the comparative ultrastructure of spermatozoa from five branchiobdellidans (annelida: clitellata)
TRANSCRIPT

The comparative ultrastructure of spermatozoa from five branchiobdellidans (Annelida: Clitellata)
M. FERRAGUTI Departimento di Biologia, Universita degli Studi di Milano, Via Celoria 26, 20133 Milano, Italy
AND
S. R. GELDER Department of Biology, University of Maine, 181 Main Street, Presque Isle, ME 04769, U.S.A.
Received July 12, 1990
FERRAGUTI, M., and GELDER, S. R. 199 1. The comparative ultrastructure of spermatozoa from five branchiobdellidans (Annelida: Clitellata). Can. J. Zool. 69: 1945-1956.
The ultrastructure of mature spermatozoa from five species of branchiobdellidans is described. The species come from four genera, and the three major families in the Branchiobdellida (Annelida: Clitellata) are all represented. The spermatozoa of Branchiobdella pentodonta and B. astaci (Branchiobdellidae) have a very long helical acrosome and a helical marginal fiber surrounding the flagellum, whereas those of Xironogiton instabilis show a twisted acrosome similar to that found in some oligochaetes. Bdellodrilus illuminatus (Bdellodrilidae) is extremely atypical, with no true acrosome, and with a long helical end- piece. Cambarincola fallax (Cambarincolidae) has an acrosome with a helical ridge on the acrosome tube, as in hirudineans, but shares with Branchiobdella spp. a helical marginal fiber around the flagellum. The spermatozoan characters in these species display a mixture of oligochaete and hirudinean features, as well as some autapomorphies, thus dispelling the idea that branchiobdellidan spermatozoa are very similar. Although a unique model cannot yet be defined for branchiobdellidan spermatozoa, a number of characters do show promise. These observations are still too few to allow any inference of evolutionary polarity in the Branchiobdellida or Clitellata, but they do suggest the possibility of using the comparative ultrastructure of the spermatozoa as an additional tool for studying branchiobdellidan phylogeny.
FERRAGUTI, M., et GELDER, S, R. 199 1. The comparative ultrastructure of spermatozoa from five branchiobdellidans (Annelida: Clitellata). Can. J. Zool. 69 : 1945-1956.
L'ultrastructure des spermatozo'ides a maturitt a t t t ttudite chez cinq espttces de ~ranchiobdellida. Les espttces appartiennent a quatre genres, et chacune des trois familles de Branchiobdellida (Annelida: Clitellata) est reprtsentke. Les spermatozo'ides de Branchiobdella pentodonta et de B. astaci (Branchiobdellidae) ont un acrosome htlicoldal tres long et une fibre marginale htlico'idale qui entoure le flagelle, alors que les spermatozoi'des de Xironogiton instabilis ont un acrosome tordu semblable a celui qui prtvaut chez certains oligochtttes. Les spermatozo'ides de Bdellodrilus illuminatus (Bdellodrilidae) sont tout a fait particuliers puisqu'il n'ont pas d'acrosome et qu'il comportent une pittce terminale longue et htlicoi'dale. Cambarincola fallax (Cambarincolidae) a un acrosome a rebord htlicoi'dal sur le tube, comme chez les hirudints, mais possttde, comme chez Branchiobdella spp., une fibre marginale htlicoi'dale autour du flagelle. Les spermatozo'ides de cette esp&ce ont des caracttristiques de spermatozoi'des d'oligochtttes et de spermatozo'ides d'hirudints, et possttdent aussi des autapomorphies, et il faut doc rejeter I'hypothttse selon laquelle les spermatozo'ides des Branchiobdellida sont tous semblables. 11 n'est pas encore possible de dtfinir un modttle type de spermatazoi'de pour ce groupe, mais un certain nombre de caracttristiques s'avttrent trtts prometteuses. Ces observations sont encore trop peu nombreuses pour permettre de supposer l'existence d'une polaritt tvolutive chez les Branchiobdellida ou les Clitellata, mais elles laissent tout de m2me entrovoir la possibilitt d'utiliser l'ultrastructure des spermatozo'ides comme outil additionnel dans les analyses phylogtnttiques des Branchiobdellida.
[Traduit par la rtdaction]
Introduction Since the pioneering work of Franzkn (1956) and the subse-
quent development of comparative spermatology (Baccetti 1970), the use of spermatozoon morphology as a tool for taxonomic and phylogenetic studies is now widely accepted, despite some continued misgivings (Mohri 1987). It has been shown that a set of spermatozoon morphological characters can be used to define various higher levels of taxa in the same way as conventional anatomical. characters.
The superclass Clitellata in the Annelida consists of four classes, Oligochaeta, Hirudinea, Branchiobdellida, and Acanth- obdellida, following the organization used by Brinkhurst and Gelder (1991). Current knowledge of clitellate spermatozoa at the ultrastructural level is based largely on a few representatives from the Oligochaeta and Hirudinea (Ferraguti 1983). The character states typical of the spermatozoa from these two taxa have been defined by Jamieson et al. (1987) and Garavaglia et al. (1974), respectively. An attempt to define branchiobdellidan spermatozoon autapomorphies on the basis of observations from Rinted in Canada i Imprime au Canada
two species proved inconclusive (Ferraguti et al. 1986), and spermatozoa from the Acanthobdellida remain to be described.
Clitellate spermatozoa are long, filiform cells with an acro- some tube, a long thin nucleus, and mitochondria interposed between the nucleus and the tail. The acrosome tube has a wide range of shapes and dimensions. It contains the acrosome vesicle with subacrosomal material in which an acrosome rod (= per- foratorium) is visible. A conventional centriole is not present in .the mature spermatozoon, as a result of the high degree of modification .that occurs during spermiogenesis (Ferraguti 1984~). The axoneme has a prominent central sheath apparently autapomorphic for clitellates, though absent in some of the more advanced forms of oligochaetes (Ferraguti 1984b), and glycogen granules. Some of these features are also found in other, higher animal taxa. For instance, an acrosome tube may be present in vestimentiferan pogonophores (Jamieson 1987), although Gardiner and Jones (1985) refute this. The positions of the mitochondria and of the central tubule of the centriole (basal cylinder sensu Ferraguti 19846) are the same as in onycopho-

1946 CAN. J. ZOOL. VOL. 69, 1991
rans, and the phylogenetic implications of these characters were discussed by Jamieson (1983). Similarly, glycogen granules are present in some molluscs, polychaetes, and arthropods (Anderson and Personne 1970).
