toward a broader understanding of aldosterone in congestive heart failure
TRANSCRIPT

http://jra.sagepub.com/Journal of Renin-Angiotensin-Aldosterone System
http://jra.sagepub.com/content/4/3/155The online version of this article can be found at:
DOI: 10.3317/jraas.2003.024
2003 4: 155Journal of Renin-Angiotensin-Aldosterone SystemKenneth J Warrington
Karl T Weber, Yao Sun, Linus A Wodi, Ahmad Munir, Eiman Jahangir, Robert A Ahokas, Ivan C Gerling, Arnold E Postlethwaite andToward a broader understanding of aldosterone in congestive heart failure
Published by:
http://www.sagepublications.com
On behalf of:
can be found at:Journal of Renin-Angiotensin-Aldosterone SystemAdditional services and information for
http://jra.sagepub.com/cgi/alertsEmail Alerts:
http://jra.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
http://jra.sagepub.com/content/4/3/155.refs.htmlCitations:
What is This?
- Sep 1, 2003Version of Record >>
by guest on September 18, 2014jra.sagepub.comDownloaded from by guest on September 18, 2014jra.sagepub.comDownloaded from

Journal ofthe Renin-Angiotensin-AldosteroneSystem(Including otherpeptidergic systems)
September 2003Volume 4Number 3
155
Toward a broader understanding of aldosterone in congestive heart failureKarl T Weber,* Yao Sun,* Linus A Wodi,* Ahmad Munir,* Eiman Jahangir,*Robert A Ahokas,† Ivan C Gerling,≠ Arnold E Postlethwaite,§ Kenneth J Warrington §
Keywords: aldosterone, magnesium,peripheral bloodmononuclearcells,oxi/nitrosativestress
Divisions of*CardiovascularDiseases,≠Endocrinology,§Connective TissueDiseases,Department ofMedicine, andDepartment of†Obstetrics &Gynecology,University of TennesseeHealth Science CenterMemphis, TNUSA
Correspondence to:Dr Karl T WeberDivision ofCardiovascularDiseases,University of TennesseeHealth Science Center,Rm. 353 DobbsResearch Institute,951 Court Avenue,Memphis, TN 38163,USATel: +1 901 448 5750Fax: +1 901 448 8084E-mail: [email protected]
Accepted for publication8th July 2003
JRAAS 2003;4:155–63
AbstractDiscovered some 50 years ago, aldosterone (ALDO)has come to be recognised as a mineralocorticoidhormone with well-known endocrine properties inepithelial cells that contribute to the pathophysiologyof congestive heart failure. This includes Na+
resorption at the expense of K+ excretion in classictarget tissues: kidneys, colon, sweat and salivaryglands. Though less well known, Mg2+ excretion islikewise enhanced by ALDO, while adrenal ALDOsecretion is regulated by extracellular Mg2+ ([Mg2+]o).An emerging body of information has and continuesto identify other endocrine actions of ALDO receptor-ligand binding. They include: promoting anefflux of cytosolic free Mg2+, or [Mg2+]i, in exchangefor Na+ in such non-epithelial cells as peripheral bloodmononuclear cells; its influence on endothelial cellfunction; and its central actions that involve regulation of cerebrospinal fluid composition produced by epithelial cells of the choroid plexus,activity of the hypothalamic paraventricular nucleusinvolved in Na+ appetite, Na+ and H2O excretion andsympathetic nerve activity, and the regulation of TNF-α production from central and/or peripheralsources. Extra-adrenal steroidogenesis and auto/paracrine properties of ALDO generated de novoin the cardiovasculature are now under investigationand preliminary findings suggest they contribute totissue repair. The past decade has witnessed a revivalof interest in this steroid molecule. In years to come,an even broader understanding of ALDO’s contribution to the pathophysiology of congestiveheart failure will undoubtedly emerge.
IntroductionAldosterone (ALDO) was discovered some 50years ago.1,2 Termed electrocortin, identification ofits 18-aldehyde steroid structure and adrenalorigin led to its being renamed aldosterone(ALDO). ALDO’s ability to alter Na+ uptake inexchange for K+ prompted Selye to refer to it as amineralocorticoid.3 During a decade of discoverythat followed its identification, sites of ALDO’saction and receptor-ligand binding were found toreside within epithelial cells of the kidney, colon,sweat and salivary glands.4,5 The importance ofthis circulating hormone in promoting salt andwater retention at each of these target tissues wasdemonstrated in such oedema-forming states ascongestive heart failure (CHF). Also confirmatoryof its importance in CHF was the efficacy of
spironolactone, an ALDO receptor antagonistintroduced into clinical practice 45 years ago, incounteracting Na+ retention and alleviatingoedema.4-8
A recent resurgence of interest in this steroidmolecule in CHF has been driven by several obser-vations.9 In brief, this included: a recognition ofALDO’s broader range of actions that extendedbeyond classic target tissues and which were asso-ciated with an adverse structural remodelling ofthe cardiovasculature that contributes to patho-physiologic expressions and progressive nature ofCHF;10 the inability of angiotensin-convertingenzyme (ACE) inhibition to provide for sustainedsuppression of plasma ALDO in patients withCHF,11,12 which indicated that the regulation of itssecretion by zona glomerulosa cells of the adrenalcortex is more complex than that governed byangiotensin II (Ang II) alone;and finally, the resultsof a controlled clinical trial (RandomisedALdactone Evaluation Study [RALES]), conductedin 19 countries on five continents in over 1,600patients with CHF, wherein spironolactone (vis-à-vis placebo), in combination with an ACE inhibitor(ACE-I) and loop diuretic, reduced the risk of all-cause and cardiac-related mortality and cardiovas-cular morbidity by 30%.13
Herein, less well recognised properties ofALDO, mediated by receptor-ligand binding, arebriefly reviewed. Each has the potential to con-tribute to the pathophysiology of CHF. Theyinclude its influence on: Mg2+ balance and the roleof extracellular Mg2+ [Mg2+]o in regulating adrenalALDO secretion; vascular remodelling andimmune cell activation; endothelial cell function;and the central nervous system. Finally, extra-adrenal steroidogenesis of ALDO by the cardiovas-culature is briefly considered. The interestedreader is referred elsewhere14 for a discussion ofnon-genomic, non-receptor-mediated actions ofALDO.
Aldosterone and Mg2+ balance The distribution of Mg2+ within body tissues is asfollows: 53% in bone, 27% in skeletal muscle and19% in other soft tissues, such as the heart; lessthan 1% of total body Mg2+ is present in blood; andcytosolic free Mg2+ [Mg2+]i, represents but 0.5–5%of total cellular Mg2+, with the remaining 80%bound to ATP and other phosphometabolitessequestered within such organelles as mitochondria
Heart failure and the renin-angiotensin-aldosterone system
Weber 155-163 7/11/03 12:15 Page 1
COPYRIG
HT JRA
AS LIM
ITED
REPR
ODUCTION PR
OHIBITE
D
by guest on September 18, 2014jra.sagepub.comDownloaded from
Journal ofthe Renin-Angiotensin-AldosteroneSystem(Including otherpeptidergic systems)
September 2003Volume 4Number 3
and endoplasmic reticulum.15 [Mg2+]i homeostasisis maintained by exchange with these intracellularstores, while [Mg2+]o is held within narrow limitsby Mg2+ efflux from tissue stores.
