the effect of altered dopaminergic activity on food intake in the rat: evidence for an optimal level...
TRANSCRIPT

Prog. Weuro-Psychoylhu~f~co1. ~01.4, pp.351-362. Pergamon Press Ltd. 1980. Printed in Great Britain.
THE EFFECI’ OF ALTERED DOPAMINERGIC AtXIWI’Y ON FOOD INTAKE IN THE RAT: EVIDENCE FOR AN OPTIMAL LEVEL OF DOPAMINERGIC
ACTMTY FOR BEHAVIOR
MICHAEL J. ZIGMOND, THOMAS G. HEFFNER' and EDWARD M. STRICKER
Departments of Biological Sciences.and Psychology and the Neuropsychobiology Program
University of Pittsburgh Pittsburgh, PA USA
(Final form, July 1980)
Abstract
1. Food intake is reduced by the acute administration of d-amphetamine. This is mimicked by other dopamine (DA) agonists and blocked by DA antagonists.
2. In higher doses, DA antagonists themselves reduce food intake, as do large lesions of the central dopaminergic projections.
3. While the reduction of food intake produced by DA agonists is accompanied by a general behavioral activation, that produced by DA antagonists and by DA-depleting lesions co- incides with a decrease in behavioral arousal.
4. These findings suggest that there is an optimal level of dopaminergic activity (and of behavioral arousal) for the mediation of feeding and that both increases and decreases from this optimum may disrupt feeding as well as other behaviors.
Keywords: amphetamine, anorexic drugs, dopamine, food intake, 6-hydroxydopamine, neuro- leptics
Abbreviations: DA: dopamine; OMI: desmethylimipramine; 6-HDA: 6-hydroxydopamine; i.p.: intraperitoneal; ivt.: intraventricular; L-DOPA: L-dihydroxyphenylalanine; NE: norepinephrine.
Amphetamine decreases food intake in rats and other animals, including humans. The drug also has a broad variety of physiological effects, both in the periphery and in the central nervous system. In the present essay we examine the role of central dopaminergic neurons in amphetamine's behavioral effects and discuss the contribution of these neurons to the central control of food intake and other motivated behaviors.
d-Amphetamine, Feeding and Dopaminergic Activation
d-Amphetamine increases the synthesis and release of dopamine (DA) in doses which also re- duce feeding (Costa and Groppeti, 1970; Chiueh and Moore, 1973; Heffner, 1976). In order to determine whether the release of DA plays an important role in the drug's inhibition of food intake, we determined whether the behavioral effect was attenuated by pretreatment with DA
antagonists. Animals were deprived of food pellets for 20 hr. each day. Intake was mea- sured during the first hour of access to food. d-Amphetamine (0.4-4.0 mg/kg,i.p.) was found to produce a dose-dependent decrease in food intake (Fig. 1). This was markedly attenuated by low doses of DA receptor antagonists, spiroperidol or haloperidol (0.1 mg/kg, i.p.) and by
'Current address: Department of Pharmacological and Physiological Sciences, University of Chicago, Abbott Hall, 947 E. 58th Street, Chicago, Illinois 60637.
351
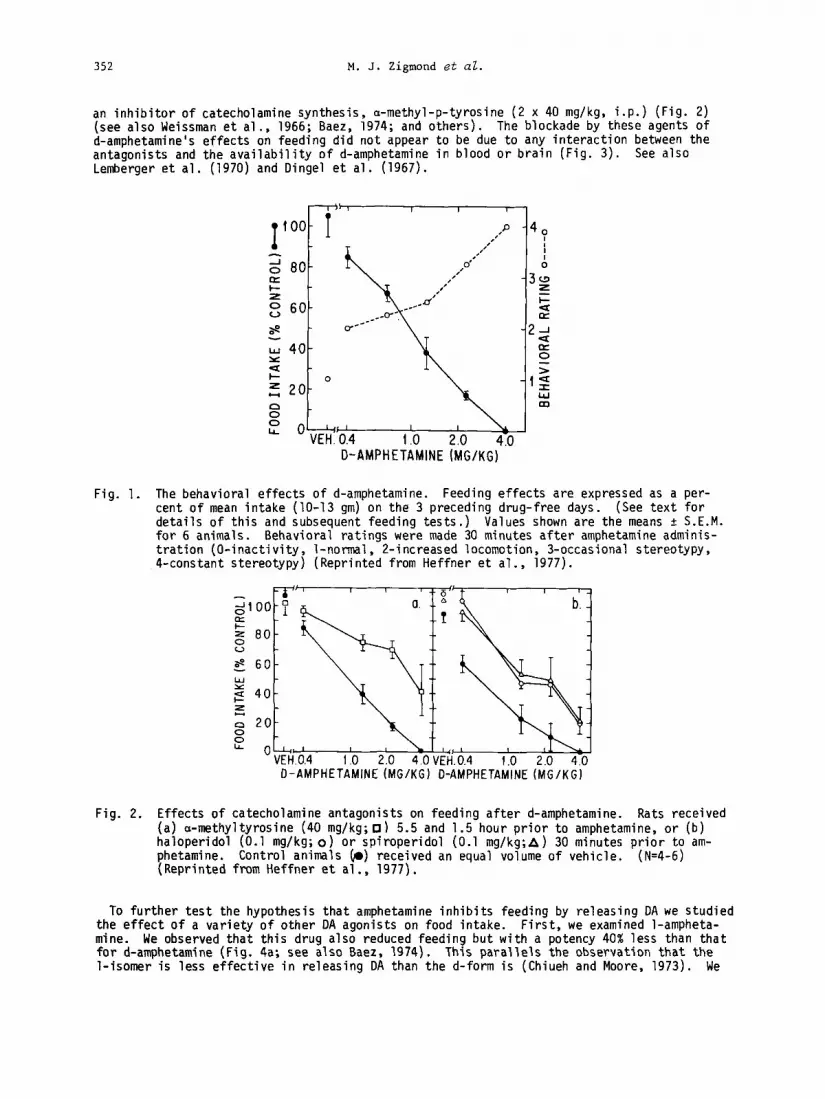
352 M. J. Zigmond et al.
an inhibitor of catecholamine synthesis, a-methyl-p-tyrosine (2 x 40 mg/kg, i.p.) (Fig. 2) (see also Weissman et al., 1966; Baez, 1974; and others). The blockade by these agents of d-amphetamine's effects on feeding did not appear to be due to any interaction between the antagonists and the availability of d-amphetamine in blood or brain (Fig. 3). See also Lemberger et al. (1970) and Oingel et al. (1967).