The Branchiobdellida contains 2 1 genera with approximately 150 species (Gelder and Brinkhurst 1990). Descriptions of spermatozoa from this taxon are few, and of these, most are from the genus Branchiobdella. At the light-microscopic level the spermatozoa of Branchiobdella parasita Henle, 1823 and B. hexodonta Gruber, 1883 (incorrectly identified as B. astaci Odier, 1823) were reported to have screw-shaped, thread-like portions (Dorner 1865). Subsequently, the B. astaci spermato- zoon was described by Ostroumoff (1883). FranzCn (1962) is the only person to have described spermatozoa from a North American species, Xironogiton instabilis (Moore, 1893). Until recently, only the spermatozoon of B. pentodonta had been studied at the ultrastructural level, and consequently was the representative spermatozoon of the taxon (Bondi and Farnesi 1976). The spermatozoon of Branchiobdella pentodonta, as de- scribed in this paper, appears to be highly aberrant with respect to the accepted clitellate pattern, e.g., the different acrosome parts and the persistence of the microtubular manchette in the ripe sperm. Subsequent investigations by Ferraguti and Lanzavecchia (1977) and Ferraguti et al. (1986) provided the first clear descriptions of B. pentodonta spermatozoa. The acrosome in this species is amongst the largest reported in the animal kingdom. The a&osomal rod was identified by Ferraguti (1983), having been described previously as the "cork" by Ferraguti and Lanzavecchia (1977). This structure is located at the base of the acrosome tube and is enclosed in a short second- ary tube. These features, together with the highly atypical marginal fiber helically wound around the axoneme, suggest a model of branchiobdellidan spermatozoon that, though possess- *ing the basic clitellate characters, is considered highly apomor- phic (Ferraguti 1983).
To assess the similarity of the B. pentodonta spermatozoon to that of one other species in the taxon, spermatozoa were studied from Cambarincola fallax Hoffman, 1963, a widely distributed North American species (Ferraguti et al. 1986). However, only a few of the apomorphisms of the B. pentodonta spermatozoon were also found in C. fallax: the marginal fiber around the axoneme, the concavity on top of the nucleus, and the extreme reduction of the secondary tube. Curiously, the pitch of the acrosome tube ridge in C. fallax is similar to that of the corkscrew-shaped acrosome tube of B. pentodonta. In general, the acrosome of C. fallax is similar to that found in oligochaetes, except that it has a helical ridge around the acrosome tube which makes it reminiscent of a hirudinean spermatozoon.
The phylogenetic position of the Branchiobdellida in the Clitellata, based on anatomical characters, is debatable. Some authorities assign the taxon to the Oligochaeta (Avel 1959; Timm 198 1 ), others assign it to the Hirudinea (Sawyer 1986) or support an independent group, equal in taxonomic rank to the former two (Holt 1989). Brinkhurst and Gelder (199 1) currently accept the last theory, that the branchiobdellidans evolved independently, along with the oligochaetes and leeches, from a common ancestral clitellate stock. The branchiobdellidans and lumbriculid oligochaetes do share a number of anatomical simi- larities in their reproductive organs (Avel 1959; Brinkhurst and Gelder 1989). However, comparable similarities were not found in the ultrastructure of the spermatozoa and muscles between the two taxa (FranzCn 1962; Ferraguti and Lanzavecchia 1977). The phylogeny within the Branchiobdellida was addressed by Holt
(1986), who arranged the genera into five families. Gelder and Brinkhurst (1990) demonstrated that the taxon was a monophyly by means of anatomical characters of the component genera in a Wagner analysis method (phylogenetic analysis using parsi- mony, of D. Swofford, Illinois Natural History Museum, Champaign). A number of the genera in that study were not resolved, owing to a lack of characters in the data base. A study of the comparative spermatology of the branchiobdellidans is expected to provide characters that will fully resolve all the genera in the taxon, either in their own right or in conjunction with existing anatomical characters.
Before this project is embarked upon, however, at least one representative species from each of the three major branchiob- dellidan families (Holt 1986) should be described at the ultra- structural level, and a preliminary character suite created. Existing preliminary information (Feraguti et al. 1986) consisted of the European B. pentodonta from the Branchiobdellidae and the North American C. fallax for the Cambarincolidae. To complete the exploratory study, the North American Bdello- drillus illuminatus (Moore, 1893) was chosen to represent the Bdellodrilidae, and Xironogiton instabilis (Moore, 1893) provided a second example from the Branchiobdellidae. The opportunity to compare intraspecific differences in spermatozoon ultrastructure was. provided when B. astaci was obtained. Only the information from Ferraguti et al. (1986) on the mature spermatozoa from B. pentodonta and C. fallax is included here, to place the new observations in context and to allow all the data to be discussed in a coherent manner.
Material and methods All the branchiobdellidans were collected from live crayfish.
Branchiobdella pentodonta were obtained on Australopotamobius pallipes collected from the River Po, Province of Piacenza, and B. astaci on the same host species from the River Rosandra, Province of Trieste, Italy. Bdellodrilus illuminatus, C. fallax, and X. instabilis were removed at various times during the summer from Cambarus bartonii bartonii collected from the Presque Isle Stream, Aroostook County, Maine, U.S.A. Specimens were removed from the host and identified, and then fixed without dissection for transmission electron microscopi- cal study, using a variety of fixatives, including Karnowsky's fixative (1965) with 0.1 M cacodylate buffer (pH 7.4) for B. pentodonta, Ernick and Eakin's (1976) fixative modified for fresh water (picric acid - paraformaldehyde - glutaraldehyde mixture) for B. astaci, and Karnowsky 's fixative diluted 1 :3 with 0.1 M cacodylate buffer (pH 7.4) for C. fallax, X. instabilis, and B. illuminatus. Additional specimens of C. fallax were fixed in McDowell's (1978) phosphate buffer (pH 7.2). Fixation lasted for 6 h for Branchiobdella spp. and ranged from 3 to 13 days for the others. Specimens were washed overnight in the respective buffer solution, postfixed in 1 % osmium tetroxide in the same buffer for 2 h, washed in distilled water, en bloc stained for a few hours in the dark in 2% aqueous uranyl acetate, and dehydrated in a graded ethanol series. Branchiobdella pentodonta was embedded in an Epon-Araldite mixture, and for all the other species Spurr's resin was used. Sections were cut with an LKB ultrotome I11 or V, and thick (about 1 ~ m ) sections were cut with an optical microscope to locate mature spermato- zoa. Thin sections were stained with lead citrate or with the triple stain following the Daddow (1 983) method, mounted on carbon-coated grids, and observed with either a Hitachi HU 11 ES, a JEOL 100B, or a JEOL 100 XS transmission electron microscope.
