utility of rdna its sequences in the systematics of teucrium section polium ( lamiaceae )
TRANSCRIPT

Pl. Syst. Evol. 215:49-70 (1999) --Plant
Systematics and
Evolution © Springer-Verlag 1999 Printed in Austria
Utility of rDNA ITS sequences in the systematics of Teucrium section Polium (Lamiaceae)
JALAL EL OUALIDI, OLWIER VERNEAU, SUZETTE PUECH, and JEAN-YVES DUBtnSSON
Received July 7, 1997; in revised version November 2, 1997
Key words: Lamiaceae, Teucrium, sect. Polium, - Internal transcribed spacer region (ITS), nuclear ribosomal DNA, polyploid complex, phylogenetic analysis.
Abstraet: A phylogenetic study based on sequence data from the complete internal transcribed spacer region (ITS) of nuclear ribosomal DNA for sect. Polium of the genus Teucrium shows both intersectional congruence and intrasectional incongruence with traditional morphologieal classifications. We attribute this incongruence !argely to problems related to the polyploid complex studie& Section Polium includes many poorly differentiated taxa of probable recent origin through hybridization followed by polyploidizationl Both on the basis of parsimony and distance (Neighbor-Joining method) analyses, T. dunense and T. thymifolium are the species that diverge most from the sampled taxa. However, unlike T. thymifolium, the morphology of T. dunense is not rauch differentiated in relation to the other taxa. Both species are, nonetheless, the only sampled species to occupy isolated, exclusive environments and which may have undergone rapid evolution by a bottleneck effect. Teucrium dunense is found only on dunes along the Spanish and French coasts and T. thymifolium, a chasmophytic species with limited endemism, is found solely on limestone and dolomite cliffs in the low mountains in south-eastern Spain. A hypothesis is presented to explain the discrepancy between the observed comparatively large amount of ITS sequence divergence and the low morphological differentiation in T. dunense.
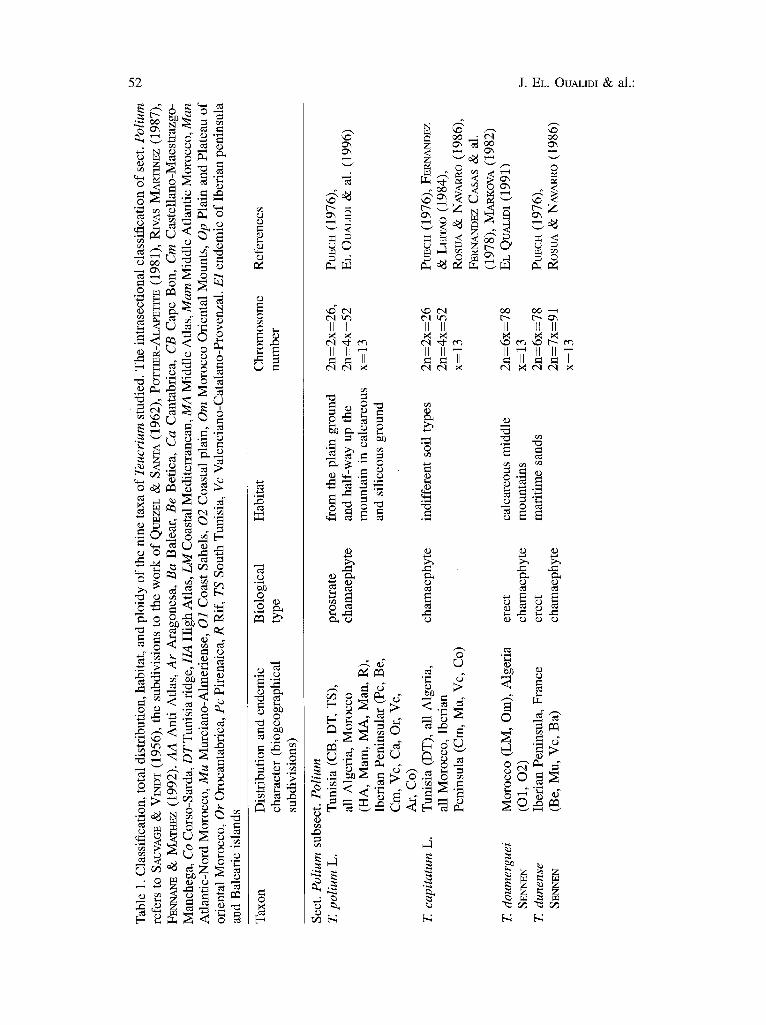
Members of the genus Teucrium L. are dwarf shrubs belonging to the Lamiaceae. They grow in open environments and are represented in the Mediterranean region by 160 taxa belonging to seven sections: Teucrium L., Chamaedrys (MmL.) SCHPa~B., Polium (MILL.) SCHREB., Isotriodon BOlSS., Scorodonia (HmL) SCHmiß., Scordium (MILL.) RCHB., and Spinularia BoIss. The division of Teucrium into sections has been largely based on morphology of inflorescence, calyx and leaves morphology (BENTHAM, 1832--1836: 684--489; WmLKOMM & LANGE 1870: 466-- 481; BOISSmR 1879: 805-822; BRIQUET 1891:141--144; LAZARO IBIZA 1896: 774-- 778; KÄSTNER 1989). Section Polium is characterized by (1) a capitate, condensed raceme inflorescence; (2) helically arranged flowers; (3) subactinomorphic calice; and (4) revolute leaves.

50 J. EL. OUALIDI & al.:
Polium, including more than half of the species of Teucrium, is the largest and most morphologically diverse section of the genus. Most are concentrated in the western Mediterranean region in north-west Africa (Morocco, Algeria and Tunisia) and southern Europe (Iberian Peninsula). In most cases the species display considerable morphological variation within and between populations and many morphological characters are known to be unstable (PuEcI-I 1976, EL OUALmI & PU~CH 1993, NAVARRO 1995). Many taxa form polyploid complexes (PUECH 1984). One documented case is that of the T. lusitanicum SEYYEY complex which comprises eight taxa having high ploidy (2n=6-7x) and which are distributed throughout the eastern Mediterranean. Only a few minor differences in floral and vegetative characters allow the species to be distinguished from each other (EL OUALm~ 1991). The greatest inter- and intraspecific variation among the western Mediterranean taxa is found in the south-eastern part of the Iberian peninsula. In this region 58 of the species are endemic and the remaining are poorly defined, offen forming species complexes. The Iberian Peninsula has been hypothesised to be the centre of evolution and subsequent species radiation for the section (HZDOE & MmLER 1977, PUECn 1984). In contrast, Morocco, with a greater number of ecological niches, has higher endemism (73%) and the wide ranging species are comparatively less variable.
In sect. Polium, the characters used by different authors for infrasectional classification are the same as those used for the intersectional classification but with the additional characters of indument type and floral morphology. In particular, corolla morphology has been given high taxonomic significance. Five different subsectional concepts have been proposed: Polium (MinE.) KÄSTt,~R, RotundifoIia COHEY ex VALDgS BERM. & SANCHEZ CP, ESPO, Simplicipilosa S. PUECH, Intermedia COHEN & Pumilum (LÄZARO IBIZA) RIVAS MARr. (LÄzAao IBIZA 1896, SAUVAGE • VINDT 1955, CoI~y 1956, RIVAS MARTI~Z 1974, P~CH 1976, KÄSZ~R 1989). Subsection. Polium is comparatively more homogeneous and the species display similar flower and inflorescence types. The species are distinguished on the basis of leaf and stem indument. Subsection Polium is to be considered the most evolved, on the basis of presence of a strongly condensed raceme inflorescence, branched hairs, a hood or mucro on calyx teeth, and high ploidy level in some taxa (EL OUALIDI & al. 1996a). The subsection also displays relatively more infraspecific variation; many species are wide-spread and polytypic.
Although several biosystematic studies on sect. Polium involving morpholog- ical and chemical characters have been undertaken, no consensus on subsectional classification nor specific delimitation with regard to wide-spread variable species has emerged (NABLI 1967; P~CH 1984; KÄsrYER 1986; ANDARY & al. 1988; KINo 1988; NAVARRO 1988; GAMMAR & al. 1989; KÄSr~R 1989; EL OUALIDI 1991; Dn~z & al. 1993; ALLAIY & al. 1994; EL OUALIDI & al. 1996a, b). The lack of reliable diagnostic characters has lead to numerous taxonomic disagreements in this group.
A new approach using modern molecular techniques was carried out to examine whether molecular characters might yield insight into phylogeny and evolution within sect. Polium. Several studies have shown that variation in the nuclear ribosomal internal transcribed spacer region (ITS) can be useful for resolving inter- and intrageneric relationships (BALDWIN 1992, SOLTIS & KUZOFF

Utility of ITS sequences in systematics of Teucrium 51
1993, WoJcI~cI-IOWSrd & al. 1993, BALDWIN & al. 1995, YUAN & al. 1996). The reason why ITS is believed to be useful for discriminating relationships among closely related species is because the sequences of spacer regions are non-coding and in general, evolve, more rapidly than coding regions, themselves more useful at the inter- and intrafamilial levels (CI-IASE & al. 1993, CI~EGG 1993, SuI-I & al. 1993, BAUM 1994, GADEK & al. 1996). To the best of our knowledge no nuclear ribosomal sequences of the large and economically important mint family, Lamiaceae, have yet been published, except an abstract on ITS results on Clerodendrum L. s. 1. (Lamiaceae) (STEAN Æ OLMSTEAD 1996).
