the amygdala: site of genomic and nongenomic arousal of aldosterone-induced sodium intake
TRANSCRIPT

Kidney International, Vol. 57 (2000), pp. 1337–1345
The amygdala: Site of genomic and nongenomic arousal ofaldosterone-induced sodium intake
RANDALL R. SAKAI, BRUCE S. MCEWEN, STEVE J. FLUHARTY, and LI YUN MA
Department of Animal Biology, University of Pennsylvania, Philadelphia, Pennsylvania, and Lab of Neuroendocrinology,Rockefeller University, New York, New York, USA
The amygdala: Site of genomic and nongenomic arousal of effects on the brain has not yet been completely eluci-aldosterone-induced sodium intake. dated, their actions include both rapid nongenomic
Background. Mineralocorticoids act on the brain to influ- mechanisms that may involve GABA receptors [4–9]ence sodium intake, and they do so via intracellular type Iand slower genomic induction incorporating cytosolicreceptors and possibly also via a direct membrane action, asmineralocorticoid receptors (MRs) [10, 11] that involvethey do in the kidney. One brain area implicated by lesion
studies investigating the regulation of sodium appetite aroused a synergy of MR with other neuropeptides, especiallyby adrenal steroids is the amygdala. angiotensin II [11–14], as well as suppression of other
Methods. To examine the mechanism by which mineralocor- neuropeptides, most notably oxytocin [15–17], that nor-ticoids act in the amygdala to arouse salt intake via a genomicmally inhibit sodium ingestion.and or membrane mode of action, rats were bilaterally fitted
with cannulae directed to terminate in the amygdala. The geno- Here, we review a series of studies that support themic action of mineralocorticoids in arousing sodium intake was findings that mineralocorticoids act on the brain to in-investigated by the administration of antisense oligodeoxy- fluence sodium intake, and that they do so via intracellu-nucleotides (ASDNs) against the mineralocorticoid receptor,
lar type I receptors and also via direct membrane action,and its effects on deoxycorticosterone (DOCA)-induced so-as they do in kidney and vascular tissue. Although thedium intake over the course of several days was examined.
The nongenomic action of mineralocorticoids on sodium intake mineralocorticoids acting through classic genomic mech-was investigated by implantation into the amygdala of DOCA, anisms to arouse sodium intake have been well docu-aldosterone (ALDO), or their A-ring–reduced tetrahydro de- mented, their novel nongenomic mode of action throughrivatives, 15 minutes prior to access to saline. Sodium intake
membrane receptors that may engage the GABA/benzo-was monitored immediately thereafter.diazepine receptor or a unique receptor system of theirResults. Treatment of rats in the amygdala with ASDN against
the mineralocorticoid receptor inhibited DOCA-induced so- own in producing a rapid increase in sodium intake isdium intake, whereas ASDN against the glucocorticoid recep- quite intriguing. Furthermore, it seems that the braintor or sense/scrambled sequences had no effect. DOCA and
nuclei in the amygdala possess the receptors that allowALDO increased saline intake within 15 minutes after steroidthe mineralocorticoids to act through both genomic andapplication. Similarly, the application of A-ring–reduced 3b,5b
tetrahydroaldosterone and 5 a-tetrahydrodeoxycorticosterone nongenomic mechanisms to arouse sodium intake.produced the same increases in sodium intake. Previous studies have used receptor antagonists to ex-
Conclusions. Together, the data imply that adrenal steroids, amine the contribution of MRs and glucocorticoid recep-in addition to acting through classic cytosolic receptors, maytor (GRs) in sodium appetite [11]. Their use has metalso act on membrane receptor systems, producing rapidwith some success by partially inhibiting sodium appetitechanges in behavior.aroused by either the exogenous administration of aldo-sterone (ALDO) or deoxycorticosterone (DOCA) or by
The arousal of salt appetite by mineralocorticoids is sodium depletion treatments, which elevate the endoge-an innate and highly specific behavior [1, 2]. Rats treated nous levels of mineralocorticoids [11, 13]. However, theirexogenously with mineralocorticoids exhibit a robust in- use has been limited because of their lack of specificitygestion of sodium-rich ingesta and will even perform for receptor subtypes, the short half-life of most antago-operant tasks to obtain saline rewards [3]. Although the nists, and the lack of regional localization or sites ofmechanism through which mineralocorticoids evoke their action when injected into the brain ventricular system
[18]. As an alternative method to investigate adrenalsteroid receptor involvement in sodium appetite, weKey words: sodium appetite, amygdala, adrenal steroids, A-ring re-
duced steroids, ALDO, DOCA, flunitrazepam. have used antisense oligodeoxynucleotides (ASDNs)that interfere with endogenous receptor activity. By their 2000 by the International Society of Nephrology
1337

Sakai et al: Genomic and nongenomic arousal of sodium intake1338
design, the ASDN sequences were designed to interfere was initiated. Before inclusion in experiments, cannulaplacement and patency were validated by a pulse ICVwith the type I MRs, thus interfering with the genomicinjection of 6 ng of angiotensin II to which a criterionactions of ALDO.response of at least 5 mL of water intake in 15 minutesThe subnuclei of the amygdala has been identified aswas required for participation in subsequent experiments.containing large concentrations of MRs and that electri-
