reversible protein phosphorylation regulates jasmonic acid-dependent and -independent wound signal...
TRANSCRIPT

The Plant Journal (1998) 13(2), 153–165
Reversible protein phosphorylation regulates jasmonic acid-dependent and -independent wound signal transductionpathways in Arabidopsis thaliana
Enrique Rojo, Elena Titarenko, Jose Leon, Susanne
Berger1, Guy Vancanneyt and Jose J. Sanchez-Serrano*
Centro Nacional de Biotecnologıa CSIC, Campus de
Cantoblanco UAM, Cta. Colmenar Viejo km. 15,500,
28049 Madrid, Spain, and1Institut fur Pflanzenbiochemie, Weinberg 3, 06120 Halle
(Saale), Germany
Summary
Plants responses to mechanical injury are complex and
include the induced expression of defence-related genes.
The phytohormone JA has been reported to mediate
some of these responses. To elucidate further the signal
transduction processes involved, the action of specific
agonists and antagonists of known signalling effectors
on the response of Arabidopsis thaliana plantlets to JA
and wounding was investigated. The identification and
characterization of a reversible protein phosphorylation
step in a transduction pathway leading to JA-induced
gene transcription is reported. This phosphorylation
event involved the opposing activities of a staurosporine-
sensitive protein kinase, negatively regulating the path-
way, and a protein phosphatase, most probably of type
2 A, which activated JA-responsive gene expression. JA
activation via this pathway was blocked in the A.
thaliana JA-insensitive mutants jin1, jin4 and coi1, and
by exogenous application of cycloheximide or auxins.
Wound-induced activation of JA-responsive genes was
also regulated by this protein phosphorylation step. An
alternative wound signalling pathway, independent of
JA, was also identified, leading to the transcriptional
activation of a different set of genes. This JA-independent
pathway was also regulated by a protein phosphorylation
switch, in which the protein kinase positively regulated
the pathway while the protein phosphatase negatively
regulated it. Moreover, a labile protein apparently
repressed the expression of these genes. One of the
genes analysed, JR3, had a complex pattern of expres-
sion, possibly because it was regulated via both of the
wound signalling pathways identified. According to the
function of an homologous gene, JR3 may be involved
in feedback inhibition of the JA response.
Received 9 May 1997; revised 2 September 1997; accepted 5 September
1997.
*For correspondence (fax 134 1585 4506).
© 1998 Blackwell Science Ltd 153
Introduction
Jasmonic acid (JA) is a cyclopentanone derivative of
linolenic acid that exhibits features of a plant hormone
(Sembdner and Parthier, 1993). A function of JA in plants
may be in regulating gene activation in response to
mechanical damage. Upon wounding, there is a rise in
endogenous levels of JA (Albrecht et al., 1993; Creelman
et al., 1992; Pena-Cortes et al., 1993), due to increased
de novo synthesis, which triggers the activation of
wound-inducible genes. Exogenously applied JA is able
to induce these wound-responsive genes in non-
damaged plants (Farmer et al., 1992; Hildmann et al.,
1992; Reinbothe et al., 1994). In addition, application of
exogenous JA also induces secondary metabolic path-
ways leading to phytoalexin production (Dittrich et al.,
1992). Elicitor-induced synthesis of phytoalexins may also
require de novo synthesis of JA (Gundlach et al., 1992),
thus establishing a link between defence responses to
injury and pathogen attack.
Novel roles for JA in the physiology of non-stressed
plants have been reported. For example, JA may be
involved in tuberization in potato, and is regarded as a
tuber-inducing factor (Koda et al., 1991; Pelacho and
Mingo-Castel, 1991). In addition, it may regulate nitrogen
partitioning in soybean, where it induces the accumulation
of vegetative storage proteins (VSP) in leaves. These
proteins act as transient nitrogen reservoirs in depoded
plants (Anderson et al., 1989; Mason and Mullet, 1990).
The recent characterization of Arabidopsis thaliana mutants
insensitive to or deficient in the production of JA has also
suggested that it is involved in pollen formation (Feys
et al., 1994; McConn and Browse, 1996).
While gene activation upon wounding requires an
increase in the endogenous JA levels due to induced de
novo synthesis, it is not established whether the effects
the hormone exerts on plant development also require
organ- or stage-specific increases of JA levels. Basal JA
levels in the different organs may play roles in plant
physiology, and differences between basal and wound-
induced JA may underlie the distinct effects this hormone
exerts in apparently unrelated processes. In this regard,
co-suppression in transgenic A. thaliana of a lipoxygenase
gene, encoding an enzyme involved in JA synthesis,
indicates that wound-induced and basal JA may be synthe-
sized through different pathways (Bell et al., 1995). More-
over, expression of the flaxseed allene oxide synthase (also
involved in JA synthesis) in transgenic potato plants results
154 Enrique Rojo et al.
in JA levels comparable to those attained upon
wounding. However, these elevated levels do not
activate the JA-responsive, wound-inducible genes (Harms
et al., 1995). Taken together, these data suggest the pres-
ence of separate JA pools that are differently perceived,
and lead to the activation of distinct physiological pro-
cesses, most probably through the induction of separate
sets of genes.
Very little is known about how JA is perceived, and of
the mechanisms which may allow cells to discriminate the
origin of JA and which specifically activate defined sets of
genes in response. Several A. thaliana mutants altered in
sensitivity to JA have been identified based on phenotypic
screenings for normal growth in the presence of JA. Among
them, three non-allelic mutants, jin1, jin4 (Berger et al.,
1996) and coi1 (Feys et al., 1994), were identified by their
ability to root in JA-containing medium in which wild-
type plants exhibited a markedly reduced root growth. In
addition, JA induction of VSP genes in these mutants was
impaired or much weaker than in wild-type plants. The
availability of JA-insensitive mutants in A. thaliana will
certainly yield highly valuable information on the
components involved in JA signalling, and on their roles
in wound-induced gene expression. However, characteriza-
tion of the mutated genes and their functions in the
signalling pathways often proves to be tedious. In addition,
screening procedures may be biased towards preferential
isolation of components of an inducible pathway, if
separate JA pools do exist.
Pharmacological approaches have proved useful in the
characterization of components of signal transduction
pathways in eukaryotes. These experiments have con-
firmed that plant intracellular signalling proceeds via the
same basic components and motifs as their counterparts
in other eukaryotic systems (Bush, 1995; Ma, 1994; Smith
and Walker, 1996). Due to its versatility in modulating the
duration of a response to a stimulus and its ability to
amplify the signals powerfully, reversible protein phos-
phorylation is a common switching mechanism in signal
transduction pathways, including hormone perception in
plants (ethylene, Kieber et al., 1993; Raz and Fluhr, 1993;
gibberellin, Kuo et al., 1996; ABA, Leung et al., 1994; Meyer
et al., 1994; cytokinin, Kakimoto, 1996). The availability of
specific inhibitors of protein kinases and phosphatases
(MacKintosh and MacKintosh, 1994) has allowed the
elucidatation of the roles of their target enzymes in signal
transduction pathways of lower eukaryotes, and in animal
and plant systems.
We have taken advantage of these pharmacological
tools to characterize wound signalling pathways in A.
thaliana. In particular, we have investigated the involve-
ment of reversible protein phosphorylation in modulating
JA signalling.
© Blackwell Science Ltd, The Plant Journal, (1998), 13, 153–165
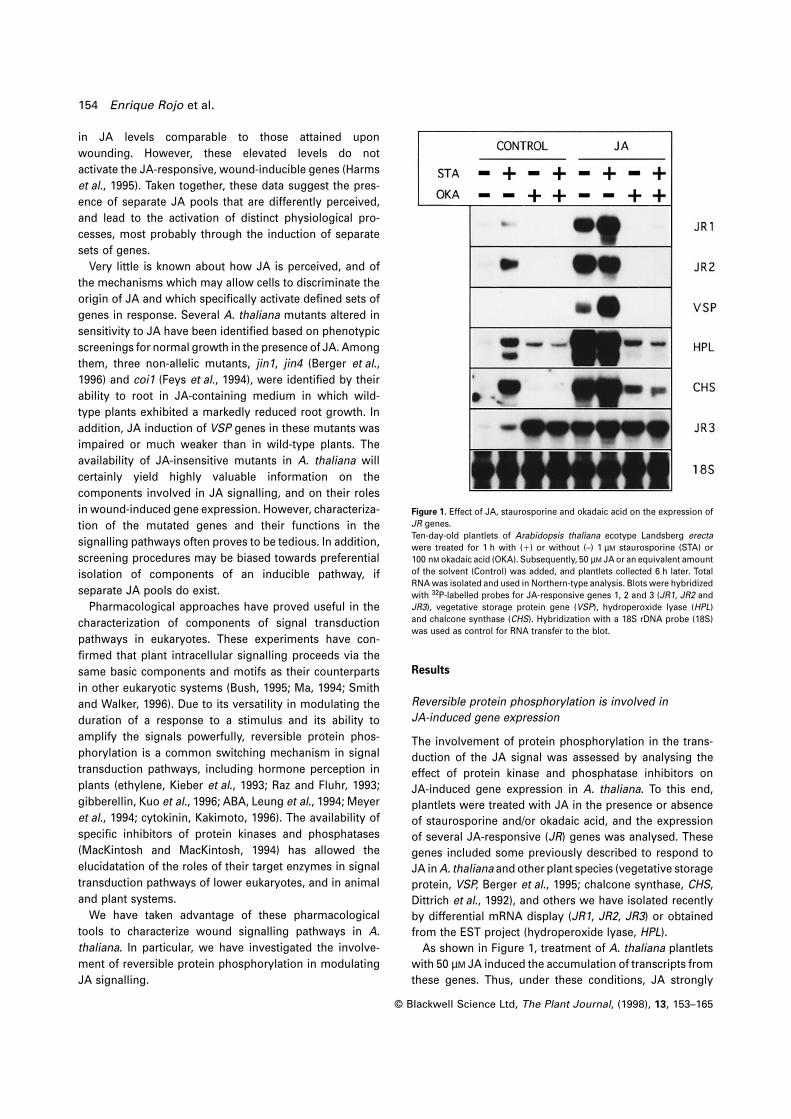
Figure 1. Effect of JA, staurosporine and okadaic acid on the expression of
JR genes.
