRac1is required for normal cell morphology of the yeast-like cell and for positioning of the septum(Mahlert et al., 2006). Umcdc42 null mutants form chains of cells that exhibit wild typemorphology, separated by a single septum (Mahlert et al., 2006). In wild type cells, there aretwo septa prior to cell separation, one on the mother side and the other on the bud side. Cellseparation occurs in the region between these two septa, the fragmentation zone (Banuett andHerskowitz, 2002; O’Donnell and McLaughlin, 1984; Weinzierl et al., 2002). In S. cerevisiaeCDC42 is essential for viability and for polarity establishment: cdc42ts and cdc24 mutantsarrest as large unbudded, multinucleate cells (reviewed in Pruyne and Bretscher, 2000a,b).Thus, U. maydis and S. cerevisiae use Cdc42 differently, a theme repeated in other filamentousfungi (see below).
Banuett et al. Page 5
Fungal Genet Biol. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

The importance of Rac1 in polarized growth is supported by the following observations:expression of an activated allele of Rac1 results in formation of balloon-like structures at thetips of yeast-like cells; and Umrac1 null mutants exhibit elongated abnormal cells with roundedends and a septum in the middle; wild type cells, in contrast, have tapered ends (Mahlert et al.,2006). UmRac1 is not required for cell viability, but depletion of UmRac1 in a strain lackingUmcdc42 is lethal: cells arrest as chains of enlarged, aberrant cells, with a single nucleus, anddelocalized cell wall material (Mahlert et al., 2006). This observation suggests that Cdc42 andRac1 share a common target. Rac1 is also necessary for the b-dependent transition from yeast-like to filamentous form: induction of Umrac1 expression in haploid cells (haploids normallylack an active b protein) causes a transition from yeast-like to filamentous form. UmRac1 isthus necessary and sufficient for the dimorphic switch that results in filament formation(Mahlert et al., 2006). The GEF and GAP regulators of the UmRac1 GTPase remain to beidentified.
In Aspergillus nidulans Cdc42 is required for hyphal morphogenesis, but it is not required forlocalization of polarisome components, the Spitzenkörper, or organization of actin, and neitheris AnRac1 required for localization of polarisome components or the Spitzenkörper (Virag etal., 2007). In contrast, S. cerevisiae Cdc42 is required for localization of polarisomecomponents and actin organization (reviewed in Chang and Peter, 2003; Pruyne and Bretscher,2000a, b). AnCdc42 and AnRac likely share a common target because double Anrac1Ancdc42 mutants are synthetic lethal: the double mutants are unable to polarize and lyse asattached rounded cells separated by septa (Virag et al., 2007). In Penicillium marneffei, Rac isrequired for normal organization of the actin cytoskeleton. Rac regulates polarized growth ofthe hypha in concert with Cdc42 but the specific steps controlled by these Rho GTPases differ(Boyce et al., 2001, 2003). The synthetic lethality of cdc42 and rac1 in U. maydis and otherfilamentous fungi supports the notion that these Rho GTPases play partially overlapping rolesin control of cell polarity.
C. U. maydis homologues of effectors of Cdc42In S. cereviasiae, downstream effectors of Cdc42 include the p21-activated kinases (PAKs)Cla4, Ste20, and Skm1, and two proteins of unknown function, Gic1 and Gic2 (reviewed inPruyne and Bretscher, 2000a, b). Ste20 and Cla4 are important for the Cdc42-dependentreorganization of the actin cytoskeleton during the cell cycle. The U. maydis genome containscoding information for three Ste20-like kinases (Table 1): Smu1, Cla4, and Don1, but not forGic1 and Gic2. Smu1 and Cla4 contain the CRIB domain present in p21-activated kinases(PAKs), whereas Don1 lacks this domain, and thus belongs to the germinal center (GCK)subfamily of Ste20-like kinases (Leveleki et al., 2004; Smith et al., 2004). (For further detailson Smu1, see the article by García-Pedrajas et al. in this issue). Both UmDon1 and UmCla4play a role in cytokinesis (Leveleki et al., 2004), just as Cla4 and Ste20 do in S. cerevisiae.Two observations support the idea that UmCla4 is likely to be a downstream effector ofUmRac1: both Δcla4 and Δrac1 mutants have a similar phenotype, and they show yeast two-hybrid interactions. Other effectors of Rac1 are likely to exist because Δcla4 does not suppressthe lethality observed upon expression of an activated allele of Rac1 (Mahlert et al., 2006).
D. U. maydis homologues of other Rho proteinsThe Rho subfamily of GTPases includes Rho, Rac, Cdc42, and others. They act as molecularswitches that play important roles in cell polarity, morphogenesis, and cell division (Jaffe andHall, 2005). S. cerevisiae contains one Cdc42 (described above) and five Rho proteins, Rho1-5.Rho1, an essential gene for viability, is required for polarity establishment. Rho3 and Rho4are also required for polarized growth. The signal from different Rho proteins (Rho1, Rho3,Rho4, and Cdc42) converges on the formin Bni1, a polarisome component, to control actinorganization (see below) (Dong et al., 2003; reviewed in Pruyne and Bretscher, 2000a,b). In
Banuett et al. Page 6
Fungal Genet Biol. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

addition, Rho1 controls bud morphogenesis via two different outputs: as a catalytic subunit ofβ-glucan synthase, the enzyme that synthesizes the major component of the cell wall, and asan activator of protein kinase C (Pkc1) that controls the cell wall integrity signal transductionpathway (reviewed in Banuett, 1998; Cabib et al., 1998; Pruyne and Bretscher, 2000a,b). Theregulators of Rho1 include two redundant GEFs, Rom1 and Rom2, and two GAPs, Sac7 andBem3. The U. maydis genome contains coding information for three Rho proteins, Rho1, Rho2,and Rho3, and for homologues of Rho1 regulators (Table 1). Their function has not beenreported. Rho proteins in other filamentous fungi are required for polarized growth and it islikely that this function will be conserved in U. maydis. In A. gossypii, rho3 mutants exhibitswelling of hyphal tips indicative of isotropic growth, and rho1 null mutants exhibit lysis ofcells in a colony (Wendland and Philippsen, 2001). In A. nidulans, RhoA, a homologue of S.cerevisiae Rho1, is necessary for polarized growth, branching pattern, and cell wall deposition(Guest et al., 2004).
E. U. maydis homologues of polarisome componentsIn S. cerevisiae, the polarity determining proteins, Spa2, Pea2, Bud6/Aip3, and Bni1, assembleat the bud tip and function as a scaffold for the assembly of other proteins. Mutants lackingeither of these proteins exhibit defects in actin organization, apical growth, and have widenednecks (reviewed in Pruyne and Bretscher, 2000a,b). The polarisome links Rho GTPases andactin cable assembly through the formin Bni1. Formins are multidomain proteins required formaintenance of cell polarity, cytokinesis, and actin organization, and are central to how inputsfrom Rho GTPases are transmitted to actin organization (Chang and Peter, 2003; Evangelistaet al., 2003; Wallar and Alberts, 2003). Bni1 binds profilin, an actin-monomer binding protein,and has been shown to nucleate actin cables. Bud6 is important for the Bni1 actin-nucleationfunction (Pruyne et al., 2002; Sagot et al., 2001). Localization of Bni1 to the bud tip requiresSpa2 and Pea2. Some polarisome components also localize to the mother-bud neck regionduring cytokinesis to participate in formation of the actomyosin ring and the septum (reviewedin Pruyne and Bretscher, 2000a,b).
The U. maydis genome contains coding information for homologues of polarisome componentsBud6, Spa2, and two formins, UmSepA and UmDia, but not of Pea2 (Table 1). The role ofthese proteins in cell polarity of the yeast-like and filamentous form in U. maydis remains tobe determined. Recent studies in A. gossypii, A. nidulans, and C. albicans (Crampin et al.,2005;Knechtle et al., 2003;Virag and Harris, 2006) are providing insights about the role of thepolarisome complex in polarized growth of the hypha. In A. nidulans, homologues of Spa2,Bud6/Aip3, and Bni1 are necessary for maintaining the axis of polarized growth: mutations ineither component result in dichotomous branching (branching at the hyphal tip), which isnormally not observed in wild type hyphae (Sharpless and Harris, 2002;Virag and Harris,2006). SpaA::GFP localizes to the hyphal tip, overlapping to some extent the localization ofthe Spitzenkörper, consistent with a role at the hyphal tip, possibly as a component of theSpitzenkörper (Virag and Harris, 2006). SpaA is not required for localization of the forminSepA (Virag and Harris, 2006), whereas S. cerevisiae Spa2 is required for localization of theformin Bni1 (reviewed in Pruyne and Bretscher, 2000a,b). BudA, the A. nidulans homologueof Bud6/Aip3, localizes to sites of septum formation. The AnbudA mutant phenotype isconsistent with a role in maintenance of the axis of polarized growth and determining the areaof growth. AnbudA null mutants exhibit reduced hyphal growth rate, increased hyphal diameterand nuclei per hyphal segment, dichotomous branching, and increased branching frequency,and potato-shaped germlings (Virag and Harris, 2006). AnSepA is required for septumformation, for maintenance of the axis of polarized growth, and determining the area of growth,and normal organization of actin at the hyphal tips and septa. SepA null mutants exhibit widerhyphal diameter, dichotomous branching, and lack septa. SepA::GFP localizes to hyphal tips,coincident with localization of the Spitzenkörper, and also to septa (Sharpless and Harris,
Banuett et al. Page 7
Fungal Genet Biol. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

2002). This localization led to the hypothesis that the Spitzenkörper acts as an actin-nucleatingcenter (Sharpless and Harris, 2002).
In A. gossypii, the Spa2 homologue, which localizes permanently to the hyphal tip and to sitesof branch initiation, appears to determine area of growth, normal growth rate, and branchingdensity (Knechtle et al., 2003). AgBNI1 is necessary for establishment of polarized growth andfor actin organization. Disruption of AgBNI1 results in formation of giant potato-shaped cellslacking actin cables and also in the inability to form hyphae (Schmitz et al., 2006). AgBni1localizes to hyphal tips. A constitutively active AgBni1 results in dichotomous hyphalbranching in young hyphae, a type of branching normally restricted to mature hyphae (Schmitzet al., 2006). The localization of these proteins at the hyphal tip supports a role in determiningpolarized growth.
In C. albicans, CaBni1 and CaMlc1 (myosin light chain) colocalize with the Spitzenkörper,whereas Spa2 and Bud6/Aip3 localize mostly as a cap at the hyphal tip that overlays theSpitzenkörper (Crampin et al., 2005). Thus, in C. albicans it seems that polarized growth ofhyphae is determined by two separate entities, the Spitzenkörper and the polarisome, whereasin pseudohyphae and yeast-like cells, the polarisome is responsible for polarized growth; thereis no Spitzenkörper in these cells.
F. U. maydis homologues of septinsSeptins are GTP binding proteins that are conserved from fungi to mammals and play a rolein cytokinesis and other cell processes (Gladfelter et al., 2001; Longtine and Bi, 2003; Pan etal., 2007). In S. cerevisiae, septins form filaments at the mother-bud neck region, arrangedlongitudinally (along the main cell axis) in an hour-glass shape or as horizontal rings in a cellcycle-dependent manner (Gladfeleter et al., 2001). Assembly of septins requires Cdc42,polarisome components, the PAK kinase, Cla4, and the actin cytoskeleton (Kadota et al.,2004). Septins are proposed to serve a scaffold role and recruit other proteins: bud site selectionlandmarks, components of the actomyosin contractile ring (CAR), chitin synthases, and otherproteins (Gladfelter et al., 2001; Longtine and Bi, 2003). Septins also act as diffusion barriersfor integral membrane proteins and have been proposed to serve as organizers of corticaldomains of the cell in addition to their scaffold role (Barral et al., 2000; Takizawa et al.,2000).
S. cerevisiae contains 7 septins, Cdc3, Cdc10, Cdc11, Cdc12, Sep7, Spr4, Spr28; the latter twoare meiosis-specific and required with other septins during spore maturation. The U. maydisgenome contains coding information for homologues of S. cereviase Cdc3, Cdc10, Cdc11, andCdc12 (Table 1), whereas in other filamentous fungi there appear to be at least five putativeseptins (see Borkovich et al., 2004). U. maydis Sep3, a homologue of Cdc11, is required fornormal cell morphogenesis (Boyce et al., 2005). Haploid Sep3 mutants exhibit aberrant cellmorphology: the cells are “lemon drop-shaped”, with a septum in the middle, becomemultinucleated, and have altered chitin deposition. Sep3 is also required for normalmorphology of the promycelium (a short filament produced upon germination and meiosis ofthe teliospore; Christensen, 1963), and the basidiospores (the haploid cells produced on thepromycelium by mitosis of the four primary meiotic products). Genetic analysis indicates thatSep3 acts in the cAMP signaling pathway (Boyce et et al., 2005). The role of other septins inU. maydis remains to be uncovered. In A. nidulans, AspB, a homologue of S. cerevisiae Cdc3,is hypothesized to act as a marker of branch formation. It localizes premitotically as a ring tosites of branching and secondary germ tube emergence. It also localizes postmitotically to sitesof septation (Westfall and Momany, 2002).
