increased fibrovascular invasion of subcutaneous polyvinyl alcohol sponges in sparc-null mice
TRANSCRIPT

Increased fibrovascular invasion of subcutaneous polyvinylalcohol sponges in SPARC-null mice
AMY D. BRADSHAW, PhDa; MAY J. REED, MDb; JULIET G. CARBON, BSca; EMMETT PINNEY, MScc;ROLF A. BREKKEN, PhDa; E. HELENE SAGE, PhDa
The expression of SPARC (secreted protein acidic and rich in cysteine/osteonectin/BM-40) is elevated in endothelialcells participating in angiogenesis in vitro and in vivo. SPARC acts on endothelial cells to elicit changes in cellshape and to inhibit cell cycle progression. In addition, SPARC binds to and diminishes the mitotic activity ofvascular endothelial growth factor. To determine the effect(s) of SPARC on angiogenic responses in vivo, weimplanted polyvinyl alcohol sponges subcutaneously into wild-type and SPARC-null mice. On days 12 and 20 followingimplantation, SPARC-null mice showed increased cellular invasion of the sponges in comparison to wild-type mice.Areas of the sponge with the highest cell density exhibited the highest numbers of vascular profiles in both wild-typeand SPARC-null animals. The endothelial component of the vessels was substantiated by immunoreactivity with threedifferent markers specific for endothelial cells. Although sponges from SPARC-null relative to wild-type mice werepopulated by significantly more cells and blood vessels, an increase in the ratio of vascular to nonvascular cells wasnot apparent. No differences in the percentage of proliferating cells within the sponge were detected between wild-type and SPARC-null sections. However, elevated levels of vascular endothelial growth factor were associated withsponges from SPARC-null versus wild-type mice. An increase in vascular endothelial growth factor production was alsoobserved in SPARC-null primary dermal fibroblasts relative to those of wild-type cells. In conclusion, we have shownthat the fibrovascular invasion of polyvinyl alcohol sponges is enhanced in mice lacking SPARC, and we propose thatincreased levels of vascular endothelial growth factor account, at least in part, for this response. (WOUND REP REG2001;9:522–530)
Secreted protein acidic and rich in cysteine (SPARC) is ELISA Enzyme-linked immunosorbent assaya member of the matricellular class of proteins, opera- FCS Fetal calf serumtionally defined as modular, matrix-associated macro- MECA Mouse endothelial cell antigen
PBS Phosphate buffered saline solutionmolecules that are thought not to contribute structurallyPCNA Proliferating cell nuclear antigento the extracellular matrix but instead to act as modula-PECAM Platelet endothelial cell adhesion moleculetors of cell-matrix interactions.1 A variety of activitiesPVA Polyvinyl alcohol
have been defined for SPARC that include counter- SPARC Secreted protein acidic and rich in cysteineVEGF Vascular endothelial growth factorVEGFR VEGF receptor
From the Department of Vascular Biologya, The HopeHeart Institute, and Department of Medicineb,University of Washington, Seattle, Washington;and Advanced Tissue Sciencesc, La Jolla, Califor-nia. adhesion and inhibition of cell cycle progression in vitro.2
Reprint requests: E. Helene Sage, PhD, The Hope Heart SPARC has been shown to affect the expression of pro-Institute, 1124 Columbia St., Ste. 723, Seattle, WA teins involved in extracellular matrix remodeling, such98104. Fax: (206) 903-2044; Email:hsage@hope
as plasminogen activator inhibitor-1, matrix metallopro-heart.orgtease-3, and fibronectin.3,4 In addition, SPARC modulatesCopyright � 2001 by The Wound Healing Society.
ISSN: 1067-1927 $15.00 � 0 the activity of vascular endothelial growth factor (VEGF)
522

WOUND REPAIR AND REGENERATIONVOL. 9, NO. 6 BRADSHAW, ET AL. 523
by binding to VEGF and subsequently preventing VEGF- with the characterization of SPARC as a modulator ofcell-matrix interaction during events that involve extra-cell surface receptor interaction.2 Expression of SPARC
is frequently associated with events that involve matrix cellular matrix remodeling.turnover such as wound healing and angiogenesis.2,5 Al-though the external appearance of SPARC-null mice is MATERIALS AND METHODSessentially indistinguishable from that of wild-type mice,
C57Bl/6 X 129SVJ mice with a targeted deletion of thea variety of tissues are affected by the absence of this
SPARC gene as described by Norose et al.6 were housedprotein. SPARC-null mice develop cataracts at an early
under modified pathogen-free conditions. All studiesage, show a progressively severe osteopenia, and display
were carried out in compliance with Institutional guide-accelerated closure of excisional dermal wounds.6–9 In
lines. Prior to implantation, circular PVA sponges of uni-addition, SPARC-null mice exhibit increased deposition
form size (10 mm diameter, clinical PVA sponges grade 3,of adipose tissue in the skin and epididymal fat pads in
M-PACT Worldwide Management Co., Eudora, KS) werethe absence of significant differences in overall body
rinsed in 70% ethanol and were boiled in sterile phos-weight.10 The mechanism responsible for several of these
phate buffered saline solution (PBS) for 15 minutes.phenotypes appears to be aberrant deposition of extra-
Sponges were hydrated overnight in sterile PBS and werecellular matrix, although further experiments for a
subsequently implanted subcutaneously in the dorsa ofproper understanding of these defects are required.
