spermatogenic disturbance induced by di-(2-ethylhexyl) phthalate is significantly prevented by...
TRANSCRIPT

Spermatogenic disturbance induced by di-(2-ethylhexyl)
phthalate is signi®cantly prevented by treatment with
antioxidant vitamins in the rat
MASARU ISHIHARA,* MASAHIRO ITOH, KENSAKU MIYAMOTO, SHIGERU SUNA,à YOSHIKI TAKEUCHI, IKUMASA TAKENAKA* and
FUMIHIKO JITSUNARIà
Departments of *Urology, Anatomy, and àHygiene and Public Health, Kagawa Medical
University, Kagawa, Japan
SummaryPhthalate esters, now regarded as endocrine disruptors, are widely used in the plastics
industry. In particular, di-(2-ethylhexyl) phthalate (DEHP) is produced in large
quantities, and is used in blood storage bags, catheters and haemodialysis instruments.
Previous studies have demonstrated that treatment of rats with DEHP induces testicular
atrophy with liver enlargement, although the precise nature and mechanism of the action
of DEHP on these organs remains unclear. In the present study, we produced an
experimental model of DEHP-induced spermatogenic disturbance in rats by feeding
them a DEHP-containing diet. Liver enlargement occurred in rats fed either a 1 or 2%
DEHP-containing diet. However, testicular atrophy accompanied by aspermatogenesis
was induced by feeding with the 2% but not with the 1% DEHP-containing diet. This
suggests that the critical DEHP dose for gonadotoxicity is higher than that for
hepatotoxicity. Using the 2% DEHP-dose, the effect of simultaneous administration of
antioxidant vitamins (� vitamins C and E) was next examined. It was found that the
vitamin supplementation signi®cantly prevented the testicular injury. The results suggest
that antioxidant vitamins can protect the testes from DEHP-toxicity.
Keywords: DNA ¯ow cytometry, phthalate, spermatogenesis, testicular toxicity,
vitamin C, vitamin E
IntroductionPhthalic acid esters are widely used as plasticizers in several
plastic formulations. Di-(2-ethylhexyl) phthalate (DEHP),
one of the most commonly used plasticizers, has been shown
to leach out from the ®nished plastics into the air, water and
ground, and thereby to enter various foods (Thomas et al.,
1978; Albro, 1987). DEHP also directly enters the human
system from blood storage bags, catheters and haemodialysis
instruments. In fact, a high DEHP residue concentration has
been found in the blood and tissues of patients after numerous
blood transfusions (Hillman et al., 1975; Sjoberg et al., 1985;
Faouzi et al., 1999; Manojkumar et al., 1999).
Recent interest in DEHP has been focused on its toxicity
to spermatogenesis (Oishi & Hiraga, 1983; Oishi, 1984).
Indeed, many studies have demonstrated that treatment of
laboratory animals with DEHP induces testicular atrophy
with liver enlargement (Oishi, 1989a, 1989b). Although the
mechanism underlying the testicular damage caused by
DEHP remains unclear, it was recently found that the
administration of vitamin B12 is effective in preventing
DEHP-induced spermatogenic disturbance in rats (Oishi,
1994). Like vitamin B12, vitamins A, C and E are also
essential for normal spermatogenesis (Mason, 1933, 1940;
Correspondence: Dr Masahiro Itoh, Department of Anatomy,
Kagawa Medical University, Miki-cho, Kita-gun, Kagawa
761±0793, Japan.
international journal of andrology, 23:85±94 (2000)
Ó 2000 Blackwell Science Ltd.

Coward et al., 1966; Mason & Mauer 1975; Chinoy et al.,
1986; Sapra et al., 1987; Bensoussan et al., 1998). This raises
the possibility that the administration of these vitamins
would also have some bene®cial effect on spermatogenesis in
DEHP-treated rats. It is well known that both vitamins C
and E are antioxidative ( � oxido-reductive) agents which
act synergistically as potent scavengers of free radicals and
terminators of free-radical chain reactions (Tappel, 1962;
Summer®eld & Tappel, 1984; McCay, 1985; Niki et al.,
1985; Niki, 1987; Burton & Ingold, 1989). Therefore, the
simultaneous administration of the two vitamins may have
some in¯uence on DEHP-induced spermatogenic lesions
through their cooperative antioxidant function. The aim of
the present study was to determine the histological appear-
ance of the seminiferous epithelium of rats fed a DEHP-
containing diet with or without supplementation of the two
antioxidant vitamins.
Materials and methods
AnimalsSprague-Dawley (SD) male rats (aged 4 weeks) were
purchased from Charles River (Kanagawa, Japan) and kept in
the Laboratory Animal Center of Kagawa Medical Univer-
sity for 1 week before use. They were maintained at
22±24 °C and 50±60% relative humidity with a 12 h light-
dark cycle. The approval of the Kagawa Medical University
Animal Committee was obtained for this study.
ChemicalsDEHP was purchased from Tokyo Chemical Industries
(Tokyo, Japan). The chemical purity of DEHP was found to
be >98% by gas-liquid chromatography. CE-2 diets (Clea,
Tokyo, Japan) containing 1 and 2% DEHP were prepared by
Oriental Yeast Company (Chiba, Japan). Vitamin powders
(Chocola EC) comprising ascorbic acid (� vitamin C) and
water-soluble d-a-tocopherol (� vitamin E), respectively,
were provided by Eizai (Tokyo, Japan). The vitamin
C:vitamin E ratio in the powders was 2:1.