Banuett et al. Page 8
Fungal Genet Biol. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

G. U. maydis homologues of the exocyst complexIn S. cerevisiae, the exocyst is a protein complex required for exocytosis and consists of Sec3,Sec5, Sec6, Sec8, Sec10, Sec15, Exo70 and Exo84 (TerBush et al., 1996). It is localized to thebud tip during polarized growth and to the neck region during cytokinesis, and is involved infusion and docking of vesicles transported on actin cables. The Sec3 protein acts as a spatiallandmark for the assembly of the exocyst complex (Finger et al., 1998). The assembly of thedifferent protein complexes described above provides a temporal and spatial link between budsite selection with organization of the cytoskeleton, and secretion resulting in cell growth.
The U. maydis genome contains coding information for all the components of the exocystcomplex (Table 1) but little is known of their role in polarized growth and cytokinesis. In A.nidulans, SecC, a homologue of S. cerevisiae Sec3, accumulates at the plasma membraneanterior to the Spitzenkörper and may function as a landmark for secretion (Taheri-Talesh etal., 2008).
IV. THE CYTOSKELETONThe cytoskeleton plays an important role in cell morphogenesis and polarized growth in diverseorganisms. In U. maydis, both actin and microtubules are required for cell morphogenesis, asin other filamentous fungi.
A. The actin cytoskeletonThe actin cytoskeleton in U. maydis consists of three structures: actin patches, actin cables,and an actin ring as observed in other fungi. The actin cytoskeleton is highly polarizedthroughout the cell cycle: actin patches concentrate at the presumptive bud site and at the tipof the growing bud, and actin cables polarize towards the actin patches and extend into themother cell (Banuett and Herskowitz, 2002). During cytokinesis, actin forms a ring in the neckregion. Actin patches persist at the growing end during this process. After cell division iscomplete, both cell ends contain actin patches, with one end having a higher concentration thanthe other. This distribution of actin is consistent with highly polarized growth at the tip of thecell throughout the cell cycle. There is no evidence for a disorganized actin cytoskeleton at anystage of the cell cycle as occurs during isotropic growth in S. cerevisiae, consistent withobservations that there is no isotropic growth during bud morphogenesis in U. maydis (Banuettand Herskowitz, 2002). U. maydis hyphae contain a prominent actin cap at the tip and actincables polarize towards the tip, as observed in other filamentous fungi (Banuett andHerskowitz, 2002).
1. U. maydis homologues of components of the actin cytoskeleton—In S.cerevisiae, the actin cytoskeleton is involved in polarized secretion. Actin cables serve as trackson which vesicles move on myosin motors towards sites of active growth and secretion (thebud tip during polarized growth and the neck region during cytokinesis) (Karpova et al., 2000;Pruyne and Bretscher, 2000a,b). All components involved in actin assembly and function inS. cerevisiae appear to be conserved in the U. maydis genome (Table 1). For example, genescoding for the Arp2/3 complex (Winter et al., 1999), responsible for nucleation of branchedactin structures, are present in the U. maydis genome, as well as genes coding for homologuesof Bee1/Las17, the activator of Arp2/3 (a homologue of the mammalian Wiskott-Aldrichsyndrome protein WASP), and Vrp1 (homologue of mammalian Wip, Wiskott-Aldrichsyndrome protein interacting-protein). In S. cerevisiae, Arp2/3, Bee1/Las17, and Vrp1 localizeto actin patches and form an actin cap during early bud morphogenesis (reviewed in Pruyneand Bretscher, 2000a,b). The S. cerevisae genome contains two tropomyosin isoforms, Tpm1and Tpm2. These proteins stabilize actin cables (reviewed in Pruyne and Bretscher, 2000a,b).The U. maydis genome contains a single tropomyosin gene, and the putative protein is similar
Banuett et al. Page 9
Fungal Genet Biol. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

to S. cerevisiae Tpm2. The function of tropomyosin in U. maydis remains to be determined.The U. maydis genome contains coding information for homologues of proteins such as actinbinding protein (Abp1), coronin (Crn1), cofilin (Cof1), profilin (Pfy1), fimbrin (Sac6), Pan1,Prk1, Srv2, Sla1 and Sla2, some of which are actin patch components (Table 1). The functionof these proteins in U. maydis has not been reported. (See Moseley and Goode, 2006, for areview on the function of these proteins in S. cerevisiae).
2. U. maydis homologues of myosins—As in other filamentous fungi, the U. maydisgenome contains coding information for 4 myosins: a class I (Myo1), a class II (Myo2), a classV (Myo5), and a class XVII (myosin-chitin synthase; Csm1) (Table 1) (see Hodge and Cope,2000, for myosin classification). Myosin-chitin synthase contains an amino-terminal myosindomain and a chitin synthase domain at the carboxy terminus. It is specific to the filamentousfungi; the genomes of S. cerevisaie or S. pombe lack this information. In the yeast-like cell,Ummyo5 is required for cell morphogenesis and cell separation: Ummyo5 null mutants areviable and exhibit slow growth and formation of clusters of shorter and wider cells, which failto separate (Weber et al., 2003). Young myo5 mutant cells still exhibit polarized growth andpolarized chitin at the cell tip, whereas older cells lose polarity and become aberrantly shaped(Weber et al, 2003). In the hypha, Ummyo5 appears to be required for the axis of polarizedgrowth and to determine the area of growth: hyphae deficient in Myo5 are wider in diameterand exhibit an irregular growth pattern (spiral-like), and contain irregular deposition of chitinalong the hyphal length (Weber et al., 2003). Myo5::GFP localizes to the incipient bud site,the bud tip in cells with small buds, and becomes more dispersed in large-budded cells; inhyphae, it localizes to the tip, consistent with a role at the cell tip (Weber et al., 2003). Thefunction of Myo1 and Myo2 has not been reported. (See below for role of Csm1).
B. The microtubule cytoskeletonIn many eukaryotes, the microtubule cytoskeleton is required for polarized growth, transportof organelles and vesicles, in addition to its role in spindle assembly and function.
The microtubule cytoskeleton in U. maydis consists of an extensive cytoplasmic arraythroughout interphase that disassembles in late G2 when a short intranuclear spindle is formedand astral microtubules are assembled from the spindle pole bodies (Banuett and Herskowitz,2002; Steinberg et al., 2001; Straube et al., 2003). These astral microtubules appear to beinvolved in nuclear movements to the bud, where the nucleus divides, and then after nucleardivision, when one of the nuclei migrates to the mother cell (Banuett and Herskowitz, 2002;Fink et al., 2006; Steinberg et al., 2001). The organization of the microtubule cytoskeleton inU. maydis is more complex than that in S. cerevisiae, and is reminiscent of that observed in S.pombe. In U. maydis yeast-like cells, nucleus-associated spindle pole bodies (SPBs) nucleateastral microtubules, and cytoplasmic microtubule organizing centers (MTOCs), assembled atthe base of the bud early during bud morphogenesis, nucleate microtubules towards the budand mother cell (Straube et al., 2003). The yeast-like cell contains additional motile cytoplasmicMTOCs (Fink and Steinberg, 2006; Straube et al., 2003). In the hypha, the majority of MTsare oriented with their plus ends towards the growing tip; some MTs are oriented with theirplus ends towards the basal end of the apical cell (Schuchart et al., 2005).
The microtubule cytoskeleton is required for normal cell morphogenesis in U. maydis(Steinberg et al., 2001), just as it is in S. pombe (Sawin and Nurse, 1998; reviewed in Chang,2001). In contrast, S. cerevisiae MTs play no role in cell morphogenesis. U. maydis thusresembles S. pombe in the organization of the microtubule cytoskeleton and in the requirementof the microtubule cytoskeleton for cell morphogenesis and for determining the axis ofpolarized growth. U. maydis may thus combine strategies used by S. cerevisiae and S.pombe in novel ways to establish and maintain polarized growth and to generate cell shape.
Banuett et al. Page 10
Fungal Genet Biol. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

1. U. maydis homologues of tubulins—The U. maydis genome contains codinginformation for an α tubulin (tub1), two β tubulins, and a γ tubulin (tub2) (Table 1). Both αand γ tubulin genes have been characterized and shown to be essential for viability (Steinberget al., 2001;Straube et al., 2003). γ tubulin is found at the SPBs and also in the cytoplasmic andpolar MTOCs and its presence in these structures varies in a cell cycle dependent manner(Straube et al., 2003). α tubulin is required for cell morphology and polarized growth. One βtubulin gene was cloned, but its role in cell polarity and cell morphogenesis was not reported(Gold et al., 1994).
2. U. maydis homologues of microtubule-plus-end-associated proteins (+Tips)—+TIPs are a highly diverse group of proteins that includes MT-dependent motors andnonmotor proteins: CLIP170 (ScBik1), Dynactin (Sc Jnm1, Nip100, Arp1, Arp10), EB1 (ScBim1), CLASPS (Sc Stu1), LIS1 (Sc Pac1), Dynein (Sc Dhc1), APC (Sc Kar9), and others(Adkhmanova and Hoogenraad, 2005).
Dynein and dynactin: Cytoplasmic dynein is a multisubunit complex with a MT minus-enddirected motor activity. In U. maydis, as in other fungi, there is a single cytoplasmic dynein.The heavy chain of cytoplasmic dynein is encoded by two unlinked genes: dyn1 codes for theputative ATPase region, and dyn2 codes for the microtubule binding region (Straube et al.,2001). Dyn1 and Dyn2 polypeptides interact to form the active protein. Both genes areessential. Analysis of conditional mutations indicates that dynein is required for cellmorphogenesis, nuclear migration, microtubule organization, retrograde endosomal transport,and organization of the endoplasmic reticulum (Straube et al., 2001; Wedlich-Soldner et al.,2002). Mutant cells in either dyn1 or dyn2 lose polarity (become rounded), contain multiplenuclei that cluster in the aberrantly shaped cells and microtubules that become disorganized,and develop long projections that exhibit polarized growth (Straube et al., 2001).
Dynactin is a multiprotein complex required for activation of cytoplamic dynein-mediatedvesicular transport. It consists of at least ten polypeptides in most eukaryotes; in yeast, fourdynactin subunits have been identified (see above). The p150Glued subunit is the largest andinteracts with the 74-kDa dynein intermediate chain. Other subunits include the actin-relatedprotein Arp1/centractin (45 kDa); the 50 kDa/dynamitin subunit; dynactin p62, dynactin p27,dynactin, p25, dynactin p24/p22, and CapZ (reviewed in Schroer, 2004; Xiang and Plaman,2003). In S. cerevisiae the dynactin complex is required for nuclear migration (Lee et al.,2003; Sheeman et al., 2003). In Neurospora crassa, it is also required for nuclear migration,in addition to a role in determining the axis of polarized growth, and position of theSptizenkörper (Plaman et al., 1994; Riquelme et al., 2000; Seiler et al., 1999). The U.maydis genome contains coding information for homologues of components of the S.cerevisiae dynactin complex (Table 1), and of other dynactin components found in filamentousfungi and other eukaryotes (data not shown), and other +TIPs (Table 1). UmDya1 (thehomologue of p150 Glued), UmClip1 (CLIP170 homologue), UmLis1 (LIS1/NudFhomologue), and UmPeb1 (homologue of EB1) localize to microtubule plus ends (Lenz et al.,2006; Straube et al., 2003). UmDya1 is required for localization of dynein to microtubule plusends. UmLis1/NudF is required for activation of dynein and this activation is necessary forretrogade endosomal movement. UmClip1 is not required for retrogade endosomal movementor binding of dynein to microtubule plus ends (Lenz et al., 2006). In S. cerevisiae, Kar9 andPac1/LIS1 are required for orientation of the mitotic spoindle and nuclear migration (Lee etal., 2003; Miller and Rose, 1998; Sheeman et al., 2003). The U. maydis genome does not containcoding information for a Kar9 homologue.
3. U. maydis homologues of kinesins—In filamentous fungi, as in many othereukaryotes, kinesins are involved in long-distance transport of vesicles. S. cerevisiae contains6 kinesins whereas filamentous fungi contain 10–12 kinesins (see Borkovich et al., 2004; Xiang
Banuett et al. Page 11
Fungal Genet Biol. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

and Plamann, 2003). The U. maydis genome contains coding information for 10 kinesins (Kin1,Kin3, Kin4, Kin6, Kin7a, Kin7b, Kin8, Kin9, and Kin14), some of which are more closelyrelated to their metazoan counterparts than to the ascomycetous fungi (Schuchardt et al.,2005). Genetic analysis has shown that all except Kin1 and Kin3 are dispensable for hyphalgrowth. Null kin1 or kin3 mutants exhibit bipolar growth and short hyphae. Double kin1kin3 null mutants exhibit the same phenotype as the single mutants, indicating that they likelyact in the same pathway (Schuchardt et al., 2005). Interestingly, kin1 and kin3 mutationsexacerbate the mutant phenotype conferred by a myo5ts mutation. The residual polar growthin the myo5ts mutant is completely abolished resulting in clusters of misshapen cells that exhibitno polar growth. This synthetic interaction suggests that Kin1 and Kin3 share a common targetwith Myo5 for polarized growth. Kin1, Kin3, and Myo5 all accumulate at the hyphal apex, theregion where the Spitzenkörper localizes; however, localization of the Spitzenkörper in thesestudies was not reported.