D-AMPHETAMINE (MG/KGI
Fig. 1. The behavioral effects of d-amphetamine. Feeding effects are expressed as a per- cent of mean intake (lo-13 gm) on the 3 preceding drug-free days. (See text for details of this and subsequent feeding tests.) Values shown are the means i: S.E.M. for 6 animals. Behavioral ratings were made 30 minutes after amphetamine adminis- tration (O-inactivity, l-normal, 2-increased locomotion, 3-occasional stereotypy, 4-constant stereotypy) (Reprinted from Heffner et al., 1977).
2100 ,, 6’
a. _. a b. E 7 = 80 z e 60 2 ;-Q 40 3 0 20
s 0
L!?ssil p
VEH.O.4 1.0 2.0 4.0VEH.0.4 1.0 2.0 4.0 O-AMPHETAMINE (MC/KG) D-AMPHETAMINE (~&/KG)-
Fig. 2. Effects of catecholamine antagonists on feeding after d-amphetamine. Rats received (a) a-methyltyrosine (40 mg/kg;n) 5.5 and 1.5 hour prior to amphetamine, or (b) haloperidol (0.1 mg/kg;o) or spiroperidol (0.1 mg/kg;A) 30 minutes prior to am- phetamine. Control animals (r) received an equal volume of vehicle. (N=4-6) (Reprinted from Heffner et al., 1977).
To further test the hypothesis that amphetamine inhibits feeding by releasing DA we studied the effect of a variety of other DA agonists on food intake. First, we examined l-ampheta- mine. We observed that this drug also reduced feeding but with a potency 40% less than that for d-amphetamine (Fig. 4a; see also Baez, 1974). This parallels the observation that the l-isomer is less effective in releasing DA than the d-form is (Chiueh and Moore, 1973). We
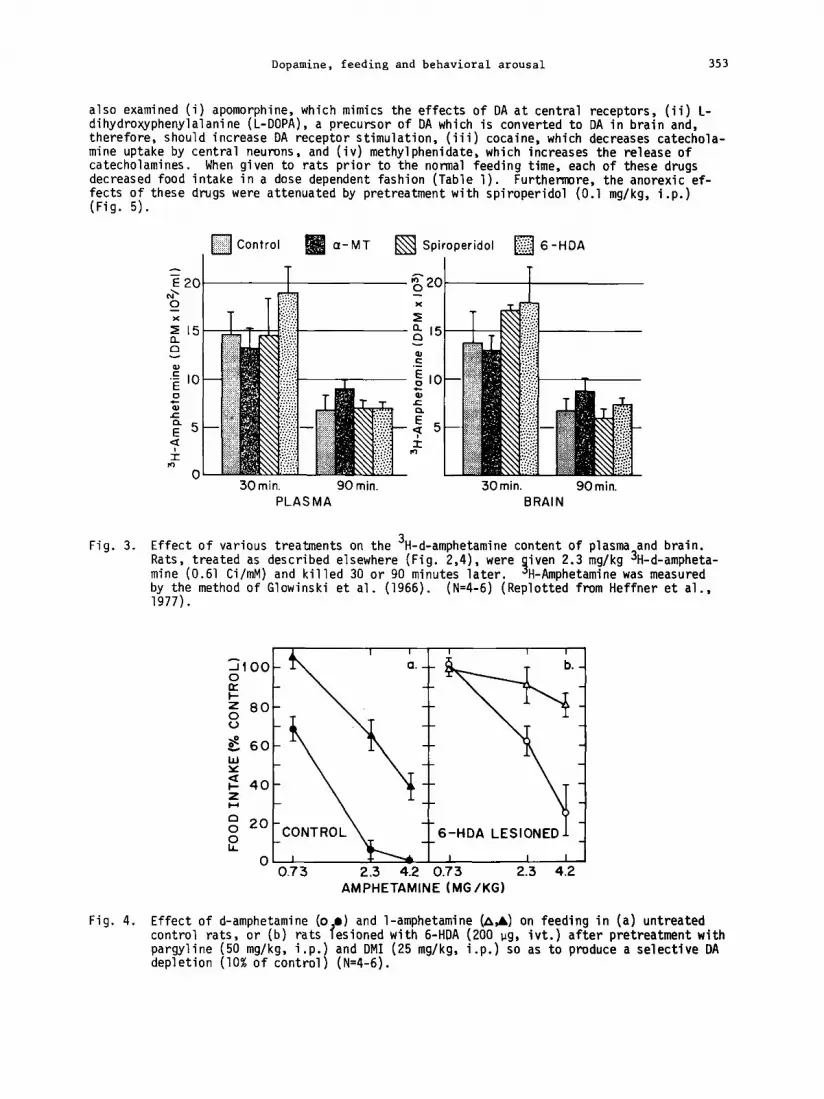
Dopamine, feeding and behavioral arousal 353
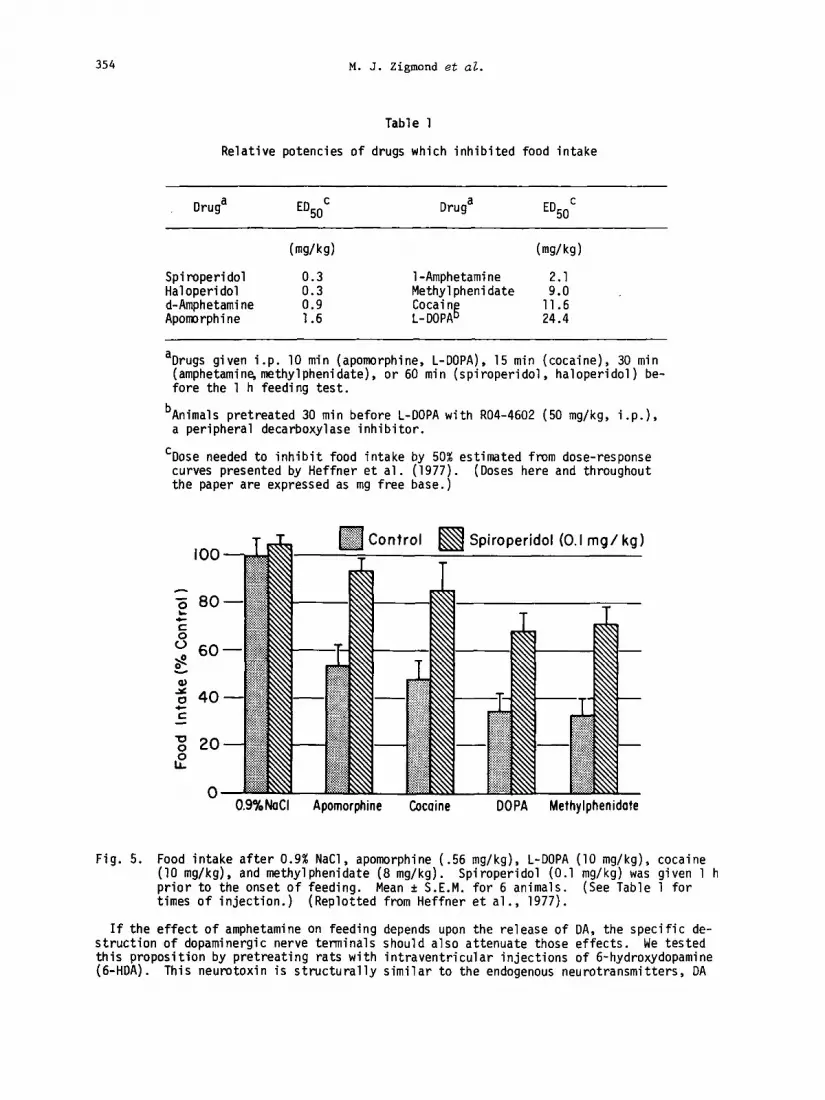
also examined (i) apomorphine, which mimics the effects of DA at central receptors, (ii) L- dihydroxyphenylalanine (L-DOPA), a precursor of DA which is converted to DA in brain and, therefore, should increase DA receptor stimulation, (iii) cocaine, which decreases catechola- mine uptake by central neurons , and (iv) methylphenidate, which increases the release of catecholamines. When given to rats prior to the normal feeding time, each of these drugs decreased food intake in a dose dependent fashion (Table 1). Furthemre, the anorexic ef- fects of these drugs were attenuated by pretreatment with spiroperidol (0.1 mg/kg, i.p.) (Fig. 5).
I’ q Control a-MT tsl Spiroperidol q 6-HDA I T
I
30min. 90 min. PLASMA
30min. 90min. BRAIN
Fig. 3. Effect of various treatments on the 3H-d-amphetamine content of plasma and brain. Rats, treated as described elsewhere (Fig. 2,4), were mine (0.61 Ci/mM) and killed 30 or 90 minutes later. 9
iven 2.3 mg/kg 3H-d-ampheta- H-Amphetamine was measured
by&Fe method of Glowinski et al. (1966). (N=4-6) (Replotted from Heffner et al.,
I I I I I * Iioo- a. b. -
i? I- f$ 80 0
E 60
e 2 40 z l-l g 20
2 0
:$!?: CONTROL 6-HDA LESIONED
0.73 2.3 4.2 0.73 2.3 4.2 AMPHETAMINE (MG /KG)
Fig. 4. Effect of d-amphetamine (o l ) and l-amphetamine (A,& on feeding in (a) untreated control rats, or (b) rats lesioned with 6-HDA (200 ug, ivt.) after pretreatment with pargyline (50 mg/kg, i.p.) and DMI (25 mg/kg, i.p.) so as to produce a selective DA depletion (10% of control) (N=4-6).