Observations When sperm maturation is completed in clitellates, the
spermatozoa crowd into the ciliated funnels of the deferent ducts. The following descriptions are based on the spermatozoa at these locations. All branchiobdellidan spermatozoa examined so far

FERRAGUTI AND GELDER
TABLE 1 . Mean dimensions of the major structures of the spermatozoa in four species of branchiobdellidans
X . instabilis C . fallax B . illurninatus B . pentodonta
Acrosome Length Diameter
Nucleus Length Diameter
Mid-piece Length Diameter
Length of flagellum
Length of end-piece
Total length
Absent
(Max.) 258.8
NOTE: All measurements are given in micrometres. The acrosomes and nuclei are tapered, so the diameters are given for both extremities
are filiform, with an acrosome, a nucleus, a mid-piece containing only mitochondria, and a tail. The total length of the spermato- zoa and the relative dimensions of the different portions vary among the species. The dimensions of these characters are given in Table 1.
Xironogiton instabilis The spermatozoon of X. instabilis has a tapering, helical
acrosome that makes 1.5 revolutions (Figs. 1,46). The acrosome tube has a characteristic wall of uneven thickness all along its length (Figs. 4-6) and the posterior end has a small acentric aperture (Fig. - 1). In longitudinal section this appears as an internal lip or shelf (= limen; Jamieson 1983) of varying length. The acrosome vesicle makes contact with the plasma membrane only through the open anterior tip of the tube (Fig. 2). The vesicle fills the anterior half of the tube, and there is a deep invagination from the vesicle's base (Figs. 3,4,6). The posterior margin of the vesicle extends posteriorly into a short secondary tube (Fig. 6). The vesicle invagination and the secondary tube surround the anterior half of the acrosomal rod. The posterior half of the rod is often bent to one side, and is surrounded by part of the elongated electron-lucent material that fills the rest of the acrosome tube (= basal chamber; Jamieson 1983) (Fig. 6). The nucleus is helical, with a pitch of 0.7 km (Fig. 1 l), and tapers from a diameter of 0.33 km posteriorly to 0.15 km anteriorly; both ends are concave (Figs. 1, 7). The nucleus is followed by four mitochondria slightly twisted around the central axis of the spermatozoon (Figs. 7, 8).
The tail consists of a flagellum and an end-piece. The flagel- lum starts with a centriolar area in which it is possible to see the remnants of the centriole (Fig. 9). The latter loses its conven- tional appearance during spermiohistogenesis (M. Ferraguti, unpublished data), to give rise in the mature spermatozoa to a short electron-dense ring in which a ninefold symmetry can be recognized. A central apparatus, similar to that of the flagella, passes through the center of the centriole remnant and ends in contact with the mitochondria (Fig. 7).
The main portion of the flagellum contains a prominent central sheath surrounding the central tubules, multiple-link heads, and pairs of glycogen granules along the external length of the axoneme (Figs. 10, 12). The tail ends in a tapering end-piece
which is composed of homogeneous electron-dense material and devoid of any axonemal structure (Fig. 13).
Cambarincola fallax In the spermatozoon of C. fallax (Fig. 47) the straight
acrosome tube tapers from posterior to anterior before bending 90" and enlarging (Fig. 14). A hollow ridge, continuous with the tube wall, winds around the tube for 1.5 revolutions (Fig. 14, arrowhead). The acrosome vesicle is enclosed by the tube except at the anterior tip, where the vesicle passes through the enlarged end and contacts the plasma membrane in an apical button (Fig. 16). Dense, fibrous "fuzzy" material (= apical corona; Ferraguti and Jamieson 1987) is sometimes present on the surface of the apical button (Fig. 16, arrowhead). An invagination starts from the posterior end of the acrosome vesicle and extends anteriorly to the level at which the helical ridge starts (Figs. 17, 47). The invagination surrounds the acrosome rod or perforatorium, which is difficult to recognize as it is "embedded" in electron-dense subacrosomal material (Fig. 18). At the distal end of the acrosome rod is a spherical structure containing disorganized, tightly packed membranes (Fig. 17). A similar structure in the form of a ring is located at the proximal end of the acrosome vesicle that surrounds the posterior invagination site (Fig. 18, arrowhead).
The nucleus has a conical concavity at the anterior end (Fig. 18) and a shallow concavity at the posterior end (Fig. 19). The four mitochondria in the mid-piece are slightly twisted about the central axis of the spermatozoon (Figs. 19, 21).
The tail consists of a flagellum and an end-piece. The flagellum starts from a short, cup-shaped electron-dense struc- ture in contact with the mitochondria (Figs. 19,21). Although a true centriole is absent in this and other branchiobdellidan species, the origin of the modified centriole has been confirmed by following the structure's development during spermiogenesis (M. Ferraguti, unpublished observations). The central apparatus of the flagellum penetrates this centriolar remnant and reaches the mitochondria (Figs. 19,21). The axoneme has a prominent central sheath, multiple-link heads, and glycogen granules. A helical structure made of electron-dense material runs around the tubules of the flagellum (pitch about 1.7 km) and under the plasma membrane (Figs. 20, 22). The helical ridge starts

1948 CAN. J. ZOOL. VOL. 69, 1991
just posterior to the mid-piece (Fig. 15, arrowhead). The tail terminates in a short non-axonemal end-piece about 1 km long (Fig. 23).
Bdellodrilus illurninatus The spermatozoon of this species (Fig. 48) is characterized by
a short conical "acrosome" filled with homogeneous electron- dense material and surrounded by the plasma membrane (Figs. 24,26,27,32). Occasionally the apex of the acrosome is charac- teristically bent to one side (Fig. 32). In the absence of any ultrastructural evidence, it is called the acrosome only because of its position anterior to the nucleus.
The nucleus is anteriorly rectilinear for about 4 km and then slightly helical for the rest of its length (Figs. 33, 28). No concavity is visible at the anterior end of the nucleus, but one is present at the posterior end (Figs. 27,25). Four mitochondria are tightly packed together and slightly twisted about the longitudi- nal axis of the spermatozoon (Figs. 25,29).