Experimental designIn the ®rst experiments, the animals were housed in three
groups (Groups I±III) of 15 animals each, and treated daily
for 2, 4 or 6 weeks in the following manner. Group I:
control rats fed the DEHP-free diet, Group II: Experimental
rats fed the diet containing 1% DEHP, Group III:
Experimental rats fed the diet containing 2% DEHP.
In the second experiment, the effects of vitamins C and E
on DEHP-treated rats were examined. The vitamin powders
were dissolved in drinking water at doses of 3.0 and 1.5 mg/mL,
respectively. Four groups (Groups A±D) composed of ®ve
animals each and were treated daily for 2 weeks, as follows.
Group A: negative control rats fed the DEHP-free diet and
vitamin-free water. Group B: Experimental rats fed the
DEHP-free diet and vitamin-supplemented water. Group C:
Experimental rats fed the 2% DEHP-containing diet and
vitamin-free water. Group D: Experimental rats fed the 2%
DEHP-containing diet and vitamin-supplemented water.
In both the ®rst and second experiments, all rats were
5 weeks of age at the time of the ®rst dosing, and the diet
and tap water were freely available. The average bodyweight
of rats at the start of the experiments was ~130 g. The
average volumes of diet taken and drinking water per rat
were approximately 50 g and 30 mL per day, respectively,
and there were no signi®cant differences in these volumes
between the DEHP-free and DEHP-administered groups, or
between the vitamin-free and vitamin-supplemented groups.
After completion of the treatments, the animals were deeply
anaesthetized with diethyl ether and their body weights were
recorded. Thereafter, the animals were killed and the testes,
kidneys and the liver of each animal were then removed and
weighed. Organ weight/body weight ´ 100 is shown as the
relative organ weight.
HistologyAfter recording the organ weights, the testes were
immediately ®xed in Bouin's solution for 2 days. Then,
the organs were washed, dehydrated in an ethanol series and
embedded in plastic (Technovit 7100; Kuizer, Germany).
The organs were sectioned at 5 lm with a microtome
(HN340E; Microme, Germany), after which the sections
were stained with Gill's haematoxylin III and 2% eosin Y.
The degree of spermatogenic disturbance was determined
according to Johnsen's scoring system, ranging from score1
(�no cells in the seminiferous tubules) to score10 (�com-
plete spermatogenesis)(Johnsen, 1970). At least 500 round or
oval sections of seminiferous tubules were examined and the
mean score was calculated for each animal.
Flow cytometryIn the second experiment, the left testis of each rat was
minced and suspended in a solution comprising RPMI1640
tissue culture medium (Biowhittaker, Maryland, USA)
containing 0.1% collagenase type IV (Sigma, Missouri,
USA) for 1 h at 37 °C. The cellular elements were then
centrifuged at 400 g for 10 min and ®xed in 70% cold
ethanol for 1 h at 4 °C. After removing the ethanol and
washing the cells with phosphate-buffered saline (pH 7.2),
the resulting pellet was resuspended in 1 mL 0.1% ribonuc-
lease A (Sigma) at 37 °C for 30 min with frequent vortex
mixing. The cells were then washed again with phosphate-
buffered saline by centrifugation under the same conditions.
The resulting pellet was stained with 1 mL of 5 lg/mL
propiduim iodide (Sigma) and then ®ltered through a nylon
mesh. Flow-cytometric analysis was performed using a
Facscan (Becton Dickinson, New Jersey, USA), ®tted with
a 488-nm argon ion laser line at 250 mW to induce the
¯uorescence of propidium iodide. Approximately 2±3 ´ 105
cells were examined for each specimen and DNA histograms
86 M. Ishihara et al.
Ó 2000 Blackwell Science Ltd, International Journal of Andrology, 23, 85±94

were generated by counting at least 1 ´ 105 nuclei per
sample. The low signal events (cell debris) were subtracted
from the histograms. The populations of cells containing
quantitatively similar amounts of DNA are represented by
discrete peaks on the histograms i.e. each of the peaks
represents a cell subpopulation with a speci®c DNA content
or ploidy. The ®rst peak for the normal control sample
represents haploid cells (1n), the second peak diploid cells
(2n), and the third peak tetraploid cells (4n). The areas under
these peaks were integrated by computer analysis to yield
quantitative information on the relative proportion of each
cell type with respect to the total cell population. Sper-
matids and spermatozoa contain haploid DNA. G0/G1
spermatogonia, Sertoli cells, interstitial cells and secondary
spermatocytes produce the diploid peak, whereas G2/M
spermatogonia and primary spermatocytes are responsible for
the tetraploid peak.
Statistical analysisValues are presented as means � standard deviation (SD).
The signi®cance of the differences between groups was
determined by ANOVA, with the aid of Statview-J-4.5
(Abacus Concepts, CA).
Results
Effect of DEHP-containing diets on the testesIn the 1% DEHP-treated rats (Group II), liver enlarge-
ment was noted at 2 and 4 weeks (p < 0.05), but not at
6 weeks (Fig. 1). However, there were no signi®cant
changes in the weights of the body, testes and kidneys
compared with controls (Group I), except that bodyweight
at 6 weeks was slightly lower than in Group I (p < 0.05)
(Fig. 1). In the 2% DEHP-treated rats (Group III), a
signi®cant decrease in bodyweight and marked testicular
atrophy were detected compared with either Group I or II
(p < 0.01) (Fig. 1). The relative weight of testes in Group III
was also dramatically decreased (p < 0.01 compared with
either Group I or II). However, the degree of liver
enlargement in Group III was not signi®cantly different
from that in Group II. Kidney weight in Group III
compared with Group I showed a slight reduction at 2 and
4 weeks (p < 0.05), however, the relative weights exhibited
no signi®cant change (Fig. 1).