anesthesized mice, as described in Reed et al.19 TwoThere are several possibilities by which SPARC could
sponges per animal were implanted into five animals ofregulate angiogenesis. For example, native SPARC inhib-
each genotype and time point for the experiment shownits proliferation and induces cell shape changes in cul-
in Table 1. One animal from each genotype exhibitedtured endothelial cells,2 and/or SPARC contains the
significant inflammation at the 20-day time point; thesepeptide KGHK which is pro-angiogenic in vivo and in
mice were not included in the analysis. One sponge fromvitro.11,12 Release of this peptide by proteolysis of SPARC
each animal was used for quantification following assess-could contribute to the induction of neovascularization
ment of equivalent invasion in both sponges from theof tissues.13 Angiogenesis is a complex process that re-
same animal. At the appropriate end point, animals werequires cell proliferation, migration, differentiation, and
euthanized and the sponges were removed for histologicextracellular matrix turnover.14,15 The different activities
processing. Sponges were immersed for 2 hours in 10%associated with SPARC and its proteolytic fragments are
neutral buffered formalin at 4�C, embedded in paraffin,thought to facilitate different steps in the formation of
and sectioned. Sponges were stained with hemotoxylinnew vessels.2
and eosin (H&E) or Masson’s trichrome reagents for anal-Implantation of polyvinyl alcohol (PVA) sponges pro-
ysis of cellular invasion.vides a permissive testbed in vivo for the analysis of
For quantification of invasion, magnified imagescellular responses such as angiogenesis.16 Subcutaneous
were viewed on a Leica microscope and were capturedplacement elicits a robust invasion of cells from the der-
by a Spot digital camera. Images were imported intomal layers into the sponges and gives rise to a highly
the National Institutes of Health (NIH) Image softwarevascularized tissue. PVA sponges have been used to ana-
program. Cell invasion within stained sponges waslyze a variety of pro- and anti-angiogenic compounds in
clearly visible on imported images. A given area wasa number of different species.17,18 In this report, PVA
outlined, and the designated area was subsequently cal-sponges were implanted into mice with a targeted dele-
culated by the NIH Image program. The sum of all areastion of the SPARC gene to ascertain whether the lack of
within the sponge occupied by cells was calculated bySPARC affected fibrovascular invasion and the subse-
use of the Excel software program (Microsoft, Seattle,quent neovascularization of the implanted material. We
WA). The area of cellular occupancy was determined asobserved an overall increase in the amount of cellular
a percentage of the total area available for invasion, i.e.,invasion in sponges of SPARC-null mice in comparison
the total area minus the area covered by sponge material.with those of wild-type animals. Although differences in
In general, three fields were required to cover the totalcell proliferation were not observed, a significant in-
area of the explanted sponge, except for two cases increase in the levels of VEGF was observed in both
which only two fields were required.sponges and in primary cells cultured from SPARC-nullmice relative to those from wild-type mice. The differ- Immunohistochemistry
For the immunolocalization of specific proteins, forma-ences observed between the invasion of sponge implantsfrom wild-type versus SPARC-null mice are consistent lin-fixed, paraffin-embedded tissue sections were rehy-

WOUND REPAIR AND REGENERATIONNOVEMBER–DECEMBER 2001524 BRADSHAW, ET AL.
drated through an ethanol gradient ending with water.Sections were incubated in 3% hydrogen peroxide for 15minutes to block endogenous peroxidases, followed by10% normal goat serum in PBS-Tween for 20 minutes toblock nonspecific protein interactions. Incubations withspecified primary antibodies were carried out for 1 hourat room temperature. The following antibodies wereused: anti-platelet endothelial cell adhesion molecule(PECAM), a rat monoclonal IgG (Pharmingen, San Diego,CA), anti-mouse endothelial cell antigen (MECA-) 32, arat monoclonal IgG,20 anti-VEGF, a rabbit polyclonal IgG(Santa Cruz Biotechnology, Santa Cruz, CA), anti-VEGFreceptor 2 (TO14), an affinity-purified rabbit polyclonalantibody,21 and antiproliferating cell nuclear antigen
Figure 1. Sponges implanted into SPARC-null mice (–/–) show(PCNA), a mouse monoclonal IgG conjugated to biotingreater vascular content than those from wild-type mice (�/(Zymed Laboratories, San Francisco, CA). When neces-�). Sponges from 8-month-old wild-type (A and C) and SPARC-sary, appropriate secondary antibodies conjugated to bi-null mice (B andD) were removed, fixed in formalin, and embed-
otin (Jackson Immuno Research, West Grove, PA) were ded in paraffin. Shown here are entire sponges embedded inincubated for 1 hour at room temperature, followed by paraffin blocks. Although no substantial hemorrhage occurred
during the removal of the sponges from the animals, SPARC-nulldetection with 3,3-diaminobenzidine (Sigma Chemicalsponges exhibited a more intense hematological componentCo., St. Louis MO). In some cases, an intermediate ampli-in comparison with wild-type sponges. Sponges are 10 mm infication with strepavidin/biotin reagent was performeddiameter.
(DAKO, Carpinteria, CA). Sections were counterstainedwith either methyl green, toluidine blue, or eosin.