The tail consists of a long flagellum and an unusual end-piece. The i'lagellum starts with a deeply modified "centriolar area" represented by a highly electron-dense, hollow cylinder with a diameter and length of 0.25 km. In the walls of this cylinder are nine electron-lucent spots arranged in the same position. as the triplets (Figs. 25, 30). The structure is derived from a conven- tional centriole and becomes modified in the course of spermio- genesis (M. Ferraguti, unpublished observations). Inside this centriolar remnant a ceptral structure is present, similar to the one described in other clitellates. This structure continues along the flagellum, forming a prominent central sheath, and glycogen granules are present in their usual position (Fig. 3 1). The twisted end-piece is a triangular prism (Figs. 34,35), with each surface. approximately 0.16 km wide and 1 10 km long, and with a pitch of about 1.7 km. Electron-dense material is arranged in a thick rod down the center and three thin rods lying along each edge of the prism.
Branchiobdella pentodonta and Branchiobdella astaci The spermatozoa of both B. pentodonta and B. astaci are very
long, the former being 258.8 km (Table 1). The Branchiobdella astaci spermatozoon, according to Ostroumoff (1 883), has an anterior spiral part, presumably the acrosome and nucleus, which is 9 km long, and a threadlike tail 3 10 km long, giving a total length of 319 km. Ultrastructural details of the B. astaci spermatozoon are reported here for the first time. Additional details for B. pentodonta (see Ferraguti et al. 1986) are given here so that a complete representative for this genus can be illustrated (Fig. 49) and compared with the other genera. The two species have the same ultrastructural characters; however, they do differ from each other as stated above.
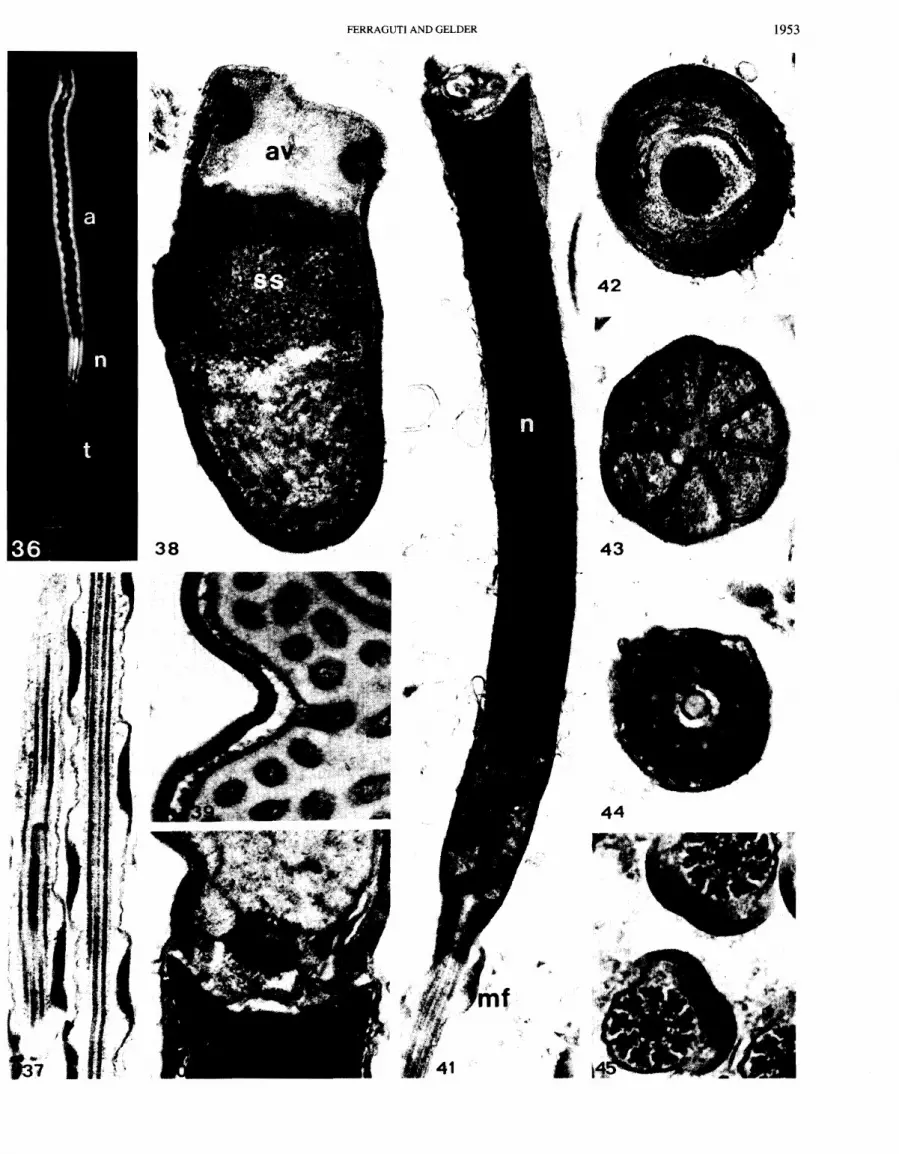
The acrosome is one of the largest in the animal kingdom (Fig. 36). It is composed of a twisted acrosome tube, about 1 km wide and 33 km long, which contains the acrosome vesicle. The posterior end of the vesicle contains a discrete concentration of multilayered membranes surrounding a central invagination (Fig. 42). Almost filling this aperture is a spherical electron-dense body (= "cork") with a very short secondary tube (Fig. 40). It is not always visible, but starts at the proximal end of the acrosome vesicle. The acrosomal vesicle extends anteriorly, with the outer membrane in contact with the acrosome tube and the inner membrane folding repeatedly. Each fold extends towards the interior to form small flattened tubules (Fig. 39). The folding ceases in the anterior region and the vesicle protrudes slightly beyond the acrosomal tube to touch the plasma membrane (Fig. 38). A ring of electron-dense material is located within the vesicle, in the slight protuberance (Fig. 38). The "subacrosomal space" is the region between the tightly packed vesicle tubules, and the material becomes more electron-dense as it approaches the anterior end of the acrosomal tube.
The nucleus is short and tapers posteriorly, with a concavity in both anterior and posterior surfaces (Fig. 41). The mid-piece contains seven short mitochondria arranged and twisted about the central axis of the spermatozoon (Figs. 41,43).