This study has as its objectives (1) the reevaluation of morphological characters traditionally used in taxonomy of Polium and (2) to test the utility of ITS analysis for resolving relationships in polyploid complexes within sect. Polium.
Materials and methods
Selection of species. The criteria governing the selection of species to be sequenced were as follows. At least several species from each subsecfion (Polium, Rotundifolia and Simplicipilosa) as defined by SAUVAGE and VIyOT (1955) and PwcH (1976) were desired as well as taxa that form known polyploid complexes. Preference was given to common species which are representative of each subsection. The lack of any PCR products was the major difficulty sometimes encountered when attempting to amplify the ITS region so that not all samples could be sequenced. This was the case for the samples of two species in subsect. Simplicipilosa and, for this reason, no data were recorded for this subsection.
Taxa typical of subsect. Polium from different habitats and having different ploidy levels were analysed. We analysed two species (T. rotundifolium and T. thymifolium) that are representative of the two major morphological groups in the subsect. Rotundifolia. Analysis of the two species addresses both the question of phylogenetic utility of ITS sequences in relation to ploidy, since T. rotundifolium is a hexaploid and T. thymifolium diploid, and also provides a measure of the usefulness of traditional morphological characters for diagnosing lineages. Finally, T. capitatum was selected because it grows throughout the range of members of sect. Polium and shows considerable intraspecific variation. We analyzed two populations from Spain and Morocco to see if there exists variation at the level of ITS sequence which corresponds to the geographic disjunction of this wide-spread species.
For outgroup analysis, we chose species from the related sections of Teucrium and Chamaedrys. On the basis of morphology these two sections are incontestably distinct from sect. Polium (CoI~EN 1956). They do, however, share with sect. Polium characters of corolla morphology.
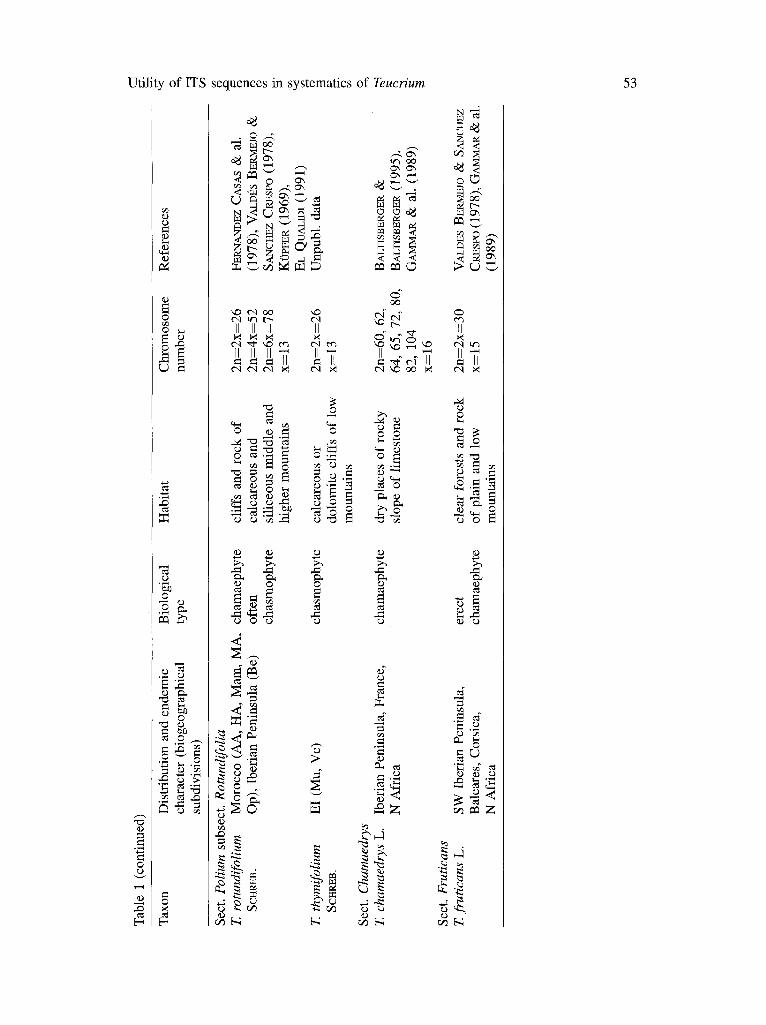
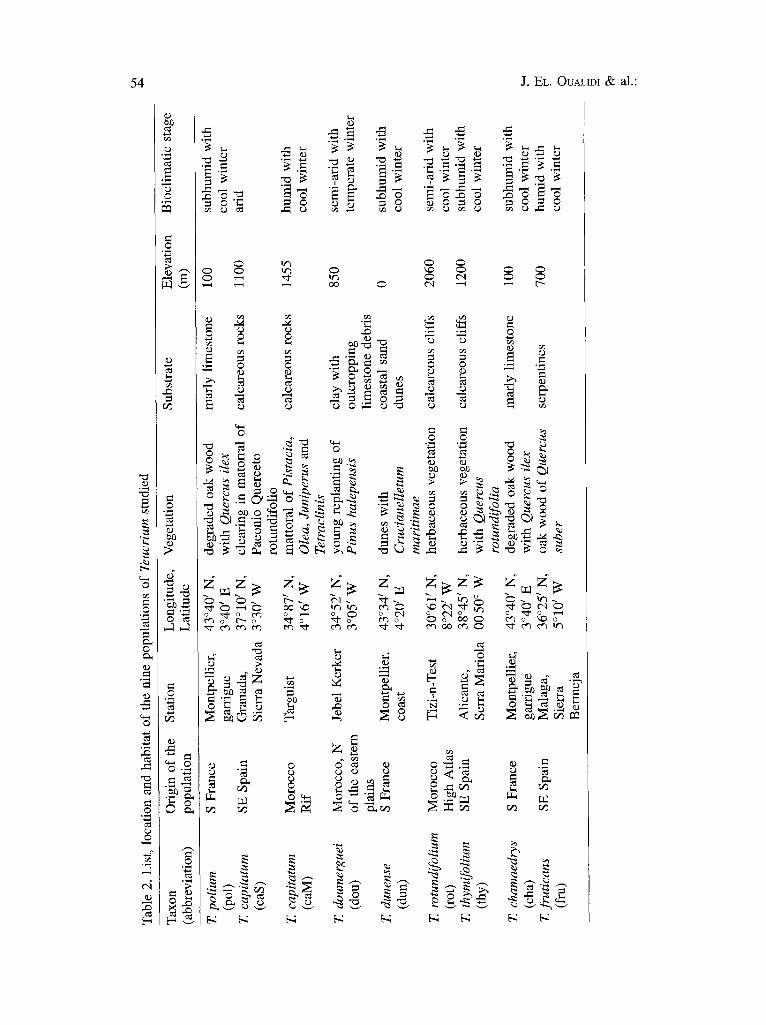
For each taxon, the habitat description, chromosome number, biological type, distribution (according to their biogeographical subdivisions), degree of endemism, and subsection and section to which they belong are shown in Table 1. The populations sampled and their main ecological features are shown in Table 2. Distribution data are limited to the western Mediterranean region and based on the examination of herbarium specimens from the following herbaria: MPU, RAB, BC, MA, R E, K, B and MGC. (see Index Herbariorum, HOLMG~N & al. 1990).
Plant samples. The seedlings were grown from a mixture of approximately 300 seeds collected from wild populations. Young germinated plantlets were transplanted into soil originating from two sites near Montpellier (France). After planting, growth was continued under controlled conditions in a culture chamber with artificial light (1500-2000 lux 12
52 J. EL. OUALIDI ~¢ al.:
• , ' ~ )
= ~ d < ~ . i
©
• ~ ~ : o 2
' U " o ~
~ ~ ~ . ~
= = . ~ d ~<~ ~-=
~~~ ~ü~Z~,
©
!. ©
ù~,«
~2
i
o,, o,,
O
N ,-4
ù,O ~ r , . D ,-, ",-,"
oo
ù--t
I"'... O', ù..«~ v
II II
II Il ¢'q C'-~ ~
C ' q ~
Il II O,I ',~" e~n
II II " = ~ 1 1
OO OO
II Il II
e',l ~ C q C q
ù'a .ü
, ~ , ~ ,-.!
O ù - - t ~ r j
ù_. -
©
< ù..-t
O
8
S
ü_,
d~
ù,.
Utility of ITS sequences in systematics of Teucrium
"m
0
0
©
©
:n
CD
o
4 £
~ cD ~
• ° ~ ~ ~
~~i ~ ~~ ~~ r~ CD
© CD
CD CD
CD e"
= 2 ~ =
b J
~ ~ ~ ~ ~,'~
53
54 J. EL. OUALIDI Æ al.:
~j
o
o ù~ c~
~« o
©
o
ù~ ù~
o
0 o
e-i
o
© »
o
o
0
ù ~ ~
tth
ù~
©
ù ~
ùt:Z +.a ù . ~
~D O O
ù , ~
~=
O © "~
: ~ o
o ~
>.
r.]3 r ~
ù.~ r.F2 r.~

Utility of ITS sequences in systematics of Teucrium 55
hours per day) at a constant temperature of 20 °C. Ten weeks after germination the plantlets were harvested and the DNA extracted. In addition, T. fruticans was collected flora a site near Malaga (Spain) and stored in silica gel for six months. Samples of T. thymifolium were obtained flora an herbarium specimen originally collected from a site near Valencia (Spain) and stored dry and untreated at RAB since 1988.
