the basal ichneumonidae (insecta, hymenoptera): 28s d2 rdna considerations of the brachycyrtinae,...
TRANSCRIPT

The basal Ichneumonidae (Insecta, Hymenoptera): 28S D2 rDNAconsiderations of the Brachycyrtinae, Labeninae,Paxylommatinae and XoridinaeDONALD L. J. QUICKE, CARLOS LOPEZ-VAAMONDE & ROBERT BELSHAW
Accepted 8 January 1999 Quicke, D. L. J., Lopez-Vaamonde, C. & Belshaw, R. (1999) The basal Ichneumonidae
(Insecta, Hymenoptera): 28S D2 rDNA considerations of the Brachycyrtinae, Labeninae,
Paxylommatinae and Xoridinae. Ð Zoologica Scripta 28, 203±210.
Determining groundplan morphological and biological features can depend on knowing
which groups are derived most basally within that group. Within the Ichneumonidae,
several subfamilies variously have been postulated as occupying a basal phylogenetic
position or possessing largely plesiomorphic morphologies and biologies (e.g. Labeninae),
and therefore potentially exemplifying ancestral life-history stategies. Sequence data from
the D2 variable region of the nuclear 28S rDNA gene have been used to examine basal
relationships in the family. Paxylommatinae and Xoridinae consistently appear as basal
within the family under both maximum parsimony and neighbour-joining analyses.
Labeninae sensu Wahl (1993) often appear near the base of the Ichneumonidae but never as
the sister group of the remainder of the family. The Brachycyrtinae, recently removed
from the Labeninae by Wahl (1993), appear as part of the informal ichneumonoid group
of subfamilies in the most parsimonious trees but this was not statistically significant using
the likelihood-based, Kishino-Hasegawa test compared with a monophyletic Labeninae +
Brachycyrtinae. Biologies of labenines and brachycyrtines are discussed in relation to those
of putative relatives.
D. L. J. Quicke, Unit of Parasitoid Systematics, CABI Bioscience UK Centre (Ascot), Department
of Biology, Imperial College at Silwood Park, Ascot, Berkshire, SL5 7PY UK, and Department of
Entomology, The Natural History Museum, Cromwell Road, London SW7 5BD UK. E-mail:
C. Lopez-Vaamonde and R.Belshaw, Unit of Parasitoid Systematics, CABI Bioscience UK Centre
(Ascot), Department of Biology, Imperial College at Silwood Park, Ascot, Berkshire, SL5 7PY UK.
IntroductionIn phylogenetic studies of relationships between higher
taxa it is frequently necessary to hypothesize groundplan
states, especially for large groups that encompass high
levels of internal homoplasy. Further, even if groundplans
are not the issue, studies are still better based on the
widest possible representation of taxa, and so knowledge of
most basal extant lineages helps ful®l this aim (Mishler
1994). Determination of groundplans should be indepen-
dent of those characters being considered for higher level
relationships so as to avoid potential circularity in the
argumentation. The parasitic wasp family Ichneumonidae
is not only one of the largest families of organisms, it is
also one typi®ed by very high levels of homoplasy (Gauld
& Mound 1982), and this makes interpretation of subfam-
ily level relationships particularly dif®cult. Therefore, we
have attempted to identify the most basal members of the
Ichneumonidae on the basis of gene sequence data and
have included representatives of the putatively least derived
groups together with a small but diverse range of other
ichneumonid and braconid species.
We recently presented analyses of the relationships
suggested by the D2 expansion region of the 28S rDNA
gene in a wide range of ichneumonid and braconid wasps
(Belshaw & Quicke 1997; Gimeno et al. 1997; Belshaw
et al. 1998), and found evidence that the subfamilies Xori-
dinae and Paxylommatinae occupy relatively basal posi-
tions in the Ichneumonidae, and that the Trachypetinae
occupied a relatively basal position in the Braconidae.