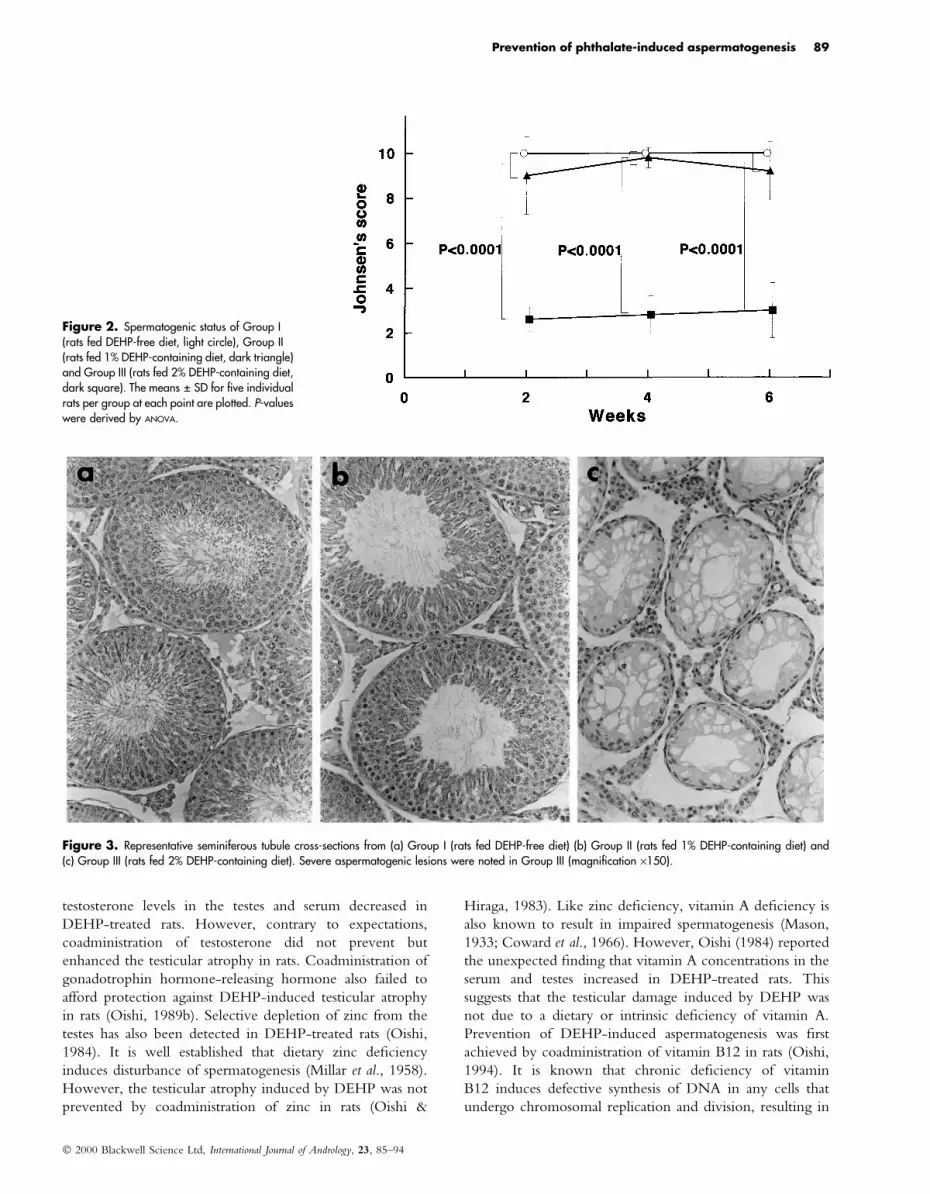
Histopathological examination revealed that testes from
Group I showed complete spermatogenesis whereas severe
aspermatogenesis was induced in Group III. Some semin-
iferous tubules in the Group II testes exhibited spermato-
genic disturbance, however, the lesions were not signi®cant
compared with Group I (Fig. 2). In Group III, marked
atrophy of the seminiferous tubules, and disappearance of
spermatids and spermatocytes were observed (Fig. 3). Sertoli
cells with no or few spermatogonia remained in most
seminiferous tubules. However, destruction of the tubular
architecture or an apparent loss of interstitial cells was not
seen. Therefore, it was found that the testicular damage was
inducible in rats by feeding a DEHP-containing diet without
injection of the chemical. Furthermore, the results showed
that 1% DEHP in the diet was enough to cause liver
enlargement, whereas a critical point for gonadotoxicity
existed with diets of between 1 and 2% DEHP.
