sparc modulates the proliferation of stromal but not melanoma cells unless endogenous sparc...
TRANSCRIPT

SPARC modulates the proliferation of stromal but not melanoma cells unless
endogenous SPARC expression is downregulated
Cynthia L�opez Haber1, Vanesa Gottifredi1, Andrea S. Llera1, Edgardo Salvatierra1, Federico Prada1, Leonardo Alonso2,Sage E. Helene3 and Osvaldo L. Podhajcer1*
1Laboratory of Molecular and Cellular Therapy, Fundaci�on Instituto Leloir-CONICET, Universidad de Buenos Aires,Buenos Aires, Argentina2Structure, Function and Protein Engineering Laboratory, Fundaci�on Instituto Leloir-CONICET, Buenos Aires, Argentina3Hope Heart Program, The Benaroya Research Institute at Virginia Mason, Seattle, WA
Cell interaction with the extracellular matrix (ECM) has profoundinfluence in cancer progression. The secreted protein, acidic andrich in cysteine (SPARC) a component of the ECM, impairs theproliferation of different cell types and modulates tumor cellaggressive features. This apparent paradox might result eitherfrom the biochemical properties of the different SPARC sourcesor from differential responses of malignant and stromal cells toSPARC. To test these hypotheses, we purified SPARC secreted bymelanoma cells (hMel-SPARC) and compared its activity with dif-ferent recombinant SPARC preparations, including a new oneproduced in insect cells. All 5 SPARC species were effective in in-hibiting bovine aortic endothelial cell proliferation, adhesion andmigration. We then used the melanoma-derived protein to assessSPARC effect on additional cell types. hMel-SPARC greatlyimpaired the proliferation of both normal and transformedhuman endothelial cells and exerted a moderate biphasic effect onhuman fetal fibroblasts proliferation, irrespective of their endoge-nous SPARC levels. However, SPARC had no effect on the prolif-eration of several human cancer cell lines regardless of their en-dogenous levels of SPARC expression. Importantly, downregula-tion of SPARC levels in melanoma cells using either an antisenseRNA or a shRNA against SPARC sensitized them to hMel-SPARCaddition in proliferation and migration assays, suggesting that ma-lignant cells developed a SPARC-resistance mechanism. This wasnot a general resistance to growth suppressing agents, as mela-noma cells with restricted SPARC expression were more resistantto chemotherapeutic agents. Thus, malignant cells expressing ornot expressing SPARC developed alternative mechanisms that, incontrary to stromal cells, rendered them SPARC-insensitive.' 2007 Wiley-Liss, Inc.
Key words: SPARC; osteonectin; cell proliferation; tumor hetero-geneity; stromal cells
Tumors grow as the result of crosstalk between the differentcellular components of the tumor mass. Malignant cells secreteproteins that reach neighboring stromal nonmalignant cells (i.e.,fibroblasts and endothelial cells) that in some cases induce them toprovide the soil in which the tumor will grow.1 Nonmalignantstromal cells might also contribute to cancer progression, since theestablishment of human cancer xenografts in mice depends on thepresence of tumor-derived fibroblasts.2 This data therefore pointstoward a central role of extracellular matrix (ECM) in tumor pro-gression.
From the many proteins secreted by tumor and stromal cells,the secreted protein, acidic and rich in cysteine, SPARC (alsoknown as BM40 and osteonectin), a nonstructural, matricellularcomponent of the ECM, has been associated with tumor growth.3
In healthy tissues, SPARC production is largely restricted to areasundergoing repair or remodeling.4 SPARC is expressed at highlevels in many tumors, and it was found to be associated with tu-mor progression in human melanoma,5,6 glioblastoma,7 meningi-oma,8 breast,9 colon,10 lung,11 pancreas,12 myeloma13 and prostatecancer.14 Among the many biological processes modulated bySPARC, its antiadhesive,5,15 promigratory,14 and antiapoptotic16
properties on certain cell types have been associated with tumorprogression. Recent evidence also indicates that SPARC producedby melanoma cells might impair the tumor attack by polymorpho-
nuclear cells, indicating a role of SPARC in tumor evasion fromimmune surveillance.17 In contrast, SPARC has also been associ-ated with antitumorigenic activities, because its overexpressioninhibited the growth of ovarian cancer cells,18 and its productionby Schwann cells was linked to angiogenesis inhibition in neuro-blastoma.19 Studies in SPARC-null mice led also to controversialresults. Indeed, pancreatic and lymphoma tumors showed acceler-ated growth in SPARC-null mice while mammary tumors wererejected through a mechanism that involved host leukocytes.20
SPARC is produced not only by the malignant cells but also bytumor-associated stromal cells.3 In most human adenocarcinomas,SPARC is expressed in tumor-associated fibroblasts and endothe-lial cells, whereas its expression is repressed in malignant cells.3,12
A study performed on human nonsmall cell lung carcinomas indi-cated that high levels of SPARC in tumor stroma correlated withpoor prognosis.11 Similarly, a recent report described the prognos-tic significance of stroma-derived SPARC on the outcome of can-cer patients.21 These evidences indicate that the expression ofSPARC by stromal cells might also contribute to tumor progres-sion.
While the evidence described earlier is suggestive of a protu-morigenic role for SPARC in many tumor models, most data indi-cate that SPARC is associated with inhibitory effects on cell pro-liferation.3,22,23 The mechanisms by which SPARC delays theonset of S phase have been partially unraveled. In fact, SPARCnegatively modulates the signal of many growth factors, includingbasic fibroblast growth factor (bFGF), VEGF165, IGF, PDGF anddelays mitogen-activated protein kinase (MAPK) and cyclins/CDKs activation.24–29
Given that SPARC effects in human cancer might depend on itsconcerted action on the different cellular components of the tumormass,30 we analyzed the effects of exogenous SPARC on the dif-ferent cell types comprising a tumor mass. Although both prokary-otic and eukaryotic systems31–37 have been used as a source ofSPARC, natural SPARC from cancer cells have been used rarelyfor this purpose38 and never for functional studies. Therefore, wepurified and characterized SPARC secreted by melanoma cells(hMel-SPARC). Comparative analyses were performed with a
This article contains supplementary material available via the Internet athttp://www.interscience.wiley.com/jpages/0020-7136/suppmat.Grant sponsors: Fundaci�on Antorchas, UBACyT, CONICET, ANPCYT,
Fundaci�on Rene Baron, AFULIC; Grant sponsor: National Institute ofHealth; Grant number: GM-40711.The first three authors contributed equally to this work.*Correspondence to: Fundaci�on Instituto Leloir, Avenida Patricias
Argentinas 435, 1405 Buenos Aires, Argentina. Fax:15491152387501.E-mail: [email protected] 29 March 2007; Accepted after revision 29 August 2007DOI 10.1002/ijc.23216Published online 4 December 2007 in Wiley InterScience (www.interscience.
wiley.com).
Abbreviations: bFGF, basic fibroblast growth factor; C-Ab, isotype-matched control antibody; ECM, extracellular matrix; FBS, fetal bovineserum; hMel-SPARC, human melanoma-derived SPARC; N-Ab, neutraliz-ing antibody; rh-SPARC, recombinant human SPARC; shRNA, short hair-pin ribonucleic acid; SPARC, secreted protein, acidic and rich in cysteins.
Int. J. Cancer: 122, 1465–1475 (2008)' 2007 Wiley-Liss, Inc.
Publication of the International Union Against Cancer

novel recombinant human SPARC expressed in Drosophila mela-nogaster cells (rh-SPARC), and with previously describedrecombinant SPARC species.35,36 Essentially, all preparations ofSPARC showed antiadhesive and antiproliferative properties onwell-known SPARC-responsive cells such as bovine aortic endo-thelial cells, indicating that the source of SPARC does not signifi-cantly alter its biological properties. We also demonstrated thatSPARC affected proliferation of additional human normal andtransformed endothelial cells and fibroblasts regardless of whethercells expressed or not SPARC. However, SPARC exhibited noeffect at all on the proliferation of various melanoma, glioma andcolon cancer cell lines. The absence of effect was not correlatedwith the production of SPARC by tumor cells. However, mela-noma cells that expressed high endogenous levels of SPARCbecame sensitive to the antiproliferative activity of exogenousSPARC following downregulation of its endogenous expressionusing either an antisense RNA or a short hairpin ribonucleic acid(shRNA) against SPARC. Similarly, only melanoma cells with re-stricted SPARC expression exhibited diminished migrationcapacity in the presence of SPARC.
Material and methods
Construction of human SPARC expression vector
Human SPARC was amplified by polymerase chain reaction(PCR) using a linearized SPARC cDNA as template. Primers weredesigned to generate a full-length SPARC flanked by SstI andBamHI restriction sites at the 50 and 30 ends, respectively.Forward primer: 50 GCATCTACGAGCTCATGCGGGCCTG
GATC 30, and reverse primers: 50 GATGGTGAGAACCGCGTGGCACCAGGATCACAAGATCC 30 and 50GCATCTGGATCCTTAATGGTGATGGTGATGGTGAGAACC 30, were used in sub-sequent PCRs to include a thrombin cleavage sequence (Leu-Val-Pro-Arg-Gly-Ser) and a 6xHis tag coding sequence immediatelyfollowing the SPARC C-terminal amino acid codon, all followed by astop codon (TAA). The PCR product, as well as the Drosophilaexpression vector pRmHa-3 (provided by K. Karjalainen, Bellinzona,Switzerland)39 were ligated, and the correct sequence of the con-struct was confirmed by automated DNA sequence analysis.
Human recombinant SPARC production and purification
Drosophila Schneider cells (SL-3) were grown in HyQ SFX-Insect MP (HyClone, Logan, UT) supplemented with 18 mM L-glutamine. Lipid-mediated transfection (Cellfectin, Invitrogen,Grand Island, NY) was carried out simultaneously with theexpression vector coding for human SPARC (pRmHa-rh-SPARC)and the selection marker plasmids pBshs-PURO (for puromycinresistance) and pBshs-BLAST (for blasticidin-S resistance, fromK. Karjalainen). After 10 days of expansion in selective medium,highly expressing clones were derived by limiting dilution.