Unfortunately, we were not at that time able to obtain
suitable specimens for sequencing of any members of the
subfamily Labeninae. This subfamily has a Gondwanan
Q The Norwegian Academy of Science and Letters . Zoologica Scripta, 28, 1±2, 1999, pp203±210 203

distribution and comprises species that are plesiomorphic
for many characters of both adult and larval morphology
(Gauld 1983; Gauld & Holloway 1986; Wahl 1993), and
which are therefore putatively basal members of the
family. We have now obtained 28S D2 gene fragment
sequences from three representatives of Labeninae sensu
lato, representing three of the tribes recognized by Gauld
(1983), viz. Brachycyrtini, Labenini and Poecilocryptinae,
though we have not managed to obtain suitable material of
the Groteini, nor of the more recently recognized
Xenothyrini (Wahl 1996). Monophyly of the Labeninae
sensu Authors has been debated by Wahl (1993) who raised
the Brachycyrtini to subfamily status and, on the basis of a
putatively derived feature of the larval head capsule,
suggested that this group was more closely related to the
Ichneumoninae + Cryptinae group of subfamilies. Wahl
(1993) did not comment on the relationships of the
remaining Labeninae though he placed them with an unre-
solved, putatively basal grouping including the `Pimpli-
formes' (Wahl 1990) group of subfamilies, together with
the Orthocentrinae, Collyriinae and the Agriotypinae.
Material and methodsWe selected a representative sample of ichneumonid and
braconid taxa for analysis. The total number was limited to
28 species to allow reasonably thorough computerized
alignments to be constructed within a practical time frame.
The ichneumonids represented the informal pimpliformes,
ichneumoniformes, tryphoniformes and ophioniformes
groups (Gauld 1991) as well as the Xoridinae, Paxylomma-
tinae, Brachycyrtinae (Brachycyrtus sp.) and Labeninae
(Poecilocryptus sp. ex galls on Banksia in South Australia;
Labena sp. Costa Rica). The Braconidae was represented
by the Rhyssalus group, other cyclostomes, helconoid and
microgastroid groups of noncyclostomes and the Trachy-
petinae which previous analyses had suggested as being a
basal member of the family (see also Quicke et al., 1999).
The aculeates, Vespa crabro (Vespoidea) and Apis mellifera
(Apoidea) were used as outgroups; their sequences were
retrieved from GENBANK where all our sequences are
also deposited (Y1853-5).
DNA was extracted from single individuals by crush-
ing and incubation at 378C for approximately 18 h in
Proteinase K, followed by sodium acetate/ethanol preci-
pitation and re-suspension in 15 mL Tris/EDTA buffer
(pH 7.5). PCRs were then carried out in 50 mL volumes
containing 0.5 mL DNA extract, 0.5 mL Boehringer Taq
polymerase, 1.25 mL 20 mm primer, 1.25 mL 10 mm
dNTPs and 5 mL buffer (1.5 mM MgCl2). PCR products
were cleaned using QIAquick PCR puri®cation kits
(Qiagen) and then sequenced in both directions using
Thermosequenase (Amersham) at half recommended
volumes on an ABI 373 automated sequencer. The
reverse primer of Campbell et al. (1993) was used with
the forward primer of Belshaw & Quicke (1997). PCR
conditions were 30 cycles of 988C denaturation (15 s),
488C annealing (30 s) and 728C extension (40 s) with an
initial denaturation of 3 min at 938C and a ®nal exten-
sion of 728C for 3 min.
Sequences were aligned using the parsimony-based
program MALIGN (version 2.7 for UNIX) (Wheeler &
Gladstein 1994) with the following parameters: Gap to
substitution ratio 2 : 1, alignaddswap, alignswap, randor-
ders = 10, build, score = 4. Subsequent tree building was
performed using a prerelease PAUP version 4.0d63 (writ-
ten by David L. Swofford) treating gaps as missing data
with 100 random additions followed by TBR. Two of the
loop regions in Megalohelcon are much longer than those of
other braconids or ichneumonids, and excess bases were
trimmed prior to alignment (the full sequence of Megalo-
helcon is presented in Fig. 1 of Belshaw et al. 1998). These
were excluded from the alignment procedure as their
inclusion led to clearly non-sensical output. The aligned
sequences are shown in Appendix I.
A major consideration in interpreting the present
sequence analysis results is the possibility that Xoridinae,
Paxylommatinae, and Trachypetinae form a link between
the other ichneumonids and braconids because each of
these taxa are on long branches and so their placements
may therefore be an effect of long branch attraction (the
Felsenstein zone). This may be further exacerbated by the
considerable base compositional differences between
Ichneumonidae and Braconidae (excluding Trachypetinae).