Epithelial cellsIn 1955, Mader and Iseri16 reported that theirpatient with adrenal adenoma had experiencedspontaneous episodes of hypomagnesaemia,together with enhanced Mg2+ excretion in urineand stool. Others17-24 would also note the presenceof hypomagnesaemia in patients with primaryaldosteronism (PAL), suggesting Mg2+ deficiencyaccompanies long-standing, ALDO-induced Mg2+
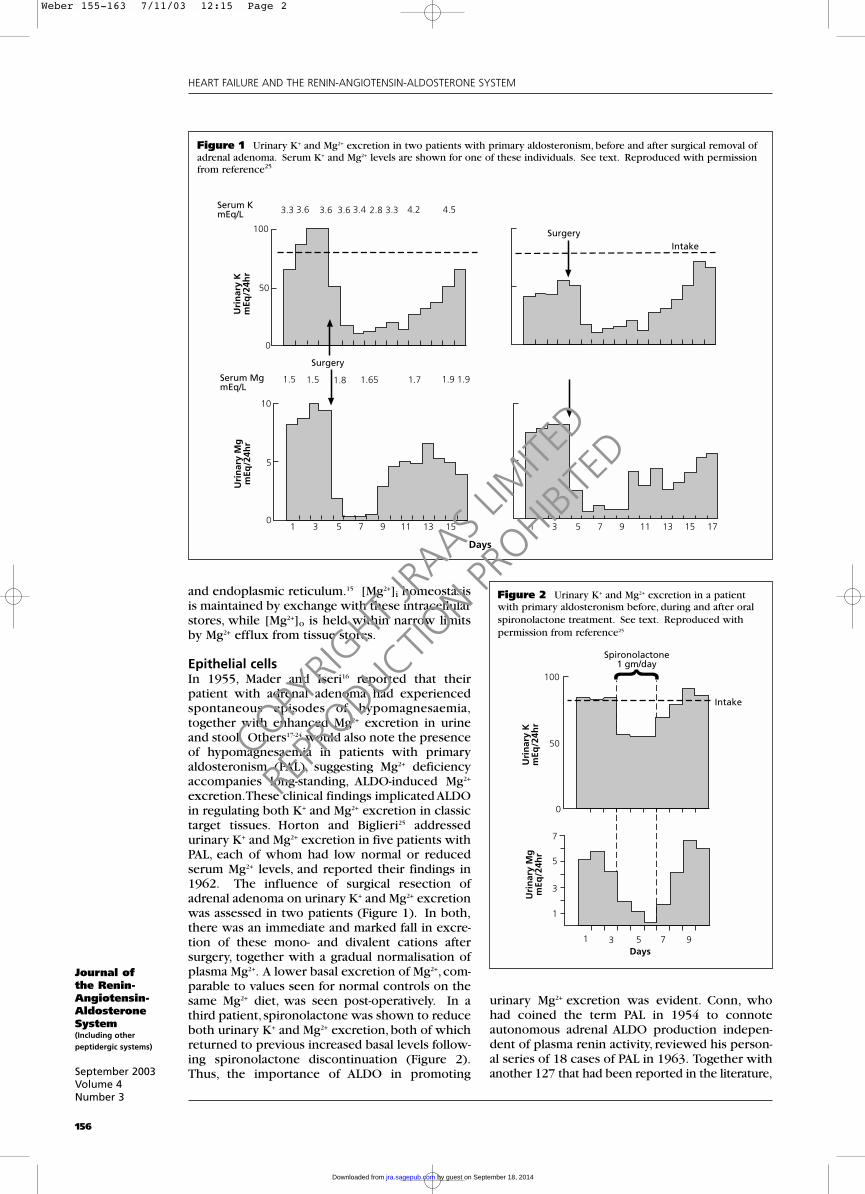
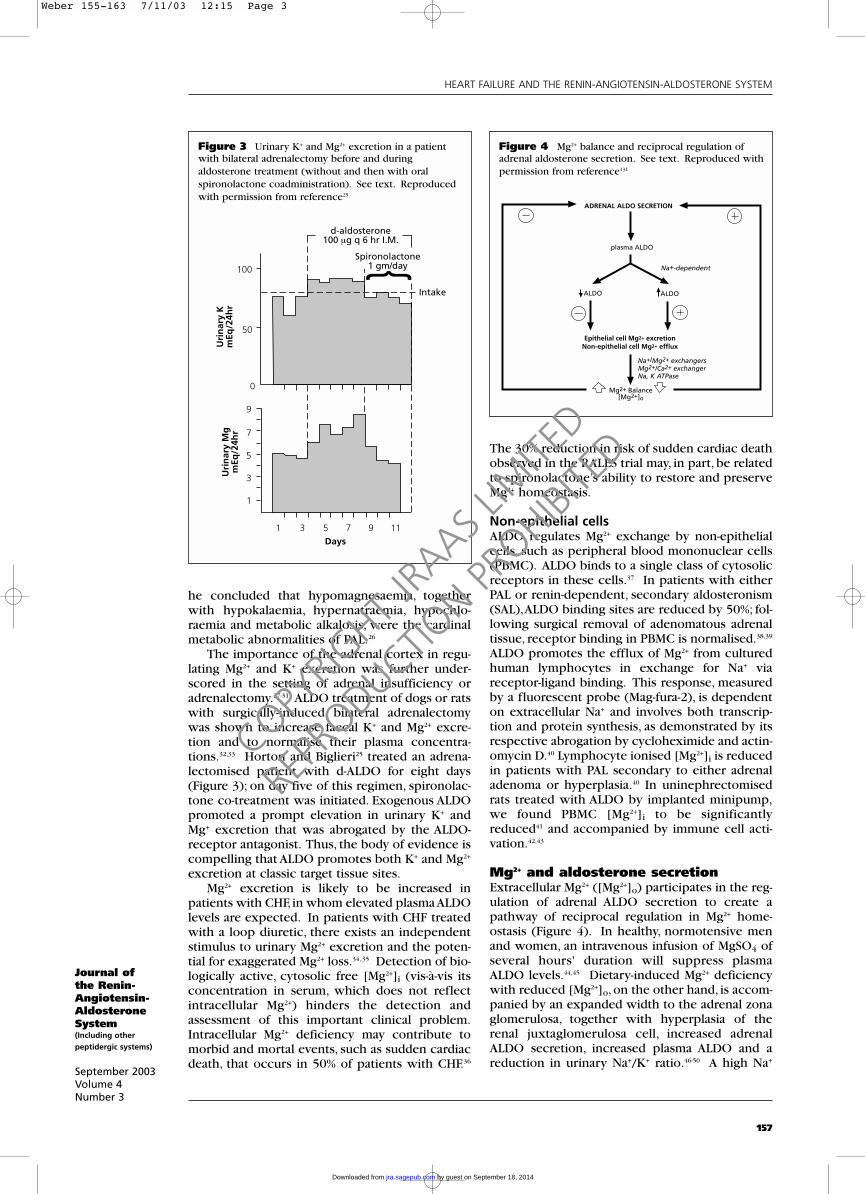
excretion.These clinical findings implicated ALDOin regulating both K+ and Mg2+ excretion in classictarget tissues. Horton and Biglieri25 addressedurinary K+ and Mg2+ excretion in five patients withPAL, each of whom had low normal or reducedserum Mg2+ levels, and reported their findings in1962. The influence of surgical resection ofadrenal adenoma on urinary K+ and Mg2+ excretionwas assessed in two patients (Figure 1). In both,there was an immediate and marked fall in excre-tion of these mono- and divalent cations aftersurgery, together with a gradual normalisation ofplasma Mg2+. A lower basal excretion of Mg2+, com-parable to values seen for normal controls on thesame Mg2+ diet, was seen post-operatively. In athird patient, spironolactone was shown to reduceboth urinary K+ and Mg2+ excretion, both of whichreturned to previous increased basal levels follow-ing spironolactone discontinuation (Figure 2).Thus, the importance of ALDO in promoting
urinary Mg2+ excretion was evident. Conn, whohad coined the term PAL in 1954 to connoteautonomous adrenal ALDO production indepen-dent of plasma renin activity, reviewed his person-al series of 18 cases of PAL in 1963. Together withanother 127 that had been reported in the literature,
156
HEART FAILURE AND THE RENIN-ANGIOTENSIN-ALDOSTERONE SYSTEM
Figure 1 Urinary K+ and Mg2+ excretion in two patients with primary aldosteronism, before and after surgical removal ofadrenal adenoma. Serum K+ and Mg2+ levels are shown for one of these individuals. See text. Reproduced with permissionfrom reference25
1 3 5 7 9 11 13 150
5
10
1.5 1.5 1.8 1.65 1.7 1.9 1.9
Surgery
Serum MgmEq/L
Uri
nar
y M
gm
Eq/2
4hr
Uri
nar
y K
mEq
/24h
r
Serum KmEq/L 3.3 3.6 3.6 3.6 3.4 2.8 3.3 4.2 4.5
100
50
0
SurgeryIntake
1 3 5 7 9 11 13 15 17
Figure 2 Urinary K+ and Mg2+ excretion in a patientwith primary aldosteronism before, during and after oralspironolactone treatment. See text. Reproduced with permission from reference25
100
50
0
7
5
3
1
1 3 5 7 9Days
Uri
nar
y M
gm
Eq/2
4hr
Uri
nar
y K
mEq
/24h
r
Spironolactone1 gm/day
Intake
Days
Weber 155-163 7/11/03 12:15 Page 2
COPYRIG
HT JRA
AS LIM
ITED
REPR
ODUCTION PR
OHIBITE
D
by guest on September 18, 2014jra.sagepub.comDownloaded from
Journal ofthe Renin-Angiotensin-AldosteroneSystem(Including otherpeptidergic systems)
September 2003Volume 4Number 3
157
he concluded that hypomagnesaemia, togetherwith hypokalaemia, hypernatraemia, hypochlo-raemia and metabolic alkalosis, were the cardinalmetabolic abnormalities of PAL.26
The importance of the adrenal cortex in regu-lating Mg2+ and K+ excretion was further under-scored in the setting of adrenal insufficiency oradrenalectomy.27-31 ALDO treatment of dogs or ratswith surgically-induced bilateral adrenalectomywas shown to increase faecal K+ and Mg2+ excre-tion and to normalise their plasma concentra-tions.32,33 Horton and Biglieri25 treated an adrena-lectomised patient with d-ALDO for eight days(Figure 3); on day five of this regimen, spironolac-tone co-treatment was initiated. Exogenous ALDOpromoted a prompt elevation in urinary K+ andMg+ excretion that was abrogated by the ALDO-receptor antagonist. Thus, the body of evidence iscompelling that ALDO promotes both K+ and Mg2+
excretion at classic target tissue sites.Mg2+ excretion is likely to be increased in
patients with CHF, in whom elevated plasma ALDOlevels are expected. In patients with CHF treatedwith a loop diuretic, there exists an independentstimulus to urinary Mg2+ excretion and the poten-tial for exaggerated Mg2+ loss.34, 35 Detection of bio-logically active, cytosolic free [Mg2+]i (vis-à-vis itsconcentration in serum, which does not reflectintracellular Mg2+) hinders the detection andassessment of this important clinical problem.Intracellular Mg2+ deficiency may contribute tomorbid and mortal events, such as sudden cardiacdeath, that occurs in 50% of patients with CHF.36
The 30% reduction in risk of sudden cardiac deathobserved in the RALES trial may, in part, be relatedto spironolactone’s ability to restore and preserveMg2+ homeostasis.
Non-epithelial cellsALDO regulates Mg2+ exchange by non-epithelialcells, such as peripheral blood mononuclear cells(PBMC). ALDO binds to a single class of cytosolicreceptors in these cells.37 In patients with eitherPAL or renin-dependent, secondary aldosteronism(SAL),ALDO binding sites are reduced by 50%; fol-lowing surgical removal of adenomatous adrenaltissue, receptor binding in PBMC is normalised.38,39
ALDO promotes the efflux of Mg2+ from culturedhuman lymphocytes in exchange for Na+ viareceptor-ligand binding. This response, measuredby a fluorescent probe (Mag-fura-2), is dependenton extracellular Na+ and involves both transcrip-tion and protein synthesis, as demonstrated by itsrespective abrogation by cycloheximide and actin-omycin D.40 Lymphocyte ionised [Mg2+]i is reducedin patients with PAL secondary to either adrenaladenoma or hyperplasia.40 In uninephrectomisedrats treated with ALDO by implanted minipump,we found PBMC [Mg2+]i to be significantlyreduced41 and accompanied by immune cell acti-vation.42,43