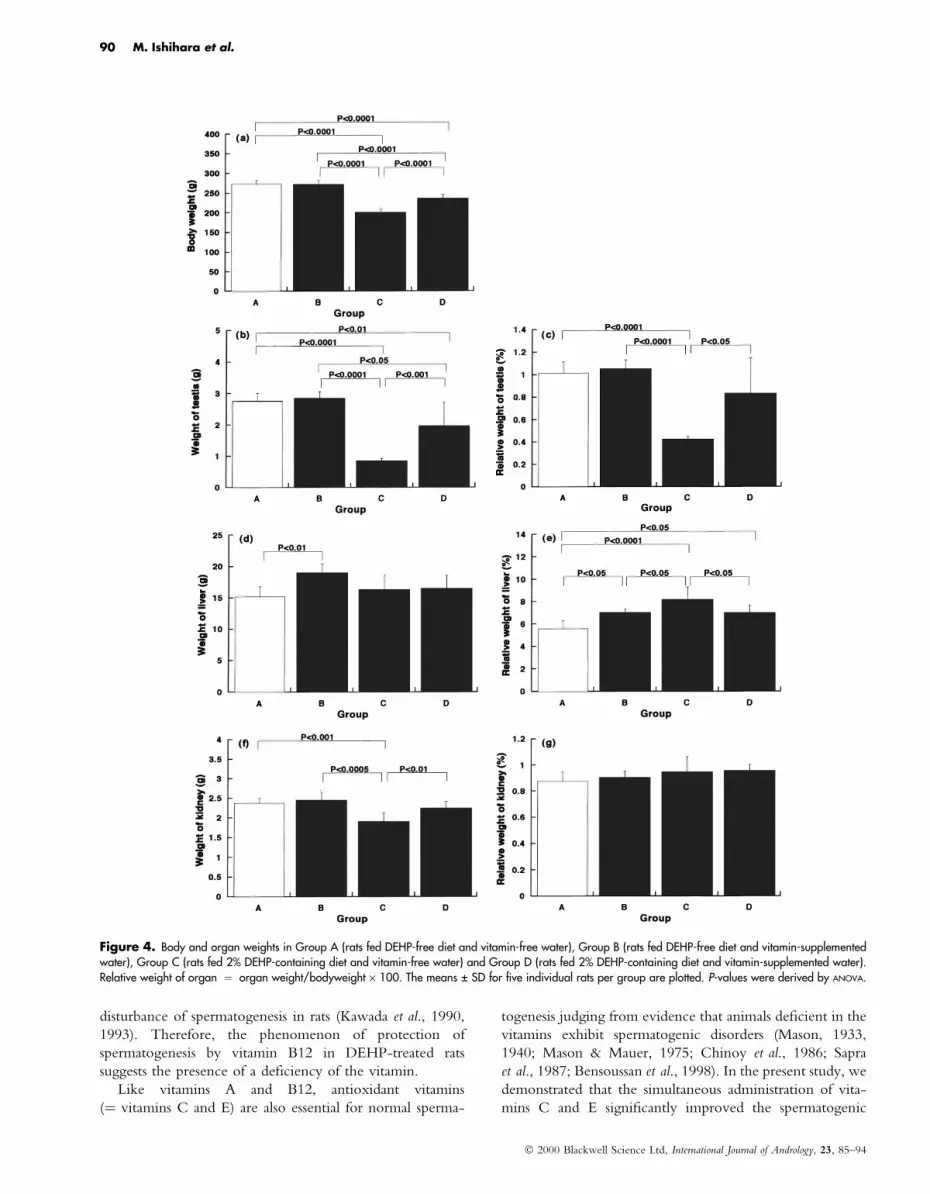
Effects of vitamins C and E on 2% DEHP-inducedtesticular damageAs seen in the ®rst experiments, experimental rats fed the
2% DEHP-containing diet for 2 weeks (Group C) exhibited
a signi®cant decrease in bodyweight and dramatical atrophy
of the testes compared with controls (Group A)
(p < 0.0001). In addition, a signi®cant increase in relative
liver weight and no signi®cant change in relative kidney
weight were also con®rmed in Group C (Fig. 4). However,
rats fed the 2% DEHP-containing diet with supplementation
of vitamins C and E (Group D) showed signi®cant
prevention of these changes (p < 0.05 compared with
Group C). The relative testis weights in Groups A and D
were not signi®cantly different from each other, although
the absolute organ weight in Group D did not reach that in
Group A (p < 0.01) (Fig. 4). Rats fed the DEHP-free diets
and vitamin-supplemented water (Group B) did not exhibit
signi®cant changes in body and organ weights except that
slight liver enlargement was unexpectedly induced (p < 0.05
compared with Group A) (Fig. 4).
Histopathological examination revealed that spermato-
genic status in Group D did not return to that in Group A
(p < 0.01) but was signi®cantly improved compared with
Group C (p < 0.0001) (Fig. 5). Severe aspermatogenesis, a
typical histopathological feature in Group C, was never seen
in Group D (Fig. 6). Group B testes exhibited complete
spermatogenesis similar to that in Group A (Fig. 6). Flow
cytometric analysis revealed apparent reductions of haploid
and tetraploid cells, and an increase of diploid cells in Group
C (p < 0.005 compared with Group A), showing that the
remaining cells were mainly G0/G1 spermatogonia, Sertoli
cells and interstitial cells (Fig. 7). However, the germ cell
populations in Group D were comparable with those in
Group A (Fig. 7), indicating that many haploid cells
(spermatozoa and spermatids) and tetraploid cells (G2/M
spermatogonia and primary spermatocytes) were continu-
ously differentiated from diploid cells (G0/G1 spermato-
gonia) in Group D. Therefore, these results showed that
supplementation of vitamins C and E could protect the testes
from DEHP-gonadotoxicity.