For protein production, 1 3 109 cells were induced with 1 mMcopper sulfate. Culture supernatant was recovered 72 hr afterinduction, supplemented with 0.01% NaN3, 1 mM PMSF and1 mM EDTA, and dialyzed against 20 volumes of PBS containing2 M NaCl. This supernatant was applied to a 1-mL NTA-Ni21 col-umn (Qiagen) previously equilibrated in PBS/2 M NaCl. The col-umn was washed with PBS/2 M NaCl and elution was performedwith 250 mM imidazole in 50 mM H2Na PO4 (pH 8.0)/300 mMNaCl.
Human recombinant-SPARC-6xH was dialyzed and furtherpurified on a HiTrapQ anion-exchange column (GE Healthcare).The column was previously equilibrated in 20 mM Tris-HCl (pH7.8), and rh-SPARC-6xH was eluted in a continuous salt gradientfrom 0 to 600 mM NaCl. Fractions containing rh-SPARC-6xHwere concentrated in a Centriprep-10 columm (Millipore, Biller-ica, MA). Yields were between 12 and 30 mg purified rh-SPARCper 1 L of culture supernatant. Protein concentration was deter-
mined by absorbance at 280 nm and a calculated extinction coeffi-cient E280nm of 0.875 mL mg21 cm21.
Histidine tag removal
One milligram of histidine-tagged protein in PBS was treatedwith 10 U thrombin (GE Healthcare) for 16 hr at 22�C and incu-bated with Ni21-NTA resin equilibrated in PBS/2 M NaCl for 1 hrat 4�C. After centrifugation, the supernatant was collected and dia-lyzed against 20 mM Tris (pH 7.8), and the protein was furtherpurified by anion-exchange chromatography as described earlier.
Purification of native human SPARC from A375culture supernatants
Serum-free media was conditioned by human melanoma cellline A375 at 80% confluence for 24 hr. Addition of 1 mM PMSF,1 mM EDTA and 0.01% sodium azide was carried out before fur-ther processing.
For protein purification, 1 L of conditioned medium was con-centrated to 50 mL in an Amicon Ultrafiltration Cell model no.202, with YM10 membrane, MWCO 10,000 (Millipore) and dia-lyzed against 20 mM Tris-HCl (pH 7.8). The sample was loadedonto a HiTrap Q column (GE Healthcare) and hMel-SPARC waseluted in a continuous gradient from 100 to 800 mM NaCl. Frac-tions with purity higher than 90% were pooled. After dialysis, thesample was chromatographed over a Superdex 200 column toachieve 98% purity. Yields were between 0.5 and 1.0 mg purifiedhMel-SPARC per liter of culture supernatant. Protein concentra-tion was determined by its absorbance at 280 nm, using a calcu-lated E280nm of 0.875 mL mg21cm21. Final eluate was dialyzedagainst cell-culture grade PBS, and concentrated to �1 mg/mL onCentricon 10 (Millipore). The purified protein was characterizedby Edman N-terminal sequencing (performed at the Protein Struc-ture Core Facility, Department of Biochemistry and MolecularBiology, Nebraska Medical Center, Omaha, NE) and peptide massfingerprinting. All SPARC preparations were routinely tested andshowed no detectable endotoxin when assayed with the Limulusamebocyte lysate kit (Sigma).
Other purified SPARC proteins used in this study were rh-SPARC (293), a human recombinant His-tagged SPARC proteinproduced in HEK (293) cells36 and rh-SPARC (SF9), a humanSPARC protein obtained by baculoviral infection of SF9 insectcells.35
Protein analysis
Conditioned medium was prepared by incubating subconfluentcell cultures in growth medium without serum for 24 hr. Condi-tioned medium was precipitated in ice-cold acetone, air-dried andresuspended in sample buffer. Proteins were resolved by SDS-PAGE and were transferred to nitrocellulose membranes. Westernblots were performed with monoclonal anti-SPARC antibodies(AON-140). Silver or Coomassie blue staining was used to assessprotein loading.
Circular dichroism
Circular dichroism (CD) spectra were monitored in the far-UV(260–190 nm) region using a Jasco J-810 spectropolarimeter. Tenscans were averaged for each measurement at (25.06 0.1)�C. Pro-tein at 5 lM was dialyzed against 10 mM Tris-HCl (pH 7.8), inthe presence of either 2 mM Ca21 or 2 mM EDTA. Changes ofellipticity during thermal denaturation were followed at 220 nmover an interval of 20–80�C.
Cell culture conditions
HMEC-1 and MEC cells were provided by I. Fidler (Houston,TX). HUVE cells were isolated as described.41 WI38, HLF1human diploid fibroblasts, LoVo colon carcinoma and U-87 gli-oma cell lines were obtained from the American Type Culture
1466 L�OPEZ ET AL.

Collection (ATCC, Manassas, VA). HCT116 cells were providedby B. Volgestein, (John Hopkins Medical Institute, Baltimore,MD). IIB-Mel-Les and IIB-Mel-J melanoma cell lines have beenpreviously described.42,43 A375, MEL-888 and SB-2 melanomacell lines were provided by E. Medrano (M.D. Anderson CancerCenter, Houston, TX). CMV and L1-D cells have been describedelsewhere.6 IIB-MEL-LES melanoma cells stably transfected witha control vector (pBLAST) or a cell clone derived from stabletransfection of IIB-MEL-LES with a specific SPARC shRNAs(cell clone L2F6) were obtained following a detailed protocoldescribed elsewhere (Sosa et al., submitted). Briefly, the 21-nucle-otide target sequence starting at base 2,175 (50-TCTTAGTCTTAGTCACCTTAT-30) of human SPARC mRNA was cloned as ashRNA expression cassette with a human U6 promoter intopBLAST, a version of pcDNA6-V5-His-B vector (Invitrogen,Carlsbad, MA), in which the CMV promoter has been deleted.The resulting construct pSP2175 or the control pBLAST vectorwas transfected into IIB-MEL-LES human melanoma cells usingLipofectamine 2000 (Invitrogen). Isolation of the L2F6 cell clonewas carried out by limiting dilution using 1–5 lg/mL blasticidin.Knockdown of SPARC expression was confirmed by real-timePCR and immunoblotting with an anti-SPARC antibody. To accu-mulate cells in the G1 phase of the cell cycle, we grew all celllines to confluence and subsequently deprived them of fetal bovineserum (FBS) in their respective medium for 48–72 hr. Cells werethen released by trypsin treatment and replated in their respectivemedia supplemented with 2.5% FBS and used in DNA synthesisassays as described later.
Induction of cell rounding
BAE cells were grown to confluence and starved for 72 hr. Cellswere plated in each well of a 8-mm multiwell slides (ICN Biomed-icals) in the presence of the indicated SPARC proteins (20 lg/mL,equivalent to 0.6 lM), and fixed at the time-points shown. Roundand spread cells were evidenced by phalloidin and phosphotyro-sine antibodies (Cell Signalling) staining and counted. Values areexpressed as the percentage of nonspread cells (round) withrespect to the total amount of cells in each field. Assays were donein triplicate, and for each data point the mean 6 SD was calcu-lated.
In vitro migration assays
In vitro cell migration of endothelial and melanoma cells wasmeasured as previously described in 48-well micro-Boyden cham-ber (Neuroprobe).44 In all assays, a fibronectin-like protein poly-mer (Sigma) coated polycarbonate filter (8-lm pores, Neuroprobe)were used. SPARC was added at a concentration of 10 lg/mL. Forendothelial cell migration, human bFGF (0.4 ng/mL; Preprotech)diluted in DMEM/ovoalbumin 0.25% BSA (Sigma) was placedin the lower compartment of the Boyden chamber. Chambers wereplaced in humidified atmosphere at 37�C, 5% CO2 for 5 hr. At theend of the incubation period, filters were removed, fixed withmethanol and stained with GIEMSA (Biopur). The filters weremounted on glass slides, and the upper surface of the filters waswiped clean. Cells were counted on a Nikon microscope witha 403 objective and 53 eyepieces. Five fields were countedand data were expressed as the mean of that value. Assays weredone in triplicate, and for each data point the mean 6 SD wascalculated.
Measurement of DNA synthesis
Incorporation of methyl [3H]-thymidine was measured as previ-ously described44,45 with slight modifications. Briefly, growth-arrested cultures were replated at subconfluent density in 96-wellplates in the presence of 2.5% FBS and SPARC proteins or che-motherapeutic agents, as described in the figure legends. Fourhours before sample collection, methyl [3H]-thymidine (15 lCi/mL) was added to the growth medium. Cells were fixed with ice-
cold 10% trichloroacetic acid for 10 min. Methyl [3H]-thymidinein samples was quantified in a scintillation counter (GE Health-care). For each data point, the mean 6 SD of triplicates was calcu-lated.
For neutralization assays, SPARC proteins were preincubated at4�C overnight with the previously described Neutralizing Antibod-ies (N-Abs) 236 and 29346 and isotype-matched control antibody(C-Ab; isotype Control IgG1 mouse Antibodies from Sigma). Pro-liferation assays were performed as described earlier by replatingcells in the presence of SPARC or SPARC 1 the indicated anti-body. Samples were collected at 24 hr, and methyl [3H]-thymidinewas performed in the last 4 hr before simple collection. For thefigures in which methyl [3H]-thymidine incorporation was plottedas percentages (%) of PBS-treated cells the values were calculatedfor each time-point as follows:
½3H�Thymidine incorporation ð% PBS� treated controlÞ ¼½3H�Thymidine incorporationof the sample ðCPMÞ
½3H�Thymidine incorporation ðCPMÞof PBS� treated control3100
Statistical analysis
One-way analysis of variance followed by a Dunnett multiplecomparison test was used for assessing the significance of themean differences in endothelial cell migration induced by PBS orSPARC and for the mean differences in thymidine incorporationbetween PBS and SPARC-treated cells in the presence or not of aN-Ab.
Two-way analysis of variance followed by Bonferroni test wasused for assessing the significance of the mean differences in thy-midine incorporation and migration of melanoma cells betweenPBS and SPARC treatments.