Mg2+ and aldosterone secretion Extracellular Mg2+ ([Mg2+]o) participates in the reg-ulation of adrenal ALDO secretion to create apathway of reciprocal regulation in Mg2+ home-ostasis (Figure 4). In healthy, normotensive menand women, an intravenous infusion of MgSO4 ofseveral hours' duration will suppress plasmaALDO levels.44,45 Dietary-induced Mg2+ deficiencywith reduced [Mg2+]o, on the other hand, is accom-panied by an expanded width to the adrenal zonaglomerulosa, together with hyperplasia of therenal juxtaglomerulosa cell, increased adrenalALDO secretion, increased plasma ALDO and areduction in urinary Na+/K+ ratio.46-50 A high Na+
HEART FAILURE AND THE RENIN-ANGIOTENSIN-ALDOSTERONE SYSTEM
Figure 3 Urinary K+ and Mg2+ excretion in a patientwith bilateral adrenalectomy before and during aldosterone treatment (without and then with oralspironolactone coadministration). See text. Reproducedwith permission from reference25
100
50
0
9
7
5
3
1
1 3 5 7 9 11Days
Uri
nar
y M
gm
Eq/2
4hr
Uri
nar
y K
mEq
/24h
rd-aldosterone
100 µg q 6 hr I.M.
Intake
Spironolactone1 gm/day
Figure 4 Mg2+ balance and reciprocal regulation ofadrenal aldosterone secretion. See text. Reproduced withpermission from reference131
ADRENAL ALDO SECRETION
plasma ALDO
Na+-dependent
ALDOALDO
Epithelial cell Mg2+ excretionNon-epithelial cell Mg2+ efflux
Na+/Mg2+ exchangersMg2+/Ca2+ exchangerNa, K ATPase
Mg2+ Balance[Mg2+]o
Weber 155-163 7/11/03 12:15 Page 3
COPYRIG
HT JRA
AS LIM
ITED
REPR
ODUCTION PR
OHIBITE
D
by guest on September 18, 2014jra.sagepub.comDownloaded from

Journal ofthe Renin-Angiotensin-AldosteroneSystem(Including otherpeptidergic systems)
September 2003Volume 4Number 3
158
diet, combined with dietary Mg2+ deficiency, atten-uates, but does not abrogate, heightened ALDOsecretion.48 In cultured zona glomerulosa cells,superfusate [Mg2+]o regulates ALDO production:high [Mg2+]o suppresses,while a [Mg2+]o free mediaaugments, their elaboration of ALDO.51 The SALthat accompanies dietary Mg2+ deficiency is asso-ciated with a time-dependent rise in [Na+]i and[Ca2+]i in heart, skeletal muscle, kidney and bone,which is suggestive of an inhibition of Na,K ATPase, a Mg2+-dependent pump, and increasedNa+/Ca2+ exchange at these sites.52-54
Vascular remodelling and immunecell activationA structural remodelling of the cardiovasculatureby fibrous tissue accompanies aldosteronismderived from either endogenous or exogenoussources.55-63 This fibrogenic phenotype includesintramural arteries of the heart, kidney, pancreas,mesentery and vasa vasorum of aorta and pul-monary artery.Co-treatment with a receptor antag-onist (e.g., spironolactone, eplerenone), in eithernon-depressor or depressor doses, prevents thisremodelling, indicating its independence of eleva-tions in blood pressure (BP).58,61-69 In a substudy tothe RALES trial, survival benefit was associatedwith a reduction in circulating markers of collagensynthesis that presumably reflected an attenuationin ongoing vascular fibrosis.70 In this connection,urinary excretion of hydroxyproline, a marker ofcollagen turnover, is increased in adrenalec-tomised rats treated with ALDO, 1% dietary NaCl,and cortisone.70 Glucocorticoids, on the otherhand, are known to reduce urinary hydroxypro-line excretion and their inhibition of collagen for-mation in bone is associated with osteoporosis.71
Campbell et al.60 and Nicoletti and co-workers61 each found that the perivascular fibrosisof the coronary vasculature that ultimatelyappears in aldosteronism is preceded by a pro-inflammatory vascular phenotype that featuresinvading monocytes/macrophages and lympho-cytes and adhesion molecule expression. Thesefindings, including their independence of BP, weremore recently confirmed by others.63,68,69 An inter-rogation of molecular responses involved in theinvasion of coronary vessels by these inflammato-ry cells were recently addressed in animal modelsof PAL and SAL.62,72 In rats receiving ALDO/salttreatment (ALDOST),where plasma renin and AngII are each suppressed, Sun et al.72 tested thehypothesis that oxidative stress was involved inthe appearance of the pro-inflammatory/fibro-genic cardiac phenotype. At week 3 of ALDOST,there was no evidence of cardiac pathology; atweeks 4 and 5, however, inflammatory cells(monocytes/macrophages and lymphocytes) werefound to have invaded intramural coronary vesselsin both ventricles. Using immunohistochemistry,invading PBMC were found to express the gp91phox
subunit of NADPH oxidase, whose activation is amajor source of superoxide in leukocytes, and 3-nitrotyrosine, a product of peroxynitrite withstable protein tyrosine residues and where perox-
ynitrite is formed by the reaction between super-oxide and nitric oxide (NO). The RelA subunit ofNFκB, a redox-sensitive transcription factor inte-gral to inflammatory responses, was likewise acti-vated in these cells. In situ hybridisation localisedincreased mRNA expression of intercellular adhe-sion molecule (ICAM)-1, monocyte chemoattrac-tant protein (MCP)-1, and a pro-inflammatorycytokine tumour necrosis factor (TNF)-α at vascu-lar sites that involved the normotensive, non-hypertrophied right atrium and ventricle and leftatrium, as well as the hypertensive, hypertrophiedleft ventricle. Co-treatment with either spirono-lactone or an antioxidant (either pyrrolidinedithiocarbamate or N-acetylcysteine) preventedthe appearance of these cells and associated mol-ecular responses, as well as the subsequentperivascular fibrosis. Thus, ALDOST inducesoxi/nitrosative stress within inflammatory cellsinvading the intramural coronary vasculature andit is this pro-inflammatory vascular phenotype thatleads to intramural coronary artery pathology andthe subsequent perivascular fibrosis. Why shouldthis be the case? Did this only occur at vascularsites or was there a prior activation of PMBC?