354 M. J. Zigmond et al.
Table 1
Relative potencies of drugs which inhibited food intake
ED50C ED50C (w/kg) h/kg)
Spiroperidol 0.3 l-Amphetamine 2.1 Haloperidol Methylphenidate d-Amphetamine i-i Cocaing 1X Apomorphine 1:6 L-DOPA 24:4
aDrugs given i.p. 10 min (apomorphine, L-DOPA), 15 min (cocaine), 30 min (amphetamine,methylphenidate), or 60 min (spiroperidol, haloperidol) be- fore the 1 h feeding test.
bAnimals pretreated 30 min before L-DOPA with R04-4602 (50 mg/kg, i.p.), a peripheral decarboxylase inhibitor.
'Dose needed to inhibit food intake by 50% estimated from dose-response curves presented by Heffner et al. (1977). (Doses here and throughout the paper are expressed as mg free base.)
100
0.9%NaCl Apomorphine Cocaine DOPA Methylphenidate
Fig. 5. Food intake after 0.9% NaCl, apomorphine (.56 mg/kg), L-DOPA (10 mg/kg), cocaine (10 mg/kg), and methylphenidate (8 mg/kg). Spiroperidol (0.1 mg/kg) was given 1 h prior to the onset of feeding. Mean + S.E.M. for 6 animals. (See Table 1 for times of injection.) (Replotted from Heffner et al., 1977).
If the effect of amphetamine on feeding depends upon the release of DA, the specific de- struction of dopaminergic nerve terminals should also attenuate those effects. We tested this proposition by pretreating rats with intraventricular injections of 6-hydroxydopamine (6-HDA). This neurotoxin is structurally similar to the endogenous neurotransmitters, DA

Dopamine, feeding and behavioral arousal 355
and norepinephrine (NE), and is believed to gain preferential access to neurons containing these catecholamines through the amine-specific uptake mechanisms in their terminals. Once within these neurons, metabolites are formed that destroy the nerve terminals. We observed that animals lesioned with 6-HDA several weeks prior to testing were significantly less af- fected by both d- and l-amphetamine (Fig. 4b; see also Samanin et al., 1972; Fibiger et al., 1973a; Hollister et al., 1975). Moreover, the residual effect of amphetamine on feeding in these rats was completely suppressed if the 6-HDA lesioned animals were given haloperidol (0.1 mg/kg, i.p.) prior to the administration of d-amphetamine (Table 2).
Table 2
Effect of decreased dopaminergic function on d-amphetamine-induced inhibition of feedinga
Treatment
Vehicle
Food Intake
Amphetamine
gmlh % vehicle
Control 8.5 f 0.4 0.0 + 0.0 6-HDA-lesion 9.2 f 0.8 4.9 i: 1.3 Ok
Haloperidol 8.8 ck 1.0 3.7 f 0.4 3 6-HDA lesion + 8.9 f 0.5 8.7 CL 0.9 98
Haloperidol
aLesioned rats received pargyline (50 mg/kg, i.p.), DMI (25 mg/kg, i.p.), and 6-HDA (200 ng, ivt.). Non-lesioned animals received pargyline, DMI and ivt. vehicle. Feeding tests were carried out several weeks later. Drug doses were: d-amphetamine, 2.3 mg/kg; haloperidol, 0.1 mg/kg. (N=6 per treatment).
bSignificant reduction in food intake (pc.05).