removal of the sponges from any of the animals, thePrimary cell isolation and ELISA sponges from SPARC-null mice appeared to contain moreIsolation of primary dermal fibroblasts was carried out as blood than those from wild-type counterparts. This vas-described by Bradshaw et al.22 The VEGF mouse-specific cular component might reflect an increase in the levelenzyme-linked immunosorbent assay (ELISA; R&D Sys- of vascularity and/or an increase in the permeability oftems, Minneapolis, MN) was carried out as specified by the vessels.the manufacturer’s instructions. No detectable VEGF ac- To determine the extent of fibrovascular invasion,tivity was present in the media control, Dulbecco’s mini- the sponges were sectioned and stained with H&E ormal essential media supplemented with 2% fetal calf Masson’s trichrome reagents. Shown in Figure 2 are rep-serum (FCS, Gibco-BRL, Rockville, MD). Similar results resentative sponges taken from wild-type (A and C) andwere found with another ELISA kit purchased from SPARC-null animals (B and D) stained with Masson’sChemicon (Temecula, CA) with primary cells grown in trichrome preparation. A and B are lower-magnification10% FCS, although higher background levels were noted images of the central region of each sponge implantedunder these conditions. for 20 days and illustrate the increased cellular invasion
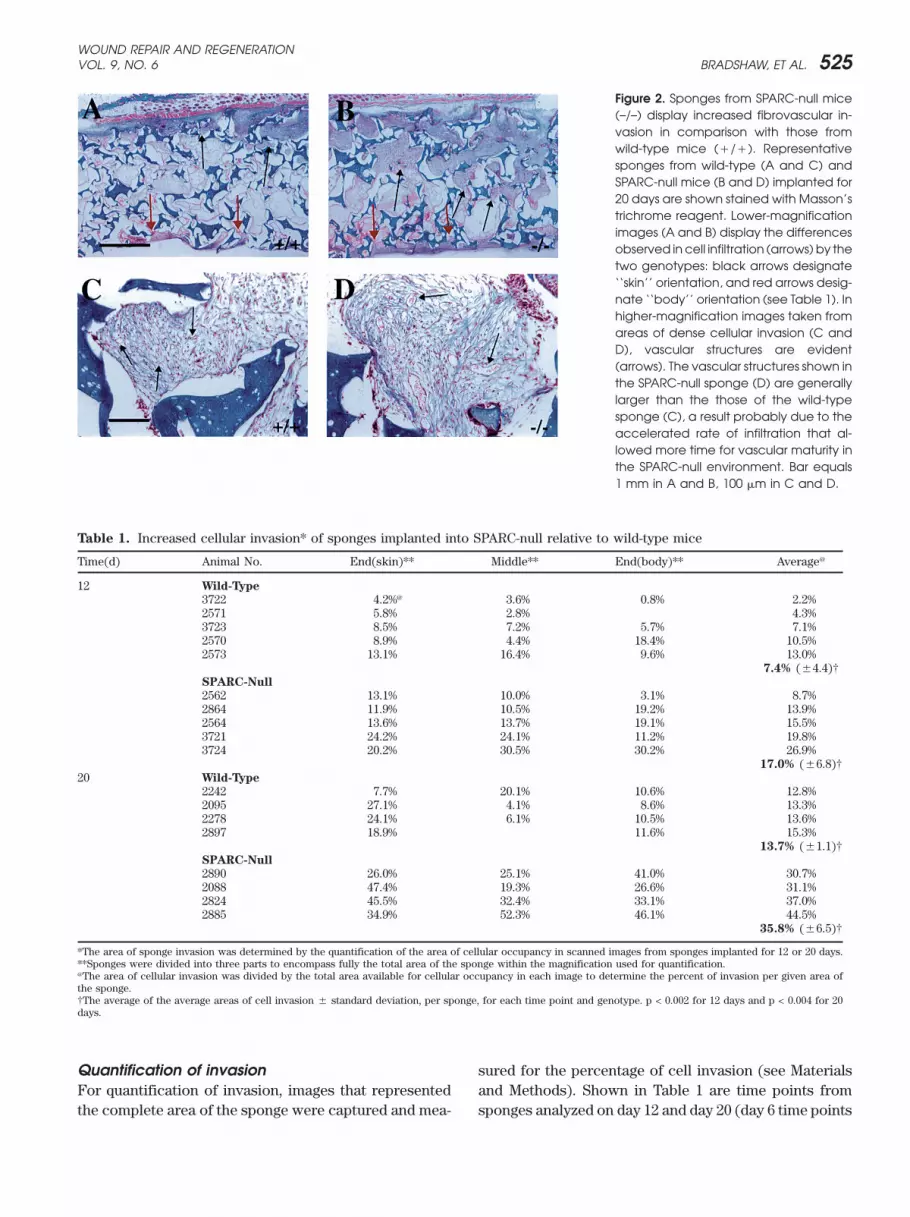
observed in sponges from SPARC-null versus wild-typemice. The arrows in A and B indicate the front of the
RESULTS invading cells (‘‘skin’’ side is designated by black arrows,and ‘‘body’’ side, by red arrows, Table 1). In some cases,Given the elevated expression of SPARC in tissues under-
going vascular remodeling, we posited that the absence there was differential invasion from the skin side of thesponge although not in every case (Table 1). Spongesof SPARC would affect the fibrovascular invasion of im-
planted PVA sponges. To this end, PVA sponges of uni- isolated from SPARC-null mice generally contained ves-sels of greater diameter (D), in comparison with thoseform size (10 mm diameter/4 mm thick) were placed
subcutaneously in age-matched wild-type and SPARC- of wild-type animals (C). The increased invasion notedin the absence of SPARC might allow for the formationnull mice. The sponges were removed at 6, 12, and 20
days following implantation. Shown in Figure 1 are low- of more mature vessels; alternatively, the apparently thin-ner walls of some of these larger vessels might contributemagnification images of sponges removed after 20 days
in wild-type (A and C) or SPARC-null animals (B and D). to the increased vascular permeability associated withsponges recovered from SPARC-null mice (Figure 1).Although no obvious hemorrhage occurred during the
WOUND REPAIR AND REGENERATIONVOL. 9, NO. 6 BRADSHAW, ET AL. 525
Figure 2. Sponges from SPARC-null mice(–/–) display increased fibrovascular in-vasion in comparison with those fromwild-type mice (�/�). Representativesponges from wild-type (A and C) andSPARC-null mice (B and D) implanted for20 days are shown stained with Masson’strichrome reagent. Lower-magnificationimages (A and B) display the differencesobserved in cell infiltration (arrows) by thetwo genotypes: black arrows designate‘‘skin’’ orientation, and red arrows desig-nate ‘‘body’’ orientation (see Table 1). Inhigher-magnification images taken fromareas of dense cellular invasion (C andD), vascular structures are evident(arrows). The vascular structures shown inthe SPARC-null sponge (D) are generallylarger than the those of the wild-typesponge (C), a result probably due to theaccelerated rate of infiltration that al-lowed more time for vascular maturity inthe SPARC-null environment. Bar equals1 mm in A and B, 100 �m in C and D.