The long tail consists of a flagellum, but an end-piece is absent. Immediately posterior to the mitochondria is the cen- triolar region consisting of a cylinder 0.4 km long and 0.22 km
FIGS. 1-13. Xironogiton instabilis spermatozoa. Fig. 1. A rare longitudinal section of the acrosome. Note the limen (1) of the acrosome tube, forming an acentric aperture (arrowhead). X 90 000. Fig. 2. The tip of the acrosome, showing dense material inside the acrosome vesicle. X 100 000. Fig. 3. Cross section of the area shown in Fig. 2. X 100 000. Fig. 4. The infolding at the base of acrosome vesicle containing the rod. X 100 000. Fig. 5. Cross section of the acrosome, showing the rod surrounded only by the acrosome tube. X 100 000. Fig. 6. Longitudinal section of two acrosomes, showing the rods (r) and the short secondary tubes (arrowhead). X 100 000. Fig. 7. The mid-piece in longitudinal section. The prominent central sheath (cs) penetrates the centriolar remnant (cr) to reach the mitochondria (m). X 100 000. Fig. 8. The four mitochondria in the mid-piece. X 90 000. Fig. 9. An area close to the centriolar remnant. X 100 000. Fig. 10. A typical cross section of the flagellum. X 100 000. Fig. 1 1. Mature spermatozoa at the funnel Cf) entrance X7000. Fig. 12. The tails in longitudinal section, showing the prominent central sheath and the radial links (arrowhead) but no marginal fiber. X 18 000. Fig. 13. End-piece of the tail X 57 000.
FIGS. 14-23. Cambarincola fallax spermatozoa. Fig. 14. Acrosome with the helical ridge of the tube (arrowhead). X 30 000. Fig. 15. Mid-piece and first tract of the flagellum. The marginal fiber (mf) starts at approximately 2.5 pm from the centriolar remnant (arrowhead). X 30 000. Fig. 16. Anterior extremity of the acrosome, with button (b) and apical corona (vowhead). X 100 000. Fig. 17. Invagination of the acrosome vesicle, with the spherical structure (s). X 100 000. Fig. 18. Acrosome base, with the confused membranous termination of the vesicle (arrowhead). Note the concavity of the nucleus (n) and the acrosomal rod (ar). X 100 000. Fig. 19. Mid-piece and centriolar remnant (arrowhead). n, nucleus; cs, central sheath. X 100 000. Fig. 20. Longitudinal section of the flagellum. Note the marginal fiber in cross section (small arrowhead) and longitudinal section (large arrowhead). X 93 000. Fig. 21. The mitochondria (m), the centriolar remnant (arrowhead), an end-piece (ep), and the cilia of the funnel (c) in cross section X 100 000. Fig. 22. A typical cross section of the flagellum, with the small marginal fiber. X 100 000. Fig. 23. End-piece of the flagellum. X 100 000.
FIGS. 24-35. Bdellodrilus illuminatus spermatozoa. Fig. 24. Anterior extremity of the nucleus (n), with the "acrosome." X 100 000. Fig. 25. Posterior extremity of the nucleus, mid-piece, and centriolar remnant (arrowhead). X 60 000. Fig. 26. Cross section of the "acrosome." X 120 000. Fig. 27. Base of the "acrosome"; even at high magnification, no substructure is visible. Only the plasma membrane of the cell surrounds the organelle. X 160 000. Fig. 28. Cross section of the nucleus. X 120 000. Fig. 29. Cross section of the mitochondria. X 120 000. Fig. 30. Centriolar remnant in cross section. X 120 000. Fig. 3 1. Cross section of the flagellum. X 120 000. Fig. 32. Some "acrosomes" are characteristically bent. x 112 000. Fig. 33. Spermatozoa at the funnel entrance. Note the straight portion of the nucleus (arrowheads) following the "acrosome." X 20 000. Fig. 34. A rare image of the transition between flagellum and end-piece (diamond). X 75 000. Fig. 35. Cross section of some transition areas. Five end-pieces are visible in cross section at the bottom. Dense material reminiscent of the marginal fibers of other branchiobdellidans forms a triangular helix (compare with Fig. 34). X 80 000.

FERRAGUTI AND GELDER 1949

1950 CAN. J . ZOOL. VOL. 69, 1991

FERRAGUTI AND GELDER 1951

1952 CAN. J. ZOOL. VOL. 69, 1991
wide (Figs. 41,44). In cross section this centriolar remnant has an indistinct ninefold symmetry. At its center, a structure continuous with the central apparatus of the flagellum extends to reach the mitochondria. The axoneme has the basic clitellate appearance, with a helical marginal fiber of electron-dense material lying just under the plasma membrane of the flagellum (Fig. 45). The marginal fiber has a pitch of about 1.6 pm (Fig. 37).
Discussion Clitellate characters in branchiobdellidan spermatozoa
There are only two autapomorphic ultrastructural characters that define a clitellate spermatozoon. The first is the acrosome tube which contains an acrosome vesicle and s~~bacrosomal material (Ferraguti 1983). Other characters, such as the position of the mitochondria between the nucleus and flagellum, are shared with the onychophorans (Jamieson 1986), and the basal cylinder penetrating the centriole is present in many oligo- chaetes. This latter feature was proposed as an additional clitellate character by Jamieson (1986), but is shared with the onychophorans. However, even if the connection between the basal cylinder and central sheath is not clear, the morphological difference between the two structures is evident. Based on current literature and supported by unpublished observations (M.F.), the basal cylinder is limited, among the clitellate groups, to the spermatozoa of oligochaetes.
The second autapomchphic character is the electron-dense, central sheath in the flagellum, which was previously named the "prominent central sheath" by Ferraguti (1984b). This structure is present in hirudineans and branchiobdellidans, and contrary to Sawyer (1 986), also in many oligochaetes. Interpretation of the phylogentic lineage of the central sheath is not easy, since it appears to be absent from the ''megadriles" and from some advanced tubificids. In these two groups it is replaced by another axonemal modification, the so-called tetragon fibers. However, both the central sheath and tetragon fibers have been observed in different sections of the same flagellum in some tubificids and during spermiohistogenesis in a branchiobdellidan (Ferraguti 1984~).
The branchiobdellidan spermatozoa so far studied are typically clitellate in that they possess an acrosome tube with various structures (except in the aberrant B. illurninatus), have mitochondria in a median position, and possess a central sheath in the flagellum.
Hirudinean characters in branchiobdellidan spermatozoa Seven characters distinguished the hirudinean spermatozoon
from the clitellate model as suggested by Wissocq and MalCcha (1975). (1) A helical portion of the acrosome is located anterior to the acrosome tube and vesicle. (2) The acrosome tube has a lateral interruption, where the acrosome vesicle contacts the plasma membrane of the spermatozoon. This is possibly homol- ogous to the apical button of oligochaete spermatozoa
(Garavaglia et al. 1974). (3) The acrosome tube has an external helical flange. (4) The nucleus forms one or more complex helices. (5) A single mitochondrion is present. (6) A conven- tional centriole, with the centriolar area penetrated by an extension of the central apparatus from the flagellum, is absent. (7) A long end-piece is present at the end of the flagellum.