In addition to using maximum parsimony, we have used
neighbour joining to construct dendrograms from our data
as this method is less susceptible to the effects of long
branch attraction.
The systematic placement of taxa included in the
analyses is shown in Appendix II.
Results and discussionBoth maximum parsimony analysis and neighbour joining
produced similar trees (Fig. 1A,B, respectively), with iden-
tical ichneumonoid backbone stucture, viz. (other Ichneu-
monidae ± Xoridinae - Paxylommatinae - Trachypetinae -
other Braconidae), and this is identical to that found by
Belshaw et al. (1998) which was based on manually aligned
sequences (gaps treated as 5th base or as missing data) and
which did not include any representatives of the Labeni-
nae. However, the rooting of the two trees shown here
differs from that found by Belshaw et al. (1998) in which
the representative aculeates joined between Paxylommati-
nae and Trachypetinae rather than between Xoridinae and
Paxylommatinae. Thus, rooting of trees from this gene
Basal Ichneumonidae ± molecular evidence . Quicke et al.
204 Zoologica Scripta, 28, 1±2, 1999, pp203±210 . Q The Norwegian Academy of Science and Letters

fragment is not stable to variations in taxonomic represen-
tation and alignment and should not therefore be relied
upon in making taxonomic decisions. Thus, although
present analyses showed high bootstrap support for the
Paxylommatinae + Braconidae clade suggesting that the
Paxylommatinae may be better considered as basal braco-
nids, until additional gene regions have been analysed and
sensitivity analyses conducted we do not recommend any
formal changes in classi®cation. Incorporation of addi-
tional species of Xoridinae, Paxylommatinae and Trachy-
petinae are also highly desirable. Further, it should be
noted that some fossils attributed to the Paxylommatinae
possess forewing vein 2 m-cu and so more closely resemble
extant Ichneumonidae sensu stricto. Interestingly, Paxylom-
matinae appeared basal to the Braconidae in some of the
analyses of Quicke et al. (1999) in which they were not
scored for the presence or absence of vein 2 m-cu because
of its presence in some fossil taxa.
Excluding the Paxylommatinae, the next most basal of
the ichneumonids, according to present analyses, is the
Xoridinae, in agreement with recent molecular analyses
that did not include labenines (Belshaw et al. 1998). The
Labeninae sensu stricto (Labena, Poecilocryptus) are placed at
the base of the Ophioniformes clade in the maximum
parsimony tree (Fig. 1A) and at the base of the Pimpli-
formes + Ichneumoniformes clade in the neighbour joining
tree (Fig. 1B) but in neither case was there strong boot-
strap support.
In neither of the analyses shown here, nor in many
others not shown, did Brachycyrtus form a clade with the
two Labeninae sensu stricto, instead in both it appeared as
the sister group of the Eucerotinae and together these
were placed within the group of taxa that we selected to
represent the ichneumoniformes group of subfamilies as
recovered by Belshaw et al. (1998) (most parsimonious
tree; Fig. 1A) or at the base of an ichneumoniformes +
pimpliformes clade (neighbour joining tree; Fig. 1B). The
®rst of these ®ndings is largely in agreement with Wahl
(1993) who considered the derived U-shaped larval salivary
ori®ce to be a character uniting a large group of ichneu-
Fig. 1 Phylogenetic af®nities of the Brachycyrtinae and labenine species (shown boxed) based on 28S D2 rRNA sequences (othersequences are from EMBL/GenBank/DDBJ databases). Ð A. Phylogram is one of four shortest trees length 1215 (CI = 0.498,RC = 0.254), a strict consensus of which merely collapses the nodes marked with an asterisk; Ð B. Neighbour joining tree (based onuncorrected distance matrix). (Bootstrap values above 50% are shown).