DNA isolation. DNA from fresh plants was extracted with a standard CTAB extraction protocol modified from DoYIùE and DoYIùE (1990) and DOYLE (1991). 1--2 g of fresh leaves were ground in liquid nitrogen. The resulting mixture was then incubated at 65 °C with 700 gl CTAB extraction buffer (2% CTAB, 1% PVP 40000 Sigma, 100 mM Tris-HC1 pH 8, 10 mM EDTA, 1.4 M NaC1, 1% 2-/3 mercaptoethanol) in a 1.5 ml microtube for 45 rninutes. After two extractions with one volume of chloroform:isoamyl alcohol (24:t), DNA was precipitated with one volume of isopropanol. After centrifugation at 13000 rpm for 3 min, the supernatant was removed and the pellet resuspended in 200 gl of sterilized water. DNA was then precipitated with 1/2 Vol of ammonium acetate 7.5M and 2 vol 95% ethanol. After centrifugation at 13000 rpm for 15 minutes, the supernatant was removed and the pellet containing the DNA was rinsed with 70% ethanol, dried, and then diluted in 50 to 100 gl of sterilized water or Tris-HC1 10 mM (pH 8.5), without Na-EDTA.
PCR amplification. Nuclear genomic DNA of the complete ITS region (including ITS- 1, 5.8S, and ITS-2 rDNA) was directly amplified by PCR using primers TEUC-1 and TEUC-2, defined for Teucrium in the flanking regions of 18S and 26S, respectively: TEUC- l=51-AAGGTTTTCCGTAGGTGAAC-3 / and TEUC-2=5r-TCCTCCGCTTATTGA - TATGC-3/. PCR was performed in a crocodile II Appligene thermal cycler in the following manner: one cycle (denaturation for 5 min at 94 °C, followed by annealing for 1 min at 51 °C, and then elongation for 2 min at 72 °C); then 35 cycles (denaturation for 45 s at 94°C, annealing for 1 min at 51 °C, and then elongation for 1 min 30 s at 72°C); and ending with 1 cycle of elongation for 10 min at 72 °C. PCR products were visualized with ethidium bromide on a 1% agarose gel (in TBE 0.5X) and directly purified with Wizard PCR Preps DNA Purification System (Promega). If more than one arnplification product was observed, the band corresponding to the ITS PCR product was cut out from a 2.5% Low Melting (Tebu Nusieve) agarose gel (in TAE 1X). The DNA was then extracted ffom the gel slice and purified with the above mentioned kit.
Cloning. PCR products were cloned in the pGEM-T vector (Promega) and transfected in Epicurian Coli XL1-Blue MRF' supercompetent cells (Stratagene) for transformation. Presence of the insert was determined by digesting the plasmid DNA with Sac I and Apa I restriction enzymes and running it out on a 1% agarose gel.
Sequeneing. Plasmids were extracted and purified using the Wizard Minipreps DNA Purification System kit (Promega). For each species, the complete ITS region was sequenced using the di-deoxy chain-termination method (SAN6ER & al. 1977), with external universal T7 and SP6 plasmidic primers and the internal primers TEUC-3 and TEUC-4 from the 5.8S region, wherein TEUC-3=51-CGCATTTCGCTACGTTCTTC-3 t and TEUC- 4 =5/-GATGAAGAACGTAGCGAAAT- 3 q
Phylogenetie analyses. Manual älignments and preparation of sequences for anälysis were performed using the MUST package (PI~mIPPE 1993). Gaps were introduced as necessary to maintain alignment while maximizing overall similarity. Phylogenetic reconstruction was conducted on the aligned ITS-region matrix using distance, maximum parsimony (MP), and maximum-likelihood (ML) approaches. Treatment by distance was performed with the Neighbor-Joining method (SAITOU & NEI 1987; program NJ in the MUST package). Robustness of branches was tested by ((bootstrap)) (1000 replicates, program NJBOOT in MUST). For MP the PAUP 3.1.1 package (SwoFFOP, D 1993) was used and the exhaustive method was selected. Nucleotide synapomorphic changes supporting

56 J. EL. OUALD~ & al.: Utility of ITS sequences in systematics of Teucrium
the produced nodes were determined a posteriori by using MacClade ver. 3.04 (MADDXSON & MADDrSON 1992). For MP, any statistical methods for measuring confidence levels of nodes (such as ((bootstrap)) or ((decay index)) analyses) were not used, assuming that a study performed only on synapomorphies is sufficient according to the reduced taxonomic sampling and sequences lengths. A {(Random Trees)) search was also applied in order to calculate the gl value (HILLIS & HUELSEYBECK 1992). This allows detection of any phylogenetic signal in the data matrix. ML was performed using the "quartet puzzling" procedure and the program PUZZLE (STRw~ER & VON HAESELER 1996), which provides a "support" index (expressed in %) for each branch. The distance and ML analyses were performed on PC 486 DX2/66 Mhz and Pentium 166 Mhz, and the MP approach on a Power Macintosh 7200/90. Trees were rooted by outgroups defined previously.
Results
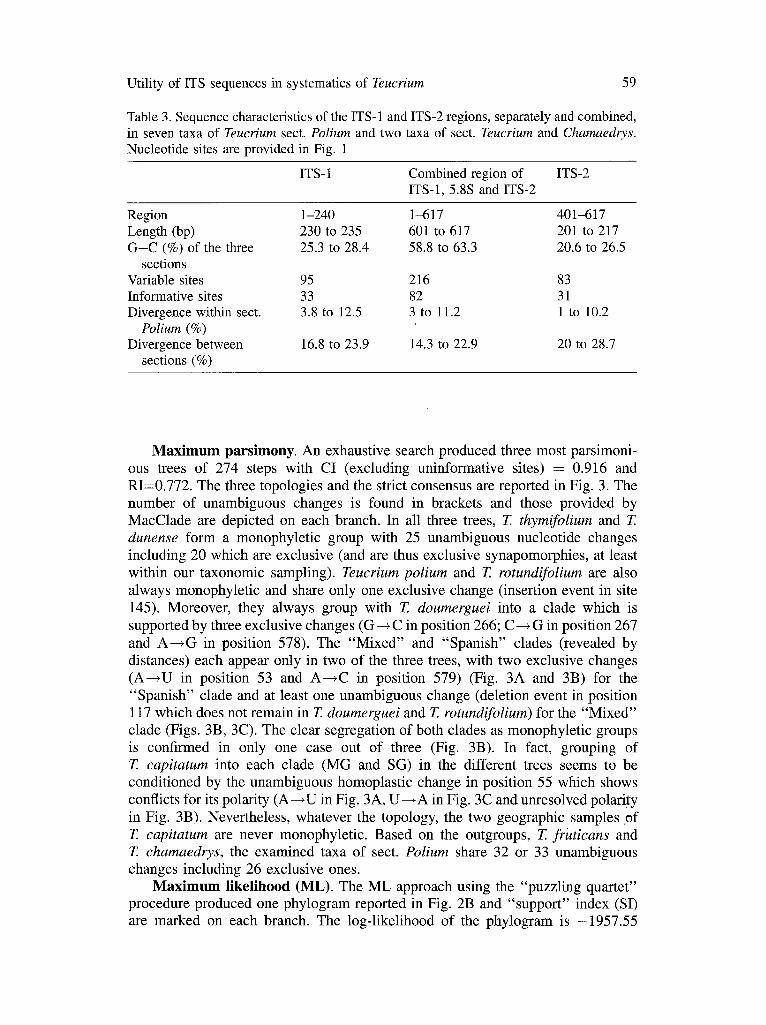
Features of ITS in sect. Polium. Length of the entire ITS-1, ITS-2, and 5.8S regions varies among samples of sect. Polium flora 601 to 617 base pairs. ITS-2 is shorter than ITS-1, ranging from 201 to 217 bp and ITS-1 from 230 to 235 bp. The 5.8S appears to be uniform in size at least among the sampled species (163 bp).
Manual alignments of all ITS region sequence posifions for analyses resulted in a matrix of 617 ¢haracters. Of the remaining, unambiguously aligned positions, 216 are variable. ITS-1 has 95 variable sites and ITS-2 has 83 variable sites. In total, 82 sites are potentially phylogenetically informative.
Percentage of G+C content ranges from 58.8% to 63.3% in the combined region of ITS-1, 5.8S and ITS-2 of sect. Polium and the two outgroups. The combined aligned nucleotide sequences of nrDNA (ITS-1, ITS-2 and 5.8S) from the nine taxa are provided in Fig. 1 and their principal characterisfics are summarized in Table 3.