A second group of male rats was surgically fitted withcal or chemical lesions of this brain area render thebilateral guide cannula stereotaxically directed to termi-animal insensitive to mineralocorticoid-induced sodiumnate above the medial area of the amygdala. Surgery wasintake [19, 20]. Similarly, we have found that local contin-performed under ketamine/acepromazine anesthesia (40uous infusions of MR antagonists into this are also effec-mg Ket, 20 mg Ace/100 g body wt). The guide cannulastive in suppressing mineralocorticoid-induced sodium in-were permanently fixed to the skull with stainless steeltake [21]. As a further extension of the studies examiningbone screws and dental acrylic. Once fitted with guidethe genomic actions of mineralocorticoids on sodium in-cannula, the animals were allowed at least one week oftake, the first series of experiments investigates whetherrecovery before the experiments were begun.ASDNs, which target the MRs, could reverse an estab-
lished DOCA-induced salt appetite. Based on the find- Oligodeoxynucleotide probesings of the identification of a brain-sensitive site (the
Antisense oligodeoxynucleotide sequences were de-amygdala) to mineralocorticoids, the effects of directsigned on sequences of the cloned rat MR [22] and GR.administration of various adrenal steroids on salt intakeThe MR antisense oligonucleotide sequence, 15 mer,and their mechanism of action are further examined inwhich was used in the experiments, corresponds to nucle-the second series of studies. Together, the two series ofotides 2 to 16 of the 59 end of the translated region ofstudies examine whether mineralocorticoids can act inthe MR mRNA and was 59 CAT-GTC-TAG-GCC-TTCthe amygdala through genomic and nongenomic mecha-39. The GR antisense oligonucleotide sequence, 18 mer,nisms to arouse sodium appetite in the rat.was also 59 directed to the ATG initiation site of theGR mRNA and was 59 GGA-TTC-TTT-GGA-GTC-
METHODS CAT 39. In addition, the control sequences used wereMR sense, 59 GAA-GGC-CTA-GAC-ATG 39; GR sense,Animals59 ATG-GAC-TCC-AAA-GAA-TCC 39; MR scram-Adult, male Harlan Sprague-Dawley rats (N 5 6 perbled, 59 CAT-TTT-GAA-GGT-TCC 39; and GR scram-experimental group) weighing 300 to 350 g at the initia-bled, 59 GGA-TTT-GAA-GGT-TCC 39. All nucleotide
tion of the experiment were individually housed in metal sequences were synthesized by Oligo’s Etc. Inc. (Wilson-cages in a temperature controlled room (22 6 28C) and ville, OR, USA) and contained phosphorothioate modi-maintained on a 12-hour light/dark cycle. The animals fications to reduced susceptibility to nuclease degrada-were given ad libitum access to Purina rodent chow, tap tion in vivo. One microliter pulse ICV (pICV) injectionswater, and 0.5 mol/L NaCl, unless otherwise noted. Daily of oligonucleotides were given on three consecutive daysfluid intakes were recorded to the nearest milliliter. to conscious animals using an injector (Plastics 1), which
terminated 1 mm past the end of the guide cannula.SurgeryControl animals received equivalent amounts of either
Rats receiving intracerebroventricular (ICV) injec- scrambled/sense oligodeoxynucleotides or isotonic salinetions were anesthetized under Ketamine HCl (intramus- vehicle. The ASDN sequences for MR and GR at thecularly 100 mg/kg body wt; Bristol Laboratories, Syra- doses used have been previously shown to decrease thecuse, NY, USA) and Acepromazine (intramuscularly targeted receptors by 40 to 60% [23].1.37 mg/kg body weight; Ayerst Laboratories, Inc., NewYork, NY, USA) anesthesia and were injected with a Steroidsprophylactic dose of Gentamicin (10 mg intramuscularly; Aldosterone, DOCA, corticosterone (CORT), 3b,5bSchering-Plough Animals Health Corp., Kenilworth, NJ, tetrahydroaldosterone (TH-ALDO), 3b,5b tetrahydro-USA) prior to surgery. Rats were fitted with an ICV corticosterone (TH-CORT), and 5 a-tetrahydrodeoxy-guide cannula (Plastics 1, Roanoke, VA, USA) that was corticosterone (TH-DOC) were acquired from Sigmastereotaxically implanted to terminate in the third ventri- Chemical Company (St. Louis, MO, USA). Allopregna-cle and fixed to the skull with dental acrylic and three nolone (3 a OH DHP) was a gift from CoCensys Inc..stainless-steel screws. The following stereotaxic coordi- Ru28362 was received as a gift from Roussel-UCLAFnates were used: 1.0 mm posterior to bregma, 1.0 mm (Romanville, France).lateral to the sinus and angled toward the midline at 108,
Adrenal steroid receptor antagonistsand 7.5 mm ventral from the dura. One week of recoverywas allowed for daily food and fluid intake, as well as The role of the cytosolic mineralocorticoid and GR
in the elicitation of the rapid increase in salt appetite bybody weight, to return to presurgery levels before testing
Sakai et al: Genomic and nongenomic arousal of sodium intake 1339
application of local steroids was examined by interferingwith their function using either the type I MR antagonistRU 28318 or the type II GR antagonist RU 486 (Roussel-UCLAF). RU 28318 or RU 486 was applied for one hourthrough the same guide cannula as the steroid implantcannula and was then removed just prior to applicationof the steroids of interest. Salt intake was measured asdescribed in each experiment.