Quicke et al. . Basal Ichneumonidae ± molecular evidence
Q The Norwegian Academy of Science and Letters . Zoologica Scripta, 28, 1±2, 1999, pp203±210 205

monid subfamilies (i.e. most ichneumonids except for the
Labeninae sensu stricto, the pimpliformes, Collyriinae,
Orthopelmatinae and Agriotypinae), and thus the presence
of this apomorphy in Brachycyrtus indicates that this genus
should be removed from the Labeninae sensu stricto, and
consequently Wahl (loc. cit.) elevated the Brachycyrtini to
subfamily status. Wahl (1993) further inferred that the
Brachycyrtinae may be the sister group of the Ichneumoni-
nae + Cryptinae (Ichneumoniformes) on the basis of the
elongate 1st metasomal sternite and pentagonal forewing
areolet. Although in our analyses, Brachycyrtinae came out
next to Eucerotinae, these together formed the sister
group of the cryptine, Dichrogaster, within a paraphyletic
Cryptinae. Thus the most parsimonious tree (Fig. 1A)
supports Wahl's (1993) association of Brachycyrtinae with
the Ichneumoniformes. However, constraining the three
species that had in the past been placed in the Labeninae
(Labena, Poecilocryptus and Brachycyrtus) to form a mono-
phyletic group, resulted in two most parsimonious trees
that are only four steps longer than the unconstrained one
(1219 vs. 1215 steps) and this difference was not signi®cant
in the Kishino-Hasegawa test as performed in paup version
4.0d63. Further molecular data are therefore desirable
before reaching ®rm conclusions about the relationships of
the Brachycyrtinae.
Wahl (1993) proposed a number of characters in support
of the monophyly of the Labeninae (excluding the Brachy-
cyrtinae), viz. the form of the notauli and the lower ovipo-
sitor valves which partly enclose the upper valve near the
apex (though a similar condition also occurs in some
Pimplinae, e.g. Delomerista). Present analyses which consis-
tently place Labena and Poecilocryptus as sister taxa therefore
supports Wahl's views at least to some extent.
The ancestral Ichneumonoidea have been postulated as
being most likely to have been idiobiont ectoparasitoids of
wood-boring insect larvae (Gauld 1988). This biology is
exhibited by the Xoridinae and many members of the
pimpliformes group of Ichneumonidae and by the Rhyssalus
group and many cyclostome braconids. The biologies of
Hybrizon (Paxylommatinae) and Megalohelcon (Trachypeti-
nae) are not known but the former are associated with ants
and the short curved ovipositor of the latter show that
neither of these have that biology. Their basal positions in
the results of these analyses and morphological data
presented in a companion paper (Quicke et al. 1999) there-
fore either suggests that generally accepted views about
ichneumonoid ancestral biology are wrong or shows that
these putatively basal lineages have very specialized biolo-
gies and do not represent the ancestral condition at all.
The Labeninae sensu lato display a range of biologies:
Labena species are idiobiont ectoparasitoids of xylophagous
coleopteran and symphytan larvae, Poecilocryptus attacks
gall formers and may be partly phytophagous. Unlike
these, Brachycyrtus attacks loosely concealed pupae of chry-
sopid lacewings and the closely related genus, Adelphion,
attacks spider egg cocoons. The biology of the Brachycyr-
tinae is thus more similar to that of typical cryptines than
to Labeninae sensu stricto, and further supports ®ndings
based on morphology (Wahl 1993) and DNA.
AcknowledgementsWe would like to thank Ian Gauld and Andy Austin for
providing specimens of brachycyrtines and labenines for
DNA extraction, and Mark Dowton for supplying the
sequence data for Toxoneuron. This work was supported by
the NERC (Natural Environment Research Council)
Initiative in Taxonomy, and prompted by the excellent
Workshop on Hymenoptera Phylogenetics organized by
Fredrik Ronquist in Uppsala in March 1998.
ReferencesBelshaw, R., Herniou, E., Gimeno, C., Fitton, M. G. & Quicke, D.
L. J. (1998). A phylogenetic reconstruction of theIchneumonoidea (Hymenoptera) based on the D2 variable region
of 28S ribosomal RNA. Systematic Entomology, 23, 109±123.Belshaw, R. & Quicke, D. L. J. (1997). A molecular phylogeny of
the Aphidiinae (Hymenoptera: Braconidae). MolecularPhylogenetics and Evolution, 7, 281±293.
Campbell, B. C., Steffen-Campbell, J. D. & Werren, J. H. (1993).Phylogeny of the Nasonia species complex (Hymenoptera
Pteromalidae) inferred from an internal transcribed spacer(ITS2) and 28SrDNA sequences. Insect Molecular Biology, 2,225±237.
Gauld, I. D. (1983). The classi®cation, evolution and distribution
of the Labeninae, an ancient southern group of Ichneumonidae(Hymenoptera). Systematic Entomology, 8, 167±178.