Table 1. Increased cellular invasion* of sponges implanted into SPARC-null relative to wild-type mice
Time(d) Animal No. End(skin)** Middle** End(body)** Average@
12 Wild-Type
3722 4.2%@ 3.6% 0.8% 2.2%2571 5.8% 2.8% 4.3%3723 8.5% 7.2% 5.7% 7.1%2570 8.9% 4.4% 18.4% 10.5%2573 13.1% 16.4% 9.6% 13.0%
7.4% (�4.4)†SPARC-Null
2562 13.1% 10.0% 3.1% 8.7%2864 11.9% 10.5% 19.2% 13.9%2564 13.6% 13.7% 19.1% 15.5%3721 24.2% 24.1% 11.2% 19.8%3724 20.2% 30.5% 30.2% 26.9%
17.0% (�6.8)†20 Wild-Type
2242 7.7% 20.1% 10.6% 12.8%2095 27.1% 4.1% 8.6% 13.3%2278 24.1% 6.1% 10.5% 13.6%2897 18.9% 11.6% 15.3%
13.7% (�1.1)†SPARC-Null
2890 26.0% 25.1% 41.0% 30.7%2088 47.4% 19.3% 26.6% 31.1%2824 45.5% 32.4% 33.1% 37.0%2885 34.9% 52.3% 46.1% 44.5%
35.8% (�6.5)†
*The area of sponge invasion was determined by the quantification of the area of cellular occupancy in scanned images from sponges implanted for 12 or 20 days.**Sponges were divided into three parts to encompass fully the total area of the sponge within the magnification used for quantification.@The area of cellular invasion was divided by the total area available for cellular occupancy in each image to determine the percent of invasion per given area ofthe sponge.†The average of the average areas of cell invasion � standard deviation, per sponge, for each time point and genotype. p < 0.002 for 12 days and p < 0.004 for 20days.
Quantification of invasion sured for the percentage of cell invasion (see Materialsand Methods). Shown in Table 1 are time points fromFor quantification of invasion, images that represented
the complete area of the sponge were captured and mea- sponges analyzed on day 12 and day 20 (day 6 time points

WOUND REPAIR AND REGENERATIONNOVEMBER–DECEMBER 2001526 BRADSHAW, ET AL.
did not exhibit significant invasion in either wild-type orSPARC-null mice). On day 12, sponges recovered fromSPARC-null mice displayed a greater amount of cellularinvasion than seen in sponges from wild-type counter-parts (17.0% vs. 7.4%). The trend toward increased inva-sion persisted, because on day 20, when wild-typesponges displayed an average of 13.7% cellular occu-pancy, sponges from SPARC-null animals exhibited anaverage value of 35.8%. Thus, SPARC-null mice showeda consistent, sustained increase in the amount of cellularinvasion of PVA sponges in comparison with wild-typemice.
Characterization of vascular structuresThe cells that populate the sponges appeared to be pri-marily fibroblastic and endothelial in nature. As shownin higher-magnification images (Figure 2, C and D), areasof dense invasion exhibited frequent vascular structures(arrows). To confirm the endothelial nature of the vascu-lar structures, we subjected sections of sponges fromwild-type and SPARC-null mice to immunostaining withantibodies against VEGF receptor 2 (VEGFR2, flk-1), acell-surface marker of endothelial cells (Figure 3). Wild-type (A) and SPARC-null sections (B) displayed immuno-reactivity for VEGFR2 in vascular structures, identifiedmorphogically in Masson’s trichrome-stained sections
Figure 3. Vascular structures in the sponges are immunoreactive(arrows). Sections were also stained with anti-PECAM with antibodies against endothelial markers. VEGFR2 immunore-and anti-MECA-32 IgG20 with similar results. In general, activity was performed to confirm the endothelial nature of theareas of the sponge that displayed augmented cell inva- vasculature within the sponges. Wild-type (�/�, A) and SPARC-
null sponges (–/–, B) are shown; immunoreactive cells are indi-sion also exhibited a more dense vascular component.catedby thearrows. Similar resultswereobtainedwithantibodiesCell invasion thus coincided with the formation of vascu-against two other endothelial markers, PECAM and MECA-32lar structures. That SPARC-null cells were able to popu-(see text). Bar equals 100 �m.