Ten-day-old plantlets of Arabidopsis thaliana ecotype Landsberg erecta
were treated for 1 h with (1) or without (–) 1 µM staurosporine (STA) or
100 nM okadaic acid (OKA). Subsequently, 50 µM JA or an equivalent amount
of the solvent (Control) was added, and plantlets collected 6 h later. Total
RNA was isolated and used in Northern-type analysis. Blots were hybridized
with 32P-labelled probes for JA-responsive genes 1, 2 and 3 (JR1, JR2 and
JR3), vegetative storage protein gene (VSP), hydroperoxide lyase (HPL)
and chalcone synthase (CHS). Hybridization with a 18S rDNA probe (18S)
was used as control for RNA transfer to the blot.
Results
Reversible protein phosphorylation is involved in
JA-induced gene expression
The involvement of protein phosphorylation in the trans-
duction of the JA signal was assessed by analysing the
effect of protein kinase and phosphatase inhibitors on
JA-induced gene expression in A. thaliana. To this end,
plantlets were treated with JA in the presence or absence
of staurosporine and/or okadaic acid, and the expression
of several JA-responsive (JR) genes was analysed. These
genes included some previously described to respond to
JA in A. thaliana and other plant species (vegetative storage
protein, VSP, Berger et al., 1995; chalcone synthase, CHS,
Dittrich et al., 1992), and others we have isolated recently
by differential mRNA display (JR1, JR2, JR3) or obtained
from the EST project (hydroperoxide lyase, HPL).
As shown in Figure 1, treatment of A. thaliana plantlets
with 50 µM JA induced the accumulation of transcripts from
these genes. Thus, under these conditions, JA strongly

Protein phosphorylation and wound signalling 155
activated the expression of a large set of genes (Titarenko
et al., 1997). In the case of HPL, the appearence of two
hybridizing mRNA bands of different size was observed
upon JA treatment.
Staurosporine, a broad range serine-threonine protein
kinase inhibitor (MacKintosh and MacKintosh, 1994;
Tamaoki et al., 1986), could mimic the effect of JA by
inducing the same set of genes (Figure 1). However, the
levels of expression attained upon treatment with 1 µM
staurosporine were lower than those observed after treat-
ment with 50 µM JA. Moreover, increasing the stauro-
sporine concentration did not result in higher levels of JR
transcripts (results not shown). Under these conditions
staurosporine treatment had a slight effect on JR1 expres-
sion and none on VSP expression. Interestingly, JR1 and
VSP transcripts accumulated to a larger extent when JA
and staurosporine were applied simultaneously compared
with incubation with JA alone. This overinduction was not
observed for the other genes analysed in the conditions
used in this experiment, but it was clearly detectable at
lower concentrations of JA (see below). These data suggest
that a protein kinase activity negatively regulates the
expression of JR genes.
To elucidate whether a protein phosphatase was involved
in JA perception as well, we examined the effects of protein
phosphatase inhibitors on the induction of JR genes by JA
and/or staurosporine. The polyether compound okadaic
acid (Bialojan and Takai, 1988; Cohen et al., 1990) is a
potent inhibitor of protein serine-threonine phosphatase
types 1 (PP1) and 2A (PP2A) at submicromolar concentra-
tions, whereas type 2B is only inhibited at high micromolar
concentrations and type 2C activity is not affected. Treat-
ment with 100 nM okadaic acid abolished JA-induced
expression of all but one (JR3) of the genes investigated,
indicating that a PP1 or PP2A activity is involved in trans-
duction of the JA signal. In addition, okadaic acid blocked
the induction of these genes caused by staurosporine,
suggesting that JA and staurosporine activate JR genes
through a common pathway that requires the activity of
this okadaic acid-sensitive protein phosphatase.
As seen in Figure 1, the expression of JR3 was induced
by treatment with JA and/or staurosporine, as were the
rest of the JR genes. However, in plantlets treated with JA
and/or staurosporine in the presence of okadaic acid, JR3
was still expressed, in contrast to the other JR genes. Since
JR3 expression was induced upon treatment with okadaic
acid alone, it was not possible to determine whether the
protein phosphatase activity previously identified was also
required for JA-induced expression of JR3.
We tested the possibility that the inhibitors used were
altering the timing of JA-induced gene activation, and not
the level of expression. As shown in Figure 2, increases in
the steady-state mRNA levels of JR genes were detectable
after 90 min of JA treatment and, for most genes, maximal
© Blackwell Science Ltd, The Plant Journal, (1998), 13, 153–165
Figure 2. Time–course of JA-induced mRNA accumulation in the presence
or absence of staurosporine and okadaic acid.
Ten-day-old plantlets of Arabidopsis thaliana ecotype Landsberg erecta
were treated for 1 h with 1 µM staurosporine (STA) or 100 nM okadaic acid
(OKA), or not treated with any of these inhibitors. Subsequently, 50 µM JA
was added, and plantlets were collected at the indicated times (in hours).
Total RNA gel blots were hybridized with the radioactive probes as described
in Figure 1.
accumulation was observed after 6 h. However, treatment
with JA and staurosporine could clearly increase accumula-
tion of JR1, VSP and CHS transcripts over the maximal
levels attained after JA treatment, suggesting that
staurosporine treatment affects the amount of transcript
accumulated but does not alter the timing of maximal
induction. Similarly, the inhibitory effect of okadaic
acid on JA-induced mRNA accumulation was observed by
90 min after treating with JA, and remained for at least
48 h, indicating that okadaic acid acts by blocking, and
not merely delaying, transduction of the JA signal. For
subsequent experiments, we took samples at 6 h after
treatments, when maximal expression of the genes was
observed.
An active protein phosphatase is required for JA-induced
gene expression
To gain further insight into the type of protein phospha-
tase activity involved in JA induction of gene expression,
a dose-dependence experiment was performed with the
inhibitors okadaic acid and calyculin A. These compounds
are structurally unrelated, and display inhibitory activity
towards PP1 and PP2A. While okadaic acid shows a
stronger inhibitory activity towards PP2A (concentration
for 50% inhibition (IC50) 5 0.1–1 nM for PP2A and 10–15 nM
for PP1; Cohen et al., 1990), calyculin A is reported to
be equally effective against both types (IC50 5 0.5–2 nM;
Ishihara et al., 1989).
As shown in Figure 3, as little as 20 nM okadaic acid (a
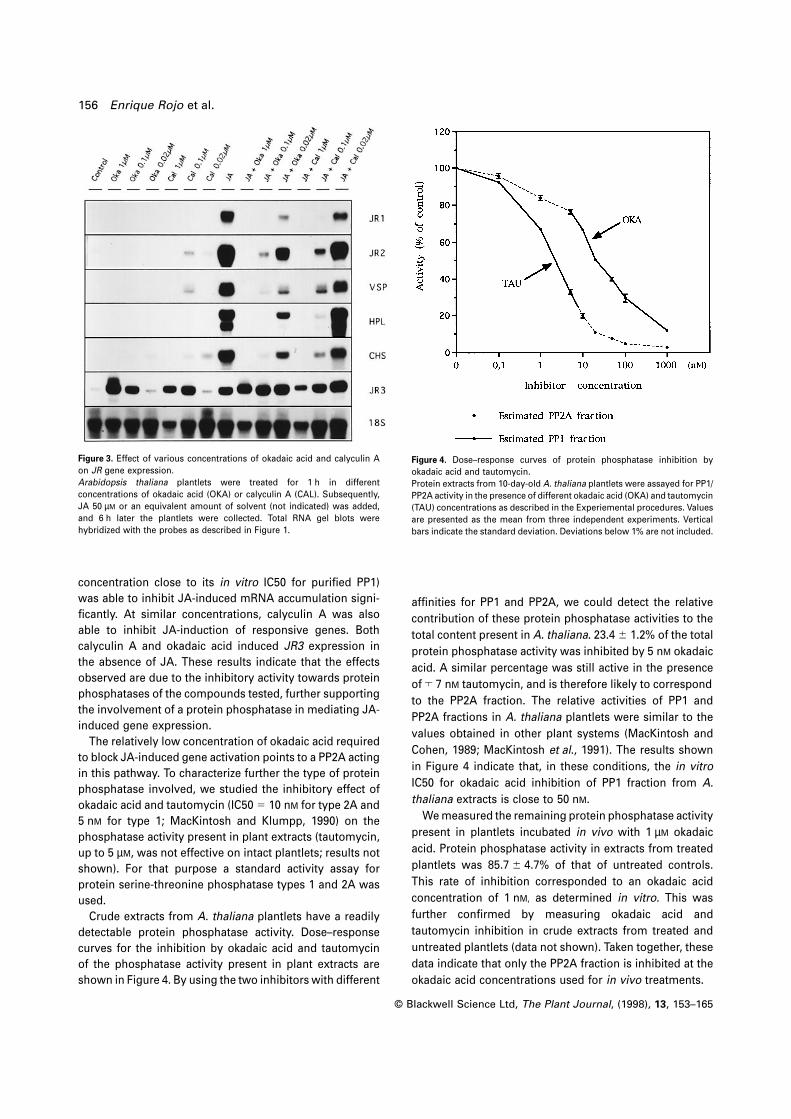
156 Enrique Rojo et al.
Figure 3. Effect of various concentrations of okadaic acid and calyculin A
on JR gene expression.
