strategies and agronomic interventions to improve the phosphorus-use efficiency of farming systems
TRANSCRIPT

REGULAR ARTICLE
Strategies and agronomic interventions to improvethe phosphorus-use efficiency of farming systems
Richard J. Simpson & Astrid Oberson & Richard A. Culvenor & Megan H. Ryan &
Erik J. Veneklaas & Hans Lambers & Jonathan P. Lynch & Peter R. Ryan &
Emmanuel Delhaize & F. Andrew Smith & Sally E. Smith & Paul R. Harvey &
Alan E. Richardson
Received: 1 March 2011 /Accepted: 21 June 2011# Springer Science+Business Media B.V. 2011
Abstract Phosphorus (P)-deficiency is a significantchallenge for agricultural productivity on many highlyP-sorbing weathered and tropical soils throughout theworld. On these soils it can be necessary to apply upto five-fold more P as fertiliser than is exported inproducts. Given the finite nature of global P resources, itis important that such inefficiencies be addressed. Forlow P-sorbing soils, P-efficient farming systems willalso assist attempts to reduce pollution associated with Plosses to the environment. P-balance inefficiency of
farms is associated with loss of P in erosion, runoff orleaching, uneven dispersal of animal excreta, andaccumulation of P as sparingly-available phosphateand organic P in the soil. In many cases it is possible tominimise P losses in runoff or erosion. Uneven dispersalof P in excreta typically amounts to ~5% of P-fertiliserinputs. However, the rate of P accumulation in moderateto highly P-sorbing soils is a major contributor toinefficient P-fertiliser use. We discuss the causaledaphic, plant and microbial factors in the context of
Plant SoilDOI 10.1007/s11104-011-0880-1
Responsible Editor: Matthias Wissuwa.
R. J. Simpson (*) :R. A. Culvenor :A. E. RichardsonCSIRO Sustainable AgricultureNational Research Flagship / CSIRO Plant Industry,GPO Box 1600, Canberra, ACT 2601, Australiae-mail: [email protected]
A. ObersonInstitute of Agricultural Sciences, Group of Plant Nutrition,Research Station Eschikon, ETH Zurich,Lindau, Switzerland
M. H. Ryan : E. J. Veneklaas :H. LambersSchool of Plant Biology, Faculty of Natural andAgricultural Sciences, and Institute of Agriculture,The University of Western Australia,35 Stirling Highway,Crawley, WA 6009, Australia
J. P. LynchDepartment of Horticulture,The Pennsylvania State University,University Park, PA 16802, USA
P. R. Ryan : E. DelhaizeCSIRO Plant Industry,GPO Box 1600, Canberra, ACT 2601, Australia
F. A. Smith : S. E. SmithSoils Group, School of Agriculture, Food and Wine,The University of Adelaide,Waite Campus DX 650 636,Adelaide, SA 5005, Australia
P. R. HarveyCSIRO Sustainable AgricultureNational Research Flagship / CSIRO Ecosystem Sciences,PMB 2,Glen Osmond, SA 5064, Australia

soil P management, P cycling and productivitygoals of farms. Management interventions that canalter P-use efficiency are explored, including bettertargeted P-fertiliser use, organic amendments, removingother constraints to yield, zone management, use ofplants with low critical-P requirements, and modifiedfarming systems. Higher productivity in low-P soils, orlower P inputs in fertilised agricultural systems can beachieved by various interventions, but it is also criticallyimportant to understand the agroecology of plant Pnutrition within farming systems for improvementsin P-use efficiency to be realised.
Keywords Fertiliser. Nutrient-use efficiency. Organicphosphorus . Phosphate fixation . Phosphorusefficiency. Soil fertility. Sparingly-available phosphate
Introduction
Phosphorus (P) is a primary limitation in most forest,weathered and tropical soils (Walker and Syers 1976;Lynch 2007) and P-deficiency is a significant chal-lenge for agricultural productivity on such soils inmany regions of the world. In this review, we draw onexamples largely, but not exclusively, from temperatefarming systems in southern Australia which arebased on moderate to highly P-sorbing soils that aredeficient in P for plant growth. In this context, weadopt Barrow’s (1999) definition of P-sorption asdescribing the adsorption of P to soil particles and thecontinuing reaction of that P with the soil that reducesits availability to plants (Barrow 1973). P-fertiliseruse in agriculture on P-deficient soils that aremoderate to highly P-sorbing, is often relativelyinefficient. For example, P-audits of Australianfarming enterprises indicate that P-inputs are typically2-fold (cropping systems) to 5-fold (grassland systems)higher than P-export in products (e.g. McLaughlin et al.1992; Weaver and Wong 2011). On low P-sorbingsoils, such large imbalances in inputs and outputs wouldbe a strong indication of potential for large P-losses tothe wider environment (Buczko and Kuchenbuch2007). However, P-balance inefficiencies in highly P-sorbing soils are more often associated with accumu-lation of P in the soil, even when P-fertiliserapplication rates are moderate and aimed only atmaintaining adequate levels of plant-available P. Weexplore some of the agronomic and management
options available for improving P-use efficiency inthese circumstances.
In many developing countries, especially in Africa,fertiliser use is low because it is a relatively expensiveinput (World Bank 2004). Many of the principlesdiscussed in the review also apply to low P-inputsystems and will become even more relevant whenfertiliser P is introduced to lift agricultural production.Crops and farming systems developed to increase Pacquisition from low P soils are contributing toimproved food security in these agroecosystems(Lynch 2007) and will also have a role in improvingthe P-efficiency of fertilised agriculture. The P effi-ciency problems for agriculture on moderate to highlyP-sorbing soils, however, do differ from the issuesexperienced in parts of Europe, the USA and Chinawhere diffuse loss of P from agriculture on P-saturatedsoils with low P-sorption capacity is a major environ-mental problem (Haygarth and Jarvis 1999; Sharpley etal. 2001; Ulén et al. 2007; Ma et al 2010; Qiu 2010).Some of the causes of inefficiency in P-use that aredescribed for highly P-sorbing soils (e.g. continuingaccumulations of P in soils fertilised to maintainadequate levels of plant-available P), no longer applyto soils where P-sorption capacity is low. However, thesolutions to P-efficiency problems are often similar. Forexample, the necessity for targeted and appropriate soilfertility management applies equally to high and lowP-sorbing soils. Plants with low external critical-Prequirements will also improve P-efficiency in bothcircumstances, albeit for different reasons. In highly P-sorbing soils they reduce the rate at which Paccumulates in soil (see below), whereas in both highand low P-sorbing soils they will help to reduce theconcentrations of dissolved P in surface runoff anddrainage because dissolved P concentrations areinfluenced by the concentration of plant-available Pin the soil (e.g. Sharpley et al. 2001; Zhao et al. 2007;Melland et al. 2008).
High quality rock phosphate reserves are finite andthere is an on-going debate about the longevity of globalP resources (Cathcart 1980; Stewart et al. 2005; Cordellet al. 2009; Van Kauwenbergh 2010). Given theimportance of P to the sustainability of agriculturalproduction and global food security, and to ensureequitable use of scarce P resources, it is important thatinefficiencies in P use in agriculture be addressed. Animportant objective is to develop productive farmingsystems in which P-fertiliser inputs are reduced. For
Plant Soil

moderate to highly P-sorbing soils, this implies systemsin which P-fertiliser input amounts are much closer toequalling the amounts of P exported in products(Helyar 1998; Syers et al. 2008). However, farmingsystems that have lower P-fertiliser requirementsbecause they export less P in products are also possible(e.g. Rose et al. 2010a; Richardson et al. 2011).
Boundaries for soil fertility managementin agriculture
P-fertilisers are applied to P-deficient soils to lift ormaintain productivity per hectare. The objective is toachieve concentrations of plant-available P in the soilthat enhance plant growth and allow the productiongoals of the farm to be met. The ‘logical’ upperboundary for soil fertility management is the ‘criticalP’ requirement of the crop-soil system (usuallydefined as the soil fertility level corresponding to90% or 95% of maximum yield (e.g. Peverill et al.1999). Managing soil fertility near the critical P levelis expected to maximise productivity. However, it isalso legitimate to manage soil fertility at levels belowcritical P for a variety of farm business or environ-mental reasons (e.g. business risk mitigation andmaintenance of ground cover, Mokany et al. 2010;native perennial grass persistence, Simpson andLangford 1996).
When P-inputs are very low, net extraction of P fromsoil occurs (Weaver andWong 2011; McIvor et al. 2011)and production will not be sustained over the longerterm. The risk of nutrient exhaustion thus sets a lower‘sustainability' boundary for soil fertility management.Even in low-production farm systems, there is a needfor P-inputs that can at least cover the amount of Pexported in products. The consequences of nutrientmining have already been observed in temperateAustralian agriculture with a widespread decline inproductivity of wheat (Triticum aestivum L.) recordedin the era before superphosphate and improved farmingpractices were adopted (e.g. Donald 1965; Angus2001; Passioura 2002) and is also observed in somelow input farming systems when inadequate amountsof fertiliser are applied (e.g. Burkitt et al. 2007).Similarly the consequence of nutrient depletion and theneed to address fertiliser inputs to enhance productiveagriculture on tropical African soils has been outlined(Sánchez 2010).
Ideal fertiliser management for a P-deficient soilwill typically involve a phase in which soil P-fertility is increased, followed by a soil fertilitymaintenance phase in which soil P levels are heldwithin a target range (Fig. 1). Variations on thispattern of management which involve periods of soilfertility increase followed by nutrient extractionwithin the logical and sustainable boundaries arealso used for a variety of pragmatic reasons (e.g.unfertilised pasture phases using residual P from apreceding crop phase, Kohn (1974); aerial fertilisa-tion of pasture at multi-year intervals in steep hillcountry, e.g. Scott 2000)
Although it is highly desirable that the P-fertiliser requirements of a production system areapproximately equal to the removal of P in productsirrespective of the soil P-fertility level being maintained(Helyar 1998; Syers et al. 2008), this is presently onlyachieved in farming systems operating with verylow productivity, or on soils with low P-sorptioncapacity where the risk of P-loss by leaching andlateral flow is often large (Weaver and Wong 2011;Johnston and Syers 2009). It is a significantchallenge to improve the P-balance efficiency ofhighly productive agricultural systems on soils withmoderate to high P-sorption capacity without com-promising productivity.
A framework for understanding P-efficiency
Although the reactions of phosphate with soil and theprocesses leading to P accumulation in soil are verycomplex (McLaughlin et al. 2011), it is possible toassess the options for improving the P-balance effi-ciency of a farming system by envisaging the mainsources, flows and effective sinks for P in farmingsystems (Fig. 2). The amount of P held in the cyclingpools of soil P (i.e. plant-available P) is the workingcapital of the soil and it is the concentration of plant-available P in this pool that determines productivity.This is often considerably above the concentration ofavailable P in natural ecosystems because of the yieldimperatives imposed by commerce, and the need to usewater and land resources efficiently. For agriculturalproduction to be maintained, the cycling pools of Pmust be maintained by fertiliser applications wheneverP is removed or lost from this pool. In a system whereplant-available P is being maintained at a stable
Plant Soil
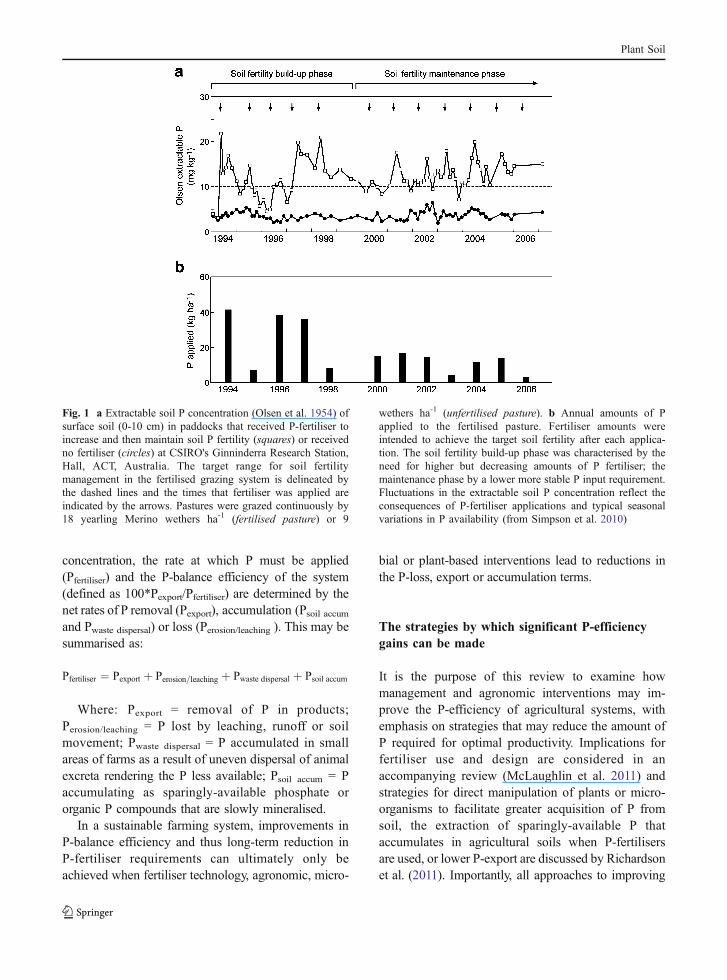
concentration, the rate at which P must be applied(Pfertiliser) and the P-balance efficiency of the system(defined as 100*Pexport/Pfertiliser) are determined by thenet rates of P removal (Pexport), accumulation (Psoil accumand Pwaste dispersal) or loss (Perosion/leaching ). This may besummarised as:
Pfertiliser ¼ Pexport þ Perosion=leaching þ Pwaste dispersal þ Psoil accum
Where: Pexport = removal of P in products;Perosion/leaching = P lost by leaching, runoff or soilmovement; Pwaste dispersal = P accumulated in smallareas of farms as a result of uneven dispersal of animalexcreta rendering the P less available; Psoil accum = Paccumulating as sparingly-available phosphate ororganic P compounds that are slowly mineralised.
In a sustainable farming system, improvements inP-balance efficiency and thus long-term reduction inP-fertiliser requirements can ultimately only beachieved when fertiliser technology, agronomic, micro-
bial or plant-based interventions lead to reductions inthe P-loss, export or accumulation terms.
The strategies by which significant P-efficiencygains can be made
It is the purpose of this review to examine howmanagement and agronomic interventions may im-prove the P-efficiency of agricultural systems, withemphasis on strategies that may reduce the amount ofP required for optimal productivity. Implications forfertiliser use and design are considered in anaccompanying review (McLaughlin et al. 2011) andstrategies for direct manipulation of plants or micro-organisms to facilitate greater acquisition of P fromsoil, the extraction of sparingly-available P thataccumulates in agricultural soils when P-fertilisersare used, or lower P-export are discussed by Richardsonet al. (2011). Importantly, all approaches to improving
Fig. 1 a Extractable soil P concentration (Olsen et al. 1954) ofsurface soil (0-10 cm) in paddocks that received P-fertiliser toincrease and then maintain soil P fertility (squares) or receivedno fertiliser (circles) at CSIRO's Ginninderra Research Station,Hall, ACT, Australia. The target range for soil fertilitymanagement in the fertilised grazing system is delineated bythe dashed lines and the times that fertiliser was applied areindicated by the arrows. Pastures were grazed continuously by18 yearling Merino wethers ha-1 (fertilised pasture) or 9
wethers ha-1 (unfertilised pasture). b Annual amounts of Papplied to the fertilised pasture. Fertiliser amounts wereintended to achieve the target soil fertility after each applica-tion. The soil fertility build-up phase was characterised by theneed for higher but decreasing amounts of P fertiliser; themaintenance phase by a lower more stable P input requirement.Fluctuations in the extractable soil P concentration reflect theconsequences of P-fertiliser applications and typical seasonalvariations in P availability (from Simpson et al. 2010)
Plant Soil