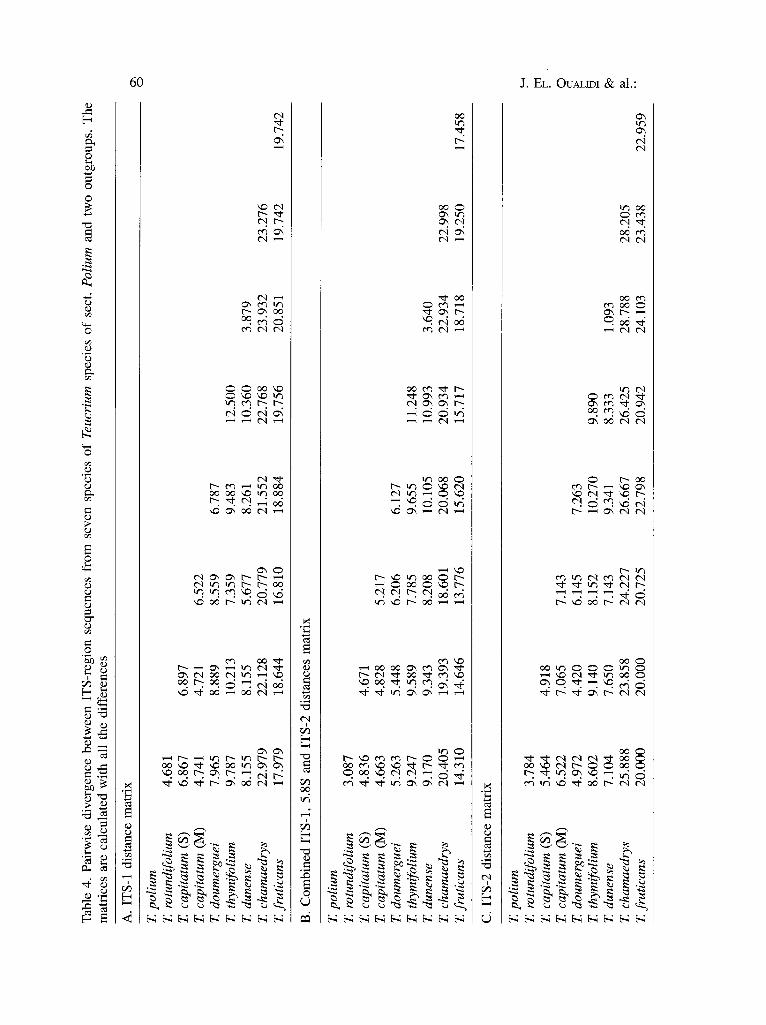
Pairwise nucleotide divergence between ITS-1 sequences of species of sect. Polium ranges from 3.8 to 12.5% (Table 4A); between species of Polium and the outgroup taxa, it ranges from 16.8 to 23.9%. Sequence divergence in the combined region of ITS-1, 5.8S and ITS-2 between pairs of species within Polium, ranges from 3 to 11.2% of nucleotides (Table 4B); between species of Polium and the outgroup taxa it ranges from 14.3 to 22.9% of nucleotides. ITS-2 sequence divergence between pairs of species within Polium ranges from 1 to 10.2% of nucleotides (Table 4C); between species of Polium and the outgroup taxa it ranges from 20 to 28.7% of nucleotides.
Distanee. The distance approach using the Neighbor-Joining method of analysis produced the tree reported in Fig. 2A. Two major groups were resolved: a "mixed group" [T. rotundifolium, T. polium, T. doumerguei and T. capitatum (M)] supported by a strong bootstrap value [94.2% of bootstrap (BP)] and a "Spanish group" related on the basis of geography [T. dunense, T. thymifolium and T. capitatum (S)] with a lower BP value (70.5). Within the "mixed group", T. polium, T. rotundifolium, and T. doumerguei cluster together with weak support (46 BP), and the group of T. rotundifolium and T. polium with a relatively higher bootstrap value (83.4 BP). Within sect. Polium, only the group containing T. dunense and T. thymifolium shows strong support (100 BP) albeit having a long common basal branch.

ITS-1 ~ 0 T. p o l T. r o t T. caS
T. caM
T. dou
T. thy
T. dun
T. cha
T. fru
T. pol
T. rot
T. caS
T. caM
T. dou
T. thy
T. dun
T. cha
T. fru
T. pol
T. rot
T. caS
T. caN
T. dou
T. thy
T. dun
T. cha
T. fru
T pol
T rot
T caS
T caM
T dou
T thy
T dun
T cha
T fru
T. pol
T. rot
T. caS
T. caM
T. dou
T. thy
T. dun
T. cha
T. fru
6O TCGAAACCTG CAAAGCAGAC CGCGAACATG TGCTCAACAAAATCGGGACG GCAGAGAGGG
................... T ....... C ..... T ................. T.T .....
........................... C ..... T .........................
............ G ............ T ....... T ............... T.T.T .....
......................... T ....... T ................. T.T .....
......... T .......... A ...... C...T.T ............ G...T ........
........................... C...T.T ............. T...G.T.T...
120 GGCTCCGTCC TCCTCCCCGC CGCCCCAACC CCGTGCGGGG CGTGCGCTCG CGCCGN-CCG
ù .- ................ A ............ CA .................. T.CT.T.
ù.- ............. GA ............. NC ...................... G...
ù .- .......................... T..CT ..... N ........... -G.T--..
ù .- ............................. -.AK... GN...-N..T ..-...AM..
...- ........... T.T ....... T ...... AC .................... TTG...
ù.- ............. T .............. AC ..................... CG...
ù .-...C.. -.GG..--.. TT...T ....... CA ..... T..A...C.. T .... TG..A
...-GT.C . . . . G...-.. T ............ GA...-A ...... TC ....... TG...
180 TGCGGGTCTA ACAAACTCGG GCGCCGGAAT GCGCCAAGGA AAACTGAAGG GATCGTCCCC
..... C ......................................... A .......... •
..... C ............... N.- ....................... A ...........
..... C ............... N.- ....................... A ...........
R ..... S ................ --....W .N.S .... A...- ..... A ...........
ù.T..C ................. -A ...................... A ...........
...T..C ................ T-A..G ................... A ...........
..... C ....... G ....... A.- ...... T ...... A ...... C..A..,G.A.T.T.
..... A ................. -..C ................. C.TA...G.A.T...
240
58s~O CA-CCCGTCG CCCGTGCGCG GTGCGATCGG CGTGGATCGG ATGTCCGTCG TAATACCATA
. -.T . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C . . . . . . . . . . . . . . . . . .
..- . . . . . . . . . . . . A . . . . . . . . . . . . . . . . . . . . . T.. ,C . . . . . . . . . . . . . . . . . .
. -.. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C.... . . . . . . . . . . . . . .
• -...A .................. G ............... C ..................
..- ............... T ..... A ............... GC ..................
..- ............... T ..... A ................ CK...K .............
.C-G..CAT ...... A.AT ...... C.-.. GTG ........ C.C..A ......... T.A.
..T...CA ....... T ..... A..,C.G.T G ............ C ........ - ..... A.
30O ACGACTCTCG GCAACGGATA TCTCGCGTCT CGCATCGATG AAGAACGTAG CGAAATGCGA
. . . . . . . . . . . . . . . . . . . . . . . ~C . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . GG . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . C . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
........ A ..C ............ GC... T ............... A .............
........ A ............... SC... T .............. TA .... A ........
..A ...... A .............. AGC... NNA..- ............... A ........
..A . . . . . . . . . . . . . . . . . . . . . . GG . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Fig. 1. Aligned nucleotide sequences of nrDNA internal transcribed spacers (ITS- 1, ITS-2, and 5.8S) from seven taxa of the section Polium and two taxa respectively of sections Teucrium and Chamaedrys (genus Teucrium, Lamiaceae; complete taxon names are provided in Table 1 and Table 2). Nucleotide sites are numbered consecutively from 1 to 617 (5' to 3'). The ITS-1 region ranges from position 1 to 240; the 5.8S region from position 241 to 400; and the ITS-2 region from position 401 to 617. N denotes uncertain nucleotide state, hyphens mark gaps required for alignment, asterisks below the alignment ambiguous regions excluded from sequence divergence calculations and the phylogenetic analyses. M = A o r C , K = G o r T , S = C o r G , W = A o r T , Y = C o r T

58
T. pol
T. rot
T. caS
T. caM
T. dou
T. thy
T. dun
T. cha
T. fru
T. pol
T. rot
T. caS
T. caM
T. dou
T. thy
T. dun
T. cha
T. fru
T. pol
T. rot
T, caS
T. caM
T. dou
T. thy
T. dun
T. cha
T. fru
T. pol
T. rot
T. caS
T. caM
T. dou
T. thy
T. dun
T. cha
T. fru
T. pol
T. rot
T. caS
T. caM
T. dou
T. thy
T. dun
T. cha
T. fru
T. pol
T. rot
T. caS
T. caM
T. dou
T. thy
T. dun
T. cha
T. fru
J. EL. OUALIDI & al.:
36O TACTTGGTGT GAATTGCAGA ATCCCGTGAA CCATCGAGTC TTTGAACGCA AGTTGCGCCC
. . . . . . . . . . . . . . . . . . . . . . . . T ° T . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
...................... T ....................................
................................. A .................... T ....
................................. A ........ A ........... T ....
............................. TG .............. T .............
ITS-2 $ ~ 420 GAAGCCATTA GCGCGAGGCG ACGTCTGCCT GGGCGTCACG CATCGCGTCG CCCCC-TCCC
. . . . . . . . . . . . . . . . . . . . . . , . . . . . . . . . . . . . . . . . . . . . . -- . . . . . . . . -- . . . .
................................ A ........... --Y ....... - ....
• . .A ....... GCT .... GC .................. T ................ C ....
ù .A...C.. .GCT..AAGC ........ T ........ TG ........... N .... C ....
....... C ....... A..- .............. A ........ A..A.T ...... - ....
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ° C ° . , T
48O CCATCGGGAG CGGGGGCGG .... AGAATGG CCTCCCGT-G CGACACCGCA CGGCTGGCCC
. . . . . . . . . . . . . . . . . . -- ------, . . . . . . . . . . . . . . -- . . . . . . . . . . ~ . . . . C . . . . .
. . . . . . . . . • °, , o, • o,, . . . . • . . . . . . . . . . . . . . -- . . . . . . . . . . G . . . . C , , • ,,
.............. -,..- ---. ...... ........ - .......... N . . C . - .....
............... N.N .... . .............. - .......... G .... C .....
..... A ........... A .... . .............. - .......... G T ..... A...
..... A ........... A .... . .............. - .......... G ...... A...