Arabidopsis thaliana plantlets were treated for 1 h in different
concentrations of okadaic acid (OKA) or calyculin A (CAL). Subsequently,
JA 50 µM or an equivalent amount of solvent (not indicated) was added,
and 6 h later the plantlets were collected. Total RNA gel blots were
hybridized with the probes as described in Figure 1.
concentration close to its in vitro IC50 for purified PP1)
was able to inhibit JA-induced mRNA accumulation signi-
ficantly. At similar concentrations, calyculin A was also
able to inhibit JA-induction of responsive genes. Both
calyculin A and okadaic acid induced JR3 expression in
the absence of JA. These results indicate that the effects
observed are due to the inhibitory activity towards protein
phosphatases of the compounds tested, further supporting
the involvement of a protein phosphatase in mediating JA-
induced gene expression.
The relatively low concentration of okadaic acid required
to block JA-induced gene activation points to a PP2A acting
in this pathway. To characterize further the type of protein
phosphatase involved, we studied the inhibitory effect of
okadaic acid and tautomycin (IC50 5 10 nM for type 2A and
5 nM for type 1; MacKintosh and Klumpp, 1990) on the
phosphatase activity present in plant extracts (tautomycin,
up to 5 µM, was not effective on intact plantlets; results not
shown). For that purpose a standard activity assay for
protein serine-threonine phosphatase types 1 and 2A was
used.
Crude extracts from A. thaliana plantlets have a readily
detectable protein phosphatase activity. Dose–response
curves for the inhibition by okadaic acid and tautomycin
of the phosphatase activity present in plant extracts are
shown in Figure 4. By using the two inhibitors with different
© Blackwell Science Ltd, The Plant Journal, (1998), 13, 153–165
Figure 4. Dose–response curves of protein phosphatase inhibition by
okadaic acid and tautomycin.
Protein extracts from 10-day-old A. thaliana plantlets were assayed for PP1/
PP2A activity in the presence of different okadaic acid (OKA) and tautomycin
(TAU) concentrations as described in the Experiemental procedures. Values
are presented as the mean from three independent experiments. Vertical
bars indicate the standard deviation. Deviations below 1% are not included.
affinities for PP1 and PP2A, we could detect the relative
contribution of these protein phosphatase activities to the
total content present in A. thaliana. 23.4 6 1.2% of the total
protein phosphatase activity was inhibited by 5 nM okadaic
acid. A similar percentage was still active in the presence
of ™ 7 nM tautomycin, and is therefore likely to correspond
to the PP2A fraction. The relative activities of PP1 and
PP2A fractions in A. thaliana plantlets were similar to the
values obtained in other plant systems (MacKintosh and
Cohen, 1989; MacKintosh et al., 1991). The results shown
in Figure 4 indicate that, in these conditions, the in vitro
IC50 for okadaic acid inhibition of PP1 fraction from A.
thaliana extracts is close to 50 nM.
We measured the remaining protein phosphatase activity
present in plantlets incubated in vivo with 1 µM okadaic
acid. Protein phosphatase activity in extracts from treated
plantlets was 85.7 6 4.7% of that of untreated controls.
This rate of inhibition corresponded to an okadaic acid
concentration of 1 nM, as determined in vitro. This was
further confirmed by measuring okadaic acid and
tautomycin inhibition in crude extracts from treated and
untreated plantlets (data not shown). Taken together, these
data indicate that only the PP2A fraction is inhibited at the
okadaic acid concentrations used for in vivo treatments.
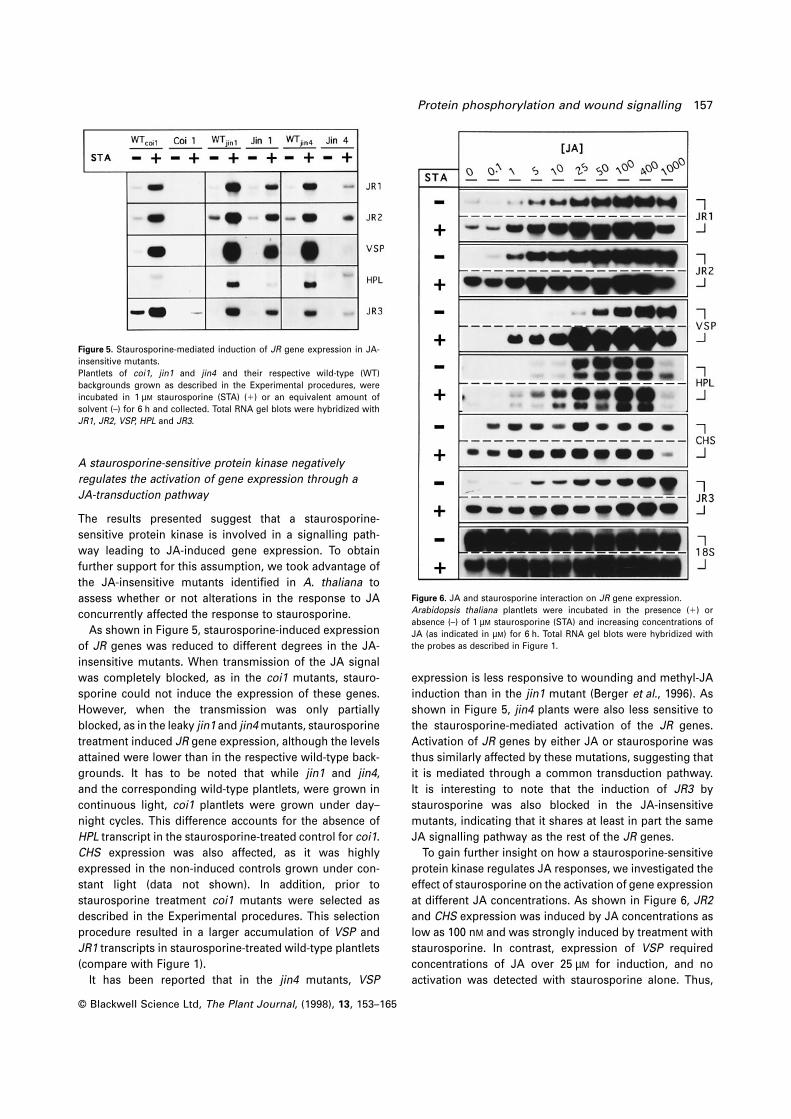
Protein phosphorylation and wound signalling 157
Figure 5. Staurosporine-mediated induction of JR gene expression in JA-
insensitive mutants.
Plantlets of coi1, jin1 and jin4 and their respective wild-type (WT)
backgrounds grown as described in the Experimental procedures, were
incubated in 1 µM staurosporine (STA) (1) or an equivalent amount of
solvent (–) for 6 h and collected. Total RNA gel blots were hybridized with
JR1, JR2, VSP, HPL and JR3.
A staurosporine-sensitive protein kinase negatively
regulates the activation of gene expression through a
JA-transduction pathway
The results presented suggest that a staurosporine-
sensitive protein kinase is involved in a signalling path-
way leading to JA-induced gene expression. To obtain
further support for this assumption, we took advantage of
the JA-insensitive mutants identified in A. thaliana to
assess whether or not alterations in the response to JA
concurrently affected the response to staurosporine.
As shown in Figure 5, staurosporine-induced expression
of JR genes was reduced to different degrees in the JA-
insensitive mutants. When transmission of the JA signal
was completely blocked, as in the coi1 mutants, stauro-
sporine could not induce the expression of these genes.
However, when the transmission was only partially
blocked, as in the leaky jin1 and jin4 mutants, staurosporine
treatment induced JR gene expression, although the levels
attained were lower than in the respective wild-type back-
grounds. It has to be noted that while jin1 and jin4,
and the corresponding wild-type plantlets, were grown in
continuous light, coi1 plantlets were grown under day–
night cycles. This difference accounts for the absence of
HPL transcript in the staurosporine-treated control for coi1.