P-use efficiency must address the need to reduce therates of P-loss, export or accumulation in agriculturalsystems if significant change is to be achieved in theeffectiveness with which scarce P resources are used.
P export in products
The objective of applying P-fertiliser is to increase ormaintain agricultural production and it is inevitablethat P will be exported in products. The amount of Pexported in products sets the minimum amount of Pthat must be replaced by fertiliser applications if thefarming system is to be sustained. In theory, it isfeasible to reduce P-fertiliser inputs by developingfarm systems that deliver products (plants, seeds,fruits, animal products) that have a low P contentleading to lower P-export (Rose et al. 2010b;Richardson et al. 2011). However, it is notable thatagricultural enterprises that export more P-intensiveproducts (e.g. grains, dairy products) tend to be thosewith higher P-balance efficiencies (McLaughlin et al.1992; Weaver and Wong 2011). Wool productionsystems are an example of systems with very lowrates of P-export (e.g. Simpson et al. 2010) becausethe main product (clean wool) contains no P andexport is only associated with animals culled at theend of their commercial life or as traces of P in thesuint of raw wool. However, on moderate to highly P-sorbing soils these farming systems tend to accumu-late P in the soil (McLaughlin et al. 1990) and havevery low P-balance efficiency (median ~15%; Weaverand Wong 2011). A number of factors may contributeto low P efficiency, including the practice of broad-
casting fertiliser on pastures (McLaughlin et al. 2011)and uneven distributions of animal excreta in paddocks(Williams and Haynes 1992), However, the generalassociation between farming systems with low P-export and low P-balance efficiency, raises the possi-bility that if high rates of P return to soil (in plantmaterial or animal waste) are a consequence of low Pexport, the expected efficiency gain from reducing Pexports may be counteracted by larger accumulationsof sparingly-available P in the soil.
P-balance efficiency and economic efficiency($-return per unit P-fertiliser) will also be lifted byany practice that addresses constraints to yield andleads to increased P-export per unit of P input.However, attempts to reduce P-export by reducingthe level of production will be counterproductivebecause production per hectare will be low, profit-ability compromised, and the farm system will not beusing land or water resources efficiently.
P-losses (erosion, runoff and leaching)
Reducing loss of P from a farming system, other thanin farm products, will directly improve P-balanceefficiency. P losses by erosion or runoff are notcovered in detail in this review as there is extensiveliterature covering actions that can be taken on farmto minimise these pathways for P loss including theuse of appropriate forms and placement of fertiliser,appropriate timing of fertiliser in relation to rain andcrop growth, use of soil amendments to reducenutrient transport, use of buffer strips and fencing ofwaterways to capture mobilised nutrients and avoid
Fig. 2 Diagrammatic repre-sentation of the inputs,flows and outputs of P inagriculture illustrating thepotential components ofthe P-balance of a farmingsystem (modified fromHelyar and Godden 1976)
Plant Soil

direct contamination, location of field access points,attention to cultivation methods and use of minimumtillage and attention to ground cover (e.g. Weaver etal. 1988; Nash and Haliwell 1999; Sharpley et al.2000; Mathers et al. 2007). In many systems, P lossdue to erosion and runoff should be a relatively smallcomponent of the P-balance of the farming system,especially when best-practice management is fol-lowed. However, losses that are small in terms of afarm’s P-balance are, nevertheless, a significantconcern for stream health and need to be addressed.Diffuse losses of P over large areas are causingsubstantial environmental problems in parts ofEurope, the USA and China (Haygarth and Jarvis1999; Sharpley et al. 2001; Buczko and Kuchenbuch2007; Ulén et al. 2007; Qiu 2010) and are issues on asmaller, but significant scale in many other parts ofthe world (e.g. Birch 1982; Hodgkin and Hamilton1993; McCulloch et al. 2003).
Losses due to leaching vary considerably with soiltype, fertiliser management and plant species and canbe a significant component of a farm’s P-balance. Forfarming systems on soils with moderate to high P-sorption capacity, losses are usually relatively small(<0.4%–<5% of applied P, McCaskill and Cayley2000; Ridley et al. 2003; Melland et al. 2008).However, those on low P-sorbing soils with poor Pretention capacity can experience very large P losses(40%–90% of applied P: Ozanne et al. 1961; Lewis etal. 1987) which need to be addressed for bothfinancial and environmental reasons. Partial solutionsto P leaching losses have been achieved by using lesssoluble forms of P-fertiliser and deep rooted pasturespecies (Weaver et al. 1988; Ozanne et al. 1961), butin some cases this also reduces the availability of Pfor plant growth (Ozanne et al. 1961).
Uneven dispersal of excreta
Accumulation of P within paddocks, or within farmsystems due to uneven distribution of excreta is aunique problem of livestock systems, especially thosewhich involve grazing. Estimates of the relative sizeof this accumulation term for grazed paddocks aretypically about 5% of the amount of P applied asfertiliser (e.g. Metherell 1994). Adjustment to farmmanagement can address this accumulation to someextent and will make a contribution, albeit sometimesrelatively small, to improved P-balance efficiency.
Uneven distribution of nutrients across farms can alsooccur and may be associated with regular patterns ofstock movement, or in the case of housed stocksystems or feed lots, when manures and slurry arespread unevenly.
P accumulation in “non-equilibrating” pools in soil
When phosphate is applied to many soils as fertiliser,it is subject to continuing reactions with the soil andincorporation into organic matter that leads ultimatelyto accumulations of P (e.g. McLaughlin et al. 1990) insparingly-available phosphate and organic P forms. Inlow-P soils the processes leading to P accumulation,also manifest themselves over time as a decline in theresidual value of freshly applied fertiliser (Barrow andCarter 1978). In early literature, the pools ofsparingly-available phosphate were sometimes re-ferred to as “irreversibly adsorbed” or “fixed”, butthese descriptors are technically incorrect. Phosphateis initially adsorbed to the surface of soil particles orprecipitates with Ca, Al and Fe (Sample et al. 1980;Pierzynski et al. 2005; McLaughlin et al. 2011). Thecontinuing reaction of phosphate with the soil mayalso involve slow diffusion into the reacting soilparticles (Barrow 1999). The forward sorption reac-tion is driven by the phosphate concentration in thesoil solution and is considerably faster than the back(desorption) reaction that will occur when the soilsolution phosphate concentration is depleted (Barrow1983a; 1983b). Conceptually, the accumulation of Pin slowly cycling organic matter is similar. Thevarious components of organic matter that arereturned to soil are mineralised at different ratesdepending on their chemical and physical protection(Krull et al. 2003). All organic matter, even the moreresistant materials (e.g. humus) turnover, albeit some-times at very slow rates (Krull et al. 2003). However, fora farming system being fertilised annually to maintainsoil P fertility, the rate of organic P accumulation will bedetermined by those components of soil organic matterthat take longer than a year to be mineralised.
P is expected to accumulate in an agricultural soilwhen P-fertiliser is applied to increase soil fertility (e.g.McLaughlin et al. 1990; Bünemann et al. 2006), butthere are relatively few reports of the amounts of P thatare accumulated in soil maintained under steady-stateP-fertility conditions, where accumulation will be thenet result of phosphate sorption/desorption reactions
Plant Soil

and the slowly cycling components of soil organicmatter that release phosphate at rates slower than therate of P supply necessary for commercial agriculturalproduction. Two long-term experiments: a pasture-wheat crop rotation (Wagga Wagga, New South Wales[NSW], Australia; Helyar et al. 1997) which accumu-lated ~4.6 kg P ha-1 year-1 (P-balance efficiency ~60%),and a permanent pasture (Hall, Australian CapitalTerritory, Australia; Simpson et al. 2010) whichaccumulated ~7.3 kg P ha-1 year-1 when managed atnear-optimal P fertility (P-balance efficiency ~20%),demonstrate how large the soil P accumulation term ofthe P-balance equation can be for moderate to highlyP-sorbing soils under relatively “stable” soil P-fertilityconditions. For accumulation of P to be reduced underthese conditions, it would be necessary to shift thebalance between phosphate sorption and desorptionrates, and/or between the rates of soil organic P inputand mineralisation.
Reducing the accumulation of sparingly-availablephosphate
Barrow (1980c) cited lack of ability in manyagricultural plants to access residual fertiliser phos-phate and the very slow rates of phosphate desorptiononce phosphate had reacted with soil over longperiods of time, as reasons for pessimism about thechances of finding plants that can usefully accesssparingly-available phosphate. However, a limitednumber of species can lower the pH of the rhizo-sphere or are able to exude organic anions to establishthe chemical conditions that increase phosphatedesorption rates, depending on soil type (Gerke1994; Barrow 2002). In the case of the much-studied white lupin (Lupinus albus L.), it is evidentthat citrate exudation from proteoid roots confers theability to access P-pools that are poorly accessible tomany other plant species (Gardner et al. 1983;Dinkelaker et al. 1989; Hocking et al. 1997) and,when grown in close association with other species,enables the companion plant to also access sparingly-available phosphate (Gardner and Boundy 1983;Horst et al. 2001; Cu et al. 2005). On the other hand,buckwheat (Fagopyrum esculentum Moench) whichproduces exudates with a lower pH in response tolow-P conditions (Amann and Amberger 1989), hashigh P-uptake efficiency in calcareous soils (Zhu et al.
2002) but is less effective in soils dominated by Feand Al phosphates (Otani and Ae 1996). Nevertheless,the general effectiveness of agronomic strategiesintended to enhance phosphate desorption remainsunclear as there are relatively few examples of cropspecies that gain substantial P nutrition benefits bythese mechanisms.
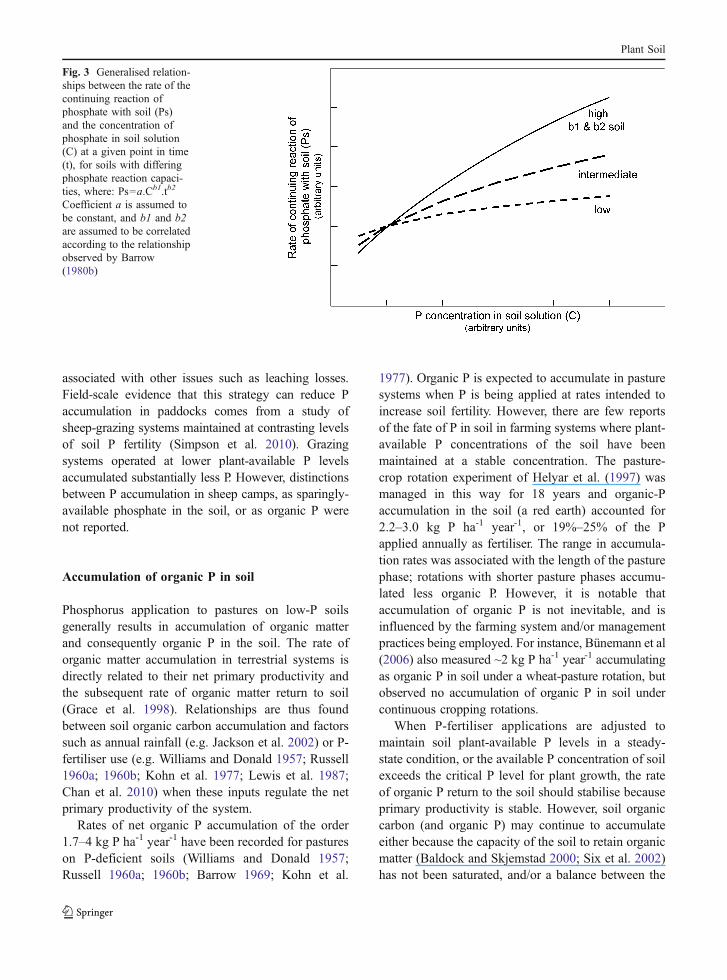
It should also be possible to reduce the accumula-tion of phosphate by slowing the rate at whichphosphate continues to react slowly with soil, makingit less available for plant growth. The reactions ofphosphate with soil are described in more detail byMcLaughlin et al. (2011), but it is helpful to considerthe empirical relationship between net phosphatesorbed by a soil (Ps) , P concentration in the soilsolution (C) and time (t) as observed for non-calcareoussoils (Barrow 1980a; 1980b).
Ps ¼ a:Cb1:tb2
where: a approximates the amount of sorbing materialin a soil, and b1 and b2 are coefficients that describe theshape of the sorption relationship. The coefficients varywidely between soils; however, b1 and b2 are reason-ably well correlated when compared across a widerange of soils (Barrow 1980a; 1980b). In calcareoussoils this equation also applies initially, but precipitationof calcium phosphates decreases the phosphate concen-tration of the soil solution to levels that are determinedby the solubility product (Barrow 1980a).
In practice, Ps is found to vary somewhat betweensoils that differ in P-sorption capacity (Barrow 1973)and is influenced strongly by the b1 and b2 exponentsthat characterise each soil. Because b1 and b2 valuestend to be correlated (Barrow 1980a; 1980b), Ps isexpected to be influenced by the phosphate concen-tration at which soil solution is maintained in thegeneralised manner shown in Fig. 3. This indicatesthat strategies that allow a farming system to beoperated at lower soil phosphate concentrationsshould reduce the rate at which slow reactions ofphosphate and soil lead to P accumulation and shouldconsequently improve P-balance efficiency. The effec-tiveness of such a strategy will be greatest in soils withhigh b1/b2 values (where continuing sorption reactionswith phosphate are slow but persistent) and least insoils with low b1/b2 values (where sorption reactionsare not as persistent). It is expected that P-inefficiencyin soils with low b1/b2 values will tend to be
Plant Soil
associated with other issues such as leaching losses.Field-scale evidence that this strategy can reduce Paccumulation in paddocks comes from a study ofsheep-grazing systems maintained at contrasting levelsof soil P fertility (Simpson et al. 2010). Grazingsystems operated at lower plant-available P levelsaccumulated substantially less P. However, distinctionsbetween P accumulation in sheep camps, as sparingly-available phosphate in the soil, or as organic P werenot reported.
Accumulation of organic P in soil
Phosphorus application to pastures on low-P soilsgenerally results in accumulation of organic matterand consequently organic P in the soil. The rate oforganic matter accumulation in terrestrial systems isdirectly related to their net primary productivity andthe subsequent rate of organic matter return to soil(Grace et al. 1998). Relationships are thus foundbetween soil organic carbon accumulation and factorssuch as annual rainfall (e.g. Jackson et al. 2002) or P-fertiliser use (e.g. Williams and Donald 1957; Russell1960a; 1960b; Kohn et al. 1977; Lewis et al. 1987;Chan et al. 2010) when these inputs regulate the netprimary productivity of the system.
Rates of net organic P accumulation of the order1.7–4 kg P ha-1 year-1 have been recorded for pastureson P-deficient soils (Williams and Donald 1957;Russell 1960a; 1960b; Barrow 1969; Kohn et al.
1977). Organic P is expected to accumulate in pasturesystems when P is being applied at rates intended toincrease soil fertility. However, there are few reportsof the fate of P in soil in farming systems where plant-available P concentrations of the soil have beenmaintained at a stable concentration. The pasture-crop rotation experiment of Helyar et al. (1997) wasmanaged in this way for 18 years and organic-Paccumulation in the soil (a red earth) accounted for2.2–3.0 kg P ha-1 year-1, or 19%–25% of the Papplied annually as fertiliser. The range in accumula-tion rates was associated with the length of the pasturephase; rotations with shorter pasture phases accumu-lated less organic P. However, it is notable thataccumulation of organic P is not inevitable, and isinfluenced by the farming system and/or managementpractices being employed. For instance, Bünemann et al(2006) also measured ~2 kg P ha-1 year-1 accumulatingas organic P in soil under a wheat-pasture rotation, butobserved no accumulation of organic P in soil undercontinuous cropping rotations.
When P-fertiliser applications are adjusted tomaintain soil plant-available P levels in a steady-state condition, or the available P concentration of soilexceeds the critical P level for plant growth, the rateof organic P return to the soil should stabilise becauseprimary productivity is stable. However, soil organiccarbon (and organic P) may continue to accumulateeither because the capacity of the soil to retain organicmatter (Baldock and Skjemstad 2000; Six et al. 2002)has not been saturated, and/or a balance between the
Fig. 3 Generalised relation-ships between the rate of thecontinuing reaction ofphosphate with soil (Ps)and the concentration ofphosphate in soil solution(C) at a given point in time(t), for soils with differingphosphate reaction capaci-ties, where: Ps=a.Cb1.tb2
Coefficient a is assumed tobe constant, and b1 and b2are assumed to be correlatedaccording to the relationshipobserved by Barrow(1980b)
Plant Soil