The statement that "branchiobdellid spermatozoa are unmis- takably hirudinean ..." (Sawyer 1986) was based entirely on a literature description of the spermatozoon of B. pentodonta, the only branchiobdellidan to have been studied at the ultrastructural level at that time. When the four species in our study are considered together, it appears that each species possesses a unique assortment of hirudinean character states: C. fallax has an acrosome tube with a helical flange, and a button of the acrosome vesicle bent to one side; C. fallax, B. illurninatus, and X. instabilis have an end-piece on their tails, though it is absent in Branchiobdella spp. The variability in dimensions among the end-pieces in the different species parallels a similar phenom- enon in leeches. It is difficult to assess whether the presence of a non-axonemal end-piece in three of the four branchiobdellidan genera studied, and in the leeches, is of phylogenetic signifi- cance. However, it may be relevant that in addition to these two taxa, some chilopods (Myriapoda) possess a comparable struc- ture (Cotelli et al. 1978; Ferraguti 1984~). Similarly, some homopteran insects (Foliot 1970) have long, complex end-pieces, but these contain a subset of axonemal microtubules, as in other spermatozoan models already known, e.g., the polychaete Tripolydora sp. (Rouse 1988). The possible roles of the end- piece in generating different patterns of spermatozoon movement have been discussed by Omoto and Brokaw (1983). It is interest- ing that a stage of spermiogenesis in B. illurninatus and in homopterans is extremely similar; even the transition between the flagellum proper and the end-piece shows identical features (M. Ferraguti, unpublished data). The axonemal portion of the tail in the 4 branchiobdellidan representatives is extremely similar to that in the 12 hirudinean species studied (Wissocq and MalCcha 1975). The prominent central sheath "embeds" the central tubules so tightly that it is hard to see them, and the radial links and link hiads assume a complex shape not resolvable with conventional electron microscopy. This flagellar pattern is shared with many oligochaetes (Ferraguti 1984~). The centriole remnant in branchiobdellidans, as well as in hirudineans, is connected to the mitochondria by a prolongation of the central apparatus.
Oligochaete characters in branchiobdellidan spermatozoa Oligochaete spermatozoa are distinguished from those of the
other clitellates by three characters. (1) A straight or slightly bent acrosome is followed by a rectilinear flange or variously spiraled nucleus. (2) There is a species-specific number of cyclindrical, sectored mitochondria (2-8) in the mid-piece. (3) The basal cylinder of the flagellar central apparatus penetrates a variable distance into the centriole remnant (Jamieson 1986). The only
FIGS. 3 6 4 5 . Branchiobdella pentodonta (Figs. 36 and 39-45) and B. astaci (Figs. 37 and 38) spermatozoa. Fig. 36. A phase-contrast view showing the long acrosome (a), the short nucleus (n), and the tail (t). X2000. Fig. 37. The marginal fiber helically wound around the tail. X 30 000. Fig. 38. Anterior extremity of the acrosome, showing the widening of the acrosome vesicle (av) at the button and the beginning of the subacrosomal space (ss). x 60 000. Fig. 39. Details of the main portion of the acrosome, showing the continuity between the flattened vesicle under the acrosome tube and the inner tubules. X 60 000. Fig. 40. The base of the acrosome, with the "cork" (= acrosome rod) closing the subacrosomal space. X 60 000. Fig. 41. Longitudinal section of the nucleus (n), mid-piece, and beginning of the flagellum, where the marginal fiber (mf) starts. x 33 000. Fig. 42. Cross section at the "cork" level (compare with Fig. 40). Note the membranes filling the base of the acrosome vesicle. X 85 000. Fig. 43. The seven mitochondria grouped in the mid-piece. X 70 000. Fig. 44. The centriolar remnant in cross section. X 75 000. Fig. 45. Two tails in cross section. X 85 000.
FERRAGUTI AND GELDER 1953

CAN. J . ZOOL. VOL. 69, 1991

FERRAGUTI AND GELDER 1955
known exception to this definition is a loosely spiraled acrosome in one species Phreodrilus observed by Jamieson ( 198 1 ).
Some of these characters are also present in branchiobdel- lidans. In particular, the acrosome of X. instabilis is surprisingly similar to the one in Phreodrilus sp. just mentioned, not only in its helical shape and size but also in the ratios between the different components of the acrosome. Even the number of gyres of the helix seems to be comparable, and it remains to be established if this similarity is due to convergence. Fertilization in Phreodrilus sp. is external, and thus contrary to that in most oligochaetes (Jamieson et al. 1987), therefore a spermatozoon model very different from that found in most oligochaetes is expected, following FranzCn's rule (FranzCn 1956). The shape of the nucleus varies in the different branchiobdellidans as well as in oligochaetes. Branchiobdella spp. have a straight nucleus, as do all "megadriles." Bdellodrilus illurninatus has a nucleus that is anteriorly straight, becoming progressively more helical towards its posterior, as found in the tubificid Kaketio sp. (Ferraguti et al. 1989). Xironogiton instabilis has a tightly coiled helical nucleus, closely resembling that of the lumbriculid Bythonornus sp. (Ferraguti and Jamieson 1987). Cambarincola fallax has a loosely coiled helical nucleus that differs from that of other oligochaetes with the possible exception of the mega- scolecid Pheretirna hawayana, for which we have only optical microscope observations (Alderete de Mayo et al. 1979).
Branchiobdellidan mitochondria are cylindrical and arranged around the central axis of the mid-pike, with a different degree of twist for each species. Four mitochondria have been reported in B. illurninatus, C. fallax, and X instabilis, as in the tubificid Lirnnodriloides sp. (Jamieson and Daddow 1979). However, Branchiobdella spp. contain seven mitochondria and the oligochaete Sparganophilus tharnensis has eight (Richards et al. 1982); these are unusually large numbers for clitellates.
The branchiobdellidan flagellum is similar to that found in the more primitive oligochaetes (Ferraguti 1984a), but differs from that found in the more advanced members. However, during flagellar morphogenesis in branchiobdellidans, a stage occurs during which the two tetragon fibers observed in the spermato- zoa of the advanced oligochaetes are present (Ferraguti 1984b; M. Ferraguti, unpublished observations).