Results
Biochemical characterization of SPARC secreted byhuman melanoma cells
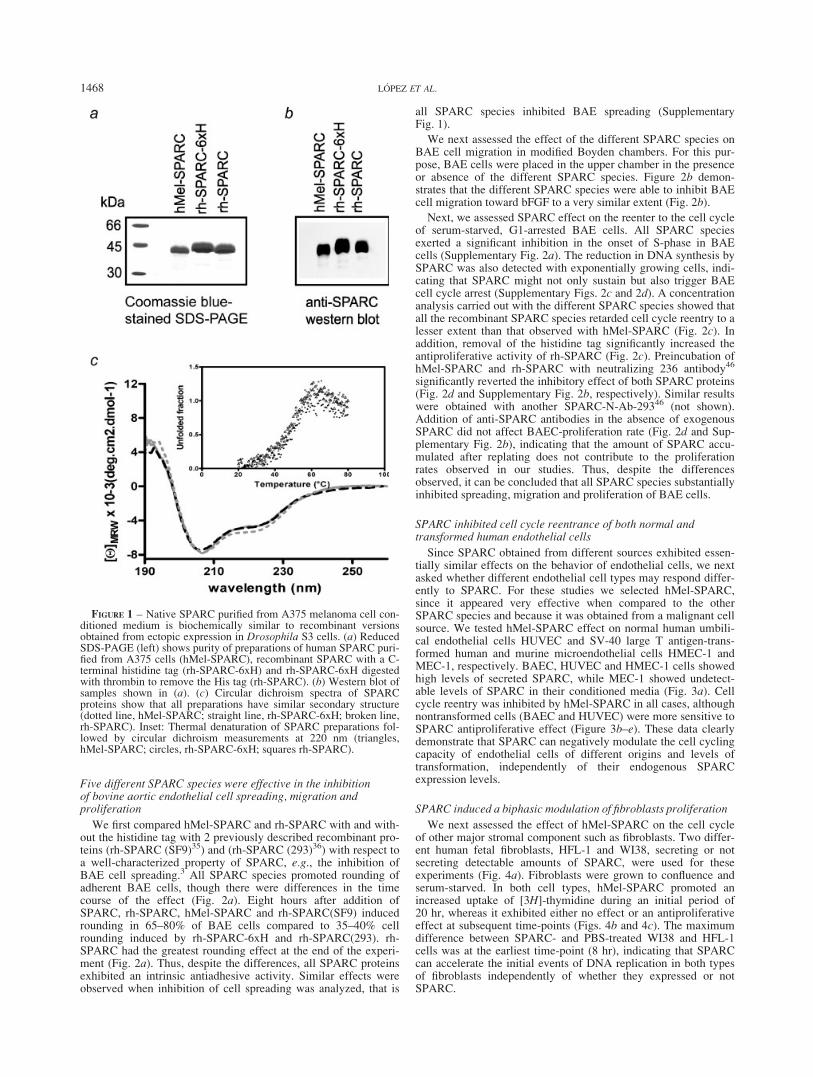
We have developed 2 new sources of biologically active humanSPARC. Native SPARC (hMel-SPARC) was purified to homoge-neity from serum-free conditioned media of the human melanomacell line A375N (Figs. 1a and 1b). Usual yields varied between0.5 and 1 mg L21 of conditioned medium. N-Terminal Edmansequencing and peptide mass fingerprinting by MALDI-TOF con-firmed the identity of hMel-SPARC. A recombinant form ofhuman SPARC with a histidine tag (rh-SPARC-6xH) was pro-duced in a Drosophila melanogaster S3 expression system andpurified by metal chelation chromatography. A tag-free rh-SPARCwas obtained through digestion of a thrombin cleavage site engi-neered between the SPARC C-terminal amino acid and the hexa-histidine tail (Fig. 1a). Final yields varied between 10 and 17 mgL21 of serum-free conditioned medium of transfected S3 insectcells. Human Mel-SPARC and rh-SPARC were recognized by arabbit anti-SPARC antiserum (Fig. 1b). rh-SPARC showed aslightly retarded electrophoretic mobility compared to hMel-SPARC, most probably due to differences in glycosylation (Figs.1a and 1b).
Human Mel-SPARC and rh-SPARC (either with or without the6xHis tag) showed superimposable CD spectra in the far-UVregion (Fig. 1c). These spectra were fully compatible with thosereported for other natural and recombinant SPARC proteins.34,35,38
Moreover, thermal denaturation curves also exhibited a similaroverall shape for hMel-SPARC and both forms of rh-SPARC, fur-ther suggesting that thermal stability of all 3 forms is similar (Fig.1c, inset). Fluorescence spectra of rh-SPARC with or without6xHis and hMel-SPARC were also coincident in their maxima at324 nm (not shown). These data indicate that native and recombi-nant SPARC have nearly identical conformation.
1467SPARC EFFECTS ON TUMOR-ASSOCIATED STROMAL CELLS
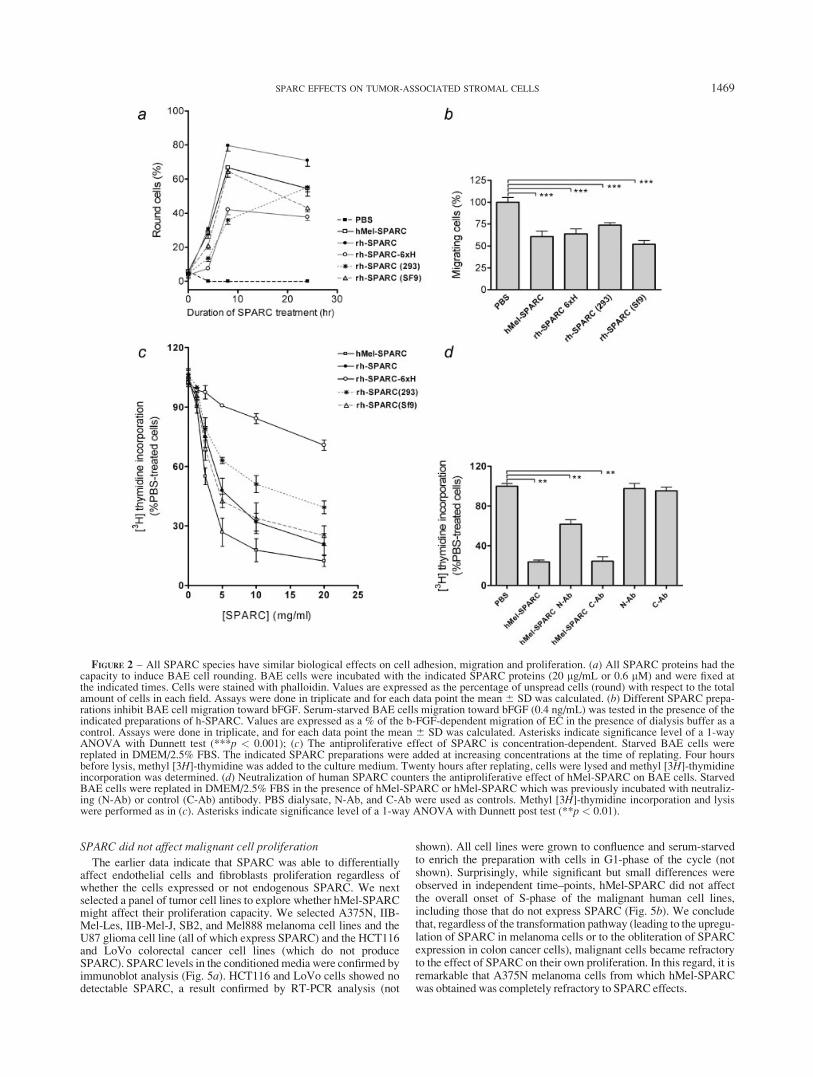
Five different SPARC species were effective in the inhibitionof bovine aortic endothelial cell spreading, migration andproliferation
We first compared hMel-SPARC and rh-SPARC with and with-out the histidine tag with 2 previously described recombinant pro-teins (rh-SPARC (SF9)35) and (rh-SPARC (293)36) with respect toa well-characterized property of SPARC, e.g., the inhibition ofBAE cell spreading.3 All SPARC species promoted rounding ofadherent BAE cells, though there were differences in the timecourse of the effect (Fig. 2a). Eight hours after addition ofSPARC, rh-SPARC, hMel-SPARC and rh-SPARC(SF9) inducedrounding in 65–80% of BAE cells compared to 35–40% cellrounding induced by rh-SPARC-6xH and rh-SPARC(293). rh-SPARC had the greatest rounding effect at the end of the experi-ment (Fig. 2a). Thus, despite the differences, all SPARC proteinsexhibited an intrinsic antiadhesive activity. Similar effects wereobserved when inhibition of cell spreading was analyzed, that is
all SPARC species inhibited BAE spreading (SupplementaryFig. 1).
We next assessed the effect of the different SPARC species onBAE cell migration in modified Boyden chambers. For this pur-pose, BAE cells were placed in the upper chamber in the presenceor absence of the different SPARC species. Figure 2b demon-strates that the different SPARC species were able to inhibit BAEcell migration toward bFGF to a very similar extent (Fig. 2b).
Next, we assessed SPARC effect on the reenter to the cell cycleof serum-starved, G1-arrested BAE cells. All SPARC speciesexerted a significant inhibition in the onset of S-phase in BAEcells (Supplementary Fig. 2a). The reduction in DNA synthesis bySPARC was also detected with exponentially growing cells, indi-cating that SPARC might not only sustain but also trigger BAEcell cycle arrest (Supplementary Figs. 2c and 2d). A concentrationanalysis carried out with the different SPARC species showed thatall the recombinant SPARC species retarded cell cycle reentry to alesser extent than that observed with hMel-SPARC (Fig. 2c). Inaddition, removal of the histidine tag significantly increased theantiproliferative activity of rh-SPARC (Fig. 2c). Preincubation ofhMel-SPARC and rh-SPARC with neutralizing 236 antibody46
significantly reverted the inhibitory effect of both SPARC proteins(Fig. 2d and Supplementary Fig. 2b, respectively). Similar resultswere obtained with another SPARC-N-Ab-29346 (not shown).Addition of anti-SPARC antibodies in the absence of exogenousSPARC did not affect BAEC-proliferation rate (Fig. 2d and Sup-plementary Fig. 2b), indicating that the amount of SPARC accu-mulated after replating does not contribute to the proliferationrates observed in our studies. Thus, despite the differencesobserved, it can be concluded that all SPARC species substantiallyinhibited spreading, migration and proliferation of BAE cells.