.-G..AA.G .......... - CGG ......... C ..... - ..... C.T.TG .... C .....
• C ....... T.T...G..G CGG ......... C ..... C. .TC.C .... G .... C .....
54O AAATGCTCTC CCCC-GGCGA ACGCACGTCG CGGCAAGTGG TGGTTGAACT CTCAACTCGC
............. - ................... T .........................
............. - .................. N ..........................
T ......... - .............................................
. . . . . . . . . . . . . --, ° T ° , - T . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
........... G.-..- ............... T .......... G ...............
........... G.- ..... - .... M ....... S .......... G ...............
.... TG ....... CA.T.. - ....... T ..... C .............. C ..........
..... G ....... - ............... T.A.C ............... T .........
6OO GTG-CTGCTG CGATCGAGGC -GTCGT-CCG TCGGGAA-AC AGAACTAGAC CCAGACGACC
• ° .-- . . . . C°
• TN .... C.
°.°-- . . . . C.
.° .-- . . . . C,
• . . - .... CA
• . . - .... CA
• °°-- . . . . C.
• . °--, æ °T° °
617
GC ..... GCC TACGACT
° -- . . . . . . . . . . . . . C
° -- . . . . ° . . . . . . . . C
• -- . . . . ° . . . . . . . . C
ùN . . . . . • . . . . . . . . C
, - . . . . . . . . . . . . . C
ùN ..... . ........ C
A. AACGT ......... C
• . AAT GC ......... C
. . . . . . . . . -- . . . . . - T . . . . . . A ° .- . . . . . . . . . . . . . . . . . . . . . .
........ N -. .... -.-T ....... -.. .T ........... AC .....
A ......... - ..... - .......... - ............... ATG... -
° A . . . . . . . . - . ° T . ,- . . . . . . . . . . - ° A . . . . . . . . . . . . . . . . . . . .
......... -...A.- . . . . . . . . . . - ............... AC .... G
. . . . . . . . . --°° .~.-- . . . . . . . . . . -- . . . . . . . . . . . . . . . A G . . . . .
• .C...ACA T ..... T.-- A.C.A..C.. GA...AG ...... -.T.G.-
ù-G.C..AG-..-..-... A ...... C.. .A...A ....... -..NN.-
Utility of ITS sequences in systematics of Teucrium 59
Table 3. Sequence characteristics of the ITS-1 and ITS-2 regions, separately and combined, in seven taxa of Teucrium sect. Polium and two taxa of sect. Teucrium and Chamaedrys. Nucleotide sites are provided in Fig. 1
ITS-1 Combined region of ITS-2 ITS-1, 5.8S and ITS-2
Region 1-240 1-617 401-617 Length (bp) 230 to 235 601 to 617 201 to 217 G+C (%) of the three 25.3 to 28.4 58.8 to 63.3 20.6 to 26.5
sections Variable sites 95 216 83 Informative sites 33 82 31 Divergence within sect. 3.8 to 12.5 3 to 11.2 1 to 10.2
Polium (%) Divergence between 16.8 to 23.9 14.3 to 22.9 20 to 28.7
sections (%)
M a x i m u m parsimony. An exhaustive search produced three most parsimoni- ous trees of 274 steps with CI (excluding uninformative sites) = 0.916 and RI=0.772. The three topologies and the strict consensus are reported in Fig. 3. The number of unambiguous changes is found in brackets and those provided by MacClade are depicted on each branch. In all three trees, T. thymifolium and T. dunense form a monophyletic group with 25 unambiguous nucleotide changes including 20 which are exclusive (and are thus exclusive synapomorphies, at least within out taxonomic sampling). Teucrium polium and T. rotundifolium are also always monophyletic and share only one exclusive change (insertion event in site 145). Moreover, they always group with T. doumerguei into a clade which is supported by three exclusive changes (G--~ C in position 266; C-~ G in position 267 and A - + G in position 578). The "Mixed" and "Spanish" clades (revealed by distances) each appear only in two of the three trees, with two exclusive changes (A-~U in position 53 and A - ~ C in position 579) (Fig. 3A and 3B) for the "Spanish" clade and at least one unambiguous change (deletion event in position 117 which does not remain in T. doumerguei and T. rotundifolium) for the "Mixed" clade (Figs. 3B, 3C). The clear segregation of both clades as monophyletic groups is confirmed in only one case out of three (Fig. 3B). In fact, grouping of T. capitatum into each clade (MG and SG) in the different trees seems to be conditioned by the unambiguous homoplastic change in position 55 wfiich shows conflicts for its polarity (A-~U in Fig. 3A, U--~A in Fig. 3C and unresolved polarity in Fig. 3B). Nevertheless, whatever the topology, the two geographic samples o f T. capitatum are never monophyletic. Based on the outgroups, T. fruticans and T. chamaedrys, the exarnined taxa of sect. Polium share 32 or 33 unambiguous changes including 26 exclusive ones.
M a x i m u m likelihood (ML). The ML approach using the "puzzling quartet" procedure produced one phylogram reported in Fig. 2B and "support" index (SI) are marked on each branch. The log-likelihood of the phylogram is -1957.55
¢1 ,x=
cd
O
O
e, ù.J
O
°,«
"p, ¢o
#, O
O
O
¢D
60
e-I
p~
t - - ' @ eq P-- ed~q eq
CNI
~ e d ¢ 5 e~ eq eq
~ - ~ - ~ ~ ~ ~ - ~
M M M M M M M M M
tt3
eä¢a eq ,--*
~ t"r3 ,-~
ee~ t'xl ,--~
¢~.~
M ~ ~ M M M M M M
J. EL. OUALIDI Æ al.:
t ~
eq eq
eq
~. ¢4 ,4 ,--~ eq e-I

Utility of ITS sequences in systematics of Teucrium 61
(+/-63.54) and any confidence intervals defined for each branch length does not contain a zero value. The topology arrived at by the ML approach does not contradict the NJ tree (Fig. 2A). Within the ingroup, only the group comprising T. thymifolium and T. dunense is supported above 90% (SI=100%). The monophyly of the "Spanish" group is confirmed with strong support (SI=87%). Conversely, the "Mixed" group and the unresolved regrouping of T. polium, T. rotundifolium and T. doumerguei also appear, with some support (SI=71% and 68%, respectively). In each case, a probable but unconfirmed phylogenetic signal is indicated.
Discussion
Variability of sect. Polium ITS sequences. The sizes of ITS-1 and ITS-2 in Teucrium are similar to those reported from other angiosperms (ITS-1, 194-268; ITS-2, 188-237), as is the size of the 5.8S coding region (VZNKAT~SWARLU & NnznR 1991, BALDWIN 1992, reviewed in BALDWIN & al. 1995). Furthermore, ITS-2 of sect. Polium is shorter than ITS-1, a condition also often reported (BnLDw~ 1992, SUH & al. 1993, BALDW~J & al. 1995, SCHmHNC & PA~RO 1996). In sect. Polium, nucleotide divergence between pairs of species ranges from 3.8% to 12.5% for ITS-1 and from 1% to 10.2% for ITS-2. This variation is comparable with that found in other families such as Asteraceae (BnLDW~ 1992), Apiaceae (DowN~ & KATZ-DOWME 1996) and, Fabaceae (WoJcmcIJowsr~ & al. 1993).
Within sect. Polium, sequence divergence in the combined region of ITS-1, 5.8S and ITS-2 between pairs of species ranged from 3 to 11.2% of nucleotides, is less than the one found between species of Polium and the outgroup taxa, which ranged from 14.3 to 22.9% of nucleotides. These values demonstrate (1) that the ITS region is quite variable within sect. Polium and (2) that there exists more intersectional than intrasectional variation. The obtained values are consistent with the expectation that the species within sect. Polium are more closely related to each other than to species in neighbouring sections.
Phylogenetic relationships within sect. Polium. The phylogenetic analysis based on ITS sequences revealed:
(1) Twenty-six unambiguous changes shared by the selected representative species of sect. Polium that support the naturalness of the group as previously suggested on the basis of morphological characters. However, analysis of additional taxa from other sections, as well as from other closely related genera, are needed to confirm the monophyly of this section.
(2) Our ITS analyses of sect. Polium closely associated taxa that are remote on the basis of existing classification. This is in contrast to similar analyses conducted on other taxa, in which the results agreed with traditional morphology-based classifications (Stm & al. 1993, DOWNIE & KaTz-DOWNm 1996, WoJ«ZCHOWSra & al. 1993). Thus, Teucrium polium and Z rotundifolium appear closely related on the basis of ITS sequence comparison, but belong to different subsections (SAUVAGE & VrNDT 1955, COHEN 1956). Characters separating them are both vegetative (leaves and indument) and reproductive (calyx and corolla). Moreover, T. dunense and T. thymifolium, always unite together into a monophyletic group having 25 unambiguous nucleotide changes including 20 which are exclusive. Teucrium

62 J. EL. OUALIDI Æ al.:
0,9240
I 7-. fruticons "1". chamaedrys
I T. capitatum (M) ~-~ I T. doumerguei I a 6 ~ T. rotundifolium
4001 ~ « T. polium I , «oo I T. dunense
z~~ f T. thymifolium v.~, T. capitatum ( S )
0,t
"11 fruticans
8
t 0 0
T. chamaedrys
T. capitatum ( S ) 4oo[~ T. thymifolium
T. dunense
B ~ T. cüpitotum(M)
T. polium oJ T. rotundifolium
T. doumerguei
Fig. 2. A. Tree obtained from the Neighbor-Joining method of SAITOU 8~ NEI (1987) for combined ITS-1, 5.8S, and ITS-2 sequences of Teucrium sect. Polium taxa. Numbers below nodes indicate bootstrap estimates for 1000 replicates. Lengths of branches are proportional to distances. (E) is a Spanish population of T. capitatum; (M) is a Morrocan population of T. capitatum. B. Cladogram obtained from the Maximum Likelihood analysis of ITS-region nucleotide sites among species of Teucrium sect. Polium and outgroups. Numbers at branch points are support indices (SI) for the clade. (S) is a Spanish population of Z capitatum; (M) is a Morrocan population of T. capitatum
dunense and T. thymifolium, have been traditionally considered distantly related on the basis of both vegetative and reproductive morphology, and have been placed in separate subsections: subsect. Polium and subsect. Rotundifolia, respectively (VALDES BEaMEJO & SAYCI-mZ Cm~spo 1978). In fact, T. polium and T. dunense share many morphological characters in c o m m o n and T. dunense has long been considered a sand-dune variety of T. polium (EL OUALIDI 1987). Teucrium polium is a tetraploid always found just inland from the dunes where T. dunense, a heptaploid, grows. This association strongly suggests that T. dunense is the product of allopolyploidization followed by fixation on the exclusive dune environment. Moreover, T. dunense strongly resembles T. polium, but is larger. Finally, the two species are parasitized by the same insect, Copium clavicorne Fotn~~. (Hemiptera, Heteroptera, HouArm 1906).