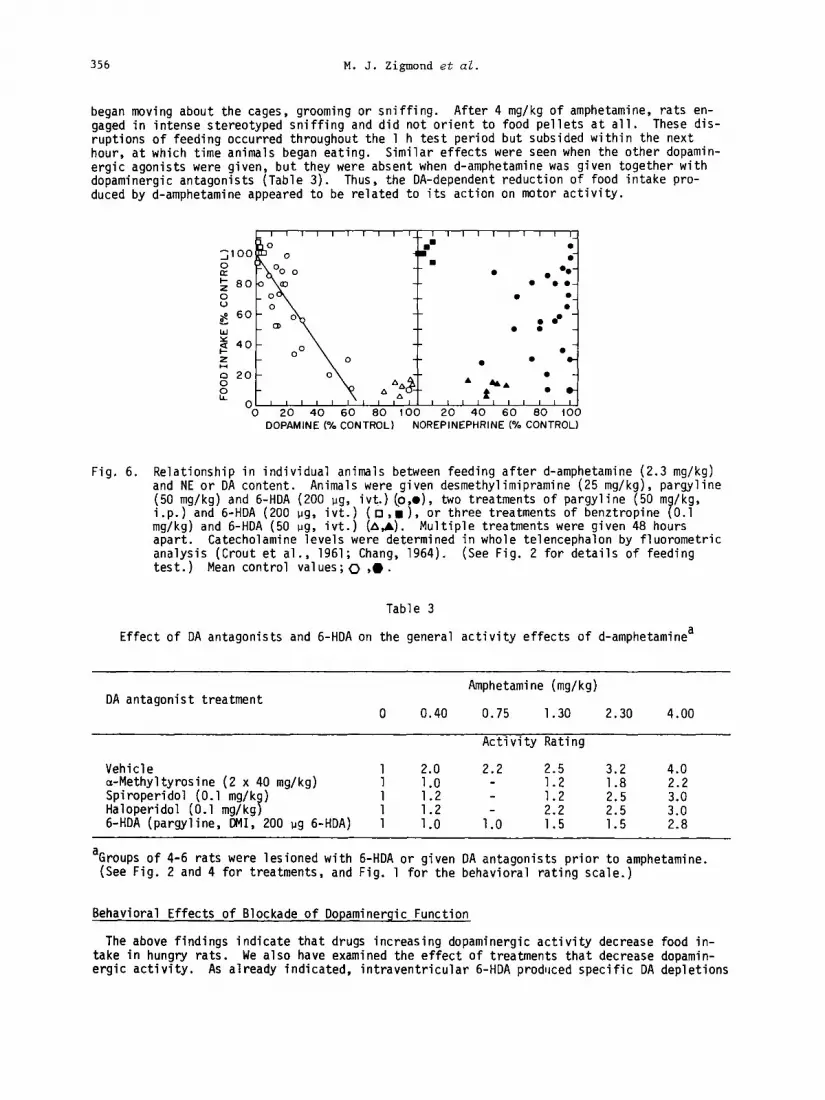
In order to separate more completely the role of DA and NE in the interaction between C-HDA and amphetamine, we varied the damage produced by the neurotoxin. Prior to C-HDA administra- tion, animals received one of several treatments: (i) pargyline (50 mg/kg, i.p.), which in- hibits monoamine oxidase and potentiates the effects of 6-HDA, (ii) pargyline together with desmethylimipramine (DMI,25 mg/kg, i.p.), a drug that prevents the uptake of 6-HDA into nor- adrenergic nerve terminals and thereby restricts its toxic effects to dopaminergic neurons, and (iii) benztropine (0.1 mg/kg, i.p.), a drug that referentially blocks u minergic neurons and can be used together with severa 7 small doses of C-HDA @e5dn$)d!ga- produce a selective loss of NE. We observed that amphetamine's effects on feeding were at- tenuated by lesions that selectively depleted DA but that little or no effect was produced by the selective loss of NE (Heffner, 1976). This is most clearly seen by comparing the be- havioral effect of d-amphetamine (2.3 mg/kg,i.p.) with the remaining concentrations of DA and NE for individual animals. While the effects of d-amphetamine were reduced in proportion to the loss of DA, no such relationship existed for NE (Fig. 6). These data, and those discus- sed above (Table 2), emphasize the importance of central DA neurons in mediating ampheta- mine's effects on feeding and fail to support a comparable role for NE cells.
Amphetamine and Motor Activity
In addition to food intake measurements, we rated the general activity of each rat given a dopaminergic agonist. d-Amphetamine produced a dose-dependent increase in behavioral activ- ity that was the mirror-image of the decrease in food intake (Fig. 1). Normally, rats adap- ted to the 4 h feeding schedule would seize food pellets when they were made available, carry them to the rear of their cages and imnediately begin eating. In contrast, whereas rats given amphetamine often initially grasped pellets, they usually dropped them and then
356 M. J. Zigmond et al.
began moving about the cages, grooming or sniffing. After 4 mg/kg of amphetamine, rats en- gaged in intense stereotyped sniffing and did not orient to food pellets at all. These dis- ruptions of feeding occurred throughout the 1 h test period but subsided within the next hour, at which time animals began eating. Similar effects were seen when the other dopamin- ergic agonists were given, but they were absent when d-amphetamine was given together with dopaminergic antagonists (Table 3). Thus, the DA-dependent reduction of food intake pro- duced by d-amphetamine appeared to be related to its action on motor activity.
0 20-
0 1' " 0 20 40 60 60 100 20 40 60 60 100
DOPAMINE We CONTROL) NOREPINEPHRINE (% CONTROL)
Fig. 6. Relationship in individual animals between feeding after d-amphetamine (2.3 mg/kg) and NE or DA content. Animals were given desmethylimipramine (25 mg/kg), pargyline (50 mg/kg) and 6-HDA (200 ug, ivt.) (o l ), two treatments of pargyline (50 i.p.) and 6-HDA (200 pg. ivt.) (cl,.),
m /kg, or three treatments of benztropine 9 0.1
mg/kg) and 6-HDA (50 ug, ivt.) (A,&. Multiple treatments were given 48 hours apart. Catecholamine levels were determined in whole telencephalon by fluorometric analysis (Crout et al., 1961; Chang, 1964). (See Fig. 2 for details of feeding test.) Mean control values; 0 ,O.
