the dehydrogenase-mediated recycling of nadph is a key antioxidant system against salt-induced...
TRANSCRIPT

Plant, Cell and Environment
(2006)
29
, 1449–1459 doi: 10.1111/j.1365-3040.2006.01530.x
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd
1449
Role of NADPH in salt-induced oxidative stress in olivesR. Valderrama
et al.
Correspondence: Francisco J. Corpas. Fax:
+
34 958129600; e-mail:[email protected]
The dehydrogenase-mediated recycling of NADPH is a key antioxidant system against salt-induced oxidative stress in olive plants
RAQUEL VALDERRAMA
1
, FRANCISCO J. CORPAS
2
, ALFONSO CARRERAS
1
, MARÍA V. GÓMEZ-RODRÍGUEZ
1
, MOUNIRA CHAKI
1
, JOSÉ R. PEDRAJAS
1
, ANA FERNÁNDEZ-OCAÑA
1
, LUIS A. DEL RÍO
2
& JUAN B. BARROSO
1
1
Grupo de Señalización Molecular y Sistemas Antioxidantes en Plantas, Unidad Asociada al CSIC (EEZ), Departamento de Bioquímica y Biología Molecular, Universidad de Jaén, and
2
Departamento de Bioquímica, Biología Celular y Molecular de Plantas, Estación Experimental del Zaidín, CSIC, Granada, Spain
ABSTRACT
NADPH is an important molecule in the redox balance ofthe cell. In this paper, using olive tissue cultures as a modelof the function of the NADPH-generating dehydrogenasesin the mechanism of oxidative stress induced by severesalinity conditions was studied. When olive (
Olea euro-paea
) plants were grown with 200 m
M
NaCl, a 40% reduc-tion in leaf fresh weight was produced. The content of non-enzymatic antioxidants such as ascorbate and glutathionewas diminished between 20% to 39%, whereas the H
2
O
2
content was increased threefold. In contrast, the analysis ofthe activity and protein contents of the main antioxidativeenzymes showed a significant increase of catalase, superox-ide dismutase and glutathione reductase. Overall, thesechanges strongly suggests that NaCl induces oxidativestress in olive plants. On the other hand, while the contentof glucose-6-phosphate was increased almost eightfold inleaves of plants grown under salt stress, the content ofNAD(P)H (reduced and oxided forms) did not show sig-nificant variations. Under salt stress conditions, the activityand protein contents of the main NADPH-recyclingenzymes, glucose-6-phosphate dehydrogenase (G6PDH),isocitrate dehydrogenase (ICDH), malic enzyme (ME) andferrodoxin-NADP reductase (FNR) showed an enhance-ment of 30–50%. In leaves of olive plants grown with200 m
M
NaCl, analysis of G6PDH by immunocytochemis-try and confocal laser scanning microscopy showed a gen-eral increase of this protein in epidermis, palisade andspongy mesophyll cells. These results indicate that in oliveplants, salinity causes reactive oxygen species (ROS)-medi-ated oxidative stress, and plants respond to this situation byinducing different antioxidative enzymes, especially theNADPH-producing dehydrogenases in order to recycleNADPH necessary for the protection against oxidativedamages. These NADP-dehydrogenases appear to be keyantioxidative enzymes in olive plants under salt stressconditions.
Key-words
:
Olea europaea
; ascorbate peroxidase; confocallaser scanning microscopy; glucose-6-phosphatedehydrogenase; glutathione reductase;
in vitro
tissueculture; monodehydroascorbate reductase; NaCl stress;NADP-dehydrogenases; salinity; superoxide dismutase.
Abbreviations
: AEBSF, 4-(2-aminoethyl)benzenesulfonylfluoride hydrochloride; APX, ascorbate peroxidase; DAR,dehydroascorbate reductase; FNR, ferredoxin-NADPreductase; G6PDH, glucose-6-phosphate dehydrogenase;GR, glutathione reductase; ICDH, NADP-isocitrate dehy-drogenase; ME, malic enzyme; MDAR, monodehy-droascorbate reductase; PMSF, phenylmethylsulfonylfluoride; PVPP, polyvinylpolypyrrolidone; SOD, superox-ide dismutase.
INTRODUCTION
Olive (
Olea europaea
L.) tree is an important species inSouthern Europe with a strong economic impact on theagricultural industry of these countries (Rugini & Fedeli1990; Salas
et al
. 2000). The olive fruit is used to obtain theoil of choice for the Mediterranean diet which is widelyrecognized to have beneficial health effects, particularly oncoronary heart diseases and certain cancer tumours (Alar-con de la Lastra
et al
. 2001; Keys 1995; Owen
et al
. 2000).As a Mediterranean tree, olives are subjected to severeclimate factors such as high solar radiation, drought, drastictemperature changes and high salinity, which can induceoxidative stress situations and important losses in crop pro-ductivity (Sofo
et al
. 2004a,b).Salinity is one of the major abiotic stresses affecting plant
productivity because of its negative effects on plant growth,ion balance and water relations (Hasegawa & Bressan2000). Higher plants have multiple protective mechanismsagainst salt stress including ion homeostasis, osmolytebiosynthesis, reactive oxygen species (ROS) scavenging,water transport and transducers of long-distance responsecoordination (Hasegawa & Bressan 2000). It is generallyaccepted that many stresses, including salinity, induce anoverproduction of ROS, such as hydrogen peroxide, super-

1450
R. Valderrama
et al
.
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd,
Plant, Cell and Environment,
29,
1449–1459
oxide radical, and hydroxyl radicals, and these species arethought to be responsible for the oxidative damage associ-ated with plant stress (Dat
et al
. 2000; Mittler 2002).However, plants possess enzymatic and non-enzymaticantioxidative defence systems to counteract oxidative dam-ages. SODs, catalases, peroxidases, and the ascorbate-glutathione cycle enzymes are examples of antioxidativeenzymes. The ascorbate-glutathione cycle is an efficientway for plant cells to dispose of H
2
O
2
and makes use of thenon-enzymatic antioxidants ascorbate and glutathione in aseries of reactions catalysed by four antioxidative enzymes(APX, MDAR, DAR and GR) (Noctor & Foyer 1998).Under normal conditions, the antioxidative defence systemof plants provides adequate protection against ROS, butwhen the generation of ROS overcomes the defence pro-vided by the plant antioxidant systems, then oxidative stressresults (Noctor & Foyer 1998; Mittler 2002).
The supply of reducing equivalents in the form ofNADPH is one of the most important factors in cell growth,proliferation and detoxification (Barroso
et al
. 1998).NADPH is one of the main end products of several meta-bolic pathways and is also an indispensable substrate forreductive biosynthetic reactions in animal and plants.NADPH is also required in the ascorbate-glutathione cyclefor protection against oxidative damage (Corpas
et al
.1998b; Noctor & Foyer 1998) by thioredoxin reductase inthe regulation of metabolic pathways via thiol group reduc-tion (Nordman
et al
. 2003), and in the formation of activecatalase tetramers, where each catalase monomer containsone bound residue of NADPH (Kirkman & Gaetani 1999).Therefore, the ultimate antioxidant capacity of the cellmust be determined by the availability of reducing equiva-lents. The main enzymes which have the capacity to gener-ate reducing power in the form of NADPH are: G6PDHand 6-phosphogluconate dehydrogenase (6PGDH) (bothbelonging to the pentose phosphate pathway); ME; ICDH;and FNR (Arakaki, Ceccarelli & Carrillo 1997; Corpas1998b, 1999b; Drincovich
et al
. 2001).The aim of this work was to study if salinity causes oxi-
dative stress in olive trees and its relationship with thestatus of the main the NADP-dependent dehydrogenases(G6PDH, ICDH and ME) and FNR. The data obtaineddemonstrate the induction of these NADP-dehydrogena-ses, which allows the recycling of NADPH as a mechanismagainst cellular oxidative damage produced by salinity.Thus, it is proposed that NADP-dehydrogenases are keyantioxidative enzymes in olive plants under salt stress, andthey could also have a protective function in other types ofabiotic stress.
MATERIALS AND METHODS
Plant material and
in vitro
tissue culture conditions
Olive (
Olea europaea
L., cv. Manzanillo) seeds were pro-vided by the Dpto. de Olivicultura y Arboricultura Frutal(CIFA, Córdoba, Spain) and germinated under
in vitro
conditions (Rugini 1984). Seedlings were grown in the darkat 13
°
C for 15 d in an embryos medium and then weretransferred to a Driver-Kuniyuki walnut (DKW) medium(Driver & Kuniyuki 1984). These seedlings were grown ina temperature-controlled chamber at 25
°
C for another30 d, with a 16 h photoperiod under Sylvania Gro-Luxlighting (Sylvania, Westfield, IN, USA) with a photon fluxdensity of 130–140
µ
mol m
−
2
s
−
1
. For the experiments withNaCl, seedlings were transferred to nutrient media with thefollowing NaCl concentrations: 0 (control), 25, 50, 100 and200 m
M
. They were kept during 21 d at the same growthconditions, and then were used for the biochemicalanalyses.