(3) In the strict consensus of the maximum-pars imony topology, the Moroccan and Spanish populations of T. capitatum are not monophylet ic and the clade

Utility of ITS sequences in systematics of Teucrium 63
32(26)
3(3)
4(0) " "
3(2] 33(26) zsczo I
Teucrium polium
Teucrium rotundifolium
Teucrium doumerguei
Teucrium capitatum ( S )
Teucrium thymifolium
Teucrium dunense
Teucrium capitatum ( M )
Teucrium chamaedrys
Teucrium fruticans
33(26~
4(0) ~ L - -
2(2)
~5(20)
Teucrium pofium ]
Teucrium rotundifofium
Teucrium doumerguei MG
Teucrium capitatum (M)
Teucrium capitatum ( S ) 7 I
Teucrium thymifolium ] SG I
Teucrium dunense _J
Teucrium chamaedrys
Teucrium fruticans
(0)
« ic • - - Teucrium pofium
3 ¢3) Teucrium rotundifolium
2 (o ~ Teucrium doumerguei
Teucrium capitatum (M)
2s (2t~ Teucrium thymifolium
Teucrium dunense
Teucrium capitatum ( S )
Teucrium chamaedrys
Teucrium fruticans
C Strict consensus
Teucrium polium
Teucrium rotundifofium
Teucrium doumerguei
Teucrium capitatum ( S )
Teucrium capitatum ( M )
Teucfium thymifolium
Teucrium dunense
Teucrium chamaedrys
Teucrium fruticans
Fig. 3. The three most parsimonious cladograms (A-C) and their strict consensus tree based on sequence data from the nuclear ribosomal internal transcribed spacer (ITS) region for Teucrium sect. Polium and outgroups. On each branch the number of unambiguous changes is depicted, with exclusive changes in parentheses. (S) is a Spanish population of T. capitatum; (M) is a Morrocan population of T. capitatum
containing them unresolved. Teucrium capitatum is sympatric with all other species of sect. Polium, and shows good reproductive isolation. For this and other reasons, it is considered by taxonomists to represent a "good" albeit polytypic species and should, therefore, appear monophyletic.
Our phylogenetic analysis based on results from ITS sequence comparison orten contradicts traditional schemes of morphological classification in sect. Polium. There are three possible explanations for this: (1) ITS sequence comparison is not always a good measure of species relatedness; (2) there are genomic particularities of the examined species; or (3) a combination of 1 and 2. We favour the third view but stress that because of the prevalence of different ploidy levels in the sampled species, unusual patterns of ITS sequences may have emerged.
The problem of polyploidy and ITS sequenee phylogeny. Except for Teucrium thymifolium, a species endemic to south-east Spain, all taxa examined display a very broad distribution, and concomitant geographical and ploidy variations. Teucrium capitatum is the most variable species. In Morocco and Spain, this species is found in all open environments and includes diploid and tetraploid

64 J. EL. OUALIDI & al.:
populations, and in the Rif mountains of Morocco a triploid population is known. Teucrium rotundifolium, an extremely variable hexaploid species (SAUVAGE ~~L VINDT 1965), is found at middle and higher elevafions of mountains and has diploid cytotypes in Spain. Teucrium polium is diploid in Morocco, and tetraploid in Spain and France. Teucrium doumerguei is a hexaploid species that is widespread in north-eastern Morocco, where it forms a complex of three morphologically similar hexaploid taxa that are biogeographically contiguous. Finally, T. dunense, a species which grows along the eastern coast of Spain and in France, has the highest ploidy level of the section (2n=7x) .
As with any other type of character used in systematics, molecular characters have their peculiarities and relative value. With regard to ITS, it has been documented in Gossypium that bidirecfional concerted evolution can occur after hybfidization and subsequent allopolyploidization (WENDEL & al. 1995), two processes that are the foundation for reticulate evolution (SCHmL~G & PANERO 1996, SOSEF 1997). Plants resulfing from hybridization or an allopolyploidization event have been shown in some instances not to form a monophyletic group with the non-hybrid patent lineage in analyses based on molecular or morphological data (SOLTIS & al. 1992, BALDWlN & al. 1995, MCDADE 1995). Thus, there may be ambiguities and lack of resolution in phylogenefic analyses in cases in which the origin of probable recent hybrids or allopolyploids is unknown, as is offen the case with members of sect. Polium.
Furthermore, DNA extracts ffom fresh material of four narrowly endemic diploid species (T. grosii PAU, T. eriocephalum WmLK., T. charidemi SANDWITH, T. turredanum LOSA & RIVAS GODAY), did not amplify although the same primers were used that worked for the other species. This result may reflect genomic divergence of these diploid species at the primer sites. The narrow endemics in sect. Polium are all diploid species and probably represent the ancestral condition of ITS sequences. In contrast to the allopolyploids, which have many potential matches to the primers, diploid species have fewer copies of ribosomal genes and, thus, offer fewer possibilities for the primers to anneal to. It may be that PCR amplification in diploid species requires different conditions of stringency.
The problem of morphological incongruence in the T. t h y m i f o l i u m - T.
d u n e n s e clade. Both in parsimony and distance (Neighbor-Joining method analyses), T. dunense and T. thymifolium emerge as the species that have diverged most from the other species. Nevertheless, unlike T. thymifolium, the morphology of T. dunense is not particularly distinctive in relation to the other taxa. This suggests that what appears as considerable molecular divergence does not necessarily correspond to a commensurate amount of morphological change (DONOGHUE & SANDERSON 1992). Changes in spacer regions, which are non-coding and presumably usually evolving by neutral mutations (HARTL & CLARK 1989), probably are not tighfly linked to morphological change.
Teucrium dunense and T. thymifolium are the only species which occupy isolated, exclusive environments, and possibly represent products of rapid evolution by bottleneck effects (SLATKIN 1987). Teucrium dunense is found only in dunes on the Spanish and French coasts and its high ploidy level (7x) is doubtless related to its substantial intra-population variability and phenotypic amplitude, a phenomenon well recognized in higher plants (HARLAN & DE WET

Utility of ITS sequences in s ystematics of Teucrium 65
1975, LEVIN 1975, Gt:z'r & al. 1989). This variability is all the more marked in nutritionally poor environments such as dunes (BRADSHAW 1965, S~ARNS 1989). Study of the growth of the polyploid complex T. lusitanicum, based on "Mahalanobis" distances, has shown that T. dunense is strongly differentiated from the other taxa by a different growth mode and, in particular, a lower branching rate, the probable result of specialization in dune environments (EL OtJALrDI & al. 1998). Indeed, late branching may have selective value in T. dunense, as growth of the primary axis is precocious and this, in turn, allows the young plant to avoid being covered by sand. To the extent that the pattem of development of this species is distinct from other members of sect. Polium, there exists congruence with the high ITS-sequence divergence obtained from the molecular analysis. However, transplant experiments demonstrate that when grown on regular soils T. dunense assumes a more typical growth pattern and closely resembles one of its hypothetical patents, T. polium. The high allopolyploidy of T. dunense might be the reason that this species is closely associated with T. thymifolium based on ITS sequence comparison, otherwise T. thymifolium or a member of its lineage was involved in the allopolyploidy event that gave rise to T. dunense, but relative to the other taxa, this still cannot explain why in T. dunênse there is an apparent lack of congruence between what is considerable ITS sequence divergence and very little morphological change.
Teucrium thymifolium, a chasrnophytic species which grows solely on lime- stone and dolomite cliffs in the low mountains in south-eastem Spain is distinctive. Among the examined species, T. thymifoIium has a unique leaf morphology and indument type. In contrast to T. dunense, the relatively large ITS sequence diver- gence found in T. thymifolium correlates well with the morphological distinctive- ness of this species.