CHS expression was also affected, as it was highly
expressed in the non-induced controls grown under con-
stant light (data not shown). In addition, prior to
staurosporine treatment coi1 mutants were selected as
described in the Experimental procedures. This selection
procedure resulted in a larger accumulation of VSP and
JR1 transcripts in staurosporine-treated wild-type plantlets
(compare with Figure 1).
It has been reported that in the jin4 mutants, VSP
© Blackwell Science Ltd, The Plant Journal, (1998), 13, 153–165
Figure 6. JA and staurosporine interaction on JR gene expression.
Arabidopsis thaliana plantlets were incubated in the presence (1) or
absence (–) of 1 µM staurosporine (STA) and increasing concentrations of
JA (as indicated in µM) for 6 h. Total RNA gel blots were hybridized with
the probes as described in Figure 1.
expression is less responsive to wounding and methyl-JA
induction than in the jin1 mutant (Berger et al., 1996). As
shown in Figure 5, jin4 plants were also less sensitive to
the staurosporine-mediated activation of the JR genes.
Activation of JR genes by either JA or staurosporine was
thus similarly affected by these mutations, suggesting that
it is mediated through a common transduction pathway.
It is interesting to note that the induction of JR3 by
staurosporine was also blocked in the JA-insensitive
mutants, indicating that it shares at least in part the same
JA signalling pathway as the rest of the JR genes.
To gain further insight on how a staurosporine-sensitive
protein kinase regulates JA responses, we investigated the
effect of staurosporine on the activation of gene expression
at different JA concentrations. As shown in Figure 6, JR2
and CHS expression was induced by JA concentrations as
low as 100 nM and was strongly induced by treatment with
staurosporine. In contrast, expression of VSP required
concentrations of JA over 25 µM for induction, and no
activation was detected with staurosporine alone. Thus,

158 Enrique Rojo et al.
Figure 7. Protein phosphatase activity is required before the step regulated
by staurosporine in the JA signalling pathway.
(a) Arabidopsis thaliana plantlets were subjected to the following
treatments: control plants (lane 1), 50 µM JA (lane 2), simultaneous
incubation in 50 µM JA, 1 µM okadaic acid and 1 µM staurosporine (lane 3),
5 min preincubation in 50 µM JA, subsequently adding 1 µM okadaic acid
and 1 µM staurosporine (lane 4) or 1 µM okadaic acid (lane 5).
(b) Arabidopsis thaliana plantlets were subjected to the following
treatments: control plants (lane 1), 1 µM staurosporine (lane 2), 50 µM JA
(lane 3), 50 µM JA and 20 nM okadaic acid (lane 4), or 50 µM JA, 20 nM
okadaic acid and 1 µM staurosporine (lane 5).
Plantlets were harvested 6 h after the initiation of the treatment, and total
RNA was isolated. Northern blots were hybridized with JR1, JR2 and
VSP probes.
the sensitivity of the genes to staurosporine treatment
correlates with their sensitivity to JA treatment, confirming
that staurosporine activates the genes via a JA transduc-
tion pathway.
When plants were incubated with JA in the presence of
staurosporine, a co-operative effect of both stimuli on the
activation of JR gene expression was observed (Figure 6).
At low JA concentrations, staurosporine strongly enhanced
the JA-induced activation of all JR genes (see for instance
the effect on VSP expression at 1 µM JA). However, JR2,
HPL, CHS and JR3 expression reached a plateau at high
JA concentrations, and incubation with staurosporine
could only enhance this effect slightly. On the other hand,
JA-mediated expression of JR1 and VSP was strongly
enhanced by staurosporine even after the response to JA
was saturated. Thus, the level of expression attained by
treatment with both JA and staurosporine could not be
reached by treatment with JA alone. This indicates that
staurosporine does not induce JA synthesis, but rather
acts on a component, probably a protein kinase, of a JA
transduction pathway. Increasing staurosporine con-
centration did not result in higher levels of JR transcripts
(data not shown), indicating that the protein kinase was
completely inhibited at 1 µM and that the effect of JA was
not mediated by the inactivation of that protein kinase.
The reduction in mRNA levels observed at 1 mM JA,
especially in the case of HPL and CHS, may be a toxic
effect due to the high concentration of the hormone.
© Blackwell Science Ltd, The Plant Journal, (1998), 13, 153–165
The dephosphorylation step affected by okadaic acid is
upstream of the phosphorylation step affected by
staurosporine in the JA-signalling pathway
We have obtained evidence for the presence of both a
protein kinase and a protein phosphatase acting with
opposite effects on a JA transduction pathway. We there-
fore sought to answer where this protein phosphatase
activity was located in the transduction pathway relative
to the step modulated by the protein kinase.
An experiment to address this point is presented in
Figure 7(a). No induction of the JR genes was observed in
plantlets incubated with 50 µM JA in the presence of 1 µM
okadaic acid and 1 µM staurosporine (shown for JR1, JR2
and VSP in Figure 7a, lane 3). This indicates that the protein
phosphatase activity was required for activation by JA and/
or staurosporine. However, if the JA signalling was allowed
to proceed for a short period of time (5 min treatment with
JA alone, in Figure 7a, lane 4), high induction of the genes
was attained upon subsequent incubation with staurospor-
ine and okadaic acid, which was not observed without
staurosporine in the incubation medium. Staurosporine
was therefore able to enhance the expression of JR genes
even in the presence of okadaic acid once the JA signal
had been transmitted for a short time lapse. Thus, the
protein phosphatase activity required for the transduction
of the JA signal probably acts prior to the protein kinase
inhibited by staurosporine, and not downstream of it. An
experiment that lends further support for this model is
shown in Figure 7(b). When plantlets were treated with JA
in the presence of 20 nM okadaic acid, a very weak activa-
tion of JR genes was observed (Figure 7b, lane 4), indicating
that the JA signal was being transduced in part. However,
if staurosporine was also included (lane 5), the signal was
amplified and JR genes were strongly activated.
The okadaic acid-sensitive protein phosphatase activity
thus appeared to be required for JA-induced accumulation
of a positive regulator of this transduction pathway, most
probably a dephosphorylated protein, which was main-
tained in an active state by inhibiting a staurosporine-
sensitive protein kinase. The effect of staurosporine on
gene induction was thus dependent on the JA-induced
accumulation of that activated protein component of the
pathway. Once this dephosphorylated protein had been
produced in sufficient amounts, staurosporine treatment
did not require the protein phosphatase activity to induce
JR gene expression.
Cycloheximide and auxins inhibit the JA-mediated
transcriptional activation of JR genes
We analysed the effect of other putative regulators of
signal transduction pathways on the activation of the
different JR genes upon JA treatment. De novo protein

Protein phosphorylation and wound signalling 159
Figure 8. Effects of cycloheximide and auxins on JA-induced expression of
JR genes.
(a) Arabidopsis thaliana plantlets were incubated for 1 h in the presence
(1) or absence (–) of 50 µM cycloheximide (CHX). 50 µM JA (1) or an
equivalent amount of solvent was subsequently added, and plants were
collected 6 h later.
(b) Arabidopsis thaliana plantlets were incubated for 1 h in the presence
(1) or absence (–) of 10 µM NAA. 50 µM JA (1) or an equivalent amount of
solvent was subsequently added, and plants were collected 6 h later.
RNA gel blots were hybridized with the probes described in Figure 1.
synthesis is required for the transduction of numerous
signals in plants, including JA in potato (Pena Cortes
et al., 1992). This reflects, in some cases, the need for the
synthesis of an inducible component or the renewal of a
labile component of the transduction pathway.
We wanted to ascertain whether protein synthesis is also
required for the transduction of the JA signal in A. thaliana.
To this end, the effect of cycloheximide on JR gene activa-
tion was examined. As shown in Figure 8(a), cycloheximide
blocked JA-mediated induction of all JR genes analysed,
with, perhaps, the exception of JR3. Moreover, cyclo-
heximide blocked staurosporine induction of JR genes
(data not shown), further supporting the involvement of a
staurosporine-sensitive protein kinase in JA signalling. As
JR3 was induced by treatment with cycloheximide alone,
it was not possible to distinguish if de novo protein
synthesis was also required for JR3 activation by JA.
Auxins have been shown to downregulate wound-
induced expression of the JA-responsive pin2 promoter–
CAT gene fusions in tobacco (Kernan and Thornburg, 1989)
and to repress methyl-JA and wound-induced VSP and
lipoxygenase A gene expression in soybean (DeWald et al.,
1994). We examined whether auxin treatment could alter
the response to JA in A. thaliana. As shown in Figure 8(b),
treatment with 10 µM NAA reduced JA-induced accumula-
© Blackwell Science Ltd, The Plant Journal, (1998), 13, 153–165
tion of all JR transcripts, except JR3. This effect was clear
in VSP and HPL, but it has been reproducibly observed for
other JR genes. Treatment with another auxin (2,4-D) gave
similar results (data not shown).
Thus, JA-induced activation of JR1, JR2, VSP, HPL and
CHS was apparently mediated through a common JA
transduction pathway sensitive to auxin treatment and
requiring de novo protein synthesis. The data obtained
with JA-insensitive mutants indicated that JA induction of
JR3 shares a common signalling pathway with other JR
genes up to the step regulated by staurosporine. As
auxins do not induce JR3 expression on their own, they
probably affect a later step in this pathway, separating
auxin-sensitive and -insensitive branches. Alternatively JR3
could be induced through this common JA transduction
pathway and, in addition, through an independent, auxin-
insensitive one.