Table 3
Effect of DA antagonists and 6-HDA on the general activity effects of d-amphetaminea
DA antagonist treatment Amphetamine (mg/kg)
0 0.40 0.75 1.30 2.30 4.00
Activity Rating
Vehicle 1 2.0 2.2 2.5 3.2 4.0 a-Methyltyrosine (2 x 40 mg/kg) 2.2 Spiroperidol (0.1 mg/k ) Haloperidol (0.1 mg/kg s
; 1:; : E 1 2:2
E 2:5
3.0 3.0
6-HDA (pargyline, DMI, 200 6-HDA) Irg 1 ;:o' 1:o 1.5 1.5 2.8
aGroups of 4-6 rats were lesioned with 6-HDA or given DA antagonists prior to amphetamine. (See Fig. 2 and 4 for treatments, and Fig. 1 for the behavioral rating scale.)
Behavioral Effects of Blockade of Dopaminergic Function
The above findings indicate that drugs increasing dopaminergic activity decrease food in- take in hungry rats. We also have examined the effect of treatments that decrease dopamin- ergic activity. As already indicated, intraventricular 6-HDA produced specific DA depletions

Dopamine, feeding and behavioral arousal 357
of 8-85% but had little apparent effect on ad libitum food intake. However, larger deple- tions (i.e. at least 90-95%) invariably produced aphagia (Zigmond and Stricker, 1972, 1973). Not only would these animals not eat the food they had been maintained on previously, they would not even accept such palatable diets as chocolate Metrecal and would starve to death unless fed by intragastric intubation. Similar results have been observed by other investi- gators utilizing a variety of procedures that presumably destroyed DA-containing neurons (Anand and Brobeck, 1951; Teitelbaum and Epstein, 1962; Ungerstedt, 1971; Fibiger et al., 1973b; Marshall et al., 1974; and others).
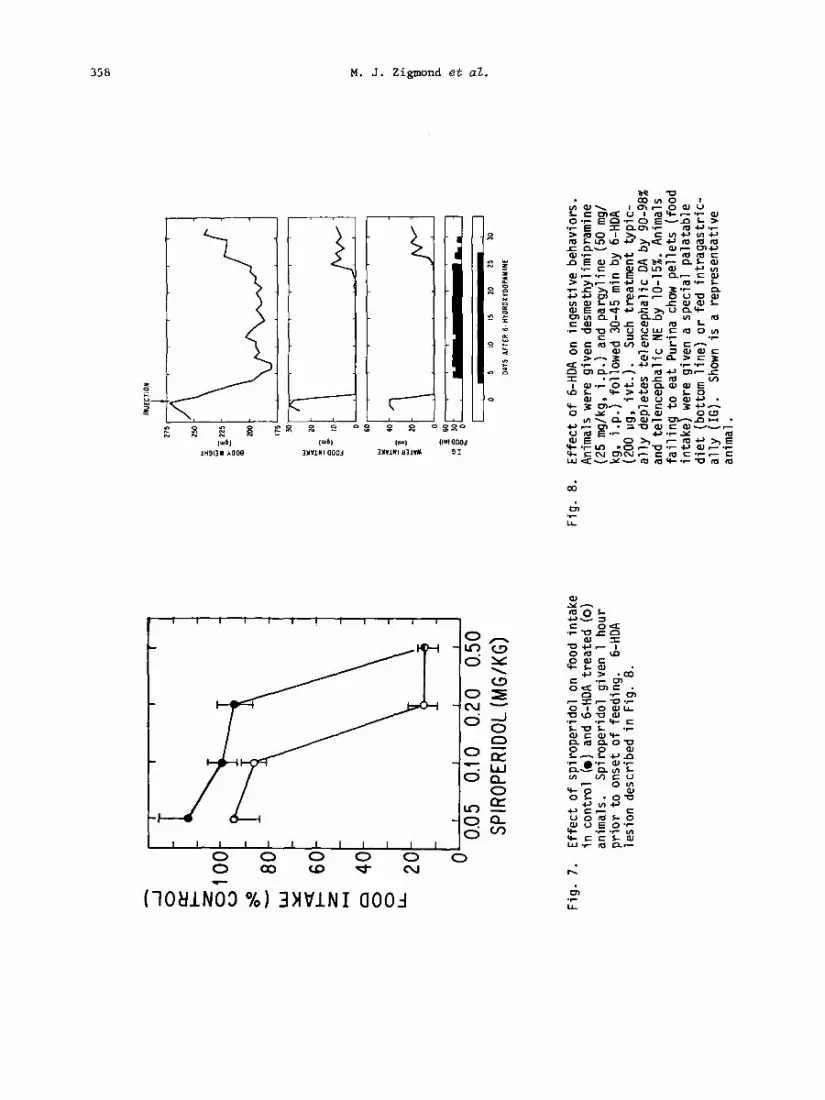
As with amphetamine's action, the effects of the large DA-depleting brain lesions were not restricted to food intake. Animals were akinetic, cataleptic, and showed pronounced sensory neolect after the lesions (Marshall et al.. 1971, 1976). Conseauentlv, in addition to aoha- gia they also show a loss of drinking, behaviorai thermoregulation, and other activities: Thus, it cannot be concluded that the loss of brain DA eliminated feeding due to a destruc- tion of central neural systems specific to food intake. Instead, it appeared to destroy some comnon activational component of all motivated behavior. This interpretation is supported by the well-known behavioral depression (and accompanying decrease in feeding) produced by high doses of a variety of drugs which reduce dopaminergic function, including spiroperidol, chlorpromazine, cr-methyltyrosine and reserpine (e.g. Fig. 7; Heffner et al., 1977).
Recovery from Large DA-Depleting Lesions
The DA depletions after 6-HDA treatment were permanent. However, lesioned animals re- ceiving intragastric feeding gradually did recover feeding and drinking behaviors (Fig. 8), a phenomenon which may be analogous to the gradual behavioral tolerance which develops to many pharmacological disruptions of DA function. At first the 6-HDA-lesioned animals ate only palatable foods and fluids, but later they ingested dry chow and finally they drank water. Sensorimotor dysfunctions diminished progressively as ingestive behaviors returned. Once these animals had recovered, the original behavioral deficits, including decreased feeding and sensorimotor dysfunctions, could be reinstated by doses of haloperidol, spiro- peridol, or a-methyltyrosine which had little effect in control rats (Fig. 7, Table 2; see also Zigmond and Stricker, 1973; Heffner et al., 1977; Stricker et al., 1979). These re- sults make it clear that residual dopaminergic function continued to participate in the con- trol of food intake despite the extensive DA depletions, and that recovery of function did not result from transfer of this function to some other neural system. Elsewhere, we have suggested that the recovery of function resulted, at least in part, from adaptive neurochem ical changes that occurred at residual dopaminergic synapses in the brain-damaged animals. These include increased release of DA from residual neurons and increased efficacy of DA due to increased receptor sensitivity and the loss of re-uptake sites for the transmitter (Stricker and Zignmnd, 1976).