V. U. maydis homologues of chitin synthasesIntegrity of the cell wall is crucial for maintenance of cell shape (Latge, 2007; Klis et al.,2006). Chitin and β-glucan are the major components of fungal cell walls (see article by Ruiz-Herrera in this issue). The U. maydis genome contains 7 genes that code for chitin synthases(Chs1, Chs2, Chs3, Chs4, Chs5, Chs6, Chs7) and one gene for a hybrid myosin-chitin synthaseprotein (Mcs1; see above) (Garcerá-Teruel et al., 2006; Gold and Kronstad, 1994; Weber etal., 2006). Deletion analysis of single genes indicates that Umchs1, Umchs2, Umchs3,Umchs4 are not required for morphology of the yeast-like cell or the hypha, whereas Δchs6results in cells with wider diameter, and Δmcs1 cells exhibit slightly wider cell diameter anda ballooning of the bud tip, indicative of a role in cell morphogenesis. The Umchs5 orUmchs7 null mutants exhibit a more pronounced phenotype indicative of a role in cellmorphology and cell separation, respectively: Δchs5 mutant cells are more elongated than wildtype cells, grow irregularly, and have a neck region that is not clearly defined, whereasΔchs7 mutant cells have a cell separation defect but the cells have normal cell morphology,though they are shorter, and in some cases, lose their normal cell shape (Weber et al., 2006).Six of the U. maydis chitin synthases (Chs3-Ch7 and Mcs1) localize to the septal region, andfour of them (Chs5, Chs6, Chs7, and Mcs1) also localize to the hyphal tip. Despite thelocalization of Chs5, 6, 7, and Mcs1 to the hyphal tip, they do not seem to be required forhyphal morphology in culture (Weber et al., 2006; see also Ruiz-Herrera et al., 2006). It islikely that the chitin synthases have partially overlapping functions and are able to substitutefor each other when one or two are absent. Deletion of multiple chitin synthases that localizeto the hyphal tip will likely be informative (Weber et al., 2006).
VI. FUTURE DIRECTIONSOur analysis indicates that most of the components of the S. cerevisae machinery for cellpolarity and cell morphogenesis, and the cytoskeleton are conserved in U. maydis. Becausethis analysis was based on S. cerevisiae and S. pombe genes, other components of the cellpolarity machinery specific to Ustilago maydis or the Basidiomycetes remain unidentified.Sequence comparisons with other filamentous fungi, where additional genes required forpolarized growth have been identified, in combination with genetic screens should prove usefulin obtaining a more comprehensive view of the machinery for cell polarity and cellmorphogenesis.
The challenge for the future is not only to identify the machinery for cell polarity, but also tounderstand how the components of the machinery are assembled and rearranged duringpolarized growth, and how these assemblies generate the diversity of forms that characterizethe life cycle of Ustilago maydis.
Banuett et al. Page 12
Fungal Genet Biol. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Supplementary MaterialRefer to Web version on PubMed Central for supplementary material.
ACKNOWLDEGEMENTSWe apologize to colleagues whose work it was not possible to cite due to space limitations. We thank anonymousreviewers for their useful comments. This work was supported by NIH grants SO6 GM63119 and 2SO6 GM063119to FB, and, in part, by CONACYT grant 53191 and SNI fellowship to CRP.
LITERATURE CITEDAkhmanova A, Hoogenraad CC. Microtubule plus-end-tracking proteins: mechanisms and functions.
Curr. Opin. Cell Biol 2005;17:47–54. [PubMed: 15661518]Banuett, F. History of the mating types in Ustilago maydis. In: Heitman, J.; Kronstad, J.; Taylor, J.;
Casselton, LA., editors. Sex in Fungi: Molecular determination and evolutionary implications.Washington, DC: ASM press; 2007. p. 351-375.
Banuett, F. Pathogenic development in Ustilago maydis: A progression of morphological transitions thatresults in tumor formation and teliospore production. In: Osiewacz, HD., editor. Molecular Biologyof Fungal Development. New York, Basel: Marcel Dekker; 2002. p. 349-398.
Banuett F. Signalling in the yeasts: an informational cascade with links to the filamentous fungi.Microbiol. Molec. Biol. Rev 1998;62:249–274. [PubMed: 9618441]
Banuett F. Genetics of Ustilago maydis, a fungal pathogen that induces tumors in maize. Annu. Rev.Genetics 1995;29:179–208. [PubMed: 8825473]
Banuett F, Herskowitz I. Bud morphogenesis and the actin and microtubule cytoskeletons during buddingin the corn smut fungus, Ustilago maydis. Fungal Genet Biol 2002;37:149–170. [PubMed: 12409100]
Banuett F, Herskowitz I. Discrete developmental stages during teliospore formation in the corn smutfungus, Ustilago maydis. Development 1996;122:2965–2976. [PubMed: 8898211]
Barral Y, Mermall V, Mooseker MS, Snyder M. Compartmentalization of the cell cortex by septins isrequired for maintenance of cell polarity in yeast. Mol Cell. 2000 2000;5:841–851.
Bartnicki-Garcia, S. Hyphal tip growth: Outstanding questions. In: Osiewacz, HD., editor. MolecularBiology of Fungal Development. New York, Basel: Marcel Dekker; 2002. p. 29-58.
Bassilana M, Arkowitz RA. Rac1 and Cdc42 have different roles in Candida albicans development.Eukaryot Cell 2006;5:321–329. [PubMed: 16467473]
Bauer Y, Knechtle P, Wendland J, Helfer H, Philippsen P. A Ras-like GTPase is involved in hyphalgrowth guidance in the filamentous fungus Ashbya gossypii. Molec. Biol. Cell 2004;15:4622–4632.[PubMed: 15282338]
Borkovich KA, Alex LA, Yarden O, Freitag M, Turner GE, Read ND, Seiler S, Bell-Pedersen D, PaiettaJ, Plesofsky N, Plamann M, Goodrich-Tanrikulu M, Schulte U, Mannhaupt G, Nargang FE, RadfordA, Selitrennikoff C, Galagan JE, Dunlap JC, Loros JJ, Catcheside D, Inoue H, Aramayo R, PolymenisM, Selker EU, Sachs MS, Marzluf GA, Paulsen I, Davis R, Ebbole DJ, Zelter A, Kalkman ER,O'Rourke R, Bowring F, Yeadon J, Ishii C, Suzuki K, Sakai W, Pratt R. Lessons from the genomesequence of Neurospora crassa: tracing the path from genomic blueprint to multicellular organism.Microbiol. Molec. Biol. Rev 2004;68:1–108. [PubMed: 15007097]
Boyce KJ, Chang H, D'Souza CA, Kronstad JW. An Ustilago maydis septin is required for filamentousgrowth in culture and for full symptom development on maize. Eukaryot Cell 2005;4:2044–2056.[PubMed: 16339722]
Boyce KJ, Hynes MJ, Andrianopoulos A. Control of morphogenesis and actin localization by thePenicillium marneffei RAC homolog. J Cell Sci 2003;1:1249–1260. [PubMed: 12615967]
Boyce KJ, Hynes MJ, Andrianopoulos A. The CDC42 homolog of the dimorphic fungus Penicilliummarneffei is required for correct cell polarization during growth but not development. J Bacteriol2001;183:3447–3457. [PubMed: 11344153]
Brachmann A, Schirawski J, Müller P, Kahmann R. An unusual MAP kinase is required for efficientpenetration of the plant surface by Ustilago maydis. EMBO J 2003;22:2199–2210. [PubMed:12727886]
Banuett et al. Page 13
Fungal Genet Biol. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Cabib E, Drgonova J, Drgon T. Role of small G proteins in yeast cell polarization and wall biosynthesis.Annu. Rev. Biochem 1998;67:307–333. [PubMed: 9759491]
Casamayor A, Snyder M. Bud-site selection and cell polarity in budding yeast. Curr. Opin. Microbiol2002;5:179–186. [PubMed: 11934615]
Chang F, Peter M. Yeasts make their mark. Nature Cell Biol 2003;5:294–299. [PubMed: 12669083]Chang F. Establishment of a cellular axis in fission yeast. Trends Genet 2001;17:273–278. [PubMed:
11335037]Chant J. Cell polarity in yeast. Annu. Rev. Cell Dev. Biol 1999;15:365–391. [PubMed: 10611966]Chen C, Ha YS, Min JY, Memmott SD, Dickman MB. Cdc42 is required for proper growth and
development in the fungal pathogen Colletotrichum trifolii. Eukaryot Cell 2006;5:155–166.[PubMed: 16400178]
Chen C, Dickman MB. Dominant active Rac and dominant negative Rac revert the dominant active Rasphenotype in Colletotrichum trifolii by distinct signaling pathways. Mol Microbiol 2004;51:1493–1507. [PubMed: 14982641]
Christensen JJ. Corn smut caused by Ustilago maydis. Am. Phytopathol. Soc. Monogr 1963;No.2:1–41.Crampin H, Finley K, Gerami-Nejad M, Court H, Galman C, Berman J, Sudbery P. Candida albicans
hyphae have a Spitzenkörper that is distinct form the polarisome found in yeast and pseudohyphae.J. Cell Sci 2005;118:2935–2947. [PubMed: 15976451]
Dong Y, Pruyne D, Bretscher A. Formin-dependent actin assembly is regulated by distinct modes of Rhosignaling in yeast. J. Cell Biol 2003;161:1081–1092. [PubMed: 12810699]
Drees BL, Sundin B, Brazeau E, Caviston JP, Chen GC, Guo W, Kozminski KG, Lau MW, Moskow JJ,Tong A, Schenkman LR, McKenzie A 3rd, Brennwald P, Longtine M, Bi E, Chan C, Novick P,Boone C, Pringle JR, Davis TN. Fields S, Drubin DG. A protein interaction map for cell polaritydevelopment. J Cell Biol 2001;154:549–571. [PubMed: 11489916]
Evangelista M, Zigmond S, Boone C. Formins: signaling effectors for assembly and polarization of actinfilaments. J. Cell Sci 2003;116:2603–2611. [PubMed: 12775772]
Finger FP, Hughes TE, Novick P. Sec3p is a spatial landmark for polarized secretion in budding yeast.Cell 1998;92:559–571. [PubMed: 9491896]
Fink G, Steinberg G. Dynein-dependent motility of microtubules and nucleation sites supportspolarization of the tubulin array in the fungus Ustilago maydis. Molec. Biol. Cell 2006;17:3242–3253. [PubMed: 16672380]
Fink G, Schuchardt I, Colombelli J, Stelzer E, Steinberg G. Dynein-mediated pulling forces drive rapidmitotic spindle elongation in Ustilago maydis. EMBO J 2006;25:4897–4908. [PubMed: 17024185]
Fischer-Parton S, Parton RM, Hickey PC, Dijksterhuis J, Atkinson A, Read ND. Confocal microscopyof FM4-64 as a tool for analysing endocytosis and vesicle trafficking in living fungal hyphae. J.Microsc 2000;198:246–259. [PubMed: 10849201]
Garcerá-Teruel A, Xoconostle-Cázares B, Rosas-Quijano R, Ortiz L, León-Ramírez C, Specht CA,Sentandreu R, Ruiz-Herrera J. Loss of virulence in Ustilago maydis by Umchs6 gene disruption. ResMicrobiol 2004;155:87–97. [PubMed: 14990260]
Gladfelter AS, Pringle JR, Lew DJ. The septin cortex at the yeast mother-bud neck. Curr Opin Microbiol2001;4:681–689. [PubMed: 11731320]
Gold SE, Kronstad JW. Disruption of two genes for chitin synthase in the phytopathogenic fungusUstilago maydis. Mol Microbiol 1994;11:897–902. [PubMed: 8022266]
Gold SE, Bakkeren G, Davies JE, Kronstad JW. Three selectable markers for transformation of Ustilagomaydis. Gene 1994;142:225–230. [PubMed: 7515016]
Guest GM, Lin X, Momany M. Aspergillus nidulans RhoA is involved in polar growth, branching, andcell wall synthesis. Fungal Genet Biol 2004;41:13–22. [PubMed: 14643255]
Harris SD, Read ND, Roberson RW, Shaw B, Seiler S, Plamann M, Momany M. Polarisome meetsSpitzenkörper: microscopy, genetics, and genomics converge. Euk. Cell 2005;4:225–229.