SPARC inhibited cell cycle reentrance of both normal andtransformed human endothelial cells
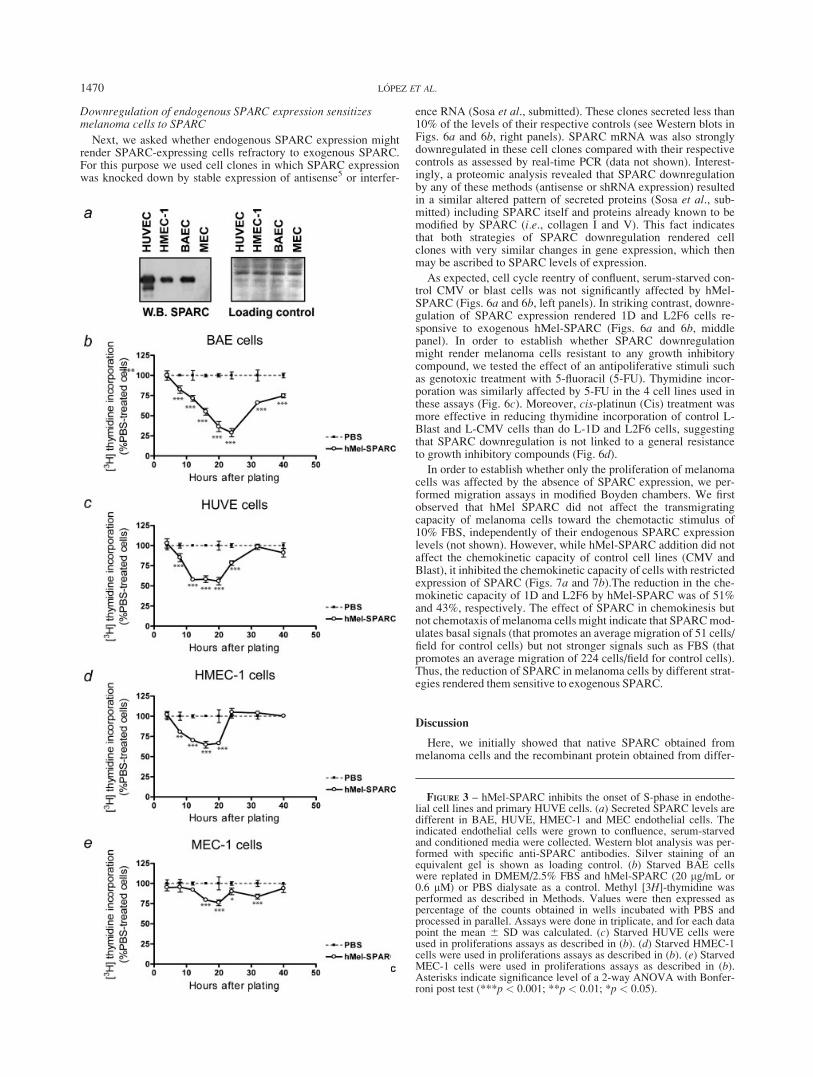
Since SPARC obtained from different sources exhibited essen-tially similar effects on the behavior of endothelial cells, we nextasked whether different endothelial cell types may respond differ-ently to SPARC. For these studies we selected hMel-SPARC,since it appeared very effective when compared to the otherSPARC species and because it was obtained from a malignant cellsource. We tested hMel-SPARC effect on normal human umbili-cal endothelial cells HUVEC and SV-40 large T antigen-trans-formed human and murine microendothelial cells HMEC-1 andMEC-1, respectively. BAEC, HUVEC and HMEC-1 cells showedhigh levels of secreted SPARC, while MEC-1 showed undetect-able levels of SPARC in their conditioned media (Fig. 3a). Cellcycle reentry was inhibited by hMel-SPARC in all cases, althoughnontransformed cells (BAEC and HUVEC) were more sensitive toSPARC antiproliferative effect (Figure 3b–e). These data clearlydemonstrate that SPARC can negatively modulate the cell cyclingcapacity of endothelial cells of different origins and levels oftransformation, independently of their endogenous SPARCexpression levels.
SPARC induced a biphasic modulation of fibroblasts proliferation
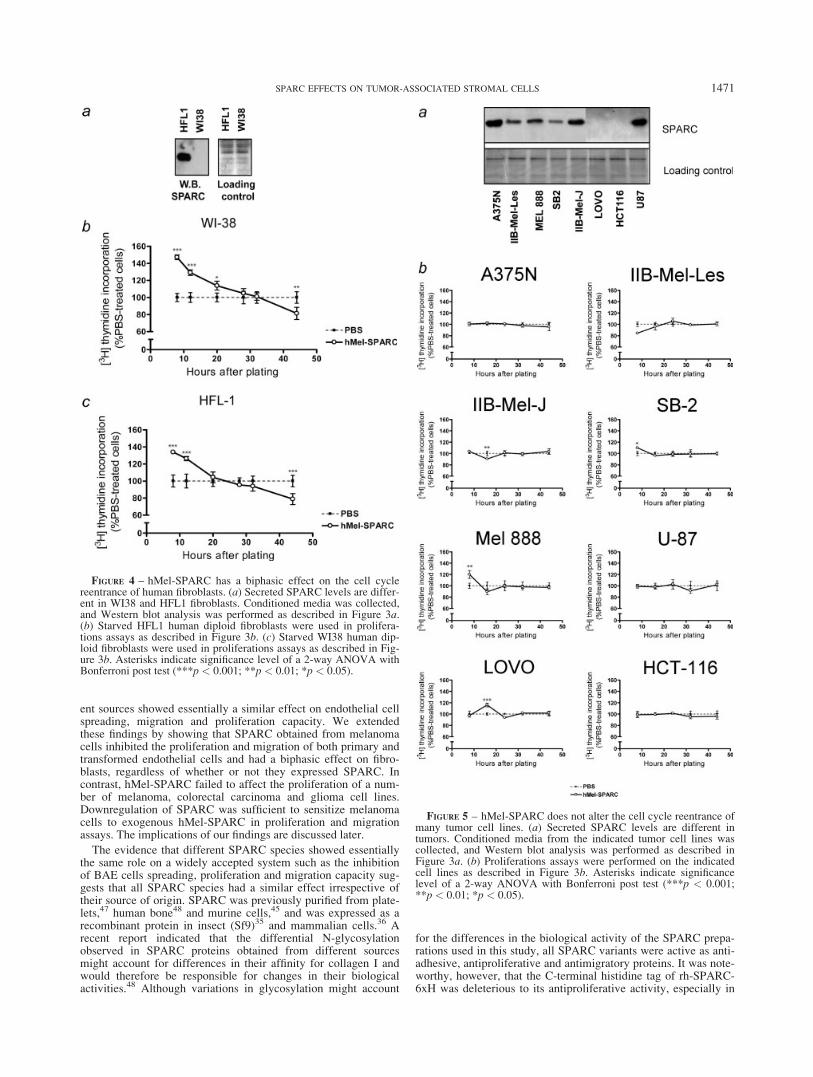
We next assessed the effect of hMel-SPARC on the cell cycleof other major stromal component such as fibroblasts. Two differ-ent human fetal fibroblasts, HFL-1 and WI38, secreting or notsecreting detectable amounts of SPARC, were used for theseexperiments (Fig. 4a). Fibroblasts were grown to confluence andserum-starved. In both cell types, hMel-SPARC promoted anincreased uptake of [3H]-thymidine during an initial period of20 hr, whereas it exhibited either no effect or an antiproliferativeeffect at subsequent time-points (Figs. 4b and 4c). The maximumdifference between SPARC- and PBS-treated WI38 and HFL-1cells was at the earliest time-point (8 hr), indicating that SPARCcan accelerate the initial events of DNA replication in both typesof fibroblasts independently of whether they expressed or notSPARC.
FIGURE 1 – Native SPARC purified from A375 melanoma cell con-ditioned medium is biochemically similar to recombinant versionsobtained from ectopic expression in Drosophila S3 cells. (a) ReducedSDS-PAGE (left) shows purity of preparations of human SPARC puri-fied from A375 cells (hMel-SPARC), recombinant SPARC with a C-terminal histidine tag (rh-SPARC-6xH) and rh-SPARC-6xH digestedwith thrombin to remove the His tag (rh-SPARC). (b) Western blot ofsamples shown in (a). (c) Circular dichroism spectra of SPARCproteins show that all preparations have similar secondary structure(dotted line, hMel-SPARC; straight line, rh-SPARC-6xH; broken line,rh-SPARC). Inset: Thermal denaturation of SPARC preparations fol-lowed by circular dichroism measurements at 220 nm (triangles,hMel-SPARC; circles, rh-SPARC-6xH; squares rh-SPARC).
1468 L�OPEZ ET AL.
SPARC did not affect malignant cell proliferation
The earlier data indicate that SPARC was able to differentiallyaffect endothelial cells and fibroblasts proliferation regardless ofwhether the cells expressed or not endogenous SPARC. We nextselected a panel of tumor cell lines to explore whether hMel-SPARCmight affect their proliferation capacity. We selected A375N, IIB-Mel-Les, IIB-Mel-J, SB2, and Mel888 melanoma cell lines and theU87 glioma cell line (all of which express SPARC) and the HCT116and LoVo colorectal cancer cell lines (which do not produceSPARC). SPARC levels in the conditioned media were confirmed byimmunoblot analysis (Fig. 5a). HCT116 and LoVo cells showed nodetectable SPARC, a result confirmed by RT-PCR analysis (not
shown). All cell lines were grown to confluence and serum-starvedto enrich the preparation with cells in G1-phase of the cycle (notshown). Surprisingly, while significant but small differences wereobserved in independent time–points, hMel-SPARC did not affectthe overall onset of S-phase of the malignant human cell lines,including those that do not express SPARC (Fig. 5b). We concludethat, regardless of the transformation pathway (leading to the upregu-lation of SPARC in melanoma cells or to the obliteration of SPARCexpression in colon cancer cells), malignant cells became refractoryto the effect of SPARC on their own proliferation. In this regard, it isremarkable that A375N melanoma cells from which hMel-SPARCwas obtained was completely refractory to SPARC effects.