Gauld, I. D. (1988). Evolutionary patterns of host utilization byichneumonoid parasitoids (Hymenoptera: Ichneumonidae and
Braconidae). Biological Journal of the Linnean Society, 35, 351±377.
Gauld, I. D. (1991). The Ichneumonidae of Costa Rica. I.Introduction, keys to subfamilies, and keys to the species of the
lower pimpliform subfamilies Rhyssinae, Pimplinae,Poemeniinae, Acaenitinae and Cylloceriinae. Memoirs of theAmerican Entomological Institute, 47, 1±589.
Gauld, I. D. & Holloway, G. A. (1986). Australian ichneumonids
of the tribes Labenini and Poecilocryptini. Bulletin of the BritishMuseum (Natural History), 53, 107±149.
Gauld, I. D. & Mound, L. A. (1982). Homoplasy and thedelineation of holophyletic genera in some insect groups.
Systematic Entomology, 7, 73±86.Gimeno, C., Belshaw, R. & Quicke, D. L. J. (1997). Phylogenetic
relationships of the Alysiinae/Opiinae (Hymenoptera:Braconidae) and the utility of cytochrome b, 16S and 28S D2
rDNA. Insect Molecular Biology, 6, 273±284.
Mishler, B. D. (1994). Cladistic analysis of molecular andmorphological data. American Journal of Physical Anthropology,94, 143±156.
Basal Ichneumonidae ± molecular evidence . Quicke et al.
206 Zoologica Scripta, 28, 1±2, 1999, pp203±210 . Q The Norwegian Academy of Science and Letters

Quicke, D. L. J., Basibuyuk, H. H., Fitton, M. G. & Rasnitsyn, A.P. (1999). Morphological, palaeontological and molecular
aspects of ichneumonoid phylogeny (Hymenoptera, Insecta).Zoologica Scripta, 28, 175±202.
Wahl, D. B. (1990). A review of the mature larvae of Diplazontinae,with notes on larvae of Acaenitinae and Orthocentrinae and
proposal of two new subfamilies (Insecta: Hymenoptera,Ichneumonidae). Journal of Natural History, 24, 27±52.
Wahl, D. B. (1993). Cladistics of the ichneumonid subfamily
Labeninae (Hymenoptera: Ichneumonidae). EntomologiaGeneralis, 18, 91±105.
Wahl, D. B. (1996). Xenothyrini, a new tribe of Labeninae, and akey to the tribes and genera of Labeninae (Hymenoptera:
Ichneumonidae). Proceedings of the Entomological Society ofWashington, 98, 158±161.
Wheeler, W. C. & Gladstein, D. S. (1994). MALIGN: amultiple sequence alignment program. Journal of Heredity, 85,417±418.
Appendix I Malign alignment of D2 region of 28S rDNA of taxa studied. Two long excised loop regions of the Megalohelcon sequence notincluded in the alignment are not shown, their positions are indicated by `[insert 1]' and `[insert 2]' and the full sequence for this taxon isshown in Figure 1 of Belshaw et al. (1998).