Crude extracts of olive leaves
All operations were performed at 0–4
°
C. Leaves wereground to a powder in a mortar with liquid nitrogen, andwere suspended in 100 m
M
Tris-HCl buffer, pH 8.0 (rela-tion 1/4; w/v) containing 1 m
M
EDTA, 1 m
M
EGTA, 0.1
M
NaCl, 7% (w/v) PVPP, 15 m
M
DTT, 15 m
M
PMSF and acommercial cocktail of proteases inhibitors (AEBSF, 1,10-phenantroline, pepstatine A, leupeptine, bestatine and E-64) (Sigma, St Louis, MO, USA). Homogenates were fil-tered through one layer of miracloth (Calbiochem, SanDiego, CA, USA) and centrifuged at 3000
g
for 5 min.Then, the supernatants were passed through NAP-10 col-umns (Amersham-Biosciences, Piscataway, NJ, USA) thatwere equilibrated and eluted with different buffers depend-ing on the enzymatic assays. Thus, for catalase, SOD, GRactivities and Western-blots, the columns were equilibratedwith 10 m
M
Na-phosphate buffer, pH 6.8, and eluted with10 m
M
K-phosphate buffer, pH 7.8. For the all the NADP-dehydrogenase activities and western-blots, the columnswere equilibrated and eluted with 10 m
M
K-phosphatebuffer, pH 7.6, containing 10 m
M
MgCl
2
, 2 m
M
sodium cit-rate, 0.15
M
NaCl and 14 m
M
β
-mercaptoethanol.
Determination of H
2
O
2
, ascorbate and glutathione
In crude extracts the hydrogen peroxide content wasdetermined by a spectrofluorometric assay, as described byCreissen
et al
. (1999) with some modifications (Romero-Puertas
et al
. 2004). For the determination of the ascorbateand glutathione contents in whole leaves, crude extractswere prepared in 5% (w/v) metaphosphoric acid and themethod of the bipyridyl and the reduction of 5,5
′
-dithiobis-(2-nitrobenzole acid) (DTNB), were used for ascorbateand glutathione, respectively (Griffith 1980; Knörzer,Durner & Böger 1996).
Determination of pyridine dinucleotides (NADPH, NADP, NADH and NAD) and glucose-6-phosphate (G6P) content
Olive leaves were ground to a powder in a mortar withliquid nitrogen, and were suspended in hot 0.1 N NaOHand 0.1 N HCl solutions (1/5; w/v). NADPH and NADH

Role of NADPH in salt-induced oxidative stress in olives
1451
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd,
Plant, Cell and Environment,
29,
1449–1459
are stable in the alkaline solution, while NAD and NADPare stable in the acid solution. The extracts were kept at100
°
C for 2 min, and then were cooled on ice and centri-fuged at 12 000
g
for 6 min. The supernatants obtained wereused to quantify the nucleotides, according to the enzymecycling method described by Matsumura & Miyachi (1980).G6P was determined spectrophotometrically according tothe method of Lang & Michal (1974).
Enzyme activity
Total SOD activity (EC 1.15.1.1) was assayed according tothe ferricytochrome
c
method of McCord & Fridovich(1969). SOD isoenzymes were separated by native-PAGEon 10% acrylamide gels and were localized by a photo-chemical method (Beauchamp & Fridovich 1971). Catalaseactivity (EC 1.11.1.6) was determined by measuring thedisappearance of H
2
O
2
, as described by Aebi (1984). GR(EC 1.6.4.2) activity was assayed by recording the NADPHoxidation, as described by Jiménez
et al
. (1997). G6PDH(EC 1.1.1.49) activity was determined by following thereduction of NADPH according to Corpas
et al
. (1998b).NADP–ICDH activity (EC 1.1.1.42) was measured by fol-lowing the NADPH reduction according to Goldberg &Ellis 1983). The cytochrome
c
reductase activity of FNR(EC 1.18.1.2) was measured spectrophotometrically at550 nm following the reduction of cytochrome
c
(Decot-tignies
et al
. 2003), and the rates were calculated using amolar absorption coefficient of 15 300
M
−
1
cm
−
1
.
Electrophoretic methods and immunoblot analyses
Polypeptides were separated by 10% or 15% SDS-PAGE,and proteins were transferred to polyvinylidene fluoride(PVDF) membranes, as described by Corpas
et al
. (1998b).For immunodetection, polyclonal antibodies against
Sac-charomyces cerevisiae
G6PDH (1/6000 dilution; Sigma), ratliver ME (1/4000 dilution) (Valverde, Benito & Lorenzo1992), cytosolic pea ICDH (1/8000 dilution) (Chen
et al
.1989), pea GR (1/1000 dilution) (Edwards, Rawsthorne &Mullineaux 1990), cucumber MDAR (1/8000 dilution)(Sano
et al
. 1995), cucumber APX (1/3000 dilution) (Corpas& Trelease 1998), cytosolic CuZn-SOD from spinach (1/3000 dilution) (Kanematsu & Asada 1989), pea Mn-SOD(1/2000 dilution), plant Fe-SOD (1/2000 dilution) andpumpkin catalase (1/2000 dilution) (Yamaguchi & Nish-imura 1984) were used. The secondary antibody and thedetection method were those reported by Corpas
et al
.(1998b), and an enhanced chemiluminescence kit (ECL-PLUS, Amersham) was used.
Localization of G6PDH by confocal laser scanning microscopy (CLSM)
Olive leaves from plants growth under control conditionsand with 200 m
M
NaCl were cut into 4–5 mm pieces and
fixed in 4% (w/v) p-formaldehyde in 0.1
M
phosphatebuffer (PB), pH 7.4, for 3 h at room temperature. Then theywere cryoprotected by immersion in 30% (w/v) sucrose inPB overnight at 4
°
C. Serial sections, 60-
µ
m thick, wereobtained by means of a cryostat (2800 Frigocut E, Reichert-Jung, Vienna, Austria). Free floating sections were incu-bated overnight at room temperature with an antibody toyeast G6PDH diluted 1/200 in 5 m
M
Tris-buffer, pH 7.6,0.9% (w/v) NaCl, containing 0.05% (w/v) sodium azide,0.1% (w/v) bovine serum albumin and 0.1% (v/v) Triton X-100 (TBSA-BSAT). After several washes with TBSA-BSAT, sections were incubated with biotinylated goat anti-rabbit IgG (Pierce, Rockford, IL, USA), diluted 1:1000 inTBSA-BSAT, for 1 h at room temperature. Then, sectionswere washed again and incubated with Cy2-streptavidin(Amersham), diluted 1:1000 in TBSA-BSAT, for 1.5 h atroom temperature. Controls for background staining,which was usually negligible, were performed by replacingthe corresponding primary antiserum by pre-immuneserum. Leaf sections were examined with a confocal laserscanning microscope (Leica TCS SL, Leicas Microsystems,Heidelberg GmbH, Wetzlar, Germany).
Other assays
The protein concentration of samples was determined bythe method of Bradford (1976) with bovine serum albumin(BSA) as standard.
RESULTS
Physiological parameters of olive plants under salinity conditions
The growth of olive plants, assessed by fresh weight of leavesand whole plants, showed a reduction proportional to theconcentration of NaCl, and the maximum growth inhibition(about 40%) was obtained at a NaCl concentration of200 m
M
(Table 1). The photosynthetic rate and the chloro-phyll content were diminished by 43% and 58%, respec-tively (Table 2). In these conditions, the oldest leavesbecame yellow and developed necrotic margins, and thelengths of the stems were shorter. All these parameters indi-cate that 200 m
M
NaCl causes a severe stress in olive plants.
Table 1.
Plant growth of
Olea europaea
after 21 d in nutrient media containing NaCl
NaCl (mM) Plant FW (g whole plant) Leaf FW (g plant
−
1
)
0 0.25
±
0.02 0.17
±
0.0225 0.25
±
0.02 0.17
±
0.0250 0.21
±
0.02 0.13
±
0.02100 0.18
±
0.02 0.11
±
0.02200 0.15
±
0.01* 0.09
±
0.01*
Data are the mean
±
SEM of at least 12 different experiments.*Differences from control values were significant at
P
<
0.01.FW, fresh weight.
1452
R. Valderrama
et al
.