input of plant residues and the rate of mineralisationhas not been reached (Baldock et al. 2007).
Fractionation of soil organic matter into physical orconceptual classes (Baldock and Skjemstad 1999;Amelung and Zech 1999) permits the prediction oforganic matter accumulation in soil because the organicmatter fractions have characteristic turnover rates(Skjemstad et al. 2004). It is clear that soil organicmatter (and consequently organic P) accumulation willequilibrate at some point, but modelling of soil organicmatter accumulation for a permanent pasture insouthern NSW, Australia predicted that soil organicmatter pools would only equilibrate after ~200 years ofregular fertiliser applications in this particular farmingsystem (Baldock et al. 2007).
The concept of eventual equilibration of organic Paccumulation is supported by empirical data fromfarming systems fertilised over very long timeperiods: e.g. Park Grass (Rothamsted, UK) whereafter 100 years of superphosphate fertilisation, onlylow proportions of accumulated P (1%–6% topsoil,10%–18% subsoil layers) were in the form of organicP (Oniani et al. 1973), and from various examples ofsoil organic carbon stabilisation as cited by Stewart etal. (2007). Unfortunately, the time frame for organic Pequilibration may be sufficiently long that relying onthe equilibration of soil organic matter concentrationsto improve P-balance efficiency will not provide atimely solution to the issue of P-imbalance.
For P-balance efficiency to be improved by reducingthe accumulation of organic P in soil, it will be necessaryto increase the rates of soil organic matter turnover sothat soil organic matter and organic P pools equilibrateat lower concentrations. Inevitably, the objective ofreducing organic P accumulations by this route willcompete directly with objectives to increase carbonstorage in soils (Dalal and Chan 2001; Lal 2004).
The stoichiometric relationship between C:N:S instabilised soil organic matter (soil humus) is relativelyconstant across a wide range of soils irrespective ofwide variability of the ratio in fresh organic orparticulate organic matter fractions. Variation in thelatter fractions depends on the source materials andcrop and soil management practices. However, C:Pand C:organic P ratios of stabilised soil organic mattercan vary considerably (Kirkby et al. 2011). Presently,it is unclear whether the variation in C:P ratios ofstabilised soil organic matter is a result of differencesbetween analytical methods or reflects real differences
in the composition of organic P in soils. If thestoichiometric ratios for C:N:S and C:P in soil organicmatter prove not to be tightly coupled, it is possiblethat enhancing release of phosphate from specificorganic-P compounds in soils by release of plant ormicrobial phosphatases (Richardson et al. 2005) couldpotentially lower net rates of organic P accumulationindependently of effects on soil organic matter accumu-lation. Orthophosphate monoesters (e.g. phytate) havebeen identified as a major form of organic P in manysoils and are accompanied by lesser amounts ofphospholipids, nucleic acids, phosphonates andother compounds (Turner et al. 2005; Smernik andDougherty 2007). However, large proportions of thesoil organic P remain poorly identified as high-molecular-weight material that is intimately associ-ated with the soil humic fraction and this currentlylimits our understanding of the opportunities formobilising organic P reserves for plant growth(Guppy and McLaughlin 2009).
Microbial contributions to organic Pmineralisation and the cycling of P in soil
Several key processes in the P cycle which affect theavailability of P to plants are mediated by microbialprocesses (Oberson and Joner 2005; Turner et al. 2006;Richardson and Simpson 2011). Most significantlymicroorganisms decompose and mineralise P fromorganic soil amendments (manures, organic waste com-posts, plant residues) and soil organic matter. The soilmicrobial biomass itself represents a P pool containingat least 0.5% to 7.5% of total P in grassland soils, and0.4% to 2.4% of total P in arable soils (Oberson andJoner 2005). The microbial P pool is highly dynamicbecause microorganisms react readily to changes in soiltemperature, moisture and carbon availability.
Amendment of a soil with a carbon sourcestimulates microbial growth and this is paralleled bya rapid decrease of soil solution P concentration (Oehlet al. 2001; Bünemann et al. 2004a; Fig. 4) and thetemporary immobilisation of P in microbial biomass.Even under low-P conditions the microbial biomasscan grow rapidly and may increase several fold whenreadily degradable carbon is available (Bünemann etal. 2004a). In the short term, this may occur incompetition with plants for available P (Olander andVitousek 2004). Plants themselves may promote micro-
Plant Soil
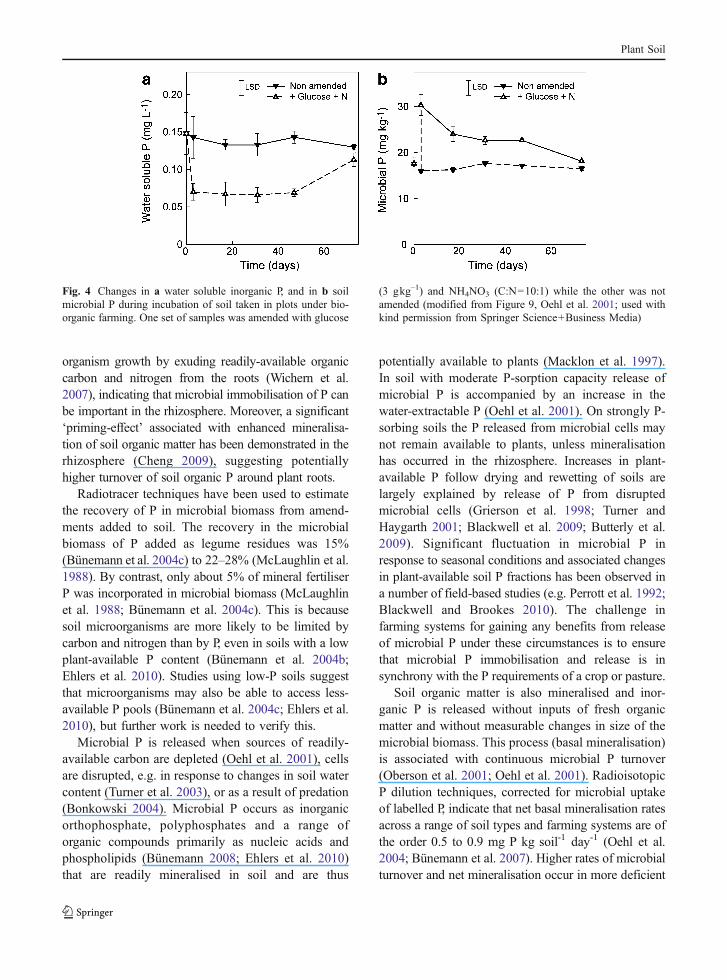
organism growth by exuding readily-available organiccarbon and nitrogen from the roots (Wichern et al.2007), indicating that microbial immobilisation of P canbe important in the rhizosphere. Moreover, a significant‘priming-effect’ associated with enhanced mineralisa-tion of soil organic matter has been demonstrated in therhizosphere (Cheng 2009), suggesting potentiallyhigher turnover of soil organic P around plant roots.
Radiotracer techniques have been used to estimatethe recovery of P in microbial biomass from amend-ments added to soil. The recovery in the microbialbiomass of P added as legume residues was 15%(Bünemann et al. 2004c) to 22–28% (McLaughlin et al.1988). By contrast, only about 5% of mineral fertiliserP was incorporated in microbial biomass (McLaughlinet al. 1988; Bünemann et al. 2004c). This is becausesoil microorganisms are more likely to be limited bycarbon and nitrogen than by P, even in soils with a lowplant-available P content (Bünemann et al. 2004b;Ehlers et al. 2010). Studies using low-P soils suggestthat microorganisms may also be able to access less-available P pools (Bünemann et al. 2004c; Ehlers et al.2010), but further work is needed to verify this.
Microbial P is released when sources of readily-available carbon are depleted (Oehl et al. 2001), cellsare disrupted, e.g. in response to changes in soil watercontent (Turner et al. 2003), or as a result of predation(Bonkowski 2004). Microbial P occurs as inorganicorthophosphate, polyphosphates and a range oforganic compounds primarily as nucleic acids andphospholipids (Bünemann 2008; Ehlers et al. 2010)that are readily mineralised in soil and are thus
potentially available to plants (Macklon et al. 1997).In soil with moderate P-sorption capacity release ofmicrobial P is accompanied by an increase in thewater-extractable P (Oehl et al. 2001). On strongly P-sorbing soils the P released from microbial cells maynot remain available to plants, unless mineralisationhas occurred in the rhizosphere. Increases in plant-available P follow drying and rewetting of soils arelargely explained by release of P from disruptedmicrobial cells (Grierson et al. 1998; Turner andHaygarth 2001; Blackwell et al. 2009; Butterly et al.2009). Significant fluctuation in microbial P inresponse to seasonal conditions and associated changesin plant-available soil P fractions has been observed ina number of field-based studies (e.g. Perrott et al. 1992;Blackwell and Brookes 2010). The challenge infarming systems for gaining any benefits from releaseof microbial P under these circumstances is to ensurethat microbial P immobilisation and release is insynchrony with the P requirements of a crop or pasture.
Soil organic matter is also mineralised and inor-ganic P is released without inputs of fresh organicmatter and without measurable changes in size of themicrobial biomass. This process (basal mineralisation)is associated with continuous microbial P turnover(Oberson et al. 2001; Oehl et al. 2001). RadioisotopicP dilution techniques, corrected for microbial uptakeof labelled P, indicate that net basal mineralisation ratesacross a range of soil types and farming systems are ofthe order 0.5 to 0.9 mg P kg soil-1 day-1 (Oehl et al.2004; Bünemann et al. 2007). Higher rates of microbialturnover and net mineralisation occur in more deficient
Fig. 4 Changes in a water soluble inorganic P, and in b soilmicrobial P during incubation of soil taken in plots under bio-organic farming. One set of samples was amended with glucose
(3 gkg–1) and NH4NO3 (C:N=10:1) while the other was notamended (modified from Figure 9, Oehl et al. 2001; used withkind permission from Springer Science+Business Media)
Plant Soil

soils, and as such may contribute substantially to plantnutrition in low-input agro-ecosystems and naturalecosystems; this highlights the importance of sustainedinput and turnover of soil organic matter.
Management options to improve the P-balanceefficiency of agricultural systems
There are a variety of options for addressing P-balanceefficiency in crop and pasture systems. P-fertiliserplacements and technology are reviewed byMcLaughlinet al. (2011). Here we examine the potential formanagement interventions that affect the cycling of P,P-availability to plants and the potential for farmingwith plants known to exhibit P-efficiency attributes.
Microbial activity and organic P cycling
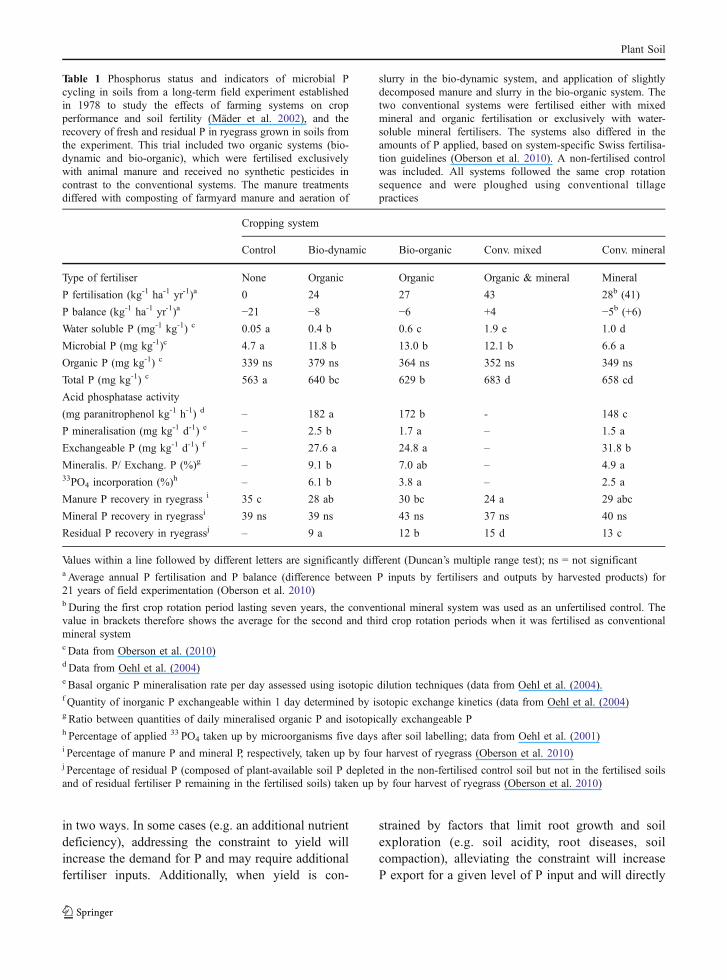
P-balance efficiency of many farming systems wouldbe improved if it were possible to increase the rates atwhich sparingly-available organic P is mineralised. Itis sometimes asserted that this may be achieved byenhancing the microbial activity of soil. Farmingsystems that receive animal manures regularly (i.e.either exclusively organic, or conventional systemsthat include manure return) maintain higher soilorganic carbon levels (Leifeld et al. 2009). Soilmicrobial biomass and activity is also consistentlyhigher in manured soils as opposed to those managedexclusively with mineral fertilisation (Table 1) (Mäderet al. 2002; Fliessbach et al. 2007; Joergensen et al.2010). This translates into a higher soil microbial Ppool with faster turnover (Oehl et al. 2001) and intohigher basal soil organic P mineralisation underorganic than conventional cropping with exclusivelymineral fertilisation (Oehl et al. 2004) (Table 1).Despite the different levels of microbial activity in thesoils under the various management regimes (Table 1),recovery of fresh manure P by ryegrass was lowerthan recovery of mineral P (24% to 35% for manureP; 37 to 43% for mineral P, Table 1) (Oberson et al.2010). Recovery of fresh manure P was affected bythe available P content of the soil and was lower insoils with higher plant-available P. Uptake of residualP was also lower from organically managed soils thanfrom conventionally managed soils. However, thisresult must be interpreted with care because the totaland available P contents of soil were depleted more in
organic systems than in the conventional systems asrevealed by their P budget deficit (Table 1) (Oehl etal. 2002). Thus, yields in the organic systems wereattained partly at the expense of soil P reserves,leaving less residual P in the soils (Oberson et al.2010). P fractionation of these soils indicated that themore residual P is depleted, the less plant-availablethe residual P will be (Oberson et al. 1993). Soilsunder organic and conventional systems did not differin NaHCO3-extractable or NaOH-extractable organicP fractions (Oberson et al. 1993). However, total soilorganic P was higher in the organic systems (Oehl etal. 2004) and as a consequence the sparingly-availableorganic P fraction tended to be higher (Oberson et al.1993).
A series of farming systems studies on highly P-sorbing tropical soils have similarly demonstratedhow enhancing soil organic P, and consequentlymicrobial activity, can increase the rate of P cyclingand the plant-availability of P in grass-legume andgrass-only pastures compared with native savannah(Oberson et al. 1999). A beneficial effect of grass-legume pastures on P cycling and P availability wasalso found in crop-pasture rotations (Gijsman et al.1997; Oberson et al. 2001). Differences in crop orforage residue decomposition and P release ratessuggest that managing the interactions of residue withsoil may help to slow strong P sorption reactions(Friesen et al. 1997) and that P held in the soil microbialbiomass is also protected, at least temporarily, fromsorption. Although these experiments demonstrated thatthe rates of organic P mineralisation and the size of themicrobial P pool could be enhanced in soils underdifferent farming regimes, there was little evidence thathigher microbial activity also rendered the sparingly-available soil P fractions more available for cropuptake. This conclusion has parallels to the study ofDann et al. (1996) who found uptake of P from rockphosphate was equally low on soils under organic orconventional cropping, suggesting that P-solubilisationactivity was also not enhanced.
Minimising inefficiencies associated with wasteand constraints to yield
Removing constraints to crop yield
Constraints to crop yield, other than those due to P-availability itself, can influence P-balance efficiency
Plant Soil
in two ways. In some cases (e.g. an additional nutrientdeficiency), addressing the constraint to yield willincrease the demand for P and may require additionalfertiliser inputs. Additionally, when yield is con-
strained by factors that limit root growth and soilexploration (e.g. soil acidity, root diseases, soilcompaction), alleviating the constraint will increaseP export for a given level of P input and will directly
Table 1 Phosphorus status and indicators of microbial Pcycling in soils from a long-term field experiment establishedin 1978 to study the effects of farming systems on cropperformance and soil fertility (Mäder et al. 2002), and therecovery of fresh and residual P in ryegrass grown in soils fromthe experiment. This trial included two organic systems (bio-dynamic and bio-organic), which were fertilised exclusivelywith animal manure and received no synthetic pesticides incontrast to the conventional systems. The manure treatmentsdiffered with composting of farmyard manure and aeration of
slurry in the bio-dynamic system, and application of slightlydecomposed manure and slurry in the bio-organic system. Thetwo conventional systems were fertilised either with mixedmineral and organic fertilisation or exclusively with water-soluble mineral fertilisers. The systems also differed in theamounts of P applied, based on system-specific Swiss fertilisa-tion guidelines (Oberson et al. 2010). A non-fertilised controlwas included. All systems followed the same crop rotationsequence and were ploughed using conventional tillagepractices
Cropping system
Control Bio-dynamic Bio-organic Conv. mixed Conv. mineral
Type of fertiliser None Organic Organic Organic & mineral Mineral
P fertilisation (kg-1 ha-1 yr-1)a 0 24 27 43 28b (41)
P balance (kg-1 ha-1 yr-1)a −21 −8 −6 +4 −5b (+6)Water soluble P (mg-1 kg-1) c 0.05 a 0.4 b 0.6 c 1.9 e 1.0 d
Microbial P (mg kg-1)c 4.7 a 11.8 b 13.0 b 12.1 b 6.6 a
Organic P (mg kg-1) c 339 ns 379 ns 364 ns 352 ns 349 ns
Total P (mg kg-1) c 563 a 640 bc 629 b 683 d 658 cd
Acid phosphatase activity
(mg paranitrophenol kg-1 h-1) d – 182 a 172 b - 148 c
P mineralisation (mg kg-1 d-1) e – 2.5 b 1.7 a – 1.5 a
Exchangeable P (mg kg-1 d-1) f – 27.6 a 24.8 a – 31.8 b
Mineralis. P/ Exchang. P (%)g – 9.1 b 7.0 ab – 4.9 a33PO4 incorporation (%)h – 6.1 b 3.8 a – 2.5 a
Manure P recovery in ryegrass i 35 c 28 ab 30 bc 24 a 29 abc
Mineral P recovery in ryegrassi 39 ns 39 ns 43 ns 37 ns 40 ns
Residual P recovery in ryegrassj – 9 a 12 b 15 d 13 c
Values within a line followed by different letters are significantly different (Duncan’s multiple range test); ns = not significanta Average annual P fertilisation and P balance (difference between P inputs by fertilisers and outputs by harvested products) for21 years of field experimentation (Oberson et al. 2010)b During the first crop rotation period lasting seven years, the conventional mineral system was used as an unfertilised control. Thevalue in brackets therefore shows the average for the second and third crop rotation periods when it was fertilised as conventionalmineral systemcData from Oberson et al. (2010)d Data from Oehl et al. (2004)e Basal organic P mineralisation rate per day assessed using isotopic dilution techniques (data from Oehl et al. (2004).f Quantity of inorganic P exchangeable within 1 day determined by isotopic exchange kinetics (data from Oehl et al. (2004)g Ratio between quantities of daily mineralised organic P and isotopically exchangeable Ph Percentage of applied 33 PO4 taken up by microorganisms five days after soil labelling; data from Oehl et al. (2001)i Percentage of manure P and mineral P, respectively, taken up by four harvest of ryegrass (Oberson et al. 2010)j Percentage of residual P (composed of plant-available soil P depleted in the non-fertilised control soil but not in the fertilised soilsand of residual fertiliser P remaining in the fertilised soils) taken up by four harvest of ryegrass (Oberson et al. 2010)
Plant Soil

improve P-balance efficiency. In reality, it is imprac-tical for the published targets for soil fertilitymanagement (e.g. Moody 2007; Gourley et al. 2007)to recognise the existence of other constraints to yieldand P will often have been supplied in excess ofactual requirements when a yield constraint of anysort exists. Thus, the alleviation of any yieldconstraint will often improve P-use efficiency.
Acid soils are a clear example of a constraint toroot foraging ability, plant yield and P-use efficiency.Acid soils are widespread (von Uexküll and Mutert1995) and limit plant production when the trivalentaluminium (Al3+) concentration in the soil solution isincreased. Al3+ inhibits root elongation and reducesthe capacity of Al-sensitive genotypes to take upnutrients and water for productive growth. Introduc-tion of Al-resistance genes alone has been shown toincrease efficiency of P uptake in barley (Hordeumvulgare L.) due primarily to improved root growthand soil exploration (Delhaize et al. 2009). However,the best yield outcomes are achieved when limeapplications are combined with the use of Al-resistantcultivars to overcome constraints associated withsurface and subsurface soil acidity in cropping (Scottet al. 1997) and pasture systems (Scott et al. 2000).This is most probably because soil acidity inhibits theroot-hair and rhizosheath development irrespective oftheir Al-resistance status (Haling et al. 2010), and thiscontinues to affect the plant's ability to capture soilnutrients under acid soil conditions.
Root diseases also impact directly on the efficiencywith which a plant can capture nutrients. This isdemonstrated by the large increase in wheat yields forAustralia that accompanied the introduction of canolaand improved break crops (Kirkegaard et al. 1997;Angus 2001). Breaking root disease cycles allowedyield responses to N-fertilisers that had previously onlygiven irregular benefits (Passioura 2002). Persistentand widespread root diseases still plague annual crops(Harvey et al. 2001; 2008) and are endemic inperennial farming systems where break crops are nota practical option (Barbetti et al. 2007). Resolvingthese issues will also improve P-use efficiency.
Many other approaches to increasing productivityper hectare will improve P-use efficiency. New grasscultivars that tolerate high grazing pressure andenable higher stocking rates per hectare (Culvenor etal. 2009) or improved animal genetics (e.g. better feedconversion efficiencies; Arthur et al. 2004; Hegarty et
al. 2007) are examples of innovative options for yieldimprovement, at least one step removed from soilfertility management, that will nevertheless improveP-use efficiency.
Targeted use of P-fertilisers
There are some obvious consequences for efficientagronomic practice revealed by considering typicalproduction responses to fertiliser application. WhenP-fertilisers are applied to nutrient-poor soil, plantyield increases to a plateau where P no longerdetermines the yield (e.g. Fig. 5). Accumulation ofsparingly-available phosphate, losses due to leaching,erosion and runoff, and accumulations due to unevendistribution of excreta will continue to increase at soilfertility levels beyond critical P, because their rates ofaccumulation or loss are P-concentration dependent.As there are no additional yield or financial benefitsto be achieved at soil P-fertility levels in excess ofcritical P, there is no justification for operatingfarming systems at P levels greater than the criticallevel appropriate to the crop-soil system.
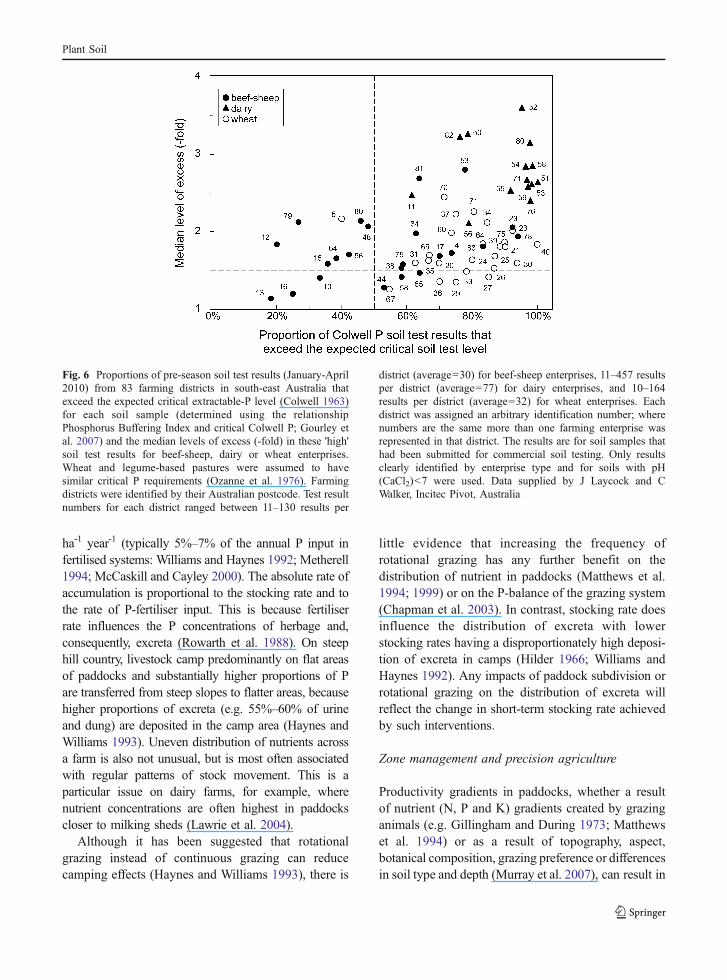
Critical P soil test values for various crop-soilsystems have been reviewed and revised recently forsouthern Australia (Moody 2007; Gourley et al.2007). These values can be used to demonstrate howwell farmer practice aligns with ideal soil P manage-ment. The fertility status of soil samples submitted forcommercial soil testing by farmers in south-easternAustralia were examined (Fig. 6). This was animperfect sample of agricultural soils, because soiltesting is used by only 40% of southern Australianfarms (ABS 2007), but it is presumed to reflect thesituation for farmers highly motivated to use P-fertilisers. In addition, soil test values do not indicatethe management reactions that they may evoke. Forexample, a high soil test value cannot always beregarded as indicating excessive fertiliser use if thereaction to it is to moderate P-fertiliser inputs. Forthese reasons we allowed relatively generous marginswhen interpreting the information in Fig. 6, and havefocussed on soil test values that are more than 1.5-fold above the expected critical value, or where morethan 50% of samples from a district are well abovethe expected critical P value. Thus, all test averagesbelow or to the left of the dashed lines are assumed toindicate acceptable practice because “high” tests areeither close to the ideal for soil fertility management
Plant Soil