late the sponge in greater numbers than the wild-typecells was paralleled by an increase in vascularity. How-ever, the percentage of vascular structures per area of centage of proliferating cells were not found at eitherdense cell invasion was not substantially different be- day 12 or day 20 between sponges from wild-type andtween sponges recovered from wild-type and SPARC- SPARC-null mice. Hence, the increase in cell invasionnull animals. observed in the absence of SPARC was not attributed to
an increase in the number of proliferating cells.Cell proliferation within the spongesSPARC is an inhibitor of cell proliferation when it is VEGF immunoreactivity and production by dermal
fibroblastsadded to cells in culture. Therefore, the absence ofSPARC might lead to increased cell proliferation in vivo. SPARC binds to and diminishes the capacity of VEGF to
bind to VEGFR1 in vitro.24 VEGF acts as a mitogen, aTo address this question, we stained sections of spongeswith anti-PCNA antibodies.23 Shown in Figure 4 are repre- chemoattractant, and a permeability factor on endothe-
lial cells. The primary observation that sponges recov-sentative sections from wild-type (A) and SPARC-nullsponges (B), in which immunoreactive cells are identi- ered from SPARC-null animals displayed an elevated
vascular component was supported by the finding thatfied by arrows. The percentage of proliferating cells wasdetermined by quantification of the number of PCNA- cellular invasion was increased with the formation of
vascular structures. An increase in the permeability ofpositive cells divided by the total cell number in separatefields from at least three animals per time point (shown these structures might also contribute to the appearance
of more vascularity in the sponges from SPARC-nullin Table 2 for day 12). Significant differences in the per-

WOUND REPAIR AND REGENERATIONVOL. 9, NO. 6 BRADSHAW, ET AL. 527
Figure 4. The percentage of proliferatingcells is not increased in sponges recov-ered from SPARC-null versus wild-typemice. Immunoreactivity for PCNA wasused as a marker of mitotically activecells (arrows) in sponges from wild-type(�/�, A) and SPARC-null mice (–/–, B).Notably, the majority of cells at the lead-ing front of invasion in both spongesweremitotically active. Bar equals 30 �m.
Table 2. Percentage of proliferating cells at day 12 in sponges tions. An increase in VEGF production by SPARC-nullfrom SPARC-null versus wild-type mice
fibroblasts in comparison with wild-type fibroblasts wasproliferating cells (%)* observed in four separate preparations of primary cells
Wild-type 19.7% (�/� 3.0)** used prior to passage five. Hence, the absence of SPARCSPARC-null 15.6% (�/� 2.0) was coincident with an increase in VEGF production in*At least 4 fields from no less than 3 sponges per time point stained with anti- vitro and in vivo. Elevated levels of VEGF might account,PCNA antibodies were quantified to determine the percentage of proliferatingcells in implants from wild-type and SPARC-null mice (410-950 total cells per in part, for the increase in fibrovascular invasion, andtime point and genotype were counted). Similar results were observed at day perhaps the increased permeability, observed in sponges20.**Standard error of the mean. p values were not significant. implanted into SPARC-null mice.
DISCUSSIONmice. To address whether the levels of VEGF in thesponges might be affected by the absence of SPARC, we The temporal and spatial expression of SPARC in endo-
thelial cells prompted us to analyze the function of thisstained sponges from both genotypes with anti-VEGFantibodies (Figure 5). The SPARC-null implants (A) dis- protein in neovascular remodeling in vivo. We report
increased fibrovascular invasion of subcutaneous PVAplayed an overall increase in VEGF immunoreactivity(black arrows), in comparison with those from wild-type sponges by cells from SPARC-null mice, in comparison to
the invasion of these implants by cells from age-matched,animals (B). In both genotypes, higher levels of VEGFimmunoreactivity were associated with fibroblastic cells wild-type mice. SPARC inhibits the proliferation of endo-
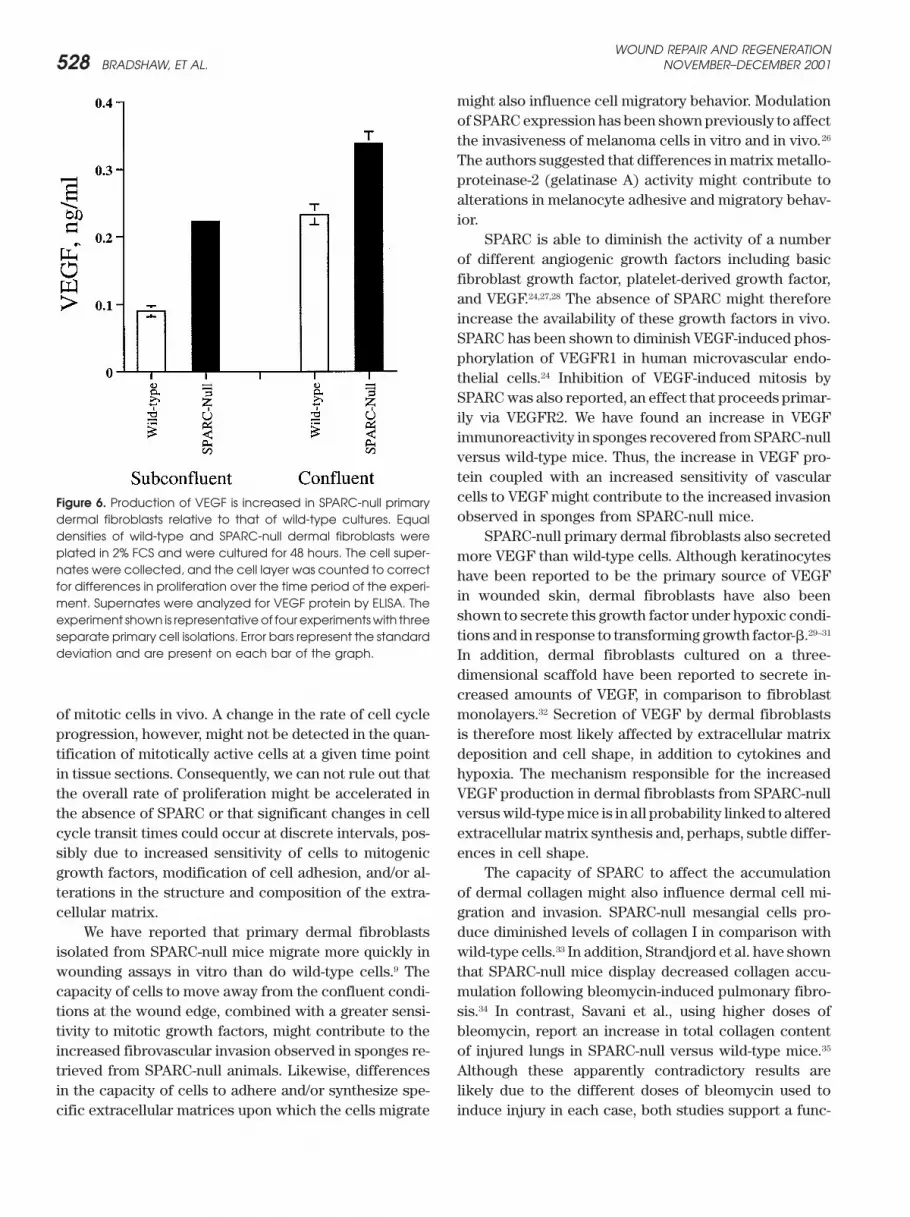
thelial cells and dermal fibroblasts in vitro.22,25 In addition,not present in vascular structures than with vascular-associated cells. primary mesenchymal cells isolated from SPARC-null
mice, including dermal fibroblasts, proliferate faster thanWild-type and SPARC-null primary dermal fibro-blasts were isolated to determine whether apparent dif- wild-type counterparts.22 In this report, anti-PCNA immu-