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd,
Plant, Cell and Environment,
29,
1449–1459
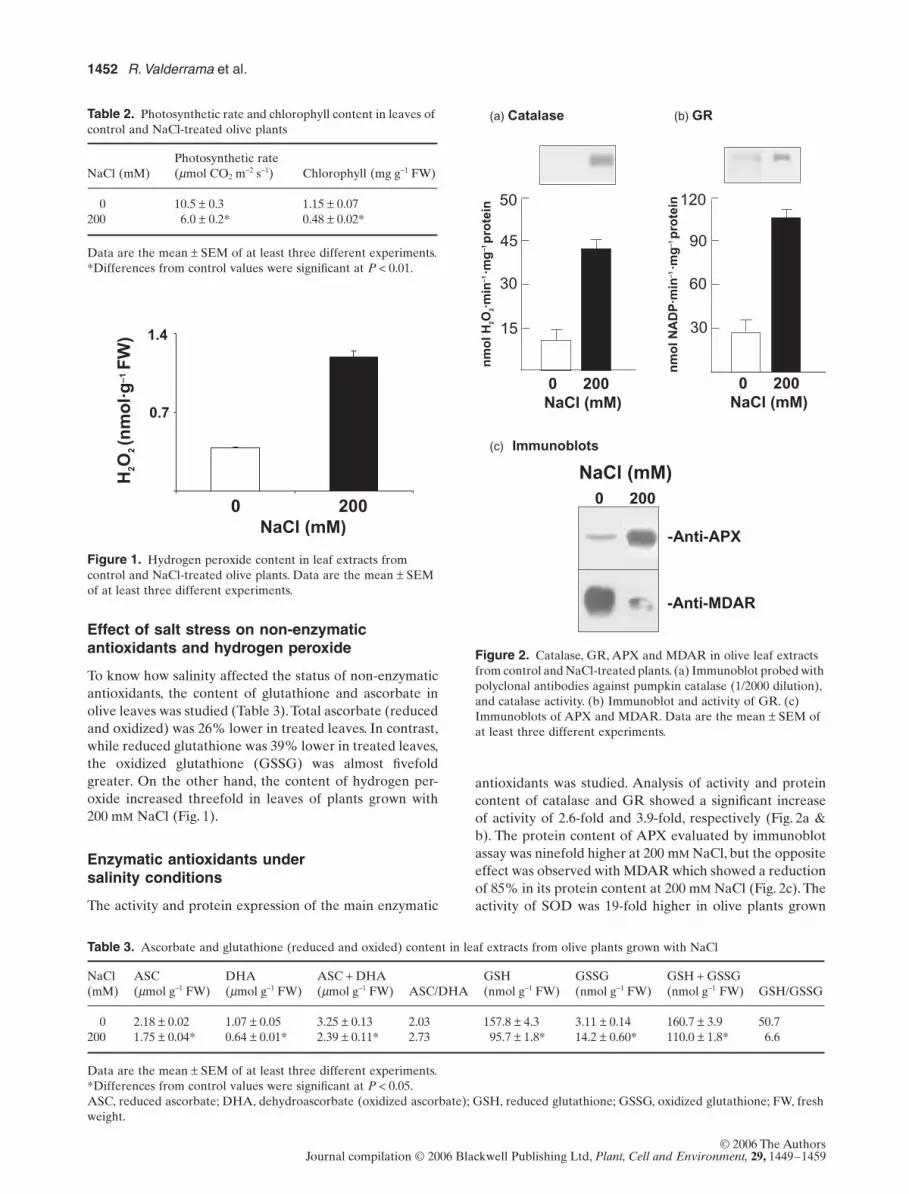
Effect of salt stress on non-enzymatic antioxidants and hydrogen peroxide
To know how salinity affected the status of non-enzymaticantioxidants, the content of glutathione and ascorbate inolive leaves was studied (Table 3). Total ascorbate (reducedand oxidized) was 26% lower in treated leaves. In contrast,while reduced glutathione was 39% lower in treated leaves,the oxidized glutathione (GSSG) was almost fivefoldgreater. On the other hand, the content of hydrogen per-oxide increased threefold in leaves of plants grown with200 m
M
NaCl (Fig. 1).
Enzymatic antioxidants under salinity conditions
The activity and protein expression of the main enzymatic
antioxidants was studied. Analysis of activity and proteincontent of catalase and GR showed a significant increaseof activity of 2.6-fold and 3.9-fold, respectively (Fig. 2a &b). The protein content of APX evaluated by immunoblotassay was ninefold higher at 200 m
M
NaCl, but the oppositeeffect was observed with MDAR which showed a reductionof 85% in its protein content at 200 m
M
NaCl (Fig. 2c). Theactivity of SOD was 19-fold higher in olive plants grown
Table 2.
Photosynthetic rate and chlorophyll content in leaves of control and NaCl-treated olive plants
NaCl (mM)Photosynthetic rate(
µmol CO2 m−2 s−1) Chlorophyll (mg g−1 FW)
0 10.5 ± 0.3 1.15 ± 0.07200 6.0 ± 0.2* 0.48 ± 0.02*
Data are the mean ± SEM of at least three different experiments.*Differences from control values were significant at P < 0.01.
Figure 1. Hydrogen peroxide content in leaf extracts from control and NaCl-treated olive plants. Data are the mean ± SEM of at least three different experiments.
H2O
2 (nm
ol·g
–1 F
W) 1.4
0 200 NaCl (mM)
0.7
Figure 2. Catalase, GR, APX and MDAR in olive leaf extracts from control and NaCl-treated plants. (a) Immunoblot probed with polyclonal antibodies against pumpkin catalase (1/2000 dilution), and catalase activity. (b) Immunoblot and activity of GR. (c) Immunoblots of APX and MDAR. Data are the mean ± SEM of at least three different experiments.
-Anti-APX
-Anti-MDAR
0 200
NaCl (mM)
45
30
15
0 200 NaCl (mM)
50
90
60
30
120
0 200 NaCl (mM)
nm
ol N
AD
P·m
in–1
·mg
–1 p
rote
in
(a) Catalase
(c) Immunoblots
(b) GR
nm
ol H
2O2·m
in–1
·mg
–1 p
rote
in
Table 3. Ascorbate and glutathione (reduced and oxided) content in leaf extracts from olive plants grown with NaCl
NaCl(mM)
ASC(µmol g−1 FW)
DHA(µmol g−1 FW)
ASC + DHA(µmol g−1 FW) ASC/DHA
GSH(nmol g−1 FW)
GSSG(nmol g−1 FW)
GSH + GSSG(nmol g−1 FW) GSH/GSSG
0 2.18 ± 0.02 1.07 ± 0.05 3.25 ± 0.13 2.03 157.8 ± 4.3 3.11 ± 0.14 160.7 ± 3.9 50.7200 1.75 ± 0.04* 0.64 ± 0.01* 2.39 ± 0.11* 2.73 95.7 ± 1.8* 14.2 ± 0.60* 110.0 ± 1.8* 6.6
Data are the mean ± SEM of at least three different experiments.*Differences from control values were significant at P < 0.05.ASC, reduced ascorbate; DHA, dehydroascorbate (oxidized ascorbate); GSH, reduced glutathione; GSSG, oxidized glutathione; FW, freshweight.
Role of NADPH in salt-induced oxidative stress in olives 1453
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Plant, Cell and Environment, 29, 1449–1459
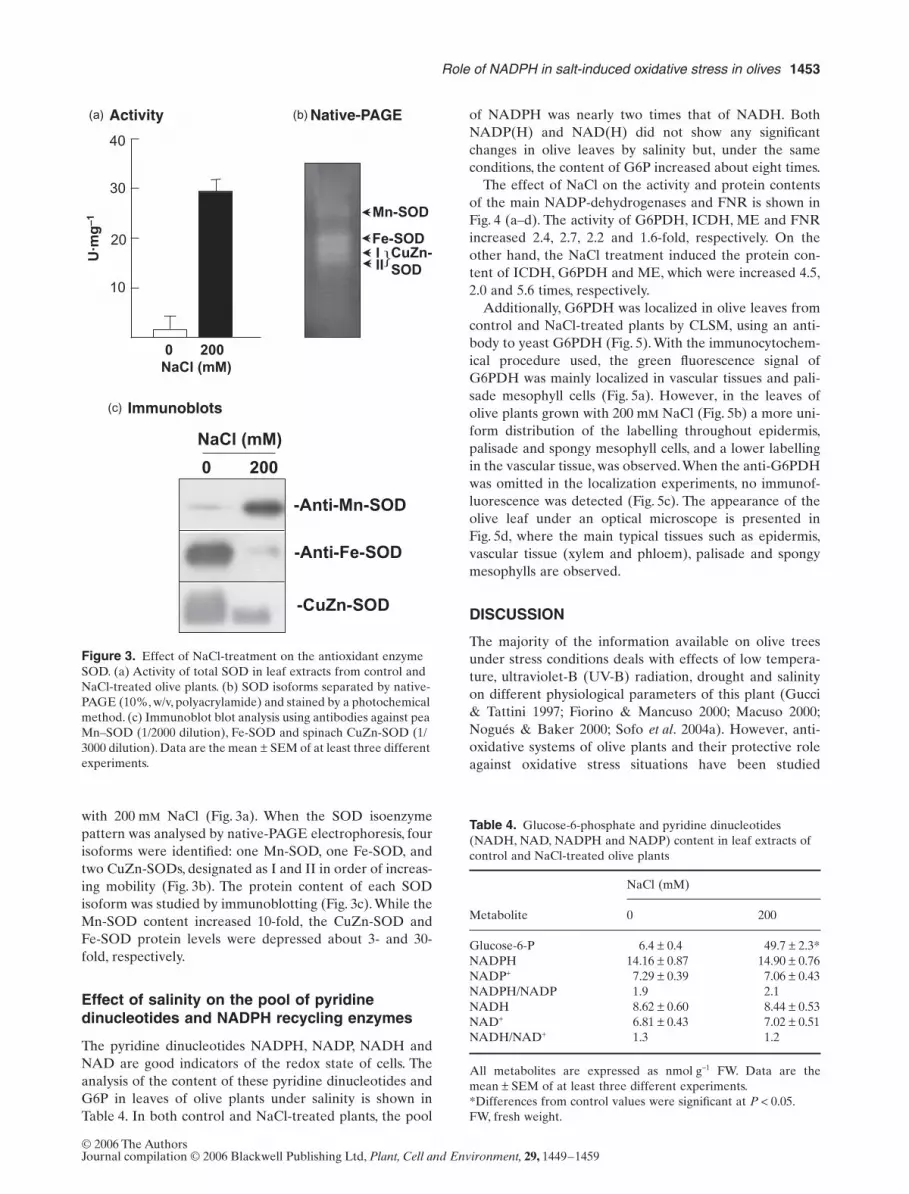
with 200 mM NaCl (Fig. 3a). When the SOD isoenzymepattern was analysed by native-PAGE electrophoresis, fourisoforms were identified: one Mn-SOD, one Fe-SOD, andtwo CuZn-SODs, designated as I and II in order of increas-ing mobility (Fig. 3b). The protein content of each SODisoform was studied by immunoblotting (Fig. 3c). While theMn-SOD content increased 10-fold, the CuZn-SOD andFe-SOD protein levels were depressed about 3- and 30-fold, respectively.