or are in the minority of test results. Although onlythree farming enterprise types were examined, thedata indicated that industry-related cultures may beinfluencing fertiliser practice. For example, soil testsfrom all dairy locations showed high proportions ofsoils testing in the excessive soil P-fertility range, aconclusion supported by other studies (Lawrie et al.2004; Burkitt and Coad 2006). Recently, Burkitt et al.(2010) have challenged the trend to excessivelyelevated soil P concentrations by demonstrating thatlower P inputs and short-term cessation of P-fertiliserapplications can be viable options when soil fertilityis in excess of critical P. The analysis of soil testresults from all three enterprise types (Fig. 6) indi-cates that there is room to reduce P-fertiliser use (and,consequently, unnecessary accumulation of P in soil)on many farms represented by this subsample ofagricultural soils. This can be achieved by usingexisting technology to promote a more targetedapproach to P-fertiliser use (e.g. Simpson et al. 2009).
Uneven dispersal of excreta
It is long established that nutrient cycling by grazinganimals is important to the overall availability ofnutrients and productivity of pasture-based systems(Haynes and Williams 1993). However, the tendencyof livestock to deposit excreta disproportionately in"camps", under shade, or close to water and feedingpoints is also a component of poor P-balanceefficiency in grazing enterprises. On flat and lowslope areas, 25%–47% of sheep dung, for example,may be deposited in only 5%–15% of the total area ofa paddock (Hilder 1966; Williams and Haynes 1992).A proportion of the P deposited in camps is returnedto the rest of the paddock, but over long periods oftime the uneven dispersal of P can lead to significantaccumulation of P in a small area (Williams and Haynes1992) effectively rendering this P unavailable forpasture growth. Where P audits of paddocks have beenconducted, the accumulations are of the order 1–2 kg P
Fig. 5 Yield responses of asilver grass (mixed swardof Vulpia myuros and V.bromoides) and subterra-nean clover (Trifoliumsubterraneum) to applica-tions of phosphate in aglasshouse experiment(after Hill et al. 2005)and b grass-dominant(nitrogen-fed) andsubterranean clover-richpasture at Bookham, NewSouth Wales, Australiashowing critical P levels(corresponding to 90%of yield maximum) foreach response function(arrow) (Simpson et al.,unpublished)
Plant Soil
ha-1 year-1 (typically 5%–7% of the annual P input infertilised systems: Williams and Haynes 1992; Metherell1994; McCaskill and Cayley 2000). The absolute rate ofaccumulation is proportional to the stocking rate and tothe rate of P-fertiliser input. This is because fertiliserrate influences the P concentrations of herbage and,consequently, excreta (Rowarth et al. 1988). On steephill country, livestock camp predominantly on flat areasof paddocks and substantially higher proportions of Pare transferred from steep slopes to flatter areas, becausehigher proportions of excreta (e.g. 55%–60% of urineand dung) are deposited in the camp area (Haynes andWilliams 1993). Uneven distribution of nutrients acrossa farm is also not unusual, but is most often associatedwith regular patterns of stock movement. This is aparticular issue on dairy farms, for example, wherenutrient concentrations are often highest in paddockscloser to milking sheds (Lawrie et al. 2004).
Although it has been suggested that rotationalgrazing instead of continuous grazing can reducecamping effects (Haynes and Williams 1993), there is
little evidence that increasing the frequency ofrotational grazing has any further benefit on thedistribution of nutrient in paddocks (Matthews et al.1994; 1999) or on the P-balance of the grazing system(Chapman et al. 2003). In contrast, stocking rate doesinfluence the distribution of excreta with lowerstocking rates having a disproportionately high deposi-tion of excreta in camps (Hilder 1966; Williams andHaynes 1992). Any impacts of paddock subdivision orrotational grazing on the distribution of excreta willreflect the change in short-term stocking rate achievedby such interventions.
Zone management and precision agriculture
Productivity gradients in paddocks, whether a resultof nutrient (N, P and K) gradients created by grazinganimals (e.g. Gillingham and During 1973; Matthewset al. 1994) or as a result of topography, aspect,botanical composition, grazing preference or differencesin soil type and depth (Murray et al. 2007), can result in
Fig. 6 Proportions of pre-season soil test results (January-April2010) from 83 farming districts in south-east Australia thatexceed the expected critical extractable-P level (Colwell 1963)for each soil sample (determined using the relationshipPhosphorus Buffering Index and critical Colwell P; Gourley etal. 2007) and the median levels of excess (-fold) in these 'high'soil test results for beef-sheep, dairy or wheat enterprises.Wheat and legume-based pastures were assumed to havesimilar critical P requirements (Ozanne et al. 1976). Farmingdistricts were identified by their Australian postcode. Test resultnumbers for each district ranged between 11–130 results per
district (average=30) for beef-sheep enterprises, 11–457 resultsper district (average=77) for dairy enterprises, and 10–164results per district (average=32) for wheat enterprises. Eachdistrict was assigned an arbitrary identification number; wherenumbers are the same more than one farming enterprise wasrepresented in that district. The results are for soil samples thathad been submitted for commercial soil testing. Only resultsclearly identified by enterprise type and for soils with pH(CaCl2)<7 were used. Data supplied by J Laycock and CWalker, Incitec Pivot, Australia
Plant Soil

ineffective use of P when it is applied uniformly acrossthe landscape. The situation is analogous to unevenyield distributions in cropping paddocks which, whenmanaged using variable rate technology, can signifi-cantly improve the net profitability of the crop (e.g.Passioura 2002). Hackney (2009) has shown that as P-fertiliser prices increase, differential fertiliser applica-tions combined with grazing management to improvepasture utilisation, will increasingly deliver economicbenefits from large paddocks that have unevenproductivity (Fig. 7). Whilst yield mapping of cropsis now an accessible technology and its use isincreasing rapidly, it is more difficult in pasture-basedsystems to assess the existence of productivity gra-dients. Where large and easily identified differences innutrient requirement can be identified (such as campareas) differential fertilisation is easy and common-place (e.g. Gillingham and During 1973). However,more complex or fragmented productivity patterns cannow also be determined using ‘passive’ or ‘active’ground level, airborne or spaceborne canopy reflec-tance sensing devices with or without pasture model-ling backup (Hill et al. 1999; Murray et al. 2007;Trotter et al. 2010). The resolution being achieved withthese technologies now far exceeds that needed forzone management in pastures (e.g. Berni et al. 2009).When combined with GPS technology, productivitymaps should enable variable rate fertiliser delivery ingrazed landscapes. However, because the plant-available nutrient status of soils cannot be mapped
remotely, the spatial mapping of nutrient gradients (e.g.Kozar et al. 2002) to underpin variable fertiliser ratedecisions is an expensive and rate-limiting step indevelopment of this technology.
Low-P farming systems
The agronomic interventions mentioned so far serveto improve current practice by eliminating waste orunguided use of P-fertilisers. However, these practiceswill still only bring farming systems up to the levelsof P-balance efficiency currently being achieved inthe most productive agricultural enterprises, with Pexport being in the range of 15% (sheep grazing)–50%(cropping) of P input (McLaughlin et al. 1992; Weaverand Wong 2011). The development of farming systemsthat are productive at low soil-P concentrations is astrategy likely to provide both environmental (lowerP-losses) and efficiency (less P-accumulation) benefits.
Impact of prior fertiliser use
It has been demonstrated that previous applications ofphosphate lower the P-sorption capacity of soil(Barrow et al. 1998; Bolland and Allen 2003; Burkittet al. 2008) and increase the availability of subsequentfertiliser applications (Fig. 8; Bolland and Baker1998). This phenomenon may have immediate sig-nificance for enterprises where there has been a longhistory of fertiliser use or over-investment in soil P
Fig. 7 Anticipated net in-come from sheep grazing ahilly, 30 ha grassland atBarraga, New South Wales,Australia which had a frag-mented productivity distribu-tion due to differences inslope, aspect, soil chemistryand soil depth. Treatmentswere: superphosphate fertil-iser applied uniformly (openbars), fertiliser applieduniformly with fencing tomanage grazing patterns(hatched bars), or fertiliserapplied differentially accord-ing to nutrient responsive-ness and fenced to managegrazing patterns (closedbars) (from Hackney 2009;used with permission)
Plant Soil

fertility. However, for many soils the amounts ofphosphate that need to have been applied previouslyto achieve a measurable improvement in the currentfertiliser requirement of a crop will be relatively large.For the example shown in Fig. 8, up to 599 kg P ha-1
had been applied in a single application 20 yearspreviously to a low P-sorbing soil (phosphorusretention index=35, Bolland and Baker 1998; Bollandand Allen 2003). It is not easy to predict whetherapplying smaller amounts of P regularly would elicitthe same change in sorption capacity (Barrow et al.1998). However, at typical rates of P application, itwould have taken up to 54 years of continuousfertiliser application to reach this level of P investmentin this soil. The quantities of P-investment required tounderpin large changes in P buffering capacity meanthat this does not appear to be a management path thatwill achieve a rapid improvement in P-balanceefficiency.
Biologically-fixed N versus fertiliser N
Legume-based pastures underpin the productivity of amajority of grazing and cropping enterprises intemperate Australia and New Zealand and are usedto a lesser extent elsewhere in the world. Inputs of Nvia biological N-fixation are economically favourable,particularly in water-limited, extensive agriculture,but tend to be less favoured in well-watered environ-ments with more intensive, high-value productionsystems (Moore 1970; White et al. 1978; Puckeridge
and French 1983). In some farming systems, fertiliser-Nhas become the preferred N source (e.g. dairying,Mundy 1996; Eckard et al. 2003). The productivity oflegume-based pastures depends on adequate nutrition ofthe legume and in many cases this translates toadequate supply of P. Forage legumes typically havethe highest P-requirements of the plants that comprise apasture system (e.g. Figs. 5 and 9; Haynes 1980; Hill etal. 2005; 2010). Figure 5b demonstrates how it wouldbe feasible to shift to N-fertilised pastures to achieve asubstantial reduction in the critical soil P fertility levelsnecessary for pasture production. The final target levelsfor soil P management would be lower and determinedby the requirements of the major grass species (e.g.Fig. 9). A change such as this has already occurred inthe dairy industry of southern Australia which wastraditionally based on white clover (Trifolium repens L.)-perennial ryegrass (Lolium perenne L.) pastures withN-fixation estimated to provide about 70% of the Nnecessary to achieve potential production (Mundy1996). However, as N applications have increased, it isnowmore likely that white clover will comprise 10–25%of the pasture mix and provide <20 kg N ha-1 year-1
(McKenzie et al. 2003). Unfortunately, the potentialimprovements in P-use efficiency that could accompanysuch a major change in nutrient management on dairyfarms has not been appreciated, and the indications arethat soil P-fertility has often been increased well beyondwhere it needs to be for a legume-based system, letalone an N-fed, grass-dominant system (Fig. 6, Lawrieet al. 2004; Burkitt and Coad 2006).
Fig. 8 Effect of phosphate applied 20 years earlier on theresponse of wheat to newly applied phosphate. The originalrates were: 0 kg P ha-1 (circles); 86 kg ha-1 (triangles); and599 kg ha-1 (squares). Lines are fits to the Mitscherlich
equation and are extended to the horizontal axis to indicatethe amounts of P available from the original application (fromBolland and Baker 1998)
Plant Soil

The feasibility of shifting to N-fertilised pasturesto gain improvements in P-balance efficiency willrest on the future relative costs of P- and N-fertilisers and the prices obtained for commoditiesproduced in such systems. It is highly likely that N-fertiliser prices will increase as the costs of energyand compliance with potential carbon emissionsconstraints rise. In addition, the use of N-fertiliserswill bring new environmental challenges. Losses ofN from N-fertilised dairy systems, for instance,have emerged relatively quickly as a major envi-ronmental issue (Ledgard et al. 1996; Ledgard et al.1999; Eckard et al. 2003) and have been a significantissue in N-fertilised systems in Europe and NorthAmerica for many years (e.g. Bussink and Oenema1998).
Farming systems that utilise soil organic P
The cycling of organic P underpins productivity andP-availability on all farms in every year. For example,the organic P cycle is estimated to contribute ~52% ofthe P used annually for pasture growth in a grasslandproduction system in NSW, Australia (Simpson et al.2007). Although opportunities to increase the rate oforganic P cycling are not easily accommodated inmany agricultural systems, soil organic matter is oftenused strategically as a nutrient resource. In leyfarming systems, soil organic matter (organic P)levels oscillate between accumulation and mineralisa-
tion across the pasture and crop phases of farmingsystems (White et al. 1978). Soil organic matterconcentrations are increased during legume-basedpasture phases (e.g. Grace et al. 1995; Dalal et al.1995; Helyar et al. 1997) and nutrients are mobilisedfrom soil organic matter and used during crop phases,bare fallows and continuous cropping (e.g. White etal. 1978; Dalal and Mayer 1986; Dalal and Chan2001; Bünemann et al. 2006). Although the focus ofsoil organic matter management is usually transfer ofN from the pasture (legume) phase to the crop phase,P supply to the crop is also a consequence of thisform of resource management and some part of thegenerally better P-balance efficiency of cereal crops(Weaver and Wong 2011) is attributable to netmineralisation of P from soil organic matter duringthe crop phase.
Intercropping and phase farming
The ability of particular grain legumes to mobilisesparingly-available P has stimulated research toexamine the possibility that P-efficient plants maybe used to improve the availability of P from othercrops by (i) mixing P-efficient with inefficient speciesin either crops or pastures (intercropping) or (ii)utilising the residues of the P-efficient crops forsubsequent P-inefficient crops (crop rotation). Thesorts of species that have been examined in thiscontext include organic anion-exuding crops such as
Fig. 9 The relationship between critical external P requirement(amount of P applied to achieve 90% of maximum growth rate)and the relative rate of root cylinder development (a measure ofroot foraging capacity calculated as the volume of soil
contacted by the roots from root length and diameter, and roothair length data) of companion species found in manytemperate pastures in south-eastern Australia (redrawn fromHill et al. 2010)
Plant Soil