We recently identified an ALDO-mediatedreduction in [Mg2+]i that appears in PBMC at week1 of ALDOST,43 long before the appearance ofcardiac pathology at week 4. The early activationof these cells is evident in their transcriptome(expressed genes) and proteome (expressed pro-teins).42,43 The interrogation of complex molecularevents that account for immune cell activation andsubsequent homing of these cells to the heart,mayexplain why cardiac pathology does not appearuntil week 4 of ALDOST and calls into questionthe prospect of an autoimmune response, notunlike that which can follow myocardial infarction(MI).73
In rodents treated with a Mg2+-deficient diet, aputative state of exaggerated aldosteronism (seeabove), lymphocyte Mg2+ is reduced to an extentcomparable to the Mg2+ depletion that appears inskeletal muscle and cardiac tissue.74 Weglicki andco-workers have identified an early (week 1)induction of oxi/nitrosative stress and depletionof antioxidant defences in PBMC and endothelialcells.75-78 The lymphocyte activation featured hereincludes their production of proinflammatorycytokines and a neurogenic peptide, substance P,together with the expression of its receptors.79
Cardiac lesions first appear in this model duringweek 3 and can be prevented by a substance Preceptor antagonist.79
The potential for the SAL that accompanieshuman CHF to likewise be accompanied byimmune cell activation remains unexplored.Nonetheless, the prospect exists that the nowrecognised ‘cytokine storm’ of CHF, which featureselevations in such circulating proinflammatorycytokines as TNF-α and IL-6, has an immune cellorigin. Cells of the monocyte-phagocyte systemare a potent source of these cytokines. An alter-native source of cytokine production in heartfailure is the central nervous system.80
HEART FAILURE AND THE RENIN-ANGIOTENSIN-ALDOSTERONE SYSTEM
Weber 155-163 7/11/03 12:15 Page 4
COPYRIG
HT JRA
AS LIM
ITED
REPR
ODUCTION PR
OHIBITE
D
by guest on September 18, 2014jra.sagepub.comDownloaded from

Journal ofthe Renin-Angiotensin-AldosteroneSystem(Including otherpeptidergic systems)
September 2003Volume 4Number 3
159
HEART FAILURE AND THE RENIN-ANGIOTENSIN-ALDOSTERONE SYSTEM
Irrespective of their origin, prolonged elevationsin these proinflammatory cytokines contribute tothe progressive systemic illness that accompaniesCHF and which features tissue wasting to eventu-ate in cardiac cachexia.81
Endothelial cell dysfunctionIn patients with PAL or renal artery stenosis withSAL, forearm vasomotor reactivity to endothelialcell-dependent acetylcholine is diminished com-pared with normotensive controls, while non-endothelial cell-dependent sodium nitroprusside-induced vasodilatation is preserved.82 Four weeksafter surgical removal of adrenal adenoma, theimpairment in endothelial cell-dependent vasodi-lation is restored. In the SAL that accompaniesCHF, Farquharson and Struthers83 found dimin-ished forearm vasomotor reactivity to acetyl-choline to be normalised after one month ofspironolactone treatment. Acetylcholine-induced,NO-dependent vasorelaxation of aortic rings isreduced in rats following MI.84 This vasomotordysfunction, together with increased superoxideformation by aortic tissue, is normalised byspironolactone, either alone or in combinationwith an ACE-I.84 In cultured aortic endothelialcells, reduced Mg2+ concentration of culturemedium is associated with increased oxidant pro-duction and reduced intracellular glutathione, anantioxidant reserve consumed in neutralisingoxi/nitrosative stress.75 Abnormal Mg2+ homeosta-sis could represent the underlying pathophysio-logical basis for endothelial dysfunction seen ineither PAL or SAL.
Central actions of aldosterone Additional sites of action of ALDO that fall outsideof its classic target tissues include the centralnervous system, where ALDO receptors are foundat diverse sites, including epithelial cells of thechoroid plexus. In this connection, a role forALDO in the genesis of idiopathic intracranialhypertension (IIH) has been proposed, given thereported association between IIH with PAL andSAL85,86 and the prevalence of headache amongpatients with PAL.26 That this proposition does notapply to all patients with IIH is underscored by itsappearance in patients with adrenal insufficiency,unless ALDO would be produced in situ withinthe central nervous system87 as is now recognisedto be the case for the cardiovasculature.88,89
The choroid plexus,a site of high-affinity ALDOreceptor binding, is involved in the production ofcerebrospinal fluid (CSF) and is a target site forALDO, spironolactone and ouabain, an endoge-nous digitalis-like substance released by theadrenals and the hypothalamic-pituitary axis.90-99
ALDO exerts its biologic actions on epithelial cellsby enhancing the activity and number of Na, KATPase pumps in their apical membrane. Aouabain-sensitive Na, K ATPase is present in themicrovilli of the plexus and is involved in the reg-ulation of CSF formation and electrolyte composi-tion (e.g., ouabain reduces CSF production).ALDO is present in CSF, where its concentration
correlates with plasma levels. Either the systemicor intracerebroventricular administration of a min-eralocorticoid (ALDO or DOC) is accompanied bya fall in CSF [K+], together with a rise in arterialpressure, without changes in blood volume,cardiac output, plasma catecholamines or vaso-pressin.99,100 This hypertensive response is abro-gated by intracerebroventricular infusion of K+ ora mineralocorticoid receptor antagonist.99,101
ALDO, therefore, has a central action involved inthe regulation of BP, as well as CSF volume andcomposition. Produced locally within the brain,ALDO’s paracrine properties may likewise con-tribute to BP regulation.87
Research by Felder et al.102-104 has broadenedthe perspective of ALDO’s central actions, whichmay contribute to the pathophysiology of CHF.The hypothalamic paraventricular nucleus (PVN),a forebrain site involved in the regulation of extra-cellular volume and sympathetic nerve activity, isgoverned by circulating neurohormones and byeffector signals originating from the brainstem. Inrats with MI induced by coronary artery ligation,the activity of the PVN is increased. Systemic orintracerebroventricular administration of spirono-lactone reduces this activity, improves baroreflexregulation of renal sympathetic nerve activity(albeit in a time-dependent manner) and preventsthe increase in Na+ appetite and decline in urinaryNa+ and H2O excretion that appear in thismodel.102,103 Plasma levels of TNF-α rise progres-sively over weeks 1–3 following MI, a responseabrogated by intracerebroventricular infusion ofspironolactone started 24 hours after coronary lig-ation, suggesting that central ALDO receptor acti-vation is involved in regulating the release of thispro-inflammatory cytokine.104 The cellular sourceof TNF-α, however, remains uncertain and mayinclude central and/or peripheral tissues.