Effect of salinity on the pool of pyridine dinucleotides and NADPH recycling enzymes
The pyridine dinucleotides NADPH, NADP, NADH andNAD are good indicators of the redox state of cells. Theanalysis of the content of these pyridine dinucleotides andG6P in leaves of olive plants under salinity is shown inTable 4. In both control and NaCl-treated plants, the pool
of NADPH was nearly two times that of NADH. BothNADP(H) and NAD(H) did not show any significantchanges in olive leaves by salinity but, under the sameconditions, the content of G6P increased about eight times.
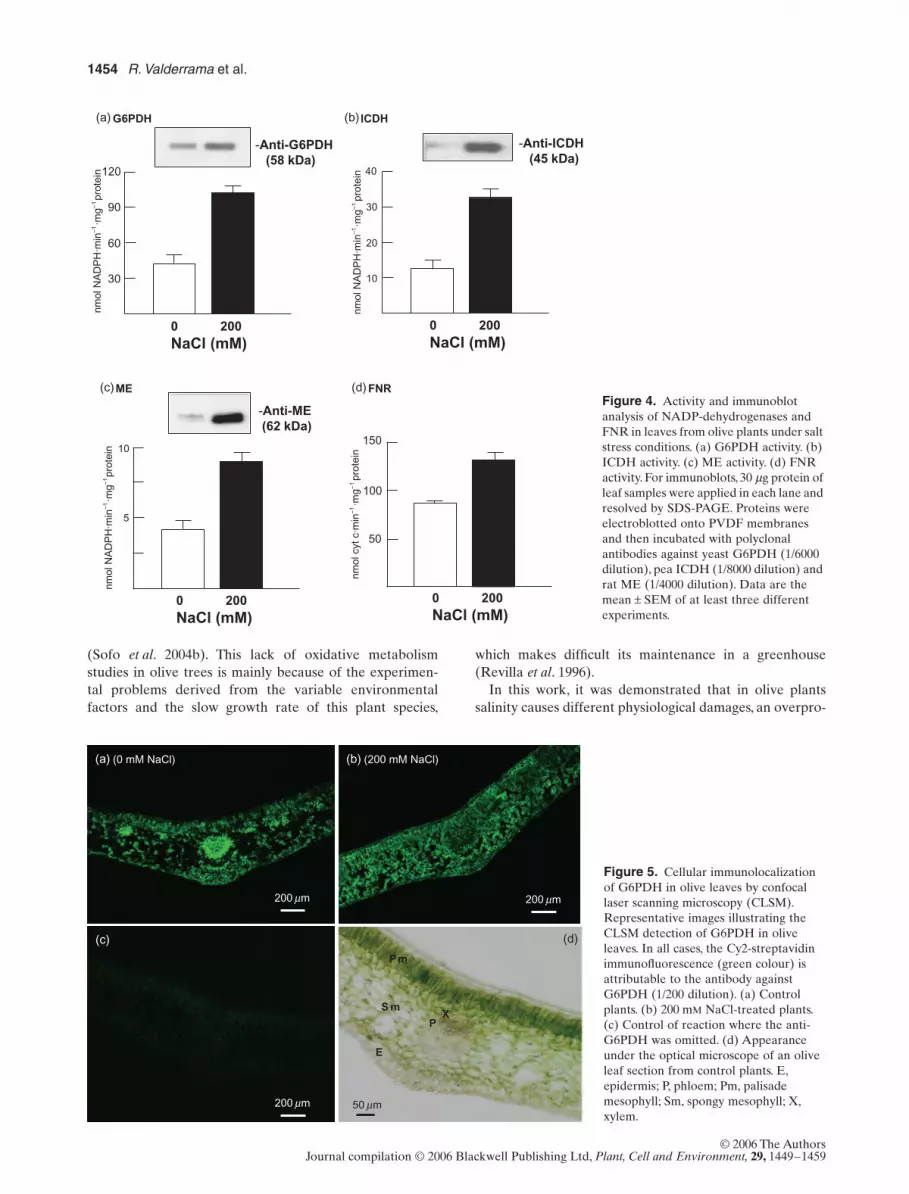
The effect of NaCl on the activity and protein contentsof the main NADP-dehydrogenases and FNR is shown inFig. 4 (a–d). The activity of G6PDH, ICDH, ME and FNRincreased 2.4, 2.7, 2.2 and 1.6-fold, respectively. On theother hand, the NaCl treatment induced the protein con-tent of ICDH, G6PDH and ME, which were increased 4.5,2.0 and 5.6 times, respectively.
Additionally, G6PDH was localized in olive leaves fromcontrol and NaCl-treated plants by CLSM, using an anti-body to yeast G6PDH (Fig. 5). With the immunocytochem-ical procedure used, the green fluorescence signal ofG6PDH was mainly localized in vascular tissues and pali-sade mesophyll cells (Fig. 5a). However, in the leaves ofolive plants grown with 200 mM NaCl (Fig. 5b) a more uni-form distribution of the labelling throughout epidermis,palisade and spongy mesophyll cells, and a lower labellingin the vascular tissue, was observed. When the anti-G6PDHwas omitted in the localization experiments, no immunof-luorescence was detected (Fig. 5c). The appearance of theolive leaf under an optical microscope is presented inFig. 5d, where the main typical tissues such as epidermis,vascular tissue (xylem and phloem), palisade and spongymesophylls are observed.
DISCUSSION
The majority of the information available on olive treesunder stress conditions deals with effects of low tempera-ture, ultraviolet-B (UV-B) radiation, drought and salinityon different physiological parameters of this plant (Gucci& Tattini 1997; Fiorino & Mancuso 2000; Macuso 2000;Nogués & Baker 2000; Sofo et al. 2004a). However, anti-oxidative systems of olive plants and their protective roleagainst oxidative stress situations have been studied
Figure 3. Effect of NaCl-treatment on the antioxidant enzyme SOD. (a) Activity of total SOD in leaf extracts from control and NaCl-treated olive plants. (b) SOD isoforms separated by native-PAGE (10%, w/v, polyacrylamide) and stained by a photochemical method. (c) Immunoblot blot analysis using antibodies against pea Mn–SOD (1/2000 dilution), Fe-SOD and spinach CuZn-SOD (1/3000 dilution). Data are the mean ± SEM of at least three different experiments.
30
20
10
40
0 200 NaCl (mM)
U·m
g–1
Fe-SODIII
Mn-SOD
CuZn-SOD
}
Native-PAGE
(c) Immunoblots
(a) (b) Activity
-Anti-Mn-SOD
-Anti-Fe-SOD
-CuZn-SOD
NaCl (mM)
0 200
Table 4. Glucose-6-phosphate and pyridine dinucleotides (NADH, NAD, NADPH and NADP) content in leaf extracts of control and NaCl-treated olive plants
Metabolite
NaCl (mM)
0 200
Glucose-6-P 6.4 ± 0.4 49.7 ± 2.3*NADPH 14.16 ± 0.87 14.90 ± 0.76NADP+ 7.29 ± 0.39 7.06 ± 0.43NADPH/NADP 1.9 2.1NADH 8.62 ± 0.60 8.44 ± 0.53NAD+ 6.81 ± 0.43 7.02 ± 0.51NADH/NAD+ 1.3 1.2
All metabolites are expressed as nmol g−1 FW. Data are themean ± SEM of at least three different experiments.*Differences from control values were significant at P < 0.05.FW, fresh weight.