Lesioned rats that had resumed voluntary food and water ingestion and were maintaining body weight nevertheless were not completely recovered. Their residual deficits provided another perspective for viewing the role of brain DA in the control of motivated behavior. Unlike normal rats, the lesioned animals did not increase food intake in response to severe cold stress or insulin-induced hypoglycemia , and their lack of response was associated with a regression in sensorimotor performance to the akinesia and sensory neglect that had char- acterized the earlier stages of recovery (Zigmond and Stricker, 1972; Stricker and Zigmond, 1974; Marshall et al., 1974; Stricker et al., 1975, 1979). Such stressors appear to increase DA turnover in certain telencephalic structures (Kennedy et al., 1979; Thierry et al., 1976). Thus, the combination of elevated basal turnover and the further increase in DA provided by stress may produce an exhaustion of releasable DA at residual terminals, thereby precluding a behavioral response.
Dopaminergic Activity and Behavior
Some of the earliest theories of amphetamine action held that the drug activated an "as- cending reticular activating system" (Moruzzi and Magoun, 1949; Bradley and Key, 1958). The reticular activating system was viewed as a network which received collateral sensory input and then facilitated transmission along primary sensory and motor pathways (Moruzzi and Magoun, 1949; Lindsley, 1951; Hebb, 1955). It was proposed that an optimal level of activa- tion existed for specific behavioral responses; too little activation would result in little or no transmission, too much in a failure to inhibit competing signals.
SP
lRO
PE
RlD
OL(
MG
/KG
)
Fig.
7. Ef
fect
of sp
irope
ridol
on
food
intak
e in
contr
ol (0)
an
d 6-
HDA
treate
d (0
) an
imals
. Sp
irope
ridol
given
1
hour
pr
ior
to on
set
of fe
eding
. 6-
HDA
lesion
de
scrib
ed
in Fig
. 8.
Fig.
8. Ef
fect
of 6-
HDA
on
inges
tive
beha
viors.
An
imals
we
re
given
de
smet
hylim
ipram
ine
(25
mg/kg
, i.p
.) an
d pa
rgyli
ne
(50
mg/
kg,
i.p.)
follo
wed
30-4
5 mi
n by
6-
HDA
(200
vg
, ivt
.).
Such
tre
atmen
t typ
ic-
ally
deple
tes
telen
ceph
alic
DA
by
90-9
8%
and
telen
ceph
alic
NE
by
lo-15
%.
Anim
als
failin
to
eat
Purin
a ch
ow
pelle
ts (fo
od
intak
e 3
were
giv
en
a sp
ecial
pa
latab
le die
t (b
ottom
lin
e)
or fed
int
raga
stric-
all
y (IG
). Sh
own
is a
repr
esen
tative
an
imal.

Dopamine, feeding and behavioral arousal 359
Subsequent studies have demonstrated that the mesencephalic DA cells and their ascending projections are located largely within the classically defined reticular formation, and have a variety of physiological and anatomical characteristics which might suit them for a role in the modulation of behavioral arousal (see reviews by Kety, 1970; Jouvet, 1972; Bolme et al., 1972; Stricker and Zigmond. 1976). This might be accomplished by the introduction of a specific dopaminergic component within the circuitry mediating each type of behavior, or by a wholly non-specific DA-containing system which is activated under all conditions requiring a behavioral response. Either possibility is consistent with the observations presented here; treatments which decreased dopaminergic activity produced a decrease in feeding and other behaviors, while drugs which increased dopaminergic activity decreased feeding but in- creased overall behavioral activity, as well. Thus, we believe that after severely decreased dopaminergic activity, feeding may be reduced because the amount of exteroceptive (food) and interoceptive (hunger) input is no longer adequate to elicit an appropriate behavioral re- sponse. Conversely, amphetamine and the other dopaminergic agonists may disrupt behavior by decreasing the stimulus intensity needed for any response. In this situation, hunger stimuli may no longer be prepotent in the food-deprived animal as feeding could not then occur.
, and a discriminative response such From such a perspective, the "anorexia" that is cotnnonly
described after the administration of amphetamine may not reflect a selective loss of appe- tite but rather an altered brain state in which animals cannot respond selectively.
Such a model has several important implications. It suggests that systemically adminis- tered drugs acting as DA agonists or antagonists throughout the brain will not have effects limited to feeding. Unfortunately, because the neurotransmitters which are involved in medi- ating the specific component of the hunger signal are not yet known, and because there is no reason to presume that, once identified, they will prove to be unique to hunger, we do not anticipate that some alternative drug with central actions will become available for use as a specific appetite depressant.
Table 4
Effects of d-amphetamine on food intake in animals not yet recovered from central DA-depleting lesions
Treatment Food Intake (gms)a
Control Lesionedb Animals Animals
Saline d-Amphetamine
0.74 mg/kg
2.2 mg/kg
10.5 f 2.8 16.1 f 5.6
8.9 f 2.5 34.5 + 7.0c
1.9 f D.6c 45.7 f 4.7c
aTwo hour intake of a mash of powdered lab chow and 10% sucrose. Values are the means of 5-6 animals f S.E.M.
bSeveral weeks prior to the experiment these animals received a bilateral electrolytic lesion (lma for 15 set) in the far lateral hypothalamus at the level of the ventromedial nucleus. When these measurement were made rats were not able to maintain themselves on a normal diet of lab chow pellets and water but would eat when given a mash of chow and 10% su- crose for 2 hours each day ("stage three", Teitelbaum and Epstein, 1962). Control animals were maintained on lab chow pellets and water.
'Food intake significantly differed frw saline treatment (pq.01).