Variability of branchiobdellidan spermatozoa (Table 1 ) The differences in spermatozoon morphology among the four
branchiobdellidan genera examined is striking, particularly in the ultrastructure of the acrosomes and tails. The functional signifi- cance of clitellate spermatozoon characters and dimensions remains unknown. However, Jamieson et al. (1983) found a good correlation between the length of the acrosome and that of the microvilli surrounding the eggs of certain lumbricid species. Our preliminary observations of B. illurninatus eggs, and those of Bondi and Facchini (1972) on B. pentodonta eggs, have revealed that both the thickness of the vitelline layer and the length of microvilli (0.4-0.5 km in both species) are quite similar. How- ever, the differnce in acrosome length between the two species is considerable, that of B. pentodonta being 32 km long, whereas the "acrosome" in B. illurninatus is only 3.6 km long. It does not appear that the correlation proposed by Jamieson et al. (1983) can be extended to these branchiobdellidans.
Can the "acrosome" of B. illurninatus be considered a real acrosome? Preliminary observations on the spermiohistogenesis of the acrosome in B. illurninatus have shown the Golgi appa- ratus during its development (M. Ferraguti, unpublished data). However, the absence of a cell membrane surrounding the electron-dense material in the mature condition clearly separates this structure from other acrosomes.
Ferraguti et al. (1986) compared B. pentodonta and C. fallax spermatozoa and suggested that the extreme reduction of the secondary tube in the acrosome, the concavity on the anterior surface of the nucleus, and the marginal fiber of the flagellum were synapomorphies for branchiobdellidan spermatozoa. The present study and a review of the literature have shown that these synapomorphies are no longer valid; the secondary tube is clearly visible only in X. instabilis, seen with difficulty in C. fallax, B. pentodonta, and B. astaci, and is absent from B. illurninatus; an anterior nuclear concavity is lacking in B. illurninatus; and a marginal fiber helically wound around the flagellum is absent from X. instabilis and B. illurninatus. However, the helical ridge wrapping around the end-piece of B. illurninatus is considered an extreme transformation of the marginal fiber.
At this time we are unable to suggest a set of synapomorphies that would define branchiobdellidan spermatozoa, which is contrary to the situation for the Oligochaeta and Hirudinea. The branchiobdellidan spermatozoon is composed of a variable mixture of characters attributed to the oligochaetes, the leeches, and others that are clearly autapomorphic. The five species examined in this study form a random sample from the three major branchiobdellidan families, but it would be unwise to propose at this time that the. spermatozoan ultrastructure of these species is representative of the respective families. The Branchi- obdellida was demonstrated to be a monophyletic group on the basis of anatomical characters (Gelder and Brinkhurst 1990), and this, together with our observations on spermatozoa ultastructure, further supports the hypothesis of an intermediate but indepen- dent taxon between the oligochaetes and the leeches (Holt 1989).
Ackndwledgements This work has been supported by a grant to M.F. from the
Consiglio Nazional delle Ricerche (Rome) as a Bilateral Project, and Faculty Development Funds to S.R.G. from the University of Maine at Presque Isle. Dr. Gasparo (Trieste) kindly supplied B. astaci.
ALDERETE DE MAYO, A. M., TOMSIC, Z., DULOUT, F., and TESAIRE, E. S. 1979. Espermatogenesis de Pheretima hawayana Rosa (Oligo- chaeta, Megascolecidae). Acta Zool. Lilloana, 35: 243-249.
ANDERSON, W. A., and PERSONNE, P. 1970. The localization of glycogen in the spermatozoa of various invertebrate and vertebrate species. J. Cell Biol. 44: 29-5 1.
AVEL, M. 1959. Classe des Annelides Oligochktes (Oligochaeta Huxley, 1875). In Trait6 de Zoologie. Vol. 5. Edited by P. P. GrassC. Masson et Cie, Editeurs, Paris. pp. 22470.
BACCE'ITI, B. 1970. Comparative spermatology. Academic Press, New York and London.
BONDI, C., and FACCHINI, L. 1972. Observations on the oocyte ultrastructure and vitellogenesis of Branchiobdella pentodonta Whitman. Acta Embryol. Exp. 2: 225-241.
FIGS. 4649. Schematic drawings of the main ultrastructural characters of branchiobdellidan spermatozoa. Note that the scale is different in the four species. Fig. 46. Xironogiton instabilis. Fig. 47. Cambarincola fallax. Fig. 48. Bdellodrilus illuminatus. Fig. 49. Branchiobdella pentodonta. a, acrosome; ac, apical corona; ar, acrosome rod; at, acrosome tube; av, acrosome vesicle; c, "cork"; cr, centriolar remnant; ep, end-piece; g, glycogen granules; m, mitochondria; mf, marginal fiber; n, nucleus; ss, subacrosomal space; st, secondary tube.

1956 CAN. J . ZOOL. VOL. 69, 1991
BONDI, C., and FARNESI, R. M. 1976. Electron microscopic studies of spermatogenesis in Branchiobdellapentodonta Whitman (Annelida, Oligochaeta). J. Morphol. 148: 65-88.
BRINKHURST, R. O., and GELDER, S. R. 1989. Did the lumbriculids provide the ancestors of the branchiobdellidans, acanthobdellidans and leeches? Hydrobiologia, 180: 7-15.
1991. Annelida: Oligochaeta and Branchiobdellida. In Ecology and classification of North American freshwater invertebrates. Edited by J. H. Thorpe and F. Covitch. Academic Press, Inc., San Diego, CA. pp. 401-435.
COTELLI, F., FERRAGUTI, M., and LORA LAMINA DONIN, C. 1978. Morphologie ultrastructurale du spermatozoide de Himantarium gabrielis Linnaeus. Abh. Verh. Naturwiss. Ver. Hamb. (NF), 21/22: 2 19-229.
DADDOW, L. Y. M. 1983. A double lead stain method for enhancing contrast of ultrathin sections in electron microscopy: a modified multiple staining technique. J. Microsc. (Oxf.), 129: 147- 153.
DORNER, H. 1865. Ueber die Gattung Branchiobdella. Z. Wiss. Zool. 15: 464494.
ERNICK, T. H., and EAKIN, R. M. 1976. Fine structure of the cerebral and pygidial ocelli in Chone ecaudata (Polychaeta: Sabellidae). J. Ultrastruct. Res. 54: 243-260.
FERRAGUTI, M. 1983. Annelida Clitellata. In Reproductive biology of invertebrates. Vol. 11. Spermatogenesis and sperm function. Edited by K. G. Adiyodi and R. G. Adiyodi. John Wiley & Sons Ltd., Chichester, U.K. pp. 343-376.