1454 R. Valderrama et al.
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Plant, Cell and Environment, 29, 1449–1459
(Sofo et al. 2004b). This lack of oxidative metabolismstudies in olive trees is mainly because of the experimen-tal problems derived from the variable environmentalfactors and the slow growth rate of this plant species,
which makes difficult its maintenance in a greenhouse(Revilla et al. 1996).
In this work, it was demonstrated that in olive plantssalinity causes different physiological damages, an overpro-
Figure 4. Activity and immunoblot analysis of NADP-dehydrogenases and FNR in leaves from olive plants under salt stress conditions. (a) G6PDH activity. (b) ICDH activity. (c) ME activity. (d) FNR activity. For immunoblots, 30 µg protein of leaf samples were applied in each lane and resolved by SDS-PAGE. Proteins were electroblotted onto PVDF membranes and then incubated with polyclonal antibodies against yeast G6PDH (1/6000 dilution), pea ICDH (1/8000 dilution) and rat ME (1/4000 dilution). Data are the mean ± SEM of at least three different experiments.
(b) ICDH
40
30
20
10
0 200 NaCl (mM)
-Anti-ICDH(45 kDa)
120
90
60
30
0 200 NaCl (mM)
(a) G6PDH
-Anti-G6PDH(58 kDa)
10
5
0 200 NaCl (mM)
(c)ME
-Anti-ME(62 kDa)
nmol
NA
DP
H·m
in–1
·mg–1
prot
ein
nmol
NA
DP
H·m
in–1
·mg–1
prot
ein
nmol
NA
DP
H·m
in–1
·mg–1
pro
tein
nmol
cyt
c·m
in–1
·mg–1
pro
tein
150
100
50
(d) FNR
0 200 NaCl (mM)
Figure 5. Cellular immunolocalization of G6PDH in olive leaves by confocal laser scanning microscopy (CLSM). Representative images illustrating the CLSM detection of G6PDH in olive leaves. In all cases, the Cy2-streptavidin immunofluorescence (green colour) is attributable to the antibody against G6PDH (1/200 dilution). (a) Control plants. (b) 200 mM NaCl-treated plants. (c) Control of reaction where the anti-G6PDH was omitted. (d) Appearance under the optical microscope of an olive leaf section from control plants. E, epidermis; P, phloem; Pm, palisade mesophyll; Sm, spongy mesophyll; X, xylem.
(a) (0 mM NaCl)
200 mm
(b) (200 mM NaCl)
200 mm
200 mm
(c) (d)
PX
Pm
S m
E
50 mm

Role of NADPH in salt-induced oxidative stress in olives 1455
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Plant, Cell and Environment, 29, 1449–1459
duction of H2O2 accompanied by a diminution of ascorbateand GSH, and an induction of catalase, SOD, and twoenzymes of the ascorbate-glutahtione cycle (GR and APX).These results indicate that salinity produces oxidative stressin olive plants, which is in agreement with data reported inliterature for pea (Hernández et al. 1993, 1995, 2000) andcitrus plants (Gueta-Dahan et al. 1997), and for differentplants under other types of abiotic stress. In all these cases,abiotic stress induced oxidative damages as a result of over-production of ROS and imbalances of the protective anti-oxidative systems (Dat et al. 2000; Mittler 2002; del Ríoet al. 2002b; Romero-Puertas et al. 2004).
The heme-enzyme catalase is localized in peroxisomes,where its function is to remove toxic hydrogen peroxidegenerated in the peroxisomal respiratory pathway by dif-ferent oxidases (Corpas et al. 1999b; del Río et al. 2002a).In olive leaves, catalase activity and catalase proteinexpression were induced by salinity and this contrasts withthe effect of NaCl in pea leaves, where catalase activity wasdiminished (Corpas et al. 1993). However, the induction ofcatalase activity by other types of abiotic stress has beenalso reported in plants (McClung 1997; Romero-Puertaset al. 1999; Dat et al. 2000; Sofo et al. 2004b). The ascorbate-glutathione cycle is an important antioxidant system whosefunction is to keep H2O2 under control using four differentantioxidative enzymes, plus ascorbate and glutathione(Noctor & Foyer 1998). The growth of olive seedlings withincreasing NaCl concentrations produced opposite effectson the enzymes of the ascorbate-glutathione cycle. TheMDAR protein was decreased by salinity whereas GR andAPX proteins were enhanced. This suggests that in leavesfrom olive plants cultured in vitro, GR and APX could bekey detoxifying enzymes under conditions of salt stress, asituation in which MDAR must have a different mechanismof regulation. In leaf chloroplasts from pea plants treatedwith NaCl, a reduction in the MDAR activity was reported,whereas in leaf mitochondria the opposite effect wasobserved (Gómez et al. 1999), indicating that the regulationof this enzyme could be associated with its subcellular local-ization. In pea plants subjected to abiotic stress by lowtemperature and wounding, an increase in the RNA expres-sion of the peroxisomal MDAR was observed, while treat-ment with the herbicide 2,4-dichlorophenoxyacetic acid (2-4-D) caused a reduction in the expression of this peroxiso-mal enzyme (Leterrier et al. 2005). The induction of GRactivity and protein in olive leaves by salinity is in agree-ment with the induction of GR activity reported in peaplants under NaCl stress (Gómez et al. 1999; Hernándezet al. 1999). On the other hand, in leaves of pea plantsexposed to different abiotic stress conditions, the GR iso-forms responded differently (Romero-Puertas et al. 2006).Thus, while the RNA expression of the chloroplastic GRwas repressed by high light, high temperature and darkness,it was induced by wounding and low temperature; the cyto-solic GR was repressed by high light and darkness and wasonly induced by low temperature. On the other hand, treat-ment of pea plants with 50 µM CdCl2 did not affect theexpression of both chloroplastic and cytosolic GR, and nei-
ther the total GR activity nor the specific protein contentwere affected by Cd (Romero-Puertas et al. 2006). On thecontrary, the activity of the peroxisomal GR was consider-ably increased by Cd (Romero-Puertas et al. 2006). In anycase, the different response of APX and GR versus MDARseems to indicate a differential regulation of these antioxi-dative enzymes and could be a result of the existence ofisoforms present in different subcellular compartments.
The SOD activity is responsible for the scavenging of O2–
radicals, and in olive leaves salinity induced a strongincrease of this activity, which accords with the datareported in other plant species, including pea (Hernándezet al. 1995, 1999), citrus and rice cell cultures (Fadzilla,Finch & Burdon 1997; Gueta-Dahan et al. 1997), andtomato (Shalata et al. 2001). Similar response has beendescribed in olive leaves during drought stress (Sofo et al.2004b). In this work, it was observed that the different SODisoenzymes had a different response to NaCl stress. Whilethe Mn-SOD increased, the CuZn-SOD and Fe-SOD werediminished, an inverse situation to that reported for theSODs of pea plants grown under Mn deficiency, where theMn-SOD activity was depressed but the activity of CuZn-SODs was enhanced (del Río et al. 1978). The reason forthis response could be also related to the different subcel-lular localization of these enzymes. In different plant spe-cies Mn-SOD has been demonstrated to be present inmitochondria and peroxisomes (del Río et al. 2002a, 2003),and the enzyme activity increase produced by NaCl in oliveleaves suggests that mitochondrial and/or peroxisomal Mn-SOD could be induced to protect against superoxide radi-cals overproduced in these cell organelles. In leaf mito-chondria from pea plants subjected to NaCl stress, a rise inthe generation of superoxide radicals, protein level andactivity of mitochondrial Mn-SOD was reported (Hernán-dez et al. 1993, 1999). In higher plants Fe-SOD is mainlylocalized in chloroplasts (Salin 1988; Van Camp et al. 1996;Navari-Izzo et al. 1999; Gómez et al. 2004) and CuZn-SOD,although it occurs chiefly in chloroplasts, is also present inthe cytosol, peroxisomes and apoplast (Ogawa, Kanematsu& Asada 1996, 1997; Sandalio et al. 1997; Corpas et al.1998a; del Río et al. 2002a; Gómez et al. 2004). The strongreduction in the Fe-SOD and CuZn-SOD protein expres-sion in olive leaves could be a result of the damages pro-duced in the thylakoidal structure of chloroplasts by theNaCl treatment (data not shown). These ultrastructuraldamages were also reported in chloroplasts from pea plantsunder salt stress (Hernández et al. 1995). However, the datareported in this paper on olive leaf Fe-SOD contrasts withthose obtained in leaves from pea plants, where there wasan increase in the activity and protein expression of thechloroplastic Fe-SOD after long-term NaCl stress (Gómezet al. 2004).