Quicke et al. . Basal Ichneumonidae ± molecular evidence
Q The Norwegian Academy of Science and Letters . Zoologica Scripta, 28, 1±2, 1999, pp203±210 207
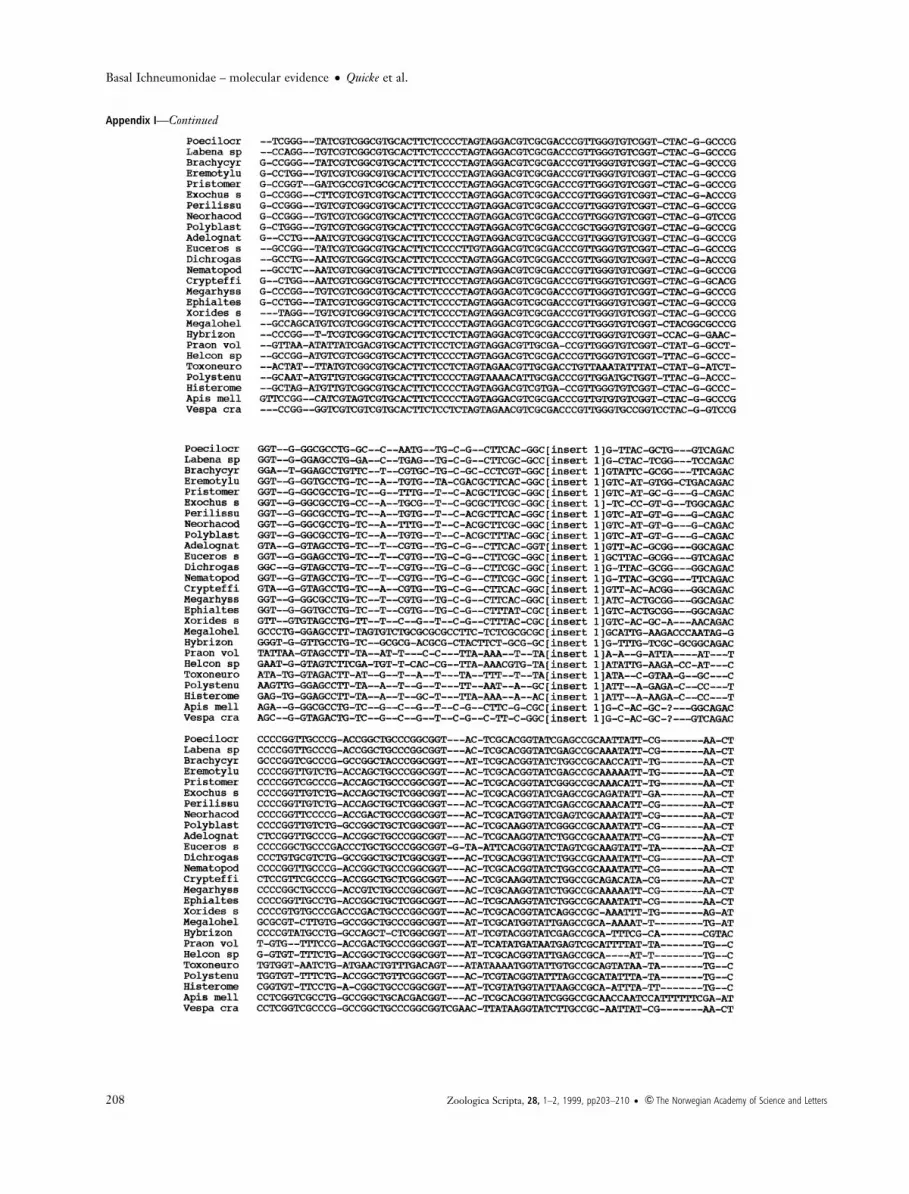
Appendix IÐContinued
Basal Ichneumonidae ± molecular evidence . Quicke et al.
208 Zoologica Scripta, 28, 1±2, 1999, pp203±210 . Q The Norwegian Academy of Science and Letters

Appendix IÐContinued
Quicke et al. . Basal Ichneumonidae ± molecular evidence
Q The Norwegian Academy of Science and Letters . Zoologica Scripta, 28, 1±2, 1999, pp203±210 209

Appendix II Classi®cation of Ichneumonoidea taxa included in study (same order as in Appendix I).
Family Subfamily Genus and species
Ichneumonidae Labeninae Poecilocryptus sp.
Labena sp.
Brachycyrtinae Brachycyrtus sp.
Ophioninae Eremotylus sp.
Cremastinae Pristomerus vulnerator
Metopiinae Exochus sp.
Ctenopelmatinae Perilissus sp.
Neorhacodinae Neorhacodes enslini
Tryphoninae Polyblastus sp.
Adelognathinae Adelognathus sp.
Eucerotinae Euceros sp.
Cryptinae Dichrogaster sp.
Nematopodius debilis
Ichneumoninae Cryptef®gies albilarvatus
Rhyssinae Megarhyssa sp.
Pimplinae Ephialtes manifestator
Xoridinae Xorides praecatorius
Paxylommatinae Hybrizon buccata
Braconidae Trachypetinae Megalohelcon ichneumonoides
Aphidiinae Praon volucre
Helconinae Helcon sp.
Cardiochilinae Toxoneuron nigriceps
Doryctinae Polystenus rugosus
Histeromerinae Histeromerus mystacinus
Basal Ichneumonidae ± molecular evidence . Quicke et al.
210 Zoologica Scripta, 28, 1±2, 1999, pp203±210 . Q The Norwegian Academy of Science and Letters