Harris SD. Cell polarity in filamentous fungi: shaping the mold. Intl. Rev. Cytol 2006;251:41–77.Heath, IB. Organization and function of actin in hyphal tip growth. In: Steiger, C.; Baluska, F.; Volkmann,
D.; Barlow, P., editors. Actin: a Dynamic Framework for Mulitple Plant Cell Functions. Dordrecht,Boston & London: Kluwer Academic; 2000. p. 275-300.
Banuett et al. Page 14
Fungal Genet Biol. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Hodge T, Cope MJ. A myosin family tree. J Cell Sci 2000;113:3353–3354. [PubMed: 10984423]Holliday, R. Handbook of Genetics. In: King, RC., editor. Ustilago maydis. vol 1. New York: Plenum;
1974. p. 575-595.Jacobs CW, Mattichak SJ, Knowles JF. Budding patterns during the cell cycle of the maize smut pathogen
Ustilago maydis. Can. J. Bot 1994;72:1675–1680.Jaffe AB, Hall A. Rho GTPases: Biochemistry and Biology. Annu. Rev. Cell Dev. Biol 2005;21:347–
369. [PubMed: 16212499]Kadota J, Yamamoto T, Yoshiuchi S, Bi E, Tanaka K. Septin ring assembly requires concerted action of
polarisome components, a PAK kinase Cla4p, and the actin cytoskeleton in Saccharomycescerevisiae. Mol Biol Cell 2004;15:5329–5345. [PubMed: 15371547]
Karpova TS, McNally JG, Moltz SL, Cooper JA. Assembly and function of the actin cytoskeleton ofyeast: relationships between cables and patches. J. Cell Biol 1998;142:1501–1517. [PubMed:9744880]
Klis FM, Boorsma A, De Groot PW. Cell wall construction in Saccharomyces cerevisiae. Yeast2006;23:185–202. [PubMed: 16498706]
Klosterman SJ, Perlin MH, García-Pedrajas M, Covert SF, Gold SE. Genetics of morphogenesis andpathogenic development of Ustilago maydis. Adv. Genet 2007;57:1–47. [PubMed: 17352901]
Knechtle P, Dietrich F, Philippsen P. Maximal growth potential depends on the polarisome componentAgSpa2 in the filamentous fungus Ashbya gossypii. Molec. Biol. Cell 2003;14:4140–4154. [PubMed:12937275]
Konzack S, Rischitor PE, Enke C, Fischer R. The role of the kinesin motor KipA in microtubuleorganization and polarized growth of Aspergillus nidulans. Mol Biol Cell 2005;16:497–506.[PubMed: 15563609]
Latge JP. The cell wall: a carbohydrate armour for the fungal cell. Molec. Microbiol 2007;66:279–290.[PubMed: 17854405]
Lee W-L, OBerle JR, Cooper JA. The role of the lissencepahly protein Pac1 during nuclear migration inbudding yeast. J. Cell Biol 2003;160:355–364. [PubMed: 12566428]
Lee IH, Kumar S, Plamann M. Null mutants of the Neurospora actin-related protein 1 pointed-endcomplex show distinct phenotypes. Mol Biol Cell 2001;12:2195–2206. [PubMed: 11452013]
Leeder AC, Turner G. Characterization of Aspergillus nidulans polarisome component BemA. FungalGenet Biol. Epub. 2007
Lenz JH, Schuhardt I, Straube A, Steinberg G. A dynein loading zone for retrograde endosome motilityat microtubule plus ends. EMBO J 2006;25:2275–2286. [PubMed: 16688221]
Leveleki L, Mahlert M, Sandrock B, Bölker M. The PAK family kinase Cla4 is required for budding andmorphogenesis in Ustilago maydis. Mol Microbiol 2004;54:396–406. [PubMed: 15469512]
Longtine MS, Bi E. Regulation of septin organization and function in yeast. Trends Cell Biol2003;13:403–409. [PubMed: 12888292]
Lopez-Franco R, Bracker CE. Diversity and dynamics of the Sptizenkörper in growing hyhal tips ofhigher fungi. Protoplasma 1996;195:90–111.
Mahlert M, Leveleki L, Hlubek A, Sandrock B, Bölker M. Rac1 and Cdc42 regulate hyphal growth andcytokinesis in the dimorphic fungus Ustilago maydis. Mol Microbiol 2006;59:567–578. [PubMed:16390450]
Miller RK, Rose MD. Kar9p is a novel cortical protein required for cytoplasmic microtubule orientationin yeast. J Cell Biol 1998;140:377–930. [PubMed: 9442113]
Moseley JB, Goode BL. The yeast actin cytoskeleton: from cellular function to biochemical mechanism.Microbiol Mol Biol Rev 2006;70:605–645. [PubMed: 16959963]
Ni L, Snyder M. A genomic study of the bipolar bud site selection pattern in Saccharomycescerevisiae. Mol. Biol. Cell 2001;12:2147–2170. [PubMed: 11452010]
O'Donnell KL, McLaughlin DJ. Postmeiotic mitosis, basidiospore development, and septation in Ustilagomaydis. Mycologia 1984;76:486–502.
Pan F, Malmberg RL, Momany M. Analysis of septins across kingdoms reveals orthology and new motifs.BMC Evol Biol 2007;7:103–119. [PubMed: 17601340]
Banuett et al. Page 15
Fungal Genet Biol. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Park HO, Bi E. Central roles of small GTPases in the development of cell polarity in yeast and beyond.Microbiol Mol Biol Rev 2007;71:48–96. [PubMed: 17347519]
Philippsen P, Kaufmann A, Schmitz H-P. Homologues of yeast polarity genes control the developmentof multinucleated hyphae in Ashbya gossypii. Curr. Opin. Microbiol 2005;8:1–8. [PubMed:15694849]
Plamann M, Minke PF, Tinsly JH, Bruno K. Cytoplasmic dynein and centractin are required for normalnuclear distribution in filamentous fungi. J. Cell Biol 1994;127:139–149. [PubMed: 7929559]
Pruyne D, Evangelista M, Yang C, Bi E, Zigmond S, Bretscher A, Boone C. Role of formins in actinassembly: nucleation and barbed-end association. Science 2002;297:612–615. [PubMed: 12052901]
Pruyne D, Bretscher A. Polarization of cell growth in yeast. I. Establishment and maintenance of polaritystates. J. Cell Sci 2000a;113:365–375. [PubMed: 10639324]
Pruyne D, Bretscher A. Polarization of cell growth in yeast. II. The role of the cortical actin cytoskeleton.2000. J. Cell Sci 2000b;113:571–585. [PubMed: 10652251]
Reynaga-Peña CG, Gierz G, Bartnicki-Garcia S. Analysis of the role of the Spitzenkörper in fungalmorphogenesis by computer simulation of apical branching in Aspergillus niger. Proc Natl Acad SciUSA 1997;94:9096–9101. [PubMed: 9256441]
Riquelme M, Bartnicki-García S, González-Prieto JM, Sánchez-León E, Verdín-Ramos JA, Beltrán-Aguilar A, Freitag M. Spitzenkörper localization and intracellular traffic of green fluoresenct protein-labeled CHS-3 and CHS_6 chitin synthases in living hyphae of Neurospora crassa. Eukaryot. Cell2007;6:1853–1864. [PubMed: 17644657]
Riquelme M, Gierz G, Bartnicki-Garcia S. Dynein and dynactin deficiencies affect the formation andfunction of the Spitzenkörper and distort hyphal morphogenesis in Neurospora crassa. Microbiol2000;146:1743–1752.
Rolke Y, Tudzynski P. The small GTPase Rac and the p21-activated kinase Cla4 in Clavicepspurpurea: interaction and impact on polarity, development and pathogenicity. Mol Microbiol2008;68:405–423. [PubMed: 18284596]
Ruiz-Herrera J, Xoconostle-Cázares B, Reynaga-Peña CG, León-Ramírez C, Cárabez-Trejo A.Immunolocalization of chitin synthases in the phytopathogenic dimorphic fungus Ustilago maydis.FEMS Yeast Res 2006;6:999–1009. [PubMed: 17042749]
Ruiz-Herrera J, León-Ramírez C, Cabrera-Ponce JL, Martínez-Espinoza AD, Herrera-Estrella L.Completion of the sexual cycle and demonstration of genetic recombination in Ustilago maydis invitro. Molec. Gen. Genet 1999;262:468–472. [PubMed: 10589834]
Rupes I, Mao WZ, Astrom H, Raudaskoski M. Effects of nocodazole and brefeldin A on microtubulecytoskeleton and membrane organization in homobasidiomycte Schizophyllum commune.Protoplasma 1995;185:212–221.
Sagot I, Klee SK, Pellman D. Yeast formins regulate cell polarity by controlling the assembly of actincables. Nature Cell Biol 2001;4:42–50. [PubMed: 11740491]
Sawin KE, Nurse P. Regulation of cell polarity by microtubules in fission yeast. J. Cell Biol1998;142:457–471. [PubMed: 9679144]
Scheffer J, Chen C, Heidrich P, Dickman MB, Tudzynski P. A CDC42 homologue in Clavicepspurpurea is involved in vegetative differentiation and is essential for pathogenicity. Eukaryot Cell2005;4:1228–1238. [PubMed: 16002649]
Scherer M, Heimel K, Starke V, Kämper J. The Clp1 protein is required for clamp formation andpathogenic development in Ustilago maydis. Plant Cell 2006 2006;18:2388–2401.
Schmitz H-P, Kaufmann A, Köhli M, Laissue PF, Philippsen P. From function to shape: a novel role ofa formin in fungal morphogenesis of the fungus Ashbya gossypii. Molec. Biol. Cell 2006;17:130–145. [PubMed: 16236798]
Schuchardt I, Assmann D, Thines E, Schuberth C, Steinberg G. Myosin-V, Kinesin-1, and Kinesin-3cooperate in hyphal growth of the fungus Ustilago maydis. Mol Biol Cell 2005;16:5191–5201.[PubMed: 16120650]
Schroer TA. Dynactin. Annu. Rev. Cell Dev. Biol 2004;20:759–779. [PubMed: 15473859]Seiler S, Plamann M. The genetic basis of cellular morphogenesis in the filamentous fungus Neurospora
crassa. Molec. Biol. Cell 2003;14:4352–4364. [PubMed: 12960438]
Banuett et al. Page 16
Fungal Genet Biol. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Seiler S, Plamann M, Schliwa M. Kinesin and dynein mutants provide novel insights into the roles ofvesicle traffic during cell morphogenesis in Neurospora. Curr Biol 1999;9:779–785. [PubMed:10469561]
Sharpless KE, Harris SD. Functional characterization and localization of the Aspergillus nidulans forminSepA. Molec. Biol. Cell 2002;13:469–479. [PubMed: 11854405]
Sheeman B, Carvalho P, Sagot I, Geiser J, Kho D, Hoyt MA, Pellman D. Determinants of S. cerevisiaedynein localization and activation: implications for the mechanism of spindle positioning. Curr Biol2003;13:364–372. [PubMed: 12620184]
Smith DG, García-Pedrajas MD, Hong W, Yu Z, Gold SE, Perlin MH. An ste20 homologue in Ustilagomaydis plays a role in mating and pathogenicity. Eukaryot Cell 2004;3:180–189. [PubMed:14871948]
Snetselaar KM, Mim CW. Light and electron microscopy of Ustilago maydis hyphae in maize. Mycol.Res 1994;98:347–355.
Steinberg G, Wedlich-Söldner R, Brill M, Schulz I. Microtubules in the fungal pathogen Ustilagomaydis are highly dynamic and determine cell polarity. J. Cell Sci 2001;114:609–622. [PubMed:11171329]
Straube A, Brill M, Oakley BR, Horio T, Steinberg G. Microtubule organization requires cell cycle-dependent nucleation at dispersed cytoplasmic sites: polar and perinuclear microtubule organizingcenters in the plant pathogen Ustilago maydis. Molec. Biol. Cell 2003;14:642–657. [PubMed:12589060]
Straube A, Enard W, Berner A, Wedlich-Söldner R, Kahmann R, Steinberg G. A split motor domain ina cytoplasmic dynein. EMBO J 2001;20:5091–6100. [PubMed: 11566874]
Taheri-Talesh N, Horio T, Araujo-Bazán L, Dou X, Espeso EA, Peñalva MA, Osmani SA, Oakley BR.The tip growth apparatus of Aspergillus nidulans. Mol Biol Cell 2008;19:1439–1449. [PubMed:18216285]
Takeshita N, Higashitsuji Y, Konzack S, Fischer R. Apical sterol-rich membranes are essential forlocalizing cell end markers that determine growth directionality in the filamentous fungus Aspergillusnidulans. Mol Biol Cell 2008;19:339–351. [PubMed: 18003978]
Takizawa PA, DeRisi JL, Wilhelm JE, Vale RD. Plasma membrane compartmentalization in yeast bymessenger RNA transport and a septin diffusion barrier. Science 2000;290:341–344. [PubMed:11030653]
TerBush DR, Maurice T, Roth D, Novick P. The Exocyst is a multiprotein complex required forexocytosis in Saccharomyces cerevisiae. EMBO J 1996;15:6483–6494. [PubMed: 8978675]
Torralba S, Raudaskoski M, Pedregosa AM. Effects of methyl benzimidazole-2-yl carbamate onmicrotubule and actin cytoskeleton in Aspergillus nidulans. Protoplasma 1998;202:54–64.