In recent years, the antioxidative nature of plant NADP-dehydrogenases resulting from their recycling of NADPH,which is essential for the functioning of the ascorbate-glu-tathione cycle, has become increasingly evident (Corpaset al. 1998b, 1999a; Noctor & Foyer 1998). In olive leavesunder salt stress, the content of NADPH did not change,

1456 R. Valderrama et al.
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Plant, Cell and Environment, 29, 1449–1459
and this is well correlated with the general induction of theactivity of the four analysed enzymes (G6PDH, ICDH, MEand FNR). Induction of ICDH and G6PDH has beenreported in pea and pepper plants subjected to Cd stress(Romero-Puertas et al. 1999; León et al. 2002), and inMesembryanthemum crystallinum plants by salt stress (Pop-ova et al. 2002). In Eucalyptus plants, the induction of MEby NaCl has been also described (Farias De Aragao et al.1997).
To evaluate if salinity could affect the cellular distribu-tion of NADP-dehydrogenases in olive leaves, G6PDH wasselected as a marker because previously this dehydroge-nase was demonstrated to play important roles in theresponse to H2O2 stress in yeast (Izawa et al. 1998) andtobacco (Debnam et al. 2004). At subcellular level thisenzyme has been reported to be present in the cytosol,chloroplasts and peroxisomes of pea leaves (Corpas et al.1998b; Wakao & Benning 2005). However, to our knowl-edge, there is a lack of information on a possible differentialdistribution of G6PDH in distinct types of cells. The cellularimmunolocalization of G6PDH in olive leaves, obtained byCLSM, revealed that, under normal conditions, this enzymewas mainly expressed in vascular tissues and palisade mes-ophyll cells, but in response to salt treatment G6PDHexhibited a more homogenous distribution in all types ofleaf cells. This is in good agreement with the increase ofactivity and protein expression of this enzyme as well as theG6P content during salt stress. To our knowledge, this is thefirst report on the localization of G6PDH in vascular tissue,and its function therein could be related with the physio-logical significance of this enzyme. Recently, two additionalantioxidant enzymes, CuZn-SOD and MDAR, have beenreported in vascular tissue of cucumber and pumpkin,respectively, and these enzymes were increased by droughtstress (Walz et al. 2002). An important function of G6PDHin vascular tissue could be connected with the NO-produc-ing nitric oxide synthase (NOS) activity. This enzyme activ-ity catalyses the oxygen- and NADPH-dependent oxidationof L-arginine to NO and citrulline in a complex reactionrequiring different cofactors (del Río, Corpas & Barroso2004). In pea leaves, the presence of a high NOS specificactivity was detected in leaf peroxisomes (Barroso et al.1999), and an important accumulation of NOS-derivednitric oxide (NO) was found in vascular tissues of leaves,stems and roots (Corpas et al. 2004, 2006). The presence ofG6PDH in these tissues would be very important to recyclethe NADPH necessary to carry out the NOS-dependentNO generation. Moreover, the pentose phosphate pathway– where G6PDH is involved – apart from the NADPHbiosynthesis, can supply other essential intermediates suchribose-5-phosphate or erytrose-4-phosphate. These sugarphosphates are needed for the biosynthesis of lignin, nucle-otides, aromatic amino acids and coenzymes, and thesecompounds perhaps could be involved in the mechanism ofresponse to salt stress.
FNR is a very well-known enzyme present in chloroplastswhich mediates the electron transfer between ferredoxinand NADP during photosynthesis (Arakaki, Ceccarelli &
Carrillo 1997). Although FNR has been reported to be amember of the cellular defence against oxidative damagein Escherichia coli (Bianchi et al. 1995), little is knownabout the response of FNR to oxidative stress in plants. Inthe facultative halophyte M. crystallinum (ice plant), it wasreported that the FNR expression did not change withsalinity (Michalowski, Schmitt & Bohnert 1989). Likewise,in wheat leaves treated with the oxidative stress-inducermethyl viologen, no effects on the expression of FNR wereobserved (Palatnik, Valle & Carrillo 1997). In contrast,results obtained in this work showed a significant increaseof the FNR activity by salt stress, indicating that thisenzyme could participate in the response of olive leaves tooxidative damage.
In conclusion, as judged by the imbalances produced inthe level of non-enzymatic and enzymatic antioxidant sys-tems, and the increase in the H2O2 production, resultsobtained in this work indicate that in olive plants NaClinduces ROS-mediated oxidative stress in leaves. More-over, there is a general induction of NADPH-producingdehydrogenases and FNR, which contribute to maintaina high NADPH/NADP ratio. This indicates that NADP-dehydrogenases are key antioxidative enzymes in oliveplants under salt stress, and suggests that the dehydroge-nase-mediated recycling system of NADPH perhaps couldbe a general mechanism of response of plants against dif-ferent abiotic and biotic stresses.
ACKNOWLEDGMENTS
This work was supported by the UJA (grant 0B.10.02.12),MCYT (grant AGL200305524) and Junta de Andalucía(research groups CVI 286 and 192), Spain. Olive seeds werekindly provided by the Dpto. de Olivicultura y Arboricul-tura Frutal, Banco de Germoplasma Mundial, CIFA, Cór-doba. The valuable help of Prof. Fernando Pliego and DrAraceli Barceló (CIFA of Churriana, Málaga, Spain) insetting up the in vitro tissue culture conditions of oliveplants is appreciated. We acknowledge Dr M. Nishimura(National Institute for Basic Biology, Okazaki, Japan), DrK. Asada (Fukuyama University, Japan), Dr P.M. Mul-lineaux (University of Essex, UK), Dr Pierre Gadal(Université de Paris-Sud, Orsay Cedex, France) and DrÁngela M. Valverde (Universidad Complutense, Madrid)for their generous gifts of antibodies against pumpkin cat-alase, cucumber MDAR, pea GR, pea ICDH and rat ME,respectively. We thank Dr Manuel Gómez (Estación Exper-imental del Zaidín, CSIC, Granada) for his advice on thephotosynthetic rate analysis and Miss Emperat Córdoba forthe maintenance of in vitro plant cultures. The CLSManalyses were carried out at the Technical Services of theUniversity of Jaén.
REFERENCES
Aebi H. (1984) Catalase in vitro. Methods Enzymology 105, 121–126.

Role of NADPH in salt-induced oxidative stress in olives 1457
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Plant, Cell and Environment, 29, 1449–1459
Alarcon de la Lastra C., Barranco M.D., Motilva V. & HerreriasJ.M. (2001) Mediterranean diet and health: biological impor-tance of olive oil. Current Pharmaceutical Design 7, 933–950.
Arakaki A.K., Ceccarelli E.A. & Carrillo N. (1997) Plant-typeferredoxin-NADP reductases: a basal structural framework anda multiplicity of functions. Federation of American Societies forExperimental Biology Journal 11, 133–140.
Barroso J.B., Peragon J., Contreras-Jurado C., Garcia-Salguero L.,Corpas F.J., Esteban F.J., Peinado M.A., De La Higuera M. &Lupiáñez J.A. (1998) Impact of starvation-refeeding on kineticsand protein expression of trout liver NADPH-production sys-tems. American Journal of Physiology 274, R1578–R1587.
Barroso J.B., Corpas F.J., Carreras A., Sandalio L.M., ValderramaR., Palma J.M., Lupiáñez J.A. & del Río L.A. (1999) Localiza-tion of nitric-oxide synthase in plant peroxisomes. Journal ofBiological Chemistry 274, 36729–36733.
Beauchamp C.O. & Fridovich I. (1971) Superoxide dismutase:improved assays and an assay applicable to acrylamide gels.Analytical Biochemistry 44, 276–287.
Bianchi V., Haggard-Ljungquist E., Pontis E. & Reichard P. (1995)Interruption of the ferredoxin (flavodoxin) NADP+ oxidoreduc-tase gene of Escherichia coli does not affect anaerobic growthbut increases sensitivity to paraquat. Journal of Bacteriology177, 4528–4531.
Bradford M.M. (1976) A rapid and sensitive method for the quan-titation of microgram quantities of protein utilizing the principleof protein-dye binding. Analytical Biochemistry 72, 248–254.
Chen R., Bismuth E., Chapigny M.L. & Gadal P. (1989) Chro-matographic and immunological evidence that chloroplastic andcytosolic pea (Pisum sativum L.) NADP-isocitrate dehydroge-nases are distinct isoenzymes. Planta 178, 157–163.
Corpas F.J. & Trelease R.N. (1998) Differential expression ofascorbate peroxidase and a putative molecular chaperone in theboundary membrane of differentiating cucumber seedling per-oxisomes. Journal of Plant Physiology 153, 332–338.
Corpas F.J., Gómez M., Hernández J.A. & del Río L.A. (1993)Metabolism of activated oxygen in peroxisomes from two Pisumsativum L cultivars with different sensitivity to sodium chloride.Journal of Plant Physiology 141, 160–165.