A wound induction pathway, independent of JA
synthesis, shows regulation by staurosporine, okadaic
acid and cycloheximide opposite to the JA-dependent
one
JA has been reported to be involved in plant responses to
mechanical damage. Upon wounding, JA biosynthesis is
induced and the resulting higher endogenous JA level
triggers activation of wound-induced genes (reviewed in
Pena-Cortes et al., 1995). All A. thaliana JR genes tested
were induced upon wounding (Titarenko et al., 1997), sug-
gesting that wound-induced synthesis of JA mediates the
subsequent upregulation of JR gene expression.
The wound response, however, includes other aspects
not mediated by JA. We have isolated, by differential
mRNA display, two genes, WR3 and choline kinase (CK),
that are induced by mechanical wounding but not by JA
treatment. The wound induction of these genes was not
blocked in the JA-insensitive mutants or by treatment with
inhibitors of JA synthesis (Titarenko et al., 1997). Thus,
wound signalling branches through at least two pathways,
one dependent on JA synthesis, and the other independent.
We analysed the effect of staurosporine, okadaic acid
and cycloheximide on wound-induced gene expression.
As shown in Figure 9, the induction of WR3 and CK
upon wounding was partially blocked by incubation with
staurosporine and amplified in the presence of okadaic
acid. Treatment with okadaic acid alone was sufficient to
activate these two genes, even at concentrations as low
as 20 nM (results not shown). This suggests that a protein
phosphatase, probably type 2A, acts as a negative regulator
in this JA-independent wound signalling pathway, and that
a protein kinase activity is needed for transduction of the
signal. The effect of okadaic acid and staurosporine on the
wound induction of JR genes was similar to the effect
these inhibitors had on their response to JA. This indicates

160 Enrique Rojo et al.
Figure 9. Effects of staurosporine and okadaic acid on wound-induced
gene expression.
Arabidopsis thaliana plantlets were incubated for 1 h in the presence (1)
or absence (–) of 1 µM staurosporine (STA) or 1 µM okadaic acid (OKA), and
subsequently wounded (W) as described above or treated with 50 µM JA
(JA). Control plants (C) were treated with an equivalent amount of the JA
solvent. Plantlets were collected after incubation for 6 h.
RNA gel blots were hybridized with the probes described in Figure 1 and
in addition with the wound-responsive gene 3 (WR3) and choline kinase
(CK) probes.
that indeed their activation by wounding is mediated
through a JA transduction pathway.
Inhibition of protein synthesis led to activation of WR3
and CK, and apparently did not affect their wound-induced
activation (Figure 10). This suggests that a labile
component of this pathway downregulated their expres-
sion in non-wounded plants. In contrast, the wound-
induced activation of JR genes, except JR3, was prevented
by inhibiting protein synthesis (shown in Figure 10 for JR2).
Treatment with okadaic acid or cycloheximide activated
the expression of JR3, in the same manner as WR3 and
CK, and did not affect its wound induction. Moreover,
treatment with inhibitors of JA biosynthesis did not inhibit
the wound-induced activation of JR3, WR3 and CK (J. Leon
et al., submitted). Thus, the wound-induced expression of
JR3 may be mediated through a JA-dependent pathway
and also through a pathway independent of JA synthesis.
© Blackwell Science Ltd, The Plant Journal, (1998), 13, 153–165
Figure 10. Effect of cycloheximide on JR2, JR3, WR3, and CK expression.
Arabidopsis thaliana plantlets were incubated for 1 h in the presence (1)
or absence (–) of 50 µM cycloheximide (CHX), and subsequently they were
(1), or were not (–), wounded (W). Plantlets were collected 4 h later.
RNA gel blots were hybridized with JR2, JR3, WR3 and CK probes, and the
18S rDNA probe as a control for RNA transfer to the blot.
This pathway is probably common to the induction path-
way of WR3 and CK.
Discussion
We have undertaken a pharmacological approach to
identify components of JA-dependent and independent
wound signalling pathways in A. thaliana. In many cases,
cell suspension cultures have been used in such
approaches (Christie and Jenkins, 1996; Felix et al., 1994;
MacKintosh et al., 1994). However, their efficacy in studying
specific hormone-induced responses is hindered by the
possibility of interfering effects due to other hormones
required to maintain culture growth. Indeed, in cell suspen-
sion cultures of A. thaliana we could not detect JA-induced
activation of JR gene expression, except a weak induction
of JR3 (data not shown). We therefore used A. thaliana
plantlets grown in liquid medium, due to the reproduci-
bility of growth conditions and the ease of application of
the compounds tested, which were readily absorbed by
the plants as judged from the effects they had on gene
expression.
Protein phosphatase and kinase inhibitors reveal
phosphorylation events in the transduction of a JA signal
The use of specific inhibitors of protein kinases and
phosphatases has proved useful in the identification of

Protein phosphorylation and wound signalling 161
reversible phosphorylation of proteins as a mechanism
for signal transduction of plant hormones (reviewed in
Smith and Walker, 1996). We have shown that this
regulatory mechanism takes part in the transduction of
a JA signal in A. thaliana.
Okadaic acid and calyculin A, two structurally unrelated
protein phosphatase inhibitors, suppress JA-induced
expresssion of JR genes. This suggests that transmission
of this JA signal in A. thaliana requires a PP1 or
PP2A activity. Similar experiments performed in potato
(Dammann et al., 1997) and tomato (E. Rojo , unpublished
results) showed that okadaic acid also blocks JA-
inducible expression of proteinase inhibitor II (pin2),
cathepsin D inhibitor, leucine aminopeptidase, and
threonine deaminase genes. This indicates that a protein
phosphatase is a component of a JA transduction
pathway conserved in different plant species. This protein
phosphatase is likely to be of type 2A. In our experiments,
the IC50 for okadaic acid inhibition of PP1 activity in
extracts of A. thaliana was ™ 50 nM. The in vivo IC50 is
probably higher due to higher concentrations of the
enzymes and reduced permeability and stability of okadaic
acid in cells. Nevertheless, concentrations as low as
20 nM okadaic acid strongly reduced the JA-induced
activation of JR genes. Specific effects of okadaic
acid concentrations below 30 nM on A. thaliana root
development have been ascribed previously to inhibition
of PP2A activity (Smith et al., 1994). Moreover, we
observed that in extracts from plantlets incubated in vivo
with 1 µM okadaic acid, a concentration exceeding that
needed to abolish JA-induced activation of JR genes,
protein phosphatase activity was inhibited to a degree
attained in vitro at 1 nM okadaic acid, a concentration at
which only PP2A activity is inhibited. A similar approach
has been used previously to provide evidence for the
involvement of a PP2A in the regulation of cdc2 gene
activity in NIH 3T3 cells (Jaramillo-Babb et al., 1996).
However, since the inhibition by okadaic acid may be
reversible (Bialojan and Takai, 1988), the values obtained
in diluted extracts may not exactly match the situation
in vivo.
Our data strongly suggest the involvement of a protein
kinase activity in negatively regulating this JA-signalling
pathway. The protein kinase inhibitor staurosporine activ-
ated the same set of genes as JA itself. This staurosporine-
mediated activation of JR genes required a protein
phosphatase activity and was impaired in the JA-
insensitive mutants coi1, jin1 and jin4, to the same
degree as the JA-mediated activation. Moreover, the
sensitivity of different JR genes to JA or staurosporine
treatment was similar. Staurosporine may therefore inhibit
a protein kinase, and thus activate JR genes through the
same signalling pathway as JA. In experiments conducted
in tomato plantlets, staurosporine activated the expres-
© Blackwell Science Ltd, The Plant Journal, (1998), 13, 153–165
sion of pin2 and the other JA-inducible genes (E. Rojo,
unpublished results), indicating that this may be another
general component of the JA transduction pathway in
different plant species.
We have analysed how JA and staurosporine interact
to activate the expression of JR genes. Maximal activation
can only be attained by incubation with staurosporine
and JA, and not by either compound alone. This indicates
that JA does not activate the pathway by inhibiting the
staurosporine-sensitive kinase, and that the effects of
staurosporine are not due to induction of JA biosynthesis.
Rather, staurosporine and JA act co-operatively on a
step downstream in the JA-signalling pathway.
Our results give some clues to how a staurosporine-
sensitive protein kinase and a protein phosphatase may
interplay in JA signal transduction. When this JA signalling
pathway is blocked initially by okadaic acid, no induction
of target genes is observed. However, if the signal is
transmitted for a short period of time, staurosporine
treatment is able to maintain the pathway and induce
gene expression even if a PP2A activity is completely
inhibited. These results indicate that treatment with JA
triggers, via a PP2A, the transduction cascade and
induces the accumulation of an activated component of
the pathway that can be maintained in the active state by
the staurosporine-mediated inhibition of a protein kinase.