GABA/benzodiazepine receptor manipulations
In a separate set of rats maintained on normal food,the effects of local application of the GABA agonistflunitrazepam (Sigma) on salt appetite were examined.Because of the high binding affinity of the A-ring–reducedTH steroids to the GABAa/benzodiazepine receptor, weexamined whether the effects of the TH steroids couldbe mimicked by this GABA agonist. To further elucidatethe role of the endogenous GABA/benzodiazepine re-ceptor complex, the GABA receptor antagonist Ro15-
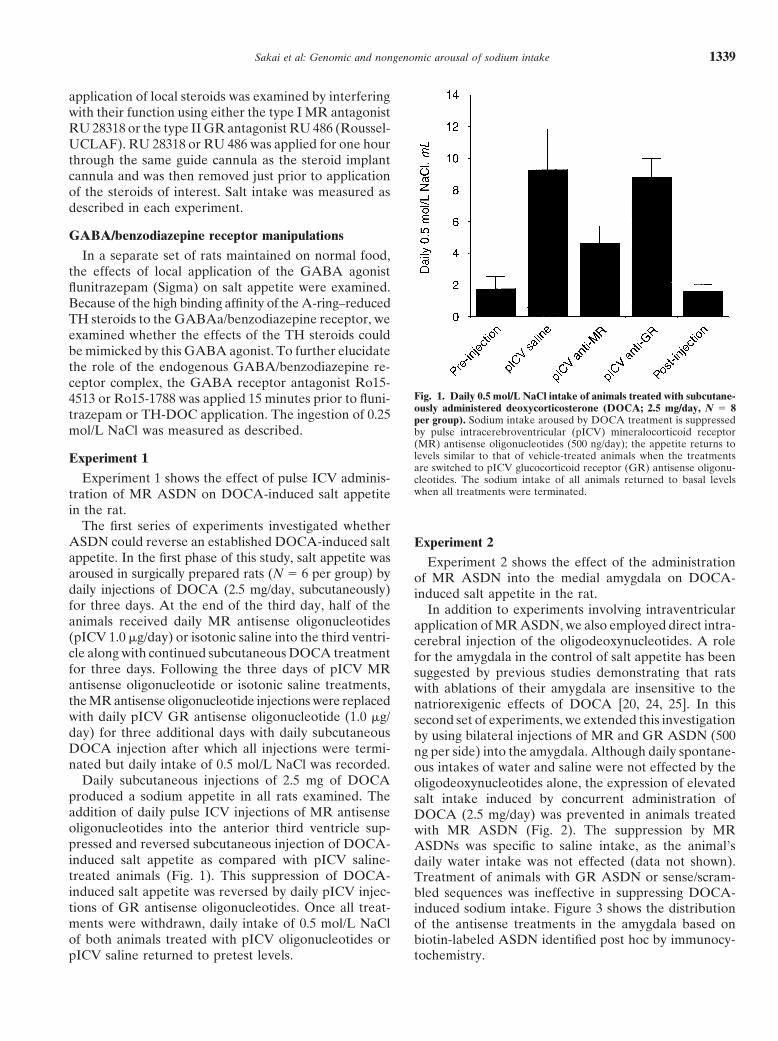
Fig. 1. Daily 0.5 mol/L NaCl intake of animals treated with subcutane-4513 or Ro15-1788 was applied 15 minutes prior to fluni-ously administered deoxycorticosterone (DOCA; 2.5 mg/day, N 5 8trazepam or TH-DOC application. The ingestion of 0.25 per group). Sodium intake aroused by DOCA treatment is suppressed
mol/L NaCl was measured as described. by pulse intracerebroventricular (pICV) mineralocorticoid receptor(MR) antisense oligonucleotides (500 ng/day); the appetite returns tolevels similar to that of vehicle-treated animals when the treatmentsExperiment 1are switched to pICV glucocorticoid receptor (GR) antisense oligonu-
Experiment 1 shows the effect of pulse ICV adminis- cleotides. The sodium intake of all animals returned to basal levelswhen all treatments were terminated.tration of MR ASDN on DOCA-induced salt appetite
in the rat.The first series of experiments investigated whether
ASDN could reverse an established DOCA-induced salt Experiment 2appetite. In the first phase of this study, salt appetite was Experiment 2 shows the effect of the administrationaroused in surgically prepared rats (N 5 6 per group) by of MR ASDN into the medial amygdala on DOCA-daily injections of DOCA (2.5 mg/day, subcutaneously) induced salt appetite in the rat.for three days. At the end of the third day, half of the In addition to experiments involving intraventricularanimals received daily MR antisense oligonucleotides application of MR ASDN, we also employed direct intra-(pICV 1.0 mg/day) or isotonic saline into the third ventri- cerebral injection of the oligodeoxynucleotides. A rolecle along with continued subcutaneous DOCA treatment for the amygdala in the control of salt appetite has beenfor three days. Following the three days of pICV MR suggested by previous studies demonstrating that ratsantisense oligonucleotide or isotonic saline treatments, with ablations of their amygdala are insensitive to thethe MR antisense oligonucleotide injections were replaced natriorexigenic effects of DOCA [20, 24, 25]. In thiswith daily pICV GR antisense oligonucleotide (1.0 mg/ second set of experiments, we extended this investigationday) for three additional days with daily subcutaneous by using bilateral injections of MR and GR ASDN (500DOCA injection after which all injections were termi- ng per side) into the amygdala. Although daily spontane-nated but daily intake of 0.5 mol/L NaCl was recorded. ous intakes of water and saline were not effected by the
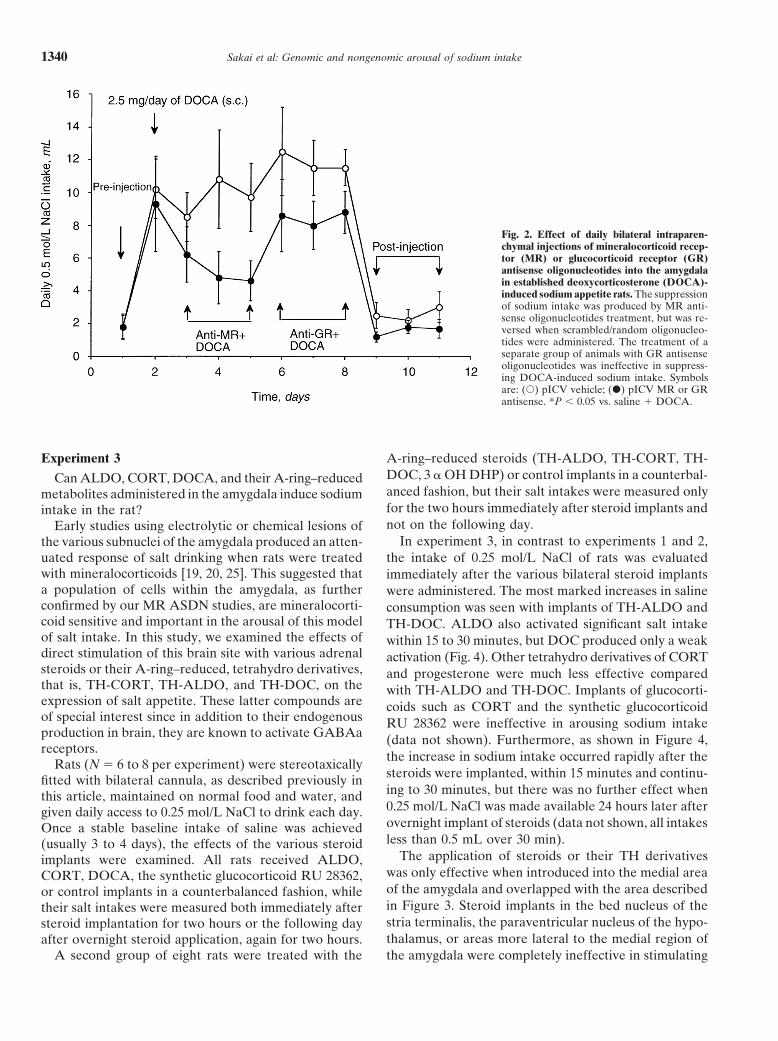
Daily subcutaneous injections of 2.5 mg of DOCA oligodeoxynucleotides alone, the expression of elevatedproduced a sodium appetite in all rats examined. The salt intake induced by concurrent administration ofaddition of daily pulse ICV injections of MR antisense DOCA (2.5 mg/day) was prevented in animals treatedoligonucleotides into the anterior third ventricle sup- with MR ASDN (Fig. 2). The suppression by MRpressed and reversed subcutaneous injection of DOCA- ASDNs was specific to saline intake, as the animal’sinduced salt appetite as compared with pICV saline- daily water intake was not effected (data not shown).treated animals (Fig. 1). This suppression of DOCA- Treatment of animals with GR ASDN or sense/scram-induced salt appetite was reversed by daily pICV injec- bled sequences was ineffective in suppressing DOCA-tions of GR antisense oligonucleotides. Once all treat- induced sodium intake. Figure 3 shows the distributionments were withdrawn, daily intake of 0.5 mol/L NaCl of the antisense treatments in the amygdala based onof both animals treated with pICV oligonucleotides or biotin-labeled ASDN identified post hoc by immunocy-
tochemistry.pICV saline returned to pretest levels.