Torralba S, Raudaskoski M, Pedregosa AM, Laborda F. Effect of cytochalasin A on apical growth, actincytoskeleton organization and enzyme secretion in Aspergillus nidulans. Microbiology 1998;144:45–53. [PubMed: 9537763]
Virag A, Lee MP, Si H, Harris SD. Regulation of hyphal morphogenesis by cdc42 and rac1 homologuesin Aspergillus nidulans. Mol Microbiol 2007;66:1579–1596. [PubMed: 18005099]
Virag A, Harris SD. Functional characterization of Aspergillus nidulans homologues of Saccharomycescerevisiae Spa2 and Bud6. Euk. Cell 2006;5:881–895.
Virag A, Griffiths AJ. A mutation in the Neurospora crassa actin gene results in multiple defects in tipgrowth and branching. Fungal Genet Biol 2004;41:213–225. [PubMed: 14732267]
Wallar BJ, Alberts AS. The formins: active scaffolds that remodel the cytoskeleton. Trends Cell Biol2003;13:435–446. [PubMed: 12888296]
Weber I, Assmann D, Thines E, Steinberg G. Polar localizing class V myosin chitin synthases are essentialduring early plant infection in the plant pathogenic fungus Ustilago maydis. Plant Cell 2006;18:225–242. [PubMed: 16314447]
Weber I, Gruber C, Steinberg G. A class V myosin required for mating, hyphal growth, and pathogenicityin the dimorphic plant pathogn Ustilago maydis. Plant Cell 2003;15:2826–2842. [PubMed:14615599]
Wedlich-Söldner R, Schulz I, Straube A, Steinberg G. Dynein supports motility of endoplasmic reticulumin the fungus Ustilago maydis. Mol Biol Cell 2002;13:965–977. [PubMed: 11907275]
Banuett et al. Page 17
Fungal Genet Biol. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Weinzierl G, Leveleki L, Hassel A, Kost G, Wanner G, Bölker M. Regulation of cell separation in thedimorphic fungus Ustilago maydis. Molec. Microbiol 2002;45:219–231. [PubMed: 12100561]
Wendland J. Analysis of the landmark protein Bud3 of Ashbya gossypii reveals a novel role in septumconstruction. EMBO Rep 2003;4:200–204. [PubMed: 12612612]
Wendland J, Philippsen P. Cell polarity and hyphal morphogenesis are controlled by multiple rho-proteinmodules in the filamentous ascomycete Ashbya gossypii. Genetics 2001;157:601–610. [PubMed:11156982]
Westfall PJ, Momany M. Aspergillus nidulans septin AspB plays pre- and postmitotic roles in septum,branch, and conidiophore development. Mol Biol Cell 2002;13:110–118. [PubMed: 11809826]
Winter DC, Choe EY, Li R. Genetic dissection of the budding yeast Arp2/3 complex: a comparison ofthe in vivo structural roles of individual subunits. Proc. Natl. Acad. Sci 1999;96:7288–7293.[PubMed: 10377407]
Xiang X, Plamann M. Cytoskeleton and motor proteins in filamentous fungi. Curr Opin Microbiol2003;6:628–633. [PubMed: 14662360]
Yaar L, Mevarech M, Koltin Y. A Candida albicans RAS-related gene (CaRSR1) is involved in budding,cell morphogenesis and hypha development. Microbiology 1997;143:3033–3044. [PubMed:9308185]
Banuett et al. Page 18
Fungal Genet Biol. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
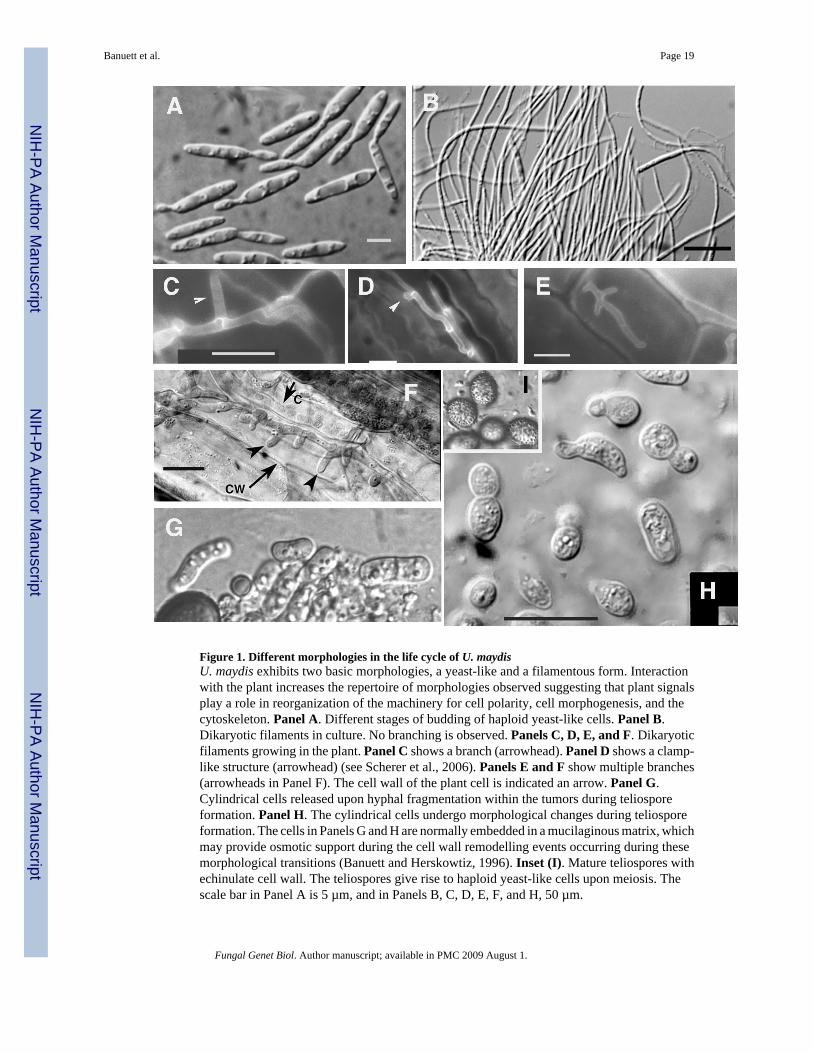
Figure 1. Different morphologies in the life cycle of U. maydisU. maydis exhibits two basic morphologies, a yeast-like and a filamentous form. Interactionwith the plant increases the repertoire of morphologies observed suggesting that plant signalsplay a role in reorganization of the machinery for cell polarity, cell morphogenesis, and thecytoskeleton. Panel A. Different stages of budding of haploid yeast-like cells. Panel B.Dikaryotic filaments in culture. No branching is observed. Panels C, D, E, and F. Dikaryoticfilaments growing in the plant. Panel C shows a branch (arrowhead). Panel D shows a clamp-like structure (arrowhead) (see Scherer et al., 2006). Panels E and F show multiple branches(arrowheads in Panel F). The cell wall of the plant cell is indicated an arrow. Panel G.Cylindrical cells released upon hyphal fragmentation within the tumors during teliosporeformation. Panel H. The cylindrical cells undergo morphological changes during teliosporeformation. The cells in Panels G and H are normally embedded in a mucilaginous matrix, whichmay provide osmotic support during the cell wall remodelling events occurring during thesemorphological transitions (Banuett and Herskowtiz, 1996). Inset (I). Mature teliospores withechinulate cell wall. The teliospores give rise to haploid yeast-like cells upon meiosis. Thescale bar in Panel A is 5 µm, and in Panels B, C, D, E, F, and H, 50 µm.
Banuett et al. Page 19
Fungal Genet Biol. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Banuett et al. Page 20Ta
ble
1U
. may
dis h
omol
ogue
s of t
he S
. cer
evisi
ae m
achi
nery
for
cell
pola
rity
, cel
l mor
phog
enes
is, a
nd th
e cy
tosk
elet
onS.
cer
evis
iae
gene
s in
volv
ed in
cel
l pol
arity
, cel
l mor
phog
enes
is, a
nd th
e cy
tosk
elet
on (c
olum
n 2)
wer
e us
ed to
pro
be th
e U
. may
dis
geno
me
with
wuB
last
at
the
MIP
S U
stila
go d
atab
ase
MU
MD
B (
http
://m
ips.g
sf.d
e/ge
nre/
proj
/ust
ilago
/), a
nd t
he U
stila
go m
aydi
sD
atab
ase a
t the
Bro
ad In
stitu
te (h
ttp://
ww
w.b
road
.mit.
edu/
anno
tatio
n/ge
nom
e/us
tilag
o_m
aydi
s/H
ome.
htm
l). W
e als
o us
ed B
last
at N
CB
I(h
ttp://
ww
w.n
cbi.n
lm.n
ih.g
ov/b
last
) (A
ltsch
ul e
t al.,
199
7). T
he g
enes
iden
tifie
d in
U. m
aydi
s wer
e th
en u
sed
to se
arch
for h
omol
ogue
sin
the
Bas
idio
myc
ota
and
Asc
omyc
ota,
and
in o
ther
euk
aryo
tes
usin
g th
e SI
MA
P to
ol a
t MIP
S (s
ee F
ig.1
S, s
uppl
emen
tary
mat
eria
l).Th
e so
urce
of
S. c
erev
isia
e ge
ne p
rodu
cts
was
Dre
es e
t al
., 20
01;
Pruy
ne a
nd B
rets
cher
, 20
00;
and
GO
ann
otat
ion
at t
heSa
ccha
rom
yces
gen
ome
data
base
(http
://w
ww
.yea
stge
nom
e.or
g/).
The
syst
emat
ic n
ame
for S
. cer
evis
iae g
enes
(col
umn
3) w
as o
btai
ned
from
(http
://w
ww
.yea
stge
nom
e.or
g/).
U. m
aydi
s gen
e nu
mbe
rs (c
olum
n 4)
wer
e ob
tain
ed fr
om (h
ttp://
mip
s.gsf
.de/
genr
e/pr
oj/u
stila
go/)
afte
r BLA
ST a
naly
sis.
E va
lues
(col
umn
5) in
dica
ted
are
as o
btai
ned
from
wuB
last
ana
lysi
s at M
IPS.
Oth
er p
aram
eter
s wer
e al
so u
sed
in o
ur B
last
ana
lysi
s, in
par
ticul
ar in
the
com
paris
on a
cros
s diff
eren
t fun
gal g
roup
s and
oth
er e
ukar
yote
s (da
ta n
ot sh
own
but a
vaila
ble
upon
requ
est).
The
last
col
umn
prov
ides
the
func
tion
of th
e S.
cer
evis
iae
gene
s (ht
tp://
ww
w.y
east
geno
me.
org/
). Th
e lis
t of g
enes
in th
isTa
ble
is n
ot c
ompr
ehen
sive
; it d
oes n
ot in
clud
e m
any
gene
s inv
olve
d in
cyt
okin
esis
, cel
l sep
arat
ion,
secr
etio
n, a
nd c
ell c
ycle
that
wer
eus
ed in
the
anal
ysis
show
n in
Fig
s 1S
(sup
plem
enta
ry m
ater
ial).
CELL
PO
LARI
TY/C
ELL
MO
RPH
OG
ENES
ISSc
gen
eSy
stem
atic
nam
eU
. may
dis M
IPS#
E v
alue
Func
tion
in S
. cer
evisi
ae1
BAG
7 Y
OR
134W
02
902
3.3e
-48
Rho
GA
P; fu
nctio
nally
rela
ted
to S
ac7
2 BE
M1
YB
R20
0W
1093
4 3.
7e-4
4 Es
tabl
ishm
ent o
f cel
l pol
arity
. Sca
ffol
d fo
r diff
eren
t pro
tein
com
plex
es
3 BE
M2
YER
155C
01
161
3.4e
-42
Rho
GA
P 4
BEM
3 Y
PL11
5C
1185
2 8.
9e-2
4 G
AP
for C
dc42
and
Rho
1 5
BEM
4 Y
PL16
1C
no
hits
In
tera
cts w
ith C
dc42
and
Rho
1. B
ud e
mer
genc
e.
6 BN
I4
YN
L233
W
no
hits
R
equi
red
for l
ocal
izat
ion
of C
HSI
II to
bud
nec
k 7
BOI1
Y
BL0
85W
03
037
1.7e
-23
Bem
1-bi
ndin
g pr
otei
n. P
olar
ized
gro
wth
8
BOI2
Y
ER11
4C
0303
7 1.