Taken together, these data suggest the presence of a
protein phosphorylation–dephosphorylation switch that
transduces a JA signal when the protein is dephos-
phorylated. The accumulation of the dephosphorylated
target protein probably occurs by a JA-induced activation
of a protein phosphatase, rather than by inducing de novo
synthesis of the target protein.
Treatment with staurosporine induces the expression of
some JR genes. This gene activation does not occur if
plantlets are treated, simultaneously, with staurosporine
and okadaic acid, suggesting that basal levels of all
components of this molecular switch are present under
non-inducing conditions. Staurosporine-induced activation
of JR genes is impaired in the JA-insensitive mutants
tested. However, combined treatment with JA and stauro-
sporine results in similar levels of JR gene expression in
wild-type plants and in the jin1 and jin4 mutants (data not
shown). A possible interpretation of these results is that
the weaker activation of JR genes upon staurosporine
treatment of jin1 and jin4 plantlets is due a reduced
perception of endogenous JA levels in these mutants,
which is largely overcome by the simultaneous application
of exogenous JA. Thus, jin1 and jin4 mutations would
affect components of the signalling pathway prior to the
phosphorylation switch, and not downstream to it as
staurosporine restores wild-type levels of induction in spite
of a reduced transmission of the JA signal.
Because of the more pleiotropic effects of the mutation,

162 Enrique Rojo et al.
coi1 has been suggested to be located before jin1 and jin4
in the JA signalling pathway (Berger et al., 1996). Thus,
mutations in any of these three loci, affecting steps
probably situated between the perception of JA and the
identified phosphorylation switch, block the staurosporine-
mediated activation of JR genes in non-inducing condi-
tions. This suggests that the presence of basal protein
phosphatase activity and/or its target protein depends on
perception of basal JA levels.
Identification of a JA-independent wound induction
pathway that shows opposite regulation to the JA-
dependent one
There is overwhelming evidence indicating that JA plays
a role in plant responses to wounding. A model that
integrates our current knowledge (Bergey et al., 1996;
Farmer and Ryan, 1992; Pena-Cortes et al., 1995; Seo et al.,
1995) suggests that upon wounding a cascade of events,
involving elements such as systemin, ABA and a MAP
kinase, leads to de novo synthesis of JA that, in turn,
regulates gene expression by a previously unknown trans-
duction pathway that may involve ethylene (O’Donnell
et al., 1996). We have shown that the A. thaliana JR genes
can also be regulated by mechanical damage of the plant
(Titarenko et al., 1997), suggesting that their wound induc-
tion is mediated by JA.
However, there is also evidence indicating that wound
responses include JA-independent effects. We have
isolated two genes from A. thaliana, WR3 and CK, that
respond to mechanical injury but not to JA treatment.
Furthermore, inhibitors of JA synthesis or mutations in
the JA transduction pathway do not affect their induction
(Titarenko et al., 1997; Leon et al., submitted), suggesting
that their activation upon wounding does not involve JA
signalling.
The wound induction of JR genes is blocked by treatment
with okadaic acid and enhanced by staurosporine, most
probably as a result of their action on the same phospho-
rylation switch that regulates JA signalling. In contrast, the
wound-inducible expression of WR3 and CK is reduced in
the presence of staurosporine, and enhanced by treatment
with okadaic acid. This indicates that a protein phos-
phorylation switch may also regulate transduction of the
wound signal through this pathway, although in this case
the signal is forwarded by the target protein in the phos-
phorylated state.
The activation of JR genes by wounding requires de
novo protein synthesis for transmission of the signal,
whereas the expression of WR3 and CK is supressed by a
labile protein. A candidate is the protein phosphatase
acting on the phosphorylation switch. Thus, wound-
induced activation of genes through the JA-dependent
and -independent pathways is regulated by protein
© Blackwell Science Ltd, The Plant Journal, (1998), 13, 153–165
Figure 11. A model for wound signalling in Arabidopsis thaliana.
Wound-responsive genes activated through a JA-dependent pathway are
shown on the top set. Genes activated through the JA-independent pathway
are shown on the bottom set. Putative components of both signalling
pathways are shown. Arrows indicate positive action; blunted lines indicate
negative regulation. For positioning the point of action of COI1, JIN1, and
JIN4 we assume in this model that the presence of basal levels of the
protein phosphatase activity, or its target protein, required for
staurosporine-induced JR gene expression depends on JA perception.
W, wounding; PP2A, protein phosphatase type 2A; PK, protein kinase; OKA,
okadaic acid; STA, staurosporine.
phosphorylation and requires protein synthesis in opposite
ways, further supporting the divergence of the wound
signal through, at least, these two pathways. A scheme
depicting the two wound signalling pathways and the sites
of action of the compounds tested is shown in Figure 11.
Other compounds affecting putative components of
these pathways, such as G proteins, PLA2 and Ca21, also
have opposite effects on transduction through them (J.
Leon et al., submitted). Thus, conditions that block one of
the pathways induce the other, suggesting that cross-talk
between them may exist, as observed for other signalling
pathways involving lipid second messengers (Liscovitch
and Cantley, 1994).
A working model for transduction of the wound signal in
which JR may modulate wound-induced gene expression
The JR3 gene is highly similar to ILR1 (Bartel and Fink,
1995), which codes for an enzyme shown to hydrolize
amino acid conjugates of indole-3 acetic acid (IAA) to active
IAA. JR3 expression is induced by wounding and by
exogenous application of JA or staurosporine, and this
induction is reduced in the JA-insensitive mutants coi1,
jin1 and jin4. This indicates that JR3 is activated through
a JA transduction pathway common to the rest of the JR
genes. However, we have obtained data that indicate that
JR3, as well as WR3 and CK, can be induced by wounding
even if de novo synthesis of JA is prevented (J. Leon et al.,
submitted). Moreover, inhibition of either protein synthesis
or protein phosphatase activity is sufficient to activate JR3,

Protein phosphorylation and wound signalling 163
CK and WR3 expression. Taken together, the evidence
suggests that JR3 could be activated by wounding through
two different pathways, one common to the other JA-
inducible genes and the other independent of JA, common
to CK and WR3.
This dual regulation of JR3 expression may provide a
way to modulate the response to mechanical damage. It
has been shown that the concentration of endogenous
auxins declines upon wounding (Thornburg and Li, 1991).
The time required for recovering the initial auxin levels
may thus define the duration of the response to a wound
stimulus. Consistent with this, application of exogenous
auxins reduced the JA-induced expression of JR genes,
except that of JR3, in A. thaliana. If JR3 has a similar
enzyme activity to ILR1, releasing active auxins from the
pool of inactive conjugates, its JA-induced expression
could play a role in feedback inhibition of the JA response
and in restoring the hormonal balance in the plant. An
injury, however, would also induce JR3 expression via a
JA-independent signalling pathway. It is interesting to note
that, in contrast to CK and WR3, the expression of JR3
induced by wounding, okadaic acid and cycloheximide is
blocked in the coi1 mutant (E. Rojo, unpublished data),
indicating that the activation of JR3 through this alternative
wound signalling pathway requires perception of JA. This
requirement would provide the fine tuning in the regulation
of JR3 expression, as it would be switched off once JA
levels return to basal values. Furthermore, the time–course
of expression of JR3 upon wounding (Titarenko et al., 1997)
mirrors the wound-induced increase in the levels of JA in
A. thaliana (Laudert et al., 1996). Thus, JR3 expression
seems to be tightly linked to the presence of levels of JA
above basal values, consistent with its proposed function
in attenuating the JA-mediated response and maintaining
the JA-to-auxin ratio in the plant.
The characterization of reversible protein phosphoryla-
tion switches has allowed the definition of two partially
divergent signalling pathways involved in wound-induced
gene expression. The wealth of sequence information and
molecular genetic tools available in A. thaliana will provide
direct access to the identified components of wound signal-
ling pathways. This will enable their use in reverse genetic
approaches to test the proposed model in an effort comple-
mentary to standard mutational analyses.
Experimental procedures
Plant material and treatments
Seeds of Arabidopsis thaliana ecotype Landsberg erecta (Lehle
Seed Co., Tucson, AZ) were surface sterilized and sown in 24-well
(20 seeds well–1) tissue culture clusters (Costar Corp., Cambridge,
MA), containing 1 ml well–1 of sterile MS medium (Murashige and
Skoog salts; ICN Hubber, Barcelona, Spain) supplemented with
0.5% sucrose, and grown with shaking (150 r.p.m.) in a culture
© Blackwell Science Ltd, The Plant Journal, (1998), 13, 153–165
room under 16-h day (26°C)/8-h night (22°C) diurnal cycles. Fresh
medium, 500 µl well–1, was added 8 days after sowing and experi-
ments were conducted 2 days later.
Jin1, jin4 mutants and their respective wild-type backgrounds
(Columbia ecotype for jin1 and Wassilewskija ecotype in the case
of jin4) were grown (12 seeds well–1) under constant light and
otherwise similar conditions to the Landsberg erecta ecotype
plantlets.
Coi1 mutant seeds were grown for selection on agar plates
containing Murashige and Skoog medium supplemented with 2%
sucrose and 10 µM methyl-JA, under 16-h day (26°C)/8-h night
(22°C) diurnal cycles. Plants were selected on the 6th day post-
germination, transferred to microplates (3 plants well–1) and grown
in the same conditions as the Landsberg erecta ecotype for 7
additional days. Seeds from the corresponding wild-type back-
ground (Columbia) were grown similarly, except that agar plates
did not contain JA.