noreactivity revealed that the percentage of proliferatingferences in VEGF production could be confirmed incultured cells. Figure 6 shows a representative experi- cells within the sponge was not significantly different
between wild-type and SPARC-null animals. Thus, thement in which dermal fibroblasts were grown in lowserum (2% FCS) under subconfluent and confluent condi- absence of SPARC does not appear to affect the profile
Figure 5 Immunoreactivity for VEGF is elevated in sponges recovered from SPARC-null versus wild-type mice. Anti-VEGF antibodieswere used to detect VEGF protein in sponges from wild-type (�/�, A) and SPARC-null mice (–/–, B). Immunoreactivity for VEGF(denoted by black arrows) was detectable in both genotypes; however, SPARC-null cells within the sponge exhibited more reactivitythan wild-type counterparts. In addition, a more enhanced reaction with anti-VEGF IgG was associated with fibroblasts (blackarrows), in comparison with vessel-associated endothelial cells (red arrows). Bar equals 50 �m.
WOUND REPAIR AND REGENERATIONNOVEMBER–DECEMBER 2001528 BRADSHAW, ET AL.
might also influence cell migratory behavior. Modulationof SPARC expression has been shown previously to affectthe invasiveness of melanoma cells in vitro and in vivo.26
The authors suggested that differences in matrix metallo-proteinase-2 (gelatinase A) activity might contribute toalterations in melanocyte adhesive and migratory behav-ior.
SPARC is able to diminish the activity of a numberof different angiogenic growth factors including basicfibroblast growth factor, platelet-derived growth factor,and VEGF.24,27,28 The absence of SPARC might thereforeincrease the availability of these growth factors in vivo.SPARC has been shown to diminish VEGF-induced phos-phorylation of VEGFR1 in human microvascular endo-thelial cells.24 Inhibition of VEGF-induced mitosis bySPARC was also reported, an effect that proceeds primar-ily via VEGFR2. We have found an increase in VEGFimmunoreactivity in sponges recovered from SPARC-nullversus wild-type mice. Thus, the increase in VEGF pro-tein coupled with an increased sensitivity of vascularcells to VEGF might contribute to the increased invasionFigure 6. Production of VEGF is increased in SPARC-null primaryobserved in sponges from SPARC-null mice.dermal fibroblasts relative to that of wild-type cultures. Equal
densities of wild-type and SPARC-null dermal fibroblasts were SPARC-null primary dermal fibroblasts also secretedplated in 2% FCS and were cultured for 48 hours. The cell super- more VEGF than wild-type cells. Although keratinocytesnates were collected, and the cell layer was counted to correct have been reported to be the primary source of VEGFfor differences in proliferation over the time period of the experi-
in wounded skin, dermal fibroblasts have also beenment. Supernates were analyzed for VEGF protein by ELISA. Theshown to secrete this growth factor under hypoxic condi-experiment shown is representativeof four experimentswith threetions and in response to transforming growth factor-�.29–31separate primary cell isolations. Error bars represent the standard
deviation and are present on each bar of the graph. In addition, dermal fibroblasts cultured on a three-dimensional scaffold have been reported to secrete in-creased amounts of VEGF, in comparison to fibroblastmonolayers.32 Secretion of VEGF by dermal fibroblastsof mitotic cells in vivo. A change in the rate of cell cycle
progression, however, might not be detected in the quan- is therefore most likely affected by extracellular matrixdeposition and cell shape, in addition to cytokines andtification of mitotically active cells at a given time point
in tissue sections. Consequently, we can not rule out that hypoxia. The mechanism responsible for the increasedVEGF production in dermal fibroblasts from SPARC-nullthe overall rate of proliferation might be accelerated in
the absence of SPARC or that significant changes in cell versus wild-type mice is in all probability linked to alteredextracellular matrix synthesis and, perhaps, subtle differ-cycle transit times could occur at discrete intervals, pos-
sibly due to increased sensitivity of cells to mitogenic ences in cell shape.The capacity of SPARC to affect the accumulationgrowth factors, modification of cell adhesion, and/or al-
terations in the structure and composition of the extra- of dermal collagen might also influence dermal cell mi-gration and invasion. SPARC-null mesangial cells pro-cellular matrix.