white lupin (Gardner and Boundy 1983; Hocking andRandall 2001; Cu et al. 2005), pigeon pea (Cajanuscajan (L.) Millsp., Ae et al. 1990), faba bean (Viciafaba L., Li et al. 2007) and chickpea (Cicer arietinumL.,Veneklaas et al. 2003), which are consideredcapable of accessing sparingly-available phosphate,or species thought capable of mobilising P fromorganic sources, e.g. chickpea (Li et al. 2003; Li et al.2004) and cowpea (Vigna unguiculata L., Makoi et al.2010). The principle that P-mobilising species canenhance growth and P uptake by cereals whenintercropped or rotated in P-deficient soil is wellestablished. However, the majority of evidence comesfrom pot experiments where the crops were not grownto maturity and periods between ‘rotations’ wereshort. Enhanced growth and P uptake were demon-strated for wheat in pots intercropped with chickpea(Li et al. 2003), lupin (Kamh et al. 1999; Cu et al.2005) and faba bean (Song et al. 2007), or byexploiting residues of white lupin and pigeon pea(Hocking and Randall 2001), faba bean and lupin(Nuruzzaman et al. 2005) as if in a rotation. Otherevidence of positive responses by cereals in potexperiments include maize (Zea mays L., Li et al.2004) and barley (Gunes et al. 2007) intercroppedwith chickpea, and maize rotated after groundnut(Arachis hypogaea L., El Dessougi et al. 2003).
Horst et al. (2001) considered that of the twomanagement options, intercropping should lead tomost efficient transfer of P, because the opportunityfor re-sorption of mobilised phosphate would be less.However, field results have been mixed. Clearevidence that cereals can gain P that would otherwisenot have been accessible has been reported (Gardnerand Boundy 1983; Li et al. 2007), but in many casesthe component species have poor yields because ofinterspecific competition (Gardner and Boundy 1983;Härdter and Horst 1991; Härdter et al. 2008). In somecases, beneficial outcomes have been attributed tofactors other than P-transfer, such as improved Nnutrition and reduced allelopathy (Horst and Härdter1994).
Intercropping is used widely in Africa and Asia,but its application for P nutrition benefits is notobvious in broadacre, highly mechanised croppingsystems. Intercropping of lucerne (Medicago sativa L.)and wheat (Harris et al. 2008), and of summer-activeperennial native grasses and wheat (Millar andBadgery 2009) have both received renewed interest
and, although feasible, crop yields are nearly alwayssubject to competitive interference (with the possibleexception of yields in wet seasons) (Harris et al. 2007a;2007b). Henry et al. (2010) have examined the impactof ‘intercropping’ different lines of the same species(i.e. ‘multilines’ of common bean, Phaseolus vulgarisL) which differed in root architecture as a way ofreducing the agronomic challenges of intercroppingand achieving improved uptake of P and water.However, they found that root growth in competitionvaried with both soil treatment and genotype mix. Themultilines did not suffer yield penalties but, againstexpectations, the root architecture differences wereoften not sustained and differences in P or wateruptake, and yield advantages were only observed in afew cases.
Using phase farming to exploit the residues of P-efficient grain legumes for subsequent cereal crop(s)is considered likely to hold more promise (Hockingand Randall 2001; Richardson et al. 2009). However,it is not clear whether P extracted by plants in onephase of a rotation will be transferred successfully toa subsequent crop under field conditions. Positiveresponses have been reported from rotating maizeafter high P-efficiency genotypes of cowpea andsoybean (Glycine max (L.) Merr.), but not after lessP-efficient genotypes (Jemo et al. 2006). However,gains to maize P nutrition following P-efficient grainlegumes in African studies were small and notconsidered sufficient to substitute for fertiliser appli-cation, particularly on high P-fixing soils (Horst et al.2001; Kamh et al. 2002).
Research is required to extend the promisingglasshouse experiments to the field where crops aregrown to maturity and translocation of P to the grainoccurs. It is necessary to determine the size of anypotential P nutrition benefit under practical farmingconditions and how long the residual P benefit fromorganic anion-exuding crops can persist in the field.Limited work to date has shown that organic anionexudation and P uptake by chickpea and lupin wasvariable across a range of low P soils from WesternAustralia (Veneklaas et al. 2003). In another study andcontrary to expectation, faba bean and chickpeagrown in other low-P soils did not mobilise P fromsparingly-soluble soil pools, pointing to a need for abetter understanding of how and when these cropsgain access to sparingly-available P (Rose et al.2010a, b).
Plant Soil

Pasture and crop varieties with low P requirements
Plants with low critical P requirements have value infarming systems that are operated with widelycontrasting soil P fertility regimes. At one end of thespectrum, plants with low P requirements can poten-tially lift production on soils where P fertility is lowand P-fertiliser use is minimal, either through choice(e.g. low-input farms, Cornish 2009), socio-economiccircumstance (e.g. subsistence agriculture Lynch2007; Sánchez 2010), or because other factors arethe primary constraint on the productivity of thefarming system (e.g. low-rainfall farming, Pang et al.2010b; extensive grazing systems, McIvor et al.2011). By contrast, the P-balance efficiency offertilised farming systems can potentially be improvedif it is possible to maintain soils at lower plant-availableP concentrations (e.g. Simpson et al. 2010) by usingproductive plants with low critical P requirements.
The agronomic potential of Australian nativeherbaceous legumes has been examined in recentyears in an effort to harness their natural adaptation tolow-P environments prone to drought stress (Table 2;Ryan et al. 2008; Dear and Ewing 2008; Ryan et al.2009; Suriyagoda et al. 2010; Bennett et al. 2011; Bellet al. 2011). Whilst a number of promising specieshave been identified, the work showed that nativespecies had few consistent productivity advantagesover exotic species in low-P soil and there was noadvantage in focusing solely on native plants adaptedto low-P environments.
Productive species with low critical P requirementshave also been identified for higher rainfall environ-ments. Numerous pasture grasses fit this category ofplants, including most of the productive, volunteerand cultivated grasses used in temperate grasslands (e.g.Hill et al. 2005). Amongst the grasses are a few specieswith exceptionally low critical P requirements (e.g.ryegrass (Lolium rigidum Gaud.), silver grass (Vulpiabromoides, V. myuros), Hill et al. 2010), grasses notedfor vigorous growth in acid, infertile soils (cocksfoot(Dactylis glomerata L.), Lolicato and Rumball 1994)and some native perennials with the capacity to capturephosphate from less soluble sources (e.g. Austro-danthonia spp., Barrett and Gifford (1999); Austrostipaspp., Marschner et al. (2006)). Unfortunately, P-efficient grasses do not have any impact on the poorP-balance efficiency of legume-based temperate pas-ture systems because they are grown in association
with legumes (e.g. subterranean clover (Trifoliumsubterraneum L.) and white clover (T. repens L.)) thathave coarse roots, short root hairs and are relativelyinefficient with respect to P acquisition (Fig. 9; Ozanneet al. 1969; 1976; Haynes 1980; Hill et al. 2006;2010). The pastures are fertilised to meet the Prequirements of the legume, because N-fixation bythe legume drives overall productivity. It will benecessary to find legumes with lower critical Prequirements to improve the P-balance efficiency ofthese pasture systems.
Interspecific differences exist in the critical Prequirements of some of the keystone temperatepasture legumes. For example, Medicago polymorphaL. has a higher critical P requirement than T.subterraneum, which has a higher requirement thanOrnithopus compressus L. (Paynter 1990; Bollandand Paynter 1992; Paynter 1992). Differences in the Prequirements of some annual medic species (e.g. M.truncatula Gaertn, M. murex Willd, M. polymorpha;Bolland 1997) and for some annual clover species (e.g.T. subterraneum, T. incarnatum L, T. hirtum All;McKell et al. 1982) have also been reported. However,the results of these and other studies are sufficientlyfragmented that it is difficult to rank species reliablyaccording to their critical P requirements. A number ofalternative pasture legume species have been releasedas cultivars in the last 15 years in response to the needfor adaptations to difficult soils and other nicheenvironments, higher levels of hardseededness, deeperroot systems, length of growing season and ease of seedproduction (e.g. Nichols et al. 2007). A few appear tohave promise for development of more P-efficientpasture systems (Table 2). The clearest example isyellow serradella (Ornithopus compressus L.), whichhad a critical P requirement ~55%–65% of that ofsubterranean clover (T. subterraneum) in field experi-ments in Western Australia (Paynter 1990; Bolland andPaynter 1992; Paynter 1992). However, P-fertiliserapplication rates recommended for serradella do notpresently reflect its lower critical P requirement(Paynter and Bolland 2006).
Intraspecific variation in the P requirements of keypasture legumes has been claimed, in a few cases, tobe as large as interspecific differences (e.g. Curll1983; Gartrell and Bolland 1987). However, closeexamination of the evidence does not support theseclaims. A number of reports are compromised bycomparisons of lines at differing physiological stages
Plant Soil

of development or at differing herbage yields. Never-theless, there are a few studies that indicate significantintraspecific variation in the critical P requirements ofsome pasture legumes (e.g. subterranean clovers whencompared at similar shoot weights, Jones et al. 1970;white clover populations from high- and low-P soils,Snaydon and Bradshaw 1962). In general, however,the P requirements of keystone pasture legumes andthe newer, alternative species are ill-defined and therehave been few demonstrations that pastures can behighly productive at lower soil P fertility levels whenP-efficient legumes are used.
There are significant differences in the critical Prequirements of some of the mainstream crops used intemperate cropping rotations. Comparisons of wheat,narrow-leaf lupin (L. angustifolius L.), canola (Brassicanapus L.) and the pasture legume, subterranean clover(T. subterranean L.) indicate that the critical P-requirement of the narrow leaf lupin is marginally(10%; Brennan and Bolland 2001) or considerablygreater (70%; Ozanne et al. 1976; Bolland 1992) than
the critical requirements of wheat and subterraneanclover (Ozanne et al. 1976). Wheat and subterraneanclover, in turn, have critical P requirements that are25%–60% larger than that of canola (Bolland 1997;Brennan and Bolland 2001; Brennan and Bolland2009). Because the critical P rankings of these cropsare reasonably consistent, it is likely that better(targeted) management of soil P-fertility can beachieved across cropping rotations than is practisedcurrently. However, experimental results occasionallyindicate wide fluctuations in the critical P requirementsof crops (e.g. Bolland 1992) suggesting that otherfactors such as root diseases, mycorrhizal status orenvironmental constraints may alter P-requirementsand may need to be considered when developingmanagement guidelines.
Substantial intraspecific variation in P-use efficiencyhas been reported for a number of major crops: e.g.wheat (Gahoonia and Nielsen 1996; Manske et al.2000; Osborne and Rengel 2002; Liao et al. 2008),barley (Gahoonia and Nielsen 1996; Górny and
Table 2 Some niche species recognised for reasonable productivity in low P soils and/or low rainfall environments
Species Herbage type Attribute(s) Reference
Ptilotus polystachyus(Gaudich.) F. Muell.
short-lived nativeperennial herb
shoot weights greater thanchicory (Chicorium intybus L.)in low-P soil
Ryan et al. 2009
Kennedia prorepens F.Muell.,K. prostrata R.Br., Glycinecanescens F.J.Herm
Australiannative legumes
good growth relative to lucernein low-P soil but does nottolerate high-P soil
Pang et al. 2010a; 2010b
Lotononis bainesii R.Br.)Bituminaria bituminosa(L.) C.H.Stirt. var.albo-marginata
exotic legumes good growth relative to lucernein low-P soil
Pang et al. 2010a; 2010b
Cullen australasicum(Schltdl.)
Australiannative legume
relatively tolerant of dry conditions;presently being developed aspasture species for low-P,low-rainfall cropping soils
Suriyagoda et al. 2010;Dear et al. 2007;Hayes et al. 2009
Lotus uliginosus Schkuhr exotic legume good tolerance of low-P soil Balocchi and Phillips1997; Kelman 2006
Trifolium ambiguum M.Bieb Caucasian clover good tolerance of low-P soil Virgona and Dear 1996
Biserrula pelecinus L. exotic legume deep-rooted species, adapted to acidsandy soils and claimed to toleratelow-P better than subterranean clover(T. suberraneum L.) and burr medic(M. polymorpha L.)
Howieson et al. 2000
Ornithopus compressus L. exotic legume substantially lower critical-Prequirement than subterraneanclover attributed to longer,finer roots; recommended primarilyfor acid, infertile sandy soils(Anon. 2007; Zurbo 2006)
Paynter 1990; Bollandand Paynter 1992;Paynter 1992
Plant Soil
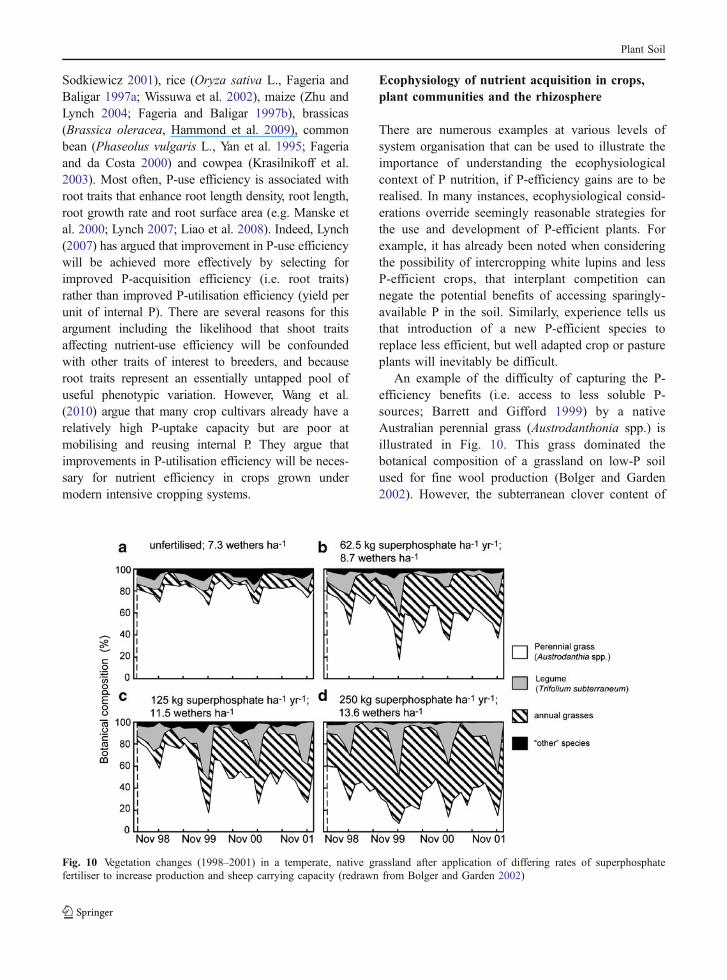
Sodkiewicz 2001), rice (Oryza sativa L., Fageria andBaligar 1997a; Wissuwa et al. 2002), maize (Zhu andLynch 2004; Fageria and Baligar 1997b), brassicas(Brassica oleracea, Hammond et al. 2009), commonbean (Phaseolus vulgaris L., Yan et al. 1995; Fageriaand da Costa 2000) and cowpea (Krasilnikoff et al.2003). Most often, P-use efficiency is associated withroot traits that enhance root length density, root length,root growth rate and root surface area (e.g. Manske etal. 2000; Lynch 2007; Liao et al. 2008). Indeed, Lynch(2007) has argued that improvement in P-use efficiencywill be achieved more effectively by selecting forimproved P-acquisition efficiency (i.e. root traits)rather than improved P-utilisation efficiency (yield perunit of internal P). There are several reasons for thisargument including the likelihood that shoot traitsaffecting nutrient-use efficiency will be confoundedwith other traits of interest to breeders, and becauseroot traits represent an essentially untapped pool ofuseful phenotypic variation. However, Wang et al.(2010) argue that many crop cultivars already have arelatively high P-uptake capacity but are poor atmobilising and reusing internal P. They argue thatimprovements in P-utilisation efficiency will be neces-sary for nutrient efficiency in crops grown undermodern intensive cropping systems.
Ecophysiology of nutrient acquisition in crops,plant communities and the rhizosphere
There are numerous examples at various levels ofsystem organisation that can be used to illustrate theimportance of understanding the ecophysiologicalcontext of P nutrition, if P-efficiency gains are to berealised. In many instances, ecophysiological consid-erations override seemingly reasonable strategies forthe use and development of P-efficient plants. Forexample, it has already been noted when consideringthe possibility of intercropping white lupins and lessP-efficient crops, that interplant competition cannegate the potential benefits of accessing sparingly-available P in the soil. Similarly, experience tells usthat introduction of a new P-efficient species toreplace less efficient, but well adapted crop or pastureplants will inevitably be difficult.
An example of the difficulty of capturing the P-efficiency benefits (i.e. access to less soluble P-sources; Barrett and Gifford 1999) by a nativeAustralian perennial grass (Austrodanthonia spp.) isillustrated in Fig. 10. This grass dominated thebotanical composition of a grassland on low-P soilused for fine wool production (Bolger and Garden2002). However, the subterranean clover content of
Fig. 10 Vegetation changes (1998–2001) in a temperate, native grassland after application of differing rates of superphosphatefertiliser to increase production and sheep carrying capacity (redrawn from Bolger and Garden 2002)
Plant Soil

the pasture (consequently legume N-inputs) andoverall production was constrained by P-deficiency.To improve stocking carrying capacity, profitability,water and land-use efficiency the system was fertilisedwith increased rates of P-fertiliser but competition fromannual species that flourish under improved soil fertility,and higher grazing pressure in the more productivesystems (e.g. Hill et al. 2005) combined to reduce theAustrodanthonia composition of the pasture andnegated much of the potential contribution to the Peconomy of the farming system that Austrodanthoniacould have made.
The importance of understanding the ecology ofnutrient acquisition also extends to the efforts thatneed to be made to improve the nutrient acquisitionefficiency of agricultural species. Plant improvementoptions fall into three broad categories: (i) strategiesfor improved soil exploration by roots, (ii) mecha-nisms that enhance the plant-availability of P fromsparingly-available phosphate and/or organic-P sour-ces, and (iii) plants that have high internal P-useefficiency. Plant improvement opportunities are dis-cussed in more detail by Richardson et al. (2011)along with an assessment of how ecophysiologicalconstraints can influence the outcome from plantbreeding.
Conclusions
It is often asserted that plants with low critical Prequirements will reduce the amount of P-fertiliserrequired in agriculture. This is true for the period inwhich a farmer is building soil fertility to a targetlevel for long-term management as the soil fertilitybuilding phase will be shorter, but the low critical Prequirement does not directly improve P-use efficiencyin the longer term. The benefit of low critical-Pplants is derived indirectly because they allow agricul-ture to be operated at lower plant-available P concen-trations. Once a target soil fertility level has beenreached (at or below the critical-P requirement of thefarming system), fertiliser use is the sum of P exportedin products, P lost to the environment and P accumu-lated through poor distribution of waste products, or insparingly-available pools of phosphate or organic-P inthe soil. Only actions that reduce losses and accumu-lations of P will reduce the P-balance efficiency ofagricultural systems.
Off-farm loss of P can be dealt with directly byimplementing best-practice measures to minimiseerosion and leaching and is often a small term in theoverall farmgate P-balance for farms on soils with amoderate to high P-buffering capacity. However, forfarms on soils with an intrinsically low P-sorptioncapacity (e.g. sands) or where P-buffering capacity islow, because of high soil-P fertility, the options forfurther reduction in the accumulation terms will below and potential losses due to leaching will be high.Inefficiencies associated with P-accumulation in stockcamps can be improved to some extent by attention tostock movements and by achieving more efficient P-fertiliser use through improved zone management.
In moderate to high P-buffering soils, phosphateaccumulation in sparingly-available pools in the soilcan be slowed by actions that reduce the concentra-tion of plant-available P at which agriculture isoperated. In the first instance, this must be addressedby eliminating the tendency for some farms to bemanaged at soil P-fertility levels well above thecritical P level appropriate to the crop-soil system.Adoption of low critical-P plants and farming systemsbased on such plants will permit agriculture to beoperated at a lower soil P concentration. The long-term benefit of low critical-P plants will be to reducethe rate at which phosphate continues to react with thesoil. There are some crop and pasture plant speciesthat fit this need, but they are limited in number, withonly a few options available for immediate triallingand implementation. It is important to improve the P-use efficiency of plant species that are keystone forfood and fibre production. Success has been achievedin a limited number of crops (e.g. Lynch 2007) butthere are many key species for which relatively littlereal progress has been made, mainly through lack ofconsistent effort. Some success has been achieved byintegrating plant species that naturally utilisesparingly-available phosphate and organic-P intoconventional farming systems (e.g. intercrops), butsuccess in other farming systems has been limited sofar by ecophysiological and agronomic practicalities.Novel agricultural varieties that mimic these speciesare needed to provide a way to reduce P accumulationin soil by enhancing extraction from, or interceptionof P destined for accumulation in the sparingly-available P pools in soil. However, it is importantthat plant improvement activities be focussed onkeystone crop and crop rotation species and, for
Plant Soil