Sakai et al: Genomic and nongenomic arousal of sodium intake1340
Fig. 2. Effect of daily bilateral intraparen-chymal injections of mineralocorticoid recep-tor (MR) or glucocorticoid receptor (GR)antisense oligonucleotides into the amygdalain established deoxycorticosterone (DOCA)-induced sodium appetite rats. The suppressionof sodium intake was produced by MR anti-sense oligonucleotides treatment, but was re-versed when scrambled/random oligonucleo-tides were administered. The treatment of aseparate group of animals with GR antisenseoligonucleotides was ineffective in suppress-ing DOCA-induced sodium intake. Symbolsare: (s) pICV vehicle; (d) pICV MR or GRantisense. *P , 0.05 vs. saline 1 DOCA.
Experiment 3 A-ring–reduced steroids (TH-ALDO, TH-CORT, TH-DOC, 3 a OH DHP) or control implants in a counterbal-Can ALDO, CORT, DOCA, and their A-ring–reducedanced fashion, but their salt intakes were measured onlymetabolites administered in the amygdala induce sodiumfor the two hours immediately after steroid implants andintake in the rat?not on the following day.Early studies using electrolytic or chemical lesions of
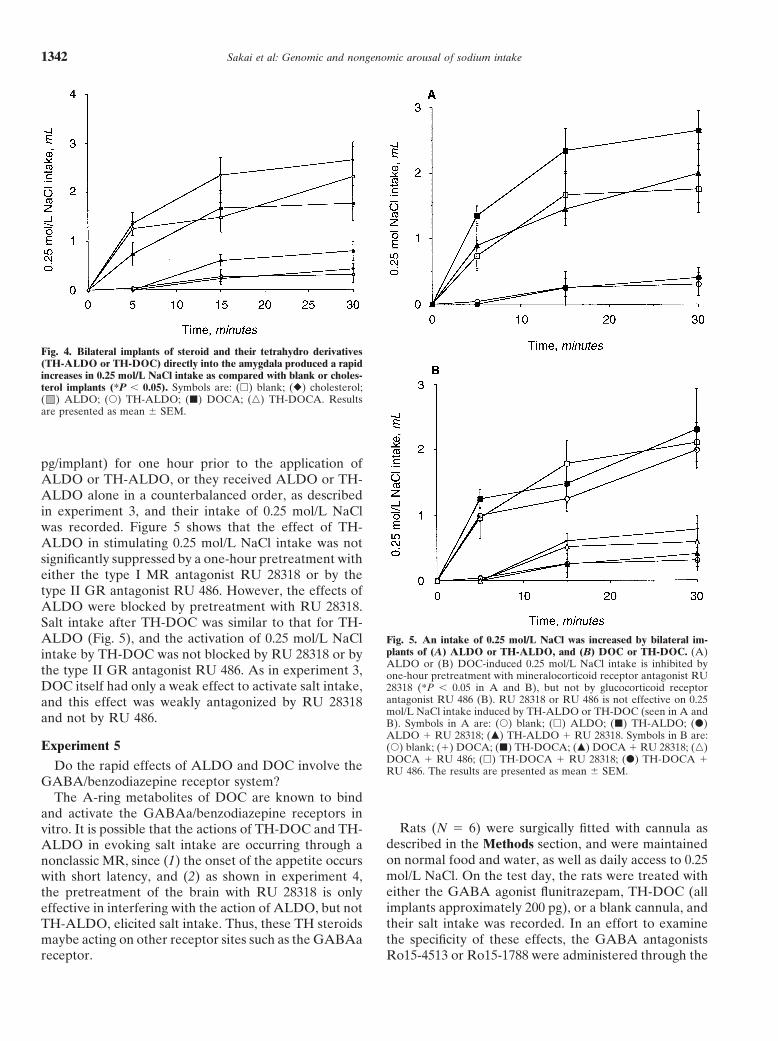
In experiment 3, in contrast to experiments 1 and 2,the various subnuclei of the amygdala produced an atten-uated response of salt drinking when rats were treated the intake of 0.25 mol/L NaCl of rats was evaluatedwith mineralocorticoids [19, 20, 25]. This suggested that immediately after the various bilateral steroid implantsa population of cells within the amygdala, as further were administered. The most marked increases in salineconfirmed by our MR ASDN studies, are mineralocorti- consumption was seen with implants of TH-ALDO andcoid sensitive and important in the arousal of this model TH-DOC. ALDO also activated significant salt intakeof salt intake. In this study, we examined the effects of within 15 to 30 minutes, but DOC produced only a weakdirect stimulation of this brain site with various adrenal activation (Fig. 4). Other tetrahydro derivatives of CORTsteroids or their A-ring–reduced, tetrahydro derivatives, and progesterone were much less effective comparedthat is, TH-CORT, TH-ALDO, and TH-DOC, on the with TH-ALDO and TH-DOC. Implants of glucocorti-expression of salt appetite. These latter compounds are coids such as CORT and the synthetic glucocorticoidof special interest since in addition to their endogenous RU 28362 were ineffective in arousing sodium intakeproduction in brain, they are known to activate GABAa (data not shown). Furthermore, as shown in Figure 4,receptors.