Conclusion and future perspectives. In Teucrium, sequence divergence in the combined regions of ITS-1, 5.8S and ITS-2, appears to discriminate well between the examined sections. In contrast, within sect. Polium, although the sequence divergence between pairs of species is significant and in the fange expected for a relatively evolved family such as the Lamiaceae, the resultant phylogeny does not provide sufficient resolution nor show congruence with traditional morphology based classifications. Much of sect. Polium is composed of taxa of probable recent hybrid origin followed by polyploidization. ITS sequence evolution may be biased by concerted ribosomal gene evolution, a phenomenon known to occur in allopoly- ploids. Notwithstanding, the latter cannot adequately explain the observation that ITS sequence divergence in T. dunense is considerable in a species that is only subtly different in morphology from one of its presumed parents, T. polium, as weil as from a number of other species. This result underlines the notion that ITS evolution is influenced by factors different from those affecting morphological change. For this reason, great care must be taken when interpreting results of ITS sequence analyses. Clearly, a good knowledge of the study plants is indispensable for a critical interpretation of phylogenetic analyses based on ITS sequence, as it is for analyses using, any other character of unknown evolutionary value.
Given that we have devoted numerous years to the study of the genus Teucrium and, in particular, sect. Polium, and thus have an intimate knowledge of the group, we look forward to sequencing the ITS region of the entire section,

66 J. EL. OUALIDI Æ al.:
and many representatives of the remaining seven sections in the hopes of gaining further insight into the evolution of this enigmatic group. Concomitant with the latter study, we propose a study specifically focused on understanding the dynamics of ITS evolution in a distinct species known to be polyploid, such as T. a u r e u m SCHREB. or T. ro tundi fo l ium, in which there exist 2x, 4x and 6x cytotypes. Such a study should be of the greatest utility understanding the value of sequence divergence in ITS regions in Teucr ium. Furthermore, it might also be worthwhile to do sequence comparison of the ITS in artificial hybrids and their known parents.
We would like to thank FRANfO~S CATZEFLIS and all the persons from "Service Commun de Biosystématique" at Montpellier for their technical assistance, and precious remarks. We are also obliged to DAWN FRAME for critical review of the manuscript. The research benefited flora support by the "Service Commun de Biosystématique" at Montpellier during a post-doctoral stay by Dr JALAL EL OUALIDI at the Institut des Sciences de l'Evolution (UMR 5554 CNRS). This work is contribution No. 96-041 of the "Institut des Sciences de l'Evolution de Montpellier" (UMR 5554 CNRS).
References
ALLAIN, P., EL OUALIDI, J., PUECH, S., PELLECUER, J. P., 1994: La variation de l'huile essentielle de Teucrium puechiae GREtrrER and BURDET. -- Biochem. Syst. Ecol. 22: 239- 248.
ANDARY, C., RASCOL, J. P., PUECH, S., ROUSSEL, J. L., PRIVAT, G., 1988: Les esters de l'acide caféique dans la chimiotaxinomie des Teucrium de la secfion Polium (Lamiaceae). - Canad. J. Bot. 66: 1007-1012.
BALDWIN, B. G., 1992: Phylogenetic utility of the Internal Transcribed Spacers of nuclear fibosomal DNA in plants: an example from the Compositae. - Molec. Phylogenet. Evol. 1: 3-16.
SANSERSON, M. J., PORTER, J. M., WOJCIECHOWSKI, M, F., CAMPBELL, C. S., DONOGHUE, M. J., 1995: The ITS region of nuclear ribosomal DNA: a valuable source of evidence on angiosperm phylogeny. - Ann. Missouri Bot. Gard. 82: 247-277.
BALT~SBZRGER, M., BALTISBZR~ZR, E., 1995: Cytological data of Albanian plants. - Candollea 50: 457-493.
BAUM, D., 1994: rbcL and seed-plants phylogeny. - Trends Ecol. Evol. 9: 39-41. BENTHAM, G., 1832-1836: Labiatarum genera et species. - London: Ridgway. BOISSlER, E E., 1879: Flora Orientalis. - Basel, Genève, Lyon: Georg. BRADSHAW, A. D., 1965: Evolutionary significance of phenotypic plasticity in plants. -
Advances Geriet. 13:115-155. BRIQUET, J., 1891: Les labiées des Alpes maritimes, 1. -Basel, Genève: Georg. CHASE, M. W., SOLTIS, D. W., OLMSTEAD, R. G., MORGAN, D., LES, D. H., MISHLER, B. D.,
DUVALL, M. R., PRICE, R. A., HILLS, H. G., QIu, Y. L., KREN, K. A., RErnNO, J. H., CONTI, E., PALMER, J. D., MANHARD, J. R., SYSTMA, K. J., MICHAELS, H. J., KRESS, W. J., KAROL, K. G., CLARK, W. D., HEDREN, M., GAUT, B. S., JANSEN, R. K., KIM, K. J., WIMPEE, C. F., SMITH, J. F., FURNIER, G. R., STRAUSS, S. H., XL~Na, Q. Y., PLUNKETT, G. M., SOLTIS, E S., SWENSEN, S. M., WILLIAMS, S. E., GADEK, P. A., QU~NN, C. J., EGUIARTE, L. E., GOLENBERG, E., LEARN, G. H. J., GRAHAM, S. W., BARRETT, S. C. H., DAYANANDAN, S., ALBERT, V. A., 1993: Phylogenetics of seed plants: an analysis of nucleotide sequences from the plastid gene rbcL. - A n n . Missouri Bot. Gard. 80: 528-580.

Utility of ITS sequences in systematics of Teucrium 67
CLEGG, M. T., 1993: Chloroplast gene sequences and the study of plant evolution. - Proc. Natl. Acad. Sci. USA 90: 363-367.
COHEN, E., 1956: Contribution ä l 'étude des Teucrium marocains de la section Polium. - Trav. Inst. Sci. Chérifien Sér. Bot. Biol. Vég. 9: 3-85.
DIEZ, M. J., OJEDA, F., COLOMER, M., 1993: Contribución a la palinológia del género Teucrium L. (Lamiaceae) en la Penfnsula Ibérica e Islas Baleares y su interés taxonómico. - Lagascalia 17:119-134.
DONOOHUE, M. J., SANDERSON, M. J., 1992: The suitability of molecular and morphological evidence in reconstructing plant phylogeny. - In SOLT~S, P. S., SOLTIS, D. E., DOYLE, J. J., (Eds): Molecular systematics of plants, pp. 340-368. - London, New York: Chapman & Hall.
DOWNIE, S. R., KATz-DowNIE, D. S., 1996: A molecular phylogeny of Apiaceae subfamily Apioideae: evidence from nuclear ribosomal DNA internal transcribed spacer sequences. - Amer. J. Bot. 83: 234-251.
DOYLE, J. J., 1991: DNA protocols for plants: CTAB total DNA extraction. - In HEwirr, G. M., JOHNSTOY, A. W. B., Yotn~G, J. P. W., (Eds): Molecular techniques in taxonomy, pp. 283-285. - Berlin, Heidelberg, New York: Springer.
DOYLE, J. L., 1990: Isolation of plant DNA from fresh tissue. - Focus 12: 13-15. EL OtJALIDI, J., 1987: Approche de l'étude biosystématique des Teucrium de la section
Polium (Lamiaceae) du Maroc. Contribution ä la "Flore pratique du Maroc". - Sciences de l"Evolution et Ecologie, Université Montpellier 2, France.
- 1991: Biosystematique et taxinomie des Teucrium de la section Polium (Lamiaceae) dans le bassin méditerranéen occidental. Differents aspects de la variation au Maroc, en France et en Espagne. - Thèse Doct., Université Montpellier 2, France.
- PUECH, S., 1993: Quelques marqueurs morphologiques des Teucrium section Polium du Maroc. Valeurs diagnostiques ä differents niveaux d'integration. - Acta Bot. Malac. 18: 163-173.
- MARTIN, A., PUECrI, S., 1996a: Le polymorphisme du calice chez Teucrium dunense (Labiatae): son maintien sur les dunes du littoral. - Acta Bot. Gall. 143: 55-63.
- RASCOL, J. P., MARTIN, A., PUECH, S., 1996b: Le poliumoside, marqueur chimique de la secfion Polium du genre Teucrium (Labiatae): l'exception du T. mideltense, espèce endémique du Maroc. - Biochem. Syst. Ecol. 24: 261-272.
EL OUALIDI, J., MARTIN, A., NAVARRO, T., PVECH, S., 1998: Growth variability of six mediterranean taxa of the Teucrium lusitanicum SCHREB. complex (Labiatae) and its systematic relationship. - Acta Bot. Gall. 145: 49-68.
FEr,~ANE, M., MArHEZ, J., 1992: Elements pour la flore pratique du Maroc. - Naturalia Monspel., Sér. Bot. 56: 171-220.
FERNAYDEZ, A., LEIXAO, M., 1984: Contribution ä l 'étude cytotaxonomique des spermatophytes du Portugal XVIII. - Mém. Soc. Brot. 27: 27-75.
FERNANDEZ CASAS, J., GONZALEZ, AGUILERA, J., Rulz REJÓN, M., 1978: Notas sobre la cariologfa de Lamiaceae. - Anales Jard. Bot. Madfid 34: 723-724.
GADEK, P. A., FERNANDO, E. S., QU~NN, C. J., HOOT, S. B., TERREZAS, T., SHEAHAN, M. C., CHASE, M. W., 1996: Sapinadales: molecular delimitation and infraordinal groups. - Amer. J. Bot. 83: 802-811.
GAMMAR, Z., NABLI, M., PUECH, S., 1989: Contribution ä l 'étude biologique et caryosystematique des Teucrium (Labiatae) de Tunisie. - Naturalia Monspel., Sér. Bot. 54: 79-92.
GvzY, M. R., EHDANE, B., WAINES, J. G., 1989: Yield and its components in diploId, tetraploid and hexaploid wheats in diverse environments. - A n n . Bot. 64: 635- 642.