pastures, the species (e.g. legumes) that drive theproductivity of the system.
Acknowledgements The authors thank Mark Conyers, PeterCornish, Keith Helyar and Peter Randall for critical discussionof the ideas expressed in this paper. FAS and SES wish toacknowledge the Australian Research Council, the SouthAustralian Grain Industry Trust and the Waite Research Institutefor research support and AO thanks Else Bünemann andEmmanuel Frossard for stimulating discussions. Preparation ofthe review was funded in part by Meat and Livestock AustraliaLtd and CSIRO’s National Research Flagships Program’sFlagship Collaboration Fund which aims to enhance collaborationbetween CSIRO’s Flagships, Australian universities and otherpublicly-funded research agencies.
References
Ae N, Arihara J, Okada K, Yoshihara T, Johansen C (1990)Phosphorus uptake by pigeon pea and its role in croppingsystems of the Indian subcontinent. Science 248:477–480
Amann C, Amberger A (1989) Phosphorus efficiency ofbuckwheat (Fagopyrum esculentum). Z Pflanz Bodenkunde152:181–189
Amelung W, Zech W (1999) Minimisation of organic matterdisruption during particle-size fractionation of grasslandepipedons. Geoderma 92:73–85
Angus JF (2001) Nitrogen supply and demand in Australianagriculture. Aust J Exp Agr 41:277–288
Arthur PF, Archer JA, Herd RM (2004) Feed intake andefficiency in beef cattle: overview of recent Australianresearch and challenges for the future. Aust J Exp Agr44:361–369
Australian Bureau of Statistics (2007) Natural resource manage-ment on Australian Farms, Australia 2004-2005 (reissue)4620.0. Australian Bureau of Statistics, Canberra, pp 1–68
Baldock JA, Skjemstad JO (1999) Soil organic carbon/soilorganic matter. In: Peverill KI, Sparrow LA, Reuter DJ(eds) Soil analysis: an interpretation manual. CSIRO,Collingwood, pp 159–170
Baldock JA, Skjemstad JO (2000) Role of the soil matrix andminerals in protecting natural organic materials againstbiological attack. Org Geochem 31:697–710
Baldock J, Skjemstad J, Bolger T (2007) Managing the carboncycle. In: Pasture systems: managing for a variableclimate. Proc. 22nd Annu. Conf. Grassl Soc N.S.W.Queanbeyan, 2007, pp 5–9
Balocchi OA, Phillips CJC (1997) The morphology anddevelopment of Lotus uliginosus and Trifolium subterraneumunder Pinus radiata in southern Chile. Agroforest Syst37:15–26
Barbetti MJ, You MP, Hua L, Xuanli M, Sivasithamparam K(2007) Management of root diseases of annual pasturelegumes in Mediterranean ecosystems—a case study ofsubterranean clover root diseases in the south-west ofWestern Australia. Phytopathol Mediterr 46:239–258
Barrett DJ, Gifford RM (1999) Increased C-gain by an endemicAustralian pasture grass at elevated atmospheric CO2
concentration when supplied with non-labile inorganicphosphorus. Aust J Plant Physiol 26:443–451
Barrow NJ (1969) The accumulation of soil organic matterunder pasture and its effects on soil properties. Aust J ExpAgric Anim Husb 9:437–444
Barrow NJ (1973) Relationship between a soils ability to absorbphosphate and the residual effectiveness of superphosphate.Aust J Agr Res 11:57–63
Barrow NJ (1980a) Differences among some North Americansoils in the rate of reaction with phosphate. J Environ Qual9:644–648
Barrow NJ (1980b) Differences amongst a wide-rangingcollection of soils in the rate of reaction with phosphate.Aust J Soil Res 8:215–224
Barrow NJ (1980c) Evaluation and utilisation of residualphosphorus in soils. In: Khasawneh FE, Sample EC,Kamprath EJ (eds) The role of phosphorus in agriculture.American Society of Agronomy, Madison, pp 333–339
Barrow NJ (1983a) A mechanistic model for describing thesorption and desorption of phosphate by soil. J Soil Sci34:733–750
Barrow NJ (1983b) On the reversibility of phosphate sorptionby soils. J Soil Sci 34:751–758
Barrow NJ (1999) The four laws of soil chemistry: the Leeperlecture 1998. Aust J Soil Res 37:787–829
Barrow NJ (2002) Influences of pH on a secondary effect ofphosphate reactions: the decrease in sorption of newlyadded phosphate. Aust J Soil Res 40:775–779
Barrow NJ, Carter ED (1978) A modified model for evaluatingresidual phosphate in soil. Aust J Agr Res 29:1011–21
Barrow NJ, Bolland MDA, Allen DG (1998) Effect of previousadditions of superphosphate on sorption of phosphate.Aust J Soil Res 36:359–372
Bell LW, Bennett RG, Ryan MH, Clarke CJ (2011) Thepotential of herbaceous native Australian legumes as graincrops: a review. Renew Agr Food Syst 26:72–91
Bennett RG, Ryan MH, Colmer TD, Real D (2011) Prioritisationof novel pasture species for use in water-limited agriculture: acase study of Cullen in the Western Australian wheatbelt.Genet Resour Crop Evol 58:391–407
Berni JAJ, Zarco-Tejada PJ, Suarez L, Fereres E (2009)Thermal and narrowband multispectral remote sensingfor vegetation monitoring from an unmanned aerialvehicle. IEEE Trans Geosci Remote Sens 47:722–738
Birch PB (1982) Phosphorus export from coastal plain drainageinto the Peel-Harvey estuarine system of Western Australia.Aust J Mar Freshwater Res 33:23–32
Blackwell MSA, Brookes PC (2010) Phosphorus solubizationand potential transfer to surface waters from soil microbialbiomass following drying-rewetting and freezing-thawing.Adv Agron 106:1–35
Blackwell M, Brookes P, de la Fuente-Martinez N, Murray P,Snars K, Williams J, Haygarth P (2009) Effects of soildrying and rate of re-wetting on concentrations and formsof phosphorus in leachate. Biol Fert Soils 45:635–643
Bolger TP, Garden DL (2002) Soil fertility, vegetation dynamicsand ecosystem sustainability in Australian temperategrasslands. In: Soil science: confronting new realities inthe 21st century. Trans 17th World Congr Soil Sci, August2002, Bangkok. Published on CD-ROM and on-line http://sfst.org/17WCSSCD/pages/MainIndex.htm
Plant Soil

Bolland MDA (1992) The phosphorus requirements of differentcrop species compared with wheat on lateritic soils. FertRes 32:27–36
Bolland MDA (1997) Comparative phosphorus requirements offive annual medics. J Plant Nutr 20:1029–1043
Bolland MDA, Allen DG (2003) Phosphorus sorption by sandysoils from Western Australia: effect of previously sorbed Pon P buffer capacity and single-point P sorption indices.Aust J Soil Res 41:1369–1388
Bolland MDA, Baker MJ (1998) Phosphate applied to soilincreases the effectiveness of subsequent applications ofphosphate for growing wheat shoots. Aust J Exp Agr38:865–869
Bolland MDA, Paynter BH (1992) Comparative responses ofannual pasture legume species to superphosphate applica-tions in medium and high rainfall areas of WesternAustralia. Fert Res 31:21–33
Bonkowski M (2004) Protozoa and plant growth: the microbialloop in soil revisited. New Phytol 162:617–631
Brennan RF, Bolland MDA (2001) Comparing fertiliserphosphorus requirements of canola, lupin and wheat. JPlant Nutr 24:1885–1900
Brennan RF, Bolland MDA (2009) Comparing the nitrogen andphosphorus requirements of canola and wheat for grainyield and quality. Crop Pasture Sci 60:566–577
Buczko U, Kuchenbuch RO (2007) Phosphorus indices as risk-assessment tools in the U.S.A. and Europe—a review. JPlant Nutr Soil Sci 170:445–460
Bünemann EK (2008) Enzyme additions as a tool to assess thepotential bioavailability of organically bound nutrients.Soil Biol Biochem 40:2116–2129
Bünemann E, Bossio DA, Smithson PC, Frossard E, Oberson A(2004a) Microbial community composition and substrateuse in a highly weathered soil as affected by crop rotationand P fertilization. Soil Biol Biochem 36:889–901
Bünemann E, Smithson PC, Jama B, Frossard E, Oberson A(2004b) Maize productivity and nutrient dynamics in maize-fallow rotations in western Kenya. Plant Soil 264:195–208
Bünemann EK, Steinebrunner F, Smithson PC, Frossard E,Oberson A (2004c) Phosphorus dynamics in a highlyweathered soil as revealed by isotopic labeling techniques.Soil Sci Soc Am J 68:645–1655
Bünemann EK, Heenan DP, Marschner P, McNeill AM (2006)Long-term effects of crop rotation, stubble managementand tillage on soil phosphorus dynamics. Aust J Soil Res44:611–618
Bünemann E, Marschner P, McNeill AM, McLaughlin MJ (2007)Measuring rates of gross and net mineralisation of organicphosphorus in soils. Soil Biol Biochem 39:900–913
Burkitt L, Coad J (2006) A study of the soil phosphorusconcentrations on every dairy farm in an intensivelygrazed catchment in north-west Tasmania, Australia. In:Proc 3 rd Int Symp Phosphorus Dyn Soil-Plant Contin,Brazil. pp 197–198.
Burkitt LL, Small DR, McDonald JW, Wales WJ, Jenkin ML(2007) Comparing irrigated, biodynamic and convention-ally managed dairy farms. 1. Soil and pasture properties.Aust J Exp Agr 47:479–488
Burkitt LL, Sale PWG, Gourley CJP (2008) Soil phosphorusbuffering measures should not be adjusted for currentphosphorus fertility. Aust J Soil Res 46:676–685
Burkitt LL, Donaghy DJ, Smethurst PJ (2010) Low rates ofphosphorus fertiliser applied strategically throughout thegrowing season under rain-fed conditions did not affectdry matter production of perennial ryegrass (Loliumperenne L.). Crop Pasture Sci 61:353–362
Bussink DW, Oenema O (1998) Ammonia volatilization fromdairy farming systems in temperate areas: a review. NutrCycl Agroecosys 51:19–33
Butterly CR, Bünemann EK, McNeill AM, Baldock JA,Marschner P (2009) Carbon pulses but not phosphoruspulses are related to decreases in microbial biomass duringrepeated drying and rewetting of soils. Soil Biol Biochem41:1406–1416
Cathcart JB (1980) World phosphate reserves and resources. In:Khasawneh FE, Sample EC, Kamprath EJ (eds) The role ofphosphorus in agriculture. American Society of Agronomy,Madison, pp 1–18
Chan KY, Oates A, Li GD, Conyers MK, Prangnell RJ, PoileJG, Liu DL, Barchia IM (2010) Soil carbon stocks underdifferent pastures and pasture management in the higherrainfall areas of south-eastern Australia. Aust J Soil Res48:7–15
Chapman DF, McCaskill MR, Quigley PE, Thompson AN,Grham JF, Borg D, Lamb J, Kearney G, Saul GR, ClarkSG (2003) Effects of grazing method and fertiliser inputson the productivity and sustainability of phalaris-basedpastures in Western Victoria. Aust J Exp Agr 43:785–798
Cheng W (2009) Rhizosphere priming effect: its functionalrelationships with microbial turnover, evapotranspiration,and C-N budgets. Soil Biol Biochem 41:1795–1801
Colwell JD (1963) The estimation of the phosphorus fertiliserrequirements of wheat in southern New South Wales bysoil analysis. Aust J Exp Agr 3:190–198
Cordell D, Drangert JO, White S (2009) The story ofphosphorus: global food security and food for thought.Global Environ Chang 19:292–305
Cornish PS (2009) Phosphorus management on extensiveorganic and low-input farms. Crop Pasture Sci 60:105–115
Cu S, Hutson J, Schuller KA (2005) Mixed culture of wheat(Triticum aestivum L.) with white lupin (Lupinus albus L.)improves the growth and phosphorus nutrition of thewheat. Plant Soil 272:143–151
Culvenor RA, Boschma SP, Reed KFM (2009) Response toselection for grazing tolerance in winter-active populations ofphalaris (Phalaris aquatica L.). 1. Persistence under grazingin three environments. Crop Pasture Sci 60:1097–1106
Curll MJ (1983) Intra-specific differences in phosphorus useefficiency. In: Phosphorus for pastures. Australian WoolCorporation, Specialist Workshop, Adelaide, pp 150–161
Dalal RC, Chan KY (2001) Soil organic matter in rainfedcropping systems of the Australian cereal belt. Aust J SoilRes 39:435–464
Dalal RC, Mayer RJ (1986) Long-term trends in fertility ofsoils under continuous cultivation and cereal cropping insouthern Queensland. II Total organic carbon and its rateof loss from the soil profile. Aust J Agr Res 24:281–292
Dalal RC, Strong WM, Weston EJ, Cooper JE, Lehane KJ,King AJ, Chicken CJ (1995) Sustaining productivity of aVertisol at Warra, Queensland, with fertilisers, no-tillage,or legumes. 1. Organic matter status. Aust J Exp Agr35:903–913
Plant Soil

Dann PR, Derrick JW, Dumaresq DC, Ryan MH (1996) Theresponse of organic and conventionally grown wheat tosuperphosphate and reactive phosphate rock. Aust J ExpAgr 36:71–78
Dear BS, Ewing MA (2008) The search for new pasture plantsto achieve more sustainable production systems in southernAustralia. Aust J Exp Agr 48:387–396
Dear BS, Li GD, Hayes RC, Hughes SJ, Charman N, BallardRA (2007) Cullen australasicum (syn Psoralea austral-asica): a review and some preliminary studies related to itspotential as a low rainfall perennial pasture legume.Rangeland J 29:121–132
Delhaize E, Taylor P, Hocking PJ, Simpson RJ, Ryan PR,Richardson AE (2009) Transgenic barley (Hordeumvulgare L.) expressing the wheat aluminium resistancegene (TaALMT1) shows enhanced phosphorus nutritionand grain production when grown on an acid soil. PlantBiotechnol J 7:391–400
Dinkelaker B, Romheld BV, Marschner H (1989) Citric acidexcretion and precipitation of Ca citrate in the rhizosphereof white lupin (Lupinus albus L.). Plant Cell Environ12:285–292
Donald CM (1965) The progress of Australian agriculture andthe role of pastures in environmental change. Aust J Sci27:187–198
Eckard RJ, Chen D, White RE, Chapman DF (2003) Gaseousnitrogen loss from temperate perennial grass and cloverdairy pastures in south-eastern Australia. Aust J Agr Res54:561–570
Ehlers K, Bakken LR, Frostegård Å, Frossard E, Bünemann EK(2010) Phosphorus limitation in a Ferralsol: impact onmicrobial activity and cell internal P pools. Soil BiolBiochem 42:558–566
El Dessougi H, zu Dreele A, Claassen N (2003) Growth andphosphorus uptake of maize cultivated alone, in mixedculture with other crops or after incorporation of theirresidues. J Plant Nutr Soil Sci 166:254–261
Fageria NK, Baligar VC (1997a) Phosphorus-use efficiency bycorn genotypes. J Plant Nutr 20:1267–1277
Fageria NK, Baligar VC (1997b) Upland rice genotypesevaluation for phosphorus use efficiency. J Plant Nutr20:499–509
Fageria NK, da Costa JGC (2000) Evaluation of common beangenotypes for phosphorus use efficiency. J Plant Nutr23:1145–1152
Fliessbach A, Oberholzer HR, Gunst L, Mader P (2007) Soilorganic matter and biological soil quality indicators after21 years of organic and conventional farming. Agr EcosystEnviron 118:273–284
Friesen DK, Rao IM, Thomas RJ, Oberson A, Sanz JI (1997)Phosphorus acquisition and cycling in crop and pasturesystems in low fertility tropical soils. Plant Soil 196:289–294
Gahoonia TS, Nielsen NE (1996) Variation in acquisition of soilphosphorus by wheat and barley genotypes. Plant Soil178:223–230
Gardner WK, Boundy KA (1983) The acquisition of phosphorusby Lupinus albus L. IV. The effect of interplanting wheat andwhite lupin on the growth and mineral composition of thetwo species. Plant Soil 70:391–402
Gardner WK, Barber DA, Parbery DG (1983) The acquisitionof phosphorus by Lupinus albus L. III. The probable
mechanism by which phosphorus movement in the soil/root interface is enhanced. Plant Soil 70:107–124
Gartrell JW, Bolland MDA (1987) Phosphorus nutrition ofpastures. In: Wheeler JL, Pearson CJ, Robards GE (eds)Temperate pastures: their production, use and manage-ment. Australian Wool Corporation/CSIRO, Australia, pp127–136
Gerke J (1994) Kinetics of soil phosphate desorption as affectedby citric acid. Z Pflanz Bodenkunde 157:17–22
Gijsman AJ, Oberson A, Friesen DK, Sanz JI, Thomas RJ(1997) Nutrient cycling through microbial biomass underrice-pasture rotations replacing native savanna. Soil BiolBiochem 29:1433–1441
Gillingham AG, During C (1973) Pasture production andtransfer of fertility within a long-established hill pasture.New Zeal J Exp Agr 1:227–232
Górny AG, Sodkiewicz T (2001) Genetic analysis of thenitrogen and phosphorus utilization efficiencies in maturespring barley plants. Plant Breeding 120:129–132
Gourley CJP, Melland AR, Waller RA, Awty IM, Smith AP,Peverill KI, Hannah MC (2007) Making better fertiliserdecisions for grazed pastures in Australia. VictorianGovernment Department of Primary Industries, Melbourne.www.asris.csiro.au/downloads/BFD/Making%20Better%20Fertiliser%20Decisions%20for%20Grazed%20Pastures%20in%20Australia.pdf (Accessed 24 May 2010)
Grace PR, Oades JM, Keith H, Hancock TW (1995) Trends inwheat yields and soil organic carbon in the permanentrotation trial at the Waite Agricultural Research institute,South Australia. Aust J Exp Agr 35:857–864
Grace PR, Post WM, Godwin DC, Bryceson KP, Truscott MA,Hennessy KJ (1998) Soil carbon dynamics in relation tosoil surface management and cropping systems in Australianagroecosystems. In: Lal R, Kimball JM, Follett RF, StewartBA (eds) Management of carbon sequestration in soil. CRC,Florida, pp 173–193
Grierson PF, Comerford NB, Jokela EJ (1998) Phosphorusmineralization kinetics and response of microbial phos-phorus to drying and rewetting in a Florida Spodosol. SoilBiol Biochem 30:1323–1331
Gunes A, Bagci EG, Inal A (2007) Interspecific facilitative rootinteractions and rhizosphere effects on phosphorus andiron nutrition between mixed grown chickpea and barley. JPlant Nutr 30:1455–1469
Guppy CN, McLaughlin MJ (2009) Options for increasing thebiological cycling of phosphorus in low-input and organicagricultural systems. Crop Pasture Sci 60:116–123
Hackney B (2009) Understanding and managing variation inpasture growth. PhD Thesis, University of Sydney,Australia
Haling RE, Simpson RJ, Delhaize E, Hocking PJ, RichardsonAE (2010) Effect of lime on root growth, morphology andthe rhizosheath of cereal seedlings growing in an acid soil.Plant Soil 327:199–212
Hammond JP, Broadley MR, White PJ, King GJ, Bowen HC,Hayden R, Meacham MC, Mead A, Overs T, SpracklenWP, Greenwood DJ (2009) Shoot yield drives phosphorususe efficiency in Brassica oleracea and correlates withroot architecture traits. J Exp Bot 60:1953–1968
Härdter R, Horst WJ (1991) Nitrogen and phosphorus use inmaize sole cropping and maize/cowpea mixed cropping
Plant Soil