FIGURE 2 – All SPARC species have similar biological effects on cell adhesion, migration and proliferation. (a) All SPARC proteins had thecapacity to induce BAE cell rounding. BAE cells were incubated with the indicated SPARC proteins (20 lg/mL or 0.6 lM) and were fixed atthe indicated times. Cells were stained with phalloidin. Values are expressed as the percentage of unspread cells (round) with respect to the totalamount of cells in each field. Assays were done in triplicate and for each data point the mean 6 SD was calculated. (b) Different SPARC prepa-rations inhibit BAE cell migration toward bFGF. Serum-starved BAE cells migration toward bFGF (0.4 ng/mL) was tested in the presence of theindicated preparations of h-SPARC. Values are expressed as a % of the b-FGF-dependent migration of EC in the presence of dialysis buffer as acontrol. Assays were done in triplicate, and for each data point the mean 6 SD was calculated. Asterisks indicate significance level of a 1-wayANOVA with Dunnett test (***p < 0.001); (c) The antiproliferative effect of SPARC is concentration-dependent. Starved BAE cells werereplated in DMEM/2.5% FBS. The indicated SPARC preparations were added at increasing concentrations at the time of replating. Four hoursbefore lysis, methyl [3H]-thymidine was added to the culture medium. Twenty hours after replating, cells were lysed and methyl [3H]-thymidineincorporation was determined. (d) Neutralization of human SPARC counters the antiproliferative effect of hMel-SPARC on BAE cells. StarvedBAE cells were replated in DMEM/2.5% FBS in the presence of hMel-SPARC or hMel-SPARC which was previously incubated with neutraliz-ing (N-Ab) or control (C-Ab) antibody. PBS dialysate, N-Ab, and C-Ab were used as controls. Methyl [3H]-thymidine incorporation and lysiswere performed as in (c). Asterisks indicate significance level of a 1-way ANOVA with Dunnett post test (**p < 0.01).
1469SPARC EFFECTS ON TUMOR-ASSOCIATED STROMAL CELLS
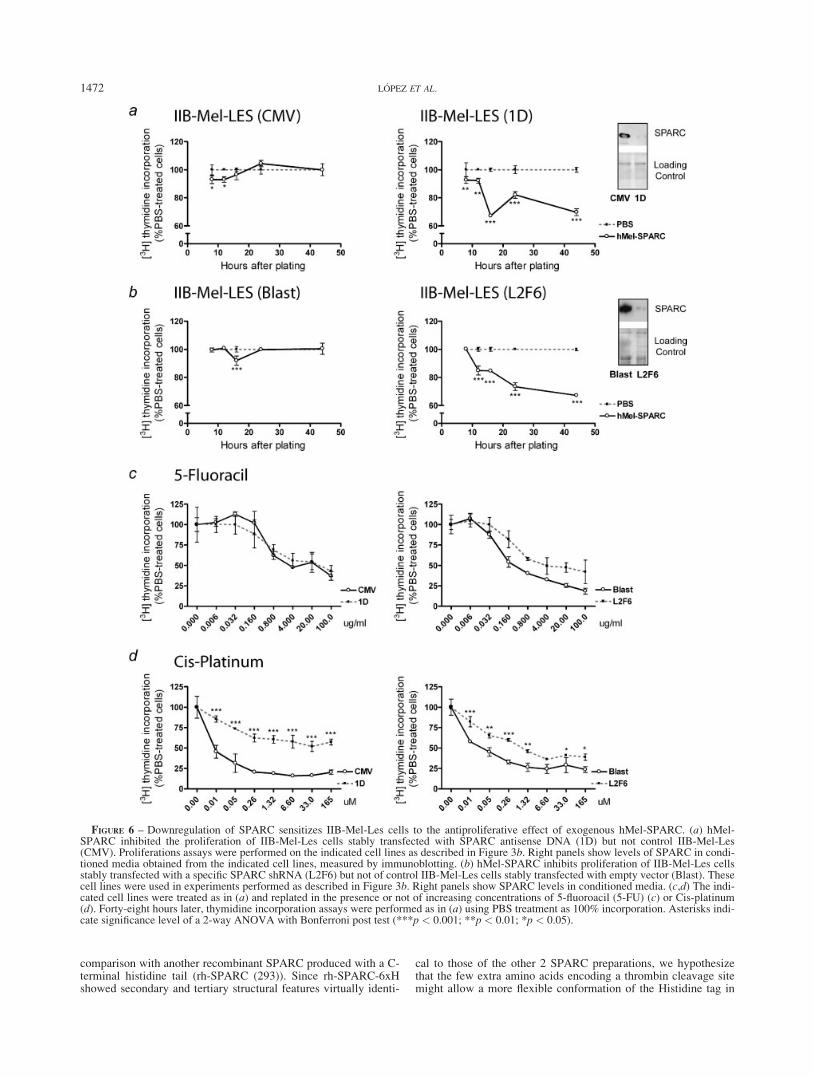
Downregulation of endogenous SPARC expression sensitizesmelanoma cells to SPARC
Next, we asked whether endogenous SPARC expression mightrender SPARC-expressing cells refractory to exogenous SPARC.For this purpose we used cell clones in which SPARC expressionwas knocked down by stable expression of antisense5 or interfer-
ence RNA (Sosa et al., submitted). These clones secreted less than10% of the levels of their respective controls (see Western blots inFigs. 6a and 6b, right panels). SPARC mRNA was also stronglydownregulated in these cell clones compared with their respectivecontrols as assessed by real-time PCR (data not shown). Interest-ingly, a proteomic analysis revealed that SPARC downregulationby any of these methods (antisense or shRNA expression) resultedin a similar altered pattern of secreted proteins (Sosa et al., sub-mitted) including SPARC itself and proteins already known to bemodified by SPARC (i.e., collagen I and V). This fact indicatesthat both strategies of SPARC downregulation rendered cellclones with very similar changes in gene expression, which thenmay be ascribed to SPARC levels of expression.
As expected, cell cycle reentry of confluent, serum-starved con-trol CMV or blast cells was not significantly affected by hMel-SPARC (Figs. 6a and 6b, left panels). In striking contrast, downre-gulation of SPARC expression rendered 1D and L2F6 cells re-sponsive to exogenous hMel-SPARC (Figs. 6a and 6b, middlepanel). In order to establish whether SPARC downregulationmight render melanoma cells resistant to any growth inhibitorycompound, we tested the effect of an antipoliferative stimuli suchas genotoxic treatment with 5-fluoracil (5-FU). Thymidine incor-poration was similarly affected by 5-FU in the 4 cell lines used inthese assays (Fig. 6c). Moreover, cis-platinun (Cis) treatment wasmore effective in reducing thymidine incorporation of control L-Blast and L-CMV cells than do L-1D and L2F6 cells, suggestingthat SPARC downregulation is not linked to a general resistanceto growth inhibitory compounds (Fig. 6d).
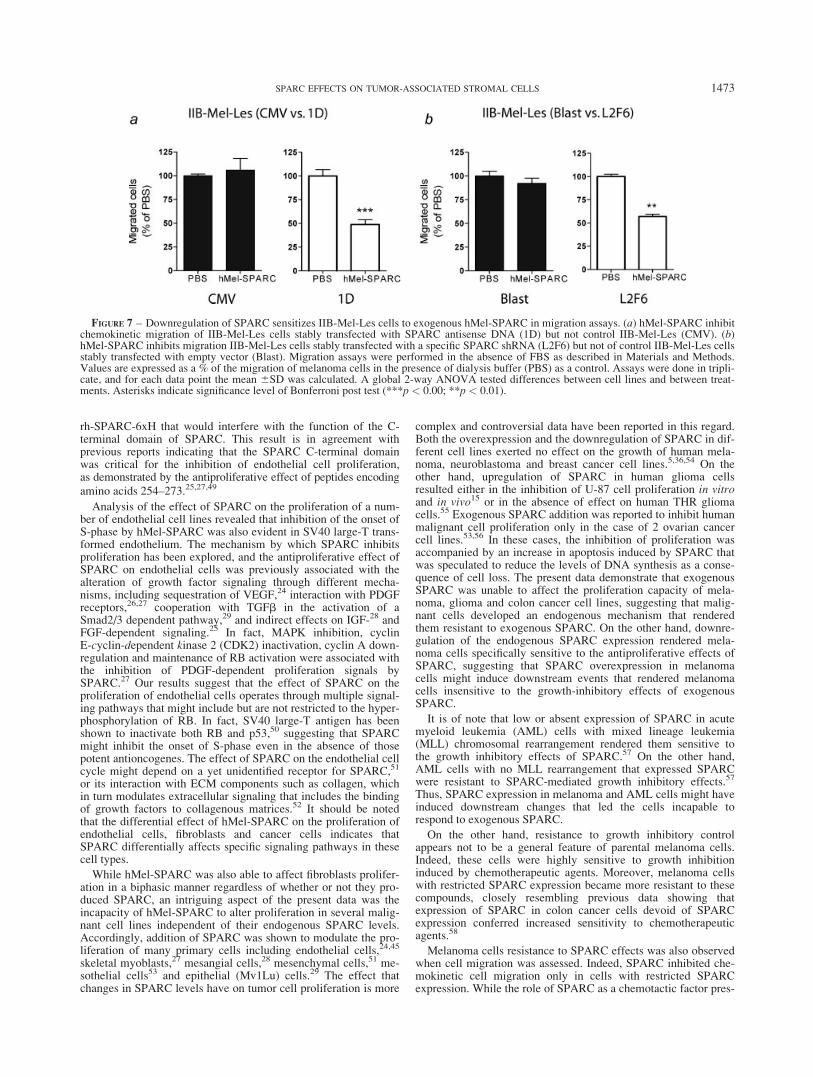
In order to establish whether only the proliferation of melanomacells was affected by the absence of SPARC expression, we per-formed migration assays in modified Boyden chambers. We firstobserved that hMel SPARC did not affect the transmigratingcapacity of melanoma cells toward the chemotactic stimulus of10% FBS, independently of their endogenous SPARC expressionlevels (not shown). However, while hMel-SPARC addition did notaffect the chemokinetic capacity of control cell lines (CMV andBlast), it inhibited the chemokinetic capacity of cells with restrictedexpression of SPARC (Figs. 7a and 7b).The reduction in the che-mokinetic capacity of 1D and L2F6 by hMel-SPARC was of 51%and 43%, respectively. The effect of SPARC in chemokinesis butnot chemotaxis of melanoma cells might indicate that SPARCmod-ulates basal signals (that promotes an average migration of 51 cells/field for control cells) but not stronger signals such as FBS (thatpromotes an average migration of 224 cells/field for control cells).Thus, the reduction of SPARC in melanoma cells by different strat-egies rendered them sensitive to exogenous SPARC.