360 M. J. Zigmond et al.
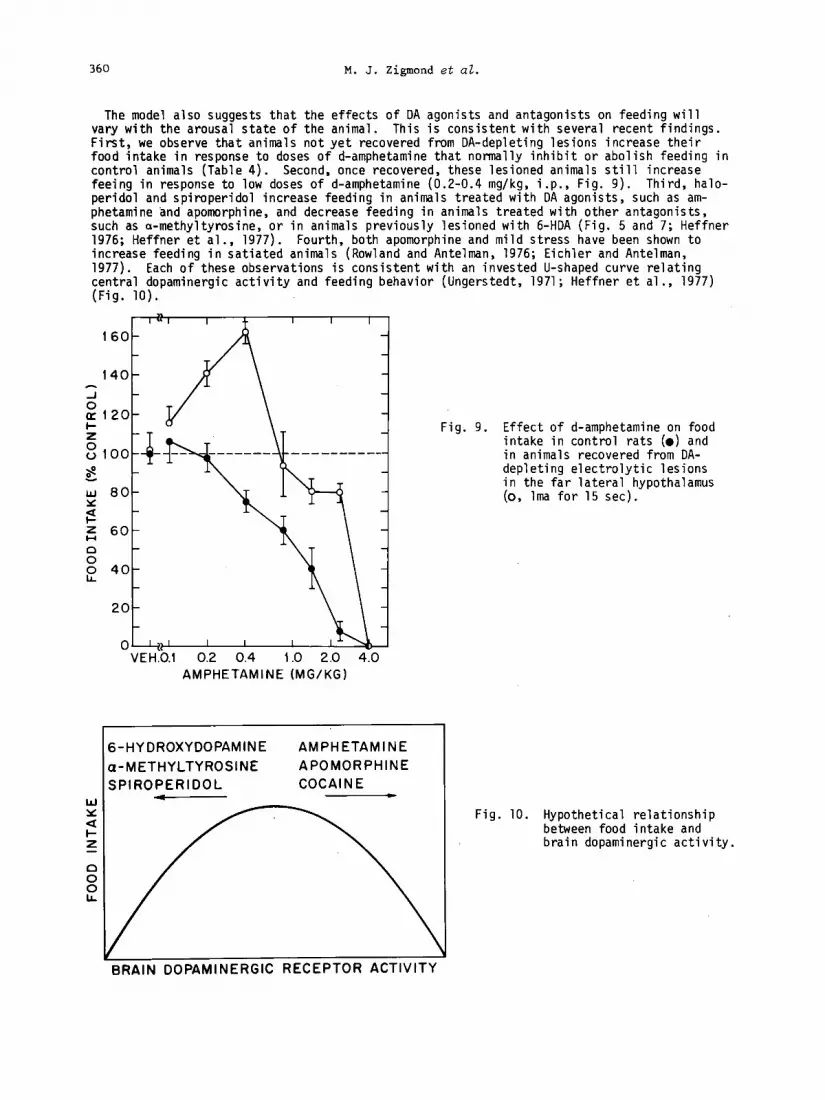
The model also suggests that the effects of DA agonists and antagonists on feeding will vary with the arousal state of the animal. This is consistent with several recent findings. First, we observe that animals not yet recovered from DA-depleting lesions increase their food intake in response to doses of d-amphetamine that normally inhibit or abolish feeding in control animals (Table 4). Second, once recovered, these lesioned animals still increase feeing in response to low doses of d-amphetamine (0.2-0.4 mg/kg, i.p., Fig. 9). Third, halo- peridol and spiroperidol increase feeding in animals treated with DA agonists, such as am- phetamine and apomorphine, and decrease feeding in animals treated with other antagonists, such as a-methyltyrosine, or in animals previously lesioned with 6-HDA (Fig. 5 and 7; Heffner 1976; Heffner et al., 1977). Fourth, both apomorphine and mild stress have been shown to increase feeding in satiated animals (Rowland and Antelman, 1976; Eichler and Antelman, 1977). Each of these observations is consistent with an invested U-shaped curve relating central dopaminergic activity and feeding behavior (Ungerstedt, 1971; Heffner et al., 1977) (Fig. 10).
160-
Fig. 9. Effect of d-amphetamine on food intake in control rats (0) and in animals recovered from DA- depleting electrolytic lesions in the far lateral hypothalamus (0, lma for 15 set).
60-
VEH:b.l 0.2 0.4 1.0 2.0 4.0 AMPHETAMINE (MG/KG)
6-HYDROXYDOPAMINE AMPHETAMINE
a-METHYLTYROSINE APOMORPHINE
SPIROPERIDOL COCAINE c
BRAIN DOPAMINERGIC RECEPTOR ACTIVITY
Fig. 10. Hypothetical relationship between food intake and brain dopaminergic activity.

Dopamine, feeding and behavioral arousal 361
References
ANAND, B.K. and BROBECK, J.R. (1951). Hypothalamic control of food intake in rats and cats. Yale J. Biol. Med. 24: 123-140.
BAEZ, L.A. (1974). Role of catecholamines in the anorexic effects of amphetamine in rats. Psychopharmacologia 35: 91-98.
BRADLEY, P. and KEY, Br(1958). The effect of drugs on arousal responses produced by elec- trical stimulation of the reticular formation of the brain. EEG Clin. Neurophysiol. 10: - 97-110.
BOLME, P., FUXE, K., and LIDBRINK, P. (1972). On the function of central catecholamine neurons -- their role in cardiovascular and arousal mechanisms. Res. Comm. Chem. Path. Pharmacol. 4: 657-697.
CHANG, C.C. (r964). A sensitive method for spectrofluorometric assay of catecholamines. Int. J. Neuropharmacol. 3: 643-649.
CHIUEH, C.C. and MOORF, K.E. (1973). Release of the endogenously synthesized catechols from the caudate nucleus by stimulation of the nigrostriatal pathway and by the administration of d-amphetamine. Brain Res. 50: 221-225.
COSTA, E. and GROPPETTI, A. (197m. Biosynthesis and storage of catecholamines in tissues of rats injected with various doses of &hetamine. In: Amphetamines and Related Com- pounds, Costa, E. and Carattini, S. (Eds.), pp. 169-204. Raven Press, New York.
COSTA, E., GROPPETTI, A. and NAIMZADA, M.K. (1972). Effects of amphetamine on the turnover rate of brain catecholamines and motor activity. Brit. J. Phannacol. 44: 742-751.
CROUT, R.J., CREVELING, C.R. and UDENFRIEND, S. (1961). Norepinephrine Metabolism in rat brain and heart. J. Pharmacol. Exp. Therap. 132: 269-277.
DINGEL, J.V., OWENS, M.L., NORVICH, M.R. and SUER, F. (1967). On the role of norepineph- rine biosynthesis in the central action of amphetamine.
EICHLER, A.J. and ANTELMAN, S.H. (1977). Apomorphine: Life Sci. 6: 1155-1162.
feeding or anorexia depending on internal state. Comm. Psychophann. 1: 533-540.