1984a. The comparative ultrastructure of sperm flagella central sheath in Clitellata reveals a new autapomorphy of the group. Zool. Scr. 13: 201-207. %
1984b. Slanted centriole and transient anchoring apparatus during spermiogenesis of an oligochaete (Annelida). Biol. Cell, 52: 175- 1 80.
FERRAGUTI, M., and JAMIESON, B. G. M. 1987. Spermiogenesis in Bythonomus lemani and the phylogenetic position of the Lumbricu- lidae (Oligochaeta, Annelida). Hydrobiologia, 155: 123- 134.
FERRAGUTI, M., and LANZAVECCHIA, G. 1977. Comparative electron microscopic studies of muscle and sperm cells in Branchiobdella pentodonta Whitman and Bythonomus lemani Grube (Annelida, Clitellata). Zoomorphologie, 88: 19-36.
FERRAGUTI, M., GELDER, S. R., and BERMARDINI, G. 1986. On the helices, sperm homologies and phylogeny: the case of branchiobdel- lids. In Biology of reproduction and cell motility in plants and animals. Edited by R. Dallai and M. Cresti. University of Siena, ' Siena. pp. 199-204.
FERRAGUTI, M., GRASSI, G. L., and ERSEUS, C. 1989. Different models of tubificid spermatozueugmata (Annelida, Oligochaeta). Hydrobio- logia, 180: 73-82.
FOLIOT, R. 1970. ~volution des structures caudales terminales des spermatozoides de divers insectes Homoptkres. C. R. Acad. Sci. Paris Ser. D, 271: 508-5 1 1.
FRANZEN, A. 1956. On spermiogenesis, morphology of the spermato- zoa, and biology of fertilization among invertebrates. Zool. Bidr. Uppsala, 31: 355482.
1962. Notes on the morphology and histology of Xironogiton instabilia (Moore 1893) (Fam. Branchiobdellidae) with special reference to the muscle cells. Zool. Bidr. Uppsala, 35: 369-383.
GARAVAGLIA, C., LORA LAMIA DONIN, C., and LANZAVECCHIA, G. 1974. Ultrastructural morphology of spermatozoa of Hirudinea. J. Submicrosc. Cytol. 6: 229-244.
GARDINER, S. L., and JONES, M. L. 1985. Ultrastructure of spermiogenesis in the vestimentiferan tube worm Rijlia pachyptila (Pogonophora: Obturata). Trans. Am. Microsc. Soc. 104: 1944.
GELDER, S. R., and BRINKHURST, R. 0 . 1990. An assessment of the phylogeny of the Branchiobdellida (Annelida: Clitellata), using PAUP. Can. J. Zool. 68: 13 18-1 326.
HOLT, P. C. 1986. Newly established families of the order Branchiob- dellida (Annelida: Clitellata) with a synopsis of the genera. Proc. Biol. Soc. Wash. 99: 676-702.
1989. Comments on the classification of Clitellata. Hydrobiolo- gia, 180: 1-5.
JAMIESON, B. G. M. 1981. Ultrastructure of spermatogenesis in Phreo- drilus (Preodrilidae, Oligoacheta, Annelida). J. Zool. (Lond.), 194: 393408.
1983. Spermatozoa1 ultrastructure: evolution and congruence with a holomorphological phylogeny of the Oligochaeta (Annelida). Zool. Scr. 12: 107-1 14.
1986. Onycophoran-euclitellate relationships: evidence from spermatozoal ultrastructure. Zool. Scr. 15: 141-155.
1987. A biological classification of sperm types, with special reference to annelid adn molluscs, and an example of spermocladis- tics. In New horizons in sperm cell research. Edited by H. Mohri. Japan Scientific Societies Press, Tokyo. pp. 31 1-332.
JAMIESON, B. G. M., and DADDOW, L. 1979. An ultrastructural study of microtubules and the acrosome in spermiogenesis of Tubificidae (Oligochaeta). J. Ultrastruct. Res. 67: 209-224.
JAMIESON, B. G. M., RICHARDS, K. S., FLEMING, T. P., and ERSEUS, C. 1983. Comparative morphometrics of oligochaete spermatozoa and egg-acrosome correlation. Gamete Res. 8: 149- 169.
JAMIESON, B. G. M., ERSEUS, C., and FERRAGUTI, M. 1987. Parsimony analysis of the phylogeny of some Oligochaeta (Annelida) using spermatozoal ultras tructure. Cladistics, 3: 145- 155.
KARNOWSKY, M. J. 1965. A formaldehyde-glutaraldehyde fixative of high osmolality for use in electron microscopy. J. Cell Biol. 27: 137A.
MCDOWELL, E. M. 1978. Fixation. In Diagnostic electron microscopy. Edited by A. Trump and J. Jones. John Wiley & Sons, New York. pp. 1 13-1 39.
MOHRI, H. 1987. New horizons in sperm cell research. Japan Scientific Societies Press, Tokyo. p. 5 16.
OMOTO, C. K., and BROKAW, C. J. 1983. The movement of distal ends of sperm flagella with and without a terminal filament. In The sperm cell. Edited by J. AndrC. Martinus Nijhoff Publishers, The Hague. pp. 380-383.
.- OSTROUMOFF, A. 1883. Ueber die Art der Gattung Branchiobdella Odier auf der Kiemer der Flusskrebs. Zool. Anz. 6: 76-78.
RICHARDS, K. S., FLEMING, T. P., and JAMIESON, B. G. M. 1982. Aberrant spermatozoa and spermatids, and observations on phagocytosis, in Sparganaphilus tamensis (Sparganophilidae, Oligochaeta, Annelida): an ultrastructural study. Int. J. Invertebr. Reprod. 4: 181-191.
ROUSE, G. W. 1988. An ultrastructural study of the spermatozoa from Prionospio cf. queenslandica and Tripolydora sp.: two spionid polychaetes with different reproductive methods. Acta Zool. (Stockh.), 69: 205-2 16.
SAWYER, R. T. 1986. Leech biology and behaviour. Clarendon Press, Oxford.
TIMM, T. 1981. On the origin and evolution of aquatic Oligochaeta. Eesti NSV Tead. Akad. Toim. Biol. 30: 174- 18 1.
WISSOCQ, J. C., and MALECHA, J. 1975. ~ t u d e des spermatozoides d'hirudinks a l'aide de la technique de coloration nCgative. J. Ultrastruct. Res. 52: 340-36 1.