DiscussionIn the present study, disruption of spermatogenesis was
induced in rats by feeding them a diet containing 2% DEHP.
The amount of DEHP producing the aspermatogenic lesions
was extremely large as the rats ingested approximately 1 g
DEHP/day. It is unlikely that such a large dose of DEHP
Ó 2000 Blackwell Science Ltd, International Journal of Andrology, 23, 85±94
Prevention of phthalate-induced aspermatogenesis 87

enters the human system. However, the spermatogenic
lesions induced by the 2% DEHP-diet could be signi®cantly
improved by supplementation of antioxidant vitamins C and
E. This shows that these vitamins can protect the testes from
the gonadotoxicity of DEHP. This is the ®rst demonstration
of prevention of DEHP-induced aspermatogenesis by
antioxidant vitamins.
DEHP is now regarded as an endocrine disrupter (Reiter
et al., 1998; Mylchreest et al., 1999). However, there has
been no direct evidence of an oestrogen-like action of the
chemical (Nakai et al., 1999). Although the basic metabolic
disturbance which elicits aspermatogenesis remains obscure,
there have been some attempts at prevention of DEHP-
induced testicular damage. Oishi (1989a) reported that
Figure 1. Body and organ weights in Group I (rats fed DEHP-free diet, light circle), Group II (rats fed 1% DEHP-containing diet, dark triangle) and Group III (ratsfed 2% DEHP-containing diet, dark square). Relative weight of organ � organ weight/bodyweight ´ 100. The means � SD for ®ve individual rats per group ateach point are plotted. P-values were derived by ANOVA.
Ó 2000 Blackwell Science Ltd, International Journal of Andrology, 23, 85±94
88 M. Ishihara et al.
testosterone levels in the testes and serum decreased in
DEHP-treated rats. However, contrary to expectations,
coadministration of testosterone did not prevent but
enhanced the testicular atrophy in rats. Coadministration of
gonadotrophin hormone-releasing hormone also failed to
afford protection against DEHP-induced testicular atrophy
in rats (Oishi, 1989b). Selective depletion of zinc from the
testes has also been detected in DEHP-treated rats (Oishi,
1984). It is well established that dietary zinc de®ciency
induces disturbance of spermatogenesis (Millar et al., 1958).
However, the testicular atrophy induced by DEHP was not
prevented by coadministration of zinc in rats (Oishi &
Hiraga, 1983). Like zinc de®ciency, vitamin A de®ciency is
also known to result in impaired spermatogenesis (Mason,
1933; Coward et al., 1966). However, Oishi (1984) reported
the unexpected ®nding that vitamin A concentrations in the
serum and testes increased in DEHP-treated rats. This
suggests that the testicular damage induced by DEHP was
not due to a dietary or intrinsic de®ciency of vitamin A.
Prevention of DEHP-induced aspermatogenesis was ®rst
achieved by coadministration of vitamin B12 in rats (Oishi,
1994). It is known that chronic de®ciency of vitamin
B12 induces defective synthesis of DNA in any cells that
undergo chromosomal replication and division, resulting in
Figure 2. Spermatogenic status of Group I(rats fed DEHP-free diet, light circle), Group II(rats fed 1% DEHP-containing diet, dark triangle)and Group III (rats fed 2% DEHP-containing diet,dark square). The means � SD for ®ve individualrats per group at each point are plotted. P-valueswere derived by ANOVA.
Figure 3. Representative seminiferous tubule cross-sections from (a) Group I (rats fed DEHP-free diet) (b) Group II (rats fed 1% DEHP-containing diet) and(c) Group III (rats fed 2% DEHP-containing diet). Severe aspermatogenic lesions were noted in Group III (magni®cation ´150).
Ó 2000 Blackwell Science Ltd, International Journal of Andrology, 23, 85±94
Prevention of phthalate-induced aspermatogenesis 89
disturbance of spermatogenesis in rats (Kawada et al., 1990,
1993). Therefore, the phenomenon of protection of
spermatogenesis by vitamin B12 in DEHP-treated rats
suggests the presence of a de®ciency of the vitamin.
Like vitamins A and B12, antioxidant vitamins
(� vitamins C and E) are also essential for normal sperma-
togenesis judging from evidence that animals de®cient in the
vitamins exhibit spermatogenic disorders (Mason, 1933,
1940; Mason & Mauer, 1975; Chinoy et al., 1986; Sapra
et al., 1987; Bensoussan et al., 1998). In the present study, we
demonstrated that the simultaneous administration of vita-
mins C and E signi®cantly improved the spermatogenic
Figure 4. Body and organ weights in Group A (rats fed DEHP-free diet and vitamin-free water), Group B (rats fed DEHP-free diet and vitamin-supplementedwater), Group C (rats fed 2% DEHP-containing diet and vitamin-free water) and Group D (rats fed 2% DEHP-containing diet and vitamin-supplemented water).Relative weight of organ � organ weight/bodyweight ´ 100. The means � SD for ®ve individual rats per group are plotted. P-values were derived by ANOVA.