We have reported that primary dermal fibroblasts duce diminished levels of collagen I in comparison withwild-type cells.33 In addition, Strandjord et al. have shownisolated from SPARC-null mice migrate more quickly in
wounding assays in vitro than do wild-type cells.9 The that SPARC-null mice display decreased collagen accu-mulation following bleomycin-induced pulmonary fibro-capacity of cells to move away from the confluent condi-
tions at the wound edge, combined with a greater sensi- sis.34 In contrast, Savani et al., using higher doses ofbleomycin, report an increase in total collagen contenttivity to mitotic growth factors, might contribute to the
increased fibrovascular invasion observed in sponges re- of injured lungs in SPARC-null versus wild-type mice.35
Although these apparently contradictory results aretrieved from SPARC-null animals. Likewise, differencesin the capacity of cells to adhere and/or synthesize spe- likely due to the different doses of bleomycin used to
induce injury in each case, both studies support a func-cific extracellular matrices upon which the cells migrate

WOUND REPAIR AND REGENERATIONVOL. 9, NO. 6 BRADSHAW, ET AL. 529
by SPARC during angiogenesis in vitro. Changes in fibronectin,tion of SPARC in the regulation of collagen productionthrombospondin-1, and plasminogen activator inhibitor-1. J Biol
or accumulation. The absence of SPARC in mice is char- Chem 1992;267:16736–45.acterized by an altered extracellular matrix, manifested 5. Reed MJ, Puolakkainen P, Lane TF, Dickerson D, Bornstein P,
Sage EH. Differential expression of SPARC and thrombospondinespecially in the quality and quantity of collagen fibrils1 in wound repair: immunolocalization and in situ hybridization.upon which the cells organize and/or mobilize. We haveJ Histochem Cytochem 1993;41:1467–77.
reported that the skin of SPARC-null mice has a reduced 6. Norose K, Clark JI, Syed NA, Basu A, Heber-Katz ES, Sage EH,Howe CC. SPARC deficiency leads to early onset cataractogenesis.collagen content, as determined by hydroxyproline anal-Invest Opthalmol Vis Sci 1998;39:2674–80.ysis, in comparison with wild-type skin.9 As visualized
7. Gilmore DT, Lyon GJ, Carlton MBL, Sanes JR, Cunningham JM,by electron microscopy, the SPARC-null dermal collagen Anderson JR, Hogan BLM, Evans MJ, Colledge WH. Mice deficientfibrils appear smaller and more uniform in diameter than for the secreted glycoprotein SPARC/osteonectin/BM 40 develop
normally but show severe age-onset cataract formation and dis-those present in wild-type dermis (A.D. Bradshaw, J.ruption of the lens. EMBO J 1998;17:1860–70.Carbon, J.M. Davidson, T.N. Wight, EH Sage, in prepara-
8. Delany AM, Amling M, Priemel M, Howe C, Baron R, Canalis E.tion). The decreased collagen content of the skin with Osteopenia and decreased bone formation in osteonectin-defi-
cient mice. J Clin Invest 2000;105:915–23.the decrease in collagen fibril diameter might result in9. Bradshaw AD, Reed MJ, Sage EH. SPARC-null mice exhibit accel-a matrix more easily degraded by extracellular matrix
erated cutaneous wound closure in the dermis, and increasedproteinases and thus facilitate cell movement to areas cellular invasion in a sponge model of angiogenesis. J. Histochemof invasion.10 Cytochem 2002; 50:1–10.
10. Bradshaw AD, Sage EH. SPARC, a matricellular protein that func-SPARC has been proposed to act as a modulator oftions in cellular differentiation and tissue response to injury. Jcell-matrix interactions. Our finding that fibrovascularClin Invest 2001;107:1049–54.
invasion of implanted PVA sponges is increased in 11. Lane TF, Iruela-Arispe ML, Johnson RS, Sage EH. SPARC is asource of copper-binding peptides that stimulate angiogenesis. JSPARC-null mice relative to the invasion seen in wild-Cell Biol 1994;125:929–43.type animals provides further evidence of the capacity
12. Iruela-Arispe ML, Lane TF, Redmond D, Reilly M, Bolender RP,of SPARC to affect cell behavior. SPARC, similar to other Kavanagh TJ, Sage EH. Expression of SPARC during developmentmatricellular proteins, appears to be particularly influen- of the chicken chorioallantoic membrane: evidence for regulated
proteolysis in vivo. Mol Biol Cell 1995;6:327–43.tial in the modulation of events that require matrix turn-13. Sage EH. Pieces of eight: bioactive fragments of extracellularover and cellular repair. Elucidation of the fundamental
proteins as regulators of angiogenesis. Trends Cell Biol 1997;7:mechanisms through which SPARC transduces its effects 182–6.
14. Vernon RB, Sage EH. Between molecules and morphology. Extra-will contribute to our understanding of a variety of pa-cellular matrix and the creation of vascular form. Am J Patholthologies and to the design of agents that facilitate their1995;147:873–83.
resolution. 15. Klagsbrun M, Moses MA. Molecular angiogenesis. Chem Biol 1999;6:R217–R224.
16. Andrade SP, Machado RDP, Teixeira AS, Belo AV, Tarso AM, Be-raldo WT. Sponge-induced angiogenesis in mice and the pharmaco-ACKNOWLEDGMENTS logical reactivity of the neovasculature quantitated by afluorimetric method. Microvasc Res 1997;54:253–61.We thank Dr. Pauli Puolakkainen and members of the
17. Andrade SP, Vieira LB, Bakhle YS, Piper PJ. Effects of plateletSage laboratory for helpful discussions. Supported byactivating factor (PAF) and other vascoconstrictors on a model
National Institutes of Health grants GM 40771 and HL of angiogenesis in the mouse. Int J Exp Pathol 1992;73:503–13.18. Majima M, Hayashi I, Muramatsu M, Katada J, Yamashina S, Katori59574 (E.H.S.), AG 15837 (M.J.R.), HL 10352 (R.A.B.),
M. Cyclo-oxygenase enhances basic fibroblast growth factor-in-and DK 07467 (A.D.B.); National Science Foundation 04-duced angiogenesis through induction of vascular endothelial
150 (E.H.S.), and a Beeson Scholar Award (M.J.R.). growth factor in rat sponge models. Br J Pharmacol 2000;130:641–9.