Prior to treatment initiation, the medium remaining in the wells
was removed and 1 ml well–1 of fresh medium was added. The
different compounds tested were diluted directly in the wells to
their final concentration from stock solutions prepared as follows:
JA (mixed isomers; Apex Organics Ltd, Devon, UK) was diluted
in N,N dimethyl-formamide to obtain 50 mM stock solutions;
okadaic acid, staurosporine and cycloheximide (Sigma-Aldrich
Quimica, Madrid, Spain) were diluted in DMSO to obtain 1 mM,
1 mM and 50 mM stock solutions, respectively.
Wounding was performed by thoroughly crushing the plantlets
with a forceps and subsequent incubation in liquid medium for
the indicated times.
All experiments were independently performed at least three
times, yielding highly reproducible results.
RNA extractions and Northern hybridization
Total RNA from A. thaliana plantlets was isolated and
separated through 1,5% agarose/formaldehyde gels (10 µg lane21)
as described elsewhere (Logemann et al., 1987). RNAs were sub-
sequently blotted onto Hybond-N membranes (Amersham Corp.)
following standard procedures (Sambrook et al., 1989). Equal RNA
loading was assayed by visualizing the ethidium bromide-stained
ribosomal RNA content (not shown), and hybridization of the blots
with a cauliflower 18S ribosomal DNA probe. The inserts of clones
to be used as probes were 32P labelled by the rediprime labelling
system (Amersham Corp.) and hybridized in 0.25 M phosphate
buffer, 0.25 M NaCl, 1 mM EDTA, 7% SDS, 10% polyethylene glycol
6000, 40% formamide, 0.2 mg ml–1 denatured salmon sperm DNA,
at 42°C. Filters were washed in 33SSC, 0.5% SDS at 65°C (13SSC:
150 mM NaCl, 15 mM sodium citrate) and exposed for autoradio-
graphy.
For hybridizations with the different probes, replica blots were
used and hybridized with at most three probes each. Previous
labelling was stripped by washing twice with 0.5% SDS in distilled
water at 65°C for 30 min. The complete removal of the radioactive
probe was assessed by autoradiography.
PP1/PP2A activity assay
Plantlets grown for 10 days in liquid culture in microplates were
frozen in liquid N2, ground to a fine powder and homogenized for
90 sec in 0.7 ml of ice-cold 50 mM Tris–HCl (pH 7.0 at 25°C), 0.1 mM
EDTA, 0.1 mM EGTA, 0.5% Triton, 1 mM PMSF, 1 µg ml–1 leupeptine,
20 µg ml–1 pepstatin and 0.2% (v/v) 2-mercaptoethanol. Extracts
were centrifuged at 15 000 g (4°C) for 10 min, and the super-

164 Enrique Rojo et al.
natant was recovered and assayed for PP1/PP2A activity using a
protein phosphatase assay system (BRL-Life Technologies, Inc.,
Madrid, Spain). Protein concentration was determined using the
protein assay reagent (Bio-Rad Laboratories SA, Madrid, Spain)
and equal amounts of protein from the different extracts were
diluted 1:150 in phosphatase assay buffer (provided in the assay
system) and incubated for 15 min at 30°C with glycogen phos-
phorylase A, freshly labelled with [γ-32P] ATP (3000 Ci mmol–1) as
described in the protocol provided by the manufacturer. Reactions
were performed within the linear range of PP1/PP2A activities
(data not shown). Okadaic acid and tautomycin (1 mM stock
solution in DMSO) were preincubated with the extracts for 10 min
before adding phosphorylase A and allowing the reaction to
proceed.
Acknowledgements
The excellent technical assistance from Tomas Cascon is
gratefully acknowledged. We also wish to thank Ines Poveda
and Angel Sanz for the photographic work. Many thanks to Drs
Isabel Merida, John Mundy and Julio Salinas for their comments
on the manuscript. The hydroperoxide lyase (stock number
94 J16T7) and vegetative storage protein (stock number
108B11T7) clones were obtained from the Arabidopsis Biological
Resource Center at the Ohio State University. The cauliflower
18S ribosomal DNA and the Arabidopsis coi1 seeds were kindly
provided by Drs Julio Salinas and John G. Turner, respectively.
This work was funded in part by the European Communities’
BIOTECH Programme, as part of the Project of Technological
Priority 1993–96. Financial support was also provided by the
Spanish Comision Interministerial de Ciencia y Tecnologıa grants
BIO93–0678-CO2–02, BIO94–1502-CE, and BIO96–0532-CO2–01.
G. Vancanneyt and J. Leon were recipients of a postdoctoral
fellowship and contract, respectively, from the Spanish Ministerio
de Educacion y Ciencia.
References
Albrecht, T., Kehlen, A., Stahl, K., Knofel, H.D., Sembdner, G. and
Weiler, E.W. (1993) Quantification of rapid, transient increases in
jasmonic acid in wounded plants using a monoclonal antibody.
Planta, 191, 86–94.
Anderson, J.M., Spilatro, S.R., Klauer, S.F. and Franceschi, V.R.
(1989) Jasmonic acid-dependent increase in the level of
vegetative storage proteins in soybean. Plant Sci. 62, 45–52.
Bartel, B. and Fink, G.R. (1995) ILR1, an amidohydrolase that
releases active indole-acetic acid from conjugates. Science 268,
1745–1748.
Bell, E., Creelman, R.A. and Mullet, J.E. (1995) A chloroplast
lipoxygenase is required for wound-induced jasmonic acid
accumulation in Arabidopsis. Proc. Natl Acad. Sci. USA, 92,
8675–8679.
Berger, S., Bell, E. and Mullet, J.E. (1996) Two methyl jasmonate-
insensitive mutants show altered expression of AtVsp in
response to methyl jasmonate and wounding. Plant Physiol.
111, 523–531.
Berger, S., Bell, E., Sadka, A. and Mullet, J.E. (1995) Arabidopsis
thaliana Atvsp is homologous to soybean VspA and VspB,
genes encoding vegetative storage protein acid phosphatases,
and is regulated similarly by methyl jasmonate, wounding,
sugars, light and phosphate. Plant Mol. Biol. 27, 933–942.
Bergey, D.R., Howe, G.A. and Ryan, C.A. (1996) Polypeptide
© Blackwell Science Ltd, The Plant Journal, (1998), 13, 153–165
signaling for plant defensive genes exhibits analogies to defence
signaling in animals. Proc. Natl Acad. Sci. USA, 93, 12053–12058.
Bialojan, C. and Takai, A. (1988) Inhibitory effect of a marine-
sponge toxin, okadaic acid, on protein phosphatases. Biochem.
J. 256, 283–290.
Bush, D.S. (1995) Calcium regulation in plant cells and its role in
signaling. Annu. Rev. Plant Physiol. Plant Mol. Biol. 46, 95–122.
Christie, J.M. and Jenkins, G.I. (1996) Distinct UV-B and UV-A/
blue light signal transduction pathways induce chalcone
synthase gene expression in Arabidopsis cells. Plant Cell, 8,
1555–1567.
Cohen, P., Holmes, C.F.B. and Tsukitani, Y. (1990) Okadaic acid: a
new probe for the study of cellular regulation. Trends Biochem.
Sci. 15, 98–102.
Creelman, R.A., Tierney, M.L. and Mullet, J.E. (1992) Jasmonic acid/
methyl jasmonate accumulate in wounded soybean hypocotyls
and modulate wound gene expression. Proc. Natl Acad. Sci.
USA, 89, 4938–4941.
Dammann, C., Rojo, E. and Sanchez-Serrano, J.J. (1997) Abscisic
acid and jasmonic acid activate wound-inducible genes in potato
through separate, organ-specific signal transduction pathways.
Plant J. 11, 773–782.
DeWald, D.B., Sadka, A. and Mullet, J.E. (1994) Sucrose modulation
of soybean Vsp gene expression is inhibited by auxin. Plant
Physiol. 104, 439–444.
Dittrich, H., Kutchan, T. and Zenk, M.H. (1992) The jasmonate
precursor 12-oxo-phytodienoic acid, induces phytoalexin
synthesis in Petroselinum hortense cell cultures. FEBS Lett. 309,
33–36.
Farmer, E.E. and Ryan, C.A. (1992) Octadecanoic precursors of
jasmonic acid activate the synthesis of wound-inducible
proteinase inhibitors. Plant Cell, 4, 129–134.
Farmer, E.E., Johnson, R.R. and Ryan, C.A. (1992) Regulation of
expression of proteinase inhibitor genes by methyl jasmonate
and jasmonic acid. Plant Physiol. 98, 995–1002.
Felix, G., Regenass, M., Spanu, P. and Boller, T. (1994) The protein
phosphatase inhibitor calyculin A mimics elicitor action in
plant cells and induces rapid hyperphosphorylation of specific
proteins as revealed by pulse labeling with [33P]phosphate.
Proc. Natl Acad. Sci. USA, 91, 952–956.