Extra-adrenal aldosterone productionand tissue repairSome 40 years ago,Lockett et al.105-110 reported thatthe beating heart and contracting soleus muscle ofcats were sites of steroid generation; the substanceresembled 18-D-aldosterone.They found this mol-ecule in coronary venous blood and demonstratedits ability to promote renal salt and water reten-tion. These findings lay fallow until the pastdecade, when Takeda111,112 identified the mRNAexpression of ALDO synthase (CYP11B2) andALDO production in rodent vascular tissue.Silvestre and coworkers113 likewise demonstratedthe expression of this enzyme, integral to thebiosynthesis of ALDO, in rodent heart, whereALDO generation was regulated by Ang II, a lowNa+ or high K+ diet, or adrenocorticotropin(ACTH). Subsequently, the expression of thisenzyme was identified in human cardiovasculartissue, where it was localised to vascular smoothmuscle and endothelial cells.112,114 Even morerecently, and in contrast to normal human hearts,ALDO production was reported to be increased inthe failing human left ventricle,based on coronarysinus levels of ALDO that exceed those found in
Weber 155-163 7/11/03 12:15 Page 5
COPYRIG
HT JRA
AS LIM
ITED
REPR
ODUCTION PR
OHIBITE
D
by guest on September 18, 2014jra.sagepub.comDownloaded from

Journal ofthe Renin-Angiotensin-AldosteroneSystem(Including otherpeptidergic systems)
September 2003Volume 4Number 3
160
HEART FAILURE AND THE RENIN-ANGIOTENSIN-ALDOSTERONE SYSTEM
the aorta.115 There exists an upregulated expres-sion of CYP11B2 in the left ventricle of the failinghuman heart of diverse aetiological origins.116
Additionally, 11β-hydroxysteroid dehydrogenase,an enzyme critical to maintaining the specificity ofthe mineralocorticoid receptor, given its equalaffinity for mineralo- and glucocorticoids, hasbeen found in human cardiac tissue.117
Following MI in rodents, macrophages andmyofibroblasts involved in tissue repair expressrenin,ACE and Ang II receptors, predominantly ofthe AT1-subtype.118-122 This has implicated locally-produced Ang II at sites of injury in regulating col-lagen turnover, which has been further suggestedby the cardioprotective actions of losartan, an AT1-receptor antagonist, in attenuating fibrous tissueformation at, and remote to, the site MI.123 Locally-produced Ang II is also involved in regulating denovo ALDO production in the infarcted heart,124
which likewise may contribute to tissue repair.Increased expression of ALDO synthase and ALDOtissue levels, together with increased concentra-tions of Ang II, have been observed in non-infarct-ed rat myocardium following coronary artery liga-tion.124,125 Treatment with losartan prevented theseresponses related to de novo ALDO production.Either losartan or spironolactone treatment pre-vented the accompanying accumulation of colla-gen at sites remote to the MI, suggesting theinvolvement of Ang II-driven local ALDO produc-tion in regulating tissue repair. Hayashi et al.126
have reported that ALDO is extracted by the heartfollowing MI, and the transcardiac ALDO gradient(between aorta and coronary sinus) is correlatedwith coronary venous effluent levels of a serolog-ical marker of collagen turnover (procollagen typeIII aminoterminal peptide, PIIINP) that has beenassociated with LV dilatation and poor functionand prognosis. ALDO is extracted by the chroni-cally failing human heart of diverse aetiologicalorigins, a response blocked by spironolactone.127
Under circumstances in which circulating ALDO isnot increased, spironolactone attenuates neointi-mal thickening following vascular barotrauma,128
tissue repair at sites of fibrous tissue formation129
and vascular injury in stroke-prone rats.130 Theseobservations further suggest that auto/paracrineproperties of locally-produced ALDO participatein tissue repair.
Future directionsSince its discovery some 50 years ago, ALDO haswell-established importance in clinical medicine,including its role in CHF. The past decade has wit-nessed a resurgence of interest in this theadrenal’s most potent mineralocorticoid, as has itsde novo production through steroidogenesiswithin the cardiovasculature and brain. An ever-expanding role for this steroid molecule in themetabolism of mono- and divalent cations byepithelial and non-epithelial cells has warrantedan even broader perspective of its portfolio ofactions. The many peripheral and central actionsof ALDO that can contribute to the pathophysiol-ogy of the CHF syndrome remain to be defined.
This is no more evident than in the adverse struc-tural remodelling of the heart and systemic organsthat accompanies chronic elevations in plasmaALDO (inappropriate relative to dietary Na+) andwhich may be secondary to PBMC activationinduced by [Mg2+]i depletion and transduced byoxi/nitrosative stress. Molecular mechanismsinvolved in immune cell responses remain to beelucidated. Today’s technologies will permit anassessment of ALDO’s role in altering the molecu-lar phenotype of these immune cells, specificallytheir transcriptome and proteome. Such insightsmay provide for the development of serologic bio-markers that address the risk, onset and progres-sion of vascular injury in CHF and could lead theway toward refined and even newer drug targets.
AcknowledgementThe editorial assistance of Richard A. Parkinson,MEd is appreciated.
References1. Tait SAS,Tait JF.The correspondence of S.A.S. Simpson andJ. F. Tait with T. Reichstein during their collaborative work onthe isolation and elucidation of the structure of electrocortin(later aldosterone). Steroids 1998;63:440-53.2. Tait JF,Tait SAS.A decade (and even more) of aldosteroneand other adrenal steroids. In: Biglieri EG, Melby JC (eds.).Endocrine Hypertension. New York: Raven Press; 1990;p. 5-27.3. Selye H.The general adaptation syndrome and the diseasesof adaptation. J Clin Endocrinol 1946;6:117-230.4. Davis JO. The physiology of congestive heart failure. In:Hamilton WF (ed.) Handbook of Physiology.Washington, DC:American Physiological Society, 1965; p. 2071-122.5. Davis JO. Mechanisms of salt and water retention in con-gestive heart failure. Am J Med 1960;29:486-506.6. Kagawa CM, Cella JA, Van Arman CG. Action of newsteroids in blocking effects of aldosterone and deoxycorticos-terone on salt. Science 1957;126:1015-16.7. Liddle GW.Aldosterone antagonists.AMA Arch Intern Med1958;102:998-1004.8. Coppage WS, Jr., Liddle GW. Mode of action and clinicalusefulness of aldosterone antagonists. Ann N Y Acad Sci 1960;88:815-21.9. Weber KT, John B. Hickam Lecture: Heart-hitting tales ofsalt and destruction. J Lab Clin Med 2000;136:7-13.10. Weber KT. Aldosterone in congestive heart failure. N EnglJ Med 2001;345:1689-97.11. Struthers AD.Aldosterone escape during angiotensin-con-verting enzyme inhibitor therapy in chronic heart failure. JCardiac Failure 1996;2:47-54.12. Jorde UP,Vittorio T, Katz SD, Colombo PC, Latif F, Le JemtelTH. Elevated plasma aldosterone levels despite complete inhi-bition of the vascular angiotensin-converting enzyme inchronic heart failure. Circulation 2002;106:1055-7.13. Pitt B, Zannad F, Remme WJ et al.The effect of spironolac-tone on morbidity and mortality in patients with severe heartfailure. Randomized Aldactone Evaluation Study Investigators.N Engl J Med 1999;341:709-17.14. Lösel RM, Feuring M, Falkenstein E, Wehling M.Nongenomic effects of aldosterone: cellular aspects and clini-cal implications. Steroids 2002;67:493-8.15. Swaminathan R. Disorders of magnesium metabolism. JInt Fed Clin Chem 1999;11(2):10-18.16. Mader IJ, Iseri LT. Spontaneous hypopotassemia, hypomag-nesemia, alkalosis and tetany due to hypersecretion of corti-costerone-like mineralocorticoid. Am J Med 1955;19:976-88.17. Milne MD, Muehrcke RC,Aird I. Primary aldosteronism. QJ Med 1957;26:317-35.18. Cope CL, Garcia-Llaurado J. The occurrence of electro-cortin in human urine. BMJ 1954;1(June 5):1290-4.19. Foye LV, Feightmeir TV.Adrenal cortical carcinoma produc-ing solely mineralocorticoid effect. Am J Med 1955;19:966-75.20. Hellem AJ. Primary aldosteronism. (Report of a case). ActaMed Scand 1956;155:271-4.21. Eales L, Linder GC. Primary aldosteronism. Some observa-tions on a case in a Cape Coloured woman. Q J Med 1956;25:539-55.