19. Reed MJ, Corsa A, Pendergrass W, Penn P, Sage EH, Abrass IB.Neovascularization in aged mice – Delayed angiogenesis is coinci-REFERENCES dent with decreased levels of transforming growth factor �1 andtype I collagen. Am J Pathol 1998;152:113–23.1. Bornstein P. Diversity of function is inherent in matricellular pro-
20. Hallmann R, Mayer DN, Berg EL, Broermann R, Butcher EC.teins: an appraisal of thrombospondin 1. J Cell Biol 1995;130:Novel mouse endothelial cell surface marker is suppressed during503–6.differentiation of the blood brain barrier. Dev Dyn 1995;202:2. Brekken RA, Sage EH. SPARC, a matricellular protein: at the325–32.crossroads of cell-matrix communication. Matrix Biol 2000;19:
21. Feng D, Nagy JA, Brekken RA, Pettersson A, Manseau EJ, Pyne569–80.K, Mulligan R, Thorpe PE, Dvorak HF, Dvorak AM. Ultrastructural3. Tremble PM, Lane TF, Sage EH, Werb Z. SPARC, a secreted proteinlocalization of the vascular permeability factor/vascular endothe-associated with morphogenesis and tissue remodeling, induceslial growth factor (VPF/VEGF) receptor-2 (flk-1, KDR) in normalexpression of metalloproteinases in fibroblasts through a novelmouse kidney and in the hyperpermeable vessels induced by VPF/extracellular matrix-dependent pathway. J Cell Biol 1993;121:VEGF-expressing tumors and adenoviral vectors. J Histochem1433–44.
4. Lane TF, Iruela-Arispe ML, Sage EH. Regulation of gene expression Cytochem 2000;48:545–55.

WOUND REPAIR AND REGENERATIONNOVEMBER–DECEMBER 2001530 BRADSHAW, ET AL.
22. Bradshaw AD, Francki A, Motamed K, Howe C, Sage EH. Primary 29. Kishimoto J, Ehama R, Ge Y, Kobayashi T, Nishiyama T, DetmarM, Burgeson RE. In vivo detection of human vascular endothelialmesenchymal cells isolated from SPARC-null mice exhibit altered
morphology and rates of proliferation. Mol Biol Cell 1999;10: growth factor promoter activity in transgenic mouse skin. Am JPathol 2000;157:103–10.1569–79.
23. Wasseem NH, Lane DP. Monoclonal antibody analysis of the prolif- 30. Detmar M, Brown LF, Berse B, Jackman RW, Elicker BM, DvorakHF, Claffey KP. Hypoxia regulates the expression of vascular per-erating cell nuclear antigen (PCNA). Structural conservation and
the detection of a nucleolar form. J Cell Sci 1990;96:121–9. meability factor/vascular endothelial growth factor (VPF/VEGF)and its receptors in human skin. J Invest Dermatol 1997;108:263–8.24. Kupprion C, Motamed K, Sage EH. SPARC (BM-40, osteonectin)
inhibits the mitogenic effect of vascular endothelial growth factor 31. Pertovaara L, Kaipainen A, Mustonen T, Orpana A, Ferrara N,Saksela O, Alitalo K. Vascular endothelial growth factor is inducedon microvascular endothelial cells. J Biol Chem 1998;273:
29635–40. in response to transforming growth factor-� in fibroblastic andepithelial cells. J Biol Chem 1994;269:6271–4.25. Funk SE, Sage EH. The Ca2� -binding glycoprotein SPARC modu-
lates cell cycle progression in bovine aortic endothelial cells. Proc 32. Pinney E, Liu K, Sheeman B, Mansbridge J. Human three-dimen-sional fibroblast cultures express angiogenic activity. J Cell PhysNatl Acad Sci USA 1991;88:2648–52.
26. Ledda MF, Adris S, Bravo AI, Kairiyama C, Bover L, Chernajovsky 2000;183:74–82.33. Francki A, Bradshaw AD, Bassuk JA, Howe CC, Couser WG, SageY, Mordoh J, Podhajcer OL. Suppression of SPARC expression by
antisense RNA abrogates the tumorgenicity of human melanoma EH. SPARC regulates the expression of collagen type I and trans-forming growth factor-�1 in mesangial cells. J Biol Chem 1999;cells. Nat Med 1997;3:171–6.
27. Hasselaar P, Sage EH. SPARC antagonizes the effect of basic 274:32145–52.34. Strandjord TP, Madtes DK, Weiss DJ, Sage EH. Collagen accumula-fibroblast growth factor on the migration of bovine aortic endothe-
lial cells. J Cell Biochem 1992;49:272–83. tion is decreased in SPARC-null mice with bleomycin-inducedpulmonary fibrosis. Am J Physiol 1999;277:L628–35.28. Raines EW, Lane TF, Iruela-Arispe ML, Ross R, Sage EH. The
extracellular glycoprotein SPARC interacts with platelet-derived 35. Savani RC, Zhou Z, Arguiri E, Wang S, Vu D, Howe CC, DeLisserHM. Bleomycin-induced pulmonary injury in mice deficient ingrowth factor (PDGF) -AB and BB and inhibits the binding of
PDGF to its receptors. Proc Natl Acad Sci USA 1992;89:1281–5. SPARC. Am J Physiol Lung Cell Mol Physiol 2000;279:L743–50.