FIBIGER, H.C., FIBIGER, H.P. and ZIS, Ti.P. (1973a). Attenuation of amphetamine-induced motor stimulation and stereotypy by 6-hydroxydopamine in the rat. Brit. J. Phatmacol. 47: 583-692.
FIEGER, H.C., ZIS, A.P. and MCGEER, E.G. (1973b). Feeding and drinking deficits after 6- hydroxydopamine administration in the rat: similarities to the lateral hypothalamic syndrome. Brain Res. 55: 135-148.
GLOWINSKI, J., AXELROD, x and IVERSEN, L. (1966). the r t brain. and 3
Regional studies of catecgolamines in IV. Effects of drugs on the disposition and metabolism of H -norepinephrine
H -dopamamine. J. Pharmacol. Exp. Therap. 153: 30-41. HEBB, D.O. (1955). Drives and the C.N.S. (conceptual nervous system). Psychol. Rev. 62: -
243-254. HEFFNER, T.G. (1976). Effects of dopaminergic agonists and antagonists on food intake in
normal and 6-hydroxydopamine-treated rats. Unpublished Doctoral Dissertation, University of Pittsburgh.
HEFFNER, T.G., ZIGMOND, M.J. and STRICKER, E.M. (1977). Effects of dopaminergic agonists and antagonists on feeding in intact and 6-hydroxydopamine-treated rats. J. Pharmacol. Exp. Therap. 201: 380-399.
HOLLISTER, A.S., ERVIN, G.N., COOPER, B.R. and BREESE, G.R. (1975). The roles of monoamine neural systems in the anorexia induced by (+)-amphetamine and related compounds. Neuro- pharmacology 14: 715-723.
JOUVET, N. (197n. The role of monoamines and acetylcholine-containing neurons in the regu- lation of the sleep-waking cycle. Ergeb. Physiol. Biol. Chem. Exp. Pharmacol. 64: 166-307.
KENNEDY, L., SALLER, C. and ZIGMOND, M.J. (1979). Stress-induced increases in diFdroxy- phenylacetic acid in rat frontal cortex. Neurosciences Abstracts 5: 339.
KETY, S.S. (1970). The biogenic amines in the central nervous system: their possible roles in arousal, emotion, and learning. In: The Neurosciences: Second Study Program, F.O. Schmitt (Ed.), pp. 324-336. Rockefeller University Press, NY.
LEMBERGER, I., WITT, E.D., DAVIS, J.M. and KOPIN, I. (1970). The effects of haloperidol and chloromazine on amphetamine metabolism and amphetamine stereotype behavior in the rat. J. Pharmacol. Exp. Therap. 174: 428-433. 7
LINDSLEY, D.B. (1951). Emotion. In: Handbook of Experimental Psychology, S. Stevens (Ed.), pp. 473-516, Wiley and Sons, New York.
MARSHALL, J., LEVITAN, D. and STRICKER, E. (1976). Activation-induced restoration of sen- sorimotor functions in rats with dopamine-depleting brain lesions. J. Comp. Physiol. Psychol. 90: 536-546. -

3$2 M. J. Zigmond et al.
MARSHALL, J., TURNER, B. and TEITELBAUM, P. (1971). Sensory neglect produced by lateral hypothalamic lesions. Science (Washington) 174: 523-525.
MARSHALL, J., RICHARDSON, J.S. and TEITELBAUM,T (1974). Nigrostriatal bundle damage and the lateral hypothalamic syndrome. J. Comp. Physiol. Psychol. 87: 808-830.
MORUZZI, 6. and MAGOUN, H. (1949). Brain stem reticular formation and activation of the EEG. EEG. Clin. Neurophysiol. 1: 455-473.
ROWLAND, N.E. and ANTELMAN.3.J. (1976). Stress-induced hyperphagia and obesity in rats: a possible model for understanding human obesity. Science (Washington) 191: 319-312.
SAMANIN, R., GHEZZI, D., VALZELLI, L. and GARATTINI, S. (1972). The effas of selective lesioniog of brain serotonin or catecholanine containing neurons on the anorectic activity of fenfluramine and amphetamine. Eur. J. Phannacol. 19: 318-322.
STRICKER, E.M. and ZIGMOND, M.J. (1974). Effects on ho%ostasis of intraventricular injec- tions of 6-hydroxydopamine in rats. J. Comp. Physiol. Psychol. &: 973-994.
STRICKER, E.M. and ZIGMOND, M.J. (1976). Recovery of function after damage to central catecholamine-containing neurons: A neurochemical model for the lateral hypothalamic syndrome. Prog. Psychobiol. Physiol. Psych. 6: 121-188.
STRICKER, E.M., COOPER. P.H., MARSHALL, J.F. and ZIGMOND, M.J. (1979). Acute homeostatic itialances reinstate sensorimotor dysfunctions in rats with lateral hypothalamic lesions. J. Comp. Physiol. Psych. 93: 512-521.
STRICKER, E.M., FRIEDMAN, Mx., and ZIGMOND, M.J. (1975). Glucoregulatory feeding by rats after intraventricular 6-hydroxydopamine or lateral hypothalamic lesions. Science (Washington) 189: 895-897.
TEITELBAUM, P. %i?i EPSTEIN, A.N. (1962). The lateral hypothalamic syndrome: recovery of feeding and drinking after lateral hypothalamic lesions. Psych. Review 69: 74-90.
THIERRY, A.M., TASSIN, J.P., BLANC, G. and GLOWINSKI, J. (1976). Selective activation of the mesocortical DA system by stress. Nature 263: 242-244.
UNDERSTEDT, U. (1971). Adipsia and aphagia after-hydroxydopamine induced degeneration of the nigro-striatal dopamine system. Acta Physiol. Stand., Suppl. 367: 95-122.
WEISSMAN, A., KOE, B. and TENEN, S. (1966). Anti-amphetamine effects following inhibition of tyrosine hydroxYlase. J. Phanacol. Exp. Therap. 151: 339-352.
ZIGMOND, M.J. and STRICKER, E.M. (1972). -.
Deficits in feeding behavior after intraventricular injection of 6-hydroxydopamine in rats. Science (Washington) 177: 1211-1214.
ZIGMOND, M.J. and STRICKER, E.M. (1973). Recovery of feeding and drinking by rats after intraventricular 6-hydroxydopamine or lateral hypothalamic lesions. Science (Washington) 182: 717-720. -
Inquiries and reprint requests should be addressed to:
Dr. Michael J. Zigmond Department of Biological Sciences University of Pittsburgh Pittsburgh, PA 15260 U.S.A.