Discussion
Here, we initially showed that native SPARC obtained frommelanoma cells and the recombinant protein obtained from differ-
FIGURE 3 – hMel-SPARC inhibits the onset of S-phase in endothe-lial cell lines and primary HUVE cells. (a) Secreted SPARC levels aredifferent in BAE, HUVE, HMEC-1 and MEC endothelial cells. Theindicated endothelial cells were grown to confluence, serum-starvedand conditioned media were collected. Western blot analysis was per-formed with specific anti-SPARC antibodies. Silver staining of anequivalent gel is shown as loading control. (b) Starved BAE cellswere replated in DMEM/2.5% FBS and hMel-SPARC (20 lg/mL or0.6 lM) or PBS dialysate as a control. Methyl [3H]-thymidine wasperformed as described in Methods. Values were then expressed aspercentage of the counts obtained in wells incubated with PBS andprocessed in parallel. Assays were done in triplicate, and for each datapoint the mean 6 SD was calculated. (c) Starved HUVE cells wereused in proliferations assays as described in (b). (d) Starved HMEC-1cells were used in proliferations assays as described in (b). (e) StarvedMEC-1 cells were used in proliferations assays as described in (b).Asterisks indicate significance level of a 2-way ANOVA with Bonfer-roni post test (***p < 0.001; **p < 0.01; *p < 0.05).
1470 L�OPEZ ET AL.
ent sources showed essentially a similar effect on endothelial cellspreading, migration and proliferation capacity. We extendedthese findings by showing that SPARC obtained from melanomacells inhibited the proliferation and migration of both primary andtransformed endothelial cells and had a biphasic effect on fibro-blasts, regardless of whether or not they expressed SPARC. Incontrast, hMel-SPARC failed to affect the proliferation of a num-ber of melanoma, colorectal carcinoma and glioma cell lines.Downregulation of SPARC was sufficient to sensitize melanomacells to exogenous hMel-SPARC in proliferation and migrationassays. The implications of our findings are discussed later.
The evidence that different SPARC species showed essentiallythe same role on a widely accepted system such as the inhibitionof BAE cells spreading, proliferation and migration capacity sug-gests that all SPARC species had a similar effect irrespective oftheir source of origin. SPARC was previously purified from plate-lets,47 human bone48 and murine cells,45 and was expressed as arecombinant protein in insect (Sf9)35 and mammalian cells.36 Arecent report indicated that the differential N-glycosylationobserved in SPARC proteins obtained from different sourcesmight account for differences in their affinity for collagen I andwould therefore be responsible for changes in their biologicalactivities.48 Although variations in glycosylation might account
for the differences in the biological activity of the SPARC prepa-rations used in this study, all SPARC variants were active as anti-adhesive, antiproliferative and antimigratory proteins. It was note-worthy, however, that the C-terminal histidine tag of rh-SPARC-6xH was deleterious to its antiproliferative activity, especially in
FIGURE 4 – hMel-SPARC has a biphasic effect on the cell cyclereentrance of human fibroblasts. (a) Secreted SPARC levels are differ-ent in WI38 and HFL1 fibroblasts. Conditioned media was collected,and Western blot analysis was performed as described in Figure 3a.(b) Starved HFL1 human diploid fibroblasts were used in prolifera-tions assays as described in Figure 3b. (c) Starved WI38 human dip-loid fibroblasts were used in proliferations assays as described in Fig-ure 3b. Asterisks indicate significance level of a 2-way ANOVA withBonferroni post test (***p < 0.001; **p < 0.01; *p < 0.05).
FIGURE 5 – hMel-SPARC does not alter the cell cycle reentrance ofmany tumor cell lines. (a) Secreted SPARC levels are different intumors. Conditioned media from the indicated tumor cell lines wascollected, and Western blot analysis was performed as described inFigure 3a. (b) Proliferations assays were performed on the indicatedcell lines as described in Figure 3b. Asterisks indicate significancelevel of a 2-way ANOVA with Bonferroni post test (***p < 0.001;**p < 0.01; *p < 0.05).
1471SPARC EFFECTS ON TUMOR-ASSOCIATED STROMAL CELLS
comparison with another recombinant SPARC produced with a C-terminal histidine tail (rh-SPARC (293)). Since rh-SPARC-6xHshowed secondary and tertiary structural features virtually identi-
cal to those of the other 2 SPARC preparations, we hypothesizethat the few extra amino acids encoding a thrombin cleavage sitemight allow a more flexible conformation of the Histidine tag in
FIGURE 6 – Downregulation of SPARC sensitizes IIB-Mel-Les cells to the antiproliferative effect of exogenous hMel-SPARC. (a) hMel-SPARC inhibited the proliferation of IIB-Mel-Les cells stably transfected with SPARC antisense DNA (1D) but not control IIB-Mel-Les(CMV). Proliferations assays were performed on the indicated cell lines as described in Figure 3b. Right panels show levels of SPARC in condi-tioned media obtained from the indicated cell lines, measured by immunoblotting. (b) hMel-SPARC inhibits proliferation of IIB-Mel-Les cellsstably transfected with a specific SPARC shRNA (L2F6) but not of control IIB-Mel-Les cells stably transfected with empty vector (Blast). Thesecell lines were used in experiments performed as described in Figure 3b. Right panels show SPARC levels in conditioned media. (c,d) The indi-cated cell lines were treated as in (a) and replated in the presence or not of increasing concentrations of 5-fluoroacil (5-FU) (c) or Cis-platinum(d). Forty-eight hours later, thymidine incorporation assays were performed as in (a) using PBS treatment as 100% incorporation. Asterisks indi-cate significance level of a 2-way ANOVA with Bonferroni post test (***p < 0.001; **p < 0.01; *p < 0.05).
1472 L�OPEZ ET AL.
rh-SPARC-6xH that would interfere with the function of the C-terminal domain of SPARC. This result is in agreement withprevious reports indicating that the SPARC C-terminal domainwas critical for the inhibition of endothelial cell proliferation,as demonstrated by the antiproliferative effect of peptides encodingamino acids 254–273.25,27,49
Analysis of the effect of SPARC on the proliferation of a num-ber of endothelial cell lines revealed that inhibition of the onset ofS-phase by hMel-SPARC was also evident in SV40 large-T trans-formed endothelium. The mechanism by which SPARC inhibitsproliferation has been explored, and the antiproliferative effect ofSPARC on endothelial cells was previously associated with thealteration of growth factor signaling through different mecha-nisms, including sequestration of VEGF,24 interaction with PDGFreceptors,26,27 cooperation with TGFb in the activation of aSmad2/3 dependent pathway,29 and indirect effects on IGF-28 andFGF-dependent signaling.25 In fact, MAPK inhibition, cyclinE-cyclin-dependent kinase 2 (CDK2) inactivation, cyclin A down-regulation and maintenance of RB activation were associated withthe inhibition of PDGF-dependent proliferation signals bySPARC.27 Our results suggest that the effect of SPARC on theproliferation of endothelial cells operates through multiple signal-ing pathways that might include but are not restricted to the hyper-phosphorylation of RB. In fact, SV40 large-T antigen has beenshown to inactivate both RB and p53,50 suggesting that SPARCmight inhibit the onset of S-phase even in the absence of thosepotent antioncogenes. The effect of SPARC on the endothelial cellcycle might depend on a yet unidentified receptor for SPARC,51
or its interaction with ECM components such as collagen, whichin turn modulates extracellular signaling that includes the bindingof growth factors to collagenous matrices.52 It should be notedthat the differential effect of hMel-SPARC on the proliferation ofendothelial cells, fibroblasts and cancer cells indicates thatSPARC differentially affects specific signaling pathways in thesecell types.
While hMel-SPARC was also able to affect fibroblasts prolifer-ation in a biphasic manner regardless of whether or not they pro-duced SPARC, an intriguing aspect of the present data was theincapacity of hMel-SPARC to alter proliferation in several malig-nant cell lines independent of their endogenous SPARC levels.Accordingly, addition of SPARC was shown to modulate the pro-liferation of many primary cells including endothelial cells,24,45
skeletal myoblasts,27 mesangial cells,28 mesenchymal cells,51 me-sothelial cells53 and epithelial (Mv1Lu) cells.29 The effect thatchanges in SPARC levels have on tumor cell proliferation is more
complex and controversial data have been reported in this regard.Both the overexpression and the downregulation of SPARC in dif-ferent cell lines exerted no effect on the growth of human mela-noma, neuroblastoma and breast cancer cell lines.5,36,54 On theother hand, upregulation of SPARC in human glioma cellsresulted either in the inhibition of U-87 cell proliferation in vitroand in vivo15 or in the absence of effect on human THR gliomacells.55 Exogenous SPARC addition was reported to inhibit humanmalignant cell proliferation only in the case of 2 ovarian cancercell lines.53,56 In these cases, the inhibition of proliferation wasaccompanied by an increase in apoptosis induced by SPARC thatwas speculated to reduce the levels of DNA synthesis as a conse-quence of cell loss. The present data demonstrate that exogenousSPARC was unable to affect the proliferation capacity of mela-noma, glioma and colon cancer cell lines, suggesting that malig-nant cells developed an endogenous mechanism that renderedthem resistant to exogenous SPARC. On the other hand, downre-gulation of the endogenous SPARC expression rendered mela-noma cells specifically sensitive to the antiproliferative effects ofSPARC, suggesting that SPARC overexpression in melanomacells might induce downstream events that rendered melanomacells insensitive to the growth-inhibitory effects of exogenousSPARC.
It is of note that low or absent expression of SPARC in acutemyeloid leukemia (AML) cells with mixed lineage leukemia(MLL) chromosomal rearrangement rendered them sensitive tothe growth inhibitory effects of SPARC.57 On the other hand,AML cells with no MLL rearrangement that expressed SPARCwere resistant to SPARC-mediated growth inhibitory effects.57
Thus, SPARC expression in melanoma and AML cells might haveinduced downstream changes that led the cells incapable torespond to exogenous SPARC.