Ó 2000 Blackwell Science Ltd, International Journal of Andrology, 23, 85±94
90 M. Ishihara et al.

status of DEHP-treated rats. Although further study is
needed to determine which vitamin is more important for
the prevention of DEHP-induced aspermatogenesis, both
vitamins have often been simultaneously used in clinical and
experimental medicine, because the two vitamins synergis-
tically exert a potent antioxidant action which affords
protection against excessive peroxidative reactions (Leung
et al., 1981; Reddy et al., 1982). Water-soluble vitamin C is
localized in the cytosol, while lipid-soluble vitamin E is
localized almost exclusively in cell membranes (Tappel et al.,
1973; Giasuddin & Diplock, 1981). The former scavenges
oxygen radicals in the aqueous phase, and the latter scavenges
oxygen radicals in the membrane (Hill & Burk, 1984; Kagan,
1989). Ginter et al. (1982) reported that the simultaneous use
of vitamins C and E synergistically increased the activities
of liver detoxi®cation enzymes dependent on cytochrome
Figure 5. Spermatogenic status of Group A(rats fed DEHP-free diet and vitamin-free water),Group B (rats fed DEHP-free diet and vitamin-supplemented water), Group C (rats fed 2%DEHP-containing diet and vitamin-free water)and Group D (rats fed 2% DEHP-containing dietand vitamin-supplemented water). The Means� SD for ®ve individual rats per group areplotted. P-values were derived by ANOVA.
Figure 6. Representative seminiferous tubule cross-sections from (a) Group B (rats fed DEHP-free diet and vitamin-supplemented water) (b) Group C (rats fed2% DEHP-containing diet and vitamin-free water) and (c) Group D (rats fed 2% DEHP-containing diet and vitamin-supplemented water) (c). Active spermatogenesiswas noted in Group D (magni®cation ´150).
Ó 2000 Blackwell Science Ltd, International Journal of Andrology, 23, 85±94
Prevention of phthalate-induced aspermatogenesis 91

P-450 in guinea pigs. Chorvatovicova et al. (1991) showed
that simultaneous treatment with vitamins C and E had a
synergistic preventive effect on lipoperoxide production in
experiments on the toxicity and mutagenicity of K2Cr2O7.
Niki et al. (1985) demonstrated that supplementation of
vitamin C signi®cantly increased vitamin E levels in the rat
liver. Further study demonstrated that vitamin C regenerates
vitamin E by reducing vitamin E radicals formed when
vitamin E scavenges oxygen radicals (Niki, 1987). In
previous studies on irradiation- or cadmium-induced
gonadotoxicity, it became evident that spermatogenesis is
very sensitive to oxidative stress (Koizumi & Li, 1992; Narra
et al., 1993, 1994). Therefore, in our experiments, it may be
that free radicals were excessively produced in the testis due
to the DEHP-treatment, with resultant de®ciency of the
antioxidant vitamins in this organ.
ConclusionsThe preventive effects of vitamins C and E on DEHP-
induced testicular damage have been demonstrated for the
®rst time in the present study. This suggests that the
cooperative redox systems comprising supplemented vita-
mins C and E play a role in maintenance of spermatogenesis.
In a further study, we will determine the concentrations of
vitamins C and E, and free radicals in the testicular tissue of
DEHP-treated rats. Besides vitamins C and E, a signi®cant
number of complementary antioxidant systems prevent
oxidative damage in the body under normal conditions.
These include superoxide dismutase, glutathione, peroxid-
ase, uric acid, cystein and catalase (Tappel et al., 1973;
Docampo et al., 1981; Harman, 1981; Reddy et al., 1982;
Hill & Burk, 1984; McCay, 1985). Therefore, it might be
that the amounts of these antioxidants in the testes also
decrease after DEHP-treatment, but that their supplemen-
tation is capable of protecting the testes from gonadotoxicity.
To our knowledge, there has been no study of histological
changes to the damaged seminiferous epithelium after
cessation of DEHP administration. Thus, in future experi-
ments we will determine whether or not the severely
damaged spermatogenesis of DEHP-treated rats recover, with
or without later supplementation with vitamins C and E.
AcknowledgementsWe thank Mrs. M. Miyawaki, Mr W. Nagata and Mrs. E.
Turumi for their secretarial and technical assistance. This
work was supported by a Grant-in-Aid for General Scienti®c
Research (11470108) from the Ministry of Education,
Science, Sports and Culture, Japan.
ReferencesAlbro, P. W. (1987) The biochemical toxicology of di(2-ethyl-
hexyl) phthalate: testicular atrophy and hepatocarcinogenesis.
Review of Biochemistry and Toxicology 8, 73±119.
Bensoussan, K., Morales, C. R. & Hermo, A. L. (1998) Vitamin E
de®ciency causes incomplete spermatogenesis and affects the
structural differentiation of epithelial cells of the epididymis
in the rat. Journal of Andrology 19, 266±288.
Burton, G. W. & Ingold, K. (1989) Vitamin E as an in vitro and in vivo
antioxidant. Annals of the New York Academy of Sciences 570, 7±22.
Chinoy, N. J.,Mehta,R.P.B. R.R., Seethalakshmi, L., Sharma, J.D.
& Chinoy, M. R. (1986) Effects of vitamin C de®ciency on
physiology of male reproductive organs of guinea pigs.
International Journal of Fertility 31, 232±239.
Chorvatovicova, D., Ginter, E., Kosinova, A. & Zloch, Z. (1991)
Effect of vitamins C and E on toxicity and mutagenicity of
hexavalent chromium in rat and guinea pig. Mutation Research
262, 41±46.
Coward, W. A., Howell, J. M. C., Pitt, G. A. J. & Thompson, J. N.
(1966) Effect of hormones on reproduction in rats fed a diet
de®cient in retinol (vitamin A alcohol) but containing methyl
retioate (vitamin A acid methyl ester). Journal of Reproduction and
Fertility 12, 309±317.