Corpas F.J., Sandalio L.M., del Río L.A. & Trelease R.N. (1998a)Copper-zinc superoxide dismutase is a constituent enzyme of thematrix of peroxisomes in the cotyledons of oilseed plants. TheNew Phytologist 138, 307–314.
Corpas F.J., Barroso J.B., Sandalio L.M., Distefano S., Palma J.M.,Lupiáñez J.A. & del Río L.A. (1998b) A dehydrogenase-mediated recycling system of NADPH in plant peroxisomes.Biochemical Journal 330, 777–784.
Corpas F.J., Barroso J.B., Sandalio L.M., Palma J.M., LupiáñezJ.A. & del Río L.A. (1999a) Peroxisomal NADP-dependentisocitrate dehydrogenase. Characterization and activity regula-tion during natural senescence. Plant Physiology 121, 921–928.
Corpas F.J., Palma J.M., Sandalio L.M., López-Huertas E.,Romero-Puertas M.C., Barroso J.B. & del Río L.A. (1999b)Purificación of catalase from pea leaf peroxisomes: Identifica-tion of five different isoforms. Free Radical Research 31(Suppl.), S235–S241.
Corpas F.J., Barroso J.B., Carreras A., et al. (2004) Cellular andsubcellular localization of endogenous nitric oxide in young andsenescent pea plants. Plant Physiology 136, 2722–2733.
Corpas F.J., Barroso J.B., Carreras A., Valderrama R., Palma J.M.,León A.M., Sandalio L.M. & del Río L.A. (2006) Constitutivearginine-dependent nitric oxide synthase activity in differentorgans of pea seedlings during plant development. Planta 233,DOI 10.1007/s00425-005-0205-9.
Creissen G., Firmin J., Fryer M., et al. (1999) Elevated glutathionebiosynthetic capacity in the chloroplasts of transgenic tobacco
plants paradoxically causes increased oxidative stress. Plant Cell11, 1277–1292.
Dat J., Vandenabeele S., Vranová E., van Montagu M., Inzé D. &van Breusegem F. (2000) Dual action of the active oxygen spe-cies during plant stress responses. Cellular and Molecular LifeScience 57, 779–795.
Debnam P.M., Fernie A.R., Leisse A., Golding A., Bowsher C.G.,Grimshaw C., Knight J.S. & Emes M.J. (2004) Altered activityof the P2 isoform of plastidic glucose 6-phosphate dehydroge-nase in tobacco (Nicotiana tabacum, cv. Samsun) causes changesin carbohydrate metabolism and response to oxidative stress inleaves. Plant Journal 38, 49–59.
Decottignies P., Flesch V., Gérard-Hirne C. & Le Maréchal P.(2003) Role of positively charged residues in Chlamydomonasreinhardtii feredoxin-NADP-reductase. Plant Physiological Bio-chemistry 41, 637–642.
Drincovich M.F., Casati P. & Andreo C.S. (2001) NADP-malicenzyme from plants: a ubiquitous enzyme involved in differentmetabolic pathways. FEBS Letters 490, 1–6.
Driver J.A. & Kuniyuki A.H. (1984) In vitro propagation of par-adox walnut root-stock. Horticultural Science 19, 507–509.
Edwards E.A., Rawsthorne S. & Mullineaux P.M. (1990) Subcel-lular distribution of multiple forms of gluthathione reductase inleaves of pea (Pisum sativum L.). Planta 180, 278–284.
Fadzilla N.M., Finch R.P. & Burdon R.H. (1997) Salinity, oxidativestress and antioxidant responses in shoot cultures of rice. Journalof Experimental Botany 48, 325–331.
Farias De Aragao M.E., Silva Lima M.G., Fernandes De Melo D.& Dizengremel P. (1997) NaCl-induced changes of NAD (P)malic enzyme activities in Eucalyptus citriodora leaves. Trees 12,66–72.
Fiorino P. & Mancuso S. (2000) Differential termal análisis, super-cooling and cell viability in organs of Olea europaea at subzerotemperatures. Advances in Horticultural Science 14, 23–27.
Goldberg D.M. & Ellis G. (1983) Isocitrate. In Methods of Enzy-matic Analysis, 3rd edn (ed. H.U. Bergmeyer), pp. 183–190.Academic Press, New York, USA.
Gómez J.M., Hernández J.A., Jiménez A., del Río L.A. & SevillaF. (1999) Differential response of antioxidative enzymes of chlo-roplasts and mitochondria to long-term NaCl stress of peaplants. Free Radical Research 31, S11–S18.
Gómez J.M., Jiménez A., Olmos E. & Sevilla F. (2004) Locationand effects of long-term NaCl stress on superoxide dismutaseand ascorbate peroxidase isoenzymes of pea (Pisum sativum cv.Puget) chloroplasts. Journal of Experimental Botany 55, 119–130.
Griffith O.W. (1980) Determination of glutathione and glutathionedisulfide using glutathione reductase and 2-vynykpyridines.Analytical Biochemistry 106, 207–212.
Gucci R. & Tattini M. (1997) Salinity tolerance in olive. Horticul-tural Review 21, 177–214.
Gueta-Dahan Y., Yaniv Z., Zilinskas B.A. & Ben-Hayyim G.(1997) Salt and oxidative stress: similar and specific responsesand their relation to salt tolerance in citrus. Planta 203, 460–469.
Hasegawa P.M. & Bressan R.A. (2000) Plant cellular and molec-ular responses to high salinity. Annual Review of Plant Physiol-ogy and Plant Molecular Biology 51, 463–499.
Hernández J.A., Corpas F.J., Gómez M., del Río L.A. & SevillaF. (1993) Salt-induced oxidative stress mediated by activatedoxygen species in pea leaf mitochondria. Physiologia Plantarum89, 103–110.
Hernández J.A., Olmos E., Corpas F.J., Sevilla F. & del Río L.A.(1995) Salt-induced oxidative stress in chloroplasts of pea plants.Plant Science 105, 151–167.
Hernández J.A., Campillo A., Jiménez A., Alarcón J.J. & Sevilla

1458 R. Valderrama et al.
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Plant, Cell and Environment, 29, 1449–1459
F. (1999) Response of antioxidant systems and leaf waterrelations to NaCl stress in pea plants. New Phytologist 141, 241–251.
Hernández J.A., Jiménez A., Mullineaux P.M. & Sevilla F. (2000)Tolerance of pea (Pisum sativum L.) to long-term salt stress isassociated with induction of antioxidant defences. Plant, Cell &Environment 23, 853–862.
Izawa S., Maeda K., Miki T., Mano J., Inoue Y. & Kimura A.(1998) Importance of glucose-6-phosphate dehydrogenase in theadaptive response to hydrogen peroxide in Saccharomyces cer-evisiae. Biochemical Journal 330, 811–817.
Jiménez A., Hernández J.A., del Río L.A. & Sevilla F. (1997)Evidence for the presence of the ascorbate-glutathione cycle inmitochondria and peroxisomes of pea leaves. Plant Physiology114, 275–284.
Kanematsu S. & Asada K. (1989) CuZn-superoxide dismutasesfrom the fern Equisetum arvense and the green alga Soirogyrasp. ocurrence of chloroplast and cytosol types of enzyme. PlantCell Physiology 30, 717–727.
Keys A. (1995) Mediterranean diet and public heath: personalreflexions. American Journal of Clinical Nutrition 61, 1321S–1323S.
Kirkman H.N. & Gaetani G.F. (1999) Catalase: a tetramericenzyme with four tightly bound molecules of NADPH. Proceed-ing of the National Academy of Sciences of the USA 81, 4343–4347.
Knörzer O.C., Durner J. & Böger P. (1996) Alterations in theantioxidative systems of suspension-cultured soybean cells (Gly-cine max) induced by oxidative stress. Physiologia Plantarum97, 399–396.
Lang G. & Michal G. (1974) D-glucose-6-phosphate and D-fruc-tose-6-phosphate. In Methods in Enzymatic Analysis, Vol. 2 (ed.H.U. Bergmeyer), pp. 1238–1242. Academic Press, Inc, NewYork, NY, USA.
León A.M., Palma J.M., Corpas F.J., Gómez M., Romero-PuertasM.C., Chatterjee D., Mateos R.M., del Río L.A. & SandalioL.M. (2002) Antioxidative enzymes in cultivars of pepper plantswith different sensitivity to cadmium. Plant Physiology Bio-chemistry 40, 813–820.
Leterrier M., Corpas F.J., Barroso J.B., Sandalio L.M. & del RíoL.A. (2005) Peroxisomal monodehydroascorbate reductase.Genomic clone characterization and functional analysis underenvironmental stress conditions. Plant Physiology 138, 2111–2123.
McClung C.R. (1997) Regulation of catalases in Arabidopsis. FreeRadical Biology & Medicine 23, 489–496.
McCord J.M. & Fridovich I. (1969) Superoxide dismutase: an enzy-mic function for erythrocuprein (hemocuprein). Journal of Bio-logical Chemistry 244, 6049–6055.