systems on an alfisol in the northern Guinea Savanna ofGhana. Biol Fert Soils 10:267–275
Härdter R, Horst WJ, Schmidt G, Frey E (2008) Yields andland-use efficiency of maize-cowpea crop rotation incomparison to mixed and monocropping on an alfisol innorthern Ghana. J Agron Crop Sci 166:326–337
Harris RH, Clune TS, Peoples MB, Swan AD, Bellotti WD,Chen W, Norng S (2007a) The importance of in-croplucerne suppression and nitrogen for cereal companioncrops in south-eastern Australia. Field Crop Res 104:31–43
Harris RH, Hirth JR, Crawford MC, Bellotti WD, Peoples MB,Norng S (2007b) Companion crop performance in theabsence and presence of agronomic manipulation. Aust JAgr Res 58:690–701
Harris RH, Crawford MC, Bellotti WD, Peoples MB, Norng S(2008) Companion crop performance in relation to annualbiomass production, resource supply, and subsoil drying.Aust J Agr 59:1–12
Harvey PR, Butterworth PJ, Hawke BG, Pankhurst CE (2001)Genetic and pathogenic variation among cereal, medic andsub-clover isolates of Pythium irregulare. Mycol Res105:85–93
Harvey PR, Warren RA, Wakelin S (2008) The Pythium-Fusarium root disease complex—an emerging constraintto irrigated maize in southern New South Wales. Aust JExp Agr 48:367–374
Hayes RC, Li GD, Dear BS, Humphries AW, Tidd JR (2009)Persistence, productivity, nutrient composition, and aphidtolerance of Cullen spp. Crop Pasture Sci 60:1184–1192
Haygarth PM, Jarvis SC (1999) Transfer of phosphorus fromagricultural soils. Adv Agron 66:195–249
Haynes RJ (1980) Competitive aspects of the grass-legumeassociation. Adv Agron 33:227–261
Haynes RJ, Williams PH (1993) Nutrient cycling and soilfertility in the grazed pasture ecosystem. Adv Agron49:119–199
Hegarty RS, Goopy JP, Herd RM, McCorkell B (2007) Cattleselected for lower residual feed intake have reduced dailymethane production. J Anim Sci 85:1479–1486
Helyar KR (1998) Efficiency of nutrient utilization andsustaining soil fertility with particular reference to phos-phorus. Field Crop Res 56:187–195
Helyar KR, Godden DP (1976) Soil phosphate as a capitalasset. In: Blair GJ (ed) Prospects for improving efficiencyof phosphorus utilisation. Reviews in Rural Science No. 3.University of New England, Armidale, Australia, pp. 23–30
Helyar KR, Cullis BR, Furniss K, Kohn D, Taylor AC (1997)Changes in the acidity and fertility of a red earth soil underwheat-annual pasture rotations. Aust J Agr Res 48:561–586
Henry A, Rosas JC, Beaver JS, Lynch JP (2010) Multiple stressresponse and belowground competition in multilines ofcommon bean (Phaseolus vulgaris L.). Field CropResearch 117:209–218
Hilder EJ (1966) Distribution of excreta by sheep at pasture.Proc X Int Grassl Congr Helsinki, Finland, pp 977–81
Hill MJ, Donald GE, Vickery PJ, Moore AD, Donnelly JR(1999) Combining satellite data with a simulation modelto describe spatial variability in pasture growth at a farmscale. Aust J Exp Agr 39:285–300
Hill JO, Simpson RJ, Wood JT, Moore AD, Chapman DF(2005) The phosphorus and nitrogen requirements oftemperate pasture species and their influence on grasslandbotanical composition. Aust J Agr Res 56:1027–1039
Hill JO, Simpson RJ, Moore AD, Chapman DF (2006)Morphology and response of roots of pasture species tophosphorus and nitrogen nutrition. Plant Soil 286:7–19
Hill JO, Simpson RJ, Ryan MJ, Chapman DF (2010) Root hairmorphology and mycorrhizal colonisation of pasturespecies in response to phosphorus and nitrogen nutrition.Crop Pasture Sci 61:122–131
Hocking PJ, Randall PJ (2001) Better growth and phosphorusnutrition of sorghum and wheat following organic acidsecreting crops. In: Horst WJ, Schenk MK, Burkert A,Claassen N, Flessa H, Frommer WB, Goldbach H, OlfsHW, Romheld V, Sattlemacher B, Schmidhalter U, SchubertS, Wiren NV, Wittenmayer L (eds) Plant nutrition—foodsecurity and sustainability of agro-ecosystems through basicand applied research. Kluwer, Dordrecht, pp 548–549
Hocking PJ, Keerthisinghe G, Smith FW, Randall PJ (1997)Comparison of the ability of different crop species toaccess poorly-available soil phosphorus. In: Ando T, FujitaK, Mae T, Matsumoto H, Mori S, Sekiya J (eds) Plantnutrition for sustainable food production and environment.Kluwer, Dordrecht, pp 305–308
Hodgkin EP, Hamilton BH (1993) Fertilizers and eutrophicationin southwestern Australia: setting the scene. Nutr CyclAgroecosyst 36:95–103
Horst WJ, Härdter R (1994) Rotation of maize with cowpeaimproves yield and nutrient use of maize compared tomaize monocropping in an alfisol in the northern GuineaSavanna of Ghana. Plant Soil 160:171–183
Horst WJ, Kamh M, Jibrin JM, Chude VO (2001) Agronomicmeasures for increasing P availability to crops. Plant Soil237:211–223
Howieson JG, O’Hara GW, Carr SJ (2000) Changing roles forlegumes in Mediterranean agriculture: developments froman Australian perspective. Field Crop Res 65:107–122
Jackson RB, Banner JL, Jobbagy EG, Pockman WT, Wall DH(2002) Ecosystem carbon loss with woody plant invasionof grasslands. Nature 418:623–626
Jemo M, Abaidoo RC, Nolte C, Tchienkoua N, Sanginga N,Horst WJ (2006) Phosphorus benefits from grain-legumecrops to subsequent maize grown on acid soils of southernCameroon. Plant Soil 284:385–397
Joergensen R, Mäder P, Fließbach A (2010) Long-term effectsof organic farming on fungal and bacterial residues inrelation to microbial energy metabolism. Biol Fert Soils46:303–307
Johnston AE, Syers JK (2009) A new approach to assessingphosphorus use efficiency in agriculture. Better Crops withPlant Food (IPNI) 93:14–16
Jones MB, Lawler PW, Ruckman JE (1970) Differences inannual clover responses to phosphorus and sulphur. AgronJ 62:439–442
Kamh M, Horst WJ, Amer F, Mostafa H, Maier P (1999)Mobilization of soil and fertilizer phosphate by covercrops. Plant Soil 211:19–27
Kamh M, Abdou M, Chude V, Wiesler F, Horst WJ (2002)Mobilization of phosphorus contributes to positive rota-tional effects of leguminous cover crops on maize grown
Plant Soil

in soils from northern Nigeria. J Plant Nutr Soil Sci165:566–572
Kelman WM (2006) The interactive effects of phosphorus,sulphur and cultivar on the early growth and condensedtannin content of greater lotus (Lotus uliginosus) andbirdsfoot trefoil (L. corniculatus). Aust J Exp Agr 46:53–58
Kirkby CA, Kirkegaard JA, Richardson AE, Wade LJ, BlanchardC, Batten G (2011) Stable soil organic matter: a comparisonof CNPS ratios in Australian and International soils. Geo-derma. doi:10.1016/j.geoderma.2011.04.010
Kirkegaard JA, Hocking PJ, Angus JF, Howe GN, Gardner PA(1997) Comparison of canola. Indian mustard and Linolain two contrasting environments. II. Break-crop andnitrogen effects on subsequent wheat crops. Field CropRes 52:179–191
Kohn GD (1974) Superphosphate utilization in clover leyfarming. I. Effects on pasture and sheep production. Aust JAgr Res 25:525–535
Kohn GD, Osbourne GJ, Batten GD, Smith AN, Lill WJ (1977)The effect of topdressed superphosphate on changes innitrogen: carbon: sulphur: phosphorus and pH on a redearth soil during a long term grazing experiment. Aust JSoil Res 15:147–158
Kozar B, Lawrence R, Long DS (2002) Soil phosphorus andpotassium mapping using a spatial correlation modelincorporating terrain slope gradient. Precis Agr 3:407–417
Krasilnikoff G, Gahoonia T, Nielsen NE (2003) Variation inphosphorus uptake efficiency by genotypes of cowpea(Vigna unguiculata) due to differences in root and root hairlength and induced rhizosphere processes. Plant Soil251:83–91
Krull ES, Baldock JA, Skjemstadt J (2003) Importance ofmechanisms and processes of stabilisation of soil organicmatter for modelling carbon turnover. Funct Plant Biol30:207–222
Lal R (2004) Soil carbon sequestration impacts on globalclimate change and food security. Science 304:1623–1627
Lawrie RA, Havilah EJ, Eldridge SM, Dougherty WJ (2004)Phosphorus budgeting and distribution on dairy farms incoastal New South Wales. In: 'Proc SuperSoil 2004, 3 rdAust N Z Soils Conf', University of Sydney, Australia 5-9Dec 2004 (ISBN 1 920842 26 8)
Ledgard SF, Sprosen MS, Brier GJ, Nemaia EKK, Clark DA(1996) Nitrogen inputs and losses from New Zealand dairyfarmlets, as affected by nitrogen fertiliser application: yearone. Plant Soil 181:65–69
Ledgard SF, Penno JW, Sprosen MS (1999) Nitrogen inputs andlosses from clover/grass pastures grazed by dairy cows, asaffected by nitrogen fertilizer application. J Agr Sci132:215–225
Leifeld J, Reiser R, Oberholzer HR (2009) Consequences ofconventional versus organic farming on soil carbon: resultsfrom a 27-year field experiment. Agron J 101:1204–1218
Lewis DC, Clarke AL, Hall WB (1987) Accumulation of plantnutrients and changes in soil properties of sandy soilsunder fertilised pastures in south-eastern South Australia.I. Phosphorus. Aust J Soil Res 25:193–202
Li L, Tang C, Rengel Z, Zhang F (2003) Chickpea facilitatesphosphorus uptake by intercropped wheat from an organicphosphorus source. Plant Soil 248:297–303
Li SM, Li L, Zhang FS, Tang C (2004) Acid phosphatase rolein chickpea/maize intercropping. Ann Bot 94:297–303
Li L, Li S, Sun J, Zhou L, Bao X, Zhang H, Zhang F (2007)Diversity enhances agricultural productivity via rhizospherephosphorus facilitation on phosphorus-deficient soils. ProcNatl Acad Sci 104:11192–11196
Liao M, Hocking PJ, Dong B, Delhaize E, Richardson AE,Ryan PR (2008) Genotypic variation in phosphorusefficiency among wheat genotypes grown on two con-trasting Australian soils. Aust J Agr Res 59:157–166
Lolicato S, Rumball W (1994) Past and present improvement ofcocskfoot (Dactylis glomerata L.) in Australia and NewZealand. New Zeal J Agr Res 37:379–390
Lynch JP (2007) Roots of the second green revolution. Aust JBot 55:1–20
Ma L, Ma WQ, Velthof GL, Wang FH, Quin W, Zhang FS,Oenema O (2010) Modeling nutrient flows in the foodchain of China. J Environ Qual 39:1279–1289
Macklon AES, Grayston SJ, Shand CA, Sim A, Sellars S, OrdBG (1997) Uptake and transport of phosphorus by Agrostiscapillaris seedlings from rapidly hydrolysed organic sourcesextracted from 32P-labelled bacterial cultures. Plant Soil190:163–167
Mäder P, Fließbach A, Dubois D, Gunst L, Fried P, Niggli U(2002) Soil fertility and biodiversity in organic farming.Science 296:1694–1697
Makoi JHJR, Chimphango SBM, Dakora FD (2010) Elevatedlevels of acid and alkaline phosphatase activity in rootsand rhizosphere of cowpea (Vigna unguiculata L. Walp.)genotypes in mixed culture and at different densities withsorghum (Sorghum bicolour L.). Crop Pasture Sci 61:279–286
Manske GGB, Ortiz-Monasterio JI, Van Ginkel M, GonzalezRM, Rajaram S, Molina E, Vlek PLG (2000) Traitsassociated with improved P-uptake efficiency in CIMMYT’ssemidwarf spring bread wheat grown on acid Andisols inMexico. Plant Soil 221:189–204
Marschner P, Solaiman Z, Rengel Z (2006) Rhizosphereproperties of Poaceae genotypes under P-limiting con-ditions. Plant Soil 283:11–24
Mathers NJ, Nash DM, Gangaiya P (2007) Nitrogen andphosphorus exports from high rainfall zone cropping inAustralia: issues and opportunities for research. J EnvironQual 36:1551–1562
Matthews BW, Tritschler JP, Carpenter JR, Sollenberger LE(1994) Soil macronutrient distribution in rotationallystocked Kikuyugrass paddocks with short and longgrazing periods. Commun Soil Sci Plan 30:557–571
Matthews BW, Sollenberger LE, Nair VD, Staples CR (1999)Impact of grazing management on soil nitrogen, phosphorus,potassium and sulfur distribution. J Environ Qual 23:1006–1013
McCaskill MR, Cayley JWD (2000) Soil audit of a long-termphosphate experiment in South-Western Victoria: totalphosphorus, sulfur, nitrogen and major cations. Aust JAgr Res 51:737–748
McCulloch M, Pailles C, Moody P, Martin CE (2003) Tracing thesource of sediment and phosphorus into the Great BarrierReef lagoon. Earth Planetary Sci Letters 210:249–258
McIvor J, Guppy C, Probert M (2011) Phosphorus requirementsof tropical grazing systems: the northern Australian
Plant Soil