Docampo, R., Moreno, S. N. J. & Stoppani, A. O. M. (1981)
Nitrofuran enhancement of microsomal electron transport,
superoxide anion production and lipid peroxidation. Archives
of Biochemistry and Biophysics 207, 316±324.
Figure 7. Flow cytometric DNA analysis of testicular cells from Group A (rats fed DEHP-free diet and vitamin-free water), Group C (rats fed 2% DEHP-containing diet and vitamin-free water), and Group D (rats fed 2% DEHP-containing diet and vitamin-supplemented water). The means � SD for ®ve individual ratsper group are plotted. P-values were derived by ANOVA. It was noted that the cell populations in Group D were comparable to those in Group A, showing thepresence of active spermatogenesis.
Ó 2000 Blackwell Science Ltd, International Journal of Andrology, 23, 85±94
92 M. Ishihara et al.

Faouzi, M. A., Dine, T., Gressier, B., Kambia, K., Luyckx, M.,
Pagniez, D., Brunet, C., Cazin, M., Belabed, A. & Cazin, J. C.
(1999) Exposure of hemodialysis patients to di-2-ethylhexyl
phthalate. International Journal of Pharmacology 180, 113±121.
Giasuddin, A. S. M. & Diplock, A. T. (1981) The in¯uence of
vitamin E on membrane lipids of mouse ®broblasts in culture.
Archives of Biochemistry and Biophysics 210, 348±362.
Ginter, E., Kosinova, A., Hudecova, A. & Madaric, A. (1982)
Synergism between vitamins C and E: effect on microsomal
hydroxylation in guinea pig liver. International Journal for Vitamin
and Nutrition Research 52, 49±55.
Harman, D. (1981) The aging process. Proceedings of the National
Academy of Sciences of the USA 78, 7124±7128.
Hill, K. E. & Burk, R. F. (1984) In¯uence of vitamin E and
selenium on glutathione-dependent protection against micro-
somal lipid peroxidation. Biochemical Pharmacology 33,
1065±1068.
Hillman, L. S., Goodwin, S. L. & Sherman, W. R. (1975)
Identi®cation and measurement of plasticizer in neonatal tissues
after umbilical catheters and blood products. New England Journal
of Medicine 292, 381±385.
Johnsen, S. G. (1970) Testicular biopsy score count-a method for
registration of spermatogenesis in human testis. normal values
and results in 335 hypogonadal males. Hormones 1, 2±25.
Kagan, V. E. (1989) Tocopherol stabilizes membrane against
phospholipase A, free fatty acids, and lysophospholipids. Annals
of the New York Academy of Sciences 570, 121±135.
Kawada, T., Ikawa, Y., Haisa, A., Hirose, Y., Fujita, A., Yamada,
K., Wada, M., Tadokoro, T., Maekawa, A. & Tanaka, N.
(1990) Effects of vitamin B12 on reproductive functions in male
rats. Vitamins 65, 192.
Kawada, T., Tamiki, I., Tahiro, A., Kan, K., Kamioka, S., Tanaka,
N., Wada, M., Yamada, K., Tadokoro, T. & Maekawa, A.
(1993) Effect of vitamin B12 de®ciency on male reproductive
functions in rats. Vitamins 67, 205.
Koizumi, T. & Li, Z. G. (1992) Role of oxidative stress in single-
dose cadmium-induced testicular cancer. Journal of Toxicology and
Environmental Health 37, 25±36.
Leung, H. W., Vang, M. J. & Mavis, R. D. (1981) The cooperative
interaction between vitamin E and vitamin C in suppression of
peroxidation of membrane phospholipids. Biochemica et Biophys-
ica Acta 664, 266±272.
Manojkumar, V., Deepadevi, K. V., Arun, P., Nair, K. G., Lakshmi,
L. R. & Kurup, P. A. (1999) Changes in the concentration of
erythrocyte membrane during storage of blood in di-(2-ethyl-
hexyl) phthalate (DEHP) plasticized poly vinyl chloride (PVC)
blood storage bags. Indian Journal of Medical Research 109, 157±163.
Mason, K. E. (1933) Differences in testis injury and repair after
vitamin A de®ciency, vitamin E de®ciency, and inanition.
American Journal of Anatomy 52, 153±239.
Mason, K. E. (1940) Minimal requirements of male and female rats
for vitamin E. American Journal of Physiology 131, 268±280.
Mason, K. E. & Mauer, S. I. (1975) Reversible testis injury in the
vitamin E-de®cient hamster. Journal of Nutrition 105, 484±490.
McCay, P. B. (1985) Vitamin E: interactions with free radicals and
ascorbate. Annual Review of Nutrition 5, 323±340.
Millar, M. J., Fischer, M. I., Elcoate, P. V. & Mawson, C. A. (1958)
The effect of dietary zinc de®ciency on the reproductive system
of male rats. Canadian Journal of Biochemistry and Biophysiology 36,
557±569.
Mylchreest, E., Sar, M., Cattley, R. C. & Foster, P. M. D. (1999)
Disruption of androgen-regulated male reproductive develop-
ment by di(n-butyl) phthalate during late gestation in rats is
different from ¯utamide. Toxicology and Applied Pharmacology
156, 81±95.