8e-1
9 B
em1-
bind
ing
prot
ein.
Pol
ariz
ed g
row
th
9 BU
D1/
RSR1
Y
GR
152C
11
909
7.8e
-49
Bud
site
sele
ctio
n. R
AS
supe
rfam
ily p
rote
in
10
BUD
2 Y
KLO
92C
01
282
6.9e
-10
Bud
site
sele
ctio
n. G
AP
for B
ud1/
Rsr
1 11
BU
D3
YC
LO14
W
no
hits
B
ud si
te se
lect
ion.
Axi
al la
ndm
ark
12
BUD
4 Y
JRO
92W
no h
its
Bud
site
sele
ctio
n. A
xial
land
mar
k 13
BU
D5
YC
RO
38C
11
476
1.3e
-12
Bud
site
sele
ctio
n. G
EF fo
r Bud
1/R
sr1
14
BUD
6/AI
P3
YLR
319C
11
657
1.6e
-70
Pola
risom
e co
mpo
nent
; int
erac
ts w
ith fo
rmin
and
act
in
15
BUD
7 Y
OR
299W
04
959
5.2e
-62
Expo
rt of
Chs
3 fr
om G
olgi
to p
lasm
a m
embr
ane.
Bip
olar
bud
ding
. 16
BU
D8
YLR
353W
no h
its
Bud
site
sele
ctio
n. B
ipol
ar la
ndm
ark
17
BUD
9 Y
GR
041W
no h
its
Bud
site
sele
ctio
n. B
ipol
ar la
ndm
ark
18
BU
D10
/AX
L2
YIL
140W
01
289
2.8e
-24
Bud
site
sele
ctio
n 19
BU
D13
Y
GL1
74W
00
191
1.1e
-10
Bud
site
sele
ctio
n. B
ipol
ar b
uddi
ng.
20
BUD
14
YA
R01
4C
0101
2 3.
4e-1
4 B
ud si
te se
lect
ion.
Bip
olar
bud
ding
. 21
BU
D15
/ED
E1
YB
L047
C
0553
3 4.
8e-6
1 B
ud si
te se
lect
ion.
Bip
olar
bud
ding
. 22
BU
D16
Y
EL02
9C
1031
0 1.
1e-4
2 B
ud si
te se
lect
ion.
Bip
olar
bud
ding
. 23
BU
D17
Y
NR
027W
10
310
1.1e
-41
Bud
site
sele
ctio
n. B
ipol
ar b
uddi
ng.
24
BUD
20
YLR
074C
04
625
2.5e
-11
Bud
site
sele
ctio
n. B
ipol
ar b
uddi
ng.
25
BUD
21
YO
R07
8W
no
hits
B
ud si
te se
lect
ion.
Bip
olar
bud
ding
. 26
BU
D22
Y
MR
014W
01
709
4.8e
-13
Bud
site
sele
ctio
n. B
ipol
ar b
uddi
ng.
27
BUD
23
YC
R04
7C
0155
1 8.
4e-6
8 B
ud si
te se
lect
ion.
Bip
olar
bud
ding
. 28
BU
D25
Y
ER01
4C-A
no h
its
Bud
site
sele
ctio
n. B
ipol
ar b
uddi
ng.
29
BUD
27
YFL
023W
no h
its
Bud
site
sele
ctio
n. B
ipol
ar b
uddi
ng.
30
BUD
31
YC
R06
3W
1005
7 1.
0e-4
6 B
ud si
te se
lect
ion.
Bip
olar
bud
ding
. 31
BU
D32
Y
GR
262C
12
089
2.3e
-20
Bud
site
sele
ctio
n. B
ipol
ar b
uddi
ng.
32
CD
C3
YLR
314C
10
503
1.1e
-117
Se
ptin
Fungal Genet Biol. Author manuscript; available in PMC 2009 August 1.
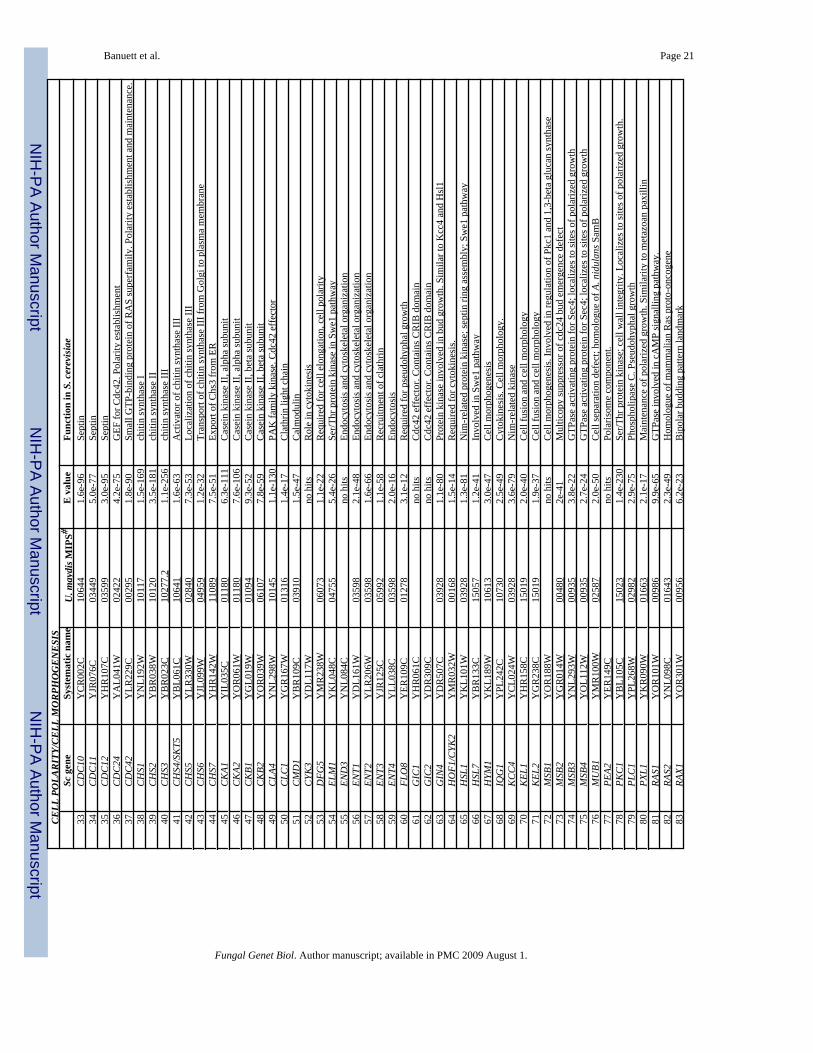
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Banuett et al. Page 21CE
LL P
OLA
RITY
/CEL
L M
ORP
HO
GEN
ESIS
Sc g
ene
Syst
emat
ic n
ame
U. m
aydi
s MIP
S#E
val
ueFu
nctio
n in
S. c
erev
isiae
33
CD
C10
Y
CR
002C
10
644
1.6e
-96
Sept
in
34
CD
C11
Y
JR07
6C
0344
9 5.
0e-7
7 Se
ptin
35
C
DC
12
YH
R10
7C
0359
9 3.
0e-9
5 Se
ptin
36
C
DC
24
YA
L041
W
0242
2 4.
2e-7
5 G
EF fo
r Cdc
42. P
olar
ity e
stab
lishm
ent
37
CD
C42
Y
LR22
9C
0029
5 1.
8e-9
0 Sm
all G
TP-b
indi
ng p
rote
in o
f RA
S su
perf
amily
. Pol
arity
est
ablis
hmen
t and
mai
nten
ance
. 38
C
HS1
Y
NL1
92W
10
117
1.5e
-169
ch
itin
synt
hase
I 39
C
HS2
Y
BR
038W
10
120
3.5e
-181
ch
itin
synt
hase
II
40
CH
S3
YB
R02
3C
1027
7.2
1.1e
-256
ch
itin
synt
hase
III
41
CH
S4/S
KT5
Y
BL0
61C
10
641
1.6e
-63
Act
ivat
or o
f chi
tin sy
ntha
se II
I 42
C
HS5
Y
LR33
0W
0284
0 7.
3e-5
3 Lo
caliz
atio
n of
chi
tin sy
ntha
se II
I 43
C
HS6
Y
JL09
9W
0495
9 1.
2e-3
2 Tr
ansp
ort o
f chi
tin sy
ntha
se II
I fro
m G
olgi
to p
lasm
a m
embr
ane
44
CH
S7
YH
R14
2W
1108
9 7.
5e-5
1 Ex
port
of C
hs3
from
ER
45
C
KA1
Y
IL03
5C
0118
0 6.
3e-1
11
Cas
ein
kina
se II
, alp
ha su
buni
t 46
C
KA2
Y
OR
061W
01
180
7.6e
-106
C
asei
n ki
nase
II, a
lpha
subu
nit
47
CK
B1
YG
L019
W
0109
4 9.
3e-5
2 C
asei
n ki
nase
II, b
eta
subu
nit
48
CK
B2
YO
R03
9W
0610
7 7.
8e-5
9 C
asei
n ki
nase
II, b
eta
subu
nit
49
CLA
4 Y
NL2
98W
10
145
1.1e
-130
PA
K fa
mily
kin
ase.
Cdc
42 e
ffec
tor
50
CLC
1 Y
GR
167W
01
316
1.4e
-17
Cla
thrin
ligh
t cha
in
51
CM
D1
YB
R10
9C
0391
0 1.
5e-4
7 C
alm
odul
in
52
CYK
3 Y
DL1
17W
no h
its
Rol
e in
cyt
okin
esis
53
D
FG5
YM
R23
8W
0607
3 1.
1e-2
2 R
equi
red
for c
ell e
long
atio
n, c
ell p
olar
ity
54
ELM
1 Y
KL0
48C
04
755
5.4e
-26
Ser/T
hr p
rote
in k
inas
e in
Sw
e1 p
athw
ay
55
END
3 Y
NL0
84C
no h
its
Endo
cyto
sis a
nd c
ytos
kele
tal o
rgan
izat
ion
56
ENT1
Y
DL1
61W
03
598
2.1e
-48
Endo
cyto
sis a
nd c
ytos
kele
tal o
rgan
izat
ion
57
ENT2
Y
LR20
6W
0359
8 1.
6e-6
6 En
docy
tosi
s and
cyt
oske
leta
l org
aniz
atio
n 58
EN
T3
YJR
125C
05
992
1.1e
-58
Rec
ruitm
ent o
f cla
thrin
59
EN
T4
YLL
038C
03
598
2.0e
-16
Endo
cyto
sis
60
FLO
8 Y
ER10
9C
0127
8 3.
1e-1
2 R
equi
red
for p
seud
ohyp
hal g
row
th
61
GIC
1 Y
HR
061C
no h
its
Cdc
42 e
ffec
tor.
Con
tain
s CR
IB d
omai
n 62
G
IC2
YD
R30
9C
no
hits
C
dc42
eff
ecto
r. C
onta
ins C
RIB
dom
ain
63
GIN
4 Y
DR
507C
03
928
1.1e
-80
Prot
ein
kina
se in
volv
ed in
bud
gro
wth
. Sim
ilar t
o K
cc4
and
Hsl
1 64
H
OF1
/CYK
2 Y
MR
032W
00
168
1.5e
-14
Req
uire
d fo
r cyt
okin
esis
. 65
H
SL1
YK
L101
W
0392
8 1.
3e-8
1 N
im-r
elat
ed p
rote
in k
inas
e; se
ptin
ring
ass
embl
y; S
we1
pat
hway
66
H
SL7
YB
R13
3C
1505
7 1.
2e-4
1 In
volv
ed in
Sw
e1 p
athw
ay
67
HYM
1 Y
KL1
89W
10
613
3.0e
-47
Cel
l mor
phog
enes
is
68
IQG
1 Y
PL24
2C
1073
0 2.
5e-4
9 C
ytok
ines
is. C
ell m
orph
olog
y.
69
KC
C4
YC
L024
W
0392
8 3.
6e-7
9 N
im-r
elat
ed k
inas
e 70
K
EL1
YH
R15
8C
1501
9 2.
0e-4
0 C
ell f
usio
n an
d ce
ll m
orph
olog
y 71
K
EL2
YG
R23
8C
1501
9 1.
9e-3
7 C
ell f
usio
n an
d ce
ll m
orph
olog
y 72
M
SB1
YO
R18
8W
no
hits
C
ell m
orph
ogen
esis
. Inv
olve
d in
regu
latio
n of
Pkc
1 an
d 1,
3-be
ta g
luca
n sy
ntha
se
73
MSB
2 Y
GR
014W
00
480
2e-4
1 M
ultic
opy
supp
ress
or o
f cdc
24 b
ud e
mer
genc
e de
fect
74
M
SB3
YN
L293
W
0093
5 3.