the increase in sodium intake occurred rapidly after theRats (N 5 6 to 8 per experiment) were stereotaxically
steroids were implanted, within 15 minutes and continu-fitted with bilateral cannula, as described previously ining to 30 minutes, but there was no further effect whenthis article, maintained on normal food and water, and0.25 mol/L NaCl was made available 24 hours later aftergiven daily access to 0.25 mol/L NaCl to drink each day.overnight implant of steroids (data not shown, all intakesOnce a stable baseline intake of saline was achievedless than 0.5 mL over 30 min).(usually 3 to 4 days), the effects of the various steroid
The application of steroids or their TH derivativesimplants were examined. All rats received ALDO,was only effective when introduced into the medial areaCORT, DOCA, the synthetic glucocorticoid RU 28362,of the amygdala and overlapped with the area describedor control implants in a counterbalanced fashion, whilein Figure 3. Steroid implants in the bed nucleus of thetheir salt intakes were measured both immediately afterstria terminalis, the paraventricular nucleus of the hypo-steroid implantation for two hours or the following daythalamus, or areas more lateral to the medial region ofafter overnight steroid application, again for two hours.
A second group of eight rats were treated with the the amygdala were completely ineffective in stimulating
Sakai et al: Genomic and nongenomic arousal of sodium intake 1341
Fig. 3. Coronal section of rat brain, taken from the Paxinos and Watson Rat Brain Atlas, which shows the location of the cannula terminatingin the amygdala. Only rats in which their implants terminated in this brain area were used in the analysis.
saline intake either immediately after steroid application the amygdala, may be dependent on prior primingor over the subsequent 24-hour period (data not shown). through a background of the genomic actions of the
adrenal steroids. That is, the TH-ALDO/ALDO-inducedExperiment 4 saline intake may be operating in the context of adrenal
steroid action, which is of genomic nature and whichCan the rapid effects of adrenal steroid-induced saltintake be blocked by agents that antagonize the classic prepares the brain to respond to the rapid salt appetite-
eliciting effects of these steroids. To interfere with theseadrenal steroid receptor?The rapid effects of ALDO and DOC, which induce genomic actions of the mineralocorticoids, rats (N 5 6)
were pretreated with the MR antagonist RU 28318 (200salt intake within 15 minutes after steroid application in
Sakai et al: Genomic and nongenomic arousal of sodium intake1342
Fig. 4. Bilateral implants of steroid and their tetrahydro derivatives(TH-ALDO or TH-DOC) directly into the amygdala produced a rapidincreases in 0.25 mol/L NaCl intake as compared with blank or choles-terol implants (*P , 0.05). Symbols are: (h) blank; (r) cholesterol;( ) ALDO; (s) TH-ALDO; (j) DOCA; (n) TH-DOCA. Resultsare presented as mean 6 SEM.
pg/implant) for one hour prior to the application ofALDO or TH-ALDO, or they received ALDO or TH-ALDO alone in a counterbalanced order, as describedin experiment 3, and their intake of 0.25 mol/L NaClwas recorded. Figure 5 shows that the effect of TH-ALDO in stimulating 0.25 mol/L NaCl intake was notsignificantly suppressed by a one-hour pretreatment witheither the type I MR antagonist RU 28318 or by thetype II GR antagonist RU 486. However, the effects ofALDO were blocked by pretreatment with RU 28318.Salt intake after TH-DOC was similar to that for TH-ALDO (Fig. 5), and the activation of 0.25 mol/L NaCl Fig. 5. An intake of 0.25 mol/L NaCl was increased by bilateral im-
plants of (A) ALDO or TH-ALDO, and (B) DOC or TH-DOC. (A)intake by TH-DOC was not blocked by RU 28318 or byALDO or (B) DOC-induced 0.25 mol/L NaCl intake is inhibited bythe type II GR antagonist RU 486. As in experiment 3, one-hour pretreatment with mineralocorticoid receptor antagonist RU
DOC itself had only a weak effect to activate salt intake, 28318 (*P , 0.05 in A and B), but not by glucocorticoid receptorantagonist RU 486 (B). RU 28318 or RU 486 is not effective on 0.25and this effect was weakly antagonized by RU 28318mol/L NaCl intake induced by TH-ALDO or TH-DOC (seen in A andand not by RU 486. B). Symbols in A are: (s) blank; (h) ALDO; (j) TH-ALDO; (d)ALDO 1 RU 28318; (m) TH-ALDO 1 RU 28318. Symbols in B are:
Experiment 5 (s) blank; (1) DOCA; (j) TH-DOCA; (m) DOCA 1 RU 28318; (n)DOCA 1 RU 486; (h) TH-DOCA 1 RU 28318; (d) TH-DOCA 1Do the rapid effects of ALDO and DOC involve the RU 486. The results are presented as mean 6 SEM.