On the other hand, resistance to growth inhibitory controlappears not to be a general feature of parental melanoma cells.Indeed, these cells were highly sensitive to growth inhibitioninduced by chemotherapeutic agents. Moreover, melanoma cellswith restricted SPARC expression became more resistant to thesecompounds, closely resembling previous data showing thatexpression of SPARC in colon cancer cells devoid of SPARCexpression conferred increased sensitivity to chemotherapeuticagents.58
Melanoma cells resistance to SPARC effects was also observedwhen cell migration was assessed. Indeed, SPARC inhibited che-mokinetic cell migration only in cells with restricted SPARCexpression. While the role of SPARC as a chemotactic factor pres-
FIGURE 7 – Downregulation of SPARC sensitizes IIB-Mel-Les cells to exogenous hMel-SPARC in migration assays. (a) hMel-SPARC inhibitchemokinetic migration of IIB-Mel-Les cells stably transfected with SPARC antisense DNA (1D) but not control IIB-Mel-Les (CMV). (b)hMel-SPARC inhibits migration IIB-Mel-Les cells stably transfected with a specific SPARC shRNA (L2F6) but not of control IIB-Mel-Les cellsstably transfected with empty vector (Blast). Migration assays were performed in the absence of FBS as described in Materials and Methods.Values are expressed as a % of the migration of melanoma cells in the presence of dialysis buffer (PBS) as a control. Assays were done in tripli-cate, and for each data point the mean 6SD was calculated. A global 2-way ANOVA tested differences between cell lines and between treat-ments. Asterisks indicate significance level of Bonferroni post test (***p < 0.00; **p < 0.01).
1473SPARC EFFECTS ON TUMOR-ASSOCIATED STROMAL CELLS

ent in bone extracts is still controversial,59,60 recent data hasshown that SPARC accelerated breast cancer cell motility in a‘‘wound healing’’ assay.60 On the other hand, SPARC was able toinhibit the migration of ovarian cancer cells induced by mesothe-lial cells through a mechanism mediated by lysophosphatidicacid.61 Interestingly, recent studies suggest that SPARC mightbind to the cell surface membrane of breast cancer cells62 as itdoes to endothelial cells,63 suggesting that SPARC effect on cellmigration might greatly depend on its capacity to bind to the targetcell through a yet unidentified receptor. Our data obtained in mela-noma cells might also indicate that the levels or availability ofSPARC receptor could be also modulated by SPARC levels. Infact, a potentially coordinated downregulation of SPARC proteinand its receptor in cancer cells that do not express SPARC mightalso account for the inability of SPARC to regulate their cellcycle. Alternatively, the possibility exists that the different cellsexhibited different capacity to cleave and inactivate SPARC.5,6
However, western analysis of cleavage of exogenous SPARC byconditioned media obtained from fibroblasts, endothelial cells,malignant cells expressing or not SPARC and melanoma cellswith restricted SPARC expression showed essentially no differ-ence, suggesting that SPARC cleavage by malignant cells is notthe mechanism by which malignant cells became SPARC-resistant(data not shown).
Our data suggest that overexpression of SPARC by melanomacells renders melanoma cells resistant to the effect of exogenousSPARC. The question whether SPARC overexpression can lead tofull receptor occupancy or induce downstream events affecting re-ceptor availability is open. Downregulation of SPARC levels can
induce qualititative and/or quantitative changes in additional pro-teins that might play a role in melanoma resistance to SPARC lev-els. In fact, proteomic analysis of secreted proteins followingdownregulation of SPARC expression in melanoma cells showedat least 13 differentially secreted proteins (Sosa et al., submitted),indicating that changes in SPARC production concomitantly alterthe expression of a number of proteins that could modulate thecapacity of melanoma cells to grow in the presence of high levelsof SPARC. In accordance with this proposal, overexpression ofSPARC in breast cancer cells was shown to mediate metastaticgrowth in the lung only when coselected with a number of otherpartners, suggesting that SPARC could be coordinately modulatedwith other proteins to facilitate cancer progression.64
Acknowledgements
We thank especially Dr. Carlos Cerve~nansky, Instituto de Inves-tigaciones Biol�ogicas Clemente Estable, Montevideo, Uruguay,for performing peptide mass fingerprinting analysis on hMel-SPARC and Dr. Alexandre Chlenski, Robert H. Lurie Comprehen-sive Cancer Center, Northwestern University, Feinberg School ofMedicine, Chicago IL, for samples of recombinant SPARC. Wealso thank Ms. Mar�ıa Romina Girotti for the obtention and initialcharacterization of melanoma cells stably transfected withshRNA. This work was supported by grants from Fundaci�onAntorchas and UBACyT (to A.S.L.), National Institute of Healthgrant GM-40711 (to E.H.S.), CONICET and ANPCYT (to O.L.P.).We also thank the permanent support of Fundaci�on Rene Baronand AFULIC (to O.L.P.).
References
1. Liotta LA, Kohn EC. The microenvironment of the tumour-hostinterface. Nature 2001;411:375–9.
2. Tuxhorn JA, McAlhany SJ, Dang TD, Ayala GE, Rowley DR.Stromal cells promote angiogenesis and growth of human prostatetumors in a differential reactive stroma (DRS) xenograft model. Can-cer Res 2002;62:3298–307.
3. Framson PE, Sage EH. SPARC and tumor growth: where the seedmeets the soil? J Cell Biochem 2004;92:679–90; and referencestherein.
4. Bradshaw AD, Sage EH. SPARC, a matricellular protein that func-tions in cellular differentiation and tissue response to injury. J ClinInvest 2001;107:1049–54.
5. Ledda MF, Adris S, Bravo AI, Kairiyama C, Bover L, ChernajovskyY, Mordoh J, Podhajcer OL. Suppression of SPARC expression byantisense RNA abrogates the tumorigenicity of human melanomacells. Nat Med 1997;3:171–6.
6. Ledda F, Bravo AI, Adris S, Bover L, Mordoh J, Podhajcer OL. Theexpression of the secreted protein acidic and rich in cysteine (SPARC)is associated with the neoplastic progression of human melanoma.J Invest Dermatol 1997;108:210–14.
7. Golembieski WA, Ge S, Nelson K, Mikkelsen T, Rempel SA.Increased SPARC expression promotes U87 glioblastoma invasion invitro. Int J Dev Neurosci 1999;17:463–72.
8. Rempel SA, Ge S, Gutierrez JA. SPARC: a potential diagnosticmarker of invasive meningiomas. Clin Cancer Res 1999;5:237–41.
9. Bellahcene A, Castronovo V. Increased expression of osteonectin andosteopontin, two bone matrix proteins, in human breast cancer. AmJ Pathol 1995;146:95–100.
10. Porte H, Chastre E, Prevot S, Nordlinger B, Empereur S, Basset P,Chambon P, Gespach C. Neoplastic progression of human colorectalcancer is associated with overexpression of the stromelysin-3 andBM-40/SPARC genes. Int J Cancer 1995;64:70–5.
11. Koukourakis MI, Giatromanolaki A, Brekken RA, Sivridis E, GatterKC, Harris AL, Sage EH. Enhanced expression of SPARC/osteonec-tin in the tumor-associated stroma of non-small cell lung cancer iscorrelated with markers of hypoxia/acidity and with poor prognosis ofpatients. Cancer Res 2003;63:5376–80.
12. Sato N, Fukushima N, Maehara N, Matsubayashi H, Koopmann J, SuGH, Hruban RH, Goggins M. SPARC/osteonectin is a frequent targetfor aberrant methylation in pancreatic adenocarcinoma and a mediatorof tumor-stromal interactions. Oncogene 2003;22:5021–30.
13. De Vos J, Thykjaer T, Tarte K, Ensslen M, Raynaud P, Requirand G,Pellet F, Pantesco V, Reme T, Jourdan M, Rossi JF, Orntoft T, et al.Comparison of gene expression profiling between malignant and
normal plasma cells with oligonucleotide arrays. Oncogene 2002;21:6848–57.
14. Thomas R, True LD, Bassuk JA, Lange PH, Vessella RL. Differentialexpression of osteonectin/SPARC during human prostate cancerprogression. Clin Cancer Res 2000;6:1140–9.
15. Rempel SA, Golembieski WA, Fisher JL, Maile M, Nakeff A. SPARCmodulates cell growth, attachment and migration of U87 glioma cellson brain extracellular matrix proteins. J Neurooncol 2001;53:149–60.
16. Shi Q, Bao S, Maxwell JA, Reese ED, Friedman HS, Bigner DD,Wang XF, Rich JN. Secreted protein acidic, rich in cysteine (SPARC),mediates cellular survival of gliomas through AKT activation. J BiolChem 2004;279:52200–9.
17. Alvarez MJ, Prada F, Salvatierra E, Bravo AI, Lutzky VP, Carbone C,Pitossi FJ, Chuluyan HE, Podhajcer OL. Secreted protein acidic andrich in cysteine produced by human melanoma cells modulates poly-morphonuclear leukocyte recruitment and antitumor cytotoxiccapacity. Cancer Res 2005;65:5123–32.
18. Mok SC, Chan WY, Wong KK, Muto MG, Berkowitz RS. SPARC,an extracellular matrix protein with tumor-suppressing activity inhuman ovarian epithelial cells. Oncogene 1996;12:1895–901.
19. Chlenski A, Liu S, Baker LJ, Yang Q, Tian Y, Salwen HR, Cohn SL.Neuroblastoma angiogenesis is inhibited with a folded synthetic mole-cule corresponding to the epidermal growth factor-like module of thefollistatin domain of SPARC. Cancer Res 2004;64:7420–5.
20. Sangaletti S, Stoppacciaro A, Guiducci C, Torrisi MR, Colombo MP.Leukocyte, rather than tumor-produced SPARC, determines stromaand collagen type IV deposition in mammary carcinoma. J Exp Med2003;198:1475–85.
21. Infante JR, Matsubayashi H, Sato N, Tonascia J, Klein AP, Riall TA,Yeo C, Iacobuzio-Donahue C, Goggins M. Peritumoral fibroblastSPARC expression and patient outcome with resectable pancreaticadenocarcinoma. J Clin Oncol 2007;25:319–25.
22. Motamed K. SPARC (osteonectin/BM-40). Int J Biochem Cell Biol1999;31:1363–6.
23. Yan Q, Sage EH. SPARC, a matricellular glycoprotein with importantbiological functions. J Histochem Cytochem 1999;47:1495–506.