8e-2
2 G
TPas
e ac
tivat
ing
prot
ein
for S
ec4;
loca
lizes
to si
tes o
f pol
ariz
ed g
row
th
75
MSB
4 Y
OL1
12W
00
935
2.7e
-24
GTP
ase
activ
atin
g pr
otei
n fo
r Sec
4; lo
caliz
es to
site
s of p
olar
ized
gro
wth
76
M
UB1
Y
MR
100W
02
587
2.0e
-50
Cel
l sep
arat
ion
defe
ct; h
omol
ogue
of A
. nid
ulan
s Sam
B
77
PEA2
Y
ER14
9C
no
hits
Po
laris
ome
com
pone
nt.
78
PKC
1 Y
BL1
05C
15
023
1.4e
-230
Se
r/Thr
pro
tein
kin
ase;
cel
l wal
l int
egrit
y. L
ocal
izes
to si
tes o
f pol
ariz
ed g
row
th.
79
PLC
1 Y
PL26
8W
0298
2 2.
9e-7
5 Ph
osph
olip
ase
C. P
seud
ohyp
hal g
row
th
80
PXL1
Y
KR
090W
01
663
2.1e
-17
Mai
nten
ance
of p
olar
ized
gro
wth
. Sim
ilarit
y to
met
azoa
n pa
xilli
n 81
RA
S1
YO
R10
1W
0098
6 9.
9e-6
5 G
TPas
e in
volv
ed in
cA
MP
sign
allin
g pa
thw
ay.
82
RAS2
Y
NL0
98C
01
643
2.3e
-49
Hom
olog
ue o
f mam
mal
ian
Ras
pro
to-o
ncog
ene
83
RAX1
Y
OR
301W
00
956
6.2e
-23
Bip
olar
bud
ding
pat
tern
land
mar
k
Fungal Genet Biol. Author manuscript; available in PMC 2009 August 1.
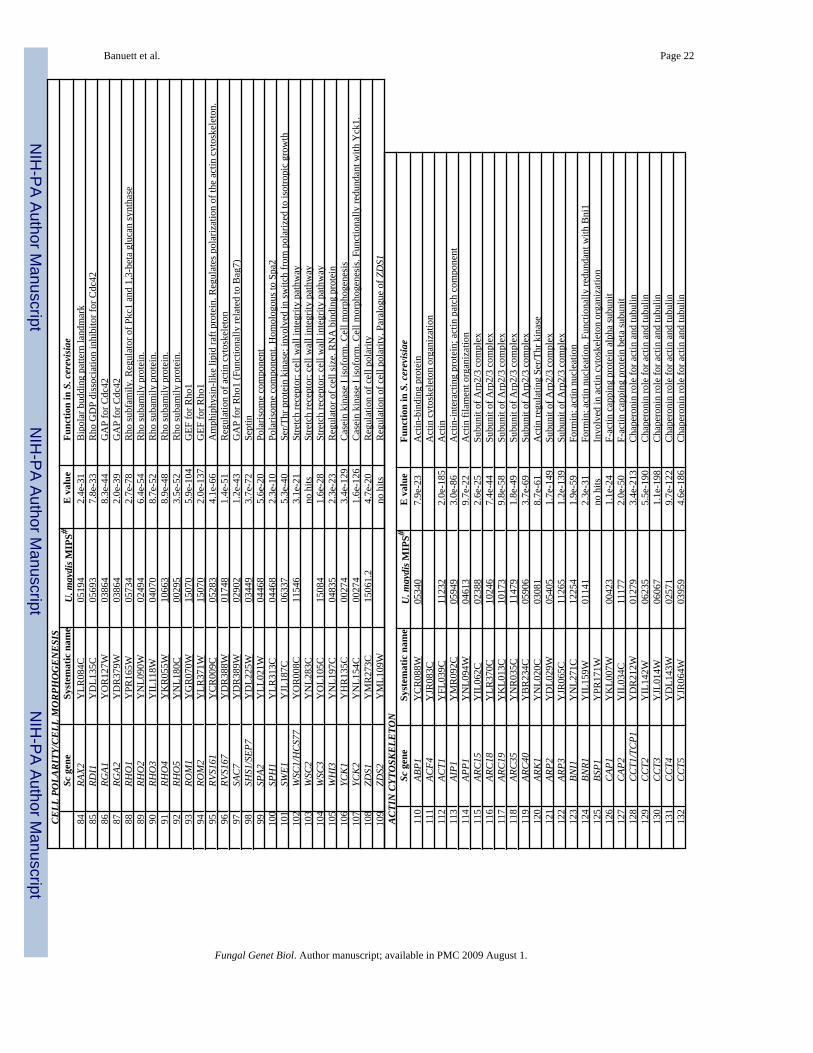
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Banuett et al. Page 22CE
LL P
OLA
RITY
/CEL
L M
ORP
HO
GEN
ESIS
Sc g
ene
Syst
emat
ic n
ame
U. m
aydi
s MIP
S#E
val
ueFu
nctio
n in
S. c
erev
isiae
84
RAX2
Y
LR08
4C
0519
4 2.
4e-3
1 B
ipol
ar b
uddi
ng p
atte
rn la
ndm
ark
85
RDI1
Y
DL1
35C
05
693
7.8e
-33
Rho
GD
P di
ssoc
iatio
n in
hibi
tor f
or C
dc42
86
RG
A1
YO
R12
7W
0386
4 8.
3e-4
4 G
AP
for C
dc42
87
RG
A2
YD
R37
9W
0386
4 2.
0e-3
9 G
AP
for C
dc42
88
RH
O1
YPR
165W
05
734
2.7e
-78
Rho
subf
amily
. Reg
ulat
or o
f Pkc
1 an
d 1,
3-be
ta g
luca
n sy
ntha
se
89
RHO
2 Y
NL0
90W
02
494
6.4e
-54
Rho
suba
mily
pro
tein
. 90
RH
O3
YIL
118W
04
070
8.7e
-52
Rho
suba
mily
pro
tein
. 91
RH
O4
YK
R05
5W
1066
3 8.
9e-4
8 R
ho su
bam
ily p
rote
in.
92
RHO
5 Y
NL1
80C
00
295
3.5e
-52
Rho
suba
mily
pro
tein
. 93
RO
M1
YG
R07
0W
1507
0 5.
9e-1
04
GEF
for R
ho1
94
ROM
2 Y
LR37
1W
1507
0 2.
0e-1
37
GEF
for R
ho1
95
RVS1
61
YC
R00
9C
0528
3 4.
1e-6
6 A
mph
iphy
sin-
like
lipid
raft
prot
ein.
Reg
ulat
es p
olar
izat
ion
of th
e ac
tin c
ytos
kele
ton.
96
RV
S167
Y
DR
388W
01
748
1.4e
-51
Reg
ulat
ion
of a
ctin
cyt
oske
leto
n 97
SA
C7
YD
R38
9W
0290
2 1.
2e-4
3 G
AP
for R
ho1
(Fun
ctio
nally
rela
ted
to B
ag7)
98
SH
S1/S
EP7
YD
L225
W
0344
9 3.
7e-7
2 Se
ptin
99
SP
A2
YLL
021W
04
468
5.6e
-20
Pola
risom
e co
mpo
nent
10
0 SP
H1
YLR
313C
04
468
2.3e
-10
Pola
risom
e co
mpo
nent
. Hom
olog
ous t
o Sp
a2
101
SWE1
Y
JL18
7C
0633
7 5.
3e-4
0 Se
r/Thr
pro
tein
kin
ase;
invo
lved
in sw
itch
from
pol
ariz
ed to
isot
ropi
c gr
owth
10
2 W
SC1/
HC
S77
YO
R00
8C
1154
6 3.
1e-2
1 St
retc
h re
cept
or; c
ell w
all i
nteg
rity
path
way
10
3 W
SC2
YN
L283
C
no
hits
St
retc
h re
cept
or; c
ell w
all i
nteg
rity
path
way
10
4 W
SC3
YO
L105
C
1508
4 1.
6e-2
8 St
retc
h re
cept
or; c
ell w
all i
nteg
rity
path
way
10
5 W
HI3
Y
NL1
97C
04
835
2.3e
-23
Reg
ulat
or o
f cel
l siz
e. R
NA
bin
ding
pro
tein
10
6 YC
K1
YH
R13
5C
0027
4 3.
4e-1
29
Cas
ein
kina
se I
isof
orm
. Cel
l mor
phog
enes
is
107
YCK
2 Y
NL1
54C
00
274
1.6e
-126
C
asei
n ki
nase
I is
ofor
m. C
ell m
orph
ogen
esis
. Fun
ctio
nally
redu
ndan
t with
Yck
1.
108
ZDS1
Y
MR
273C
15
061.
2 4.
7e-2
0 R
egul
atio
n of
cel
l pol
arity
10
9 ZD
S2
YM
L109
W
no
hits
R
egul
atio
n of
cel
l pol
arity
. Par
alog
ue o
f ZD
S1
ACTI
N C
YTO
SKEL
ETO
NSc
gen
eSy
stem
atic
nam
eU
. may
dis M
IPS#
E v
alue
Func
tion
in S
. cer
evisi
ae11
0 AB
P1
YC
R08
8W
0534
0 7.
9e-2
3 A
ctin
-bin
ding
pro
tein
11
1 AC
F4
YJR
083C
A
ctin
cyt
oske
leto
n or
gani
zatio
n 11
2 AC
T1
YFL
039C
11
232
2.0e
-185
A
ctin
11
3 AI
P1
YM
R09
2C
0594
9 3.
0e-8
6 A
ctin
-inte
ract
ing
prot
ein;
act
in p
atch
com
pone
nt
114
APP1
Y
NL0
94W
04
613
9.7e
-22
Act
in fi
lam
ent o
rgan
izat
ion
115
ARC
15
YIL
062C
02
388
2.6e
-25
Subu
nit o
f Arp
2/3
com
plex
11
6 AR
C18
Y
LR37
0C
1024
6 7.
4e-4
4 Su
buni
t of A
rp2/
3 co
mpl
ex
117
ARC
19
YK
L013
C
1017
3 9.
8e-5
8 Su
buni
t of A
rp2/
3 co
mpl
ex
118
ARC
35
YN
R03
5C
1147
9 1.
8e-4
9 Su
buni
t of A
rp2/
3 co
mpl
ex
119
ARC
40
YB
R23
4C
0590
6 3.
7e-6
9 Su
buni
t of A
rp2/
3 co
mpl
ex
120
ARK
1 Y
NL0
20C
03
081
8.7e
-61
Act
in re
gula
ting
Ser/T
hr k
inas
e 12
1 AR
P2
YD
L029
W
0540
5 1.
7e-1
49
Subu
nit o
f Arp
2/3
com
plex
12
2 AR
P3
YJR
065C
11
265
1.2e
-139
Su
buni
t of A
rp2/
3 co
mpl
ex
123
BNI1
Y
NL2
71C
12
254
1.9e
-59
Form
in; a
ctin
nuc
leat
ion
124
BNR1
Y
IL15
9W
0114
1 2.
3e-3
1 Fo
rmin
; act
in n
ucle
atio
n. F
unct
iona
lly re
dund
ant w
ith B
ni1
125
BSP1
Y
PR17
1W
no
hits
In
volv
ed in
act
in c
ytos
kele
ton
orga
niza
tion
126
CAP
1 Y
KL0
07W
00
423
1.1e
-24
F-ac
tin c
appi
ng p
rote
in a
lpha
subu
nit
127
CAP
2 Y
IL03
4C
1117
7 2.
0e-5
0 F-
actin
cap
ping
pro
tein
bet
a su
buni
t 12
8 C
CT1
/TC
P1
YD
R21
2W
0127
9 3.
4e-2
13
Cha
pero
nin
role
for a
ctin
and
tubu
lin
129
CC
T2
YIL
142W
06
235
5.5e
-190
C
hape
roni
n ro
le fo
r act
in a
nd tu
bulin
13
0 C
CT3
Y
JL01
4W
0606
7 1.
1e-1
98
Cha
pero
nin
role
for a
ctin
and
tubu
lin
131
CC
T4
YD
L143
W
0257
1 9.
7e-1
22
Cha
pero
nin
role
for a
ctin
and
tubu
lin
132
CC
T5
YJR
064W
03
959
4.6e
-186
C
hape
roni
n ro
le fo
r act
in a
nd tu
bulin
Fungal Genet Biol. Author manuscript; available in PMC 2009 August 1.