GABA/benzodiazepine receptor system?The A-ring metabolites of DOC are known to bind
and activate the GABAa/benzodiazepine receptors inRats (N 5 6) were surgically fitted with cannula asvitro. It is possible that the actions of TH-DOC and TH-
described in the Methods section, and were maintainedALDO in evoking salt intake are occurring through aon normal food and water, as well as daily access to 0.25nonclassic MR, since (1) the onset of the appetite occursmol/L NaCl. On the test day, the rats were treated withwith short latency, and (2) as shown in experiment 4,either the GABA agonist flunitrazepam, TH-DOC (allthe pretreatment of the brain with RU 28318 is onlyimplants approximately 200 pg), or a blank cannula, andeffective in interfering with the action of ALDO, but nottheir salt intake was recorded. In an effort to examineTH-ALDO, elicited salt intake. Thus, these TH steroidsthe specificity of these effects, the GABA antagonistsmaybe acting on other receptor sites such as the GABAa
receptor. Ro15-4513 or Ro15-1788 were administered through the
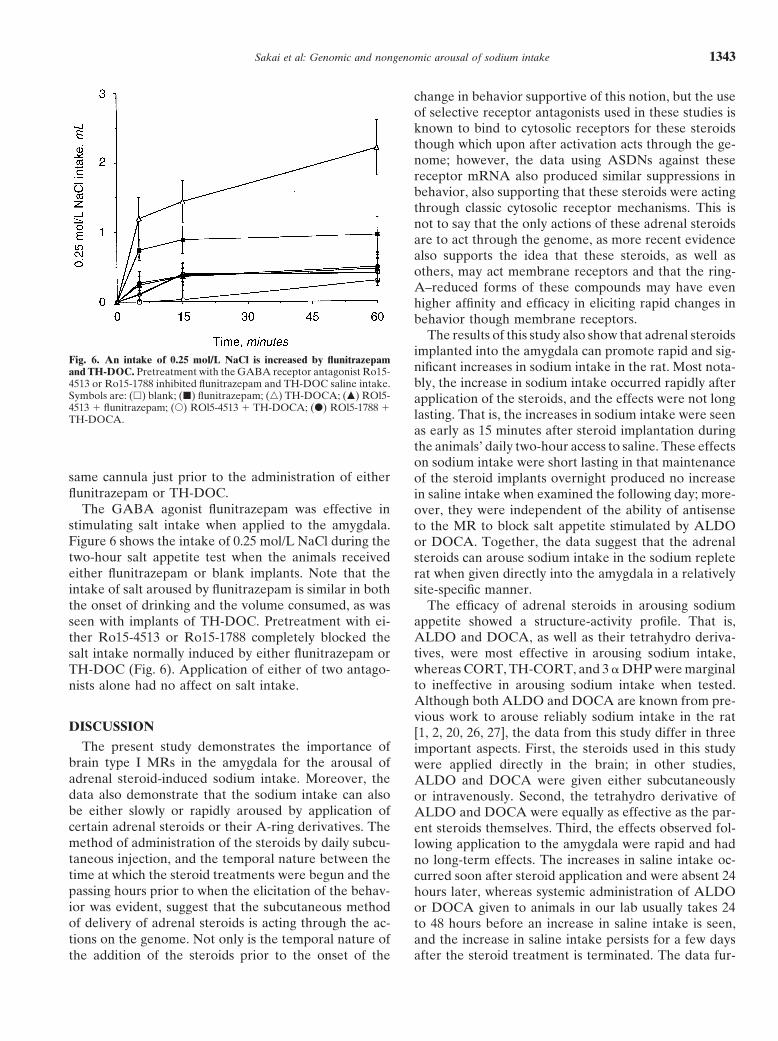
Sakai et al: Genomic and nongenomic arousal of sodium intake 1343
change in behavior supportive of this notion, but the useof selective receptor antagonists used in these studies isknown to bind to cytosolic receptors for these steroidsthough which upon after activation acts through the ge-nome; however, the data using ASDNs against thesereceptor mRNA also produced similar suppressions inbehavior, also supporting that these steroids were actingthrough classic cytosolic receptor mechanisms. This isnot to say that the only actions of these adrenal steroidsare to act through the genome, as more recent evidencealso supports the idea that these steroids, as well asothers, may act membrane receptors and that the ring-A–reduced forms of these compounds may have evenhigher affinity and efficacy in eliciting rapid changes inbehavior though membrane receptors.
The results of this study also show that adrenal steroidsimplanted into the amygdala can promote rapid and sig-
Fig. 6. An intake of 0.25 mol/L NaCl is increased by flunitrazepamnificant increases in sodium intake in the rat. Most nota-and TH-DOC. Pretreatment with the GABA receptor antagonist Ro15-bly, the increase in sodium intake occurred rapidly after4513 or Ro15-1788 inhibited flunitrazepam and TH-DOC saline intake.