Feys, B.J.F., Benedetti, C.E., Penfold, C.N. and Turner, J.G. (1994)
Arabidopsis mutants selected for resistance to the phytotoxin
coronatine are male sterile, insensitive to methyl-jasmonate,
and resistant to a bacterial pathogen. Plant Cell, 6, 751–759.
Gundlach, H., Muller, M.J., Kutchan, T.M. and Zenk, M.H. (1992)
Jasmonic acid is a signal transducer in elicitor-induced plant
cell cultures. Proc. Natl Acad. Sci. USA, 89, 2389–2393.
Harms, K., Atzorn, R., Brash, A., Kuhn, H., Wasternack, C.,
Willmitzer, L. and Pena-Cortes, H. (1995) Expression of a flax
allene oxide synthase cDNA leads to increased endogenous
jasmonic acid (JA) levels in transgenic potato plants but not to
a corresponding activation of JA-responding genes. Plant Cell,
7, 1645–1654.
Hildmann, T., Ebneth, M., Pena-Cortes, H., Sanchez-Serrano, J.J.,
Willmitzer, L. and Prat, S. (1992) General roles of abscisic acid
and jasmonic acid in gene activation as a result of mechanical
damage. Plant Cell, 4, 1157–1170.
Ishihara, H., Martin, B.L., Brautigan, D.L. et al. (1989) Calyculin A
and okadaic acid, inhibitors of protein phosphatases activity.
Biochem. Biophys. Res. Commun. 159, 871–877.
Jaramillo-Babb, V.L., Sugarman, J.L., Scavetta, R., Wang, S.J.,
Berndt, N., Born, T.L., Glass, C.K. and Schoenthal, A.H. (1996)
Positive regulation of cdc2 gene activity by protein phosphatase
type 2A. J. Biol. Chem. 271, 5988–5992.

Protein phosphorylation and wound signalling 165
Kakimoto, T. (1996) CKI1, a histidine kinase homolog implicated
in cytokinin signal transduction. Science, 274, 982–985.
Kernan, A. and Thornburg, R.W. (1989) Auxin levels regulate
the expression of a wound-inducible proteinase inhibitor II-
chloramphenicol acetyl transferase gene fusion in vitro and
in vivo. Plant Physiol. 91, 73–78.
Kieber, J.J., Rothenberg, M., Roman, G., Feldmann, K.A. and
Ecker, J.R. (1993) CTR1, a negative regulator of the ethylene
response pathway in Arabidopsis, encodes a member of the
Raf family of protein kinases. Cell, 72, 427–441.
Koda, Y., Kikuta, Y., Tazaki, H., Tsujino, Y., Sakamura, S. and
Yoshihara, T. (1991) Potato tuber-inducing activities of jasmonic
acid and related compounds. Phytochem. 30, 1435–1438.
Kuo, A., Cappelluti, S., Cervantes-Cervantes, M., Rodriguez, M.
and Bush, D.S. (1996) Okadaic acid, a protein phosphatase
inhibitor, blocks calcium changes, gene expression, and cell
death induced by gibberellin in wheat aleurone cells. Plant Cell,
8, 259–269.
Laudert, D., Pfannschmidt, U., Hollander-Czytko, H. and Weiler,
E.W. (1996) Cloning, molecular and functional characterization
of Arabidopsis thaliana allene oxide synthase (CYP 74), the first
enzyme of the octadecanoid pathway to jasmonates. Plant Mol.
Biol. 31, 323–335.
Leung, J., Bouvier-Durand, M., Morris, P., Guerrier, D., Chefdor, F.
and Giraudat, J. (1994) Arabidopsis ABA response gene ABI1:
features of a calcium-modulated protein phosphatase. Science,
264, 1448–1452.
Liscovitch, M. and Cantley, L.C. (1994) Lipid second messengers.
Cell, 77, 329–334.
Logemann, J., Schell, J. and Willmitzer, L. (1987) Improved method
for the isolation of RNA from plant tissues. Anal. Biochem. 163,
16–20.
Ma, H. (1994) GTP-binding proteins in plants: new members of an
old family. Plant Mol. Biol. 26, 1611–1636.
McConn, M. and Browse, J. (1996) The critical requirement for
linolenic acid is pollen development, not photosynthesis, in an
arabidopsis mutant. Plant Cell, 8, 403–416.
MacKintosh, C. and Cohen, P. (1989) Identification of high levels
of type 1 and type 2A protein phosphatases in higher plants.
Biochem. J. 262, 335–339.
MacKintosh, C. and Klumpp, S. (1990) Tautomycin from the
bacterium Streptomyces verticillatus. Another potent and
specific inhibitor of protein phosphatases 1 and 2A. FEBS Lett.
277, 137–140.
MacKintosh, C. and MacKintosh, R.W. (1994) Inhibitors of protein
kinases and phosphatases. Trends Biochem. Sci. 19, 444–448.
MacKintosh, C., Coggings, J. and Cohen, P. (1991) Plant protein
phosphatases: subcellular distribution, detection of protein
phosphatase 2C and identification of protein phosphatase 2A
as the major quinate dehydrogenase phosphatase. Biochem. J.
273, 733–738.
MacKintosh, C., Lyon, G.D. and MacKintosh, R.W. (1994) Protein
phosphatase inhibitors activate anti-fungal defence responses
of soybean cotyledons and cell cultures. Plant J. 5, 137–147.
Mason, H.S. and Mullet, J.E. (1990) Expression of two soybean
© Blackwell Science Ltd, The Plant Journal, (1998), 13, 153–165
vegetative storage protein genes during development and in
response to water deficit, wounding and jasmonic acid. Plant
Cell, 2, 569–579.
Meyer, K., Leube, M.P. and Grill, E. (1994) A protein phosphatase
2C involved in ABA signal transduction in Arabidopsis thaliana.
Science, 264, 1452–1455.
O’Donnell, P.J., Calvert, C., Atzorn, R., Wasternack, C., Leyser,
H.M.O. and Bowles, D.J. (1996) Ethylene as a signal mediating
the wound response of tomato plants. Science, 274, 1914–1917.
Pelacho, A.M. and Mingo-Castel, A.M. (1991) Jasmonic acid
induces tuberisation of potato stolons cultured in vitro. Plant
Physiol. 97, 1253–1255.
Pena-Cortes, H., Albrecht, T., Prat, S., Weiler, E.W. and Willmitzer,
L. (1993) Aspirin prevents wound-induced gene expression in
tomato leaves by blocking jasmonic acid biosynthesis. Planta,
191, 123–128.
Pena-Cortes, H., Fisahn, J. and Willmitzer, L. (1995) Signals
involved in wound-induced proteinase inhibitor II gene
expression in tomato and potato plants. Proc. Natl Acad. Sci.
USA, 92, 4106–4113.
Pena-Cortes, H., Liu, X., Sanchez-Serrano, J.J., Schmid, R. and
Willmitzer, L. (1992) Factors affecting gene expression of patatin
and proteinase-inhibitor-II gene families in detached potato
leaves: implications for their co-expression in developing tubers.
Planta, 186, 495–502.
Raz, V. and Fluhr, R. (1993) Ethylene signal is transduced via
protein phosphorylation events in plants. Plant Cell, 5, 523–530.
Reinbothe, S., Mollenhauer, B. and Reinbothe, C. (1994) JIPs and
RIPs: the regulation of plant gene expression by jasmonates in
response to environmental cues and pathogens. Plant Cell, 6,
1197–1209.
Sambrook, J., Fritsch, E.F. and Maniatis, T. (1989) Molecular
Cloning. A Laboratory Manual, 2nd Edn. NY: Cold Spring Harbor
Laboratory Press.
Sembdner, G. and Parthier, B. (1993) The biochemistry and the
physiological and molecular actions of jasmonates. Annu. Rev.
Plant Physiol. Plant Mol. Biol. 44, 569–589.
Seo, S., Okamoto, M., Seto, H., Ishizuka, K., Sano, H. and Ohashi,
Y. (1995) Tobacco MAP kinase: a possible mediator in wound
signal transduction pathways. Science 270, 1988–1992.
Smith, R.D. and Walker, J.C. (1996) Plant protein phosphatases.
Annu. Rev. Plant Physiol. Plant Mol. Biol. 47, 101–125.
Smith, R.D., Wilson, J.E., Walker, J.C. and Baskin, T.I. (1994)
Protein phosphatase inhibitors block root hair growth and alter
cortical cell shape of Arabidopsis roots. Planta, 194, 516–524.
Tamaoki, T., Nomoto, H., Takahashi, I., Kato, Y., Morimoto, M. and
Tomita, F. (1986) Staurosporin, a potent inhibitor of
phospholipid/Ca21 dependent protein kinase. Biochem. Biophys.
Res. Commun. 135, 397–402.
Thornburg, R.W. and Li, X. (1991) Wounding Nicotiana tabacum
leaves causes a decline in endogenous indole-3-acetic acid.
Plant Physiol. 96, 802–805.
Titarenko, E., Rojo, E., Leon, J. and Sanchez-Serrano, J.J. (1997)
JA-dependent and independent signalling pathways control
wound-induced gene activation in Arabidopsis thaliana. Plant
Physiol. 115, 817–826.