Weber 155-163 7/11/03 12:15 Page 6
COPYRIG
HT JRA
AS LIM
ITED
REPR
ODUCTION PR
OHIBITE
D
by guest on September 18, 2014jra.sagepub.comDownloaded from

Journal ofthe Renin-Angiotensin-AldosteroneSystem(Including otherpeptidergic systems)
September 2003Volume 4Number 3
161
HEART FAILURE AND THE RENIN-ANGIOTENSIN-ALDOSTERONE SYSTEM
22. van Buchem FSP, Doorenbos H, Elings HS. Primary aldos-teronism due to adrenocortical hyperplasia.Lancet 1956;2(Aug18):335-7.23. Campbell CH, Nicolaides N, Steinbeck AW.Adrenocorticaltumour with hypokalæmia and flaccid muscle paralysis. Lancet1956;2:553-5.24. Chalmers TM, FitzGerald MG, James AH, Scarborough H.Conn's syndrome with severe hypertension. Lancet 1956;1:127-32.25. Horton R, Biglieri EG. Effect of aldosterone on the metab-olism of magnesium. J Clin Endocrinol Metab 1962;22:1187-92.26. Conn JW.Aldosteronism in man. Some clinical and clima-tological aspects. Part II. JAMA 1963;183:871-8.27. Zwemer RL, Sullivan RC. Blood chemistry of adrenal insuf-ficiency in cats. Endocrinology 1934;18:97-106.28. Loeb RF.Effect of sodium chloride in treatment of patientswith Addison's disease.Proc Soc Exp Biol Med 1933;30:808-12.29. Loeb RF, Atchley DW, Benedict EM, Leland J. Electrolytebalance studies in adrenalectomized dogs with particular ref-erence to the excretion of sodium. J Exp Med 1933;57:775-92.30. Harrop GA, Soffer LJ, Ellsworth R,Trescher JH. Studies onthe suprarenal cortex. III. Plasma electrolytes and electrolyteexcretion during suprarenal insufficiency in the dog. J Exp Med1933;58:17-38.31. Conway EJ, Hingerty D. The influence of adrenalectomyon muscle constituents. Biochemistry 1946;40:561-8.32. DaVanzo JP, Crossfield HC, Swingle WW. Effect of variousadrenal steroids on plasma magnesium and the electrocardio-gram of adrenalectomized dogs. Endocrinology 1958;63:825-30.33. Hanna S, MacIntyre I. The influence of aldosterone onmagnesium metabolism. Lancet 1960;2(Aug 13):348-50.34. Wester PO, Dyckner T. Diuretic treatment and magnesiumlosses. Acta Med Scand Suppl 1981;647:145-52.35. Leier CV, Dei Cas L, Metra M. Clinical relevance and man-agement of the major electrolyte abnormalities in congestiveheart failure: hyponatremia, hypokalemia, and hypomagne-semia. Am Heart J 1994;128:564-74.36. Wilson JR. Use of antiarrhythmic drugs with heart failure:clinical efficacy, hemodynamic results, and relation to survival.Circulation 1987;75:IV64-IV73.37. Armanini D, Strasser T, Weber PC. Characterization ofaldosterone binding sites in circulating human mononuclearleukocytes. Am J Physiol 1985;248:E388-E390.38. Armanini D, Witzgall H, Wehling M, Kuhnle U, Weber PC.Aldosterone receptors in different types of primary hyperal-dosteronism. J Clin Endocrinol Metab 1987;65:101-04.39. Armanini D, Zennaro CM, Martella L, Pratesi C, Scali M,Zampollo V. Regulation of aldosterone receptors in hyperten-sion. Steroids 1993;58:611-13.40. Delva P, Pastori C, Degan M, Montesi G, Brazzarola P, LechiA. Intralymphocyte free magnesium in patients with primaryaldosteronism: aldosterone and lymphocyte magnesium home-ostasis. Hypertension 2000;35:113-17.41. Wodi LA,Ahokas RA, Sun Y,Weber KT. Oxidative stress andperipheral blood mononuclear cell free cytosolic magnesiumin rats treated with aldosterone/salt [abstract]. J Investig Med2003;51(suppl 1):S283.42. Munir A, Gerling IC, Sun Y, Ali C, Weber KT. Proteomicanalysis of peripheral blood mononuclear cell activation inaldosterone-treated rats [abstract]. J Investig Med 2003;51(suppl 1):S283.43. Gerling IC, Sun Y, Ahokas RA et al. Aldosteronism. Animmunostimulatory state precedes the proinflammatory/fibro-genic cardiac phenotype. Am J Physiol Heart Circ Physiol2003;285:H813-H821.44. Ichihara A, Suzuki H, Saruta T. Effects of magnesium on therenin-angiotensin-aldosterone system in human subjects. J LabClin Med 1993;122:432-40.45. Corica F, Allegra A, Ientile R et al. Effects of the intra-venous magnesium administration on aldosterone and atrialnatriuretic factor plasma concentrations. Nephron 1996;73:739-41.46. Cantin M. Relationship of juxtaglomerular apparatus andadrenal cortex to biochemical and extracellular fluid volumechanges in magnesium deficiency. Lab Invest 1970;22:558-68.47. Ginn HE, Cade R, McCallum T, Fregley M. Aldosteronesecretion in magnesium-deficient rats. Endocrinology1967;80:969-71.48. Solounias BM, Schwartz R.The effect of magnesium defi-ciency on serum aldosterone in rats fed two levels of sodium.Life Sci 1975;17:1211-17.49. Nadler JL, Buchanan T, Natarajan R,Antonipillai I, Bergman
R, Rude R. Magnesium deficiency produces insulin resistanceand increased thromboxane synthesis. Hypertension1993;21:1024-9.50. Laurant P, Dalle M, Berthelot A, Rayssiguier Y.Time-courseof the change in blood pressure level in magnesium-deficientWistar rats. Br J Nutr 1999;82:243-51.51. Atarashi K, Matsuoka H,Takagi M, Sugimoto T. Magnesiumion: a possible physiological regulator of aldosterone produc-tion. Life Sci 1989;44:1483-9.52. MacIntyre I, Davidsson D. The production of secondarypotassium depletion, sodium retention, nephrocalcinosis andhypercalcinemia by magnesium deficiency. Biochem J 1958;70:456-62.53. Madden JA, Smith GA, Llaurado JG. Myocardial K kineticsin rats on Mg-deficient diet. J Am Coll Nutr 1982;1:323-9.54. Chang C, Bloom S. Interrelationship of dietary Mg intakeand electrolyte homeostasis in hamsters: I. Severe Mg deficien-cy, electrolyte homeostasis, and myocardial necrosis. J Am CollNutr 1985;4:173-85.55. Hall CE, Hall O. Hypertension and hypersalimentation. I.Aldosterone hypertension. Lab Invest 1965;14:285-94.56. Brilla CG, Pick R,Tan LB, Janicki JS,Weber KT. Remodelingof the rat right and left ventricle in experimental hypertension.Circ Res 1990;67:1355-64.57. Sun Y, Ratajska A, Zhou G,Weber KT.Angiotensin convert-ing enzyme and myocardial fibrosis in the rat receivingangiotensin II or aldosterone. J Lab Clin Med 1993;122:395-403.58. Robert V,Van Thiem N, Cheav SL, Mouas C, SwynghedauwB,Delcayre C. Increased cardiac types I and III collagen mRNAsin aldosterone-salt hypertension. Hypertension 1994;24:30-6.59. Sun Y, Ramires FJA, Weber KT. Fibrosis of atria and greatvessels in response to angiotensin II or aldosterone infusion.Cardiovasc Res 1997;35:138-47.60. Campbell SE, Janicki JS,Weber KT.Temporal differences infibroblast proliferation and phenotype expression in responseto chronic administration of angiotensin II or aldosterone. JMol Cell Cardiol 1995;27:1545-60.61. Nicoletti A, Mandet C, Challah M, Barity J, Michel J-B.Mediators of perivascular