Symbols are: (h) blank; (j) flunitrazepam; (n) TH-DOCA; (m) ROl5- application of the steroids, and the effects were not long4513 1 flunitrazepam; (s) ROl5-4513 1 TH-DOCA; (d) ROl5-1788 1
lasting. That is, the increases in sodium intake were seenTH-DOCA.as early as 15 minutes after steroid implantation duringthe animals’ daily two-hour access to saline. These effectson sodium intake were short lasting in that maintenance
same cannula just prior to the administration of either of the steroid implants overnight produced no increaseflunitrazepam or TH-DOC. in saline intake when examined the following day; more-
The GABA agonist flunitrazepam was effective in over, they were independent of the ability of antisensestimulating salt intake when applied to the amygdala. to the MR to block salt appetite stimulated by ALDOFigure 6 shows the intake of 0.25 mol/L NaCl during the or DOCA. Together, the data suggest that the adrenaltwo-hour salt appetite test when the animals received steroids can arouse sodium intake in the sodium repleteeither flunitrazepam or blank implants. Note that the rat when given directly into the amygdala in a relativelyintake of salt aroused by flunitrazepam is similar in both site-specific manner.the onset of drinking and the volume consumed, as was The efficacy of adrenal steroids in arousing sodium
appetite showed a structure-activity profile. That is,seen with implants of TH-DOC. Pretreatment with ei-ALDO and DOCA, as well as their tetrahydro deriva-ther Ro15-4513 or Ro15-1788 completely blocked thetives, were most effective in arousing sodium intake,salt intake normally induced by either flunitrazepam orwhereas CORT, TH-CORT, and 3 a DHP were marginalTH-DOC (Fig. 6). Application of either of two antago-to ineffective in arousing sodium intake when tested.nists alone had no affect on salt intake.Although both ALDO and DOCA are known from pre-vious work to arouse reliably sodium intake in the rat
DISCUSSION [1, 2, 20, 26, 27], the data from this study differ in threeThe present study demonstrates the importance of important aspects. First, the steroids used in this study
brain type I MRs in the amygdala for the arousal of were applied directly in the brain; in other studies,adrenal steroid-induced sodium intake. Moreover, the ALDO and DOCA were given either subcutaneouslydata also demonstrate that the sodium intake can also or intravenously. Second, the tetrahydro derivative ofbe either slowly or rapidly aroused by application of ALDO and DOCA were equally as effective as the par-certain adrenal steroids or their A-ring derivatives. The ent steroids themselves. Third, the effects observed fol-method of administration of the steroids by daily subcu- lowing application to the amygdala were rapid and hadtaneous injection, and the temporal nature between the no long-term effects. The increases in saline intake oc-time at which the steroid treatments were begun and the curred soon after steroid application and were absent 24passing hours prior to when the elicitation of the behav- hours later, whereas systemic administration of ALDOior was evident, suggest that the subcutaneous method or DOCA given to animals in our lab usually takes 24of delivery of adrenal steroids is acting through the ac- to 48 hours before an increase in saline intake is seen,tions on the genome. Not only is the temporal nature of and the increase in saline intake persists for a few days
after the steroid treatment is terminated. The data fur-the addition of the steroids prior to the onset of the

Sakai et al: Genomic and nongenomic arousal of sodium intake1344
ther suggest that the population of cells stimulated by behavior. The amygdala is one brain area in which popu-lations of both of these receptors coexist. The importancethe steroids may be acting via nonclassic cytosolic steroid
receptors. The inability of the type I (MR) antagonist of intact cell bodies in the amygdala is necessary for thenormal expression of mineralocorticoid-induced salineRU 28318 to block the effects of TH-ALDO–induced
sodium intake, along with the rapid elicitation of intake intake when the steroids are given systemically and arethen thought to exert their actions through genomicsoon after steroid treatment, suggests that these local
and rapid effects of steroids and their metabolites in mechanisms. As demonstrated here, direct stimulationof these same cells by application of adrenal hormonesthe amygdala may be acting at membrane, rather than
intracellular receptors. A-ring reduction of the steroids evokes a rapid arousal of sodium intake in the rat, whichbecause of the quick elicitation of behavior may be actingcan occur within the brain, and the enzymes, primarily
5 a and 3 b reductase, are present in the amygdala [5, 28]. through membrane receptors rather than cytosolic recep-tors.The TH steroids are thought to act by binding to GABAa/
benzodiazepine receptor complexes to evoke changes inchloride channels [29, 30]. It is possible then that the ACKNOWLEDGMENTSeffects seen here may be engaging a GABAa/benzodiaz- This work was supported by DK48061 (R.R.S.) and MH43787epine system, but in a steroid-specific manner [31], with (R.R.S., S.J.F., and B.M.).ALDO and DOCA and their metabolites being the most
Reprint requests to Randall R. Sakai, Ph.D., Department of Psychia-effective and CORT, TH-CORT and 3 b DHP being try, University of Cincinnati, College of Medicine, P.O. Box 670559,ineffective in evoking sodium intake. Cincinnati, Ohio 45267, USA.
E-mail: [email protected] involvement of a GABAa/benzodiazepine recep-tor system was supported by pharmacological studies,which demonstrated that flunitrazepam was equally ef- APPENDIXfective as TH-ALDO or TH-DOC in stimulating a rapid
Abbreviations used in this article are: ALDO, aldosterone; 3 alphasalt intake. Furthermore, the effects of TH-ALDO andOH DHP, alloprenanolone; ASDN, antisense oligodeoxynucleotides;
TH-DOC were inhibited by pretreating the amygdala CORT, corticosterone; DOCA, deoxycorticosterone acetate; GR, glu-cocorticoid receptors; ICV, intracerebroventricular; MR, mineralocor-with the reverse agonists Ro15-4513 or Ro15-1788. To-ticoid receptors; pICV, pulse intracerebroventricular; TH-ALDO,gether, these data suggest that GABA plays a role in3b,5b tetrahydroaldosterone; TH-CORT, 3b,5b tetrahydrocorticoste-
this rapid induction of sodium intake by these ring-A– rone; TH-DOC, 5 a-tetrahydrodeoxycorticosterone.reduced steroids. The data, however, do not preclude aGABAa/benzodiazepine receptor downstream from a REFERENCESmembrane receptor, although the most parsimonious ex-