68 J. EL. OUALmI & al.:
HARLAN, J. R., DE WET, J. M. J., 1975: On Ö. WINGE and a prayer: the origins of polyploidy. - Bot. Rev. 41 361-390.
HARTE, D. L., CEARK, A. G., 1989: Principles of population genetics. 2nd edn. - Sundefland: Mass.: Sinauer.
HEDGE, I. C., MILEER, A. G., 1977: New and interesting taxa from N.E. tropical Africa. - Notes Roy. Bot. Gard. Edinburgh 35: 179-191.
HILEIS, D. M., HUEESENBECK, J. P., 1992: Signal, noise, and reliability in molecular phylogenefic analyses. - J. Heredity 83: 189-195.
HOEMGREN, P. K., HOLMGREN, N. H., BARNETT, L. C., 1990: Index Herbariorum. Part I: The herbaria of the world. 8th edn. - Regnum Veg. 120. - Bronx, New York: Botanical Garden.
HOUARD, C., 1906: Modificafions histologiques produites par des Copium dans les fleurs des Teucrium. - Marcellia 5: 83-101.
KÄSTNER, A., 1986: Beiträge zur Wuchsformendifferenzierung und systematischen Gliederung von Teucrium L. V. Wuchsformen und Verbreitung von Arten der Verwandtschaftskreise von T. marum und T. chamaedrys, sect. Chamaedrys. - Flora 178: 111-138.
- 1989: Übersicht zur systematischen Gliederung der Gattung Teucrium L. - Biocosme Mésogéen 6: 63-78.
KI~G, R., 1988: Studies in the flora of Arabia XIX. Teucrium in the Arabian Peninsula and Socotra. - Notes Roy. Bot. Gard. Edinburgh. 45: 21-42.
KÜPFER, P., 1969: Recherches cytotaxinomique sur la flore des montagnes de la Peninsule Ibérique.- Bull. Soc. Neuchateloise Sci. Nat. 92: 31-48.
LÄZARO IBIZA, B., 1896: Botänica descriptiva. Compendio de la Flora Espafiola, 2. - Madrid: Hemendo.
LEVlN, D. A., 1975: Minority cytotype exclusion in local plant populations. - Taxon 24: 35- 43.
MADDISON, W. P., MADDISON, D. R., 1992: MacClade: analysis of phylogeny and character evolution. Version 3.04. - Sunderland, Mass.: Sinauer.
MARKOVA, M. L., 1982: Chromosome number reports. - Taxon 31: 776-777. MCDADE, L. A., 1995: Hybridization and phylogenetics. - In HocH, P. C., STEmENSOY, A.
G., (Eds): Experimental and molecular approaches to plant biosystematics, pp. 305- 331. - St. Louis: Missouri Botanical Garden.
NAßEr, M. A., 1967: Contribution ä l 'étude taxinomique et écologique des espèces tunisiennes du genre Teucrium L. - Thèse 3ème Cycle, Université Montpellier 2, France.
NAVARRO, T., 1988: Estudios biosistemäticos en el género Teucrium (sección Polium
subsección Polium) en la Peninsula Ibérica (Lamiaceae). - Tesis doctoral, Universidad Granada, Espafia.
- 1995: Revisión del género Teucrium sect. Polium (MILL.) SCrIREB., en la Peninsula Ibérica e Baleares. - Acta Bot. Malac. 20: 163-275.
PHmIPPE, H., 1993: MUST: A computer package of management utilities for sequences and trees. - Nucl. Acids Res. 21: 5264-5272.
POTTIZR-ALAPErrrrE, G., 1981: Teucrium L. Flore de la Tunisie 2, pp. 764-776. Ministére de l'Enseignement Supérieur et de la Recherche Scientifique of Ministére de l'Agriculture, Tunis.
PVECH, S., 1976: Recherches de biosystématique sur les Teucrium (Labiées) de la section Polium du bassin méditerranéen occidental (Espagne et France). - Thèse Doct. Sci., Université Montpellier 2, France.

Utility of ITS sequences in systematics of Teucrium 69
- 1984: Les Teucrium (Labiées) de la sect. Polium du bassin Méditerranéen occidental (France et Peninsule Ibérique). - Naturalia Monspel., Sér AS.
QugzEL, E, SANXA, S., 1962: Nouvelle flore de l'Algerie et des régions désertiques méridionales. - Paris: C. N. R. S.
RIVAS MARTINEZ, S., 1974: Sobre el Teucrium pumi lum L. (Labiatae) y sus especies afines. - Anales Jard. Bot. Madrid 31: 79-96.
- 1987: Memoria del mapa de series de vegetación de Espafia. - Madrid. ROSÜA, J. L., NAVARRO, T., 1986: N6meros cromosomäticos para la flora espafiola. -
Lagascalia 14: 282-286. SAITOU, N., NEI, M., 1987: The Neighbor-Joining method: a new method for reconstructing
phylogenetic trees. - Molec. Biol. Evol. 4: 406-425. SAN~ER, E, N~CKLEN, S., COULSON, A. R., 1977: DNA sequencing with chain terminating
inhibitors. - Proc. Natl. Acad. Sci. USA 74: 5463-5467. SAUVAGE, CH., VINDT~ J., 1955: Synopsis du genre Teucrium sect. Polium au Maroc. - Bull.
Soc. Phys. Maroc 35: 283-293. - 1965: Synopsis du genre Teucrium sect. Polium au Maroc (suite et fin). - Naturalia
Monspel., Ser. Bot. 16: 195-200. SCIJmLIN~, E. E., PAN~RO, J. L., 1996: Phylogenetic reticulation in subtribe Helianthinae. -
Amer. J. Bot. 83: 939-948. SLATKIN, M. K., 1987: Gene flow and the geographic structure of natural populations. -
Science 236: 787-792. SOLT~S, E S., Kuzow, R. K., 1993: ITS sequence variation within and among populations
of Lomat ium grayi and L. laevigatum (UmbelIi ferae). - Molec. Phylogenet. Evol. 2: 166-170.
DOYLE, J. J., SOLTIS, D. E., 1992: Molecular data and polyploid evolution in plants in reconstructing plant phylogeny. - In SOLTIS, E S., SOLXIS, D. E., DOYLE, J. J., (Eds): Molecular systematics of plants, pp. 177-201. - New York, London: Chapman & Hall.
SOSEF, M. S. M., 1997: Hierarchical models, reticulate evolution and the inevitability of paraphyletic supraspecific taxa. - Taxon 46: 75-85.
STEAY, D. A., OLMSTEAD, R. G., 1996: Molecular systematics of Clerodendrum L. s. 1. (Lamiaceae). - Amer. J. Bot. 83(6, Suppl.): 194.
SXEARNS, S. C., 1989: The evolutionary significance of phenotypic plasticity. Phenotypic sources of variation among organisms can be described by developmental switches and reaction norms. - BioScience 39: 436--445.
STRIMM~R, K., Von HAZSELER, A., 1996: Quartet Puzzling: a quartet Maximum-Likelihood method for reconstructing tree topologies. - Molec. Biol. Evol. 13: 964-969.
Sun, Y., THIZN, L. B., REEW, H. E., ZIM~vxn~, E. A., 1993: Molecular evolution and phylogenetic implications of internal transcribed spacer sequences of ribosomal DNA in W i n t e r a c e a e . - Amer. J. Bot. 80: 1042-1055.
SWOFFORD, D., 1993: PAUP 3.1.1: phylogenetic analysis using parsimony. - Champaign, II1.: Illinois Natural History Survey.
VALDgS BZRMEJO, E., SÄNCI-mZ Cp, zsPo, A., 1978: Datos cariológicos sobre el género Teuerium L. (Labiatae) en la Peninsula Ibérica. - Acta Bot. Malac. 4: 27-54.
VENKATESWARLU, K., NAZAR, R., 1991: A conserved core structure in the 18-25S rRNA intergenic region from tobacco, Nieot iana rustica. - Pl. Molec. Biol. 17: 189-194.
WBI~EL, J. F., SCHNABEL, A., SEELANAN, T., 1995: Bidirectional interlocus concerted evolution follwing allopolyploid speciation in cotton (Gossypium). - Proc. Natl. Acad. Sci. USA 92: 280-284.

70 J. EL. OUALIDI Æ al.: Utility of ITS sequences in systematics of Teucrium
WILLKOMM, H. M., LANGE, J., 1870: Prodromus florae hispanicae, 2. - Stuttgart: Schweizerbart.
WOJCIECHOWSKI, M. E, SANDERSON, M. J., BALDWlN, B. G., DONOGHUE, M. J., 1993: Monophyly of aneuploid Astragalus (Fabaceae): evidence from nuclear ribosomal DNA intemal transcribed spacer sequences. - Amer. J. Bot. 80: 711-722.
YtJAN, Y. M., KüP~R, P., DoYLz, J. J., 1996: Infrageneric phylogeny of the genus Gentiana (Gentianaceae) inferred from nucleotide sequences of the intemal Transcribed Spacers (ITS) of nuclear ribosomal DNA. - Amer. J. Bot. 83: 641-652.
Addresses of the authors: JALAL EL OUALIDI (correspondence), Institut Scientifique, Département de Botanique et d'Ecologie Végétale, Université Mohammed V, B.R 703, Rabat, Marocco. - OLIVIER VERNEAU, SUZETTE PUECH, JEAN-Yvzs Du8uISSON, Institut des Sciences de l'Evolution, UMR 5554 CNRS, Université de Montpellier II, Place E. Bataillon, F-34095 Montpellier Cedex 05, France.
Accepted November 4, 1997 by K. BmaM~R