inflammation in the left ventricle ofrenovascular hypertensive rats. Cardiovasc Res 1996;31:585-95.62. Fiebeler A, Schmidt F, Müller DN et al. Mineralocorticoidreceptor affects AP-1 and nuclear factor-kB activation inangiotensin II-induced cardiac injury. Hypertension 2001;37:787-93.63. Rocha R, Stier CT, Jr., Kifor I et al.Aldosterone: a mediatorof myocardial necrosis and renal arteriopathy. Endocrinology2000;141:3871-8.64. Selye H. Protection by a steroid-spironolactone againstcertain types of cardiac necroses. Proc Soc Exp Biol Med1960;104:212-13.65. Young M, Head G, Funder J. Determinants of cardiac fibro-sis in experimental hypermineralocorticoid states. Am JPhysiol 1995;269:E657-E662.66. Benetos A, Lacolley P, Safar ME. Prevention of aortic fibro-sis by spironolactone in spontaneously hypertensive rats.Arterioscler Thromb Vasc Biol 1997;17:1152-6.67. Lacolley P, Safar ME, Lucet B, Ledudal K, Labat C, BenetosA. Prevention of aortic and cardiac fibrosis by spironolactonein old normotensive rats. J Am Coll Cardiol 2001;37:662-7.68. Martinez DV, Rocha R, Matsumura M et al. Cardiac damageprevention by eplerenone: comparison with low sodium dietor potassium loading. Hypertension 2002;39:614-18.69. Rocha R, Rudolph AE, Frierdich GE et al. Aldosteroneinduces a vascular inflammatory phenotype in the rat heart.Am J Physiol 2002;283:H1802-H1810.70. Zannad F, Alla F, Dousset B, Perez A, Pitt B. Limitation ofexcessive extracellular matrix turnover may contribute to sur-vival benefit of spironolactone therapy in patients with con-gestive heart failure: insights from the randomized aldactoneevaluation study (RALES). Rales Investigators. Circulation2000;102:2700-06.71. Thompson JS, Palmieri GM, Eliel LP, Crawford RL. Theeffect of porcine calcitonin on osteoporosis induced byadrenal cortical steroids. J Bone Joint Surg Am 1972;54:1490-500.72. Sun Y, Zhang J, Lu L, Chen SS, Quinn MT, Weber KT.Aldosterone-induced inflammation in the rat heart. Role ofoxidative stress. Am J Pathol 2002;161:1773-81.73. Maisel A,Cesario D,Baird S,Rehman J,Haghighi P,Carter S.Experimental autoimmune myocarditis produced by adoptivetransfer of splenocytes after myocardial infarction. Circ Res1998;82:458-63.
Weber 155-163 7/11/03 12:15 Page 7
COPYRIG
HT JRA
AS LIM
ITED
REPR
ODUCTION PR
OHIBITE
D
by guest on September 18, 2014jra.sagepub.comDownloaded from

Journal ofthe Renin-Angiotensin-AldosteroneSystem(Including otherpeptidergic systems)
September 2003Volume 4Number 3
162
HEART FAILURE AND THE RENIN-ANGIOTENSIN-ALDOSTERONE SYSTEM
74. Ryan MF, Ryan MP. Lymphocyte electrolyte alterationsduring magnesium deficiency in the rat. Ir J Med Sci 1979;148:108-09.75. Wiles ME,Wagner TL,Weglicki WB. Effect of acute magne-sium deficiency (MgD) on aortic endothelial cell (EC) oxidantproduction. Life Sci 1997;60:221-36.76. Weglicki WB, Mak IT, Stafford RE, Dickens BF, Cassidy MM,Phillips TM. Neurogenic peptides and the cardiomyopathy ofmagnesium-deficiency: effects of substance P-receptor inhibi-tion. Mol Cell Biochem 1994;130:103-09.77. Weglicki WB, Dickens BF, Wagner TL, Chmielinska JJ,Phillips TM. Immunoregulation by neuropeptides in magne-sium deficiency: ex vivo effect of enhanced substance P pro-duction on circulating T lymphocytes from magnesium-defi-cient mice. Magnes Res 1996;9:3-11.78. Mak IT, Komarov AM,Wagner TL, Stafford RE, Dickens BF,Weglicki WB. Enhanced NO production during Mg deficiencyand its role in mediating red blood cell glutathione loss. Am JPhysiol 1996;271:C385-C390.79. Weglicki WB, Mak IT, Phillips TM. Blockade of cardiacinflammation in Mg2+ deficiency by substance P receptor inhi-bition. Circ Res 1994;74:1009-13.80. Felder RB, Francis J, Zhang ZH,Wei SG,Weiss RM, JohnsonAK.Heart failure and the brain:new perspectives.Am J Physiol2003;284:R259-R276.81. Anker SD, Chua TP, Ponikowski P et al. Hormonal changesand catabolic/anabolic imbalance in chronic heart failure andtheir importance for cardiac cachexia. Circulation 1997;96:526-34.82. Taddei S,Virdis A, Mattei P, Salvetti A.Vasodilation to acetyl-choline in primary and secondary forms of human hyperten-sion. Hypertension 1993;21:929-33.83. Farquharson CA, Struthers AD. Spironolactone increasesnitric oxide bioactivity, improves endothelial vasodilator dys-function, and suppresses vascular angiotensin I/angiotensin IIconversion in patients with chronic heart failure. Circulation2000;101:594-7.84. Bauersachs J, Heck M, Fraccarollo D et al. Addition ofspironolactone to angiotensin-converting enzyme inhibition inheart failure improves endothelial vasomotor dysfunction: roleof vascular superoxide anion formation and endothelial nitricoxide synthase expression. J Am Coll Cardiol 2002;39:351-8.85. Weber KT,Singh KD,Hey JC. Idiopathic intracranial hyper-tension with primary aldosteronism. Report of two cases. Am JMed Sci 2002;324:45-50.86. Mendonça E, Albuquerque L, Jácombe L, Luís MLS, RosaFC. Pseudotumor cerebri in a patient with Bartter's syndromeand obesity. Nephron 1996;72:98-9.87. Gomez-Sanchez CE, Zhou MY, Cozza EN, Morita H,Foecking MF, Gomez-Sanchez EP. Aldosterone biosynthesis inthe rat brain. Endocrinology 1997;138:3369-73.88. Slight SH, Joseph J, Ganjam VK, Weber KT. Extra-adrenalmineralocorticoids and cardiovascular tissue. J Mol CellCardiol 1999;31:1175-84.89. Delcayre C, Swynghedauw B. Molecular mechanisms ofmyocardial remodeling. The role of aldosterone. J Mol CellCardiol 2002;34:1577-84.90. Birmingham MK, Sar M, Stumpf WE. Localization of aldos-terone and corticostorone in the central nervous system,assessed by quantitative autoradiography. Neurochem Res1984;9:333-50.91. Wright EM. Transport processes in the formation of thecerebrospinal fluid.Rev Physiol Biochem Pharmacol 1978;83:1-34.92. Vates TS, Jr., Bonting SL, Oppelt WW. Na-K activated adeno-sine triphosphatase formation of cerebrospinal fluid in the cat.Am J Physiol 1964;206:1165-72.93. Johanson CE, Reed DJ,Woodbury DM.Active transport ofsodium and potassium by the choroid plexus of the rat. JPhysiol 1974;241:359-72.94. Loffreda N, Eldin P, Auzou G, Frelin C, Claire M.Corticosteroid receptors in cells derived from rat brainmicrovessels: mRNA identification and aldosterone binding.Am J Physiol 1992;262:C156-C163.95. Ames A, III, Higashi K, Nesbett FB. Effects of PCO2, aceta-zolamide and ouabain on volume and composition of choroid-plexus fluid. J Physiol 1965;181:516-24.96. Davson H, Segal MB. The effects of some inhibitors andaccelerators of sodium transport on the turnover of 22Na in thecerebrospinal fluid and the brain. J Physiol 1970;209:131-53.97. Kageyama Y, Suzuki H, Saruta T. Presence of aldosterone-like immunoreactivity in cerebrospinal fluid in normotensivesubjects. Acta Endocrinol 1992;126:501-04.98. Masuzawa T, Saito T, Sato F. Cytochemical study on enzyme