experience. Plant Soil (submitted to Special Issue S43—Phosphorus)
McKell CM, Wilson AW, Williams WA (1982) Effect oftemperature on phosphorus utilization by native andintroduced legumes. Agron J 54:109–113
McKenzie FR, Jacobs JL, Riffkin P, Kearney G, McCaskill M(2003) Long-term effects of multiple applications ofnitrogen fertiliser on grazed dryland perennial ryegrass/whiteclover dairy pastures in south-west Victoria. 1. Nitrogenfixation by white clover. Aust J Agr Res 54:461–469
McLaughlin MJ, Alston AM, Martin JK (1988) Phosphoruscycling in wheat-pasture rotations II the role of themicrobial biomass in phosphorus cycling. Aust J SoilRes 26:333–342
McLaughlin MJ, Baker TG, James TR, Rundle JA (1990)Distribution and forms of phosphorus and aluminum inacidic topsoils under pastures in south-eastern Australia.Aust J Soil Res 28:371–385
McLaughlin MJ, Fillery IR, Till AR (1992) Operation of thephosphorus, sulphur and nitrogen cycles. In: Australia’srenewable resources: sustainability and global change. BurRural Resour Proc 14:67–116
McLaughlin MJ, McBeath TM, Smernik R, Stacey SP, Ajiboye S.Guppy, C (2011) The chemical nature of P-accumulation inagricultural soils - implications for fertiliser management anddesign: an Australian perspective. (accepted and in press forSpecial Issue S43—Phosphorus)
Melland AR, McCaskill MR, White RE, Chapman DF (2008)Loss of phosphorus and nitrogen in runoff and subsurfacedrainage from high and low input pastures grazed bysheep in Australia. Aust J Soil Res 46:161–172
Metherell, AK (1994) A model for phosphate fertiliser require-ments for pastures—incorporating dynamics and economics.In: Currie LD, Loganathon P (eds) The efficient use offertilisers in a changing environment: reconciling productivityand sustainability. Occasional Report No. 7. Fertiliser andLime Research Centre, Massey University, New Zealand, pp18-37
Millar GD, Badgery WB (2009) Pasture cropping: a newapproach to integrate crop and livestock farming systems.Anim Prod Sci 49:777–787
Mokany K, Moore AD, Graham P, Simpson RJ (2010) Optimalmanagement of fertiliser and stocking rate in temperategrazing systems. Anim Prod Sci 50:6–16
Moody PW (2007) Interpretation of a single-point P bufferingindex for adjusting critical levels of the Colwell soil P test.Aust J Soil Res 45:55–62
Moore RM (1970) Australian grasslands. Australian NationalUniversity Press, Canberra
Mundy G (1996) Nitrogen fertiliser—what are the upper limits?In: Proc Large Herds Conf, Aust. Rebecca LedgerProduction Centre, McMillan College, University ofMelbourne, Australia, pp182–193
Murray IR, Yule IJ, Gillingham AG (2007) Developing variablerate application technology: modelling annual pastureproduction on hill country. New Zeal J Agr Res 50:41–52
Nash DM, Haliwell DJ (1999) Fertilisers and phosphorus loss fromproductive grazing systems. Aust J Soil Res 37:403–429
Nichols PGH, Loi A, Nutt BJ, Evans PM, Craig AD, PengellyBC, Dear BS, Lloyd DL, Revell CK, Nair RM, EwingMA, Howieson JG, Auricht GA, Howie JH, Sandral GA,
Carr SJ, de Koning CT, Hackney BF, Crocker GJ,Snowball R, Hughes SJ, Hall EJ, Foster KJ, Skinner PW,Barbetti MJ, You MP (2007) New annual and short-livedperennial pasture legumes for Australian agriculture—15 years of revolution. Field Crop Res 104:10–23
Nuruzzaman M, Lambers H, Bolland MDA, Veneklaas EJ(2005) Phosphorus benefits of different legume crops tosubsequent wheat grown in different soils of WesternAustralia. Plant Soil 271:175–187
Oberson A, Joner EJ (2005) Microbial turnover of phosphorusin soil. In: Turner BL, Frossard E, Baldwin DS (eds)Organic phosphorus in the environment. CABI, Wallingford,pp 133–164
Oberson A, Fardeau JC, Besson JM, Sticher H (1993) Soil-phosphorus dynamics in cropping systems managedaccording to conventional and biological agriculturalmethods. Biol Fert Soils 16:111–117
Oberson A, Friesen DK, Tiessen H, Morel C, Stahel W (1999)Phosphorus status and cycling in native savanna andimproved pastures on an acid low-P Colombian Oxisol.Nutr Cycl Agroecosys 55:77–88
Oberson A, Friesen DK, Rao IM, Bühler S, Frossard E (2001)Phosphorus transformations in an Oxisol under contrastingland- use systems: the role of the soil microbial biomass.Plant Soil 237:197–210
Oberson A, Tagmann HU, Langmeier M, Dubois D, Mäder P,Frossard E (2010) Fresh and residual phosphorus uptakeby ryegrass from soils with different fertilization histories.Plant Soil 334:391–407
Oehl F, Oberson A, Probst M, Andreas Fliessbach A, Roth H-R,Frossard E (2001) Kinetics of microbial phosphorusuptake in cultivated soils. Biol Fert Soils 34:31–41
Oehl F, Oberson A, Tagmann HU, Besson JM, Dubois D,Mäder P, Roth H-R, Frossard E (2002) Phosphorus budgetand phosphorus availability in soils under organic andconventional farming. Nutr Cycl Agroecosys 62:25–35
Oehl F, Frossard E, Fliessbach A, Dubois D, Oberson A (2004)Basal organic phosphorus mineralization in soils underdifferent farming systems. Soil Biol Biochem 36:667–675
Olander LP, Vitousek PM (2004) Biological and geochemicalsinks for phosphorus in soil from a wet tropical forest.Ecosystems 7:404–419
Olsen SR, Cole C, Watanabe CV, Dean LA (1954) Estimation ofavailable phosphorus in soils by extraction with sodiumbicarbonate. USDA Circular No. 939
Oniani OG, Chater M, Mattingly GEG (1973) Some effects offertilizers and farmyard manure on the organic phosphorusin soils. J Soil Sci 24:1–9
Osborne LD, Rengel Z (2002) Screening cereals for genotypicvariation in efficiency of phosphorus uptake and utilisation.Aust J Agr Res 53:295–303
Otani T, Ae N (1996) Sensitivity of phosphorus uptake to changesin root length and soil volume. Agron J 88:371–375
Ozanne PG, Kirton DJ, Shaw TC (1961) The loss ofphosphorus from sandy soils. Aust J Agr Res 11:927–924
Ozanne PG, Keay J, Biddiscombe EF (1969) The comparativeapplied phosphate requirements of eight annual pasturespecies. Aust J Agr Res 20:809–818
Ozanne PG, Howes KMW, Petch A (1976) The comparativephosphate requirements of four annual pastures and twocrops. Aust J Agr Res 27:479–488
Plant Soil

Pang J, Ryan MH, Tibbett M, Cawthray GR, Siddique KHM,Bolland MDA, Denton MD, Lambers H (2010a) Variationin morphological and physiological parameters in herba-ceous perennial legumes in response to phosphorus supply.Plant Soil 331:241–255
Pang J, Tibbett M, Denton MD, Lambers H, Siddique KHM,Bolland MDA, Revell CK, Ryan MH (2010b) Variation inseedling growth of 11 perennial legumes in response tophosphorus supply. Plant Soil 328:133–143
Passioura JB (2002) Environmental biology and crop improve-ment. Funct Plant Biol 29:537–546
Paynter BH (1990) Comparative phosphate requirements ofyellow serradella (Ornithopus compressus), burr medic(Medicago polymorpha var. brevispina) and subterraneanclover (Trifolium subterraneum). Aust J Exp Agr 30:507–514
Paynter BH (1992) Comparison of the phosphate requirementsof burr medic and yellow serradella with subterraneanclover in the low rainfall wheatbelt of Western Australia.Aust J Exp Agr 32:1077–1086
Paynter B, Bolland M (2006) Phosphorus deficiency in burrmedic and yellow serradella. Farmnote 93/96, Depart FoodAgric West Aust
Perrott KW, Sarathchandra SU, Dow BW (1992) Seasonal andfertilizer effects on the organic cycle and microbialbiomass in a hill country soil under pasture. Aust J SoilRes 30:383–394
Peverill KI, Sparrow LA, Reuter DJ (1999) Soil analysis: aninterpretation manual. CSIRO, Melbourne
Pierzynski GM, McDowell RW, Sims JT (2005) Chemistry,cycling and potential movement of inorganic phosphorusin soils. In: Sims JT, Sharpley AN, Pierzynski GM,Westermann DT, Cabrera ML, Powell JM, Daniel TC,Withers PJA (eds) Phosphorus: agriculture and theenvironment. American Society of Agronomy, CropScience Society of America, Soil Science Society ofAmerica, Madison, pp 53–86
Puckeridge DW, French RJ (1983) The annual legume pasturein cereal-ley farming systems of southern Australia: areview. Agr Ecosyst Environ 9:229–267
Qiu J (2010) Phosphate fertiliser warning for China. Naturenews.doi:10.1038/news.2010.498
Richardson AE, Simpson RJ (2011) Soil microorganisms mediat-ing phosphorus availability. Plant Physiol. doi:10.1104/pp.111.175448
Richardson AE, George TS, Hens M, Simpson RJ (2005)Utilisation of soil organic phosphorus by higher plants. In:Turner BL, Frossard E, Baldwin DS (eds) Organicphosphorus in the environment. CABI, Wallingford, pp165–184
Richardson AE, Hocking PJ, Simpson RJ, George TS (2009)Plant mechanisms to optimise access to soil phosphorus.Crop Pasture Sci 60:124–143
Richardson AE, Lynch JP, Ryan PR, Delhaize E, Smith FA,Smith SE, Harvey PR, Ryan MH, Veneklaas EJ, LambersH, Oberson A, Culvenor RA, Simpson RJ (2011) Plantand microbial strategies to improve the phosphorusefficiency of agriculture. Plant Soil (submitted to SpecialIssue S43—Phosphorus)
Ridley AM, Christy BP, White RE, McLean T, Green R (2003)North-east Victoria SGS National Experiment site: water
and nutrient losses from grazing systems on contrastingsoil types and levels of inputs. Aust J Exp Agr 43:799–815
Rose TJ, Hardiputra B, Rengel Z (2010a) Wheat, canola andgrain legume access to soil phosphorus fractions differs insoils with contrasting phosphorus dynamics. Plant Soil326:159–170
Rose TJ, Pariasca-Tanaka J, Rose MJ, Fukuta Y, Wissuwa M(2010b) Genotypic variation in grain phosphorus concen-tration, and opportunities to improve P-use efficiency inrice. Field Crops Res 119:154–160
Rowarth JS, Gillingham AG, Tillman RW, Syers JK (1988)Effects of season and fertiliser rate on phosphorusconcentrations in pasture and sheep faeces in hill country.New Zeal J Agr Res 31:187–193
Russell JS (1960a) Soil fertility changes in the long-termexperimental plots at Kybybolite, South Australia. I.Changes in pH total nitrogen, organic carbon, and bulkdensity. Aust J Agr Res 11:902–926
Russell JS (1960b) Soil fertility changes in the long-termexperimental plots at Kybybolite, South Australia. II.Changes in phosphorus. Aust J Agr Res 11:926–947
Ryan MH, Bennett RG, Denton M, Hughes S, Mitchell M,Carmody B, Edmonds-Tibbett T, Nicol D, Kroiss L,Snowball R (2008) Searching for native perennial legumeswith pasture potential. Proc 14th Aust Agron Conf, Sept2008. Adelaide. www.agronomy.org.au/ (Accessed 31January 2011)
Ryan MH, Ehrenberg S, Bennett RG, Tibbett M (2009) Puttingthe P in Ptilotus: a phosphorus-accumulating herb nativeto Australia. Ann Bot 103:901–911
Sample EC, Soper RJ, Racz GJ (1980) Reactions of phosphatefertilizers in soils. In: Khasawneh FE, Sample EC,Kamprath EJ (eds) The role of phosphorus in agriculture.American Society of Agronomy, Crop Science Society ofAmerica, Soil Science Society of America, Madison, pp263–310
Sánchez PA (2010) Tripling crop yields in tropical Africa. NatGeosci 3:299–300
Scott D (2000) Sustainability of New Zealand high-countrypastures under contrasting development inputs 6. Fertiliserefficiency. New Zeal J Agr Res 43:525–532
Scott BJ, Conyers MK, Poile GJ, Cullis BR (1997) Subsurfaceacidity and liming affect yield of cereals. Aust J Agr Res48:843–854
Scott BJ, Ridley AM, Conyers MK (2000) Management of soilacidity in long-term pastures of south-eastern Australia: areview. Aust J Exp Agr 40:1173–1198
Sharpley A, Foy B, Withers P (2000) Practical and innovativemeasures for the control of agricultural phosphorus lossesto water: an overview. J Environ Qual 29:1–9
Sharpley AN, McDowell RW, Kleinman PJA (2001) Phosphorusloss from land to water: integrating agricultural andenvironmental management. Plant Soil 237:287–307
Simpson P, Langford C (1996) Managing high rainfall nativepastures on a whole farm basis. NSWAgriculture, ISBN: 07310 5778 3.
Simpson R, Richardson A, Salmon L, Graham P, McKay A,Riley I (2007) Soil biology—some good and bad impactson pasture production. In: From the Ground Up. Proc 48thAnnu Conf Grassl Soc South Aust. pp 69–80
Plant Soil

Simpson R, Graham P, Davies L, Zurcher E (2009) Five easysteps to ensure you are making money from superphos-phate. CSIRO, Industry and Investment N.S.W. Farmadvisory booklet and computer decision-support tool.39 pp, ISBN: 978 0 643 10084 8, www.mla.com.au(Accessed 31 January 2011)
Simpson RJ, Stefanski A, Marshall DJ, Moore AD, RichardsonAE (2010) The farm-gate phosphorus balance of sheepgrazing systems maintained at three contrasting soil fertilitylevels. In: Food Security from Sustainable Agriculture. Proc15th Aust Soc Agron Conf, 15-18 Nov 2010, Lincoln, NewZealand. www.agronomy.org.au/ (Accessed 31 January2011)
Six J, Conant RT, Paul EA, Paustian K (2002) Stabilizationmechanisms of soil organic matter: Implications for C-saturation of soils. Plant Soil 241:155–176
Skjemstad JO, Spouncer LR, Cowie B, Swift RS (2004)Calibration of the Rothamsted organic carbon turnovermodel (RothC ver. 26.3), using measurable soil organiccarbon pools. Aust J Soil Res 42:79–88
Smernik RJ, Dougherty WJ (2007) Identification of phytate inphosphorus-31 nuclear magnetic resonance spectra: theneed for spiking. Soil Sci Soc Am J 71:1045–1050
Snaydon RW, Bradshaw AD (1962) Differences betweennatural populations of Trifolium repens L. in response tomineral nutrients. J Exp Bo 13:422–434
Song YN, Zhang FS, Marschner P, Fan FL, Gao HM, Bao XG,Sun JH, Li L (2007) Effect of intercropping on crop yieldand chemical and microbiological properties in rhizosphereof wheat (Triticum aestivum L.), maize (Zea mays L.), andfaba bean (Vicia faba L.). Biol Fert Soils 43:565–574
Stewart WM, Hammond LL, Van Kauwenbergh SJ (2005)Phosphorus as a natural resource. In: Sims JT, SharpleyAN, Pierzynski GM, Westermann DT, Cabrera ML, PowellJM, Daniel TC, Withers PJA (eds) Phosphorus: agricultureand the environment. American Society of Agronomy,Crop Science Society of America, Soil Science Society ofAmerica, Madison, WI, USA, pp 53–86
Stewart CE, Paustian K, Conant RT, Plante AF, Six J (2007)Soil carbon saturation: concept, evidence and evaluation.Biogeochemistry 86:19–31
Suriyagoda LDB, Ryan MH, Renton M, Lambers H (2010)Multiple adaptive responses of Australian native perenniallegumes with pasture potential to grow in phosphorus- andmoisture-limited environments. Ann Bot 105:755–767
Syers JK, Johnston AE, Curtin D (2008) Efficiency of soil andfertilizer phosphorus use—reconciling changing conceptsof soil phosphorus behaviour with agronomic information.FAO Fertilizer and Plant Nutrition Bulletin 18. Food andAgriculture Organisation of the United Nations, Rome2008
Trotter MG, Lamb DW, Donald GE, Schneider DA (2010)Evaluating an active optical sensor for quantifying andmapping green herbage mass and growth in a perennialgrass pasture. Crop Pasture Sci 61:389–398
Turner BL, Haygarth PM (2001) Phosphorus solubilization inrewetted soils. Nature 411:258
Turner BL, Driessen JP, Haygarth PM, McKelvie ID (2003)Potential contribution of lysed bacterial cells to phosphorussolubilisation in two rewetted Australian pasture soils. SoilBiol Biochem 35:187–189
Turner BL, Cade-Menun BL, Condron LM, Newman S(2005) Extraction of soil organic phosphorus. Talanta66:294–306
Turner BL, Frossard E, Oberson A (2006) Enhancing phospho-rus availability in low-fertility soils. In: Uphoff N, BallAS, Fernandes E, Herren H, Husson O, Laing M, Palm C,Pretty J, Sanchez P, Sanginga N, Thies J (eds) Biologicalapproaches to sustainable soil systems. CRC, Boca Raton,pp 191–205
Ulén B, Bechmann M, Fölster J, Jarvie HP, Tunney H (2007)Agriculture as a phosphorus source for eutrophication inthe north-west European countries, Norway, Sweden,United Kingdom and Ireland: a review. Soil Use Manage-ment 23:5–15
Van Kauwenbergh SJ (2010) World phosphate rock reserves andresources. International Fertilizer Development Center,Muscle Shoals, Alabama
Veneklaas EJ, Stevens J, Cawthray GR, Turner S, Grigg AM,Lambers H (2003) Chickpea and white lupin rhizospherecarboxylates vary with soil properties and enhancephosphorus uptake. Plant Soil 248:187–197
Virgona JM, Dear BS (1996) Comparative performance ofCaucasian clover (Trifolium ambiguum cv. Monaro) after11 years under low-input conditions in south-easternAustralia. New Zeal J Agr Res 39:245–253
von Uexküll HR, Mutert E (1995) Global extent, develop-ment and economic impact of acid soils. Plant Soil171:1–15
Walker TW, Syers JK (1976) The fate of phosphorus duringpedogenesis. Geoderma 15:1–19
Wang X, Shen J, Liao H (2010) Acquisition or utilization,which is more critical for enhancing phosphorus efficiencyin modern crops? Plant Sci 179:302–306
Weaver DM, Wong MTF (2011) Phosphorus balance efficiencyand P status in crop and pasture soils with contrasting Pbuffer indices: scope for improvement. Plant Soil (submittedto Special Issue S43—Phosphorus)
Weaver DM, Ritchie GSP, Anderson GC, Deeley DM (1988)Phosphorus leaching in sandy soils. II. Laboratory studiesof the long-term effects of the phosphorus source. Aust JSoil Res 26:191–200
White DH, Elliot BR, Sharkey MJ, Reeves TG (1978)Efficiency of land-use systems involving crops andpastures. J Aust Inst Agr Sci 44:21–27
Wichern F, Mayer J, Joergensen RG, Muller T (2007) Releaseof C and N from roots of peas and oats and theiravailability to soil microorganisms. Soil Biol Biochem39:2829–2839
Williams CH, Donald CM (1957) Changes in organic matterand pH in a podzolic soil as influenced by subterraneanclover and superphosphate. Aust J Agr Res 8:179–189
Williams PH, Haynes RJ (1992) Balance sheet of phospho-rus, sulphur and potassium in a long-term grazedpasture supplied with superphosphate. Fert Res 31:51–60
Wissuwa M, Wegner J, Ae N, Yano M (2002) Substitutionmapping of Pup1: a major QTL increasing phosphorusuptake of rice from a phosphorus-deficient soil. TheoretAppl Genet 105:890–897
World Bank (2004) World development indicators. The WorldBank, Washington
Plant Soil

Yan X, Beebe SE, Lynch JP (1995) Genetic variation forphosphorus efficiency of common bean in contrasting soiltypes. II. Yield response. Crop Sci 35:1094–1099
Zhao XR, Zhong XY, Bao HJ, Li HH, Li GT, Tuo DB, Lin QM,Brookes PC (2007) Relating soil P concentrations at whichP movement occurs to soil properties in Chinese agriculturalsoils. Geoderma 142:237–244
Zhu J, Lynch JP (2004) The contribution of lateral rooting tophosphorus acquisition efficiency in maize (Zea mays L.)seedlings. Funct Plant Biol 31:949–958
Zhu Y-G, He Y-Q, Smith SE, Smith FA (2002) Buckwheat(Fagopyrum esculentum Moench) has high capacity totake up phosphorus (P) from a calcium (Ca)-bound source.Plant Soil 239:1–8
Plant Soil