1. Rice KK, Richter CP: Increased sodium chloride and water intakeplanation involves a steroid-sensitive GABAa receptor of normal rats treated with deoxycorticosterone acetate. Endocri-with unique steroid specificity. nology 33:106–115, 1943
2. Fregly MJ, Waters IW: Effects of mineralocorticoids on spontane-It is important to note that the methodology used hereous sodium chloride appetite of adrenalectomized rats. Physiolto arouse sodium intake in the rat involved primarily Behav 1:65–74, 1966
the adrenal steroids and most probably did not involve 3. Schulkin J: Behavior of sodium deficient rats: The search for asalty taste. J Comp Physiol Psychol 96:628–634, 1982the renin-angiotensin system. It is known that high circu-
4. Baulieu EE: Cell membrane, a target for steroid hormones. Mollating levels of ALDO will suppress plasma renin activity Cell Endocrinol 12:247–254, 1978such that the angiotensin system in these rats was proba- 5. Mellon SH, Deschepper CF: Neurosteroid biosynthesis: Genes
of adrenal steroidogenic enzymes are expressed in the brain. Brainbly working at its nadir [32, 33]. Thus, the appetite forRes 629:283–291, 1993sodium aroused by these treatments was due primarily 6. Morris DJ, Souness GW, Saccoccio NA: The effects of infusions
to the actions of the adrenal steroids acting alone. More- of ring-A-reduced derivatives of aldosterone on the antinatriureticand kaliuretic action of aldosterone. Steroids 53:21–26, 1988over, previous studies have shown that sodium appetite
7. Orchinik M, Murray TF, Moore FL: A corticosteroid receptoraroused by mineralocorticoids is not affected by agents in neuronal membranes. Science 252:1848–1851, 1991that interfere with the brain angiotensin system [26]. 8. Sakai RR, Ma LY, McEwen BS: Rapid arousal of sodium intake
by adrenal steroid implants in the amygdala of the rat brain. (sub-However, that is not to say that both systems work inde-mitted for publication)pendently in arousing sodium intake. During natural epi-
9. Wehling M, Christ M, Theisen K: Membrane receptors for aldo-sodes of sodium depletion, both the renin-angiotensin sterone: A novel pathway for mineralocorticoid action. Am J Phys-
iol 263:E974–E979, 1992system and the adrenal steroids ALDO and CORT10. de Kloet ER, Versteed DHG, Kovacs GL: Aldosterone blockswould be elevated and work in concert to arouse sodium
the response to corticosterone in the raphe-hippocampal serotoninintake. system. Brain Res 264:323–327, 1983
11. Sakai RR, Nicolaidis S, Epstein AN: Salt appetite is suppressedIn conclusion, in addition to the cytosolic receptorsby interference with angiotensin II and aldosterone. Am J Physiolthat bind mineralocorticoids, there may also be mem-251:R762–R768, 1986
brane receptors that also bind mineralocorticoid metabo- 12. Fluharty SJ, Epstein AN: Sodium appetite elicited by intracere-broventricular infusion of angiotensin II in the rat. II. Synergisticlites and that can evoke rapid changes in salt intake

Sakai et al: Genomic and nongenomic arousal of sodium intake 1345
interaction with systemic mineralocorticoids. Behav Neurosci receptor complementary DNA: Structural and functional kinshipwith the glucocorticoid receptor. Science 237:268–275, 198797:746–758, 1983
23. Ma LY, Itharat P, Fluharty SJ, Sakai RR: Intracerebroventricu-13. Janiak P, Brody MJ: Central interactions between aldosteronelar administration of mineralocorticoid receptor antisense oligonu-and vasopressin on cardiovascular system. Am J Physiol 255(1 Ptcleotides attenuates salt appetite in the rat. Stress 2:37–50, 19971):R166–R173, 1988
24. Schulkin J, Marini J, Epstein AN: A role for the medial region14. Sumners C, Gault TR, Fregly MJ: Potentiation of angiotensinof the amygdala in mineralocorticoid-induced salt hunger. BehavII-induced drinking by glucocorticoids is a specific glucocorticoidNeurosci 103:178–185, 1989type II receptor (GR)-mediated event. Brain Res 552:283–290,
25. Zhang DM, Epstein AN, Schulkin J: Medial region of the amyg-1991dala: Involvement in adrenal-steroid-induced salt appetite. Brain15. Blackburn RE, Demko AD, Hoffman GE, Stricker EM, Ver-Res 600:20–26, 1993balis JG: Central oxytocin inhibition of angiotensin-induced salt
26. Ma LY, McEwen BS, Sakai RR, Schulkin J: Glucocorticoidsappetite in rats. Am J Physiol 263:R1347–R1353, 1992facilitate mineralocorticoid-induced sodium intake in the rat. Horm16. Stricker E, Verbalis J: Central inhibitory control of sodium appe-Behav 27:240–250, 1993tite in rats: Correlation with pituitary oxytocin secretion. Behav
27. Wolf G, Handal PJ: Aldosterone induced sodium appetite: DoseNeurosci 101:560–567, 1987response and specificity. Endocrinology 78:1120–1124, 196617. Verbalis JG, Blackburn RE, Olson BR, Stricker EM: Central
28. Penning TM, Sharp RB, Krieger NR: Purification and propertiesoxytocin inhibition of food and salt ingestion: A mechanism forof 3 alpha hydroxysteroid dehydrogenase from rat brain cytosol.intake regulation of solute homeostasis. Regul Pept 45:149–154,J Biol Chem 260:15266–15272, 19851993
29. Gee KW: Steroid modulation of the Gaba benzodiazepine receptor18. Torreli V, Hardy M, Nedelec C, Tournemine C, Deraedt R,linked chloride ionophore. Mol Neurobiol 2:291–317, 1988Philibert D: 7-Alkyl spironolactones as potent aldosterone antag-
30. Gee KW, McCauley LD, Lan NC: A putative receptor for neurost-onist. J Steroid Biochem 17(Suppl III):17, 1982 eroids on the GABAA receptor complex: The pharmacological19. Galaverna O, DeLuca LA, Schulkin J, Yao SZ, Epstein AN: properties and therapeutic potential of epalons. Crit Rev NeurobiolDeficits in NaCl ingestion after damage to the central nucleus of 9:207–227, 1995the amygdala in the rat. Brain Res Bull 28:89–98, 1992 31. Lan NC, Chen JS, Belelli D, Pritchett DB, Seeburg PH, Gee
20. Nitabach MN, Schulkin J, Epstein AN: The medial amygdala is KW: A steroid recognition site is functionally coupled to an ex-part of a mineralocorticoid-sensitive circuit controlling NaCl intake pressed GABAA benzodiazepine receptor. Eur J Pharmacol Molin the rat. Behav Brain Res 35:127–134, 1989 Pharmacol 188:403–406, 1990
21. Ma LY, McEwen BS, Sakai RR: Effect of central infusion of 32. Ganong WF, Van Brunt EE, Lee TC, Mulrow PJ: Inhibition ofmineralocorticoid and/or glucocorticoid receptor antagonists on aldosterone-stimulating activity of hog and dog renin by the plasmaadrenal steroid-induced sodium appetite in the rat. (submitted for of dogs immunized with hog renin. Proc Soc Exp Biol (NY)publication) 112:1062–1067, 1963
22. Arriza JL, Weinberger C, Cerelli G, Glaser TM, Handelin BL, 33. Pettinger WA, Marchelle M, Augusto L: Renin suppression byDOC and NaCl in the rat. Am J Physiol 221:1071–1074, 1971Housman DE, Evans RM: Cloning of human mineralocorticoid