activity asociated with cerebrospinal fluid secretion in thechoroid plexus and ventricular ependyma. Brain Res 1981;222:309-22.99. Klarr SA, Ulanski LJ, II, Stummer W, Xiang J, Betz AL, KeepRF. The effects of hypo- and hyperkalemia on choroid plexuspotassium transport. Brain Res 1997;758:39-44.100. Kageyama Y, Bravo EL. Hypertensive mechanisms associat-ed with centrally administered aldosterone in dogs.Hypertension 1988;11:750-3.101. Gómez-Sánchez EP. Intracerebroventricular infusion ofaldosterone induces hypertension in rats. Endocrinology1986;118:819-23.102. Zhang ZH, Francis J, Weiss RM, Felder RB. The renin-angiotensin-aldosterone system excites hypothalamic paraven-tricular nucleus neurons in heart failure. Am J Physiol2002;283:H423-H433.103. Francis J,Weiss RM,Wei SG et al.Central mineralocorticoidreceptor blockade improves volume regulation and reducessympathetic drive in heart failure. Am J Physiol 2001;281:H2241-H2251.104. Francis J,Weiss RM, Johnson AK, Felder RB. Central miner-alocorticoid receptor blockade decreases plasma TNF-α aftercoronary artery ligation in rats. Am J Physiol 2003;284:R328-R335.105. Lockett MF.Hormonal actions of the heart and of lungs onthe isolated kidney. J Physiol 1967;193:661-9.106. Lockett MF,Retallack RW.The isolation of a substance veryclosely resembling the 18-monoacetate of D-aldosterone fromthe venous blood of activated muscle and from contractingmuscle. J Physiol 1969;204:435-42.107. Lockett MF. Changes in the flow and composition of theurine induced by the 18 monoacetate of D-aldosterone, in cats.J Physiol 1969;202:671-82.108. Lockett MF, Retallack RW. The influence of heart rate onthe secretion of a substance closely resembling the 18-monoac-etate of D-aldosterone by the hearts of cats under chloraloseanaesthesia. J Physiol 1970;208:21-32.109. Lockett MF,Retallack RW.Factors controlling the secretionof a substance biologically resembling the 18-monoacetate ofD-aldosterone by heart muscle. J Physiol 1970;210:717-25.110. Lockett MF, Retallack RW.The isolation of a renally activesubstance from arterial blood. J Physiol 1972;223:49-57.111. Takeda Y, Miyamori I, Yoneda T et al. Production of aldos-terone in isolated rat blood vessels. Hypertension 1995;25:170-3.112. Takeda Y, Miyamori I,Yoneda T et al. Regulation of aldos-terone synthase in human vascular endothelial cells byangiotensin II and adrenocorticotropin. J Clin EndocrinolMetab 1996;81:2797-800.113. Silvestre J-S, Robert V, Heymes C et al. Myocardial produc-tion of aldosterone and corticosterone in the rat. J Biol Chem1998;273:4883-91.114. Hatakeyama H, Miyamori I, Fujita T, Takeda Y, Takeda R,Yamamoto H. Vascular aldosterone. Biosynthesis and a link toangiotensin II-induced hypertrophy of vascular smooth musclecells. J Biol Chem 1994;269:24316-20.115. Mizuno Y, Yoshimura M, Yasue H et al. Aldosterone pro-duction is activated in failing ventricle in humans. Circulation2001;103:72-7.116. Yoshimura M, Nakamura S, Ito T et al. Expression of aldos-terone synthase gene in failing human heart: quantitative analy-sis using modified real-time polymerase chain reaction. J ClinEndocrinol Metab 2002;87:3936-40.117. Slight SH, Ganjam VK, Gómez-Sánchez CE, Zhou M-Y,Weber KT. High affinity NAD+-dependent 11β-hydroxysteroiddehydrogenase in the human heart. J Mol Cell Cardiol1996;28:781-7.118. Sun Y, Cleutjens JPM, Diaz-Arias AA, Weber KT. Cardiacangiotensin converting enzyme and myocardial fibrosis in therat. Cardiovasc Res 1994;28:1423-32.119. Sun Y,Weber KT.Angiotensin II receptor binding followingmyocardial infarction in the rat. Cardiovasc Res 1994;28:1623-8.120. Sun Y,Weber KT. Cells expressing angiotensin II receptorsin fibrous tissue of rat heart. Cardiovasc Res 1996;31:518-25.121. Passier RC, Smits JF,Verluyten MJ, Daemen MJ. Expressionand localization of renin and angiotensinogen in rat heart aftermyocardial infarction. Am J Physiol 1996;271:H1040-H1048.122. Lefroy DC, Wharton J, Crake T et al. Regional changes inangiotensin II receptor density after experimental myocardialinfarction. J Mol Cell Cardiol 1996;28:429-40.123. Sun Y,Zhang JQ,Zhang J,Ramires FJA.Angiotensin II, trans-forming growth factor-β1 and repair in the infarcted heart. JMol Cell Cardiol 1998;30:1559-69.124. Silvestre J-S, Heymes C, Oubénaïssa A et al. Activation of
Weber 155-163 7/11/03 12:15 Page 8
COPYRIG
HT JRA
AS LIM
ITED
REPR
ODUCTION PR
OHIBITE
D
by guest on September 18, 2014jra.sagepub.comDownloaded from

Journal ofthe Renin-Angiotensin-AldosteroneSystem(Including otherpeptidergic systems)
September 2003Volume 4Number 3
163
HEART FAILURE AND THE RENIN-ANGIOTENSIN-ALDOSTERONE SYSTEM
cardiac aldosterone production in rat myocardial infarction.Effect of angiotensin II receptor blockade and role in cardiacfibrosis. Circulation 1999;99:2694-701.125. Delcayre C, Silvestre JS, Garnier A et al. Cardiac aldos-terone production and ventricular remodeling. Kidney Int2000;57:1346-51.126. Hayashi M, Tsutamoto T, Wada A et al. Relationshipbetween transcardiac extraction of aldosterone and left ven-tricular remodeling in patients with first acute myocardialinfarction: extracting aldosterone through the heart promotesventricular remodeling after acute myocardial infarction. J AmColl Cardiol 2001;38:1375-82.127. Tsutamoto T, Wada A, Maeda K et al. Spironolactoneinhibits the transcardiac extraction of aldosterone in patients
with congestive heart failure. J Am Coll Cardiol 2000;36:838-44.128. Van Belle E, Bauters C,Wernert N et al. Neointimal thick-ening after balloon denudation is enhanced by aldosterone andinhibited by spironolactone, and aldosterone antagonist.Cardiovasc Res 1995;29:27-32.129. Slight SH,Chilakamarri VK,Nasr S et al. Inhibition of tissuerepair by spironolactone: role of mineralocorticoids in fibroustissue formation. Mol Cell Biochem 1998;189:47-54.130. Rocha R,Chander PN,Khanna K,Zuckerman A,Stier CT, Jr.Mineralocorticoid blockade reduces vascular injury in stroke-prone hypertensive rats. Hypertension 1998;31(part 2):451-8.131. Weber KT. Aldosteronism revisited. Perspectives on lesswell-recognized actions of aldosterone. J Lab Clin Med2003:142:71-82.
Weber 155-163 7/11/03 12:15 Page 9
COPYRIG
HT JRA
AS LIM
ITED
REPR
ODUCTION PR
OHIBITE
D
by guest on September 18, 2014jra.sagepub.comDownloaded from