Nakai, M., Tabira, Y., Asai, D., Yakabe, Y., Shimyozu, T.,
Noguchi, M., Takatsuki, M. & Shimohigashi, Y. (1999)
Binding characteristics of dialkyl phthalates for the estrogen
receptor. Biochemical and Biophysical Research Communications 254,
311±314.
Narra, V. R., Howell, R. W., Sastry, K. S. R. & Rao, D. V. (1993)
Vitamin C as a radioprotector against 131I in vivo. Journal of
Nuclear Medicine 34, 637±640.
Narra, V. R., Harapanhalli, R. S., Howell, R. W., Sastry, K. S. R.
& Rao, D. V. (1994) Vitamins as radioprotectors in vivo. I.
Protection by vitamin C against internal radionuclides in mouse
testes: Implications to the mechanism of damage caused by the
auger effect. Radiation Research 137, 394±399.
Niki, E., Saito, T., Kawakami, A. & Kamiya, Y. (1985) Inhibition
of oxidation of methyl linoleate in solution by vitamin E and
vitamin C. Journal of Biological Chemistry 259, 4177±4182.
Niki, E. (1987) Interaction of ascorbate and a-tocopherol. Annals of
the New York Academy of Sciences 498, 186±198.
Oishi, S. & Hiraga, K. (1983) Testicular atrophy induced by
di-2-ethylhexyl phthalate: Effect of Zinc Supplement. Toxicology
and Applied Pharmacology 70, 43±48.
Oishi, S. (1984) Testicular atrophy of rats induced by di-2-
ethylhexyl phthalate: Effects of vitamin A and zinc concentra-
tions in the testis, liver and serum. Toxicology Letters 20, 75±78.
Oishi, S. (1989a) Effects of co-administration of di-(2-ethylhexyl)
phthalate and testosterone on several parameters in the testis and
pharmacokinetics of its mono-de-esteri®ed metabolite. Archives
of Toxicology 63, 289±295.
Oishi, S. (1989b) Enhancing effects of luteinizing hormone-
releasing hormone on testicular damage induced by di-(2-ethyl-
hexyl) phthalate in rats. Toxicology Letters 47, 271±277.
Oishi, S. (1994) Prevention of di-(2-ethylhexyl) phthalate-induced
testicular atrophy in rats by co-administration of the vitamin
B12 derivative adenosylcobalamin. Archives of Environmental
Contamination and Toxicology 26, 497±503.
Reddy, C. C., Scholz, R. W., Thomas, C. E. & Massaro, E. J.
(1982) Vitamin E dependent reduced gluthathione inhibition of
rat liver microsomal lipid peroxidation. European Journal of Life
Sciences 31, 571±576.
Reiter, L. W., De-Rosa, C., Kavlock, R. J., Lucier, G., Mac M.
J., Melillo, J., Melnick, R. L., Sinks, T. & Walton, B. T.
(1998) The U.S. federal framework for research on endocrine
disruptors and an analysis of research programs supported
during ®scal year 1996. Environmental Health Perspectives 106,
105±113.
Sapra, M., Sharma, P. & Kothari, L. K. (1987) Effect of vitamin C
de®ciency on testicular structure in the guinea pig. Journal of
Postgraduate Medicine 33, 69±73.
Sjoberg, P. O. J., Bondesson, U. G., Sedin, E. G. & Gustaffson,
J. P. (1985) Exposure of newborn infants to plasticizers. Plasma
level of di-(2-ethylhexyl) phthalate and mono-(2-ethylhexyl)
phthalate during exchange transfusion. Transfusion 25, 424±
428.
Summer®eld, F. W. & Tappel, A. L. (1984) Effects of dietary
polyunsaturated fats and vitamin E on aging and peroxidative
damage to DNA. Archives of Biochemistry and Biophysics 233,
408±416.
Tappel, A. L. (1962) Vitamin E as the biological lipid antioxidant.
Vitamins and Hormones 20, 493±510.
Ó 2000 Blackwell Science Ltd, International Journal of Andrology, 23, 85±94
Prevention of phthalate-induced aspermatogenesis 93

Tappel, A L., Fletcher, B. & Deamer, D. (1973) Effect of
antioxidants and nutrients on lipid peroxidation ¯uorescent
products and aging parameters in the mouse. Journal of
Gerontology 28, 415±424.
Thomas, J. A., Darby, T. D., Wallin, R. F., Garvin, P. J. &
Martis, L. (1978) A. review of the biological effects of
di(2-ethylhexyl) phthalate. Toxicology and Applied Pharmacology
45, 1±27.
Received 29 September 1999; accepted 27 November 1999
1. VII International Congress of Andrology, hosted by the American Society of Andrology
(also the 26th Annual Meeting of the ASA)
Montreal, Quebec, Canada
Friday 15 June 2001 ± Tuesday 19 June 2001
2. 27th Annual Meeting of the American Society of Andrology
Sheraton Seattle Hotel and Towers, Seattle, Washington, USA
Thursday 26 April ± Saturday 29 April
(Postgraduate course: 29 April 2002. Laboratory workshop: 24 April 2002
For more information on all meetings or to request a 2000 abstract submission form please contact:
American Society of Andrology
74 New Montgomery, Suite 230
San Francisco, CA 94105
USA
phone: (415) 764 4823
fax: (415) 764 4915
email: [email protected]
homepage: www.cvm.uiuc.edu/HomePages/rhess/asa/front
Ó 2000 Blackwell Science Ltd, International Journal of Andrology, 23, 85±94
94 M. Ishihara et al.