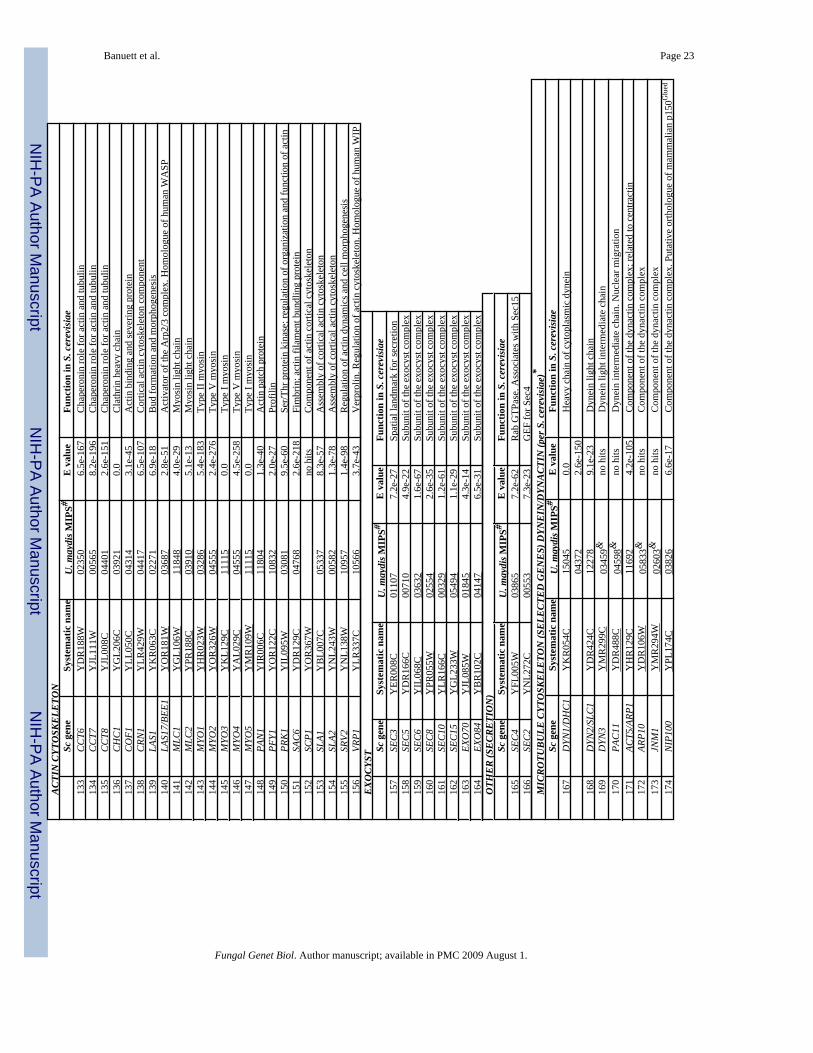
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Banuett et al. Page 23AC
TIN
CYT
OSK
ELET
ON
Sc g
ene
Syst
emat
ic n
ame
U. m
aydi
s MIP
S#E
val
ueFu
nctio
n in
S. c
erev
isiae
133
CC
T6
YD
R18
8W
0235
0 6.
5e-1
67
Cha
pero
nin
role
for a
ctin
and
tubu
lin
134
CC
T7
YJL
111W
00
565
8.2e
-196
C
hape
roni
n ro
le fo
r act
in a
nd tu
bulin
13
5 C
CT8
Y
JL00
8C
0440
1 2.
6e-1
51
Cha
pero
nin
role
for a
ctin
and
tubu
lin
136
CH
C1
YG
L206
C
0392
1 0.
0 C
lath
rin h
eavy
cha
in
137
CO
F1
YLL
050C
04
314
3.1e
-45
Act
in b
indi
ng a
nd se
verin
g pr
otei
n 13
8 C
RN1
YLR
429W
04
417
6.5e
-107
C
ortic
al a
ctin
cyt
oske
leto
n co
mpo
nent
13
9 LA
S1
YK
R06
3C
0227
1 6.
9e-1
8 B
ud fo
rmat
ion
and
mor
phog
enes
is
140
LAS1
7/BE
E1
YO
R18
1W
0368
7 2.
8e-5
1 A
ctiv
ator
of t
he A
rp2/
3 co
mpl
ex. H
omol
ogue
of h
uman
WA
SP
141
MLC
1 Y
GL1
06W
11
848
4.0e
-29
Myo
sin
light
cha
in
142
MLC
2 Y
PR18
8C
0391
0 5.
1e-1
3 M
yosi
n lig
ht c
hain
14
3 M
YO1
YH
R02
3W
0328
6 5.
4e-1
83
Type
II m
yosi
n 14
4 M
YO2
YO
R32
6W
0455
5 2.
4e-2
76
Type
V m
yosi
n 14
5 M
YO3
YK
L129
C
1111
5 0.
0 Ty
pe I
myo
sin
146
MYO
4 Y
AL0
29C
04
555
4.5e
-258
Ty
pe V
myo
sin
147
MYO
5 Y
MR
109W
11
115
0.0
Type
I m
yosi
n 14
8 PA
N1
YIR
006C
11
804
1.3e
-40
Act
in p
atch
pro
tein
14
9 PF
Y1
YO
R12
2C
1083
2 2.
0e-2
7 Pr
ofili
n 15
0 PR
K1
YIL
095W
03
081
9.5e
-60
Ser/T
hr p
rote
in k
inas
e; re
gula
tion
of o
rgan
izat
ion
and
func
tion
of a
ctin
15
1 SA
C6
YD
R12
9C
0476
8 2.
6e-2
18
Fim
brin
; act
in fi
lam
ent b
undl
ing
prot
ein
152
SCP1
Y
OR
367W
no h
its
Com
pone
nt o
f act
in c
ortic
al c
ytos
kele
ton
153
SLA1
Y
BL0
07C
05
337
8.3e
-57
Ass
embl
y of
cor
tical
act
in c
ytos
kele
ton
154
SLA2
Y
NL2
43W
00
582
1.3e
-78
Ass
embl
y of
cor
tical
act
in c
ytos
kele
ton
155
SRV2
Y
NL1
38W
10
957
1.4e
-98
Reg
ulat
ion
of a
ctin
dyn
amic
s and
cel
l mor
phog
enes
is
156
VRP1
Y
LR33
7C
1056
6 3.
7e-4
3 V
erpr
olin
. Reg
ulat
ion
of a
ctin
cyt
oske
leto
n. H
omol
ogue
of h
uman
WIP
EX
OCY
STSc
gen
eSy
stem
atic
nam
eU
. may
dis M
IPS#
E v
alue
Func
tion
in S
. cer
evisi
ae15
7 SE
C3
YER
008C
01
107
7.2e
-27
Spat
ial l
andm
ark
for s
ecre
tion
158
SEC
5 Y
DR
166C
00
710
4.9e
-22
Subu
nit o
f the
exo
cyst
com
plex
15
9 SE
C6
YIL
068C
03
632
1.6e
-67
Subu
nit o
f the
exo
cyst
com
plex
16
0 SE
C8
YPR
055W
02
554
2.6e
-35
Subu
nit o
f the
exo
cyst
com
plex
16
1 SE
C10
Y
LR16
6C
0032
9 1.
2e-6
1 Su
buni
t of t
he e
xocy
st c
ompl
ex
162
SEC
15
YG
L233
W
0549
4 1.
1e-2
9 Su
buni
t of t
he e
xocy
st c
ompl
ex
163
EXO
70
YJL
085W
01
845
4.3e
-14
Subu
nit o
f the
exo
cyst
com
plex
16
4 EX
O84
Y
BR
102C
04
147
6.5e
-31
Subu
nit o
f the
exo
cyst
com
plex
O
THER
(SEC
RETI
ON
)Sc
gen
eSy
stem
atic
nam
eU
. may
dis M
IPS#
E v
alue
Func
tion
in S
. cer
evisi
ae16
5 SE
C4
YFL
005W
03
865
7.2e
-62
Rab
GTP
ase.
Ass
ocia
tes w
ith S
ec15
16
6 SE
C2
YN
L272
C
0055
3 7.
3e-2
3 G
EF fo
r Sec
4 M
ICRO
TUBU
LE C
YTO
SKEL
ETO
N (S
ELEC
TED
GEN
ES) D
YNEI
N/D
YNAC
TIN
(per
S. c
erev
isiae
)*Sc
gen
eSy
stem
atic
nam
eU
. may
dis M
IPS#
E v
alue
Func
tion
in S
. cer
evisi
ae16
7 D
YN1/
DH
C1
YK
R05
4C
1504
5 0.
0 H
eavy
cha
in o
f cyt
opla
smic
dyn
ein
04
372
2.6e
-150
168
DYN
2/SL
C1
YD
R42
4C
1227
8 9.
1e-2
3 D
ynei
n lig
ht c
hain
16
9 D
YN3
YM
R29
9C
0345
9&
no h
its
Dyn
ein
light
inte
rmed
iate
cha
in
170
PAC
11
YD
R48
8C
0459
8&
no h
its
Dyn
ein
inte
rmed
iate
cha
in. N
ucle
ar m
igra
tion
171
ACT5
/ARP
1 Y
HR
129C
11
692
4.2e
-105
C
ompo
nent
of t
he d
ynac
tin c
ompl
ex; r
elat
ed to
cen
tract
in
172
ARP1
0 Y
DR
106W
05
833&
no
hits
C
ompo
nent
of t
he d
ynac
tin c
ompl
ex
173
JNM
1 Y
MR
294W
02
603&
no
hits
C
ompo
nent
of t
he d
ynac
tin c
ompl
ex
174
NIP
100
YPL
174C
03
826
6.6e
-17
Com
pone
nt o
f the
dyn
actin
com
plex
. Put
ativ
e or
thol
ogue
of m
amm
alia
n p1
50G
lued
Fungal Genet Biol. Author manuscript; available in PMC 2009 August 1.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Banuett et al. Page 24KI
NES
INS
(per
S. c
erev
isiae
)#Sc
gen
eSy
stem
atic
nam
eU
. may
dis M
IPS#
E v
alue
Func
tion
in S
. cer
evisi
ae17
5 C
IN8
YEL
O61
C
1067
8 1.
0e-9
6 K
ines
in. M
itotic
spin
dle
asse
mbl
y; c
hrom
osom
e se
greg
atio
n. R
elat
ed to
Kip
1
176
KAR
3 Y
PR14
1C
1198
6 1.
6e-8
0 K
ines
in (-
end
dire
cted
) Rel
ated
to U
m K
in14
17
7 K
IP1
YB
L063
W
1067
8 2.
5e-1
07
Kin
esin
. Fun
ctio
nally
redu
ndan
t with
Cin
8 17
8 K
IP2
YPL
155C
00
896
5.4e
-52
Kin
esin
rela
ted
mot
or p
rote
in R
elat
ed to
Um
Kin
7a a
nd K
in7b
17
9 K
IP3
YG
L216
W
0156
0 1.
7e-1
18
Kin
esin
rela
ted
mot
or p
rote
in R
elat
ed to
Um
Kin
8 18
0 SM
Y1
YK
L079
W
0421
8 3.
4e-4
5 In
tera
cts w
ith M
yo2.
N-te
rmin
al d
omai
n re
late
d to
mot
or d
omai
n of
kin
esin
s. +T
IPs
Sc g
ene
Syst
emat
ic n
ame
U. m
aydi
s MIP
S#E
val
ueFu
nctio
n in
S. c
erev
isiae
181
BIK
1 Y
CL0
29C
06
338
2.5e
-13
MT
plus
end
ass
ocia
ted
prot
ein.
Orth
olog
ue o
f mam
mal
ian
CLI
P170
18
2 BI
M1/
YEB1
Y
ER01
6W
0576
1 7.
5e-5
1 To
geth
er w
ih K
ar9
mak
es u
p th
e M
T ca
ptur
e si
te a
t the
cel
l cor
tex
183
KAR
9 Y
PL26
9W
no
hits
N
ucle
ar m
igra
tion
184
PAC
1 Y
OR
269W
03
164
7.8e
-36
LIS1
/Nud
F ho
mol
ogue
. Nuc
lear
mig
ratio
n TU
BULI
N Sc g
ene
Syst
emat
ic n
ame
U. m
aydi
s MIP
S#E
val
ueFu
nctio
n in
S. c
erev
isiae
185
TUB1
Y
ML0
85C
01
221
4.4e
-181
α–
tubu
lin
186
TUB2
Y
FL03
7W
0582
8 6.
7e-1
85
β–tu
bulin
1055
8 2.
0e-1
60
β–tu
bulin
18
7 TU
B3
YM
L124
C
0122
1 1.
5e-1
80
α–tu
bulin
18
8 TU
B4
YLR
212C
03
803
1.1e
-90
γ–tu
bulin
* C
ompo
nent
s of t
he U
. may
dis d
ynac
tin c
ompl
ex sh
own
are
per c
ompa
rison
with
S. c
erev
isia
e.
&B
last
ana
lysi
s ind
icat
es th
at th
ere
are
no h
its fo
r the
se g
enes
, how
ever
, the
U. m
aydi
s gen
e nu
mbe
rs p
rovi
ded
appe
ars t
o co
rres
pond
to th
e ye
ast g
ene.
# Kin
esin
s sho
wn
are
per c
ompa
rison
with
S. c
erev
isia
e ki
nesi
ns. S
ee a
rticl
e by
Sch
ucha
rdt e
t al.,
200
5 fo
r det
ails
of t
he k
ines
in su
perf
amily
in U
. may
dis.
Fungal Genet Biol. Author manuscript; available in PMC 2009 August 1.