Macuso S. (2000) Electrical resistance changes during exposure tolow temperature measure chilling and freezing tolerance in olivetree (Olea europaea L.) plants. Plant, Cell & Environment 23,291–299.
Matsumura H. & Miyachi S. (1980) Cycling assay for nicotinamideadenine dinucleotides. Methods Enzymology 69, 465–470.
Michalowski C.B., Schmitt J.M. & Bohnert H.J. (1989) Expressionduring salt stress and nucleotide-sequence of cDNA for ferre-doxin-NADP reductase from. Mesembryanthemum-Crystalli-num Plant Physiology 89, 817–822.
Mittler R. (2002) Oxidative stress, antioxidants and stress toler-ance. Trends in Plant Science 7, 405–410.
Navari-Izzo F., Pinzino C., Quartacci M.F. & Sgherri L.M. (1999)Superoxide and hydroxyl radical generation, and superoxidedismutase in PSII membrane fragments from wheat. Free Radi-cal Research 31 (Suppl.), S3–S9.
Noctor G. & Foyer C.H. (1998) Ascorbate and glutathione: keep-
ing active oxygen under control. Annual Review of Plant Phys-iology Plant Molecular Biology 49, 249–279.
Nogués S. & Baker N.R. (2000) Effects of drought on photosyn-thesis in Mediterranean plants grown under enhanced UV-Bradiation. Journal of Experimental Botany 51, 1309–1317.
Nordman T., Xia L., Bjorkhem-Bergman L., Damdimopoulos A.,Nalvarte I., Arner E.S., Spyrou G., Eriksson L.C., BjornstedtM. & Olsson J.M. (2003) Regeneration of the antioxidantubiquinol by lipoamide dehydrogenase, thioredoxin reductaseand glutathione reductase. Biofactors 18, 45–50.
Ogawa K., Kanematsu S. & Asada K. (1996) Intra- and extra-cellular localization of ‘cytosolic’ CuZn-superoxide dismutase inspinach leaf and hypocotyls. Plant Cell Physiology 37, 790–799.
Ogawa K., Kanematsu S. & Asada K. (1997) Generation of super-oxide anion and localization of CuZn-superoxide dismutase inthe vascular tissue of spinach hypocotyls: their association withlignification. Plant Cell Physiology 38, 1118.1126.
Owen R.W., Giacosa A., Hull W.E., Haubner R., Wurtele G.,Spiegelhalder B. & Bartsch H. (2000) Olive-oil consumptionand health: the possible role of antioxidants. Lancet Oncology1, 107–112.
Palatnik J.F., Valle E.M. & Carrillo N. (1997) Oxidative stresscauses ferredoxin-NADP reductase solubilization from thethylakoid membranes in methyl viologen-treated plants. PlantPhysiology 115, 1721–1727.
Popova O.V., Ismailov S.F., Popova T.N., Dietz K.J. & GolldackD. (2002) Salt-induced expression of NADP-dependentisocitrate dehydrogenase and ferredoxin-dependent glutamatesynthase in Mesembryanthemum crystallinum. Planta 215, 906–913.
Revilla M.A., Pacheco J., Casares A. & Rodríguez R. (1996) Invitro reinvigoration of mature olive trees (Olea europaea L.)through micrografting. In Vitro Cellular & Developmental Biol-ogy Plant 32, 257–261.
del Río L.A., Sevilla F., Gómez M., Yáñez J. & López-Gorgé J.(1978) Superoxide dismutase: an enzyme system for the studyof micronutrient interactions in plants. Planta 140, 221–225.
del Río L.A., Corpas F.J., Sandalio L.M., Palma J.M., Gómez M.& Barroso J.B. (2002a) Reactive oxygen species, antioxidantsystems and nitric oxide in peroxisomes. Journal of Experimen-tal Botany 53, 1255–1272.
del Río L.A., Sandalio L.M., Palma J.M., Corpas F.J., López-Huertas E., Romero-Puertas M.C. & McCarthy I. (2002b) Per-oxisomes, reactive oxygen metabolism, and stress-relatedenzyme activities. In Plant Peroxisomes Biochemistry, CellBiology and Biotechnological Applications (eds A. Baker & I.Graham), pp. 221–258. Kluwer Academic Publishers, Dor-drecht, The Netherlands.
del Río L.A., Sandalio L.M., Altomare D.A. & Zilinskas B.A.(2003) Mitochondrial and peroxisomal manganese superoxidedismutase: differential expression during leaf senescence. Jour-nal of Experimental Botany 54, 923–933.
del Río L.A., Corpas F.J. & Barroso J.B. (2004) Nitric oxide andnitric oxide synthase activity in plants. Phytochemistry 65, 783–792.
Romero-Puertas M.C., McCarthy I., Sandalio L.M., Palma J.M.,Corpas F.J., Gómez M. & del Río L.A. (1999) Cadmium toxicityand oxidative metabolism of pea leaf peroxisomes. Free RadicalResearch 31 (Suppl.), S25–S31.
Romero-Puertas M.C., Rodríguez-Serrano M., Corpas F.J.,Gómez M., del Río L.A. & Sandalio L.M. (2004) Cadmium-induced subcellular accumulation of O2
· − and H2O2 in pealeaves. Plant Cell & Environment 27, 1122–1134.
Romero-Puertas M., Corpas F.J., Sandalio L.M., Leterrier M.,Rodríguez-Serrano M., del Río L.A. & Palma J.M. (2006) Glu-tathione reductase from pea leaves: response to abiotic stress

Role of NADPH in salt-induced oxidative stress in olives 1459
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Plant, Cell and Environment, 29, 1449–1459
and characterization of the peroxisomal isozyme. New Phytolo-gist 170, 43–52.
Rugini E. (1984) In vitro propagation of some olive (Olea europaeasativa L.) cultivars with different root ability, and medium devel-opment using analytical data from developing shoots andembryos. Scientia Horticulturae 24, 123–134.
Rugini E. & Fedeli E. (1990) Olive (Olea europaea L.) as anoilseed crop. In Biotechnology in Agriculture & ForestryLegumes & Oilseed I (ed. Y.P.S. Bajaj), pp. 593–641. Springer-Verlag, Berlin, Germany.
Salas J.J., Sánchez J., Ramli U.S., Manaf A.M., Williams M. &Harwood J.L. (2000) Biochemistry of lipid metabolism in oliveand other oil fruits. Progress in Lipid Research 39, 151–180.
Salin M.L. (1988) Toxic oxygen species and protective systems ofthe chloroplast. Physiologia Plantarum 72, 681–689.
Sandalio L.M., López-Huertas E., Bueno P. & del Río L.A. (1997)Immunocytochemical localization of copper,zinc superoxide dis-mutase in peroxisomes from watermelon (Citrullus vulgarisSchrad) Free Radical Research 26, 187–194.
Sano S., Miyake C., Mikami B. & Asada K. (1995) Molecularcharacterization of monodehydroascorbate radical reductasefrom cucumber highly expressed in Escherichia coli. Journal ofBiological Chemistry 270, 21354–21361.
Shalata A., Mittova V., Volokita M., Guy M. & Tal M. (2001)Response of the cultivated tomato and its wild salt-tolerant rel-ative Lycopersicon pennellii to salt-dependent oxidative stress:the root antioxidative system. Physiologia Plantarum 112, 487–494.
Sofo A., Dichio B., Xiloyannis C. & Masia A. (2004a) Lipoxyge-
nase activity and proline accumulation in leaves and roots ofolive trees in response to drought stress. Physiologia Plantarum121, 58–65.
Sofo A., Dichio B., Xiloyannis C. & Masia A. (2004b) Antioxidantdefences in olive tree during drought stress: changes in activityof some antioxidant enzymes. Functional Plant Biology 32, 45–53.
Valverde A.M., Benito M. & Lorenzo M. (1992) Hormonal regu-lation of malic enzyme and glucose-6-phosphate dehydrogenaseexpression in fetal brown adipocyte primary cultures under non-proliferative conditions. European Journal of Biochemistry 203,313–319.
Van Camp W., Capiau K., van Montagu M., Inzé D. & Slooten L.(1996) Enhancement of oxidative stress tolerance in transgenictobacco plants overexpressing Fe-superoxide dismutase in chlo-roplasts. Plant Physiology 112, 1703–1714.
Wakao S. & Benning C. (2005) Genome-wide analysis of glucose-6-phosphate dehydrogenases in Arabidopsis. Plant Journal 41,243–256.
Walz C., Juenger M., Schad M. & Kelr J. (2002) Evidence for thepresence and activity of a complete antioxidant defence systemin mature sieve tubes. Plant Journal 31, 189–197.
Yamaguchi J. & Nishimura M. (1984) Purification of glyoxysomalcatalase and immunochemical comparison of glyoxysomal andleaf peroxisomal catalase in germinating pumpkin cotyledons.Plant Physiology 262, 261–267.
Received 24 November 2005; received in revised form 6 February2006; accepted for publication 13 February 2006