24. Kupprion C, Motamed K, Sage EH. SPARC (BM-40, osteonectin)inhibits the mitogenic effect of vascular endothelial growth factor onmicrovascular endothelial cells. J Biol Chem 1998;273:29635–40.
25. Motamed K, Blake DJ, Angello JC, Allen BL, Rapraeger AC,Hauschka SD, Sage EH. Fibroblast growth factor receptor-1 mediatesthe inhibition of endothelial cell proliferation and the promotion ofskeletal myoblast differentiation by SPARC: a role for protein kinaseA. J Cell Biochem 2003;90:408–23.
1474 L�OPEZ ET AL.

26. Raines EW, Lane TF, Iruela-Arispe ML, Ross R, Sage EH. Theextracellular glycoprotein SPARC interacts with platelet-derivedgrowth factor (PDGF)-AB and -BB and inhibits the binding of PDGFto its receptors. Proc Natl Acad Sci USA 1992;89:1281–5.
27. Motamed K, Funk SE, Koyama H, Ross R, Raines EW, Sage EH. In-hibition of PDGF-stimulated and matrix-mediated proliferation ofhuman vascular smooth muscle cells by SPARC is independent ofchanges in cell shape or cyclin-dependent kinase inhibitors. J CellBiochem 2002;84:759–71.
28. Francki A, Motamed K, McClure TD, Kaya M, Murri C, Blake DJ,Carbon JG, Sage EH. SPARC regulates cell cycle progression in mes-angial cells via its inhibition of IGF-dependent signaling. J Cell Bio-chem 2003;88:802–11.
29. Schiemann BJ, Neil JR, Schiemann WP. SPARC inhibits epithelialcell proliferation in part through stimulation of the transforminggrowth factor- b-signaling system. Mol Biol Cell 2003;14:3977–88.
30. Bos TJ, Cohn SL, Kleinman HK, Murphy-Ulrich JE, Podhajcer OL,Rempel SA, Rich JN, Rutka JT, Sage EH, Thompson EW. Interna-tional Hermelin brain tumor symposium on matricellular proteins innormal and cancer cell-matrix interactions. Matrix Biol 2004;23:63–9.
31. Kelm RJ, Jr., Hair GA, Mann KG, Grant BW. Characterization ofhuman osteoblast and megakaryocyte-derived osteonectin (SPARC).Blood 1992;80:3112–19.
32. Nischt R, Pottgiesser J, Krieg T, Mayer U, Aumailley M, Timpl R.Recombinant expression and properties of the human calcium-bindingextracellular matrix protein BM-40. Eur J Biochem 1991;200:529–36.
33. Yost JC, Bell A, Seale R, Sage EH. Purification of biologically activeSPARC expressed in Saccharomyces cerevisiae. Arch Biochem Bio-phys 1994;314:50–63.
34. Bassuk JA, Baneyx F, Vernon RB, Funk SE, Sage EH. Expression ofbiologically active human SPARC in Escherichia coli. Arch BiochemBiophys 1996;325:8–19.
35. Bradshaw AD, Bassuk JA, Francki A, Sage EH. Expression and puri-fication of recombinant human SPARC produced by baculovirus. MolCell Biol Res Commun 2000;3:345–51.
36. Chlenski A, Liu S, Guerrero LJ, Yang Q, Tian Y, Salwen HR, Zage P,Cohn SL. SPARC expression is associated with impaired tumorgrowth, inhibited angiogenesis and changes in the extracellular ma-trix. Int J Cancer 2006;118:310–6.
37. Sage H, Vernon RB, Funk SE, Everitt EA, Angello J. SPARC, asecreted protein associated with cellular proliferation, inhibits cellspreading in vitro and exhibits Ca21-dependent binding to the extrac-ellular matrix. J Cell Biol 1989;109:341–56.
38. Engel J, Taylor W, Paulsson M, Sage H, Hogan B. Calcium-bindingdomains and calcium-induced transition in SPARC (osteonectin/BM40), an extracellular glycoprotein expressed in mineralized boneand nonmineralized tissues. Biochemistry 1987;26:6958–65.
39. Bunch TA, Grinblat Y, Goldstein LS. Characterization and use of theDrosophila metallothionein promoter in cultured Drosophila mela-nogaster cells. Nucleic Acids Res 1988;16:1043–61.
40. Bolander ME, Robey PG, Fisher LW, Conn KM, Prabhakar BS,Termine JD. Monoclonal antibodies against osteonectin show conser-vation of epitopes across species. Calcif Tissue Int 1989;45:74–80.
41. Jaffe EA, Nachman RL, Becker CG, Minick CR. Culture of humanendothelial cells derived from umbilical veins. Identification by mor-phologic and immunologic criteria. J Clin Invest 1973;52:2745–56.
42. Kairiyama C, Slavutsky I, Larripa I, Morvillo V, Bravo AI, Bover L,Podhajcer OL, Mordoh J. Biologic, immunocytochemical, and cyto-genetic characterization of two new human melanoma cell lines: IIB-MEL-LES and IIB-MEL-IAN. Pigment Cell Res 1995;8:121–31.
43. Guerra L, Mordoh J, Slavutsky I, Larripa I, Medrano EE. Characteri-zation of IIB-MEL-J: a new and highly heterogenous human mela-noma cell line. Pigment Cell Res 1989;2:504–9.
44. Funk SE, Sage EH. Differential effects of SPARC and cationicSPARC peptides on DNA synthesis by endothelial cells and fibro-blasts. J Cell Physiol 1993;154:53–63.
45. Funk SE, Sage EH. The Ca2(1)-binding glycoprotein SPARC modu-lates cell cycle progression in bovine aortic endothelial cells. ProcNatl Acad Sci USA 1991;88:2648–52.
46. Sweetwyne MT, Brekken RA, Workman G, Bradshaw AD, Carbon J,Siadak AW, Murri C, Sage EH. Functional analysis of the matricellu-lar protein SPARC with novel monoclonal antibodies. J HistochemCytochem 2004;52:723–33.
47. Chlenski A, Liu S, Crawford SE, Volpert OV, DeVries GH, Evangel-ista A, Yang Q, Salwen HR, Farrer R, Bray J, Cohn SL. SPARC is akey Schwannian-derived inhibitor controlling neuroblastoma tumorangiogenesis. Cancer Res 2002;62:7357–63.
48. Kaufmann B, Muller S, Hanisch FG, Hartmann U, Paulsson M,Maurer P, Zaucke F. Structural variability of BM-40/SPARC/osteo-nectin glycosylation: implications for collagen affinity. Glycobiology2004;14:609–19.
49. Sage EH, Reed M, Funk SE, Truong T, Steadele M, Puolakkainen P,Maurice DH, Bassuk JA. Cleavage of the matricellular proteinSPARC by matrix metalloproteinase 3 produces polypeptides thatinfluence angiogenesis. J Biol Chem 2003;278:37849–57.
50. Ahuja D, Saenz-Robles MT, Pipas JM. SV40 large T antigen targetsmultiple cellular pathways to elicit cellular transformation. Oncogene2005;24:7729–45.
51. Bradshaw AD, Francki A, Motamed K, Howe C, Sage EH. Primarymesenchymal cells isolated from SPARC-null mice exhibit alteredmorphology and rates of proliferation. Mol Biol Cell 1999;10:1569–79.
52. Ramirez F, Rifkin DB. Cell signaling events: a view from the matrix.Matrix Biol 2003;22:101–7.
53. Yiu GK, Chan WY, Ng SW, Chan PS, Cheung KK, Berkowitz RS,Mok SC. SPARC (secreted protein acidic and rich in cysteine) indu-ces apoptosis in ovarian cancer cells. Am J Pathol 2001;159:609–22.
54. Koblinski JE, Kaplan-Singer BR, VanOsdol SJ, Wu M, Engbring JA,Wang S, Goldsmith CM, Piper JT, Vostal JG, Harms JF, Welch DR,Kleinman HK. Endogenous osteonectin/SPARC/BM-40 expressioninhibits MDA-MB-231 breast cancer cell metastasis. Cancer Res2005;65:7370–7.
55. Rich JN, Shi Q, Hjelmeland M, Cummings TJ, Kuan CT, Bigner DD,Counter CM, Wang XF. Bone-related genes expressed in advancedmalignancies induce invasion and metastasis in a genetically definedhuman cancer model. J Biol Chem 2003;278:15951–7.
56. Said N, Motamed K. Absence of host-secreted protein acidic and richin cysteine (SPARC) augments peritoneal ovarian carcinomatosis.Am J Pathol 2005;167:1739–52.
57. DiMartino JF, Lacayo NJ, Varadi M, Li L, Saraiya C, RavindranathY, Yu R, Sikic BI, Raimondi SC, Dahl GV. Low or absent SPARCexpression in acute myeloid leukemia with MLL rearrangements isassociated with sensitivity to growth inhibition by exogenous SPARCprotein. Leukemia 2006;20:426–32.
58. Tai IT, Dai M, Owen DA, Chen LB. Genome-wide expression analy-sis of therapy-resistant tumors reveals SPARC as a novel target forcancer therapy. J Clin Invest 2005;115:1492–502.
59. De S, Chen J, Narizhneva NV, Heston W, Brainard J, Sage EH,Byzova TV. Molecular pathway for cancer metastasis to bone. J BiolChem 2003;278:39044–50.
60. Campo McKnight DA, Sosnoski DM, Koblinski JE, Gay CV. Rolesof osteonectin in the migration of breast cancer cells into bone. J CellBiochem 2006;97:288–302.
61. Said NA, Najwer I, Socha MJ, Fulton DJ, Mok SC, Motamed K.SPARC inhibits LPA-mediated mesothelial-ovarian cancer cell cross-talk. Neoplasia 2007;9:23–35.
62. Gradishar WJ. Albumin-bound paclitaxel: a next-generation taxane.Expert Opin Pharmacother 2006;7:1041–53.
63. Yost JC, Sage EH. Specific interaction of SPARC with endothelialcells is mediated through a carboxyl-terminal sequence containing acalcium-binding EF hand. J Biol Chem 1993;268:25790–6.
64. Minn AJ, Gupta GP, Siegel PM, Bos PD, Shu W, Giri DD, Viale A,Olshen AB, Gerald WL, Massague J. Genes that mediate breast can-cer metastasis to lung. Nature 2005;436:518–24.
1475SPARC EFFECTS ON TUMOR-ASSOCIATED STROMAL CELLS